Embed Size (px)
Citation preview

554 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, MARCH–APRIL 2009
RESEARCH
Agriculture is an integrated biological production system, in which several crops are grown in combination (variously
called polyculture, intercropping, or associated cropping), in suc-cession (crop rotation), or in a combination of both approaches. The transition from hunting–gathering to agriculture therefore not only involved the domestication of individual crops (Zeder et al., 2006) but led also to the development of cropping systems that combined several crops in space and time (and animal hus-bandry systems as well). Little is known about how cropping sys-tems appeared in relation to the domestication of individual crops, more specifi cally with regard to the geographic origin and timing of appearance of these systems. We attempt to address the ques-tion of geographic origin by inferring a presumed Mesoameri-can domestication center of common bean (Phaseolus vulgaris L.) and comparing this location with the proposed location of maize domestication in the same country. Mexico is one of the most important centers of agricultural origins in the world (Hancock, 2004; Smith, 1995). Among the important food crops that were domesticated in Mexico is the trilogy of maize (Zea mays L.), beans (Phaseolus spp., most often common bean, P. vulgaris), and squash (Cucurbita spp.). Wild relatives of these three crops are found in Mexico and adjacent regions (maize: Wilkes, 2004; beans: Freytag and Debouck, 2002; and squash: Montes-Hernandez and Egui-arte, 2002). Furthermore, both archaeological and genetic data
The Putative Mesoamerican Domestication Center of Phaseolus vulgaris Is Located in the
Lerma–Santiago Basin of Mexico
Myounghai Kwak, James A. Kami, and Paul Gepts*
ABSTRACT
Mesoamerican food agriculture is defi ned by the
milpa cropping system, consisting of maize (Zea
mays L.), common bean (Phaseolus vulgaris L.),
and squash (Cucurbita spp.). In recent years, a
domestication center for maize has been pro-
posed in the Balsas River basin in west-cen-
tral Mexico, raising the question whether the
Balsas basin was also the center of origin for
Mesoamerican food agriculture in general. We
conducted a survey of genetic diversity for 26
microsatellite markers in a representative sam-
ple of 155 wild and domesticated common bean
from its Mesoamerican gene pool. Microsatel-
lite diversity was analyzed with STRUCTURE,
neighbor-joining tree construction, and princi-
pal coordinate analysis. Most Mesoamerican
domesticated accessions clustered in a single
group, suggesting a single domestication. Fur-
thermore, the most closely related wild beans
to the domesticated clade originated from a
restricted region in the Rio Lerma–Rio Grande
de Santiago basin in west-central Mexico, dis-
tinct from the Balsas basin. Although wild maize
and Phaseolus beans grow together in the wild,
they appear to have been domesticated in dif-
ferent regions to be reunited later on in a single
cropping system. Crop domestications in Meso-
america may therefore have a diffuse rather
than a single geographic origin. Archaeological
and ecological investigations into the origins of
agriculture should be refocused from the arid
eastern half of Mexico to the west-central part
of the country.
Univ. of California, Dep. of Plant Sciences/MS1, Section of Crop and
Ecosystem Sciences, 1 Shields Ave., Davis, CA 95616-8780. Received
19 July 2008. *Corresponding author ([email protected]).
Abbreviations: NJ, neighbor-joining; PCR, polymerase chain reac-
tion; SSC, simulation similarity coeffi cient.
Published in Crop Sci. 49:554–563 (2009).doi: 10.2135/cropsci2008.07.0421© Crop Science Society of America677 S. Segoe Rd., Madison, WI 53711 USA
All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher.

CROP SCIENCE, VOL. 49, MARCH–APRIL 2009 WWW.CROPS.ORG 555
point to Mexico as a center of domestication of the three crops (Smith, 1995; Doebley, 2004; Gepts, 1988; Gepts et al., 1986; Piperno and Flannery, 2001). (For common bean, there is an additional center of domestication in the Andes [Gepts et al., 1986; Gepts 1998]; however, this cen-ter is not discussed further in this report.)
