Embed Size (px)
Citation preview

INVESTIGATION
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Buddedagger and Mark B Rothdagger1
Molecular and Cellular Biology Graduate Program University of Washington Seattle Washington 98195 and daggerDivision of BasicSciences Fred Hutchinson Cancer Research Center Seattle Washington 98109
ABSTRACT Hydrogen sulfide (H2S) an endogenously produced small molecule protects animals from various stresses Recent studiesdemonstrate that animals exposed to H2S are long lived resistant to hypoxia and resistant to ischemiandashreperfusion injury Weperformed a forward genetic screen to gain insights into the molecular mechanisms Caenorhabditis elegans uses to appropriatelyrespond to H2S At least two distinct pathways appear to be important for this response including the H2S-oxidation pathway and thehydrogen cyanide (HCN)-assimilation pathway The H2S-oxidation pathway requires two distinct enzymes important for the oxidationof H2S the sulfidequinone reductase sqrd-1 and the dioxygenase ethe-1 The HCN-assimilation pathway requires the cysteinesynthase homologs cysl-1 and cysl-2 A low dose of either H2S or HCN can activate hypoxia-inducible factor 1 (HIF-1) which is requiredfor C elegans to respond to either gas sqrd-1 and cysl-2 represent the entry points in the H2S-oxidation and HCN-assimilationpathways respectively and expression of both of these enzymes is highly induced by HIF-1 in response to both H2S and HCN Inaddition to their role in appropriately responding to H2S and HCN we found that cysl-1 and cysl-2 are both essential mediators ofinnate immunity against fast paralytic killing by Pseudomonas Furthermore in agreement with these data we showed that growingworms in the presence of H2S is sufficient to confer resistance to Pseudomonas fast paralytic killing Our results suggest the hypoxia-independent hif-1 response in C elegans evolved to respond to the naturally occurring small molecules H2S and HCN
HYDROGEN sulfide (H2S) is an endogenously producedmolecule with profound physiological effects (Kabil
and Banerjee 2010) While first noted for its potent toxicityit is now clear that organisms and H2S have long had anintimate relationship (Philippot et al 2007) Prior to theappearance of oxygen as a component of the earthrsquos atmo-sphere H2S was abundant at the earthrsquos surface (Reinhardet al 2009) Sulfur was likely important for redox bio-chemistry in early life similar to the role of oxygen today(Philippot et al 2007) Sulfur-containing redox reactions arestill essential in modern organisms in the form of glutathi-one and thioredoxin (Holmgren et al 2005) and in ironndashsulfur clusters in the electron transport chain and aconitasein the tricarboxylic acid (TCA) cycle (Rouault and Tong2008) Recent molecular phylogenetic data confirm thatH2S-interacting enzymes predated oxygen-binding enzymes
(David and Alm 2011) H2S is also essential for the existenceof sulfur-containing amino acids because cysteine synthaseuses H2S as the sulfur source during de novo cysteine bio-synthesis (Feldman-Salit et al 2009)
In animals H2S is generated by dissimilating H2S fromcysteine (Kabil and Banerjee 2010) The numerous activitiesascribed to H2S in animals include its function as a neuromo-dulator in the brain and as a mediator of vascular tone andsmooth muscle of the heart (Li et al 2011) The ability of H2Sto affect mammalian tissue has recently been exploited fortherapeutic uses (Mustafa et al 2009) When rodents areexposed to H2S they enter a reversible state of decreasedmetabolic rate and core body temperature (Blackstone et al2005) Exogenous H2S has shown benefit in a number ofanimal-disease models For example H2S protects mice fromhypoxic death (Blackstone and Roth 2007) helps rats survivesevere hemorrhage (Morrison et al 2008) and diminishesischemiandashreperfusion injury (Elrod et al 2007)
Our understanding of the mechanism by which H2Saffects biology is incomplete To fill these gaps we havedeveloped C elegans as a genetically amenable model sys-tem for studying the response of animals to H2S Previouswork has shown that C elegans becomes longer lived and
Copyright copy 2011 by the Genetics Society of Americadoi 101534genetics111129841Manuscript received April 19 2011 accepted for publication July 28 2011Available freely online through the author-supported open access optionSupporting information is available online at httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC11Corresponding author Fred Hutchinson Cancer Research Center A3-015 1100Fairview Ave N Seattle WA 98109 E-mail mrothfhcrcorg
Genetics Vol 189 521ndash532 October 2011 521
thermotolerant when grown in H2S (Miller and Roth2007) We showed that H2S induces the accumulation ofthe hypoxia-inducible factor 1 (HIF-1) (Budde and Roth2010) In turn hif-1 is required for survival in the presenceof H2S and keeping HIF-1 levels constitutively high confersresistance to otherwise lethal H2S concentrations HIF-1 isa transcription factor and thus the observed protectiveeffects are likely to be mediated by increased expressionof genes regulated by HIF-1 To determine the molecularmechanism by which tolerance to H2S is conferred weperformed a forward genetic screen for C elegans strainssensitive to H2S and report on two hif-1 target genes thatfunction in the first steps of two distinct pathways impor-tant for survival in H2S
Materials and Methods
Strains
C elegans were grown at room temperature on nematodegrowth medium plates seeded with live Escherichia coliOP50 food (Brenner 1974) The following mutant strainswere obtained from the Caenorhabditis Genetics Centercysl-1(ok762) cysl-2(ok3516) cysl-4(ok3359) cbs-2(ok666)hif-1 (ia04) sqrd-2 (ok3440) and CB4856 The followingstrains were provided by Shohei Mitani of the National Bio-Resource Project sqrd-1(tm3378) and ethe-1(tm4104) Pseu-domonas aeruginosa strain PA01 was obtained from ColinManoil (University of Washington Seattle WA)
Atmospheric chambers and viability tests
H2S-containing atmospheric chambers were generated bymixing air with small amounts of defined compressed H2Sbalanced with N2 (Budde and Roth 2010) The resulting gasmixture is essentially room air containing hydrogen sulfide(RAH2S) Unless otherwise stated RAH2S is room air con-taining 50 ppm of H2S All survival experiments were per-formed in 1000-ml chambers with a gas flow of 1000 mlminHCN-containing atmospheres were created in the same wayas the H2S atmospheres except HCN balanced with N2 wasused and they are referred to as RAHCN (essentially roomair containing hydrogen cyanide) Unless otherwise statedRAHCN is room air containing 5 ppm of HCN
Viability tests were performed by picking L4 larval stageworms onto OP50-seeded NGM plates and placing them intothe gaseous atmosphere for 24 hr (Budde and Roth 2010)For RAH2S exposures all animals observed after 24 hrwere either healthy adults or appeared to have perishedshortly after initiation of the experiment For RAHCN expo-sures developmentally arrested L4 animals with very slowmovement were occasionally observed If maintained inRAHCN these animals invariably perished after severaldays as L4 If removed from RAHCN after 24 hr some ofthese animals eventually recovered to become fertile adultsFor the data in this article animals with this developmentalarrest were scored as not surviving the RAHCN exposureAll error values presented are standard deviation (SD)
Video acquisition and automated worm speed analysis
A Quickcam Communicate STX (Logitech) webcamwas affixedto the eyepiece of a stereomicroscope Images were acquiredevery 12 sec and analyzed with the Multi-Worm Tracker(MWT) (Swierczek et al 2011) The video was analyzed threetimes on the MWT and the speed was averaged every 25 min
Genetic screen for H2S-sensitive mutants
Ethyl methanesulfonate (EMS) mutagenesis was performedas previously described (Brenner 1974) and was optimizedto maximize the number of unique mutations observed(Shaham 2007) N2 animals at the L4 or young-adult stagewere incubated with EMS for 4 hr The worms were thenwashed once aliquoted onto four OP50-seeded NGM platesand allowed to lay eggs overnight The next day the adultswere washed off the plates and the F1 animals were grownto adults Each pool of 25000 F1 adults was bleached togenerate synchronized F2 animals A total of 100000 F2L4 animals were screened (25000 from each pool)
The screen for nematodes that are sensitive to H2S wasperformed as follows When the F2 worms entered the L4larval stage the plate of worms was exposed to RAH2S for4 hr The worms were moved to room air for 5 min and thenall paralyzed worms were picked from the plate within15 min of removal from RAH2S From this screen 791 F2animals were singled out to generate 544 viable strains Ofthese viable strains 124 strains exhibited a lethality pheno-type during a 24-hr exposure to RAH2S Of the 124 sensi-tive strains 23 were 100 lethal when exposed to RAH2Sfor 24 hr as L4 and were annotated as strongly penetrant(Figure 2)
Mapping mutations to chromosomes andcomplementation tests
Prior to complementation testing strongly penetrant strainswere crossed to the Hawaiian strain CB4856 and SNP enrich-ment analysis was performed (Davis et al 2005) MaleCB4856 worms were mated into the 23 mutant strains F1animals were isolated and synchronous F2 populations werecreated by egg lays When F2 animals reached L4 larval stagethe plates were exposed to RAH2S for 4 hr Thirty paralyzedanimals were picked as phenotypic and 30 motile animalswere picked as aphenotypic In most cases 25 of F2worms became paralyzed when exposed to RAH2S and wereannotated as recessive In four cases75 of worms becameparalyzed and these mutants were annotated as dominant
After the restriction digests were separated by gel electro-phoreses the gel was visually examined for genomic regionswhere the DNA from phenotypic and aphenotypic animals didnot cut equally well with DraI Ethidium bromide fluores-cence was quantified with Quantity One 1-D analysis soft-ware (Bio-Rad) and a fluorescence ratio of cutuncut DNAwas generated for each SNP on the chromosome of interestThe SNPs on two neighboring chromosomes are presented asa background control Strains that appeared to show similar
522 M W Budde and M B Roth
enrichment patterns were grouped together as suspectedcomplementation groups (Figure 3)
Complementation tests were carried out by crossing homo-zygous males of one strain into another strain and testing theF1 progeny for survival in RAH2S Sequencing of cysl-1 andsqrd-1 was accomplished by PCR amplification of genomicDNA and then using nested primers for the labeling reactionAfter the point mutations were identified SIFT-BLink (Ngand Henikoff 2001) (httpsiftbiia-staredusgwwwSIFT_BLink_submithtml) was used to determine whether the muta-tions are predicted to be tolerated or not tolerated In parallelall splice mutations were annotated as not tolerated
Protein homology
Proteinndashprotein Basic Local and Alignment and Search Tool(BLASTp) was used to locate homologous genes Potentialhomologs were reciprocally BLAST searched to ensure speci-ficity Amino acid alignment was performed with ClustalX2using the following National Center for Biotechnology Infor-mation reference sequence IDs Arabidopsis thaliana CASNP_1917031 A thaliana CYS NP_1932241 Solanum tuber-osum CAS BAB187601 S tuberosum CYS AAC256351 Zeamays CAS ADG602361 Z mays CYS NP_0011054691Mus musculus CBS NP_6591041 Homo sapiens CBSNP_0000621 Ciona intestinalis CBS XP_0021202471 andDrosophila melanogaster CBS NP_6084241
Psuedomonas aeruginosa dependent paralysis
Pseudomonas experiments were performed with strain PA01as previously described (Shao et al 2010) Briefly a fulllawn of P aeruginosa was grown on brain heart infusionagar (37 g Difco BHI 20 g agar per liter) on 35-cm platesfor 24 hr at 37 The plates were cooled to room tempera-ture and 10 L4 animals were placed onto the bacterial lawnand the plate was sealed with parafilm The number of ani-mals moving and not moving after 6 hr was recorded
mRNA detection by QRTndashPCR
Quantitative reverse transcriptase PCR was performed ona Bio-Rad C1000 Thermal Cycler with a CFX96 real-timesystem with iQ SYBR green supermix (Bio-Rad) as pre-viously described (Budde and Roth 2010) ExperimentalmRNA was compared to the average mRNA level of thehousekeeping genes tba-1 and tbb-2 The primer sequencesare listed in Supporting Information Table S1
Western blotting
Antibody was generated and affinity purified using theamino acids 419ndash431 of SQRD-1 (wormbaseorg WPCE17629) amino acids AMETTPFDQSKPTY by ProSci In-corporated (San Diego CA) Western blotting was per-formed as previously described (Budde and Roth 2010)
Figure 1 Wild-type nematodes exposedto 50 ppm RAH2S exhibit a brief in-crease in movement initially and thencontinue to move (A black line) Wheneither wild-type or hif-1 mutant animalsare exposed to 150 ppm RAH2S or50 ppm RAH2S respectively they stopmoving and when the RAH2S is re-moved they resume movement (blueand red lines A and B) Examples ofHif-1 moving stopped and reanimatedworms are shown in C
The Response of C elegans to H2S and HCN 523
with the following changes primary antibody was diluted1500 Goat antirabbit IgG (H+L)-horseradish peroxidase(catalog no 111-035-003 Jackson ImmunoReseach Labora-tories West Grove PA) was used as the secondary antibody
Fosmid recombineering
GFP was inserted into fosmid WRM066CH11 as previouslydescribed (Dolphin and Hope 2006) The oligonucleotidesused to insert the RT cassette were
RT cassette forward -gtttgattaaaggatactggaatggaccagctacactcagaaattgtacaTCGCTGTCGAGATATGACGGTG
RT cassette reverse -acaaataaaaacgacaaagtgggaaatatctatttagactttaccaatcgGATGATAAGCTGTCAAACATGAG
GFP forward -acaaataaaaacgacaaagtgggaaatatctatttagactttaccaatcgTTTGTATAGTTCATCCATGCCATGTG
GFP reverse -gtttgattaaaggatactggaatggaccagctacactcagaaattgtacaATGAGTAAAGGAGAAGAACTTTTC
Uppercase letters indicate primers for the PCR reaction
Generation of transgenic animals
Wild-type C elegans were injected with 50 ngml fosmidtogether with 100 ngml pRF4 which confers a dominantroller [rol-6(su1006)] phenotype (Mello et al 1991) Roll-ing F1 animals were picked onto single plates to generatetransgenic lines Animals were exposed to 50 ppm RAH2Sprior to confocal fluorescent microscopy as previously de-scribed (Budde and Roth 2010)
RNAi experiments
RNAi was performed as previously described (Frazier andRoth 2009) using WBRNAi00013759 to target egl-9 L4 ani-mals were moved onto the RNAi food and the progeny were
moved onto NGM-OP50 or BHI-PA01 plates and immedi-ately assayed
Results
We showed previously that hypoxia-inducible factor hif-1 isrequired for growth and survival in H2S (Budde and Roth2010) HIF-1 protein concentration and activity are in-creased when C elegans are moved into RAH2S and egl-9mutant animals which have constitutively high HIF-1 ex-pression and activity are resistant to H2S
During our investigations with hif-1 mutants we noticedthat although hif-1 mutant animals are unable to surviveprolonged exposure to RAH2S (24 hr) a brief exposure(4 hr) causes a reversible paralysis (Figure 1A) Roughly 2hr after entering an environment containing RAH2S hif-1mutant L4 animals enter a state of suspended animationcharacterized by a complete cessation of movement andfeeding (Figure 1B) and the animals lose their characteristicsinusoidal shape (Figure 1C) If removed to fresh air duringthis suspended state the animals resume moving after20 min and proceed to become fertile adults This pheno-type is also present in wild-type animals exposed to 150 ppmRAH2S (Figure 1 A and B File S1) We used this pheno-type as a basis for screening mutant C elegans that are sen-sitive to H2S in an attempt to find genes that conferresistance to H2S (Figure 2A)
About 200000 mutant haploid genomes were screenedfor H2S sensitivity Through screening we obtained 124
Figure 3 Enrichment of H2S-sensitive phenotype after crossing mutantlines with the Hawaiian strain CB4856 SNP mapping data show fiveapparent complementation groups For suls-1 and suls-2 the apparentlocation of mutation is indicated by an arrow The known locations ofsqrd-1 cysl-1 and hif-1 are also indicated by arrows Black indicates anN2HI ratio 5
Figure 2 H2S sensitivity screen method and results (A) Schematic ofinitial paralysis screen and lethality rescreen (B) Histogram of the numberof strains isolated for each penetrance bin The strains with zero survivorsafter sulfide exposure are highlighted in red These strains were subse-quently characterized
524 M W Budde and M B Roth
lines that were sensitive to H2S (Figure 2B) Of these lines23 were completely penetrant where penetrance is definedas the percentage of worms unable to survive a 24-hr RAH2S exposure These mutant strains were chosen for furthercharacterization
The 23 strongly penetrant mutant lines were crossed withthe Hawaiian CB4856 strain for mapping the sensitivity traitto chromosomal regions using enrichment of SNPs The SNPanalysis revealed that 22 mutants map to only five chromo-somal regions (Figure 3 Table 1) only mr37 showed noMendelian inheritance of the trait These mutants were thencrossed to confirm that each chromosomal region containeda single complementation group (Figure 4) One group onchromosome V (mr18 mr27 mr35 and mr38) exhibitsdominant genetics and thus could not be further tested forcomplementation We named this gene suls-1 (sulfide sen-sitive) and suls-1 (mr35) was mapped to the 4-cM regionbetween SNP pkP5097 and pkP5068 The two alleles onchromosome I were named suls-2 The genes mutated insuls-1 and suls-2 strains remain to be identified Two alleleson chromosome V (mr20 and mr22) mapped near the hif-1locus and failed to complement the hif-1(ia04) mutationwhich confirms that we successfully screened for our phe-notype of interest
The 23 strongest mutant lines were next tested forsurvival in room air containing lower H2S concentrationsin an effort to phenotypically discriminate between the com-
plementation groups Table 1 shows that complementationgroups have differences in H2S sensitivity with the groupmapping to the X chromosome being the most sensitive andsuls-2 being the least sensitive In contrast to the other com-plementation groups suls-1 alleles vary in sensitivity to H2S
A cysteine synthase homolog cysl-1 is required forsurvival in H2S
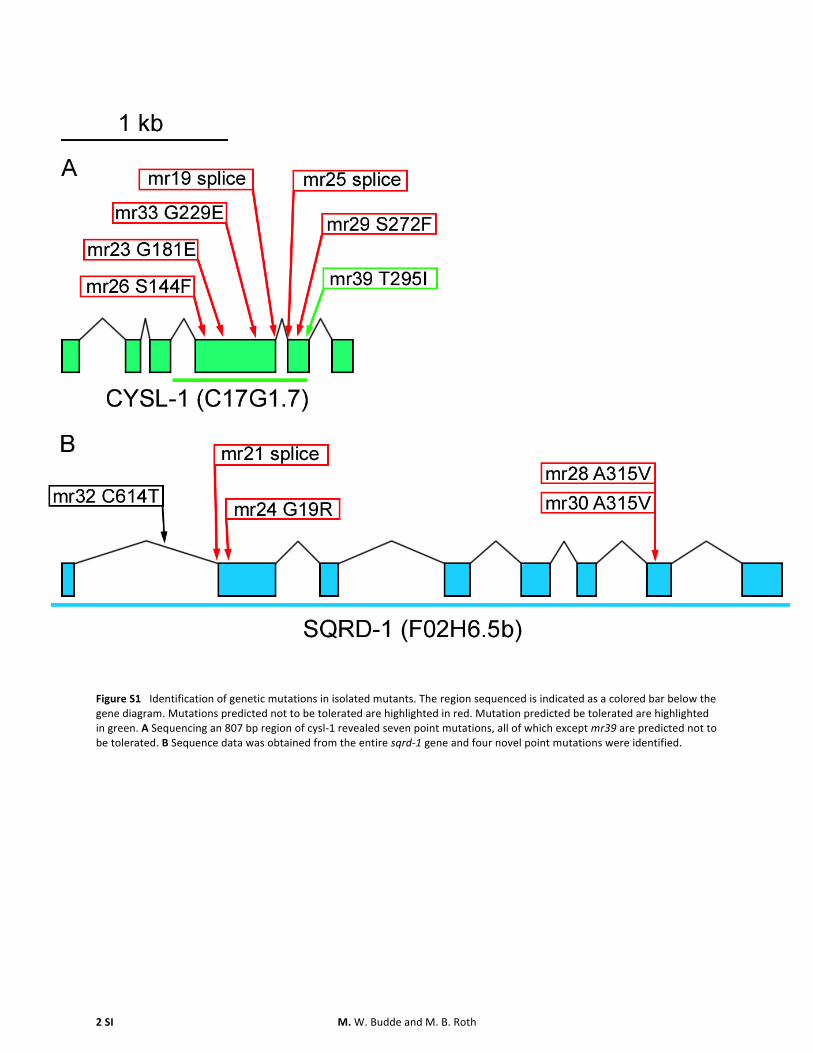
mr23 was used as a representative allele of the group ofalleles on the X chromosome (mr19 mr23 mr25 mr26mr29 mr33 mr34 mr39 and mr40) and was mapped toa 02-cM region between SNPs pas23398 and snp_F41E7[3] This region contains 70 predicted genes Of the deletionmutants available for that region ok762 phenocopies andfails to complement these H2S-sensitive mutations Theok762 allele deletes five of the six exons of the geneC17G17 Sequencing a portion of C17G17 of our isolatedstrains revealed adenine-to-guanine point mutations whichare characteristic of EMS mutagenesis (Figure S1) All iden-tified mutations were either missense or splice mutations
C17G17 is a previously uncharacterized gene encodinga member of the cystathionine b-synthase (CBS)-like sub-group which is made up of enzymes important in cysteineproduction CBS and cysteine synthase (CYS) (OAS-TL andCysK) (Feldman-Salit et al 2009) Animals generally haveCBS homologs and bacteria and plants tend to have CYShomologs (Hell et al 2008) CBS is critical for generating
Table 1 Alleles unable to survive in RAH2S
Alivetotal ( surviving)
Gene name Chromosome Allele 32 ppm RAH2S 15 ppm RAH2S 75 ppm RAH2S Dominance
N2 mdash mdash 1212 (100) 1212 (100) 1313 (100) mdash
cysl-1 X ok762 020 (0) 018 (0) 011 (0) mdash
cysl-1 X mr19 013 (0) 012 (0) 010 (0) Recessivecysl-1 X mr23 016 (0) 010 (0) 010 (0) Recessivecysl-1 X mr25 013 (0) 010 (0) 06 (0) Recessivecysl-1 X mr26 012 (0) 010 (0) 011 (0) Recessivecysl-1 X mr33 015 (0) 59 (56) 1010 (100) Recessivecysl-1 X mr34 010 (0) 011 (0) 110 (10) Recessivecysl-1 X mr29 011 (0) 010 (0) 113 (8) Recessivecysl-1 X mr39 010 (0) 010 (0) 1313 (100) Recessivecysl-1 X mr40 010 (0) 010 (0) 111 (9) Recessivesqrd-1 IV tm3378 510 (50) 1010 (100) 1010 (100) mdash
sqrd-1 IV mr24 1014 (71) 1111 (100) mdash Recessivesqrd-1 IV mr30 213 (15) 99 (100) mdash Recessivesqrd-1 IV mr32 1213 (92) 1010 (100) mdash Recessivesqrd-1 IV mr21 615 (40) 1010 (100) mdash Recessivesqrd-1 IV mr28 59 (56) 1010 (100) 1111 (100) Recessivehif-1 V ia04 015 (0) 011 (0) 1010 (100) mdash
hif-1 V mr20 014 (0) 010 (0) 1013 (77) Recessivehif-1 V mr22 015 (0) 111 (9) 810 (80) Recessivesuls-1 V mr18 612 (50) 1112 (92) 1010 (100) Dominantsuls-1 V mr27 29 (22) 1010 (100) mdash Dominantsuls-1 V mr35 05 (0) 010 (0) 010 (0) Dominantsuls-1 V mr38 09 (0) 88 (100) mdash Dominantsuls-2 I mr31 78 (88) 88 (100) mdash Recessivesuls-2 I mr36 89 (89) 1111 (100) mdash Recessivemdash mdash mr37 1112 (92) 1111 (100) mdash mdash
The Response of C elegans to H2S and HCN 525
cysteine in animals which make cysteine from dietary me-thionine (Finkelstein et al 1988) CBS is also one of the onlyH2S-generating enzymes in mammals (Kabil and Banerjee2010) In plants and bacteria which are cysteine proto-trophs CYS combines H2S with O-acetylserine (OAS) togenerate cysteine (Figure 5A) (Wirtz et al 2010) Humansand mice each have one CBS-like family member In con-trast C elegans has six C17G17 K10H102 R08E52F59A79 and F54A34 (Table 2)
C17G17 (cysl-1) K10H102 (cysl-2) R08E52 (cysl-3) andF59A79 (cysl-4) are more similar to CYS from plants andbacteria whereas ZC3731 (cbs-1) and F54A34 (cbs-2) aremore similar to CBS from animals (Figure 5C) A BLASTsearch reveals that the genus Caenorhabditis contains the onlyknown animal CYS homologs We have renamed these genescysl-1 (cysteine synthase-like) cysl-2 cysl-3 cysl-4 cbs-1 andcbs-2 to reflect this homology Depletion of H2S levels as it isused as a substrate by CYS may explain why cysl-1 mutantanimals are sensitive to H2S Nematodes appear to have ac-quired the ability to assimilate H2S and cysl-1 is likely usingthis reaction to drive down internal H2S levels
cysl-2 mediates immunity to fast paralytic killingby Pseudomonas
The existence of three additional Cysl homologs in C elegansraises the possibility that these other Cysl enzymes are alsoimportant in H2S detoxification This possibility is especiallyintriguing because we have shown a hif-1ndashdependent in-crease in cysl-2 transcript levels during RAH2S exposure(Budde and Roth 2010) We obtained the available knock-out strains of cysl-2 cysl-4 and cbs-2 and tested them forRAH2S sensitivity but none showed a RAH2S sensitivityphenotype (Table 2)
In plants CYS and cyanoalanine synthase (CAS) areclosely related and have overlapping catalytic activities(Figure 5 A and B) The CAS reaction combines cysteineand HCN to produce b-cyanoalanine and H2S lowering cel-lular HCN levels (Figure 5B) Furthermore cysl-2 mRNA waspreviously shown to be upregulated by HIF-1 (Shen et al2005 Shao et al 2010 Budde and Roth 2010) and highHIF-1 activity has been shown to create resistance to HCN-mediated fast paralytic killing by P aeruginosa (Darby et al1999 Gallagher and Manoil 2001 Shao et al 2010) There-fore cysl-2 may be induced not to effect H2S levels but in-stead may be the mediator of resistance to HCN-dependentfast paralytic killing To test this hypothesis we exposed cysl-1 cysl-2 and cysl-4 mutant animals to RAHCN and foundthat both cysl-1 and cysl-2 mutant animals are sensitive toRAHCN (Table 2) The specificity of cysl-2 mutant sensitivityto cyanide suggests that this enzyme is directly acting to di-minish HCN levels rather than to affect H2S levels The sen-sitivity of cysl-1 mutants to both RAH2S and RAHCNsuggest that cysl-1 acts downstream of cysl-2 and cysl-1mutants are being overwhelmed by the H2S produced bycysl-2 during HCN detoxification (Figure 8)
Next we tested whether these enzymes are responsiblefor HIF-1ndashmediated innate immunity to P aeruginosa Weshowed that similar to previous studies (Gallagher andManoil 2001 Shao et al 2010) egl-9ndashdeficient animals haveincreased innate immunity to P aeruginosa (Table 3) Nextwe demonstrated that cysl-1 and cysl-2 are both requiredfor the observed innate immunity to P aeruginosa demon-strating a mechanism of innate immunity to Pseudomonas-dependent fast paralytic killing Since we previously showedthat animals grown in RAH2S have increased cysl-2 mRNAexpression we assayed whether growth in H2S confers
Figure 4 Complementation datashowing that each genetic locusconsists of only one complemen-tation group (blue green yellowand pink shades) suls-1 is notshown because it is dominantsqrd-1 cysl-1 double heterozy-gotes are slightly sensitive toRAH2S
526 M W Budde and M B Roth
resistance to P aeruginosa Indeed our results show thatgrowth in H2S increases the innate immunity of C elegansto P aeruginosa (Table 3)
sqrd-1 is required for survival in H2S
The group of alleles on chromosome IV mr21 mr24 mr28mr30 and mr32 were mapped near the SNP pkP4095 lo-cated at the genetic location IV12 Analysis of the regionrevealed a gene F02H65 encoding an apparent homolog ofthe protein sulfidequinone reductase dehydrogenase (SQRD)which we have named sqrd-1 to reflect this homology SQRDwas initially described in prokaryotes as oxidizing sulfideand transferring the electrons to ubiquinone for generationof ATP during oxidative phosphorylation (Vande Weghe andOw 1999) The available deletion allele of sqrd-1 (tm3378)phenocopies and fails to complement these mutants Se-
quence analysis revealed A-to-G transitions in our isolatedmutants (Figure S1B)
The C elegans gene sqrd-1 encodes a member of thegroup II SQRD enzymes which also includes human mouseand Drosophila homologs The C elegans genome is predictedto encode another SQRD homolog Y9C9A16 (sqrd-2)which was initially classified as a pseudogene We obtainedthe deletion allele of sqrd-2 and found that it is not sensitiveto RAH2S
Transcriptional regulation of identified genes
We undertook this screen to determine the mechanism bywhich HIF-1 confers resistance to H2S Therefore we wereinterested to learn whether either cysl-1 or sqrd-1 is tran-scriptionally regulated by HIF-1 in response to RAH2S ex-posure Figure 6A shows that sqrd-1 and cysl-2 mRNA levels
Figure 5 (A) Cysteine synthase (CYS) catalyzes the formation of cysteine and acetate from OAS and H2S (B) Cyanoalanine synthase (CAS) catalyzes theformation of b-cyanoalanine and H2S from cysteine and HCN (C) Amino acid alignment of a selected region of CYS and CBS homologs C elegans Cyslgenes are homologous to CYS whereas C elegans Cbs genes are homologous to cystathionine b-synthase (CBS)
Table 2 Genes named in this article
Gene name Gene ID Allele tested 50 ppm RAH2S survival () 5 ppm RAHCN survival ()
cysl-1 C17G17 ok762 0 6 0 0 6 0cysl-2 K10H102 ok3516 100 6 0 0 6 0cysl-3 R08E52 mdash mdash mdash
cysl-4 F59A79 ok3359 92 6 11 100 6 0cbs-1 ZC3731 mdash mdash mdash
cbs-2 F54A34 ok666 100 6 0 100 6 0sqrd-1 F02H65 tm3378 0 6 0 97 6 5ethe-1 C33A127 tm4101 0 6 0 100 6 0
P 005 as compared to N2 wild-type animals
The Response of C elegans to H2S and HCN 527
increase during RAH2S and RAHCN exposure but cysl-1mRNA levels remain unchanged Furthermore hif-1(ia04)mutant animals fail to increase sqrd-1 and cysl-2 mRNA lev-els during RAH2S exposure (Figure 6B) suggesting thatupregulation of sqrd-1 by HIF-1 is a mechanism for survivingin the presence of H2S
To determine whether or not SQRD-1 protein levels in-crease during RAH2S exposure we performed a Westernblot to determine relative protein concentrations Figure 6Cshows that SQRD-1 protein levels increase during RAH2Sexposure in the parental N2 Bristol strain No SQRD-1 pro-tein accumulation is observed in the sqrd-1(tm3378) or inthe hif-1(ia04) mutant animals egl-9(sa307) mutant ani-mals have constitutively high levels of SQRD-1 proteinwhich may explain their ability to survive in 100 timeshigher RAH2S concentration than hif-1(ia04) mutantanimals
SQRD-1GFP protein is expressed during H2S exposure
Next we analyzed the expression pattern of SQRD-1 in Celegans We created a SQRD-1GFP fusion construct becausethe antibody we created was unable to stain worms by im-munofluorescence The presence of a predicted SL2 alterna-tive splice site upstream of sqrd-1 indicated that a standardpromoter fusion might not accurately recapitulate the en-dogenous expression pattern Therefore we used the recom-
bineering approach for GFP fusion construction whichincludes a 35-kb region surrounding the 45-kb SQRD-1gene RAH2S exposure causes a fluorescence increase inrecombinant animals that is most apparent in the anteriorof the worm (Figure 7) Expression appears limited to thehypodermis the body wall muscle and the anterior bulb ofthe pharynx (Figure 7C) A cross section through the wormconfirms that expression is limited to the most exposed partsof the worm (Figure 7D) It is possible that this SQRD-1expression pattern reflects a function of SQRD-1 to preventH2S from reaching the interior tissue
We conclude from these experiments that HIF-1 acts toinitiate the response of C elegans to H2S by upregulating theSQRD-1 protein levels SQRD-1 is an oxidoreductase oxidiz-ing H2S to polysulfide and reducing ubiquinone (Theissenet al 2003) The oxidation of sulfide generates polysulfide(Marcia et al 2009) which can then be further oxidized viathe H2S oxidation pathway to yield sulfate and thiosulfate(Kabil and Banerjee 2010) The decrease in H2S caused bySQRD-1 is likely to reduce internal H2S levels to a toleratedrange
A second enzyme required for the H2S oxidation pathwayis ETHE1 (Tiranti et al 2009) ETHE1 is a sulfur dioxyge-nase required in mammals for oxidation of sulfur to sulfate(Kabil and Banerjee 2010) C elegans has a single homologof Ethe1 C33A127 which we named ethe-1 We obtainedthe available ethe-1 (tm4101) deletion allele and tested forH2S sensitivity (Table 2) Similar to sqrd-1 (tm3378) ethe-1(tm4101) is sensitive to RAH2S which suggests that sqrd-1confers resistance to H2S through the sulfurndashoxidationpathway
Epistasis with egl-9
Egl-9ndashdeficient animals have been shown to be resistant toboth H2S and HCN and our data suggest that resistance ismediated by HIF-1 upregulating sqrd-1 and cysl-2 respec-tively To test this hypothesis we used RNAi to knockdownegl-9 in mutant animals which we have described as H2Sand HCN sensitive and tested them for resistance to H2S andHCN Table 4 shows that egl-9 knockdown does not mediateresistance to HCN in cysl-1 and cysl-2 mutant backgrounds
Table 3 Innate immunity to P aeroginosa requires hif-1 cysl-1and cysl-2
Strain Pretreatment AlleleSurvival on
P aeruginosa ()
N2 Control RNAi mdash 0 6 0N2 Egl-9 RNAi mdash 89 6 19hif-1 Egl-9 RNAi ia04 10 6 16egl-9 Egl-9 RNAi sa307 100 6 0cysl-1 Egl-9 RNAi ok762 4 6 7cysl-2 Egl-9 RNAi ok3516 0 6 0sqrd-1 Egl-9 RNAi tm3378 100 6 0ethe-1 Egl-9 RNAi tm4101 100 6 0N2 H2S grown mdash 83 6 29y
P 005 as compared to egl-9 knockdown in N2 animals yP 005 as comparedto untreated control RNAi N2 animals
Figure 6 (A) Both sqrd-1and cysl-2 mRNA levels increaserelative to untreated animals as measured by quantitativereverse transciptase PCR Y axis is relative mRNA concen-tration compared to room air-treated control animalsP 005 as compared to animals grown in room air(B) hif-1 is required for sqrd-1 and cysl-2 mRNA inductionwhen exposed to RAH2S or RAHCN Arrowheads indicatethat mRNA levels were observed at very low levels P 005 when comparing wild-type and hif-1mutant animals(C) Western blot shows SQRD-1 protein induction in RAH2S This induction requires hif-1 No immunoreactivity isobserved in sqrd-1 mutant animals SQRD-1 protein isabundant in egl-9 mutant animals Circles indicate proteinsize markers of 116 82 62 and 49 kDa from top tobottom The calculated molecular weight of 53 kDa cor-responds well with the observed relative migration
528 M W Budde and M B Roth
and therefore cysl-1 and cysl-2 are epistatic to egl-9 Sqrd-1and ethe-1 mutant animals with knocked down egl-9 arepartially resistant to H2S
Discussion
C elegans likely encounters H2S in the soil as a result ofbreakdown of organic sulfur (Morra and Dick 1991) andHCN produced by both bacteria (Gallagher and Manoil2001) and plants (Garciacutea et al 2010) This study has exam-ined the mechanisms that C elegans uses to respond to highlevels of H2S and HCN RAH2S and RAHCN both cause an
increase in HIF-1 activity which increases the expression ofsqrd-1 and cysl-2 to catalyze the first steps of the metabolismof H2S and HCN respectively sqrd-1 acts to oxidize H2Sultimately preventing internal H2S levels from exceedingthe maximum tolerated concentration cysl-2 acts in parallelto keep HCN levels low however equimolar amounts of H2Sare generated as a consequence of cysl-2 activity cysl-1 isrequired to reduce H2S levels that are produced by cysl-2during RAHCN exposure (Figure 8)
Here we provide direct evidence that H2S acts throughhif-1 to increase both SQRD-1 mRNA and protein expressionThis initiates the sulfidendashoxidation pathway which lowers in-ternal H2S levels (Figure 8) SQRD-1 expression appears to belocalized to the cells most readily exposed to exogenous H2S inthe worm In mammals colon epithelial cells that express highlevels of SQRDL (Ponteacuten et al 2008) are likely to do so be-cause a prominent enteric bacteria Desulfovibrio piger produ-ces high levels of H2S in the lower intestine (Loubinoux et al2002) The requirement for ethe-1 during RAH2S exposureconfirms that the sulfidendashoxidation pathway is the essentialfunction of sqrd-1 The sulfidendashoxidation pathway requires ox-ygen both to maintain a functional electron transport chain (togenerate oxidized ubiquinone) and to directly oxidize sulfurvia ethe-1 Exposure of mammals to exogenous H2S results inmany alterations in physiology including the induction of a sus-pended-animationndashlike state and improved outcome in ische-mia reperfusion injury It will be interesting to learn whetherSQRDL expression is induced upon exposure to H2S inmammals
The question remains as to why SQRD-1 is inducible atall Surrounded by decaying matter C elegans must encoun-ter areas of high H2S in its natural environment Havingconstitutively high SQRD-1 levels as in egl-9 mutant ani-mals allows survival when wild-type animals with lowerHIF-1 activity would perish However low levels of H2Sare important for biological function so perhaps nematodesare using SQRD-1 to deliberately titrate the cellular concen-tration of H2S by controlling its rate of destruction In mam-mals the H2S-generating enzymes CBS CSE and 3MPcontrol sulfide levels (Kabil and Banerjee 2010) Howeverour study suggests that mammals may also modulate cellu-lar H2S levels by regulating SQRDL-dependent destruction
In addition to furthering our understanding of the sulfideoxidation pathway this study also provides insight into two
Figure 7 SQRD-1GFP expression is induced by exposure to RAH2S (A)SQRD-1GFP is not expressed in room air (B) RAH2S at 50 ppm inducesexpression of SQRD-1GFP (C) Confocal microscopy at middot40 magnifica-tion shows that the expression is brightest in the muscle dense bodies(tailed arrows) and the hypodermis (tail-less arrow) (D) Cross-section(Z-stack) shows expression is most intense in the hypodermis
Table 4 Epistasis relationships with egl-9
15 ppm RAHCN 250 ppm RAH2S
Empty vector () egl-9 RNAi () Empty vector () egl-9 RNAi ()
N2 0 6 0 92 6 10 0 6 0 98 6 4hif-1 (ia04) 0 6 0 0 6 0 0 6 0 0 6 0egl-9 (sa307) 100 6 0 100 6 0 100 6 0 100 6 0sqrd-1 (tm3378) 0 6 0 87 6 16 0 6 0 47 6 15ethe-1 (tm4101) 0 6 0 89 6 14 0 6 0 73 6 10cysl-1 (ok762) 0 6 0 27 6 35 0 6 0 68 6 30cysl-2 (ok3516) 0 6 0 0 6 0 0 6 0 94 6 6
P 005 as compared to egl-9 RNAi knockdown in wild-type animals Data are presented as percentage of survival
The Response of C elegans to H2S and HCN 529
new metabolic activities previously unknown in animalscysteine synthase and cyanoalanine synthase cysl-1 mutantanimals are sensitive to both RAH2S and RAHCN whilecysl-2 mutant animals are only sensitive to RAHCN Thissuggests that cysl-1 is a cysteine synthase acting downstreamof cysl-2 a cyanoalanine synthase (Figure 8) We hypothe-size that cysl-1 mutant animals could be succumbing to H2Sproduced by CYSL-2 as a result of cyanide assimilation Inthis way each molecule of HCN assimilated by CYSL-2 wouldproduce one molecule of H2S In this reaction mechanismCYSL-1 might regenerate cysteine used in cyanide assimila-tion Alternatively it is possible that CYSL-1 is catalyzing theassimilation of both H2S and HCN a hypothesis supportedby in vitro plant studies (Hell et al 2002) The function ofthe two remaining cysteine synthase homologs cysl-3 andcysl-4 remains to be elucidated but they may also haveimportant CYSCAS activity during development or in spe-cific tissues
To our knowledge cysl-1 is the first reported functionalCYS homolog in animals Indeed while no orthologs appearto be present in animals outside of the genus Caenorhabditisin plants and bacteria CYS uses OAS as the H2S-acceptingsubstrate (Feldman-Salit et al 2009) Plants and bacteriagenerate OAS by acetylating serine with the enzyme serineacetyltransferase (SAT) (Wirtz et al 2010) which has nohomolog in C elegans How might C elegans be obtainingOAS As with many other amino acids C elegans is likelyobtaining OAS from its bacterial food source AlternativelyC elegans might have evolved a mechanism of generatingOAS using alternative enzymes CYS activity is essential forcysteine biosynthesis in prototrophs (Feldman-Salit et al2009) Because the critical aspect of CYS appears to be theelimination of H2S instead of the generation of cysteine
perhaps C elegans is freed from using OAS as the H2S ac-ceptor and is using an alternative H2S acceptor that does notgenerate cysteine In mammals the closest mammalian ho-molog to CYS is CBS Consistent with the idea that mammalshave CYS activity there is evidence that radiolabeled H2Scan be incorporated into cysteine in germ-free rats (Huovinenand Gustafsson 1967)
The protective effect of cysl-2 to HCN explains the pre-vious observation that egl-9 mutant animals are resistant tothe pathogenic Pseudomonas fast paralytic killing P aerugi-nosa is a clinically relevant pathogen and can produce toxiccyanide levels in the sputum of cystic fibrosis patients (Andersonet al 2010) P fluorescens is a cyanide-producing soil bacte-ria that has also been shown to exhibit cyanide-dependentfast paralytic killing effect on C elegans (Romanowski et al2011) HIF-1 activates resistance to hypoxia and this hasbeen proposed to also confer resistance to the cytochromec oxidase inhibiting effects of cyanide (Gallagher andManoil 2001 Shao et al 2010 Romanowski et al 2011)While this certainly may play some role we provide evidencehere that the resistance of egl-9 mutant animals to P aerugi-nosa requires cysl-2 likely by directly detoxifying cyanide tob-cyanoalanine (Figure 8) This reaction produces H2S whichcan be further detoxified by cysl-1 and sqrd-1
While we have demonstrated critical components of theRAH2S-induced hif-1 response the mechanism of hif-1 in-duction remains unclear The overlapping gene expressionpatterns between both H2S and HCN suggests that C eleganshas a shared response to these two molecules In fact ourstudies suggest that C elegans is unable to discriminate be-tween H2S and HCN leading them to activate the degrada-tion pathways of both molecules when either is presentedThus C elegans treated with RAH2S or RAHCN increases
Figure 8 Proposed mechanismBoth H2S and HCN can inhibitelectron transport Electron trans-port inhibition causes inductionof HIF-1 activity resulting in high ex-pression of SQRD-1 and CYSL-2SQRD-1 catalyzes the first step inthe H2S-oxidation pathway ulti-mately resulting in production ofsulfate and thiosulfate CYSL-2catalyzes the first step in the HCNassimilation pathway producingH2S The resultant H2S is detoxifiedby CYSL-1
530 M W Budde and M B Roth
both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841

thermotolerant when grown in H2S (Miller and Roth2007) We showed that H2S induces the accumulation ofthe hypoxia-inducible factor 1 (HIF-1) (Budde and Roth2010) In turn hif-1 is required for survival in the presenceof H2S and keeping HIF-1 levels constitutively high confersresistance to otherwise lethal H2S concentrations HIF-1 isa transcription factor and thus the observed protectiveeffects are likely to be mediated by increased expressionof genes regulated by HIF-1 To determine the molecularmechanism by which tolerance to H2S is conferred weperformed a forward genetic screen for C elegans strainssensitive to H2S and report on two hif-1 target genes thatfunction in the first steps of two distinct pathways impor-tant for survival in H2S
Materials and Methods
Strains
C elegans were grown at room temperature on nematodegrowth medium plates seeded with live Escherichia coliOP50 food (Brenner 1974) The following mutant strainswere obtained from the Caenorhabditis Genetics Centercysl-1(ok762) cysl-2(ok3516) cysl-4(ok3359) cbs-2(ok666)hif-1 (ia04) sqrd-2 (ok3440) and CB4856 The followingstrains were provided by Shohei Mitani of the National Bio-Resource Project sqrd-1(tm3378) and ethe-1(tm4104) Pseu-domonas aeruginosa strain PA01 was obtained from ColinManoil (University of Washington Seattle WA)
Atmospheric chambers and viability tests
H2S-containing atmospheric chambers were generated bymixing air with small amounts of defined compressed H2Sbalanced with N2 (Budde and Roth 2010) The resulting gasmixture is essentially room air containing hydrogen sulfide(RAH2S) Unless otherwise stated RAH2S is room air con-taining 50 ppm of H2S All survival experiments were per-formed in 1000-ml chambers with a gas flow of 1000 mlminHCN-containing atmospheres were created in the same wayas the H2S atmospheres except HCN balanced with N2 wasused and they are referred to as RAHCN (essentially roomair containing hydrogen cyanide) Unless otherwise statedRAHCN is room air containing 5 ppm of HCN
Viability tests were performed by picking L4 larval stageworms onto OP50-seeded NGM plates and placing them intothe gaseous atmosphere for 24 hr (Budde and Roth 2010)For RAH2S exposures all animals observed after 24 hrwere either healthy adults or appeared to have perishedshortly after initiation of the experiment For RAHCN expo-sures developmentally arrested L4 animals with very slowmovement were occasionally observed If maintained inRAHCN these animals invariably perished after severaldays as L4 If removed from RAHCN after 24 hr some ofthese animals eventually recovered to become fertile adultsFor the data in this article animals with this developmentalarrest were scored as not surviving the RAHCN exposureAll error values presented are standard deviation (SD)
Video acquisition and automated worm speed analysis
A Quickcam Communicate STX (Logitech) webcamwas affixedto the eyepiece of a stereomicroscope Images were acquiredevery 12 sec and analyzed with the Multi-Worm Tracker(MWT) (Swierczek et al 2011) The video was analyzed threetimes on the MWT and the speed was averaged every 25 min
Genetic screen for H2S-sensitive mutants
Ethyl methanesulfonate (EMS) mutagenesis was performedas previously described (Brenner 1974) and was optimizedto maximize the number of unique mutations observed(Shaham 2007) N2 animals at the L4 or young-adult stagewere incubated with EMS for 4 hr The worms were thenwashed once aliquoted onto four OP50-seeded NGM platesand allowed to lay eggs overnight The next day the adultswere washed off the plates and the F1 animals were grownto adults Each pool of 25000 F1 adults was bleached togenerate synchronized F2 animals A total of 100000 F2L4 animals were screened (25000 from each pool)
The screen for nematodes that are sensitive to H2S wasperformed as follows When the F2 worms entered the L4larval stage the plate of worms was exposed to RAH2S for4 hr The worms were moved to room air for 5 min and thenall paralyzed worms were picked from the plate within15 min of removal from RAH2S From this screen 791 F2animals were singled out to generate 544 viable strains Ofthese viable strains 124 strains exhibited a lethality pheno-type during a 24-hr exposure to RAH2S Of the 124 sensi-tive strains 23 were 100 lethal when exposed to RAH2Sfor 24 hr as L4 and were annotated as strongly penetrant(Figure 2)
Mapping mutations to chromosomes andcomplementation tests
Prior to complementation testing strongly penetrant strainswere crossed to the Hawaiian strain CB4856 and SNP enrich-ment analysis was performed (Davis et al 2005) MaleCB4856 worms were mated into the 23 mutant strains F1animals were isolated and synchronous F2 populations werecreated by egg lays When F2 animals reached L4 larval stagethe plates were exposed to RAH2S for 4 hr Thirty paralyzedanimals were picked as phenotypic and 30 motile animalswere picked as aphenotypic In most cases 25 of F2worms became paralyzed when exposed to RAH2S and wereannotated as recessive In four cases75 of worms becameparalyzed and these mutants were annotated as dominant
After the restriction digests were separated by gel electro-phoreses the gel was visually examined for genomic regionswhere the DNA from phenotypic and aphenotypic animals didnot cut equally well with DraI Ethidium bromide fluores-cence was quantified with Quantity One 1-D analysis soft-ware (Bio-Rad) and a fluorescence ratio of cutuncut DNAwas generated for each SNP on the chromosome of interestThe SNPs on two neighboring chromosomes are presented asa background control Strains that appeared to show similar
522 M W Budde and M B Roth
enrichment patterns were grouped together as suspectedcomplementation groups (Figure 3)
Complementation tests were carried out by crossing homo-zygous males of one strain into another strain and testing theF1 progeny for survival in RAH2S Sequencing of cysl-1 andsqrd-1 was accomplished by PCR amplification of genomicDNA and then using nested primers for the labeling reactionAfter the point mutations were identified SIFT-BLink (Ngand Henikoff 2001) (httpsiftbiia-staredusgwwwSIFT_BLink_submithtml) was used to determine whether the muta-tions are predicted to be tolerated or not tolerated In parallelall splice mutations were annotated as not tolerated
Protein homology
Proteinndashprotein Basic Local and Alignment and Search Tool(BLASTp) was used to locate homologous genes Potentialhomologs were reciprocally BLAST searched to ensure speci-ficity Amino acid alignment was performed with ClustalX2using the following National Center for Biotechnology Infor-mation reference sequence IDs Arabidopsis thaliana CASNP_1917031 A thaliana CYS NP_1932241 Solanum tuber-osum CAS BAB187601 S tuberosum CYS AAC256351 Zeamays CAS ADG602361 Z mays CYS NP_0011054691Mus musculus CBS NP_6591041 Homo sapiens CBSNP_0000621 Ciona intestinalis CBS XP_0021202471 andDrosophila melanogaster CBS NP_6084241
Psuedomonas aeruginosa dependent paralysis
Pseudomonas experiments were performed with strain PA01as previously described (Shao et al 2010) Briefly a fulllawn of P aeruginosa was grown on brain heart infusionagar (37 g Difco BHI 20 g agar per liter) on 35-cm platesfor 24 hr at 37 The plates were cooled to room tempera-ture and 10 L4 animals were placed onto the bacterial lawnand the plate was sealed with parafilm The number of ani-mals moving and not moving after 6 hr was recorded
mRNA detection by QRTndashPCR
Quantitative reverse transcriptase PCR was performed ona Bio-Rad C1000 Thermal Cycler with a CFX96 real-timesystem with iQ SYBR green supermix (Bio-Rad) as pre-viously described (Budde and Roth 2010) ExperimentalmRNA was compared to the average mRNA level of thehousekeeping genes tba-1 and tbb-2 The primer sequencesare listed in Supporting Information Table S1
Western blotting
Antibody was generated and affinity purified using theamino acids 419ndash431 of SQRD-1 (wormbaseorg WPCE17629) amino acids AMETTPFDQSKPTY by ProSci In-corporated (San Diego CA) Western blotting was per-formed as previously described (Budde and Roth 2010)
Figure 1 Wild-type nematodes exposedto 50 ppm RAH2S exhibit a brief in-crease in movement initially and thencontinue to move (A black line) Wheneither wild-type or hif-1 mutant animalsare exposed to 150 ppm RAH2S or50 ppm RAH2S respectively they stopmoving and when the RAH2S is re-moved they resume movement (blueand red lines A and B) Examples ofHif-1 moving stopped and reanimatedworms are shown in C
The Response of C elegans to H2S and HCN 523
with the following changes primary antibody was diluted1500 Goat antirabbit IgG (H+L)-horseradish peroxidase(catalog no 111-035-003 Jackson ImmunoReseach Labora-tories West Grove PA) was used as the secondary antibody
Fosmid recombineering
GFP was inserted into fosmid WRM066CH11 as previouslydescribed (Dolphin and Hope 2006) The oligonucleotidesused to insert the RT cassette were
RT cassette forward -gtttgattaaaggatactggaatggaccagctacactcagaaattgtacaTCGCTGTCGAGATATGACGGTG
RT cassette reverse -acaaataaaaacgacaaagtgggaaatatctatttagactttaccaatcgGATGATAAGCTGTCAAACATGAG
GFP forward -acaaataaaaacgacaaagtgggaaatatctatttagactttaccaatcgTTTGTATAGTTCATCCATGCCATGTG
GFP reverse -gtttgattaaaggatactggaatggaccagctacactcagaaattgtacaATGAGTAAAGGAGAAGAACTTTTC
Uppercase letters indicate primers for the PCR reaction
Generation of transgenic animals
Wild-type C elegans were injected with 50 ngml fosmidtogether with 100 ngml pRF4 which confers a dominantroller [rol-6(su1006)] phenotype (Mello et al 1991) Roll-ing F1 animals were picked onto single plates to generatetransgenic lines Animals were exposed to 50 ppm RAH2Sprior to confocal fluorescent microscopy as previously de-scribed (Budde and Roth 2010)
RNAi experiments
RNAi was performed as previously described (Frazier andRoth 2009) using WBRNAi00013759 to target egl-9 L4 ani-mals were moved onto the RNAi food and the progeny were
moved onto NGM-OP50 or BHI-PA01 plates and immedi-ately assayed
Results
We showed previously that hypoxia-inducible factor hif-1 isrequired for growth and survival in H2S (Budde and Roth2010) HIF-1 protein concentration and activity are in-creased when C elegans are moved into RAH2S and egl-9mutant animals which have constitutively high HIF-1 ex-pression and activity are resistant to H2S
During our investigations with hif-1 mutants we noticedthat although hif-1 mutant animals are unable to surviveprolonged exposure to RAH2S (24 hr) a brief exposure(4 hr) causes a reversible paralysis (Figure 1A) Roughly 2hr after entering an environment containing RAH2S hif-1mutant L4 animals enter a state of suspended animationcharacterized by a complete cessation of movement andfeeding (Figure 1B) and the animals lose their characteristicsinusoidal shape (Figure 1C) If removed to fresh air duringthis suspended state the animals resume moving after20 min and proceed to become fertile adults This pheno-type is also present in wild-type animals exposed to 150 ppmRAH2S (Figure 1 A and B File S1) We used this pheno-type as a basis for screening mutant C elegans that are sen-sitive to H2S in an attempt to find genes that conferresistance to H2S (Figure 2A)
About 200000 mutant haploid genomes were screenedfor H2S sensitivity Through screening we obtained 124
Figure 3 Enrichment of H2S-sensitive phenotype after crossing mutantlines with the Hawaiian strain CB4856 SNP mapping data show fiveapparent complementation groups For suls-1 and suls-2 the apparentlocation of mutation is indicated by an arrow The known locations ofsqrd-1 cysl-1 and hif-1 are also indicated by arrows Black indicates anN2HI ratio 5
Figure 2 H2S sensitivity screen method and results (A) Schematic ofinitial paralysis screen and lethality rescreen (B) Histogram of the numberof strains isolated for each penetrance bin The strains with zero survivorsafter sulfide exposure are highlighted in red These strains were subse-quently characterized
524 M W Budde and M B Roth
lines that were sensitive to H2S (Figure 2B) Of these lines23 were completely penetrant where penetrance is definedas the percentage of worms unable to survive a 24-hr RAH2S exposure These mutant strains were chosen for furthercharacterization
The 23 strongly penetrant mutant lines were crossed withthe Hawaiian CB4856 strain for mapping the sensitivity traitto chromosomal regions using enrichment of SNPs The SNPanalysis revealed that 22 mutants map to only five chromo-somal regions (Figure 3 Table 1) only mr37 showed noMendelian inheritance of the trait These mutants were thencrossed to confirm that each chromosomal region containeda single complementation group (Figure 4) One group onchromosome V (mr18 mr27 mr35 and mr38) exhibitsdominant genetics and thus could not be further tested forcomplementation We named this gene suls-1 (sulfide sen-sitive) and suls-1 (mr35) was mapped to the 4-cM regionbetween SNP pkP5097 and pkP5068 The two alleles onchromosome I were named suls-2 The genes mutated insuls-1 and suls-2 strains remain to be identified Two alleleson chromosome V (mr20 and mr22) mapped near the hif-1locus and failed to complement the hif-1(ia04) mutationwhich confirms that we successfully screened for our phe-notype of interest
The 23 strongest mutant lines were next tested forsurvival in room air containing lower H2S concentrationsin an effort to phenotypically discriminate between the com-
plementation groups Table 1 shows that complementationgroups have differences in H2S sensitivity with the groupmapping to the X chromosome being the most sensitive andsuls-2 being the least sensitive In contrast to the other com-plementation groups suls-1 alleles vary in sensitivity to H2S
A cysteine synthase homolog cysl-1 is required forsurvival in H2S
mr23 was used as a representative allele of the group ofalleles on the X chromosome (mr19 mr23 mr25 mr26mr29 mr33 mr34 mr39 and mr40) and was mapped toa 02-cM region between SNPs pas23398 and snp_F41E7[3] This region contains 70 predicted genes Of the deletionmutants available for that region ok762 phenocopies andfails to complement these H2S-sensitive mutations Theok762 allele deletes five of the six exons of the geneC17G17 Sequencing a portion of C17G17 of our isolatedstrains revealed adenine-to-guanine point mutations whichare characteristic of EMS mutagenesis (Figure S1) All iden-tified mutations were either missense or splice mutations
C17G17 is a previously uncharacterized gene encodinga member of the cystathionine b-synthase (CBS)-like sub-group which is made up of enzymes important in cysteineproduction CBS and cysteine synthase (CYS) (OAS-TL andCysK) (Feldman-Salit et al 2009) Animals generally haveCBS homologs and bacteria and plants tend to have CYShomologs (Hell et al 2008) CBS is critical for generating
Table 1 Alleles unable to survive in RAH2S
Alivetotal ( surviving)
Gene name Chromosome Allele 32 ppm RAH2S 15 ppm RAH2S 75 ppm RAH2S Dominance
N2 mdash mdash 1212 (100) 1212 (100) 1313 (100) mdash
cysl-1 X ok762 020 (0) 018 (0) 011 (0) mdash
cysl-1 X mr19 013 (0) 012 (0) 010 (0) Recessivecysl-1 X mr23 016 (0) 010 (0) 010 (0) Recessivecysl-1 X mr25 013 (0) 010 (0) 06 (0) Recessivecysl-1 X mr26 012 (0) 010 (0) 011 (0) Recessivecysl-1 X mr33 015 (0) 59 (56) 1010 (100) Recessivecysl-1 X mr34 010 (0) 011 (0) 110 (10) Recessivecysl-1 X mr29 011 (0) 010 (0) 113 (8) Recessivecysl-1 X mr39 010 (0) 010 (0) 1313 (100) Recessivecysl-1 X mr40 010 (0) 010 (0) 111 (9) Recessivesqrd-1 IV tm3378 510 (50) 1010 (100) 1010 (100) mdash
sqrd-1 IV mr24 1014 (71) 1111 (100) mdash Recessivesqrd-1 IV mr30 213 (15) 99 (100) mdash Recessivesqrd-1 IV mr32 1213 (92) 1010 (100) mdash Recessivesqrd-1 IV mr21 615 (40) 1010 (100) mdash Recessivesqrd-1 IV mr28 59 (56) 1010 (100) 1111 (100) Recessivehif-1 V ia04 015 (0) 011 (0) 1010 (100) mdash