Beans, maize, and squash are often cultivated together in traditional cropping systems in Mexico (e.g., milpa cropping system) and other countries of Latin America and in eastern Africa because they complement each other agronomically (e.g., plant nitrogen balance; Cardoso et al., 2007) and nutritionally (e.g., balance of essential amino acids; Bressani, 1983). The actual area of domes-tication of these crops within Mexico—and particularly, whether these domestications can be traced back to a single region—is still the subject of both archaeologi-cal and genetic research. Maize was domesticated from teosinte (Zea mays subsp. parviglumis), its closest wild relative (Doebley, 2004), in a region presumably located in west-central Mexico around the Balsas River basin, based on genetic data (Matsuoka et al., 2002). The old-est archaeological remains of incipient maize domestica-tion have been identifi ed in the Guilá Naquitz cave to the south of the Balsas basin (Piperno and Flannery, 2001), as were 8000- to 10,000-yr-old squash (C. pepo) seeds (Smith, 1997). Given the close association of maize and beans before and after domestication (Delgado Salinas et al., 1988), these fi ndings raise the question whether the common bean was domesticated in the same general area as maize. Recent experimental developments prompted us to examine the question of the origin of domestication of common bean in its Mesoamerican center of domestica-tion using a broadly distributed range of wild populations (Freytag and Debouck, 2002) and a set of 26 putatively neutral microsatellite markers distributed over the 11 linkage groups of common bean (Blair et al., 2003).
MATERIAL AND METHODS
Microsatellite Marker AnalysisTwenty-six microsatellite markers from all 11 linkage groups
were selected on the basis of their map location (Supplemental
Table S1; Blair et al., 2003). Forward primers were designed
with a 5′-TGTAAACGACGGCCAGTATGC M-13 reverse
sequence tail added to the 5′ end of the forward primer. Genomic
DNA was extracted from young leaves using the cetyl trimethyl
ammonium bromide (CTAB) method (Doyle and Doyle, 1987).
Except for simple sequence repeat markers BM146 and BM157,
two independent polymerase chain reactions (PCRs) were per-
formed. For the primary PCR, the pairs of forward and reverse
primer were used to amplify microsatellite fragments. Thus,
the fragments amplifi ed in the primary PCR included the M13
sequence extension at forward primer site. The secondary PCR
reactions were performed with the reverse primer and the M-13
primer labeled with the 6-FAM, NED, PET, or VIC fl uorescence
dyes (Applied Biosystems, Foster City, CA). For the primary
PCR reaction, PCR reaction mixtures contained approximately
30 ng of total genomic DNA, 200 μM of dNTP, 0.2 μM of
forward primer and reverse primer, the standard Taq buff er with
1.5 mM MgCl2, and 1 unit of Taq polymerase (New England
Biololabs, Ipswich, MA) in a 20 μL total reaction volume. The
primary PCR cycle consisted of 2 min at 94°C and 35 cycles of
30 s at 94°C, 1 min at 47°C (BMd45, BMd10, BMd1, Pv-ctt001,
BMd53, BMd37, BMd12, BMd25, BMd42, and BMd41), 49°C
(Pv-ag003, BM143, BM172, Pv-ag004, BM151, Pv-at007, and
BM212), 52°C (GATS91, BM160, and BM210), 55°C (BM188),
57°C (Pv-ag001), or 60°C (BM53 and BMd20), and then 40 s
at 72°C followed by a 3-min extension at 72°C. For the second-
ary PCR reaction, the PCR reaction mixtures contained 1 μL
of primary PCR product, 0.2 μM of fl uorescence-labeled M13
universal primer and reverse primer, 0.34 μM of forward primer
and standard Taq buff er with 1.5 mM MgCl2, and 1 unit of Taq
polymerase in a total volume 20 μL reaction. For M13 primer
labeling, the 6-FAM, NED, PET, or VIC dye was attached to
the 5′ end of the 5′-TGTAAAACGACGGCCAGT-3′ M-13
universal primer sequence. The secondary PCR cycle consisted
of 2 min at 94°C and 30 cycles of 30 s at 94°C, 45 s at 56°C, and
45 s at 72°C followed by 8 cycles of 30 s at 94°C, 45 s at 53°C,
and 45 s at 72°C, and then 3 min at 72°C for the fi nal exten-
sion. For the BM146 and BM157 amplifi cation, PCR reaction
mixtures contained approximately 30 ng of total genomic DNA,
200 μM of dNTP, 0.16 μM of labeled M-13 universal primer
and reverse primer, 0.04 μM of reverse primer, standard Taq buf-
fer with 1.5 mM MgCl2, and 1 unit of Taq polymerase (New
England Biololabs) in a 20 μL total reaction volume. Polymerase
chain reaction cycles consisted of 5 min at 94°C and 30 cycles of
30 s at 94°C, 45 s at 56°C, and 45 s at 72°C followed by 8 cycles
of 30 s at 94°C, 45 c at 53°C, and 45 s at 72°C, and a fi nal exten-
sion of 3 min at 72°C. The amplifi ed fragments were multiplexed
depending on their size variation and analyzed in an ABI 3730
(Applied Biosystems). Marker genotypes were determined using
the GeneMarker program (version1.51; SoftGenetics LLC, State
College, PA) and are included in Supplemental Table S2.