hif-1 V mr20 014 (0) 010 (0) 1013 (77) Recessivehif-1 V mr22 015 (0) 111 (9) 810 (80) Recessivesuls-1 V mr18 612 (50) 1112 (92) 1010 (100) Dominantsuls-1 V mr27 29 (22) 1010 (100) mdash Dominantsuls-1 V mr35 05 (0) 010 (0) 010 (0) Dominantsuls-1 V mr38 09 (0) 88 (100) mdash Dominantsuls-2 I mr31 78 (88) 88 (100) mdash Recessivesuls-2 I mr36 89 (89) 1111 (100) mdash Recessivemdash mdash mr37 1112 (92) 1111 (100) mdash mdash
The Response of C elegans to H2S and HCN 525
cysteine in animals which make cysteine from dietary me-thionine (Finkelstein et al 1988) CBS is also one of the onlyH2S-generating enzymes in mammals (Kabil and Banerjee2010) In plants and bacteria which are cysteine proto-trophs CYS combines H2S with O-acetylserine (OAS) togenerate cysteine (Figure 5A) (Wirtz et al 2010) Humansand mice each have one CBS-like family member In con-trast C elegans has six C17G17 K10H102 R08E52F59A79 and F54A34 (Table 2)
C17G17 (cysl-1) K10H102 (cysl-2) R08E52 (cysl-3) andF59A79 (cysl-4) are more similar to CYS from plants andbacteria whereas ZC3731 (cbs-1) and F54A34 (cbs-2) aremore similar to CBS from animals (Figure 5C) A BLASTsearch reveals that the genus Caenorhabditis contains the onlyknown animal CYS homologs We have renamed these genescysl-1 (cysteine synthase-like) cysl-2 cysl-3 cysl-4 cbs-1 andcbs-2 to reflect this homology Depletion of H2S levels as it isused as a substrate by CYS may explain why cysl-1 mutantanimals are sensitive to H2S Nematodes appear to have ac-quired the ability to assimilate H2S and cysl-1 is likely usingthis reaction to drive down internal H2S levels
cysl-2 mediates immunity to fast paralytic killingby Pseudomonas
The existence of three additional Cysl homologs in C elegansraises the possibility that these other Cysl enzymes are alsoimportant in H2S detoxification This possibility is especiallyintriguing because we have shown a hif-1ndashdependent in-crease in cysl-2 transcript levels during RAH2S exposure(Budde and Roth 2010) We obtained the available knock-out strains of cysl-2 cysl-4 and cbs-2 and tested them forRAH2S sensitivity but none showed a RAH2S sensitivityphenotype (Table 2)
In plants CYS and cyanoalanine synthase (CAS) areclosely related and have overlapping catalytic activities(Figure 5 A and B) The CAS reaction combines cysteineand HCN to produce b-cyanoalanine and H2S lowering cel-lular HCN levels (Figure 5B) Furthermore cysl-2 mRNA waspreviously shown to be upregulated by HIF-1 (Shen et al2005 Shao et al 2010 Budde and Roth 2010) and highHIF-1 activity has been shown to create resistance to HCN-mediated fast paralytic killing by P aeruginosa (Darby et al1999 Gallagher and Manoil 2001 Shao et al 2010) There-fore cysl-2 may be induced not to effect H2S levels but in-stead may be the mediator of resistance to HCN-dependentfast paralytic killing To test this hypothesis we exposed cysl-1 cysl-2 and cysl-4 mutant animals to RAHCN and foundthat both cysl-1 and cysl-2 mutant animals are sensitive toRAHCN (Table 2) The specificity of cysl-2 mutant sensitivityto cyanide suggests that this enzyme is directly acting to di-minish HCN levels rather than to affect H2S levels The sen-sitivity of cysl-1 mutants to both RAH2S and RAHCNsuggest that cysl-1 acts downstream of cysl-2 and cysl-1mutants are being overwhelmed by the H2S produced bycysl-2 during HCN detoxification (Figure 8)
Next we tested whether these enzymes are responsiblefor HIF-1ndashmediated innate immunity to P aeruginosa Weshowed that similar to previous studies (Gallagher andManoil 2001 Shao et al 2010) egl-9ndashdeficient animals haveincreased innate immunity to P aeruginosa (Table 3) Nextwe demonstrated that cysl-1 and cysl-2 are both requiredfor the observed innate immunity to P aeruginosa demon-strating a mechanism of innate immunity to Pseudomonas-dependent fast paralytic killing Since we previously showedthat animals grown in RAH2S have increased cysl-2 mRNAexpression we assayed whether growth in H2S confers
Figure 4 Complementation datashowing that each genetic locusconsists of only one complemen-tation group (blue green yellowand pink shades) suls-1 is notshown because it is dominantsqrd-1 cysl-1 double heterozy-gotes are slightly sensitive toRAH2S
526 M W Budde and M B Roth
resistance to P aeruginosa Indeed our results show thatgrowth in H2S increases the innate immunity of C elegansto P aeruginosa (Table 3)
sqrd-1 is required for survival in H2S
The group of alleles on chromosome IV mr21 mr24 mr28mr30 and mr32 were mapped near the SNP pkP4095 lo-cated at the genetic location IV12 Analysis of the regionrevealed a gene F02H65 encoding an apparent homolog ofthe protein sulfidequinone reductase dehydrogenase (SQRD)which we have named sqrd-1 to reflect this homology SQRDwas initially described in prokaryotes as oxidizing sulfideand transferring the electrons to ubiquinone for generationof ATP during oxidative phosphorylation (Vande Weghe andOw 1999) The available deletion allele of sqrd-1 (tm3378)phenocopies and fails to complement these mutants Se-
quence analysis revealed A-to-G transitions in our isolatedmutants (Figure S1B)
The C elegans gene sqrd-1 encodes a member of thegroup II SQRD enzymes which also includes human mouseand Drosophila homologs The C elegans genome is predictedto encode another SQRD homolog Y9C9A16 (sqrd-2)which was initially classified as a pseudogene We obtainedthe deletion allele of sqrd-2 and found that it is not sensitiveto RAH2S
Transcriptional regulation of identified genes
We undertook this screen to determine the mechanism bywhich HIF-1 confers resistance to H2S Therefore we wereinterested to learn whether either cysl-1 or sqrd-1 is tran-scriptionally regulated by HIF-1 in response to RAH2S ex-posure Figure 6A shows that sqrd-1 and cysl-2 mRNA levels
Figure 5 (A) Cysteine synthase (CYS) catalyzes the formation of cysteine and acetate from OAS and H2S (B) Cyanoalanine synthase (CAS) catalyzes theformation of b-cyanoalanine and H2S from cysteine and HCN (C) Amino acid alignment of a selected region of CYS and CBS homologs C elegans Cyslgenes are homologous to CYS whereas C elegans Cbs genes are homologous to cystathionine b-synthase (CBS)
Table 2 Genes named in this article
Gene name Gene ID Allele tested 50 ppm RAH2S survival () 5 ppm RAHCN survival ()
cysl-1 C17G17 ok762 0 6 0 0 6 0cysl-2 K10H102 ok3516 100 6 0 0 6 0cysl-3 R08E52 mdash mdash mdash
cysl-4 F59A79 ok3359 92 6 11 100 6 0cbs-1 ZC3731 mdash mdash mdash
cbs-2 F54A34 ok666 100 6 0 100 6 0sqrd-1 F02H65 tm3378 0 6 0 97 6 5ethe-1 C33A127 tm4101 0 6 0 100 6 0
P 005 as compared to N2 wild-type animals
The Response of C elegans to H2S and HCN 527
increase during RAH2S and RAHCN exposure but cysl-1mRNA levels remain unchanged Furthermore hif-1(ia04)mutant animals fail to increase sqrd-1 and cysl-2 mRNA lev-els during RAH2S exposure (Figure 6B) suggesting thatupregulation of sqrd-1 by HIF-1 is a mechanism for survivingin the presence of H2S
To determine whether or not SQRD-1 protein levels in-crease during RAH2S exposure we performed a Westernblot to determine relative protein concentrations Figure 6Cshows that SQRD-1 protein levels increase during RAH2Sexposure in the parental N2 Bristol strain No SQRD-1 pro-tein accumulation is observed in the sqrd-1(tm3378) or inthe hif-1(ia04) mutant animals egl-9(sa307) mutant ani-mals have constitutively high levels of SQRD-1 proteinwhich may explain their ability to survive in 100 timeshigher RAH2S concentration than hif-1(ia04) mutantanimals
SQRD-1GFP protein is expressed during H2S exposure
Next we analyzed the expression pattern of SQRD-1 in Celegans We created a SQRD-1GFP fusion construct becausethe antibody we created was unable to stain worms by im-munofluorescence The presence of a predicted SL2 alterna-tive splice site upstream of sqrd-1 indicated that a standardpromoter fusion might not accurately recapitulate the en-dogenous expression pattern Therefore we used the recom-
bineering approach for GFP fusion construction whichincludes a 35-kb region surrounding the 45-kb SQRD-1gene RAH2S exposure causes a fluorescence increase inrecombinant animals that is most apparent in the anteriorof the worm (Figure 7) Expression appears limited to thehypodermis the body wall muscle and the anterior bulb ofthe pharynx (Figure 7C) A cross section through the wormconfirms that expression is limited to the most exposed partsof the worm (Figure 7D) It is possible that this SQRD-1expression pattern reflects a function of SQRD-1 to preventH2S from reaching the interior tissue
We conclude from these experiments that HIF-1 acts toinitiate the response of C elegans to H2S by upregulating theSQRD-1 protein levels SQRD-1 is an oxidoreductase oxidiz-ing H2S to polysulfide and reducing ubiquinone (Theissenet al 2003) The oxidation of sulfide generates polysulfide(Marcia et al 2009) which can then be further oxidized viathe H2S oxidation pathway to yield sulfate and thiosulfate(Kabil and Banerjee 2010) The decrease in H2S caused bySQRD-1 is likely to reduce internal H2S levels to a toleratedrange
A second enzyme required for the H2S oxidation pathwayis ETHE1 (Tiranti et al 2009) ETHE1 is a sulfur dioxyge-nase required in mammals for oxidation of sulfur to sulfate(Kabil and Banerjee 2010) C elegans has a single homologof Ethe1 C33A127 which we named ethe-1 We obtainedthe available ethe-1 (tm4101) deletion allele and tested forH2S sensitivity (Table 2) Similar to sqrd-1 (tm3378) ethe-1(tm4101) is sensitive to RAH2S which suggests that sqrd-1confers resistance to H2S through the sulfurndashoxidationpathway
Epistasis with egl-9
Egl-9ndashdeficient animals have been shown to be resistant toboth H2S and HCN and our data suggest that resistance ismediated by HIF-1 upregulating sqrd-1 and cysl-2 respec-tively To test this hypothesis we used RNAi to knockdownegl-9 in mutant animals which we have described as H2Sand HCN sensitive and tested them for resistance to H2S andHCN Table 4 shows that egl-9 knockdown does not mediateresistance to HCN in cysl-1 and cysl-2 mutant backgrounds
Table 3 Innate immunity to P aeroginosa requires hif-1 cysl-1and cysl-2
Strain Pretreatment AlleleSurvival on
P aeruginosa ()
N2 Control RNAi mdash 0 6 0N2 Egl-9 RNAi mdash 89 6 19hif-1 Egl-9 RNAi ia04 10 6 16egl-9 Egl-9 RNAi sa307 100 6 0cysl-1 Egl-9 RNAi ok762 4 6 7cysl-2 Egl-9 RNAi ok3516 0 6 0sqrd-1 Egl-9 RNAi tm3378 100 6 0ethe-1 Egl-9 RNAi tm4101 100 6 0N2 H2S grown mdash 83 6 29y
P 005 as compared to egl-9 knockdown in N2 animals yP 005 as comparedto untreated control RNAi N2 animals
Figure 6 (A) Both sqrd-1and cysl-2 mRNA levels increaserelative to untreated animals as measured by quantitativereverse transciptase PCR Y axis is relative mRNA concen-tration compared to room air-treated control animalsP 005 as compared to animals grown in room air(B) hif-1 is required for sqrd-1 and cysl-2 mRNA inductionwhen exposed to RAH2S or RAHCN Arrowheads indicatethat mRNA levels were observed at very low levels P 005 when comparing wild-type and hif-1mutant animals(C) Western blot shows SQRD-1 protein induction in RAH2S This induction requires hif-1 No immunoreactivity isobserved in sqrd-1 mutant animals SQRD-1 protein isabundant in egl-9 mutant animals Circles indicate proteinsize markers of 116 82 62 and 49 kDa from top tobottom The calculated molecular weight of 53 kDa cor-responds well with the observed relative migration
528 M W Budde and M B Roth
and therefore cysl-1 and cysl-2 are epistatic to egl-9 Sqrd-1and ethe-1 mutant animals with knocked down egl-9 arepartially resistant to H2S
Discussion
C elegans likely encounters H2S in the soil as a result ofbreakdown of organic sulfur (Morra and Dick 1991) andHCN produced by both bacteria (Gallagher and Manoil2001) and plants (Garciacutea et al 2010) This study has exam-ined the mechanisms that C elegans uses to respond to highlevels of H2S and HCN RAH2S and RAHCN both cause an
increase in HIF-1 activity which increases the expression ofsqrd-1 and cysl-2 to catalyze the first steps of the metabolismof H2S and HCN respectively sqrd-1 acts to oxidize H2Sultimately preventing internal H2S levels from exceedingthe maximum tolerated concentration cysl-2 acts in parallelto keep HCN levels low however equimolar amounts of H2Sare generated as a consequence of cysl-2 activity cysl-1 isrequired to reduce H2S levels that are produced by cysl-2during RAHCN exposure (Figure 8)
Here we provide direct evidence that H2S acts throughhif-1 to increase both SQRD-1 mRNA and protein expressionThis initiates the sulfidendashoxidation pathway which lowers in-ternal H2S levels (Figure 8) SQRD-1 expression appears to belocalized to the cells most readily exposed to exogenous H2S inthe worm In mammals colon epithelial cells that express highlevels of SQRDL (Ponteacuten et al 2008) are likely to do so be-cause a prominent enteric bacteria Desulfovibrio piger produ-ces high levels of H2S in the lower intestine (Loubinoux et al2002) The requirement for ethe-1 during RAH2S exposureconfirms that the sulfidendashoxidation pathway is the essentialfunction of sqrd-1 The sulfidendashoxidation pathway requires ox-ygen both to maintain a functional electron transport chain (togenerate oxidized ubiquinone) and to directly oxidize sulfurvia ethe-1 Exposure of mammals to exogenous H2S results inmany alterations in physiology including the induction of a sus-pended-animationndashlike state and improved outcome in ische-mia reperfusion injury It will be interesting to learn whetherSQRDL expression is induced upon exposure to H2S inmammals
The question remains as to why SQRD-1 is inducible atall Surrounded by decaying matter C elegans must encoun-ter areas of high H2S in its natural environment Havingconstitutively high SQRD-1 levels as in egl-9 mutant ani-mals allows survival when wild-type animals with lowerHIF-1 activity would perish However low levels of H2Sare important for biological function so perhaps nematodesare using SQRD-1 to deliberately titrate the cellular concen-tration of H2S by controlling its rate of destruction In mam-mals the H2S-generating enzymes CBS CSE and 3MPcontrol sulfide levels (Kabil and Banerjee 2010) Howeverour study suggests that mammals may also modulate cellu-lar H2S levels by regulating SQRDL-dependent destruction
In addition to furthering our understanding of the sulfideoxidation pathway this study also provides insight into two
Figure 7 SQRD-1GFP expression is induced by exposure to RAH2S (A)SQRD-1GFP is not expressed in room air (B) RAH2S at 50 ppm inducesexpression of SQRD-1GFP (C) Confocal microscopy at middot40 magnifica-tion shows that the expression is brightest in the muscle dense bodies(tailed arrows) and the hypodermis (tail-less arrow) (D) Cross-section(Z-stack) shows expression is most intense in the hypodermis
Table 4 Epistasis relationships with egl-9
15 ppm RAHCN 250 ppm RAH2S
Empty vector () egl-9 RNAi () Empty vector () egl-9 RNAi ()
N2 0 6 0 92 6 10 0 6 0 98 6 4hif-1 (ia04) 0 6 0 0 6 0 0 6 0 0 6 0egl-9 (sa307) 100 6 0 100 6 0 100 6 0 100 6 0sqrd-1 (tm3378) 0 6 0 87 6 16 0 6 0 47 6 15ethe-1 (tm4101) 0 6 0 89 6 14 0 6 0 73 6 10cysl-1 (ok762) 0 6 0 27 6 35 0 6 0 68 6 30cysl-2 (ok3516) 0 6 0 0 6 0 0 6 0 94 6 6
P 005 as compared to egl-9 RNAi knockdown in wild-type animals Data are presented as percentage of survival
The Response of C elegans to H2S and HCN 529
new metabolic activities previously unknown in animalscysteine synthase and cyanoalanine synthase cysl-1 mutantanimals are sensitive to both RAH2S and RAHCN whilecysl-2 mutant animals are only sensitive to RAHCN Thissuggests that cysl-1 is a cysteine synthase acting downstreamof cysl-2 a cyanoalanine synthase (Figure 8) We hypothe-size that cysl-1 mutant animals could be succumbing to H2Sproduced by CYSL-2 as a result of cyanide assimilation Inthis way each molecule of HCN assimilated by CYSL-2 wouldproduce one molecule of H2S In this reaction mechanismCYSL-1 might regenerate cysteine used in cyanide assimila-tion Alternatively it is possible that CYSL-1 is catalyzing theassimilation of both H2S and HCN a hypothesis supportedby in vitro plant studies (Hell et al 2002) The function ofthe two remaining cysteine synthase homologs cysl-3 andcysl-4 remains to be elucidated but they may also haveimportant CYSCAS activity during development or in spe-cific tissues
To our knowledge cysl-1 is the first reported functionalCYS homolog in animals Indeed while no orthologs appearto be present in animals outside of the genus Caenorhabditisin plants and bacteria CYS uses OAS as the H2S-acceptingsubstrate (Feldman-Salit et al 2009) Plants and bacteriagenerate OAS by acetylating serine with the enzyme serineacetyltransferase (SAT) (Wirtz et al 2010) which has nohomolog in C elegans How might C elegans be obtainingOAS As with many other amino acids C elegans is likelyobtaining OAS from its bacterial food source AlternativelyC elegans might have evolved a mechanism of generatingOAS using alternative enzymes CYS activity is essential forcysteine biosynthesis in prototrophs (Feldman-Salit et al2009) Because the critical aspect of CYS appears to be theelimination of H2S instead of the generation of cysteine
perhaps C elegans is freed from using OAS as the H2S ac-ceptor and is using an alternative H2S acceptor that does notgenerate cysteine In mammals the closest mammalian ho-molog to CYS is CBS Consistent with the idea that mammalshave CYS activity there is evidence that radiolabeled H2Scan be incorporated into cysteine in germ-free rats (Huovinenand Gustafsson 1967)
The protective effect of cysl-2 to HCN explains the pre-vious observation that egl-9 mutant animals are resistant tothe pathogenic Pseudomonas fast paralytic killing P aerugi-nosa is a clinically relevant pathogen and can produce toxiccyanide levels in the sputum of cystic fibrosis patients (Andersonet al 2010) P fluorescens is a cyanide-producing soil bacte-ria that has also been shown to exhibit cyanide-dependentfast paralytic killing effect on C elegans (Romanowski et al2011) HIF-1 activates resistance to hypoxia and this hasbeen proposed to also confer resistance to the cytochromec oxidase inhibiting effects of cyanide (Gallagher andManoil 2001 Shao et al 2010 Romanowski et al 2011)While this certainly may play some role we provide evidencehere that the resistance of egl-9 mutant animals to P aerugi-nosa requires cysl-2 likely by directly detoxifying cyanide tob-cyanoalanine (Figure 8) This reaction produces H2S whichcan be further detoxified by cysl-1 and sqrd-1
While we have demonstrated critical components of theRAH2S-induced hif-1 response the mechanism of hif-1 in-duction remains unclear The overlapping gene expressionpatterns between both H2S and HCN suggests that C eleganshas a shared response to these two molecules In fact ourstudies suggest that C elegans is unable to discriminate be-tween H2S and HCN leading them to activate the degrada-tion pathways of both molecules when either is presentedThus C elegans treated with RAH2S or RAHCN increases
Figure 8 Proposed mechanismBoth H2S and HCN can inhibitelectron transport Electron trans-port inhibition causes inductionof HIF-1 activity resulting in high ex-pression of SQRD-1 and CYSL-2SQRD-1 catalyzes the first step inthe H2S-oxidation pathway ulti-mately resulting in production ofsulfate and thiosulfate CYSL-2catalyzes the first step in the HCNassimilation pathway producingH2S The resultant H2S is detoxifiedby CYSL-1
530 M W Budde and M B Roth
both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841
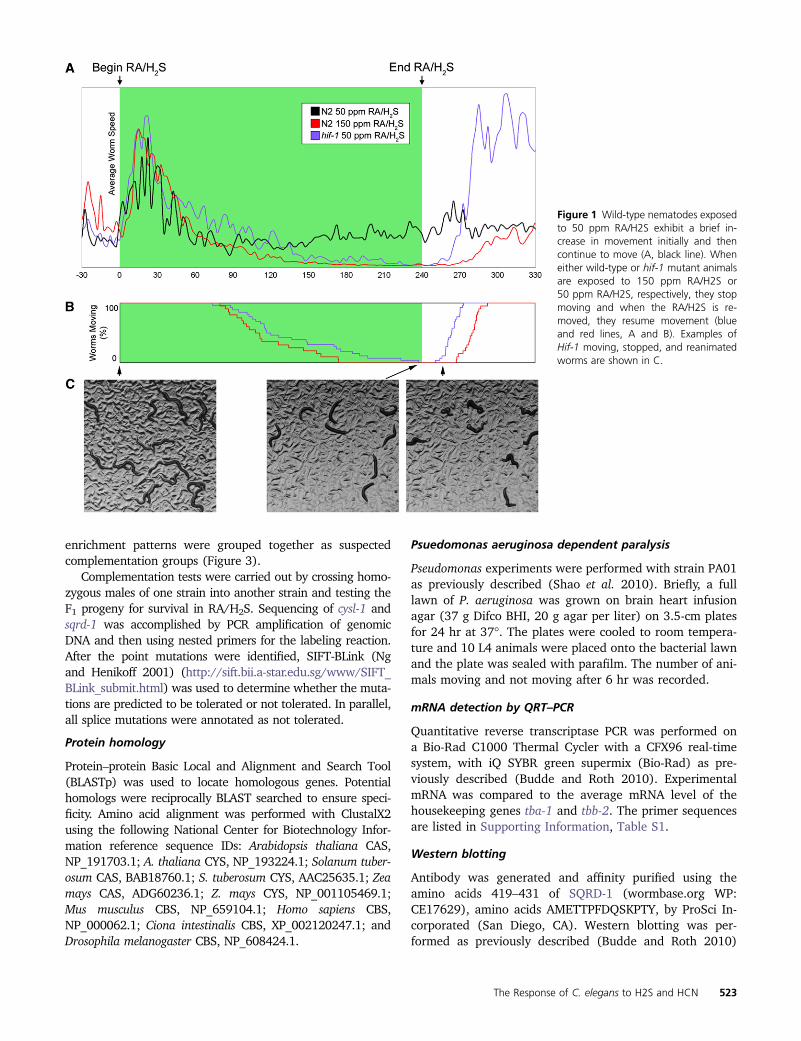
enrichment patterns were grouped together as suspectedcomplementation groups (Figure 3)
Complementation tests were carried out by crossing homo-zygous males of one strain into another strain and testing theF1 progeny for survival in RAH2S Sequencing of cysl-1 andsqrd-1 was accomplished by PCR amplification of genomicDNA and then using nested primers for the labeling reactionAfter the point mutations were identified SIFT-BLink (Ngand Henikoff 2001) (httpsiftbiia-staredusgwwwSIFT_BLink_submithtml) was used to determine whether the muta-tions are predicted to be tolerated or not tolerated In parallelall splice mutations were annotated as not tolerated
Protein homology
Proteinndashprotein Basic Local and Alignment and Search Tool(BLASTp) was used to locate homologous genes Potentialhomologs were reciprocally BLAST searched to ensure speci-ficity Amino acid alignment was performed with ClustalX2using the following National Center for Biotechnology Infor-mation reference sequence IDs Arabidopsis thaliana CASNP_1917031 A thaliana CYS NP_1932241 Solanum tuber-osum CAS BAB187601 S tuberosum CYS AAC256351 Zeamays CAS ADG602361 Z mays CYS NP_0011054691Mus musculus CBS NP_6591041 Homo sapiens CBSNP_0000621 Ciona intestinalis CBS XP_0021202471 andDrosophila melanogaster CBS NP_6084241
Psuedomonas aeruginosa dependent paralysis
Pseudomonas experiments were performed with strain PA01as previously described (Shao et al 2010) Briefly a fulllawn of P aeruginosa was grown on brain heart infusionagar (37 g Difco BHI 20 g agar per liter) on 35-cm platesfor 24 hr at 37 The plates were cooled to room tempera-ture and 10 L4 animals were placed onto the bacterial lawnand the plate was sealed with parafilm The number of ani-mals moving and not moving after 6 hr was recorded
mRNA detection by QRTndashPCR
Quantitative reverse transcriptase PCR was performed ona Bio-Rad C1000 Thermal Cycler with a CFX96 real-timesystem with iQ SYBR green supermix (Bio-Rad) as pre-viously described (Budde and Roth 2010) ExperimentalmRNA was compared to the average mRNA level of thehousekeeping genes tba-1 and tbb-2 The primer sequencesare listed in Supporting Information Table S1
Western blotting
Antibody was generated and affinity purified using theamino acids 419ndash431 of SQRD-1 (wormbaseorg WPCE17629) amino acids AMETTPFDQSKPTY by ProSci In-corporated (San Diego CA) Western blotting was per-formed as previously described (Budde and Roth 2010)
Figure 1 Wild-type nematodes exposedto 50 ppm RAH2S exhibit a brief in-crease in movement initially and thencontinue to move (A black line) Wheneither wild-type or hif-1 mutant animalsare exposed to 150 ppm RAH2S or50 ppm RAH2S respectively they stopmoving and when the RAH2S is re-moved they resume movement (blueand red lines A and B) Examples ofHif-1 moving stopped and reanimatedworms are shown in C
The Response of C elegans to H2S and HCN 523
with the following changes primary antibody was diluted1500 Goat antirabbit IgG (H+L)-horseradish peroxidase(catalog no 111-035-003 Jackson ImmunoReseach Labora-tories West Grove PA) was used as the secondary antibody
Fosmid recombineering
GFP was inserted into fosmid WRM066CH11 as previouslydescribed (Dolphin and Hope 2006) The oligonucleotidesused to insert the RT cassette were
RT cassette forward -gtttgattaaaggatactggaatggaccagctacactcagaaattgtacaTCGCTGTCGAGATATGACGGTG
RT cassette reverse -acaaataaaaacgacaaagtgggaaatatctatttagactttaccaatcgGATGATAAGCTGTCAAACATGAG
GFP forward -acaaataaaaacgacaaagtgggaaatatctatttagactttaccaatcgTTTGTATAGTTCATCCATGCCATGTG
GFP reverse -gtttgattaaaggatactggaatggaccagctacactcagaaattgtacaATGAGTAAAGGAGAAGAACTTTTC
Uppercase letters indicate primers for the PCR reaction
Generation of transgenic animals
Wild-type C elegans were injected with 50 ngml fosmidtogether with 100 ngml pRF4 which confers a dominantroller [rol-6(su1006)] phenotype (Mello et al 1991) Roll-ing F1 animals were picked onto single plates to generatetransgenic lines Animals were exposed to 50 ppm RAH2Sprior to confocal fluorescent microscopy as previously de-scribed (Budde and Roth 2010)
RNAi experiments
RNAi was performed as previously described (Frazier andRoth 2009) using WBRNAi00013759 to target egl-9 L4 ani-mals were moved onto the RNAi food and the progeny were
moved onto NGM-OP50 or BHI-PA01 plates and immedi-ately assayed
Results
We showed previously that hypoxia-inducible factor hif-1 isrequired for growth and survival in H2S (Budde and Roth2010) HIF-1 protein concentration and activity are in-creased when C elegans are moved into RAH2S and egl-9mutant animals which have constitutively high HIF-1 ex-pression and activity are resistant to H2S
During our investigations with hif-1 mutants we noticedthat although hif-1 mutant animals are unable to surviveprolonged exposure to RAH2S (24 hr) a brief exposure(4 hr) causes a reversible paralysis (Figure 1A) Roughly 2hr after entering an environment containing RAH2S hif-1mutant L4 animals enter a state of suspended animationcharacterized by a complete cessation of movement andfeeding (Figure 1B) and the animals lose their characteristicsinusoidal shape (Figure 1C) If removed to fresh air duringthis suspended state the animals resume moving after20 min and proceed to become fertile adults This pheno-type is also present in wild-type animals exposed to 150 ppmRAH2S (Figure 1 A and B File S1) We used this pheno-type as a basis for screening mutant C elegans that are sen-sitive to H2S in an attempt to find genes that conferresistance to H2S (Figure 2A)
About 200000 mutant haploid genomes were screenedfor H2S sensitivity Through screening we obtained 124
Figure 3 Enrichment of H2S-sensitive phenotype after crossing mutantlines with the Hawaiian strain CB4856 SNP mapping data show fiveapparent complementation groups For suls-1 and suls-2 the apparentlocation of mutation is indicated by an arrow The known locations ofsqrd-1 cysl-1 and hif-1 are also indicated by arrows Black indicates anN2HI ratio 5
Figure 2 H2S sensitivity screen method and results (A) Schematic ofinitial paralysis screen and lethality rescreen (B) Histogram of the numberof strains isolated for each penetrance bin The strains with zero survivorsafter sulfide exposure are highlighted in red These strains were subse-quently characterized
524 M W Budde and M B Roth
lines that were sensitive to H2S (Figure 2B) Of these lines23 were completely penetrant where penetrance is definedas the percentage of worms unable to survive a 24-hr RAH2S exposure These mutant strains were chosen for furthercharacterization
The 23 strongly penetrant mutant lines were crossed withthe Hawaiian CB4856 strain for mapping the sensitivity traitto chromosomal regions using enrichment of SNPs The SNPanalysis revealed that 22 mutants map to only five chromo-somal regions (Figure 3 Table 1) only mr37 showed noMendelian inheritance of the trait These mutants were thencrossed to confirm that each chromosomal region containeda single complementation group (Figure 4) One group onchromosome V (mr18 mr27 mr35 and mr38) exhibitsdominant genetics and thus could not be further tested forcomplementation We named this gene suls-1 (sulfide sen-sitive) and suls-1 (mr35) was mapped to the 4-cM regionbetween SNP pkP5097 and pkP5068 The two alleles onchromosome I were named suls-2 The genes mutated insuls-1 and suls-2 strains remain to be identified Two alleleson chromosome V (mr20 and mr22) mapped near the hif-1locus and failed to complement the hif-1(ia04) mutationwhich confirms that we successfully screened for our phe-notype of interest
The 23 strongest mutant lines were next tested forsurvival in room air containing lower H2S concentrationsin an effort to phenotypically discriminate between the com-
plementation groups Table 1 shows that complementationgroups have differences in H2S sensitivity with the groupmapping to the X chromosome being the most sensitive andsuls-2 being the least sensitive In contrast to the other com-plementation groups suls-1 alleles vary in sensitivity to H2S
A cysteine synthase homolog cysl-1 is required forsurvival in H2S
mr23 was used as a representative allele of the group ofalleles on the X chromosome (mr19 mr23 mr25 mr26mr29 mr33 mr34 mr39 and mr40) and was mapped toa 02-cM region between SNPs pas23398 and snp_F41E7[3] This region contains 70 predicted genes Of the deletionmutants available for that region ok762 phenocopies andfails to complement these H2S-sensitive mutations Theok762 allele deletes five of the six exons of the geneC17G17 Sequencing a portion of C17G17 of our isolatedstrains revealed adenine-to-guanine point mutations whichare characteristic of EMS mutagenesis (Figure S1) All iden-tified mutations were either missense or splice mutations
C17G17 is a previously uncharacterized gene encodinga member of the cystathionine b-synthase (CBS)-like sub-group which is made up of enzymes important in cysteineproduction CBS and cysteine synthase (CYS) (OAS-TL andCysK) (Feldman-Salit et al 2009) Animals generally haveCBS homologs and bacteria and plants tend to have CYShomologs (Hell et al 2008) CBS is critical for generating
Table 1 Alleles unable to survive in RAH2S
Alivetotal ( surviving)
Gene name Chromosome Allele 32 ppm RAH2S 15 ppm RAH2S 75 ppm RAH2S Dominance
N2 mdash mdash 1212 (100) 1212 (100) 1313 (100) mdash
cysl-1 X ok762 020 (0) 018 (0) 011 (0) mdash
cysl-1 X mr19 013 (0) 012 (0) 010 (0) Recessivecysl-1 X mr23 016 (0) 010 (0) 010 (0) Recessivecysl-1 X mr25 013 (0) 010 (0) 06 (0) Recessivecysl-1 X mr26 012 (0) 010 (0) 011 (0) Recessivecysl-1 X mr33 015 (0) 59 (56) 1010 (100) Recessivecysl-1 X mr34 010 (0) 011 (0) 110 (10) Recessivecysl-1 X mr29 011 (0) 010 (0) 113 (8) Recessivecysl-1 X mr39 010 (0) 010 (0) 1313 (100) Recessivecysl-1 X mr40 010 (0) 010 (0) 111 (9) Recessivesqrd-1 IV tm3378 510 (50) 1010 (100) 1010 (100) mdash
sqrd-1 IV mr24 1014 (71) 1111 (100) mdash Recessivesqrd-1 IV mr30 213 (15) 99 (100) mdash Recessivesqrd-1 IV mr32 1213 (92) 1010 (100) mdash Recessivesqrd-1 IV mr21 615 (40) 1010 (100) mdash Recessivesqrd-1 IV mr28 59 (56) 1010 (100) 1111 (100) Recessivehif-1 V ia04 015 (0) 011 (0) 1010 (100) mdash
hif-1 V mr20 014 (0) 010 (0) 1013 (77) Recessivehif-1 V mr22 015 (0) 111 (9) 810 (80) Recessivesuls-1 V mr18 612 (50) 1112 (92) 1010 (100) Dominantsuls-1 V mr27 29 (22) 1010 (100) mdash Dominantsuls-1 V mr35 05 (0) 010 (0) 010 (0) Dominantsuls-1 V mr38 09 (0) 88 (100) mdash Dominantsuls-2 I mr31 78 (88) 88 (100) mdash Recessivesuls-2 I mr36 89 (89) 1111 (100) mdash Recessivemdash mdash mr37 1112 (92) 1111 (100) mdash mdash
The Response of C elegans to H2S and HCN 525
cysteine in animals which make cysteine from dietary me-thionine (Finkelstein et al 1988) CBS is also one of the onlyH2S-generating enzymes in mammals (Kabil and Banerjee2010) In plants and bacteria which are cysteine proto-trophs CYS combines H2S with O-acetylserine (OAS) togenerate cysteine (Figure 5A) (Wirtz et al 2010) Humansand mice each have one CBS-like family member In con-trast C elegans has six C17G17 K10H102 R08E52F59A79 and F54A34 (Table 2)
C17G17 (cysl-1) K10H102 (cysl-2) R08E52 (cysl-3) andF59A79 (cysl-4) are more similar to CYS from plants andbacteria whereas ZC3731 (cbs-1) and F54A34 (cbs-2) aremore similar to CBS from animals (Figure 5C) A BLASTsearch reveals that the genus Caenorhabditis contains the onlyknown animal CYS homologs We have renamed these genescysl-1 (cysteine synthase-like) cysl-2 cysl-3 cysl-4 cbs-1 andcbs-2 to reflect this homology Depletion of H2S levels as it isused as a substrate by CYS may explain why cysl-1 mutantanimals are sensitive to H2S Nematodes appear to have ac-quired the ability to assimilate H2S and cysl-1 is likely usingthis reaction to drive down internal H2S levels
cysl-2 mediates immunity to fast paralytic killingby Pseudomonas
The existence of three additional Cysl homologs in C elegansraises the possibility that these other Cysl enzymes are alsoimportant in H2S detoxification This possibility is especiallyintriguing because we have shown a hif-1ndashdependent in-crease in cysl-2 transcript levels during RAH2S exposure(Budde and Roth 2010) We obtained the available knock-out strains of cysl-2 cysl-4 and cbs-2 and tested them forRAH2S sensitivity but none showed a RAH2S sensitivityphenotype (Table 2)
In plants CYS and cyanoalanine synthase (CAS) areclosely related and have overlapping catalytic activities(Figure 5 A and B) The CAS reaction combines cysteineand HCN to produce b-cyanoalanine and H2S lowering cel-lular HCN levels (Figure 5B) Furthermore cysl-2 mRNA waspreviously shown to be upregulated by HIF-1 (Shen et al2005 Shao et al 2010 Budde and Roth 2010) and highHIF-1 activity has been shown to create resistance to HCN-mediated fast paralytic killing by P aeruginosa (Darby et al1999 Gallagher and Manoil 2001 Shao et al 2010) There-fore cysl-2 may be induced not to effect H2S levels but in-stead may be the mediator of resistance to HCN-dependentfast paralytic killing To test this hypothesis we exposed cysl-1 cysl-2 and cysl-4 mutant animals to RAHCN and foundthat both cysl-1 and cysl-2 mutant animals are sensitive toRAHCN (Table 2) The specificity of cysl-2 mutant sensitivityto cyanide suggests that this enzyme is directly acting to di-minish HCN levels rather than to affect H2S levels The sen-sitivity of cysl-1 mutants to both RAH2S and RAHCNsuggest that cysl-1 acts downstream of cysl-2 and cysl-1mutants are being overwhelmed by the H2S produced bycysl-2 during HCN detoxification (Figure 8)
Next we tested whether these enzymes are responsiblefor HIF-1ndashmediated innate immunity to P aeruginosa Weshowed that similar to previous studies (Gallagher andManoil 2001 Shao et al 2010) egl-9ndashdeficient animals haveincreased innate immunity to P aeruginosa (Table 3) Nextwe demonstrated that cysl-1 and cysl-2 are both requiredfor the observed innate immunity to P aeruginosa demon-strating a mechanism of innate immunity to Pseudomonas-dependent fast paralytic killing Since we previously showedthat animals grown in RAH2S have increased cysl-2 mRNAexpression we assayed whether growth in H2S confers
Figure 4 Complementation datashowing that each genetic locusconsists of only one complemen-tation group (blue green yellowand pink shades) suls-1 is notshown because it is dominantsqrd-1 cysl-1 double heterozy-gotes are slightly sensitive toRAH2S
526 M W Budde and M B Roth
resistance to P aeruginosa Indeed our results show thatgrowth in H2S increases the innate immunity of C elegansto P aeruginosa (Table 3)
sqrd-1 is required for survival in H2S
The group of alleles on chromosome IV mr21 mr24 mr28mr30 and mr32 were mapped near the SNP pkP4095 lo-cated at the genetic location IV12 Analysis of the regionrevealed a gene F02H65 encoding an apparent homolog ofthe protein sulfidequinone reductase dehydrogenase (SQRD)which we have named sqrd-1 to reflect this homology SQRDwas initially described in prokaryotes as oxidizing sulfideand transferring the electrons to ubiquinone for generationof ATP during oxidative phosphorylation (Vande Weghe andOw 1999) The available deletion allele of sqrd-1 (tm3378)phenocopies and fails to complement these mutants Se-
quence analysis revealed A-to-G transitions in our isolatedmutants (Figure S1B)
The C elegans gene sqrd-1 encodes a member of thegroup II SQRD enzymes which also includes human mouseand Drosophila homologs The C elegans genome is predictedto encode another SQRD homolog Y9C9A16 (sqrd-2)which was initially classified as a pseudogene We obtainedthe deletion allele of sqrd-2 and found that it is not sensitiveto RAH2S
Transcriptional regulation of identified genes
We undertook this screen to determine the mechanism bywhich HIF-1 confers resistance to H2S Therefore we wereinterested to learn whether either cysl-1 or sqrd-1 is tran-scriptionally regulated by HIF-1 in response to RAH2S ex-posure Figure 6A shows that sqrd-1 and cysl-2 mRNA levels
Figure 5 (A) Cysteine synthase (CYS) catalyzes the formation of cysteine and acetate from OAS and H2S (B) Cyanoalanine synthase (CAS) catalyzes theformation of b-cyanoalanine and H2S from cysteine and HCN (C) Amino acid alignment of a selected region of CYS and CBS homologs C elegans Cyslgenes are homologous to CYS whereas C elegans Cbs genes are homologous to cystathionine b-synthase (CBS)
Table 2 Genes named in this article
Gene name Gene ID Allele tested 50 ppm RAH2S survival () 5 ppm RAHCN survival ()
cysl-1 C17G17 ok762 0 6 0 0 6 0cysl-2 K10H102 ok3516 100 6 0 0 6 0cysl-3 R08E52 mdash mdash mdash
cysl-4 F59A79 ok3359 92 6 11 100 6 0cbs-1 ZC3731 mdash mdash mdash
cbs-2 F54A34 ok666 100 6 0 100 6 0sqrd-1 F02H65 tm3378 0 6 0 97 6 5ethe-1 C33A127 tm4101 0 6 0 100 6 0
P 005 as compared to N2 wild-type animals
The Response of C elegans to H2S and HCN 527
increase during RAH2S and RAHCN exposure but cysl-1mRNA levels remain unchanged Furthermore hif-1(ia04)mutant animals fail to increase sqrd-1 and cysl-2 mRNA lev-els during RAH2S exposure (Figure 6B) suggesting thatupregulation of sqrd-1 by HIF-1 is a mechanism for survivingin the presence of H2S
To determine whether or not SQRD-1 protein levels in-crease during RAH2S exposure we performed a Westernblot to determine relative protein concentrations Figure 6Cshows that SQRD-1 protein levels increase during RAH2Sexposure in the parental N2 Bristol strain No SQRD-1 pro-tein accumulation is observed in the sqrd-1(tm3378) or inthe hif-1(ia04) mutant animals egl-9(sa307) mutant ani-mals have constitutively high levels of SQRD-1 proteinwhich may explain their ability to survive in 100 timeshigher RAH2S concentration than hif-1(ia04) mutantanimals
SQRD-1GFP protein is expressed during H2S exposure
Next we analyzed the expression pattern of SQRD-1 in Celegans We created a SQRD-1GFP fusion construct becausethe antibody we created was unable to stain worms by im-munofluorescence The presence of a predicted SL2 alterna-tive splice site upstream of sqrd-1 indicated that a standardpromoter fusion might not accurately recapitulate the en-dogenous expression pattern Therefore we used the recom-
bineering approach for GFP fusion construction whichincludes a 35-kb region surrounding the 45-kb SQRD-1gene RAH2S exposure causes a fluorescence increase inrecombinant animals that is most apparent in the anteriorof the worm (Figure 7) Expression appears limited to thehypodermis the body wall muscle and the anterior bulb ofthe pharynx (Figure 7C) A cross section through the wormconfirms that expression is limited to the most exposed partsof the worm (Figure 7D) It is possible that this SQRD-1expression pattern reflects a function of SQRD-1 to preventH2S from reaching the interior tissue
We conclude from these experiments that HIF-1 acts toinitiate the response of C elegans to H2S by upregulating theSQRD-1 protein levels SQRD-1 is an oxidoreductase oxidiz-ing H2S to polysulfide and reducing ubiquinone (Theissenet al 2003) The oxidation of sulfide generates polysulfide(Marcia et al 2009) which can then be further oxidized viathe H2S oxidation pathway to yield sulfate and thiosulfate(Kabil and Banerjee 2010) The decrease in H2S caused bySQRD-1 is likely to reduce internal H2S levels to a toleratedrange
A second enzyme required for the H2S oxidation pathwayis ETHE1 (Tiranti et al 2009) ETHE1 is a sulfur dioxyge-nase required in mammals for oxidation of sulfur to sulfate(Kabil and Banerjee 2010) C elegans has a single homologof Ethe1 C33A127 which we named ethe-1 We obtainedthe available ethe-1 (tm4101) deletion allele and tested forH2S sensitivity (Table 2) Similar to sqrd-1 (tm3378) ethe-1(tm4101) is sensitive to RAH2S which suggests that sqrd-1confers resistance to H2S through the sulfurndashoxidationpathway
Epistasis with egl-9
Egl-9ndashdeficient animals have been shown to be resistant toboth H2S and HCN and our data suggest that resistance ismediated by HIF-1 upregulating sqrd-1 and cysl-2 respec-tively To test this hypothesis we used RNAi to knockdownegl-9 in mutant animals which we have described as H2Sand HCN sensitive and tested them for resistance to H2S andHCN Table 4 shows that egl-9 knockdown does not mediateresistance to HCN in cysl-1 and cysl-2 mutant backgrounds
Table 3 Innate immunity to P aeroginosa requires hif-1 cysl-1and cysl-2
Strain Pretreatment AlleleSurvival on
P aeruginosa ()
N2 Control RNAi mdash 0 6 0N2 Egl-9 RNAi mdash 89 6 19hif-1 Egl-9 RNAi ia04 10 6 16egl-9 Egl-9 RNAi sa307 100 6 0cysl-1 Egl-9 RNAi ok762 4 6 7cysl-2 Egl-9 RNAi ok3516 0 6 0sqrd-1 Egl-9 RNAi tm3378 100 6 0ethe-1 Egl-9 RNAi tm4101 100 6 0N2 H2S grown mdash 83 6 29y
P 005 as compared to egl-9 knockdown in N2 animals yP 005 as comparedto untreated control RNAi N2 animals
Figure 6 (A) Both sqrd-1and cysl-2 mRNA levels increaserelative to untreated animals as measured by quantitativereverse transciptase PCR Y axis is relative mRNA concen-tration compared to room air-treated control animalsP 005 as compared to animals grown in room air(B) hif-1 is required for sqrd-1 and cysl-2 mRNA inductionwhen exposed to RAH2S or RAHCN Arrowheads indicatethat mRNA levels were observed at very low levels P 005 when comparing wild-type and hif-1mutant animals(C) Western blot shows SQRD-1 protein induction in RAH2S This induction requires hif-1 No immunoreactivity isobserved in sqrd-1 mutant animals SQRD-1 protein isabundant in egl-9 mutant animals Circles indicate proteinsize markers of 116 82 62 and 49 kDa from top tobottom The calculated molecular weight of 53 kDa cor-responds well with the observed relative migration
528 M W Budde and M B Roth
and therefore cysl-1 and cysl-2 are epistatic to egl-9 Sqrd-1and ethe-1 mutant animals with knocked down egl-9 arepartially resistant to H2S
Discussion
C elegans likely encounters H2S in the soil as a result ofbreakdown of organic sulfur (Morra and Dick 1991) andHCN produced by both bacteria (Gallagher and Manoil2001) and plants (Garciacutea et al 2010) This study has exam-ined the mechanisms that C elegans uses to respond to highlevels of H2S and HCN RAH2S and RAHCN both cause an
increase in HIF-1 activity which increases the expression ofsqrd-1 and cysl-2 to catalyze the first steps of the metabolismof H2S and HCN respectively sqrd-1 acts to oxidize H2Sultimately preventing internal H2S levels from exceedingthe maximum tolerated concentration cysl-2 acts in parallelto keep HCN levels low however equimolar amounts of H2Sare generated as a consequence of cysl-2 activity cysl-1 isrequired to reduce H2S levels that are produced by cysl-2during RAHCN exposure (Figure 8)
Here we provide direct evidence that H2S acts throughhif-1 to increase both SQRD-1 mRNA and protein expressionThis initiates the sulfidendashoxidation pathway which lowers in-ternal H2S levels (Figure 8) SQRD-1 expression appears to belocalized to the cells most readily exposed to exogenous H2S inthe worm In mammals colon epithelial cells that express highlevels of SQRDL (Ponteacuten et al 2008) are likely to do so be-cause a prominent enteric bacteria Desulfovibrio piger produ-ces high levels of H2S in the lower intestine (Loubinoux et al2002) The requirement for ethe-1 during RAH2S exposureconfirms that the sulfidendashoxidation pathway is the essentialfunction of sqrd-1 The sulfidendashoxidation pathway requires ox-ygen both to maintain a functional electron transport chain (togenerate oxidized ubiquinone) and to directly oxidize sulfurvia ethe-1 Exposure of mammals to exogenous H2S results inmany alterations in physiology including the induction of a sus-pended-animationndashlike state and improved outcome in ische-mia reperfusion injury It will be interesting to learn whetherSQRDL expression is induced upon exposure to H2S inmammals
The question remains as to why SQRD-1 is inducible atall Surrounded by decaying matter C elegans must encoun-ter areas of high H2S in its natural environment Havingconstitutively high SQRD-1 levels as in egl-9 mutant ani-mals allows survival when wild-type animals with lowerHIF-1 activity would perish However low levels of H2Sare important for biological function so perhaps nematodesare using SQRD-1 to deliberately titrate the cellular concen-tration of H2S by controlling its rate of destruction In mam-mals the H2S-generating enzymes CBS CSE and 3MPcontrol sulfide levels (Kabil and Banerjee 2010) Howeverour study suggests that mammals may also modulate cellu-lar H2S levels by regulating SQRDL-dependent destruction
In addition to furthering our understanding of the sulfideoxidation pathway this study also provides insight into two
Figure 7 SQRD-1GFP expression is induced by exposure to RAH2S (A)SQRD-1GFP is not expressed in room air (B) RAH2S at 50 ppm inducesexpression of SQRD-1GFP (C) Confocal microscopy at middot40 magnifica-tion shows that the expression is brightest in the muscle dense bodies(tailed arrows) and the hypodermis (tail-less arrow) (D) Cross-section(Z-stack) shows expression is most intense in the hypodermis
Table 4 Epistasis relationships with egl-9
15 ppm RAHCN 250 ppm RAH2S
Empty vector () egl-9 RNAi () Empty vector () egl-9 RNAi ()
N2 0 6 0 92 6 10 0 6 0 98 6 4hif-1 (ia04) 0 6 0 0 6 0 0 6 0 0 6 0egl-9 (sa307) 100 6 0 100 6 0 100 6 0 100 6 0sqrd-1 (tm3378) 0 6 0 87 6 16 0 6 0 47 6 15ethe-1 (tm4101) 0 6 0 89 6 14 0 6 0 73 6 10cysl-1 (ok762) 0 6 0 27 6 35 0 6 0 68 6 30cysl-2 (ok3516) 0 6 0 0 6 0 0 6 0 94 6 6
P 005 as compared to egl-9 RNAi knockdown in wild-type animals Data are presented as percentage of survival
The Response of C elegans to H2S and HCN 529
new metabolic activities previously unknown in animalscysteine synthase and cyanoalanine synthase cysl-1 mutantanimals are sensitive to both RAH2S and RAHCN whilecysl-2 mutant animals are only sensitive to RAHCN Thissuggests that cysl-1 is a cysteine synthase acting downstreamof cysl-2 a cyanoalanine synthase (Figure 8) We hypothe-size that cysl-1 mutant animals could be succumbing to H2Sproduced by CYSL-2 as a result of cyanide assimilation Inthis way each molecule of HCN assimilated by CYSL-2 wouldproduce one molecule of H2S In this reaction mechanismCYSL-1 might regenerate cysteine used in cyanide assimila-tion Alternatively it is possible that CYSL-1 is catalyzing theassimilation of both H2S and HCN a hypothesis supportedby in vitro plant studies (Hell et al 2002) The function ofthe two remaining cysteine synthase homologs cysl-3 andcysl-4 remains to be elucidated but they may also haveimportant CYSCAS activity during development or in spe-cific tissues
To our knowledge cysl-1 is the first reported functionalCYS homolog in animals Indeed while no orthologs appearto be present in animals outside of the genus Caenorhabditisin plants and bacteria CYS uses OAS as the H2S-acceptingsubstrate (Feldman-Salit et al 2009) Plants and bacteriagenerate OAS by acetylating serine with the enzyme serineacetyltransferase (SAT) (Wirtz et al 2010) which has nohomolog in C elegans How might C elegans be obtainingOAS As with many other amino acids C elegans is likelyobtaining OAS from its bacterial food source AlternativelyC elegans might have evolved a mechanism of generatingOAS using alternative enzymes CYS activity is essential forcysteine biosynthesis in prototrophs (Feldman-Salit et al2009) Because the critical aspect of CYS appears to be theelimination of H2S instead of the generation of cysteine
perhaps C elegans is freed from using OAS as the H2S ac-ceptor and is using an alternative H2S acceptor that does notgenerate cysteine In mammals the closest mammalian ho-molog to CYS is CBS Consistent with the idea that mammalshave CYS activity there is evidence that radiolabeled H2Scan be incorporated into cysteine in germ-free rats (Huovinenand Gustafsson 1967)
The protective effect of cysl-2 to HCN explains the pre-vious observation that egl-9 mutant animals are resistant tothe pathogenic Pseudomonas fast paralytic killing P aerugi-nosa is a clinically relevant pathogen and can produce toxiccyanide levels in the sputum of cystic fibrosis patients (Andersonet al 2010) P fluorescens is a cyanide-producing soil bacte-ria that has also been shown to exhibit cyanide-dependentfast paralytic killing effect on C elegans (Romanowski et al2011) HIF-1 activates resistance to hypoxia and this hasbeen proposed to also confer resistance to the cytochromec oxidase inhibiting effects of cyanide (Gallagher andManoil 2001 Shao et al 2010 Romanowski et al 2011)While this certainly may play some role we provide evidencehere that the resistance of egl-9 mutant animals to P aerugi-nosa requires cysl-2 likely by directly detoxifying cyanide tob-cyanoalanine (Figure 8) This reaction produces H2S whichcan be further detoxified by cysl-1 and sqrd-1
While we have demonstrated critical components of theRAH2S-induced hif-1 response the mechanism of hif-1 in-duction remains unclear The overlapping gene expressionpatterns between both H2S and HCN suggests that C eleganshas a shared response to these two molecules In fact ourstudies suggest that C elegans is unable to discriminate be-tween H2S and HCN leading them to activate the degrada-tion pathways of both molecules when either is presentedThus C elegans treated with RAH2S or RAHCN increases
Figure 8 Proposed mechanismBoth H2S and HCN can inhibitelectron transport Electron trans-port inhibition causes inductionof HIF-1 activity resulting in high ex-pression of SQRD-1 and CYSL-2SQRD-1 catalyzes the first step inthe H2S-oxidation pathway ulti-mately resulting in production ofsulfate and thiosulfate CYSL-2catalyzes the first step in the HCNassimilation pathway producingH2S The resultant H2S is detoxifiedby CYSL-1
530 M W Budde and M B Roth
both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841
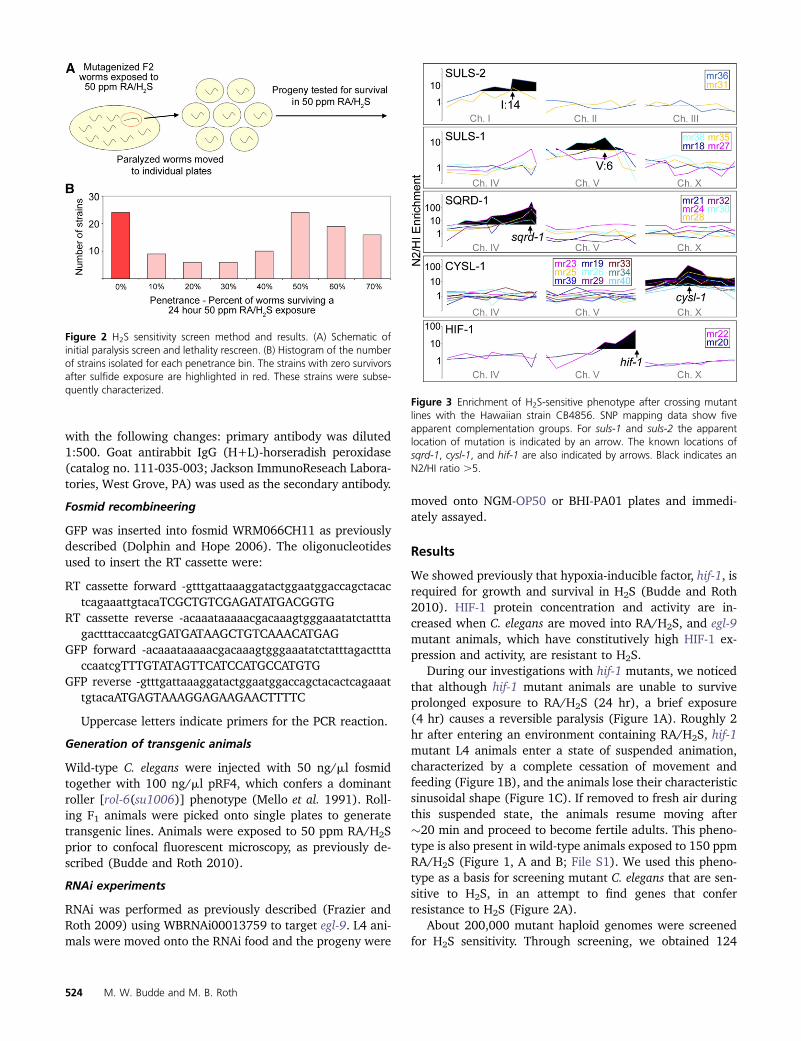
with the following changes primary antibody was diluted1500 Goat antirabbit IgG (H+L)-horseradish peroxidase(catalog no 111-035-003 Jackson ImmunoReseach Labora-tories West Grove PA) was used as the secondary antibody
Fosmid recombineering
GFP was inserted into fosmid WRM066CH11 as previouslydescribed (Dolphin and Hope 2006) The oligonucleotidesused to insert the RT cassette were
RT cassette forward -gtttgattaaaggatactggaatggaccagctacactcagaaattgtacaTCGCTGTCGAGATATGACGGTG
RT cassette reverse -acaaataaaaacgacaaagtgggaaatatctatttagactttaccaatcgGATGATAAGCTGTCAAACATGAG
GFP forward -acaaataaaaacgacaaagtgggaaatatctatttagactttaccaatcgTTTGTATAGTTCATCCATGCCATGTG
GFP reverse -gtttgattaaaggatactggaatggaccagctacactcagaaattgtacaATGAGTAAAGGAGAAGAACTTTTC
Uppercase letters indicate primers for the PCR reaction
Generation of transgenic animals
Wild-type C elegans were injected with 50 ngml fosmidtogether with 100 ngml pRF4 which confers a dominantroller [rol-6(su1006)] phenotype (Mello et al 1991) Roll-ing F1 animals were picked onto single plates to generatetransgenic lines Animals were exposed to 50 ppm RAH2Sprior to confocal fluorescent microscopy as previously de-scribed (Budde and Roth 2010)
RNAi experiments
RNAi was performed as previously described (Frazier andRoth 2009) using WBRNAi00013759 to target egl-9 L4 ani-mals were moved onto the RNAi food and the progeny were
moved onto NGM-OP50 or BHI-PA01 plates and immedi-ately assayed
Results
We showed previously that hypoxia-inducible factor hif-1 isrequired for growth and survival in H2S (Budde and Roth2010) HIF-1 protein concentration and activity are in-creased when C elegans are moved into RAH2S and egl-9mutant animals which have constitutively high HIF-1 ex-pression and activity are resistant to H2S
During our investigations with hif-1 mutants we noticedthat although hif-1 mutant animals are unable to surviveprolonged exposure to RAH2S (24 hr) a brief exposure(4 hr) causes a reversible paralysis (Figure 1A) Roughly 2hr after entering an environment containing RAH2S hif-1mutant L4 animals enter a state of suspended animationcharacterized by a complete cessation of movement andfeeding (Figure 1B) and the animals lose their characteristicsinusoidal shape (Figure 1C) If removed to fresh air duringthis suspended state the animals resume moving after20 min and proceed to become fertile adults This pheno-type is also present in wild-type animals exposed to 150 ppmRAH2S (Figure 1 A and B File S1) We used this pheno-type as a basis for screening mutant C elegans that are sen-sitive to H2S in an attempt to find genes that conferresistance to H2S (Figure 2A)
About 200000 mutant haploid genomes were screenedfor H2S sensitivity Through screening we obtained 124
Figure 3 Enrichment of H2S-sensitive phenotype after crossing mutantlines with the Hawaiian strain CB4856 SNP mapping data show fiveapparent complementation groups For suls-1 and suls-2 the apparentlocation of mutation is indicated by an arrow The known locations ofsqrd-1 cysl-1 and hif-1 are also indicated by arrows Black indicates anN2HI ratio 5
Figure 2 H2S sensitivity screen method and results (A) Schematic ofinitial paralysis screen and lethality rescreen (B) Histogram of the numberof strains isolated for each penetrance bin The strains with zero survivorsafter sulfide exposure are highlighted in red These strains were subse-quently characterized
524 M W Budde and M B Roth
lines that were sensitive to H2S (Figure 2B) Of these lines23 were completely penetrant where penetrance is definedas the percentage of worms unable to survive a 24-hr RAH2S exposure These mutant strains were chosen for furthercharacterization
The 23 strongly penetrant mutant lines were crossed withthe Hawaiian CB4856 strain for mapping the sensitivity traitto chromosomal regions using enrichment of SNPs The SNPanalysis revealed that 22 mutants map to only five chromo-somal regions (Figure 3 Table 1) only mr37 showed noMendelian inheritance of the trait These mutants were thencrossed to confirm that each chromosomal region containeda single complementation group (Figure 4) One group onchromosome V (mr18 mr27 mr35 and mr38) exhibitsdominant genetics and thus could not be further tested forcomplementation We named this gene suls-1 (sulfide sen-sitive) and suls-1 (mr35) was mapped to the 4-cM regionbetween SNP pkP5097 and pkP5068 The two alleles onchromosome I were named suls-2 The genes mutated insuls-1 and suls-2 strains remain to be identified Two alleleson chromosome V (mr20 and mr22) mapped near the hif-1locus and failed to complement the hif-1(ia04) mutationwhich confirms that we successfully screened for our phe-notype of interest
The 23 strongest mutant lines were next tested forsurvival in room air containing lower H2S concentrationsin an effort to phenotypically discriminate between the com-
plementation groups Table 1 shows that complementationgroups have differences in H2S sensitivity with the groupmapping to the X chromosome being the most sensitive andsuls-2 being the least sensitive In contrast to the other com-plementation groups suls-1 alleles vary in sensitivity to H2S
A cysteine synthase homolog cysl-1 is required forsurvival in H2S
mr23 was used as a representative allele of the group ofalleles on the X chromosome (mr19 mr23 mr25 mr26mr29 mr33 mr34 mr39 and mr40) and was mapped toa 02-cM region between SNPs pas23398 and snp_F41E7[3] This region contains 70 predicted genes Of the deletionmutants available for that region ok762 phenocopies andfails to complement these H2S-sensitive mutations Theok762 allele deletes five of the six exons of the geneC17G17 Sequencing a portion of C17G17 of our isolatedstrains revealed adenine-to-guanine point mutations whichare characteristic of EMS mutagenesis (Figure S1) All iden-tified mutations were either missense or splice mutations
C17G17 is a previously uncharacterized gene encodinga member of the cystathionine b-synthase (CBS)-like sub-group which is made up of enzymes important in cysteineproduction CBS and cysteine synthase (CYS) (OAS-TL andCysK) (Feldman-Salit et al 2009) Animals generally haveCBS homologs and bacteria and plants tend to have CYShomologs (Hell et al 2008) CBS is critical for generating
Table 1 Alleles unable to survive in RAH2S
Alivetotal ( surviving)