Defi ning Mesoamerican Accessions Using STRUCTUREThree hundred forty-nine wild, landrace, and commercial
cultivars and advanced germplasm accessions of common bean
were analyzed in this study (Supplemental Table S3). These
accessions were obtained from the Phaseolus World Collec-
tion at CIAT, Cali, Colombia, or from the Bean Collection of
the USDA National Plant Germplasm System. Because crop
varieties can be disseminated across diff erent regions or con-
tinents, the geographic origin of individual accessions is not
a reliable indicator of the origin of domestication (Andean vs.
Mesoamerican; Gepts et al., 1986; Gepts 1998). Thus, the pro-
gram STRUCTURE (Pritchard et al., 2000) was run with a
clustering number (K) = 2 setting. Five independent simula-
tions were performed using the admixture model and 5000
replicates for burn-in and 50000 replicates during analysis. A
between-simulation similarity coeffi cient (SSC) was calculated
using the STRUCTURE sum program (Ehrich 2006), which
showed that the clustering in diff erent runs was almost identi-
cal (SSC = 0.9969). Among the fi ve simulations, the run with
the highest likelihood value was selected, and the accessions

556 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, MARCH–APRIL 2009
from at least two domestications from an already diverged ancestor, wild common bean. A common origin for these two gene pools may reside in the wild common bean of Ecuador and northern Peru (Kami et al., 1995). Because of exchanges after domestication between the Andean and the Mesoamerican gene pools (Gepts et al., 1988; Paredes and Gepts, 1995), geographic origin is an ambiguous indicator of gene pool membership. Neutral molecular markers such as microsatellites provide the opportunity to assess the gene pool origin of common bean accessions, independently of geographic information.
To identify a more specifi c region for the Meso-american domestication of common bean, we followed a two-step strategy. First, we identifi ed the presump-tive Mesoamerican accessions in a large representative sample following a model-based approach of microsatel-lite diversity, implemented in the STRUCTURE soft-ware. Next, we analyzed genetic distances among these Mesoamerican accessions by a STRUCTURE again, combined with NJ and principal coordinate analyses of microsatellite diversity.
A representative sample of 349 wild and domesti-cated accessions of common bean was developed, which comprised 100 wild and 249 domesticated types. Twenty-six microsatellite markers, distributed on the 11 linkage groups of the common bean genome, were chosen for this analysis (Blair et al., 2003; Yu et al., 2000; Gaitán-Solís et al., 2002). Genetic diversity was analyzed with a model-based approach implemented in STRUCTURE (Pritchard et al., 2000) assuming two populations (K = 2) (see “Materials and Methods”). This procedure assigned 155 accessions to the Mesoamerican gene pool, includ-ing 76 wild and 79 domesticated types, and 199 Andean gene pool materials. Further discussion of results in this article addresses only the 155 accessions assigned to the Mesoamerican gene pool. The wild-types in this Meso-american sample included 8 accessions from the ancestral populations in Ecuador and Peru, 8 from Colombia, 3 from Guatemala, and 57 from Mexico. The 79 domes-ticated entries included 2 from Bolivia, 6 from Brazil, 1 from Chile, 5 from Colombia, 1 each from Costa Rica and Ecuador, 5 from Guatemala, 3 from Honduras, 35 from Mexico, 2 from Nicaragua, 1 each from Panama and Peru, 14 from the United States, and 2 from Venezuela.