Gene name Chromosome Allele 32 ppm RAH2S 15 ppm RAH2S 75 ppm RAH2S Dominance
N2 mdash mdash 1212 (100) 1212 (100) 1313 (100) mdash
cysl-1 X ok762 020 (0) 018 (0) 011 (0) mdash
cysl-1 X mr19 013 (0) 012 (0) 010 (0) Recessivecysl-1 X mr23 016 (0) 010 (0) 010 (0) Recessivecysl-1 X mr25 013 (0) 010 (0) 06 (0) Recessivecysl-1 X mr26 012 (0) 010 (0) 011 (0) Recessivecysl-1 X mr33 015 (0) 59 (56) 1010 (100) Recessivecysl-1 X mr34 010 (0) 011 (0) 110 (10) Recessivecysl-1 X mr29 011 (0) 010 (0) 113 (8) Recessivecysl-1 X mr39 010 (0) 010 (0) 1313 (100) Recessivecysl-1 X mr40 010 (0) 010 (0) 111 (9) Recessivesqrd-1 IV tm3378 510 (50) 1010 (100) 1010 (100) mdash
sqrd-1 IV mr24 1014 (71) 1111 (100) mdash Recessivesqrd-1 IV mr30 213 (15) 99 (100) mdash Recessivesqrd-1 IV mr32 1213 (92) 1010 (100) mdash Recessivesqrd-1 IV mr21 615 (40) 1010 (100) mdash Recessivesqrd-1 IV mr28 59 (56) 1010 (100) 1111 (100) Recessivehif-1 V ia04 015 (0) 011 (0) 1010 (100) mdash
hif-1 V mr20 014 (0) 010 (0) 1013 (77) Recessivehif-1 V mr22 015 (0) 111 (9) 810 (80) Recessivesuls-1 V mr18 612 (50) 1112 (92) 1010 (100) Dominantsuls-1 V mr27 29 (22) 1010 (100) mdash Dominantsuls-1 V mr35 05 (0) 010 (0) 010 (0) Dominantsuls-1 V mr38 09 (0) 88 (100) mdash Dominantsuls-2 I mr31 78 (88) 88 (100) mdash Recessivesuls-2 I mr36 89 (89) 1111 (100) mdash Recessivemdash mdash mr37 1112 (92) 1111 (100) mdash mdash
The Response of C elegans to H2S and HCN 525
cysteine in animals which make cysteine from dietary me-thionine (Finkelstein et al 1988) CBS is also one of the onlyH2S-generating enzymes in mammals (Kabil and Banerjee2010) In plants and bacteria which are cysteine proto-trophs CYS combines H2S with O-acetylserine (OAS) togenerate cysteine (Figure 5A) (Wirtz et al 2010) Humansand mice each have one CBS-like family member In con-trast C elegans has six C17G17 K10H102 R08E52F59A79 and F54A34 (Table 2)
C17G17 (cysl-1) K10H102 (cysl-2) R08E52 (cysl-3) andF59A79 (cysl-4) are more similar to CYS from plants andbacteria whereas ZC3731 (cbs-1) and F54A34 (cbs-2) aremore similar to CBS from animals (Figure 5C) A BLASTsearch reveals that the genus Caenorhabditis contains the onlyknown animal CYS homologs We have renamed these genescysl-1 (cysteine synthase-like) cysl-2 cysl-3 cysl-4 cbs-1 andcbs-2 to reflect this homology Depletion of H2S levels as it isused as a substrate by CYS may explain why cysl-1 mutantanimals are sensitive to H2S Nematodes appear to have ac-quired the ability to assimilate H2S and cysl-1 is likely usingthis reaction to drive down internal H2S levels
cysl-2 mediates immunity to fast paralytic killingby Pseudomonas
The existence of three additional Cysl homologs in C elegansraises the possibility that these other Cysl enzymes are alsoimportant in H2S detoxification This possibility is especiallyintriguing because we have shown a hif-1ndashdependent in-crease in cysl-2 transcript levels during RAH2S exposure(Budde and Roth 2010) We obtained the available knock-out strains of cysl-2 cysl-4 and cbs-2 and tested them forRAH2S sensitivity but none showed a RAH2S sensitivityphenotype (Table 2)
In plants CYS and cyanoalanine synthase (CAS) areclosely related and have overlapping catalytic activities(Figure 5 A and B) The CAS reaction combines cysteineand HCN to produce b-cyanoalanine and H2S lowering cel-lular HCN levels (Figure 5B) Furthermore cysl-2 mRNA waspreviously shown to be upregulated by HIF-1 (Shen et al2005 Shao et al 2010 Budde and Roth 2010) and highHIF-1 activity has been shown to create resistance to HCN-mediated fast paralytic killing by P aeruginosa (Darby et al1999 Gallagher and Manoil 2001 Shao et al 2010) There-fore cysl-2 may be induced not to effect H2S levels but in-stead may be the mediator of resistance to HCN-dependentfast paralytic killing To test this hypothesis we exposed cysl-1 cysl-2 and cysl-4 mutant animals to RAHCN and foundthat both cysl-1 and cysl-2 mutant animals are sensitive toRAHCN (Table 2) The specificity of cysl-2 mutant sensitivityto cyanide suggests that this enzyme is directly acting to di-minish HCN levels rather than to affect H2S levels The sen-sitivity of cysl-1 mutants to both RAH2S and RAHCNsuggest that cysl-1 acts downstream of cysl-2 and cysl-1mutants are being overwhelmed by the H2S produced bycysl-2 during HCN detoxification (Figure 8)
Next we tested whether these enzymes are responsiblefor HIF-1ndashmediated innate immunity to P aeruginosa Weshowed that similar to previous studies (Gallagher andManoil 2001 Shao et al 2010) egl-9ndashdeficient animals haveincreased innate immunity to P aeruginosa (Table 3) Nextwe demonstrated that cysl-1 and cysl-2 are both requiredfor the observed innate immunity to P aeruginosa demon-strating a mechanism of innate immunity to Pseudomonas-dependent fast paralytic killing Since we previously showedthat animals grown in RAH2S have increased cysl-2 mRNAexpression we assayed whether growth in H2S confers
Figure 4 Complementation datashowing that each genetic locusconsists of only one complemen-tation group (blue green yellowand pink shades) suls-1 is notshown because it is dominantsqrd-1 cysl-1 double heterozy-gotes are slightly sensitive toRAH2S
526 M W Budde and M B Roth
resistance to P aeruginosa Indeed our results show thatgrowth in H2S increases the innate immunity of C elegansto P aeruginosa (Table 3)
sqrd-1 is required for survival in H2S
The group of alleles on chromosome IV mr21 mr24 mr28mr30 and mr32 were mapped near the SNP pkP4095 lo-cated at the genetic location IV12 Analysis of the regionrevealed a gene F02H65 encoding an apparent homolog ofthe protein sulfidequinone reductase dehydrogenase (SQRD)which we have named sqrd-1 to reflect this homology SQRDwas initially described in prokaryotes as oxidizing sulfideand transferring the electrons to ubiquinone for generationof ATP during oxidative phosphorylation (Vande Weghe andOw 1999) The available deletion allele of sqrd-1 (tm3378)phenocopies and fails to complement these mutants Se-
quence analysis revealed A-to-G transitions in our isolatedmutants (Figure S1B)
The C elegans gene sqrd-1 encodes a member of thegroup II SQRD enzymes which also includes human mouseand Drosophila homologs The C elegans genome is predictedto encode another SQRD homolog Y9C9A16 (sqrd-2)which was initially classified as a pseudogene We obtainedthe deletion allele of sqrd-2 and found that it is not sensitiveto RAH2S
Transcriptional regulation of identified genes
We undertook this screen to determine the mechanism bywhich HIF-1 confers resistance to H2S Therefore we wereinterested to learn whether either cysl-1 or sqrd-1 is tran-scriptionally regulated by HIF-1 in response to RAH2S ex-posure Figure 6A shows that sqrd-1 and cysl-2 mRNA levels
Figure 5 (A) Cysteine synthase (CYS) catalyzes the formation of cysteine and acetate from OAS and H2S (B) Cyanoalanine synthase (CAS) catalyzes theformation of b-cyanoalanine and H2S from cysteine and HCN (C) Amino acid alignment of a selected region of CYS and CBS homologs C elegans Cyslgenes are homologous to CYS whereas C elegans Cbs genes are homologous to cystathionine b-synthase (CBS)
Table 2 Genes named in this article
Gene name Gene ID Allele tested 50 ppm RAH2S survival () 5 ppm RAHCN survival ()
cysl-1 C17G17 ok762 0 6 0 0 6 0cysl-2 K10H102 ok3516 100 6 0 0 6 0cysl-3 R08E52 mdash mdash mdash
cysl-4 F59A79 ok3359 92 6 11 100 6 0cbs-1 ZC3731 mdash mdash mdash
cbs-2 F54A34 ok666 100 6 0 100 6 0sqrd-1 F02H65 tm3378 0 6 0 97 6 5ethe-1 C33A127 tm4101 0 6 0 100 6 0
P 005 as compared to N2 wild-type animals
The Response of C elegans to H2S and HCN 527
increase during RAH2S and RAHCN exposure but cysl-1mRNA levels remain unchanged Furthermore hif-1(ia04)mutant animals fail to increase sqrd-1 and cysl-2 mRNA lev-els during RAH2S exposure (Figure 6B) suggesting thatupregulation of sqrd-1 by HIF-1 is a mechanism for survivingin the presence of H2S
To determine whether or not SQRD-1 protein levels in-crease during RAH2S exposure we performed a Westernblot to determine relative protein concentrations Figure 6Cshows that SQRD-1 protein levels increase during RAH2Sexposure in the parental N2 Bristol strain No SQRD-1 pro-tein accumulation is observed in the sqrd-1(tm3378) or inthe hif-1(ia04) mutant animals egl-9(sa307) mutant ani-mals have constitutively high levels of SQRD-1 proteinwhich may explain their ability to survive in 100 timeshigher RAH2S concentration than hif-1(ia04) mutantanimals
SQRD-1GFP protein is expressed during H2S exposure
Next we analyzed the expression pattern of SQRD-1 in Celegans We created a SQRD-1GFP fusion construct becausethe antibody we created was unable to stain worms by im-munofluorescence The presence of a predicted SL2 alterna-tive splice site upstream of sqrd-1 indicated that a standardpromoter fusion might not accurately recapitulate the en-dogenous expression pattern Therefore we used the recom-
bineering approach for GFP fusion construction whichincludes a 35-kb region surrounding the 45-kb SQRD-1gene RAH2S exposure causes a fluorescence increase inrecombinant animals that is most apparent in the anteriorof the worm (Figure 7) Expression appears limited to thehypodermis the body wall muscle and the anterior bulb ofthe pharynx (Figure 7C) A cross section through the wormconfirms that expression is limited to the most exposed partsof the worm (Figure 7D) It is possible that this SQRD-1expression pattern reflects a function of SQRD-1 to preventH2S from reaching the interior tissue
We conclude from these experiments that HIF-1 acts toinitiate the response of C elegans to H2S by upregulating theSQRD-1 protein levels SQRD-1 is an oxidoreductase oxidiz-ing H2S to polysulfide and reducing ubiquinone (Theissenet al 2003) The oxidation of sulfide generates polysulfide(Marcia et al 2009) which can then be further oxidized viathe H2S oxidation pathway to yield sulfate and thiosulfate(Kabil and Banerjee 2010) The decrease in H2S caused bySQRD-1 is likely to reduce internal H2S levels to a toleratedrange
A second enzyme required for the H2S oxidation pathwayis ETHE1 (Tiranti et al 2009) ETHE1 is a sulfur dioxyge-nase required in mammals for oxidation of sulfur to sulfate(Kabil and Banerjee 2010) C elegans has a single homologof Ethe1 C33A127 which we named ethe-1 We obtainedthe available ethe-1 (tm4101) deletion allele and tested forH2S sensitivity (Table 2) Similar to sqrd-1 (tm3378) ethe-1(tm4101) is sensitive to RAH2S which suggests that sqrd-1confers resistance to H2S through the sulfurndashoxidationpathway
Epistasis with egl-9
Egl-9ndashdeficient animals have been shown to be resistant toboth H2S and HCN and our data suggest that resistance ismediated by HIF-1 upregulating sqrd-1 and cysl-2 respec-tively To test this hypothesis we used RNAi to knockdownegl-9 in mutant animals which we have described as H2Sand HCN sensitive and tested them for resistance to H2S andHCN Table 4 shows that egl-9 knockdown does not mediateresistance to HCN in cysl-1 and cysl-2 mutant backgrounds
Table 3 Innate immunity to P aeroginosa requires hif-1 cysl-1and cysl-2
Strain Pretreatment AlleleSurvival on
P aeruginosa ()
N2 Control RNAi mdash 0 6 0N2 Egl-9 RNAi mdash 89 6 19hif-1 Egl-9 RNAi ia04 10 6 16egl-9 Egl-9 RNAi sa307 100 6 0cysl-1 Egl-9 RNAi ok762 4 6 7cysl-2 Egl-9 RNAi ok3516 0 6 0sqrd-1 Egl-9 RNAi tm3378 100 6 0ethe-1 Egl-9 RNAi tm4101 100 6 0N2 H2S grown mdash 83 6 29y
P 005 as compared to egl-9 knockdown in N2 animals yP 005 as comparedto untreated control RNAi N2 animals
Figure 6 (A) Both sqrd-1and cysl-2 mRNA levels increaserelative to untreated animals as measured by quantitativereverse transciptase PCR Y axis is relative mRNA concen-tration compared to room air-treated control animalsP 005 as compared to animals grown in room air(B) hif-1 is required for sqrd-1 and cysl-2 mRNA inductionwhen exposed to RAH2S or RAHCN Arrowheads indicatethat mRNA levels were observed at very low levels P 005 when comparing wild-type and hif-1mutant animals(C) Western blot shows SQRD-1 protein induction in RAH2S This induction requires hif-1 No immunoreactivity isobserved in sqrd-1 mutant animals SQRD-1 protein isabundant in egl-9 mutant animals Circles indicate proteinsize markers of 116 82 62 and 49 kDa from top tobottom The calculated molecular weight of 53 kDa cor-responds well with the observed relative migration
528 M W Budde and M B Roth
and therefore cysl-1 and cysl-2 are epistatic to egl-9 Sqrd-1and ethe-1 mutant animals with knocked down egl-9 arepartially resistant to H2S
Discussion
C elegans likely encounters H2S in the soil as a result ofbreakdown of organic sulfur (Morra and Dick 1991) andHCN produced by both bacteria (Gallagher and Manoil2001) and plants (Garciacutea et al 2010) This study has exam-ined the mechanisms that C elegans uses to respond to highlevels of H2S and HCN RAH2S and RAHCN both cause an
increase in HIF-1 activity which increases the expression ofsqrd-1 and cysl-2 to catalyze the first steps of the metabolismof H2S and HCN respectively sqrd-1 acts to oxidize H2Sultimately preventing internal H2S levels from exceedingthe maximum tolerated concentration cysl-2 acts in parallelto keep HCN levels low however equimolar amounts of H2Sare generated as a consequence of cysl-2 activity cysl-1 isrequired to reduce H2S levels that are produced by cysl-2during RAHCN exposure (Figure 8)
Here we provide direct evidence that H2S acts throughhif-1 to increase both SQRD-1 mRNA and protein expressionThis initiates the sulfidendashoxidation pathway which lowers in-ternal H2S levels (Figure 8) SQRD-1 expression appears to belocalized to the cells most readily exposed to exogenous H2S inthe worm In mammals colon epithelial cells that express highlevels of SQRDL (Ponteacuten et al 2008) are likely to do so be-cause a prominent enteric bacteria Desulfovibrio piger produ-ces high levels of H2S in the lower intestine (Loubinoux et al2002) The requirement for ethe-1 during RAH2S exposureconfirms that the sulfidendashoxidation pathway is the essentialfunction of sqrd-1 The sulfidendashoxidation pathway requires ox-ygen both to maintain a functional electron transport chain (togenerate oxidized ubiquinone) and to directly oxidize sulfurvia ethe-1 Exposure of mammals to exogenous H2S results inmany alterations in physiology including the induction of a sus-pended-animationndashlike state and improved outcome in ische-mia reperfusion injury It will be interesting to learn whetherSQRDL expression is induced upon exposure to H2S inmammals
The question remains as to why SQRD-1 is inducible atall Surrounded by decaying matter C elegans must encoun-ter areas of high H2S in its natural environment Havingconstitutively high SQRD-1 levels as in egl-9 mutant ani-mals allows survival when wild-type animals with lowerHIF-1 activity would perish However low levels of H2Sare important for biological function so perhaps nematodesare using SQRD-1 to deliberately titrate the cellular concen-tration of H2S by controlling its rate of destruction In mam-mals the H2S-generating enzymes CBS CSE and 3MPcontrol sulfide levels (Kabil and Banerjee 2010) Howeverour study suggests that mammals may also modulate cellu-lar H2S levels by regulating SQRDL-dependent destruction
In addition to furthering our understanding of the sulfideoxidation pathway this study also provides insight into two
Figure 7 SQRD-1GFP expression is induced by exposure to RAH2S (A)SQRD-1GFP is not expressed in room air (B) RAH2S at 50 ppm inducesexpression of SQRD-1GFP (C) Confocal microscopy at middot40 magnifica-tion shows that the expression is brightest in the muscle dense bodies(tailed arrows) and the hypodermis (tail-less arrow) (D) Cross-section(Z-stack) shows expression is most intense in the hypodermis
Table 4 Epistasis relationships with egl-9
15 ppm RAHCN 250 ppm RAH2S
Empty vector () egl-9 RNAi () Empty vector () egl-9 RNAi ()
N2 0 6 0 92 6 10 0 6 0 98 6 4hif-1 (ia04) 0 6 0 0 6 0 0 6 0 0 6 0egl-9 (sa307) 100 6 0 100 6 0 100 6 0 100 6 0sqrd-1 (tm3378) 0 6 0 87 6 16 0 6 0 47 6 15ethe-1 (tm4101) 0 6 0 89 6 14 0 6 0 73 6 10cysl-1 (ok762) 0 6 0 27 6 35 0 6 0 68 6 30cysl-2 (ok3516) 0 6 0 0 6 0 0 6 0 94 6 6
P 005 as compared to egl-9 RNAi knockdown in wild-type animals Data are presented as percentage of survival
The Response of C elegans to H2S and HCN 529
new metabolic activities previously unknown in animalscysteine synthase and cyanoalanine synthase cysl-1 mutantanimals are sensitive to both RAH2S and RAHCN whilecysl-2 mutant animals are only sensitive to RAHCN Thissuggests that cysl-1 is a cysteine synthase acting downstreamof cysl-2 a cyanoalanine synthase (Figure 8) We hypothe-size that cysl-1 mutant animals could be succumbing to H2Sproduced by CYSL-2 as a result of cyanide assimilation Inthis way each molecule of HCN assimilated by CYSL-2 wouldproduce one molecule of H2S In this reaction mechanismCYSL-1 might regenerate cysteine used in cyanide assimila-tion Alternatively it is possible that CYSL-1 is catalyzing theassimilation of both H2S and HCN a hypothesis supportedby in vitro plant studies (Hell et al 2002) The function ofthe two remaining cysteine synthase homologs cysl-3 andcysl-4 remains to be elucidated but they may also haveimportant CYSCAS activity during development or in spe-cific tissues
To our knowledge cysl-1 is the first reported functionalCYS homolog in animals Indeed while no orthologs appearto be present in animals outside of the genus Caenorhabditisin plants and bacteria CYS uses OAS as the H2S-acceptingsubstrate (Feldman-Salit et al 2009) Plants and bacteriagenerate OAS by acetylating serine with the enzyme serineacetyltransferase (SAT) (Wirtz et al 2010) which has nohomolog in C elegans How might C elegans be obtainingOAS As with many other amino acids C elegans is likelyobtaining OAS from its bacterial food source AlternativelyC elegans might have evolved a mechanism of generatingOAS using alternative enzymes CYS activity is essential forcysteine biosynthesis in prototrophs (Feldman-Salit et al2009) Because the critical aspect of CYS appears to be theelimination of H2S instead of the generation of cysteine
perhaps C elegans is freed from using OAS as the H2S ac-ceptor and is using an alternative H2S acceptor that does notgenerate cysteine In mammals the closest mammalian ho-molog to CYS is CBS Consistent with the idea that mammalshave CYS activity there is evidence that radiolabeled H2Scan be incorporated into cysteine in germ-free rats (Huovinenand Gustafsson 1967)
The protective effect of cysl-2 to HCN explains the pre-vious observation that egl-9 mutant animals are resistant tothe pathogenic Pseudomonas fast paralytic killing P aerugi-nosa is a clinically relevant pathogen and can produce toxiccyanide levels in the sputum of cystic fibrosis patients (Andersonet al 2010) P fluorescens is a cyanide-producing soil bacte-ria that has also been shown to exhibit cyanide-dependentfast paralytic killing effect on C elegans (Romanowski et al2011) HIF-1 activates resistance to hypoxia and this hasbeen proposed to also confer resistance to the cytochromec oxidase inhibiting effects of cyanide (Gallagher andManoil 2001 Shao et al 2010 Romanowski et al 2011)While this certainly may play some role we provide evidencehere that the resistance of egl-9 mutant animals to P aerugi-nosa requires cysl-2 likely by directly detoxifying cyanide tob-cyanoalanine (Figure 8) This reaction produces H2S whichcan be further detoxified by cysl-1 and sqrd-1
While we have demonstrated critical components of theRAH2S-induced hif-1 response the mechanism of hif-1 in-duction remains unclear The overlapping gene expressionpatterns between both H2S and HCN suggests that C eleganshas a shared response to these two molecules In fact ourstudies suggest that C elegans is unable to discriminate be-tween H2S and HCN leading them to activate the degrada-tion pathways of both molecules when either is presentedThus C elegans treated with RAH2S or RAHCN increases
Figure 8 Proposed mechanismBoth H2S and HCN can inhibitelectron transport Electron trans-port inhibition causes inductionof HIF-1 activity resulting in high ex-pression of SQRD-1 and CYSL-2SQRD-1 catalyzes the first step inthe H2S-oxidation pathway ulti-mately resulting in production ofsulfate and thiosulfate CYSL-2catalyzes the first step in the HCNassimilation pathway producingH2S The resultant H2S is detoxifiedby CYSL-1
530 M W Budde and M B Roth
both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841
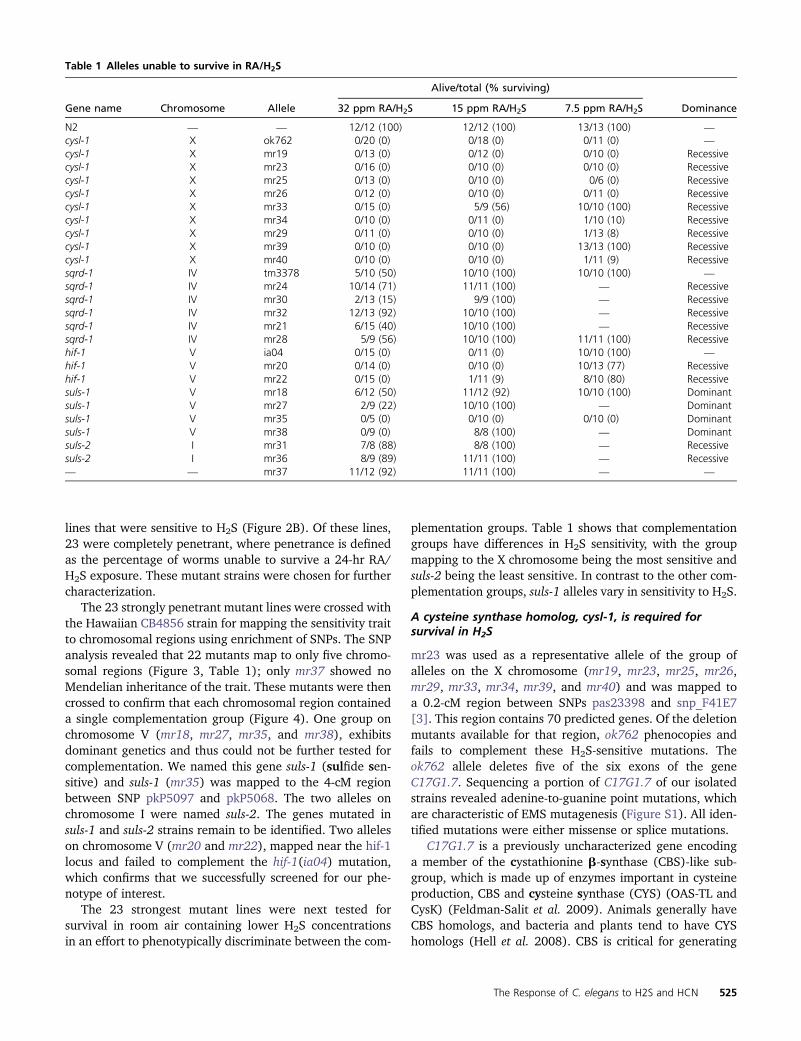
lines that were sensitive to H2S (Figure 2B) Of these lines23 were completely penetrant where penetrance is definedas the percentage of worms unable to survive a 24-hr RAH2S exposure These mutant strains were chosen for furthercharacterization
The 23 strongly penetrant mutant lines were crossed withthe Hawaiian CB4856 strain for mapping the sensitivity traitto chromosomal regions using enrichment of SNPs The SNPanalysis revealed that 22 mutants map to only five chromo-somal regions (Figure 3 Table 1) only mr37 showed noMendelian inheritance of the trait These mutants were thencrossed to confirm that each chromosomal region containeda single complementation group (Figure 4) One group onchromosome V (mr18 mr27 mr35 and mr38) exhibitsdominant genetics and thus could not be further tested forcomplementation We named this gene suls-1 (sulfide sen-sitive) and suls-1 (mr35) was mapped to the 4-cM regionbetween SNP pkP5097 and pkP5068 The two alleles onchromosome I were named suls-2 The genes mutated insuls-1 and suls-2 strains remain to be identified Two alleleson chromosome V (mr20 and mr22) mapped near the hif-1locus and failed to complement the hif-1(ia04) mutationwhich confirms that we successfully screened for our phe-notype of interest
The 23 strongest mutant lines were next tested forsurvival in room air containing lower H2S concentrationsin an effort to phenotypically discriminate between the com-
plementation groups Table 1 shows that complementationgroups have differences in H2S sensitivity with the groupmapping to the X chromosome being the most sensitive andsuls-2 being the least sensitive In contrast to the other com-plementation groups suls-1 alleles vary in sensitivity to H2S
A cysteine synthase homolog cysl-1 is required forsurvival in H2S
mr23 was used as a representative allele of the group ofalleles on the X chromosome (mr19 mr23 mr25 mr26mr29 mr33 mr34 mr39 and mr40) and was mapped toa 02-cM region between SNPs pas23398 and snp_F41E7[3] This region contains 70 predicted genes Of the deletionmutants available for that region ok762 phenocopies andfails to complement these H2S-sensitive mutations Theok762 allele deletes five of the six exons of the geneC17G17 Sequencing a portion of C17G17 of our isolatedstrains revealed adenine-to-guanine point mutations whichare characteristic of EMS mutagenesis (Figure S1) All iden-tified mutations were either missense or splice mutations
C17G17 is a previously uncharacterized gene encodinga member of the cystathionine b-synthase (CBS)-like sub-group which is made up of enzymes important in cysteineproduction CBS and cysteine synthase (CYS) (OAS-TL andCysK) (Feldman-Salit et al 2009) Animals generally haveCBS homologs and bacteria and plants tend to have CYShomologs (Hell et al 2008) CBS is critical for generating
Table 1 Alleles unable to survive in RAH2S
Alivetotal ( surviving)
Gene name Chromosome Allele 32 ppm RAH2S 15 ppm RAH2S 75 ppm RAH2S Dominance
N2 mdash mdash 1212 (100) 1212 (100) 1313 (100) mdash
cysl-1 X ok762 020 (0) 018 (0) 011 (0) mdash
cysl-1 X mr19 013 (0) 012 (0) 010 (0) Recessivecysl-1 X mr23 016 (0) 010 (0) 010 (0) Recessivecysl-1 X mr25 013 (0) 010 (0) 06 (0) Recessivecysl-1 X mr26 012 (0) 010 (0) 011 (0) Recessivecysl-1 X mr33 015 (0) 59 (56) 1010 (100) Recessivecysl-1 X mr34 010 (0) 011 (0) 110 (10) Recessivecysl-1 X mr29 011 (0) 010 (0) 113 (8) Recessivecysl-1 X mr39 010 (0) 010 (0) 1313 (100) Recessivecysl-1 X mr40 010 (0) 010 (0) 111 (9) Recessivesqrd-1 IV tm3378 510 (50) 1010 (100) 1010 (100) mdash
sqrd-1 IV mr24 1014 (71) 1111 (100) mdash Recessivesqrd-1 IV mr30 213 (15) 99 (100) mdash Recessivesqrd-1 IV mr32 1213 (92) 1010 (100) mdash Recessivesqrd-1 IV mr21 615 (40) 1010 (100) mdash Recessivesqrd-1 IV mr28 59 (56) 1010 (100) 1111 (100) Recessivehif-1 V ia04 015 (0) 011 (0) 1010 (100) mdash
hif-1 V mr20 014 (0) 010 (0) 1013 (77) Recessivehif-1 V mr22 015 (0) 111 (9) 810 (80) Recessivesuls-1 V mr18 612 (50) 1112 (92) 1010 (100) Dominantsuls-1 V mr27 29 (22) 1010 (100) mdash Dominantsuls-1 V mr35 05 (0) 010 (0) 010 (0) Dominantsuls-1 V mr38 09 (0) 88 (100) mdash Dominantsuls-2 I mr31 78 (88) 88 (100) mdash Recessivesuls-2 I mr36 89 (89) 1111 (100) mdash Recessivemdash mdash mr37 1112 (92) 1111 (100) mdash mdash
The Response of C elegans to H2S and HCN 525
cysteine in animals which make cysteine from dietary me-thionine (Finkelstein et al 1988) CBS is also one of the onlyH2S-generating enzymes in mammals (Kabil and Banerjee2010) In plants and bacteria which are cysteine proto-trophs CYS combines H2S with O-acetylserine (OAS) togenerate cysteine (Figure 5A) (Wirtz et al 2010) Humansand mice each have one CBS-like family member In con-trast C elegans has six C17G17 K10H102 R08E52F59A79 and F54A34 (Table 2)
C17G17 (cysl-1) K10H102 (cysl-2) R08E52 (cysl-3) andF59A79 (cysl-4) are more similar to CYS from plants andbacteria whereas ZC3731 (cbs-1) and F54A34 (cbs-2) aremore similar to CBS from animals (Figure 5C) A BLASTsearch reveals that the genus Caenorhabditis contains the onlyknown animal CYS homologs We have renamed these genescysl-1 (cysteine synthase-like) cysl-2 cysl-3 cysl-4 cbs-1 andcbs-2 to reflect this homology Depletion of H2S levels as it isused as a substrate by CYS may explain why cysl-1 mutantanimals are sensitive to H2S Nematodes appear to have ac-quired the ability to assimilate H2S and cysl-1 is likely usingthis reaction to drive down internal H2S levels
cysl-2 mediates immunity to fast paralytic killingby Pseudomonas
The existence of three additional Cysl homologs in C elegansraises the possibility that these other Cysl enzymes are alsoimportant in H2S detoxification This possibility is especiallyintriguing because we have shown a hif-1ndashdependent in-crease in cysl-2 transcript levels during RAH2S exposure(Budde and Roth 2010) We obtained the available knock-out strains of cysl-2 cysl-4 and cbs-2 and tested them forRAH2S sensitivity but none showed a RAH2S sensitivityphenotype (Table 2)
In plants CYS and cyanoalanine synthase (CAS) areclosely related and have overlapping catalytic activities(Figure 5 A and B) The CAS reaction combines cysteineand HCN to produce b-cyanoalanine and H2S lowering cel-lular HCN levels (Figure 5B) Furthermore cysl-2 mRNA waspreviously shown to be upregulated by HIF-1 (Shen et al2005 Shao et al 2010 Budde and Roth 2010) and highHIF-1 activity has been shown to create resistance to HCN-mediated fast paralytic killing by P aeruginosa (Darby et al1999 Gallagher and Manoil 2001 Shao et al 2010) There-fore cysl-2 may be induced not to effect H2S levels but in-stead may be the mediator of resistance to HCN-dependentfast paralytic killing To test this hypothesis we exposed cysl-1 cysl-2 and cysl-4 mutant animals to RAHCN and foundthat both cysl-1 and cysl-2 mutant animals are sensitive toRAHCN (Table 2) The specificity of cysl-2 mutant sensitivityto cyanide suggests that this enzyme is directly acting to di-minish HCN levels rather than to affect H2S levels The sen-sitivity of cysl-1 mutants to both RAH2S and RAHCNsuggest that cysl-1 acts downstream of cysl-2 and cysl-1mutants are being overwhelmed by the H2S produced bycysl-2 during HCN detoxification (Figure 8)
Next we tested whether these enzymes are responsiblefor HIF-1ndashmediated innate immunity to P aeruginosa Weshowed that similar to previous studies (Gallagher andManoil 2001 Shao et al 2010) egl-9ndashdeficient animals haveincreased innate immunity to P aeruginosa (Table 3) Nextwe demonstrated that cysl-1 and cysl-2 are both requiredfor the observed innate immunity to P aeruginosa demon-strating a mechanism of innate immunity to Pseudomonas-dependent fast paralytic killing Since we previously showedthat animals grown in RAH2S have increased cysl-2 mRNAexpression we assayed whether growth in H2S confers
Figure 4 Complementation datashowing that each genetic locusconsists of only one complemen-tation group (blue green yellowand pink shades) suls-1 is notshown because it is dominantsqrd-1 cysl-1 double heterozy-gotes are slightly sensitive toRAH2S
526 M W Budde and M B Roth
resistance to P aeruginosa Indeed our results show thatgrowth in H2S increases the innate immunity of C elegansto P aeruginosa (Table 3)
sqrd-1 is required for survival in H2S
The group of alleles on chromosome IV mr21 mr24 mr28mr30 and mr32 were mapped near the SNP pkP4095 lo-cated at the genetic location IV12 Analysis of the regionrevealed a gene F02H65 encoding an apparent homolog ofthe protein sulfidequinone reductase dehydrogenase (SQRD)which we have named sqrd-1 to reflect this homology SQRDwas initially described in prokaryotes as oxidizing sulfideand transferring the electrons to ubiquinone for generationof ATP during oxidative phosphorylation (Vande Weghe andOw 1999) The available deletion allele of sqrd-1 (tm3378)phenocopies and fails to complement these mutants Se-
quence analysis revealed A-to-G transitions in our isolatedmutants (Figure S1B)
The C elegans gene sqrd-1 encodes a member of thegroup II SQRD enzymes which also includes human mouseand Drosophila homologs The C elegans genome is predictedto encode another SQRD homolog Y9C9A16 (sqrd-2)which was initially classified as a pseudogene We obtainedthe deletion allele of sqrd-2 and found that it is not sensitiveto RAH2S
Transcriptional regulation of identified genes
We undertook this screen to determine the mechanism bywhich HIF-1 confers resistance to H2S Therefore we wereinterested to learn whether either cysl-1 or sqrd-1 is tran-scriptionally regulated by HIF-1 in response to RAH2S ex-posure Figure 6A shows that sqrd-1 and cysl-2 mRNA levels
Figure 5 (A) Cysteine synthase (CYS) catalyzes the formation of cysteine and acetate from OAS and H2S (B) Cyanoalanine synthase (CAS) catalyzes theformation of b-cyanoalanine and H2S from cysteine and HCN (C) Amino acid alignment of a selected region of CYS and CBS homologs C elegans Cyslgenes are homologous to CYS whereas C elegans Cbs genes are homologous to cystathionine b-synthase (CBS)
Table 2 Genes named in this article
Gene name Gene ID Allele tested 50 ppm RAH2S survival () 5 ppm RAHCN survival ()
cysl-1 C17G17 ok762 0 6 0 0 6 0cysl-2 K10H102 ok3516 100 6 0 0 6 0cysl-3 R08E52 mdash mdash mdash
cysl-4 F59A79 ok3359 92 6 11 100 6 0cbs-1 ZC3731 mdash mdash mdash
cbs-2 F54A34 ok666 100 6 0 100 6 0sqrd-1 F02H65 tm3378 0 6 0 97 6 5ethe-1 C33A127 tm4101 0 6 0 100 6 0
P 005 as compared to N2 wild-type animals
The Response of C elegans to H2S and HCN 527
increase during RAH2S and RAHCN exposure but cysl-1mRNA levels remain unchanged Furthermore hif-1(ia04)mutant animals fail to increase sqrd-1 and cysl-2 mRNA lev-els during RAH2S exposure (Figure 6B) suggesting thatupregulation of sqrd-1 by HIF-1 is a mechanism for survivingin the presence of H2S
To determine whether or not SQRD-1 protein levels in-crease during RAH2S exposure we performed a Westernblot to determine relative protein concentrations Figure 6Cshows that SQRD-1 protein levels increase during RAH2Sexposure in the parental N2 Bristol strain No SQRD-1 pro-tein accumulation is observed in the sqrd-1(tm3378) or inthe hif-1(ia04) mutant animals egl-9(sa307) mutant ani-mals have constitutively high levels of SQRD-1 proteinwhich may explain their ability to survive in 100 timeshigher RAH2S concentration than hif-1(ia04) mutantanimals
SQRD-1GFP protein is expressed during H2S exposure
Next we analyzed the expression pattern of SQRD-1 in Celegans We created a SQRD-1GFP fusion construct becausethe antibody we created was unable to stain worms by im-munofluorescence The presence of a predicted SL2 alterna-tive splice site upstream of sqrd-1 indicated that a standardpromoter fusion might not accurately recapitulate the en-dogenous expression pattern Therefore we used the recom-
bineering approach for GFP fusion construction whichincludes a 35-kb region surrounding the 45-kb SQRD-1gene RAH2S exposure causes a fluorescence increase inrecombinant animals that is most apparent in the anteriorof the worm (Figure 7) Expression appears limited to thehypodermis the body wall muscle and the anterior bulb ofthe pharynx (Figure 7C) A cross section through the wormconfirms that expression is limited to the most exposed partsof the worm (Figure 7D) It is possible that this SQRD-1expression pattern reflects a function of SQRD-1 to preventH2S from reaching the interior tissue
We conclude from these experiments that HIF-1 acts toinitiate the response of C elegans to H2S by upregulating theSQRD-1 protein levels SQRD-1 is an oxidoreductase oxidiz-ing H2S to polysulfide and reducing ubiquinone (Theissenet al 2003) The oxidation of sulfide generates polysulfide(Marcia et al 2009) which can then be further oxidized viathe H2S oxidation pathway to yield sulfate and thiosulfate(Kabil and Banerjee 2010) The decrease in H2S caused bySQRD-1 is likely to reduce internal H2S levels to a toleratedrange
A second enzyme required for the H2S oxidation pathwayis ETHE1 (Tiranti et al 2009) ETHE1 is a sulfur dioxyge-nase required in mammals for oxidation of sulfur to sulfate(Kabil and Banerjee 2010) C elegans has a single homologof Ethe1 C33A127 which we named ethe-1 We obtainedthe available ethe-1 (tm4101) deletion allele and tested forH2S sensitivity (Table 2) Similar to sqrd-1 (tm3378) ethe-1(tm4101) is sensitive to RAH2S which suggests that sqrd-1confers resistance to H2S through the sulfurndashoxidationpathway
Epistasis with egl-9
Egl-9ndashdeficient animals have been shown to be resistant toboth H2S and HCN and our data suggest that resistance ismediated by HIF-1 upregulating sqrd-1 and cysl-2 respec-tively To test this hypothesis we used RNAi to knockdownegl-9 in mutant animals which we have described as H2Sand HCN sensitive and tested them for resistance to H2S andHCN Table 4 shows that egl-9 knockdown does not mediateresistance to HCN in cysl-1 and cysl-2 mutant backgrounds
Table 3 Innate immunity to P aeroginosa requires hif-1 cysl-1and cysl-2
Strain Pretreatment AlleleSurvival on
P aeruginosa ()
N2 Control RNAi mdash 0 6 0N2 Egl-9 RNAi mdash 89 6 19hif-1 Egl-9 RNAi ia04 10 6 16egl-9 Egl-9 RNAi sa307 100 6 0cysl-1 Egl-9 RNAi ok762 4 6 7cysl-2 Egl-9 RNAi ok3516 0 6 0sqrd-1 Egl-9 RNAi tm3378 100 6 0ethe-1 Egl-9 RNAi tm4101 100 6 0N2 H2S grown mdash 83 6 29y
P 005 as compared to egl-9 knockdown in N2 animals yP 005 as comparedto untreated control RNAi N2 animals
Figure 6 (A) Both sqrd-1and cysl-2 mRNA levels increaserelative to untreated animals as measured by quantitativereverse transciptase PCR Y axis is relative mRNA concen-tration compared to room air-treated control animalsP 005 as compared to animals grown in room air(B) hif-1 is required for sqrd-1 and cysl-2 mRNA inductionwhen exposed to RAH2S or RAHCN Arrowheads indicatethat mRNA levels were observed at very low levels P 005 when comparing wild-type and hif-1mutant animals(C) Western blot shows SQRD-1 protein induction in RAH2S This induction requires hif-1 No immunoreactivity isobserved in sqrd-1 mutant animals SQRD-1 protein isabundant in egl-9 mutant animals Circles indicate proteinsize markers of 116 82 62 and 49 kDa from top tobottom The calculated molecular weight of 53 kDa cor-responds well with the observed relative migration
528 M W Budde and M B Roth
and therefore cysl-1 and cysl-2 are epistatic to egl-9 Sqrd-1and ethe-1 mutant animals with knocked down egl-9 arepartially resistant to H2S
Discussion
C elegans likely encounters H2S in the soil as a result ofbreakdown of organic sulfur (Morra and Dick 1991) andHCN produced by both bacteria (Gallagher and Manoil2001) and plants (Garciacutea et al 2010) This study has exam-ined the mechanisms that C elegans uses to respond to highlevels of H2S and HCN RAH2S and RAHCN both cause an
increase in HIF-1 activity which increases the expression ofsqrd-1 and cysl-2 to catalyze the first steps of the metabolismof H2S and HCN respectively sqrd-1 acts to oxidize H2Sultimately preventing internal H2S levels from exceedingthe maximum tolerated concentration cysl-2 acts in parallelto keep HCN levels low however equimolar amounts of H2Sare generated as a consequence of cysl-2 activity cysl-1 isrequired to reduce H2S levels that are produced by cysl-2during RAHCN exposure (Figure 8)
Here we provide direct evidence that H2S acts throughhif-1 to increase both SQRD-1 mRNA and protein expressionThis initiates the sulfidendashoxidation pathway which lowers in-ternal H2S levels (Figure 8) SQRD-1 expression appears to belocalized to the cells most readily exposed to exogenous H2S inthe worm In mammals colon epithelial cells that express highlevels of SQRDL (Ponteacuten et al 2008) are likely to do so be-cause a prominent enteric bacteria Desulfovibrio piger produ-ces high levels of H2S in the lower intestine (Loubinoux et al2002) The requirement for ethe-1 during RAH2S exposureconfirms that the sulfidendashoxidation pathway is the essentialfunction of sqrd-1 The sulfidendashoxidation pathway requires ox-ygen both to maintain a functional electron transport chain (togenerate oxidized ubiquinone) and to directly oxidize sulfurvia ethe-1 Exposure of mammals to exogenous H2S results inmany alterations in physiology including the induction of a sus-pended-animationndashlike state and improved outcome in ische-mia reperfusion injury It will be interesting to learn whetherSQRDL expression is induced upon exposure to H2S inmammals
The question remains as to why SQRD-1 is inducible atall Surrounded by decaying matter C elegans must encoun-ter areas of high H2S in its natural environment Havingconstitutively high SQRD-1 levels as in egl-9 mutant ani-mals allows survival when wild-type animals with lowerHIF-1 activity would perish However low levels of H2Sare important for biological function so perhaps nematodesare using SQRD-1 to deliberately titrate the cellular concen-tration of H2S by controlling its rate of destruction In mam-mals the H2S-generating enzymes CBS CSE and 3MPcontrol sulfide levels (Kabil and Banerjee 2010) Howeverour study suggests that mammals may also modulate cellu-lar H2S levels by regulating SQRDL-dependent destruction
In addition to furthering our understanding of the sulfideoxidation pathway this study also provides insight into two
Figure 7 SQRD-1GFP expression is induced by exposure to RAH2S (A)SQRD-1GFP is not expressed in room air (B) RAH2S at 50 ppm inducesexpression of SQRD-1GFP (C) Confocal microscopy at middot40 magnifica-tion shows that the expression is brightest in the muscle dense bodies(tailed arrows) and the hypodermis (tail-less arrow) (D) Cross-section(Z-stack) shows expression is most intense in the hypodermis
Table 4 Epistasis relationships with egl-9
15 ppm RAHCN 250 ppm RAH2S
Empty vector () egl-9 RNAi () Empty vector () egl-9 RNAi ()
N2 0 6 0 92 6 10 0 6 0 98 6 4hif-1 (ia04) 0 6 0 0 6 0 0 6 0 0 6 0egl-9 (sa307) 100 6 0 100 6 0 100 6 0 100 6 0sqrd-1 (tm3378) 0 6 0 87 6 16 0 6 0 47 6 15ethe-1 (tm4101) 0 6 0 89 6 14 0 6 0 73 6 10cysl-1 (ok762) 0 6 0 27 6 35 0 6 0 68 6 30cysl-2 (ok3516) 0 6 0 0 6 0 0 6 0 94 6 6
P 005 as compared to egl-9 RNAi knockdown in wild-type animals Data are presented as percentage of survival
The Response of C elegans to H2S and HCN 529
new metabolic activities previously unknown in animalscysteine synthase and cyanoalanine synthase cysl-1 mutantanimals are sensitive to both RAH2S and RAHCN whilecysl-2 mutant animals are only sensitive to RAHCN Thissuggests that cysl-1 is a cysteine synthase acting downstreamof cysl-2 a cyanoalanine synthase (Figure 8) We hypothe-size that cysl-1 mutant animals could be succumbing to H2Sproduced by CYSL-2 as a result of cyanide assimilation Inthis way each molecule of HCN assimilated by CYSL-2 wouldproduce one molecule of H2S In this reaction mechanismCYSL-1 might regenerate cysteine used in cyanide assimila-tion Alternatively it is possible that CYSL-1 is catalyzing theassimilation of both H2S and HCN a hypothesis supportedby in vitro plant studies (Hell et al 2002) The function ofthe two remaining cysteine synthase homologs cysl-3 andcysl-4 remains to be elucidated but they may also haveimportant CYSCAS activity during development or in spe-cific tissues
To our knowledge cysl-1 is the first reported functionalCYS homolog in animals Indeed while no orthologs appearto be present in animals outside of the genus Caenorhabditisin plants and bacteria CYS uses OAS as the H2S-acceptingsubstrate (Feldman-Salit et al 2009) Plants and bacteriagenerate OAS by acetylating serine with the enzyme serineacetyltransferase (SAT) (Wirtz et al 2010) which has nohomolog in C elegans How might C elegans be obtainingOAS As with many other amino acids C elegans is likelyobtaining OAS from its bacterial food source AlternativelyC elegans might have evolved a mechanism of generatingOAS using alternative enzymes CYS activity is essential forcysteine biosynthesis in prototrophs (Feldman-Salit et al2009) Because the critical aspect of CYS appears to be theelimination of H2S instead of the generation of cysteine
perhaps C elegans is freed from using OAS as the H2S ac-ceptor and is using an alternative H2S acceptor that does notgenerate cysteine In mammals the closest mammalian ho-molog to CYS is CBS Consistent with the idea that mammalshave CYS activity there is evidence that radiolabeled H2Scan be incorporated into cysteine in germ-free rats (Huovinenand Gustafsson 1967)
The protective effect of cysl-2 to HCN explains the pre-vious observation that egl-9 mutant animals are resistant tothe pathogenic Pseudomonas fast paralytic killing P aerugi-nosa is a clinically relevant pathogen and can produce toxiccyanide levels in the sputum of cystic fibrosis patients (Andersonet al 2010) P fluorescens is a cyanide-producing soil bacte-ria that has also been shown to exhibit cyanide-dependentfast paralytic killing effect on C elegans (Romanowski et al2011) HIF-1 activates resistance to hypoxia and this hasbeen proposed to also confer resistance to the cytochromec oxidase inhibiting effects of cyanide (Gallagher andManoil 2001 Shao et al 2010 Romanowski et al 2011)While this certainly may play some role we provide evidencehere that the resistance of egl-9 mutant animals to P aerugi-nosa requires cysl-2 likely by directly detoxifying cyanide tob-cyanoalanine (Figure 8) This reaction produces H2S whichcan be further detoxified by cysl-1 and sqrd-1
While we have demonstrated critical components of theRAH2S-induced hif-1 response the mechanism of hif-1 in-duction remains unclear The overlapping gene expressionpatterns between both H2S and HCN suggests that C eleganshas a shared response to these two molecules In fact ourstudies suggest that C elegans is unable to discriminate be-tween H2S and HCN leading them to activate the degrada-tion pathways of both molecules when either is presentedThus C elegans treated with RAH2S or RAHCN increases
Figure 8 Proposed mechanismBoth H2S and HCN can inhibitelectron transport Electron trans-port inhibition causes inductionof HIF-1 activity resulting in high ex-pression of SQRD-1 and CYSL-2SQRD-1 catalyzes the first step inthe H2S-oxidation pathway ulti-mately resulting in production ofsulfate and thiosulfate CYSL-2catalyzes the first step in the HCNassimilation pathway producingH2S The resultant H2S is detoxifiedby CYSL-1
530 M W Budde and M B Roth
both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841
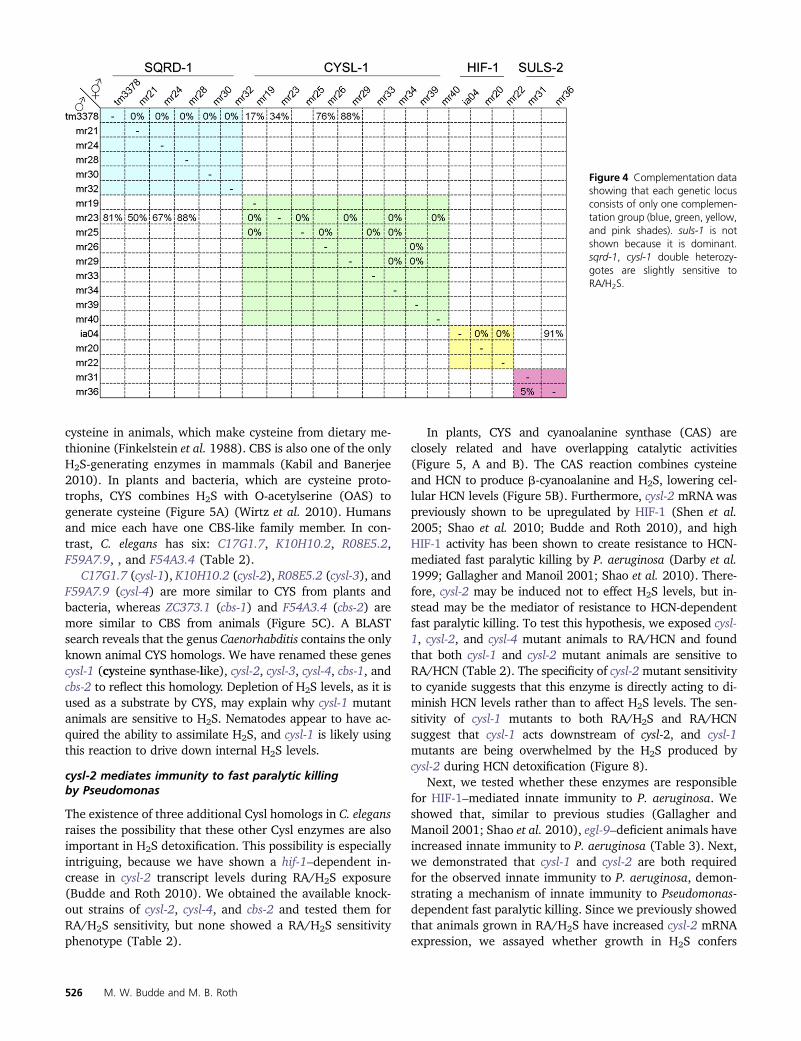
cysteine in animals which make cysteine from dietary me-thionine (Finkelstein et al 1988) CBS is also one of the onlyH2S-generating enzymes in mammals (Kabil and Banerjee2010) In plants and bacteria which are cysteine proto-trophs CYS combines H2S with O-acetylserine (OAS) togenerate cysteine (Figure 5A) (Wirtz et al 2010) Humansand mice each have one CBS-like family member In con-trast C elegans has six C17G17 K10H102 R08E52F59A79 and F54A34 (Table 2)
C17G17 (cysl-1) K10H102 (cysl-2) R08E52 (cysl-3) andF59A79 (cysl-4) are more similar to CYS from plants andbacteria whereas ZC3731 (cbs-1) and F54A34 (cbs-2) aremore similar to CBS from animals (Figure 5C) A BLASTsearch reveals that the genus Caenorhabditis contains the onlyknown animal CYS homologs We have renamed these genescysl-1 (cysteine synthase-like) cysl-2 cysl-3 cysl-4 cbs-1 andcbs-2 to reflect this homology Depletion of H2S levels as it isused as a substrate by CYS may explain why cysl-1 mutantanimals are sensitive to H2S Nematodes appear to have ac-quired the ability to assimilate H2S and cysl-1 is likely usingthis reaction to drive down internal H2S levels
cysl-2 mediates immunity to fast paralytic killingby Pseudomonas
The existence of three additional Cysl homologs in C elegansraises the possibility that these other Cysl enzymes are alsoimportant in H2S detoxification This possibility is especiallyintriguing because we have shown a hif-1ndashdependent in-crease in cysl-2 transcript levels during RAH2S exposure(Budde and Roth 2010) We obtained the available knock-out strains of cysl-2 cysl-4 and cbs-2 and tested them forRAH2S sensitivity but none showed a RAH2S sensitivityphenotype (Table 2)
In plants CYS and cyanoalanine synthase (CAS) areclosely related and have overlapping catalytic activities(Figure 5 A and B) The CAS reaction combines cysteineand HCN to produce b-cyanoalanine and H2S lowering cel-lular HCN levels (Figure 5B) Furthermore cysl-2 mRNA waspreviously shown to be upregulated by HIF-1 (Shen et al2005 Shao et al 2010 Budde and Roth 2010) and highHIF-1 activity has been shown to create resistance to HCN-mediated fast paralytic killing by P aeruginosa (Darby et al1999 Gallagher and Manoil 2001 Shao et al 2010) There-fore cysl-2 may be induced not to effect H2S levels but in-stead may be the mediator of resistance to HCN-dependentfast paralytic killing To test this hypothesis we exposed cysl-1 cysl-2 and cysl-4 mutant animals to RAHCN and foundthat both cysl-1 and cysl-2 mutant animals are sensitive toRAHCN (Table 2) The specificity of cysl-2 mutant sensitivityto cyanide suggests that this enzyme is directly acting to di-minish HCN levels rather than to affect H2S levels The sen-sitivity of cysl-1 mutants to both RAH2S and RAHCNsuggest that cysl-1 acts downstream of cysl-2 and cysl-1mutants are being overwhelmed by the H2S produced bycysl-2 during HCN detoxification (Figure 8)
Next we tested whether these enzymes are responsiblefor HIF-1ndashmediated innate immunity to P aeruginosa Weshowed that similar to previous studies (Gallagher andManoil 2001 Shao et al 2010) egl-9ndashdeficient animals haveincreased innate immunity to P aeruginosa (Table 3) Nextwe demonstrated that cysl-1 and cysl-2 are both requiredfor the observed innate immunity to P aeruginosa demon-strating a mechanism of innate immunity to Pseudomonas-dependent fast paralytic killing Since we previously showedthat animals grown in RAH2S have increased cysl-2 mRNAexpression we assayed whether growth in H2S confers
Figure 4 Complementation datashowing that each genetic locusconsists of only one complemen-tation group (blue green yellowand pink shades) suls-1 is notshown because it is dominantsqrd-1 cysl-1 double heterozy-gotes are slightly sensitive toRAH2S
526 M W Budde and M B Roth
resistance to P aeruginosa Indeed our results show thatgrowth in H2S increases the innate immunity of C elegansto P aeruginosa (Table 3)
sqrd-1 is required for survival in H2S
The group of alleles on chromosome IV mr21 mr24 mr28mr30 and mr32 were mapped near the SNP pkP4095 lo-cated at the genetic location IV12 Analysis of the regionrevealed a gene F02H65 encoding an apparent homolog ofthe protein sulfidequinone reductase dehydrogenase (SQRD)which we have named sqrd-1 to reflect this homology SQRDwas initially described in prokaryotes as oxidizing sulfideand transferring the electrons to ubiquinone for generationof ATP during oxidative phosphorylation (Vande Weghe andOw 1999) The available deletion allele of sqrd-1 (tm3378)phenocopies and fails to complement these mutants Se-
quence analysis revealed A-to-G transitions in our isolatedmutants (Figure S1B)
The C elegans gene sqrd-1 encodes a member of thegroup II SQRD enzymes which also includes human mouseand Drosophila homologs The C elegans genome is predictedto encode another SQRD homolog Y9C9A16 (sqrd-2)which was initially classified as a pseudogene We obtainedthe deletion allele of sqrd-2 and found that it is not sensitiveto RAH2S
Transcriptional regulation of identified genes
We undertook this screen to determine the mechanism bywhich HIF-1 confers resistance to H2S Therefore we wereinterested to learn whether either cysl-1 or sqrd-1 is tran-scriptionally regulated by HIF-1 in response to RAH2S ex-posure Figure 6A shows that sqrd-1 and cysl-2 mRNA levels
Figure 5 (A) Cysteine synthase (CYS) catalyzes the formation of cysteine and acetate from OAS and H2S (B) Cyanoalanine synthase (CAS) catalyzes theformation of b-cyanoalanine and H2S from cysteine and HCN (C) Amino acid alignment of a selected region of CYS and CBS homologs C elegans Cyslgenes are homologous to CYS whereas C elegans Cbs genes are homologous to cystathionine b-synthase (CBS)
Table 2 Genes named in this article
Gene name Gene ID Allele tested 50 ppm RAH2S survival () 5 ppm RAHCN survival ()
cysl-1 C17G17 ok762 0 6 0 0 6 0cysl-2 K10H102 ok3516 100 6 0 0 6 0cysl-3 R08E52 mdash mdash mdash
cysl-4 F59A79 ok3359 92 6 11 100 6 0cbs-1 ZC3731 mdash mdash mdash
cbs-2 F54A34 ok666 100 6 0 100 6 0sqrd-1 F02H65 tm3378 0 6 0 97 6 5ethe-1 C33A127 tm4101 0 6 0 100 6 0
P 005 as compared to N2 wild-type animals
The Response of C elegans to H2S and HCN 527
increase during RAH2S and RAHCN exposure but cysl-1mRNA levels remain unchanged Furthermore hif-1(ia04)mutant animals fail to increase sqrd-1 and cysl-2 mRNA lev-els during RAH2S exposure (Figure 6B) suggesting thatupregulation of sqrd-1 by HIF-1 is a mechanism for survivingin the presence of H2S
To determine whether or not SQRD-1 protein levels in-crease during RAH2S exposure we performed a Westernblot to determine relative protein concentrations Figure 6Cshows that SQRD-1 protein levels increase during RAH2Sexposure in the parental N2 Bristol strain No SQRD-1 pro-tein accumulation is observed in the sqrd-1(tm3378) or inthe hif-1(ia04) mutant animals egl-9(sa307) mutant ani-mals have constitutively high levels of SQRD-1 proteinwhich may explain their ability to survive in 100 timeshigher RAH2S concentration than hif-1(ia04) mutantanimals
SQRD-1GFP protein is expressed during H2S exposure
Next we analyzed the expression pattern of SQRD-1 in Celegans We created a SQRD-1GFP fusion construct becausethe antibody we created was unable to stain worms by im-munofluorescence The presence of a predicted SL2 alterna-tive splice site upstream of sqrd-1 indicated that a standardpromoter fusion might not accurately recapitulate the en-dogenous expression pattern Therefore we used the recom-
bineering approach for GFP fusion construction whichincludes a 35-kb region surrounding the 45-kb SQRD-1gene RAH2S exposure causes a fluorescence increase inrecombinant animals that is most apparent in the anteriorof the worm (Figure 7) Expression appears limited to thehypodermis the body wall muscle and the anterior bulb ofthe pharynx (Figure 7C) A cross section through the wormconfirms that expression is limited to the most exposed partsof the worm (Figure 7D) It is possible that this SQRD-1expression pattern reflects a function of SQRD-1 to preventH2S from reaching the interior tissue
We conclude from these experiments that HIF-1 acts toinitiate the response of C elegans to H2S by upregulating theSQRD-1 protein levels SQRD-1 is an oxidoreductase oxidiz-ing H2S to polysulfide and reducing ubiquinone (Theissenet al 2003) The oxidation of sulfide generates polysulfide(Marcia et al 2009) which can then be further oxidized viathe H2S oxidation pathway to yield sulfate and thiosulfate(Kabil and Banerjee 2010) The decrease in H2S caused bySQRD-1 is likely to reduce internal H2S levels to a toleratedrange
A second enzyme required for the H2S oxidation pathwayis ETHE1 (Tiranti et al 2009) ETHE1 is a sulfur dioxyge-nase required in mammals for oxidation of sulfur to sulfate(Kabil and Banerjee 2010) C elegans has a single homologof Ethe1 C33A127 which we named ethe-1 We obtainedthe available ethe-1 (tm4101) deletion allele and tested forH2S sensitivity (Table 2) Similar to sqrd-1 (tm3378) ethe-1(tm4101) is sensitive to RAH2S which suggests that sqrd-1confers resistance to H2S through the sulfurndashoxidationpathway
Epistasis with egl-9
Egl-9ndashdeficient animals have been shown to be resistant toboth H2S and HCN and our data suggest that resistance ismediated by HIF-1 upregulating sqrd-1 and cysl-2 respec-tively To test this hypothesis we used RNAi to knockdownegl-9 in mutant animals which we have described as H2Sand HCN sensitive and tested them for resistance to H2S andHCN Table 4 shows that egl-9 knockdown does not mediateresistance to HCN in cysl-1 and cysl-2 mutant backgrounds
Table 3 Innate immunity to P aeroginosa requires hif-1 cysl-1and cysl-2
Strain Pretreatment AlleleSurvival on
P aeruginosa ()
N2 Control RNAi mdash 0 6 0N2 Egl-9 RNAi mdash 89 6 19hif-1 Egl-9 RNAi ia04 10 6 16egl-9 Egl-9 RNAi sa307 100 6 0cysl-1 Egl-9 RNAi ok762 4 6 7cysl-2 Egl-9 RNAi ok3516 0 6 0sqrd-1 Egl-9 RNAi tm3378 100 6 0ethe-1 Egl-9 RNAi tm4101 100 6 0N2 H2S grown mdash 83 6 29y
P 005 as compared to egl-9 knockdown in N2 animals yP 005 as comparedto untreated control RNAi N2 animals
Figure 6 (A) Both sqrd-1and cysl-2 mRNA levels increaserelative to untreated animals as measured by quantitativereverse transciptase PCR Y axis is relative mRNA concen-tration compared to room air-treated control animalsP 005 as compared to animals grown in room air(B) hif-1 is required for sqrd-1 and cysl-2 mRNA inductionwhen exposed to RAH2S or RAHCN Arrowheads indicatethat mRNA levels were observed at very low levels P 005 when comparing wild-type and hif-1mutant animals(C) Western blot shows SQRD-1 protein induction in RAH2S This induction requires hif-1 No immunoreactivity isobserved in sqrd-1 mutant animals SQRD-1 protein isabundant in egl-9 mutant animals Circles indicate proteinsize markers of 116 82 62 and 49 kDa from top tobottom The calculated molecular weight of 53 kDa cor-responds well with the observed relative migration
528 M W Budde and M B Roth
and therefore cysl-1 and cysl-2 are epistatic to egl-9 Sqrd-1and ethe-1 mutant animals with knocked down egl-9 arepartially resistant to H2S
Discussion
C elegans likely encounters H2S in the soil as a result ofbreakdown of organic sulfur (Morra and Dick 1991) andHCN produced by both bacteria (Gallagher and Manoil2001) and plants (Garciacutea et al 2010) This study has exam-ined the mechanisms that C elegans uses to respond to highlevels of H2S and HCN RAH2S and RAHCN both cause an
increase in HIF-1 activity which increases the expression ofsqrd-1 and cysl-2 to catalyze the first steps of the metabolismof H2S and HCN respectively sqrd-1 acts to oxidize H2Sultimately preventing internal H2S levels from exceedingthe maximum tolerated concentration cysl-2 acts in parallelto keep HCN levels low however equimolar amounts of H2Sare generated as a consequence of cysl-2 activity cysl-1 isrequired to reduce H2S levels that are produced by cysl-2during RAHCN exposure (Figure 8)