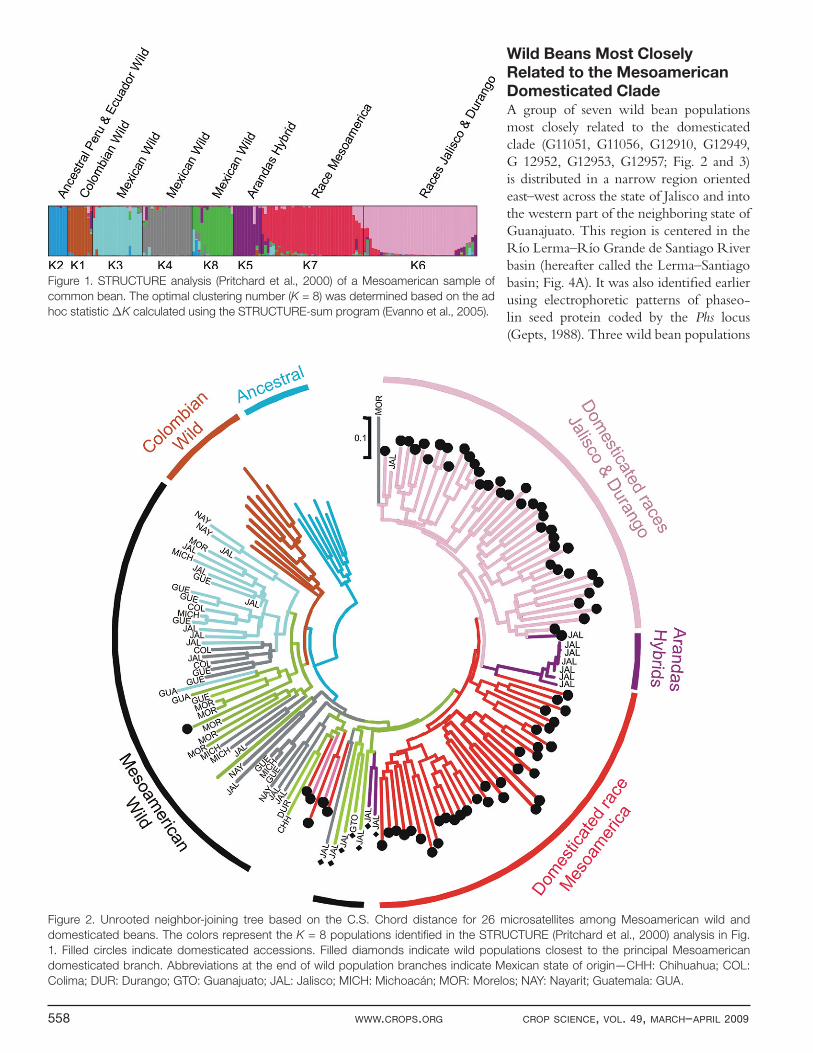
Genetic Diversity in the Mesoamerican Gene PoolThe organization of this Mesoamerican sample was fur-ther analyzed with STRUCTURE in a stepwise fashion from 2 to 1 populations (K = 2 to K = 11). The fi nal num-ber of populations of K = 8 was chosen based on the ΔK statistical test (Evanno et al., 2005) showing no further signifi cant subdivisions for larger K values. At K = 8, two domesticated groups were identifi ed, one corresponding
with more than 50% of membership coeffi cient for the Meso-
american cluster were assigned to the Mesoamerican gene pool.
Through this process, 155 accessions out of 349 were defi ned
as belonging to the Mesoamerican gene pool. Among these 155
accessions, 88% had a membership coeffi cient above 90% and
95% above 80%. The remaining 5% had membership coeffi -
cients between 50 and 80%.
Population Structure in Mesoamerican Gene PoolTo defi ne the population structure in the Mesoamerican
gene pool, the 155 accessions were further analyzed with the
STRUCTURE program (Pritchard et al., 2000). For each of
the K = 2 to K = 13 settings, 20 independent simulations were
performed using the admixture model. To compare likelihoods
of each assumption, a Wilcoxon two-sample test was performed
(Rosenberg et al., 2001). A between-simulation similarity coef-
fi cient was calculated using the STRUCTURE-sum program
(Ehrich, 2006). To detect the optimal clustering number (K),
the ad hoc statistic ΔK, which is based on the rate of change
in the log probability of data between successive K values, was
calculated using the STRUCTURE-sum program (Evanno et
al., 2005). ΔK identifi ed the optimal K number as 8 based on
the lowest ΔK value and no further signifi cant decrease beyond
K = 8. The STRUCTURE bar graph was established with the
software Distruct (Rosenberg, 2002).
Neighbor-Joining Tree Construction and Principal Coordinate AnalysisThe genetic distance among 155 Mesoamerican accessions was
calculated using the C.S. Chord distance (Cavalli-Sforza and
Edwards, 1967). The C.S. Chord distance does not require
any mutation model for microsatellite evolution and is free
from bottleneck eff ect (Takezaki and Nei, 1996). Based on this
genetic distance, an unrooted neighbor-joining (NJ) tree was
constructed in PowerMarker (Liu and Muse, 2005). Principal
coordinate analysis was performed using the GenAlEx 6 Pro-
gram (Peakall and Smouse, 2006).