Here we provide direct evidence that H2S acts throughhif-1 to increase both SQRD-1 mRNA and protein expressionThis initiates the sulfidendashoxidation pathway which lowers in-ternal H2S levels (Figure 8) SQRD-1 expression appears to belocalized to the cells most readily exposed to exogenous H2S inthe worm In mammals colon epithelial cells that express highlevels of SQRDL (Ponteacuten et al 2008) are likely to do so be-cause a prominent enteric bacteria Desulfovibrio piger produ-ces high levels of H2S in the lower intestine (Loubinoux et al2002) The requirement for ethe-1 during RAH2S exposureconfirms that the sulfidendashoxidation pathway is the essentialfunction of sqrd-1 The sulfidendashoxidation pathway requires ox-ygen both to maintain a functional electron transport chain (togenerate oxidized ubiquinone) and to directly oxidize sulfurvia ethe-1 Exposure of mammals to exogenous H2S results inmany alterations in physiology including the induction of a sus-pended-animationndashlike state and improved outcome in ische-mia reperfusion injury It will be interesting to learn whetherSQRDL expression is induced upon exposure to H2S inmammals
The question remains as to why SQRD-1 is inducible atall Surrounded by decaying matter C elegans must encoun-ter areas of high H2S in its natural environment Havingconstitutively high SQRD-1 levels as in egl-9 mutant ani-mals allows survival when wild-type animals with lowerHIF-1 activity would perish However low levels of H2Sare important for biological function so perhaps nematodesare using SQRD-1 to deliberately titrate the cellular concen-tration of H2S by controlling its rate of destruction In mam-mals the H2S-generating enzymes CBS CSE and 3MPcontrol sulfide levels (Kabil and Banerjee 2010) Howeverour study suggests that mammals may also modulate cellu-lar H2S levels by regulating SQRDL-dependent destruction
In addition to furthering our understanding of the sulfideoxidation pathway this study also provides insight into two
Figure 7 SQRD-1GFP expression is induced by exposure to RAH2S (A)SQRD-1GFP is not expressed in room air (B) RAH2S at 50 ppm inducesexpression of SQRD-1GFP (C) Confocal microscopy at middot40 magnifica-tion shows that the expression is brightest in the muscle dense bodies(tailed arrows) and the hypodermis (tail-less arrow) (D) Cross-section(Z-stack) shows expression is most intense in the hypodermis
Table 4 Epistasis relationships with egl-9
15 ppm RAHCN 250 ppm RAH2S
Empty vector () egl-9 RNAi () Empty vector () egl-9 RNAi ()
N2 0 6 0 92 6 10 0 6 0 98 6 4hif-1 (ia04) 0 6 0 0 6 0 0 6 0 0 6 0egl-9 (sa307) 100 6 0 100 6 0 100 6 0 100 6 0sqrd-1 (tm3378) 0 6 0 87 6 16 0 6 0 47 6 15ethe-1 (tm4101) 0 6 0 89 6 14 0 6 0 73 6 10cysl-1 (ok762) 0 6 0 27 6 35 0 6 0 68 6 30cysl-2 (ok3516) 0 6 0 0 6 0 0 6 0 94 6 6
P 005 as compared to egl-9 RNAi knockdown in wild-type animals Data are presented as percentage of survival
The Response of C elegans to H2S and HCN 529
new metabolic activities previously unknown in animalscysteine synthase and cyanoalanine synthase cysl-1 mutantanimals are sensitive to both RAH2S and RAHCN whilecysl-2 mutant animals are only sensitive to RAHCN Thissuggests that cysl-1 is a cysteine synthase acting downstreamof cysl-2 a cyanoalanine synthase (Figure 8) We hypothe-size that cysl-1 mutant animals could be succumbing to H2Sproduced by CYSL-2 as a result of cyanide assimilation Inthis way each molecule of HCN assimilated by CYSL-2 wouldproduce one molecule of H2S In this reaction mechanismCYSL-1 might regenerate cysteine used in cyanide assimila-tion Alternatively it is possible that CYSL-1 is catalyzing theassimilation of both H2S and HCN a hypothesis supportedby in vitro plant studies (Hell et al 2002) The function ofthe two remaining cysteine synthase homologs cysl-3 andcysl-4 remains to be elucidated but they may also haveimportant CYSCAS activity during development or in spe-cific tissues
To our knowledge cysl-1 is the first reported functionalCYS homolog in animals Indeed while no orthologs appearto be present in animals outside of the genus Caenorhabditisin plants and bacteria CYS uses OAS as the H2S-acceptingsubstrate (Feldman-Salit et al 2009) Plants and bacteriagenerate OAS by acetylating serine with the enzyme serineacetyltransferase (SAT) (Wirtz et al 2010) which has nohomolog in C elegans How might C elegans be obtainingOAS As with many other amino acids C elegans is likelyobtaining OAS from its bacterial food source AlternativelyC elegans might have evolved a mechanism of generatingOAS using alternative enzymes CYS activity is essential forcysteine biosynthesis in prototrophs (Feldman-Salit et al2009) Because the critical aspect of CYS appears to be theelimination of H2S instead of the generation of cysteine
perhaps C elegans is freed from using OAS as the H2S ac-ceptor and is using an alternative H2S acceptor that does notgenerate cysteine In mammals the closest mammalian ho-molog to CYS is CBS Consistent with the idea that mammalshave CYS activity there is evidence that radiolabeled H2Scan be incorporated into cysteine in germ-free rats (Huovinenand Gustafsson 1967)
The protective effect of cysl-2 to HCN explains the pre-vious observation that egl-9 mutant animals are resistant tothe pathogenic Pseudomonas fast paralytic killing P aerugi-nosa is a clinically relevant pathogen and can produce toxiccyanide levels in the sputum of cystic fibrosis patients (Andersonet al 2010) P fluorescens is a cyanide-producing soil bacte-ria that has also been shown to exhibit cyanide-dependentfast paralytic killing effect on C elegans (Romanowski et al2011) HIF-1 activates resistance to hypoxia and this hasbeen proposed to also confer resistance to the cytochromec oxidase inhibiting effects of cyanide (Gallagher andManoil 2001 Shao et al 2010 Romanowski et al 2011)While this certainly may play some role we provide evidencehere that the resistance of egl-9 mutant animals to P aerugi-nosa requires cysl-2 likely by directly detoxifying cyanide tob-cyanoalanine (Figure 8) This reaction produces H2S whichcan be further detoxified by cysl-1 and sqrd-1
While we have demonstrated critical components of theRAH2S-induced hif-1 response the mechanism of hif-1 in-duction remains unclear The overlapping gene expressionpatterns between both H2S and HCN suggests that C eleganshas a shared response to these two molecules In fact ourstudies suggest that C elegans is unable to discriminate be-tween H2S and HCN leading them to activate the degrada-tion pathways of both molecules when either is presentedThus C elegans treated with RAH2S or RAHCN increases
Figure 8 Proposed mechanismBoth H2S and HCN can inhibitelectron transport Electron trans-port inhibition causes inductionof HIF-1 activity resulting in high ex-pression of SQRD-1 and CYSL-2SQRD-1 catalyzes the first step inthe H2S-oxidation pathway ulti-mately resulting in production ofsulfate and thiosulfate CYSL-2catalyzes the first step in the HCNassimilation pathway producingH2S The resultant H2S is detoxifiedby CYSL-1
530 M W Budde and M B Roth
both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841

resistance to P aeruginosa Indeed our results show thatgrowth in H2S increases the innate immunity of C elegansto P aeruginosa (Table 3)
sqrd-1 is required for survival in H2S
The group of alleles on chromosome IV mr21 mr24 mr28mr30 and mr32 were mapped near the SNP pkP4095 lo-cated at the genetic location IV12 Analysis of the regionrevealed a gene F02H65 encoding an apparent homolog ofthe protein sulfidequinone reductase dehydrogenase (SQRD)which we have named sqrd-1 to reflect this homology SQRDwas initially described in prokaryotes as oxidizing sulfideand transferring the electrons to ubiquinone for generationof ATP during oxidative phosphorylation (Vande Weghe andOw 1999) The available deletion allele of sqrd-1 (tm3378)phenocopies and fails to complement these mutants Se-
quence analysis revealed A-to-G transitions in our isolatedmutants (Figure S1B)
The C elegans gene sqrd-1 encodes a member of thegroup II SQRD enzymes which also includes human mouseand Drosophila homologs The C elegans genome is predictedto encode another SQRD homolog Y9C9A16 (sqrd-2)which was initially classified as a pseudogene We obtainedthe deletion allele of sqrd-2 and found that it is not sensitiveto RAH2S
Transcriptional regulation of identified genes
We undertook this screen to determine the mechanism bywhich HIF-1 confers resistance to H2S Therefore we wereinterested to learn whether either cysl-1 or sqrd-1 is tran-scriptionally regulated by HIF-1 in response to RAH2S ex-posure Figure 6A shows that sqrd-1 and cysl-2 mRNA levels
Figure 5 (A) Cysteine synthase (CYS) catalyzes the formation of cysteine and acetate from OAS and H2S (B) Cyanoalanine synthase (CAS) catalyzes theformation of b-cyanoalanine and H2S from cysteine and HCN (C) Amino acid alignment of a selected region of CYS and CBS homologs C elegans Cyslgenes are homologous to CYS whereas C elegans Cbs genes are homologous to cystathionine b-synthase (CBS)
Table 2 Genes named in this article
Gene name Gene ID Allele tested 50 ppm RAH2S survival () 5 ppm RAHCN survival ()
cysl-1 C17G17 ok762 0 6 0 0 6 0cysl-2 K10H102 ok3516 100 6 0 0 6 0cysl-3 R08E52 mdash mdash mdash
cysl-4 F59A79 ok3359 92 6 11 100 6 0cbs-1 ZC3731 mdash mdash mdash
cbs-2 F54A34 ok666 100 6 0 100 6 0sqrd-1 F02H65 tm3378 0 6 0 97 6 5ethe-1 C33A127 tm4101 0 6 0 100 6 0
P 005 as compared to N2 wild-type animals
The Response of C elegans to H2S and HCN 527
increase during RAH2S and RAHCN exposure but cysl-1mRNA levels remain unchanged Furthermore hif-1(ia04)mutant animals fail to increase sqrd-1 and cysl-2 mRNA lev-els during RAH2S exposure (Figure 6B) suggesting thatupregulation of sqrd-1 by HIF-1 is a mechanism for survivingin the presence of H2S
To determine whether or not SQRD-1 protein levels in-crease during RAH2S exposure we performed a Westernblot to determine relative protein concentrations Figure 6Cshows that SQRD-1 protein levels increase during RAH2Sexposure in the parental N2 Bristol strain No SQRD-1 pro-tein accumulation is observed in the sqrd-1(tm3378) or inthe hif-1(ia04) mutant animals egl-9(sa307) mutant ani-mals have constitutively high levels of SQRD-1 proteinwhich may explain their ability to survive in 100 timeshigher RAH2S concentration than hif-1(ia04) mutantanimals
SQRD-1GFP protein is expressed during H2S exposure
Next we analyzed the expression pattern of SQRD-1 in Celegans We created a SQRD-1GFP fusion construct becausethe antibody we created was unable to stain worms by im-munofluorescence The presence of a predicted SL2 alterna-tive splice site upstream of sqrd-1 indicated that a standardpromoter fusion might not accurately recapitulate the en-dogenous expression pattern Therefore we used the recom-
bineering approach for GFP fusion construction whichincludes a 35-kb region surrounding the 45-kb SQRD-1gene RAH2S exposure causes a fluorescence increase inrecombinant animals that is most apparent in the anteriorof the worm (Figure 7) Expression appears limited to thehypodermis the body wall muscle and the anterior bulb ofthe pharynx (Figure 7C) A cross section through the wormconfirms that expression is limited to the most exposed partsof the worm (Figure 7D) It is possible that this SQRD-1expression pattern reflects a function of SQRD-1 to preventH2S from reaching the interior tissue
We conclude from these experiments that HIF-1 acts toinitiate the response of C elegans to H2S by upregulating theSQRD-1 protein levels SQRD-1 is an oxidoreductase oxidiz-ing H2S to polysulfide and reducing ubiquinone (Theissenet al 2003) The oxidation of sulfide generates polysulfide(Marcia et al 2009) which can then be further oxidized viathe H2S oxidation pathway to yield sulfate and thiosulfate(Kabil and Banerjee 2010) The decrease in H2S caused bySQRD-1 is likely to reduce internal H2S levels to a toleratedrange
A second enzyme required for the H2S oxidation pathwayis ETHE1 (Tiranti et al 2009) ETHE1 is a sulfur dioxyge-nase required in mammals for oxidation of sulfur to sulfate(Kabil and Banerjee 2010) C elegans has a single homologof Ethe1 C33A127 which we named ethe-1 We obtainedthe available ethe-1 (tm4101) deletion allele and tested forH2S sensitivity (Table 2) Similar to sqrd-1 (tm3378) ethe-1(tm4101) is sensitive to RAH2S which suggests that sqrd-1confers resistance to H2S through the sulfurndashoxidationpathway
Epistasis with egl-9
Egl-9ndashdeficient animals have been shown to be resistant toboth H2S and HCN and our data suggest that resistance ismediated by HIF-1 upregulating sqrd-1 and cysl-2 respec-tively To test this hypothesis we used RNAi to knockdownegl-9 in mutant animals which we have described as H2Sand HCN sensitive and tested them for resistance to H2S andHCN Table 4 shows that egl-9 knockdown does not mediateresistance to HCN in cysl-1 and cysl-2 mutant backgrounds
Table 3 Innate immunity to P aeroginosa requires hif-1 cysl-1and cysl-2
Strain Pretreatment AlleleSurvival on
P aeruginosa ()
N2 Control RNAi mdash 0 6 0N2 Egl-9 RNAi mdash 89 6 19hif-1 Egl-9 RNAi ia04 10 6 16egl-9 Egl-9 RNAi sa307 100 6 0cysl-1 Egl-9 RNAi ok762 4 6 7cysl-2 Egl-9 RNAi ok3516 0 6 0sqrd-1 Egl-9 RNAi tm3378 100 6 0ethe-1 Egl-9 RNAi tm4101 100 6 0N2 H2S grown mdash 83 6 29y
P 005 as compared to egl-9 knockdown in N2 animals yP 005 as comparedto untreated control RNAi N2 animals
Figure 6 (A) Both sqrd-1and cysl-2 mRNA levels increaserelative to untreated animals as measured by quantitativereverse transciptase PCR Y axis is relative mRNA concen-tration compared to room air-treated control animalsP 005 as compared to animals grown in room air(B) hif-1 is required for sqrd-1 and cysl-2 mRNA inductionwhen exposed to RAH2S or RAHCN Arrowheads indicatethat mRNA levels were observed at very low levels P 005 when comparing wild-type and hif-1mutant animals(C) Western blot shows SQRD-1 protein induction in RAH2S This induction requires hif-1 No immunoreactivity isobserved in sqrd-1 mutant animals SQRD-1 protein isabundant in egl-9 mutant animals Circles indicate proteinsize markers of 116 82 62 and 49 kDa from top tobottom The calculated molecular weight of 53 kDa cor-responds well with the observed relative migration
528 M W Budde and M B Roth
and therefore cysl-1 and cysl-2 are epistatic to egl-9 Sqrd-1and ethe-1 mutant animals with knocked down egl-9 arepartially resistant to H2S
Discussion
C elegans likely encounters H2S in the soil as a result ofbreakdown of organic sulfur (Morra and Dick 1991) andHCN produced by both bacteria (Gallagher and Manoil2001) and plants (Garciacutea et al 2010) This study has exam-ined the mechanisms that C elegans uses to respond to highlevels of H2S and HCN RAH2S and RAHCN both cause an
increase in HIF-1 activity which increases the expression ofsqrd-1 and cysl-2 to catalyze the first steps of the metabolismof H2S and HCN respectively sqrd-1 acts to oxidize H2Sultimately preventing internal H2S levels from exceedingthe maximum tolerated concentration cysl-2 acts in parallelto keep HCN levels low however equimolar amounts of H2Sare generated as a consequence of cysl-2 activity cysl-1 isrequired to reduce H2S levels that are produced by cysl-2during RAHCN exposure (Figure 8)
Here we provide direct evidence that H2S acts throughhif-1 to increase both SQRD-1 mRNA and protein expressionThis initiates the sulfidendashoxidation pathway which lowers in-ternal H2S levels (Figure 8) SQRD-1 expression appears to belocalized to the cells most readily exposed to exogenous H2S inthe worm In mammals colon epithelial cells that express highlevels of SQRDL (Ponteacuten et al 2008) are likely to do so be-cause a prominent enteric bacteria Desulfovibrio piger produ-ces high levels of H2S in the lower intestine (Loubinoux et al2002) The requirement for ethe-1 during RAH2S exposureconfirms that the sulfidendashoxidation pathway is the essentialfunction of sqrd-1 The sulfidendashoxidation pathway requires ox-ygen both to maintain a functional electron transport chain (togenerate oxidized ubiquinone) and to directly oxidize sulfurvia ethe-1 Exposure of mammals to exogenous H2S results inmany alterations in physiology including the induction of a sus-pended-animationndashlike state and improved outcome in ische-mia reperfusion injury It will be interesting to learn whetherSQRDL expression is induced upon exposure to H2S inmammals
The question remains as to why SQRD-1 is inducible atall Surrounded by decaying matter C elegans must encoun-ter areas of high H2S in its natural environment Havingconstitutively high SQRD-1 levels as in egl-9 mutant ani-mals allows survival when wild-type animals with lowerHIF-1 activity would perish However low levels of H2Sare important for biological function so perhaps nematodesare using SQRD-1 to deliberately titrate the cellular concen-tration of H2S by controlling its rate of destruction In mam-mals the H2S-generating enzymes CBS CSE and 3MPcontrol sulfide levels (Kabil and Banerjee 2010) Howeverour study suggests that mammals may also modulate cellu-lar H2S levels by regulating SQRDL-dependent destruction
In addition to furthering our understanding of the sulfideoxidation pathway this study also provides insight into two
Figure 7 SQRD-1GFP expression is induced by exposure to RAH2S (A)SQRD-1GFP is not expressed in room air (B) RAH2S at 50 ppm inducesexpression of SQRD-1GFP (C) Confocal microscopy at middot40 magnifica-tion shows that the expression is brightest in the muscle dense bodies(tailed arrows) and the hypodermis (tail-less arrow) (D) Cross-section(Z-stack) shows expression is most intense in the hypodermis
Table 4 Epistasis relationships with egl-9
15 ppm RAHCN 250 ppm RAH2S
Empty vector () egl-9 RNAi () Empty vector () egl-9 RNAi ()
N2 0 6 0 92 6 10 0 6 0 98 6 4hif-1 (ia04) 0 6 0 0 6 0 0 6 0 0 6 0egl-9 (sa307) 100 6 0 100 6 0 100 6 0 100 6 0sqrd-1 (tm3378) 0 6 0 87 6 16 0 6 0 47 6 15ethe-1 (tm4101) 0 6 0 89 6 14 0 6 0 73 6 10cysl-1 (ok762) 0 6 0 27 6 35 0 6 0 68 6 30cysl-2 (ok3516) 0 6 0 0 6 0 0 6 0 94 6 6
P 005 as compared to egl-9 RNAi knockdown in wild-type animals Data are presented as percentage of survival
The Response of C elegans to H2S and HCN 529
new metabolic activities previously unknown in animalscysteine synthase and cyanoalanine synthase cysl-1 mutantanimals are sensitive to both RAH2S and RAHCN whilecysl-2 mutant animals are only sensitive to RAHCN Thissuggests that cysl-1 is a cysteine synthase acting downstreamof cysl-2 a cyanoalanine synthase (Figure 8) We hypothe-size that cysl-1 mutant animals could be succumbing to H2Sproduced by CYSL-2 as a result of cyanide assimilation Inthis way each molecule of HCN assimilated by CYSL-2 wouldproduce one molecule of H2S In this reaction mechanismCYSL-1 might regenerate cysteine used in cyanide assimila-tion Alternatively it is possible that CYSL-1 is catalyzing theassimilation of both H2S and HCN a hypothesis supportedby in vitro plant studies (Hell et al 2002) The function ofthe two remaining cysteine synthase homologs cysl-3 andcysl-4 remains to be elucidated but they may also haveimportant CYSCAS activity during development or in spe-cific tissues
To our knowledge cysl-1 is the first reported functionalCYS homolog in animals Indeed while no orthologs appearto be present in animals outside of the genus Caenorhabditisin plants and bacteria CYS uses OAS as the H2S-acceptingsubstrate (Feldman-Salit et al 2009) Plants and bacteriagenerate OAS by acetylating serine with the enzyme serineacetyltransferase (SAT) (Wirtz et al 2010) which has nohomolog in C elegans How might C elegans be obtainingOAS As with many other amino acids C elegans is likelyobtaining OAS from its bacterial food source AlternativelyC elegans might have evolved a mechanism of generatingOAS using alternative enzymes CYS activity is essential forcysteine biosynthesis in prototrophs (Feldman-Salit et al2009) Because the critical aspect of CYS appears to be theelimination of H2S instead of the generation of cysteine
perhaps C elegans is freed from using OAS as the H2S ac-ceptor and is using an alternative H2S acceptor that does notgenerate cysteine In mammals the closest mammalian ho-molog to CYS is CBS Consistent with the idea that mammalshave CYS activity there is evidence that radiolabeled H2Scan be incorporated into cysteine in germ-free rats (Huovinenand Gustafsson 1967)
The protective effect of cysl-2 to HCN explains the pre-vious observation that egl-9 mutant animals are resistant tothe pathogenic Pseudomonas fast paralytic killing P aerugi-nosa is a clinically relevant pathogen and can produce toxiccyanide levels in the sputum of cystic fibrosis patients (Andersonet al 2010) P fluorescens is a cyanide-producing soil bacte-ria that has also been shown to exhibit cyanide-dependentfast paralytic killing effect on C elegans (Romanowski et al2011) HIF-1 activates resistance to hypoxia and this hasbeen proposed to also confer resistance to the cytochromec oxidase inhibiting effects of cyanide (Gallagher andManoil 2001 Shao et al 2010 Romanowski et al 2011)While this certainly may play some role we provide evidencehere that the resistance of egl-9 mutant animals to P aerugi-nosa requires cysl-2 likely by directly detoxifying cyanide tob-cyanoalanine (Figure 8) This reaction produces H2S whichcan be further detoxified by cysl-1 and sqrd-1
While we have demonstrated critical components of theRAH2S-induced hif-1 response the mechanism of hif-1 in-duction remains unclear The overlapping gene expressionpatterns between both H2S and HCN suggests that C eleganshas a shared response to these two molecules In fact ourstudies suggest that C elegans is unable to discriminate be-tween H2S and HCN leading them to activate the degrada-tion pathways of both molecules when either is presentedThus C elegans treated with RAH2S or RAHCN increases
Figure 8 Proposed mechanismBoth H2S and HCN can inhibitelectron transport Electron trans-port inhibition causes inductionof HIF-1 activity resulting in high ex-pression of SQRD-1 and CYSL-2SQRD-1 catalyzes the first step inthe H2S-oxidation pathway ulti-mately resulting in production ofsulfate and thiosulfate CYSL-2catalyzes the first step in the HCNassimilation pathway producingH2S The resultant H2S is detoxifiedby CYSL-1
530 M W Budde and M B Roth
both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841
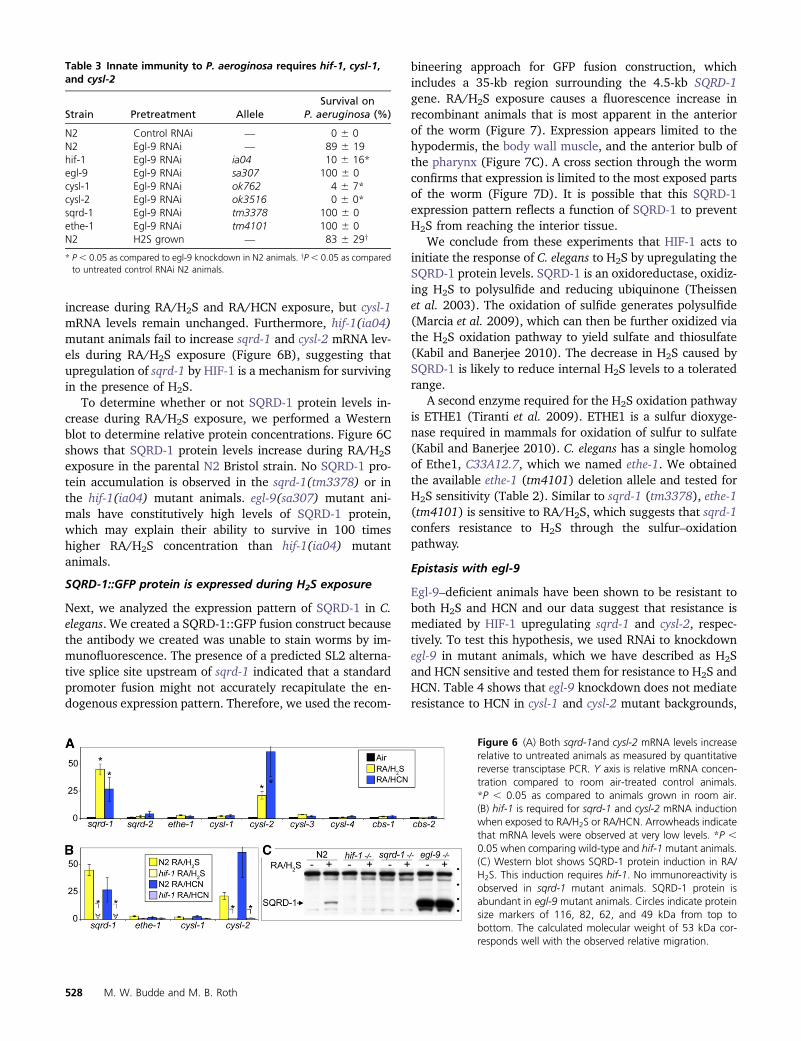
increase during RAH2S and RAHCN exposure but cysl-1mRNA levels remain unchanged Furthermore hif-1(ia04)mutant animals fail to increase sqrd-1 and cysl-2 mRNA lev-els during RAH2S exposure (Figure 6B) suggesting thatupregulation of sqrd-1 by HIF-1 is a mechanism for survivingin the presence of H2S
To determine whether or not SQRD-1 protein levels in-crease during RAH2S exposure we performed a Westernblot to determine relative protein concentrations Figure 6Cshows that SQRD-1 protein levels increase during RAH2Sexposure in the parental N2 Bristol strain No SQRD-1 pro-tein accumulation is observed in the sqrd-1(tm3378) or inthe hif-1(ia04) mutant animals egl-9(sa307) mutant ani-mals have constitutively high levels of SQRD-1 proteinwhich may explain their ability to survive in 100 timeshigher RAH2S concentration than hif-1(ia04) mutantanimals
SQRD-1GFP protein is expressed during H2S exposure
Next we analyzed the expression pattern of SQRD-1 in Celegans We created a SQRD-1GFP fusion construct becausethe antibody we created was unable to stain worms by im-munofluorescence The presence of a predicted SL2 alterna-tive splice site upstream of sqrd-1 indicated that a standardpromoter fusion might not accurately recapitulate the en-dogenous expression pattern Therefore we used the recom-
bineering approach for GFP fusion construction whichincludes a 35-kb region surrounding the 45-kb SQRD-1gene RAH2S exposure causes a fluorescence increase inrecombinant animals that is most apparent in the anteriorof the worm (Figure 7) Expression appears limited to thehypodermis the body wall muscle and the anterior bulb ofthe pharynx (Figure 7C) A cross section through the wormconfirms that expression is limited to the most exposed partsof the worm (Figure 7D) It is possible that this SQRD-1expression pattern reflects a function of SQRD-1 to preventH2S from reaching the interior tissue
We conclude from these experiments that HIF-1 acts toinitiate the response of C elegans to H2S by upregulating theSQRD-1 protein levels SQRD-1 is an oxidoreductase oxidiz-ing H2S to polysulfide and reducing ubiquinone (Theissenet al 2003) The oxidation of sulfide generates polysulfide(Marcia et al 2009) which can then be further oxidized viathe H2S oxidation pathway to yield sulfate and thiosulfate(Kabil and Banerjee 2010) The decrease in H2S caused bySQRD-1 is likely to reduce internal H2S levels to a toleratedrange
A second enzyme required for the H2S oxidation pathwayis ETHE1 (Tiranti et al 2009) ETHE1 is a sulfur dioxyge-nase required in mammals for oxidation of sulfur to sulfate(Kabil and Banerjee 2010) C elegans has a single homologof Ethe1 C33A127 which we named ethe-1 We obtainedthe available ethe-1 (tm4101) deletion allele and tested forH2S sensitivity (Table 2) Similar to sqrd-1 (tm3378) ethe-1(tm4101) is sensitive to RAH2S which suggests that sqrd-1confers resistance to H2S through the sulfurndashoxidationpathway
Epistasis with egl-9
Egl-9ndashdeficient animals have been shown to be resistant toboth H2S and HCN and our data suggest that resistance ismediated by HIF-1 upregulating sqrd-1 and cysl-2 respec-tively To test this hypothesis we used RNAi to knockdownegl-9 in mutant animals which we have described as H2Sand HCN sensitive and tested them for resistance to H2S andHCN Table 4 shows that egl-9 knockdown does not mediateresistance to HCN in cysl-1 and cysl-2 mutant backgrounds
Table 3 Innate immunity to P aeroginosa requires hif-1 cysl-1and cysl-2
Strain Pretreatment AlleleSurvival on
P aeruginosa ()
N2 Control RNAi mdash 0 6 0N2 Egl-9 RNAi mdash 89 6 19hif-1 Egl-9 RNAi ia04 10 6 16egl-9 Egl-9 RNAi sa307 100 6 0cysl-1 Egl-9 RNAi ok762 4 6 7cysl-2 Egl-9 RNAi ok3516 0 6 0sqrd-1 Egl-9 RNAi tm3378 100 6 0ethe-1 Egl-9 RNAi tm4101 100 6 0N2 H2S grown mdash 83 6 29y
P 005 as compared to egl-9 knockdown in N2 animals yP 005 as comparedto untreated control RNAi N2 animals
Figure 6 (A) Both sqrd-1and cysl-2 mRNA levels increaserelative to untreated animals as measured by quantitativereverse transciptase PCR Y axis is relative mRNA concen-tration compared to room air-treated control animalsP 005 as compared to animals grown in room air(B) hif-1 is required for sqrd-1 and cysl-2 mRNA inductionwhen exposed to RAH2S or RAHCN Arrowheads indicatethat mRNA levels were observed at very low levels P 005 when comparing wild-type and hif-1mutant animals(C) Western blot shows SQRD-1 protein induction in RAH2S This induction requires hif-1 No immunoreactivity isobserved in sqrd-1 mutant animals SQRD-1 protein isabundant in egl-9 mutant animals Circles indicate proteinsize markers of 116 82 62 and 49 kDa from top tobottom The calculated molecular weight of 53 kDa cor-responds well with the observed relative migration
528 M W Budde and M B Roth
and therefore cysl-1 and cysl-2 are epistatic to egl-9 Sqrd-1and ethe-1 mutant animals with knocked down egl-9 arepartially resistant to H2S
Discussion
C elegans likely encounters H2S in the soil as a result ofbreakdown of organic sulfur (Morra and Dick 1991) andHCN produced by both bacteria (Gallagher and Manoil2001) and plants (Garciacutea et al 2010) This study has exam-ined the mechanisms that C elegans uses to respond to highlevels of H2S and HCN RAH2S and RAHCN both cause an