RESULTS
Identifi cation of Mesoamerican Accessions
Extensive research has shown that common bean consists of two major ecogeographic gene pools. The existence of Andean and Mesoamerican gene pools was initially sug-gested on the basis of the geographic distribution of hybrid lethality genes based on electrophoretic patterns of phaseo-lin seed protein (Gepts and Bliss, 1985; Gepts et al., 1986). This reproductive isolation preexisted domestication (Koin-ange and Gepts, 1992). Additional evidence for these two gene pools was provided by morphological (Gepts et al., 1986; Singh et al., 1991a,b), molecular marker (Singh et al., 1991c; Khairallah et al., 1990, 1992; Becerra-Velásquez and Gepts 1994; Tohme et al., 1996; Chacón et al., 2007), and phytopathological (Guzmán et al., 1995; Kelly and Vallejo, 2004) data. Thus, domesticated common bean resulted

CROP SCIENCE, VOL. 49, MARCH–APRIL 2009 WWW.CROPS.ORG 557
to the ecogeographic race Mesoamerica (K7) and the other including ecogeographic races Jalisco and Durango (K6) (Singh et al., 1991a). A close relationship between races Jalisco and Durango had already been suggested with earlier data (Pallottini et al., 2004). Wild populations were subdivided by STRUCTURE into fi ve groups, in part along geographic lines. In the Andes, the ancestral wild group from northern Peru and Ecuador (K2) (Kami et al., 1995) was distinct from the Colombian wild beans (K1) (Fig. 1). Wild beans from Mesoamerica were divided into three groups although no geographic, topographic or ecological pattern could be identifi ed as yet for this subdivision (K3, K4, and K8). The eighth group included wild beans from two populations near the municipality of Arandas in eastern Jalisco (K5). Segregation for seed size and color among these populations suggests they resulted from recent outcrossing events between wild and domes-ticated types in these locations.
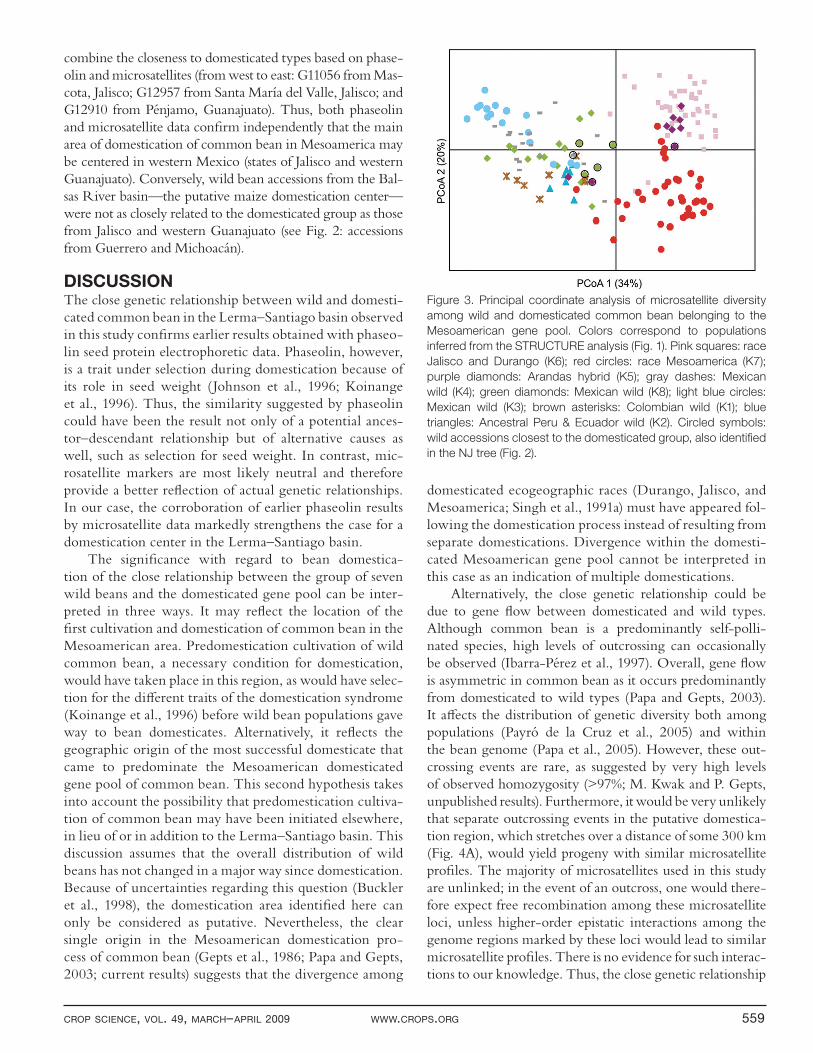
To better understand the genetic relatedness among and within these diff erent groups, genetic distances were calculated using the C.S. Chord distance, which does not assume a specifi c mutation model for microsatellite evo-lution and is free from bottleneck eff ect (Cavalli-Sforza and Edwards, 1967; Takezaki and Nei, 1996). A cluster analysis using these genetic distances was conducted based on the NJ method as implemented in Powermarker ver-sion 3.25 (Liu and Muse, 2005). The resulting tree (Fig. 2) identifi ed a predominantly single-cluster origin for the Mesoamerican domestication of common bean, with 95% of Mesoamerican domesticates grouped in a single clade, confi rming earlier evidence based on phaseolin data (Gepts et al., 1986) and amplifi ed fragment length polymorphism (Papa and Gepts, 2003). The tree further confi rms the subdivisions identifi ed by the STRUCTURE analysis. These include a geographic distribution of genetic diver-sity in wild beans belonging to the Mesoamerican gene pool. Specifi cally, wild common bean from Ecuador and northern Peru (a group that is the presumed ancestor of P. vulgaris; Kami et al., 1995), Colombia, and Mexico and Guatemala were included in diff erent branches (Fig. 2).