increase in HIF-1 activity which increases the expression ofsqrd-1 and cysl-2 to catalyze the first steps of the metabolismof H2S and HCN respectively sqrd-1 acts to oxidize H2Sultimately preventing internal H2S levels from exceedingthe maximum tolerated concentration cysl-2 acts in parallelto keep HCN levels low however equimolar amounts of H2Sare generated as a consequence of cysl-2 activity cysl-1 isrequired to reduce H2S levels that are produced by cysl-2during RAHCN exposure (Figure 8)
Here we provide direct evidence that H2S acts throughhif-1 to increase both SQRD-1 mRNA and protein expressionThis initiates the sulfidendashoxidation pathway which lowers in-ternal H2S levels (Figure 8) SQRD-1 expression appears to belocalized to the cells most readily exposed to exogenous H2S inthe worm In mammals colon epithelial cells that express highlevels of SQRDL (Ponteacuten et al 2008) are likely to do so be-cause a prominent enteric bacteria Desulfovibrio piger produ-ces high levels of H2S in the lower intestine (Loubinoux et al2002) The requirement for ethe-1 during RAH2S exposureconfirms that the sulfidendashoxidation pathway is the essentialfunction of sqrd-1 The sulfidendashoxidation pathway requires ox-ygen both to maintain a functional electron transport chain (togenerate oxidized ubiquinone) and to directly oxidize sulfurvia ethe-1 Exposure of mammals to exogenous H2S results inmany alterations in physiology including the induction of a sus-pended-animationndashlike state and improved outcome in ische-mia reperfusion injury It will be interesting to learn whetherSQRDL expression is induced upon exposure to H2S inmammals
The question remains as to why SQRD-1 is inducible atall Surrounded by decaying matter C elegans must encoun-ter areas of high H2S in its natural environment Havingconstitutively high SQRD-1 levels as in egl-9 mutant ani-mals allows survival when wild-type animals with lowerHIF-1 activity would perish However low levels of H2Sare important for biological function so perhaps nematodesare using SQRD-1 to deliberately titrate the cellular concen-tration of H2S by controlling its rate of destruction In mam-mals the H2S-generating enzymes CBS CSE and 3MPcontrol sulfide levels (Kabil and Banerjee 2010) Howeverour study suggests that mammals may also modulate cellu-lar H2S levels by regulating SQRDL-dependent destruction
In addition to furthering our understanding of the sulfideoxidation pathway this study also provides insight into two
Figure 7 SQRD-1GFP expression is induced by exposure to RAH2S (A)SQRD-1GFP is not expressed in room air (B) RAH2S at 50 ppm inducesexpression of SQRD-1GFP (C) Confocal microscopy at middot40 magnifica-tion shows that the expression is brightest in the muscle dense bodies(tailed arrows) and the hypodermis (tail-less arrow) (D) Cross-section(Z-stack) shows expression is most intense in the hypodermis
Table 4 Epistasis relationships with egl-9
15 ppm RAHCN 250 ppm RAH2S
Empty vector () egl-9 RNAi () Empty vector () egl-9 RNAi ()
N2 0 6 0 92 6 10 0 6 0 98 6 4hif-1 (ia04) 0 6 0 0 6 0 0 6 0 0 6 0egl-9 (sa307) 100 6 0 100 6 0 100 6 0 100 6 0sqrd-1 (tm3378) 0 6 0 87 6 16 0 6 0 47 6 15ethe-1 (tm4101) 0 6 0 89 6 14 0 6 0 73 6 10cysl-1 (ok762) 0 6 0 27 6 35 0 6 0 68 6 30cysl-2 (ok3516) 0 6 0 0 6 0 0 6 0 94 6 6
P 005 as compared to egl-9 RNAi knockdown in wild-type animals Data are presented as percentage of survival
The Response of C elegans to H2S and HCN 529
new metabolic activities previously unknown in animalscysteine synthase and cyanoalanine synthase cysl-1 mutantanimals are sensitive to both RAH2S and RAHCN whilecysl-2 mutant animals are only sensitive to RAHCN Thissuggests that cysl-1 is a cysteine synthase acting downstreamof cysl-2 a cyanoalanine synthase (Figure 8) We hypothe-size that cysl-1 mutant animals could be succumbing to H2Sproduced by CYSL-2 as a result of cyanide assimilation Inthis way each molecule of HCN assimilated by CYSL-2 wouldproduce one molecule of H2S In this reaction mechanismCYSL-1 might regenerate cysteine used in cyanide assimila-tion Alternatively it is possible that CYSL-1 is catalyzing theassimilation of both H2S and HCN a hypothesis supportedby in vitro plant studies (Hell et al 2002) The function ofthe two remaining cysteine synthase homologs cysl-3 andcysl-4 remains to be elucidated but they may also haveimportant CYSCAS activity during development or in spe-cific tissues
To our knowledge cysl-1 is the first reported functionalCYS homolog in animals Indeed while no orthologs appearto be present in animals outside of the genus Caenorhabditisin plants and bacteria CYS uses OAS as the H2S-acceptingsubstrate (Feldman-Salit et al 2009) Plants and bacteriagenerate OAS by acetylating serine with the enzyme serineacetyltransferase (SAT) (Wirtz et al 2010) which has nohomolog in C elegans How might C elegans be obtainingOAS As with many other amino acids C elegans is likelyobtaining OAS from its bacterial food source AlternativelyC elegans might have evolved a mechanism of generatingOAS using alternative enzymes CYS activity is essential forcysteine biosynthesis in prototrophs (Feldman-Salit et al2009) Because the critical aspect of CYS appears to be theelimination of H2S instead of the generation of cysteine
perhaps C elegans is freed from using OAS as the H2S ac-ceptor and is using an alternative H2S acceptor that does notgenerate cysteine In mammals the closest mammalian ho-molog to CYS is CBS Consistent with the idea that mammalshave CYS activity there is evidence that radiolabeled H2Scan be incorporated into cysteine in germ-free rats (Huovinenand Gustafsson 1967)
The protective effect of cysl-2 to HCN explains the pre-vious observation that egl-9 mutant animals are resistant tothe pathogenic Pseudomonas fast paralytic killing P aerugi-nosa is a clinically relevant pathogen and can produce toxiccyanide levels in the sputum of cystic fibrosis patients (Andersonet al 2010) P fluorescens is a cyanide-producing soil bacte-ria that has also been shown to exhibit cyanide-dependentfast paralytic killing effect on C elegans (Romanowski et al2011) HIF-1 activates resistance to hypoxia and this hasbeen proposed to also confer resistance to the cytochromec oxidase inhibiting effects of cyanide (Gallagher andManoil 2001 Shao et al 2010 Romanowski et al 2011)While this certainly may play some role we provide evidencehere that the resistance of egl-9 mutant animals to P aerugi-nosa requires cysl-2 likely by directly detoxifying cyanide tob-cyanoalanine (Figure 8) This reaction produces H2S whichcan be further detoxified by cysl-1 and sqrd-1
While we have demonstrated critical components of theRAH2S-induced hif-1 response the mechanism of hif-1 in-duction remains unclear The overlapping gene expressionpatterns between both H2S and HCN suggests that C eleganshas a shared response to these two molecules In fact ourstudies suggest that C elegans is unable to discriminate be-tween H2S and HCN leading them to activate the degrada-tion pathways of both molecules when either is presentedThus C elegans treated with RAH2S or RAHCN increases
Figure 8 Proposed mechanismBoth H2S and HCN can inhibitelectron transport Electron trans-port inhibition causes inductionof HIF-1 activity resulting in high ex-pression of SQRD-1 and CYSL-2SQRD-1 catalyzes the first step inthe H2S-oxidation pathway ulti-mately resulting in production ofsulfate and thiosulfate CYSL-2catalyzes the first step in the HCNassimilation pathway producingH2S The resultant H2S is detoxifiedby CYSL-1
530 M W Budde and M B Roth
both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841
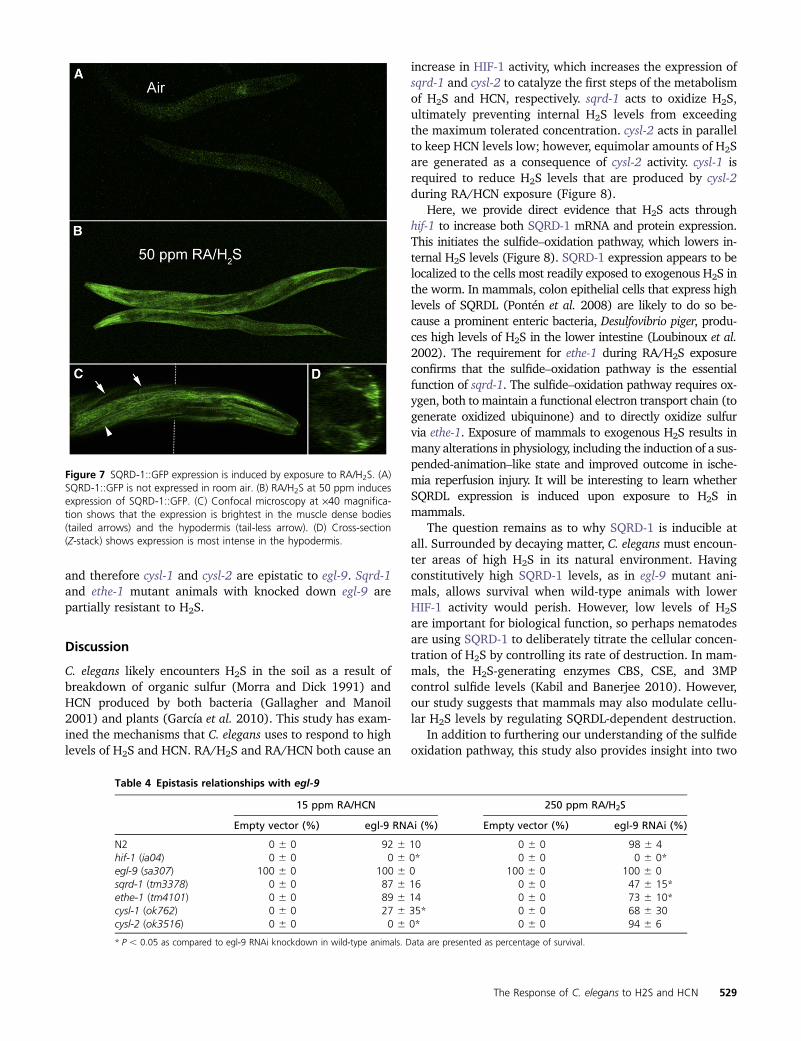
and therefore cysl-1 and cysl-2 are epistatic to egl-9 Sqrd-1and ethe-1 mutant animals with knocked down egl-9 arepartially resistant to H2S
Discussion
C elegans likely encounters H2S in the soil as a result ofbreakdown of organic sulfur (Morra and Dick 1991) andHCN produced by both bacteria (Gallagher and Manoil2001) and plants (Garciacutea et al 2010) This study has exam-ined the mechanisms that C elegans uses to respond to highlevels of H2S and HCN RAH2S and RAHCN both cause an
increase in HIF-1 activity which increases the expression ofsqrd-1 and cysl-2 to catalyze the first steps of the metabolismof H2S and HCN respectively sqrd-1 acts to oxidize H2Sultimately preventing internal H2S levels from exceedingthe maximum tolerated concentration cysl-2 acts in parallelto keep HCN levels low however equimolar amounts of H2Sare generated as a consequence of cysl-2 activity cysl-1 isrequired to reduce H2S levels that are produced by cysl-2during RAHCN exposure (Figure 8)
Here we provide direct evidence that H2S acts throughhif-1 to increase both SQRD-1 mRNA and protein expressionThis initiates the sulfidendashoxidation pathway which lowers in-ternal H2S levels (Figure 8) SQRD-1 expression appears to belocalized to the cells most readily exposed to exogenous H2S inthe worm In mammals colon epithelial cells that express highlevels of SQRDL (Ponteacuten et al 2008) are likely to do so be-cause a prominent enteric bacteria Desulfovibrio piger produ-ces high levels of H2S in the lower intestine (Loubinoux et al2002) The requirement for ethe-1 during RAH2S exposureconfirms that the sulfidendashoxidation pathway is the essentialfunction of sqrd-1 The sulfidendashoxidation pathway requires ox-ygen both to maintain a functional electron transport chain (togenerate oxidized ubiquinone) and to directly oxidize sulfurvia ethe-1 Exposure of mammals to exogenous H2S results inmany alterations in physiology including the induction of a sus-pended-animationndashlike state and improved outcome in ische-mia reperfusion injury It will be interesting to learn whetherSQRDL expression is induced upon exposure to H2S inmammals
The question remains as to why SQRD-1 is inducible atall Surrounded by decaying matter C elegans must encoun-ter areas of high H2S in its natural environment Havingconstitutively high SQRD-1 levels as in egl-9 mutant ani-mals allows survival when wild-type animals with lowerHIF-1 activity would perish However low levels of H2Sare important for biological function so perhaps nematodesare using SQRD-1 to deliberately titrate the cellular concen-tration of H2S by controlling its rate of destruction In mam-mals the H2S-generating enzymes CBS CSE and 3MPcontrol sulfide levels (Kabil and Banerjee 2010) Howeverour study suggests that mammals may also modulate cellu-lar H2S levels by regulating SQRDL-dependent destruction
In addition to furthering our understanding of the sulfideoxidation pathway this study also provides insight into two
Figure 7 SQRD-1GFP expression is induced by exposure to RAH2S (A)SQRD-1GFP is not expressed in room air (B) RAH2S at 50 ppm inducesexpression of SQRD-1GFP (C) Confocal microscopy at middot40 magnifica-tion shows that the expression is brightest in the muscle dense bodies(tailed arrows) and the hypodermis (tail-less arrow) (D) Cross-section(Z-stack) shows expression is most intense in the hypodermis
Table 4 Epistasis relationships with egl-9
15 ppm RAHCN 250 ppm RAH2S
Empty vector () egl-9 RNAi () Empty vector () egl-9 RNAi ()
N2 0 6 0 92 6 10 0 6 0 98 6 4hif-1 (ia04) 0 6 0 0 6 0 0 6 0 0 6 0egl-9 (sa307) 100 6 0 100 6 0 100 6 0 100 6 0sqrd-1 (tm3378) 0 6 0 87 6 16 0 6 0 47 6 15ethe-1 (tm4101) 0 6 0 89 6 14 0 6 0 73 6 10cysl-1 (ok762) 0 6 0 27 6 35 0 6 0 68 6 30cysl-2 (ok3516) 0 6 0 0 6 0 0 6 0 94 6 6
P 005 as compared to egl-9 RNAi knockdown in wild-type animals Data are presented as percentage of survival
The Response of C elegans to H2S and HCN 529
new metabolic activities previously unknown in animalscysteine synthase and cyanoalanine synthase cysl-1 mutantanimals are sensitive to both RAH2S and RAHCN whilecysl-2 mutant animals are only sensitive to RAHCN Thissuggests that cysl-1 is a cysteine synthase acting downstreamof cysl-2 a cyanoalanine synthase (Figure 8) We hypothe-size that cysl-1 mutant animals could be succumbing to H2Sproduced by CYSL-2 as a result of cyanide assimilation Inthis way each molecule of HCN assimilated by CYSL-2 wouldproduce one molecule of H2S In this reaction mechanismCYSL-1 might regenerate cysteine used in cyanide assimila-tion Alternatively it is possible that CYSL-1 is catalyzing theassimilation of both H2S and HCN a hypothesis supportedby in vitro plant studies (Hell et al 2002) The function ofthe two remaining cysteine synthase homologs cysl-3 andcysl-4 remains to be elucidated but they may also haveimportant CYSCAS activity during development or in spe-cific tissues
To our knowledge cysl-1 is the first reported functionalCYS homolog in animals Indeed while no orthologs appearto be present in animals outside of the genus Caenorhabditisin plants and bacteria CYS uses OAS as the H2S-acceptingsubstrate (Feldman-Salit et al 2009) Plants and bacteriagenerate OAS by acetylating serine with the enzyme serineacetyltransferase (SAT) (Wirtz et al 2010) which has nohomolog in C elegans How might C elegans be obtainingOAS As with many other amino acids C elegans is likelyobtaining OAS from its bacterial food source AlternativelyC elegans might have evolved a mechanism of generatingOAS using alternative enzymes CYS activity is essential forcysteine biosynthesis in prototrophs (Feldman-Salit et al2009) Because the critical aspect of CYS appears to be theelimination of H2S instead of the generation of cysteine
perhaps C elegans is freed from using OAS as the H2S ac-ceptor and is using an alternative H2S acceptor that does notgenerate cysteine In mammals the closest mammalian ho-molog to CYS is CBS Consistent with the idea that mammalshave CYS activity there is evidence that radiolabeled H2Scan be incorporated into cysteine in germ-free rats (Huovinenand Gustafsson 1967)
The protective effect of cysl-2 to HCN explains the pre-vious observation that egl-9 mutant animals are resistant tothe pathogenic Pseudomonas fast paralytic killing P aerugi-nosa is a clinically relevant pathogen and can produce toxiccyanide levels in the sputum of cystic fibrosis patients (Andersonet al 2010) P fluorescens is a cyanide-producing soil bacte-ria that has also been shown to exhibit cyanide-dependentfast paralytic killing effect on C elegans (Romanowski et al2011) HIF-1 activates resistance to hypoxia and this hasbeen proposed to also confer resistance to the cytochromec oxidase inhibiting effects of cyanide (Gallagher andManoil 2001 Shao et al 2010 Romanowski et al 2011)While this certainly may play some role we provide evidencehere that the resistance of egl-9 mutant animals to P aerugi-nosa requires cysl-2 likely by directly detoxifying cyanide tob-cyanoalanine (Figure 8) This reaction produces H2S whichcan be further detoxified by cysl-1 and sqrd-1
While we have demonstrated critical components of theRAH2S-induced hif-1 response the mechanism of hif-1 in-duction remains unclear The overlapping gene expressionpatterns between both H2S and HCN suggests that C eleganshas a shared response to these two molecules In fact ourstudies suggest that C elegans is unable to discriminate be-tween H2S and HCN leading them to activate the degrada-tion pathways of both molecules when either is presentedThus C elegans treated with RAH2S or RAHCN increases
Figure 8 Proposed mechanismBoth H2S and HCN can inhibitelectron transport Electron trans-port inhibition causes inductionof HIF-1 activity resulting in high ex-pression of SQRD-1 and CYSL-2SQRD-1 catalyzes the first step inthe H2S-oxidation pathway ulti-mately resulting in production ofsulfate and thiosulfate CYSL-2catalyzes the first step in the HCNassimilation pathway producingH2S The resultant H2S is detoxifiedby CYSL-1
530 M W Budde and M B Roth
both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841

new metabolic activities previously unknown in animalscysteine synthase and cyanoalanine synthase cysl-1 mutantanimals are sensitive to both RAH2S and RAHCN whilecysl-2 mutant animals are only sensitive to RAHCN Thissuggests that cysl-1 is a cysteine synthase acting downstreamof cysl-2 a cyanoalanine synthase (Figure 8) We hypothe-size that cysl-1 mutant animals could be succumbing to H2Sproduced by CYSL-2 as a result of cyanide assimilation Inthis way each molecule of HCN assimilated by CYSL-2 wouldproduce one molecule of H2S In this reaction mechanismCYSL-1 might regenerate cysteine used in cyanide assimila-tion Alternatively it is possible that CYSL-1 is catalyzing theassimilation of both H2S and HCN a hypothesis supportedby in vitro plant studies (Hell et al 2002) The function ofthe two remaining cysteine synthase homologs cysl-3 andcysl-4 remains to be elucidated but they may also haveimportant CYSCAS activity during development or in spe-cific tissues
To our knowledge cysl-1 is the first reported functionalCYS homolog in animals Indeed while no orthologs appearto be present in animals outside of the genus Caenorhabditisin plants and bacteria CYS uses OAS as the H2S-acceptingsubstrate (Feldman-Salit et al 2009) Plants and bacteriagenerate OAS by acetylating serine with the enzyme serineacetyltransferase (SAT) (Wirtz et al 2010) which has nohomolog in C elegans How might C elegans be obtainingOAS As with many other amino acids C elegans is likelyobtaining OAS from its bacterial food source AlternativelyC elegans might have evolved a mechanism of generatingOAS using alternative enzymes CYS activity is essential forcysteine biosynthesis in prototrophs (Feldman-Salit et al2009) Because the critical aspect of CYS appears to be theelimination of H2S instead of the generation of cysteine
perhaps C elegans is freed from using OAS as the H2S ac-ceptor and is using an alternative H2S acceptor that does notgenerate cysteine In mammals the closest mammalian ho-molog to CYS is CBS Consistent with the idea that mammalshave CYS activity there is evidence that radiolabeled H2Scan be incorporated into cysteine in germ-free rats (Huovinenand Gustafsson 1967)
The protective effect of cysl-2 to HCN explains the pre-vious observation that egl-9 mutant animals are resistant tothe pathogenic Pseudomonas fast paralytic killing P aerugi-nosa is a clinically relevant pathogen and can produce toxiccyanide levels in the sputum of cystic fibrosis patients (Andersonet al 2010) P fluorescens is a cyanide-producing soil bacte-ria that has also been shown to exhibit cyanide-dependentfast paralytic killing effect on C elegans (Romanowski et al2011) HIF-1 activates resistance to hypoxia and this hasbeen proposed to also confer resistance to the cytochromec oxidase inhibiting effects of cyanide (Gallagher andManoil 2001 Shao et al 2010 Romanowski et al 2011)While this certainly may play some role we provide evidencehere that the resistance of egl-9 mutant animals to P aerugi-nosa requires cysl-2 likely by directly detoxifying cyanide tob-cyanoalanine (Figure 8) This reaction produces H2S whichcan be further detoxified by cysl-1 and sqrd-1
While we have demonstrated critical components of theRAH2S-induced hif-1 response the mechanism of hif-1 in-duction remains unclear The overlapping gene expressionpatterns between both H2S and HCN suggests that C eleganshas a shared response to these two molecules In fact ourstudies suggest that C elegans is unable to discriminate be-tween H2S and HCN leading them to activate the degrada-tion pathways of both molecules when either is presentedThus C elegans treated with RAH2S or RAHCN increases
Figure 8 Proposed mechanismBoth H2S and HCN can inhibitelectron transport Electron trans-port inhibition causes inductionof HIF-1 activity resulting in high ex-pression of SQRD-1 and CYSL-2SQRD-1 catalyzes the first step inthe H2S-oxidation pathway ulti-mately resulting in production ofsulfate and thiosulfate CYSL-2catalyzes the first step in the HCNassimilation pathway producingH2S The resultant H2S is detoxifiedby CYSL-1
530 M W Budde and M B Roth
both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841

both H2S-detoxification enzymes and cyanide-detoxificationenzymes (Figure 6A) A recent study showed that inhibitionof electron transport through reduction of expression ofelectron transport proteins is sufficient to induce HIF-1 ac-tivity (Lee et al 2010) H2S and HCN are natural effectors ofelectron transport and cause increased HIF-1 activity there-fore H2S and HCN might be natural regulators of electrontransport
Acknowledgments
We thank members of the Roth lab for helpful discussions JimPriess and Sue Biggins for allowing gratuitous reagent useColin Manoil and Larry Gallagher for Pseudomonas aeruginosastrains and Shohei Mitani at Tokyo Womens Medical Univer-sity School of Medicine for providing knockout strains
Literature Cited
Anderson R D L F Roddam S Bettiol K Sanderson and D WReid 2010 Biosignificance of bacterial cyanogenesis in the CFlung J Cyst Fibros 9 158ndash164
Blackstone E and M B Roth 2007 Suspended animation-likestate protects mice from lethal hypoxia Shock 27 370ndash372
Blackstone E M Morrison and M B Roth 2005 H2S inducesa suspended animation-like state in mice Science 308 518
Brenner S 1974 The genetics of Caenorhabditis elegans Genetics77 71ndash94
Budde M W and M B Roth 2010 Hydrogen sulfide increaseshypoxia-inducible factor-1 activity independently of von Hippel-Lindau tumor suppressor-1 in C elegans Mol Biol Cell 21 212ndash217
Darby C C L Cosma J H Thomas and C Manoil 1999 Lethalparalysis of Caenorhabditis elegans by Pseudomonas aeruginosaProc Natl Acad Sci USA 96 15202ndash15207
David L A and E J Alm 2011 Rapid evolutionary innovationduring an Archaean genetic expansion Nature 469 93ndash96
Davis M W M Hammarlund T Harrach P Hullett S Olsenet al 2005 Rapid single nucleotide polymorphism mappingin C elegans BMC Genomics 6 118
Dolphin C T and I A Hope 2006 Caenorhabditis elegans re-porter fusion genes generated by seamless modification of largegenomic DNA clones Nucleic Acids Res 34 e72
Elrod J W J W Calvert J Morrison J E Doeller D W Krauset al 2007 Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial functionProc Natl Acad Sci USA 104 15560ndash15565
Feldman-Salit A M Wirtz R Hell and R C Wade 2009 Amechanistic model of the cysteine synthase complex J MolBiol 386 37ndash59
Finkelstein J D J J Martin and B J Harris 1988 Methioninemetabolism in mammals The methionine-sparing effect of cys-tine J Biol Chem 263 11750ndash11754
Frazier H N 3rd and M B Roth 2009 Adaptive sugar provi-sioning controls survival of C elegans embryos in adverse envi-ronments Curr Biol 19 859ndash863
Gallagher L A and C Manoil 2001 Pseudomonas aeruginosaPAO1 kills Caenorhabditis elegans by cyanide poisoning J Bac-teriol 183 6207ndash6214
Garciacutea I J M Castellano B Vioque R Solano C Gotor et al2010 Mitochondrial beta-cyanoalanine synthase is essential
for root hair formation in Arabidopsis thaliana Plant Cell 223268ndash3279
Hell R R Jost O Berkowitz and M Wirtz 2002 Molecular andbiochemical analysis of the enzymes of cysteine biosynthesis inthe plant Arabidopsis thaliana Amino Acids 22 245ndash257
Hell R C Dahl D B Knaff and T Leustek 2008 Sulfur Metab-olism in Phototrophic Organisms Springer-Verlag New York
Holmgren A C Johansson C Berndt M E Loumlnn C Hudemannet al 2005 Thiol redox control via thioredoxin and glutare-doxin systems Biochem Soc Trans 33 1375ndash1377
Huovinen J A and B E Gustafsson 1967 Inorganic sulphatesulphite and sulphide as sulphur donors in the biosynthesis ofsulphur amino acids in germ-free and conventional rats Bio-chim Biophys Acta 136 441ndash447
Kabil O and R Banerjee 2010 Redox biochemistry of hydrogensulfide J Biol Chem 285 21903ndash21907
Lee S-J A B Hwang and C Kenyon 2010 Inhibition of respi-ration extends C elegans life span via reactive oxygen speciesthat increase HIF-1 activity Curr Biol 20 2131ndash2136
Li L P Rose and P K Moore 2011 Hydrogen sulfide and cellsignaling Annu Rev Pharmacol Toxicol 51 169ndash187
Loubinoux J F M A Valente I A C Pereira A Costa P A DGrimont et al 2002 Reclassification of the only species of thegenus Desulfomonas Desulfomonas pigra as Desulfovibrio pigercomb nov Int J Syst Evol Microbiol 52 1305ndash1308
Marcia M U Ermler G Peng and H Michel 2009 The structureof Aquifex aeolicus sulfidequinone oxidoreductase a basis tounderstand sulfide detoxification and respiration Proc NatlAcad Sci USA 106 9625ndash9630
Mello C C J M Kramer D Stinchcomb and V Ambros1991 Efficient gene transfer in Celegans extrachromosomalmaintenance and integration of transforming sequences EMBOJ 10 3959ndash3970
Miller D L and M B Roth 2007 Hydrogen sulfide increasesthermotolerance and lifespan in Caenorhabditis elegans ProcNatl Acad Sci USA 104 20618ndash20622
Morra M J and W A Dick 1991 Mechanisms of H2S produc-tion from cysteine and cystine by microorganisms isolated fromsoil by selective enrichment Appl Environ Microbiol 571413ndash1417
Morrison M L J E Blackwood S L Lockett A Iwata R K Winnet al 2008 Surviving blood loss using hydrogen sulfide JTrauma 65 183ndash188
Mustafa A K M M Gadalla and S H Snyder 2009 Signalingby gasotransmitters Sci Signal 2 re2
Ng P C and S Henikoff 2001 Predicting deleterious amino acidsubstitutions Genome Res 11 863ndash874
Philippot P M Van Zuilen K Lepot C Thomazo J Farquharet al 2007 Early Archaean microorganisms preferred elemen-tal sulfur not sulfate Science 317 1534ndash1537
Ponteacuten F K Jirstroumlm and M Uhlen 2008 The Human ProteinAtlasndasha tool for pathology J Pathol 216 387ndash393
Reinhard C T R Raiswell C Scott A D Anbar and T W Lyons2009 A late Archean sulfidic sea stimulated by early oxidativeweathering of the continents Science 326 713ndash716
Romanowski A M L Migliori C Valverde and D A Golombek2011 Circadian variation in Pseudomonas fluorescens (CHA0)-mediated paralysis of Caenorhabditis elegans Microb Pathog50 23ndash30
Rouault T A and W H Tong 2008 Iron-sulfur cluster biogen-esis and human disease Trends Genet 24 398ndash407
Shaham S 2007 Counting mutagenized genomes and optimizinggenetic screens in Caenorhabditis elegans PLoS ONE 2 e1117
Shao Z Y Zhang Q Ye J N Saldanha and J A Powell-Coffman2010 C elegans SWAN-1 Binds to EGL-9 and regulates HIF-1-mediated resistance to the bacterial pathogen Pseudomonas aer-uginosa PAO1 PLoS Pathog 6pii e1001075
The Response of C elegans to H2S and HCN 531
Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841

Shen C D Nettleton M Jiang S K Kim and J A Powell-Coffman2005 Roles of the HIF-1 hypoxia-inducible factor during hyp-oxia response in Caenorhabditis elegans J Biol Chem 28020580ndash20588
Swierczek N A Giles A C Rankin C H Kerr R A 2011 High-throughput behavioral analysis in C elegans Nat Methods 8592ndash598
Theissen U M Hoffmeister M Grieshaber and W Martin2003 Single eubacterial origin of eukaryotic sulfide quinoneoxidoreductase a mitochondrial enzyme conserved from theearly evolution of eukaryotes during anoxic and sulfidic timesMol Biol Evol 20 1564ndash1574
Tiranti V C Viscomi T Hildebrandt I Di Meo R Mineri et al2009 Loss of ETHE1 a mitochondrial dioxygenase causes fa-tal sulfide toxicity in ethylmalonic encephalopathy Nat Med15 200ndash205
Vande Weghe J G and D W Ow 1999 A fission yeast gene formitochondrial sulfide oxidation J Biol Chem 274 13250ndash13257
Wirtz M H Birke C Heeg C Muumlller F Hosp et al2010 Structure and function of the hetero-oligomeric cysteinesynthase complex in plants J Biol Chem 285 32810ndash32817
Communicating editor D I Greenstein
532 M W Budde and M B Roth
GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841

GENETICSSupporting Information
httpwwwgeneticsorgcontentsuppl20110812genetics111129841DC1
The Response of Caenorhabditis elegansto Hydrogen Sulfide and Hydrogen Cyanide
Mark W Budde and Mark B Roth
Copyright copy 2011 by the Genetics Society of AmericaDOI 101534genetics111129841