The tree also identifi ed two major domesticated sub-groups in the domesticated cluster, one coinciding with race Mesoamerica with small seeds (generally <25–30 g/100 seeds) and the other including races Jalisco and Durango, with medium- to large-size seeds (>30 g/100 seeds) (Singh et al., 1991a). There were two exceptions to this single-cluster pattern of Mesoamerican domestication. First, a smaller (n = 5) group of domesticated accessions was sepa-rate from the main group of domesticated accessions and which included three cultivars from the STRUCTURE K7 group (G01344; G03807; and G05059), representing the Mesoamerica ecogeographic race, and two cultivars from the STRUCTURE K6 group (G01796 and G05024), representing races Durango and Jalisco. One possibility is
that this small group resulted from hybridization Meso-american and Andean domesticated types. Membership coeffi cients in the Mesoamerican gene pool, as calculated with STRUCTURE for K = 2, were G01344 (0.991), G03807 (0.989), G05059 (0.997), G01796 (0.994), and G05024 (0.995). These high values for membership coef-fi cients suggest either that these accessions are not Andean × Mesoamerican hybrids or that introgression has aff ected the genome of these accessions in a minimal way. The latter is the case for accession G03807 (Brasil2), which resulted from the introgression of an Andean determi-nacy allele into a Mesoamerican background (Singh et al., 1991a; M. Kwak and P. Gepts, unpublished results) yet shows a high Mesoamerican membership coeffi cient. Another possibility is that this small group consists of intra-Mesoamerican gene pool hybrids. This appears to be the case for accessions G01796 (K6: races Jalisco & Durango: 0.437; K4: Mexican wild: 0.28; K7: race Meso-america: 0.185) and G05024 (K6: races Jalisco & Durango: 0.449; K7: race Mesoamerica: 0.448). This leaves acces-sions G01344 (unnamed from Nicaragua) and G05059 (H6 Mulatinho from Brazil), which are apparently nonhybrid and belong to the K7 (race Mesoamerica) group (mem-bership coeffi cients: 0.921 and 0.964, respectively). Both accessions are small-seeded (~20–23 g/100 seeds accord-ing to the CIAT database, http://isa.ciat.cgiar.org/urg/main.do?language=en). Gepts and Bliss (1986) and Beebe et al. (2000) speculated on the existence of an additional, minor center of domestication or ecogeographic race in Central America and lowland South America. Additional materials are needed from these regions to better under-stand their origin.
The second exception to the single-cluster pattern of Mesoamerican domestication was a cluster of acces-sions separating the two main domesticated subgroups (race Mesoamerica; races Durango and Jalisco). This cluster of accessions (Fig. 2: Arandas hybrids: accessions G12922–G12935) represents a cross between wild and domesticated types, all originating from El Tule, munici-pality of Arandas, in Jalisco.
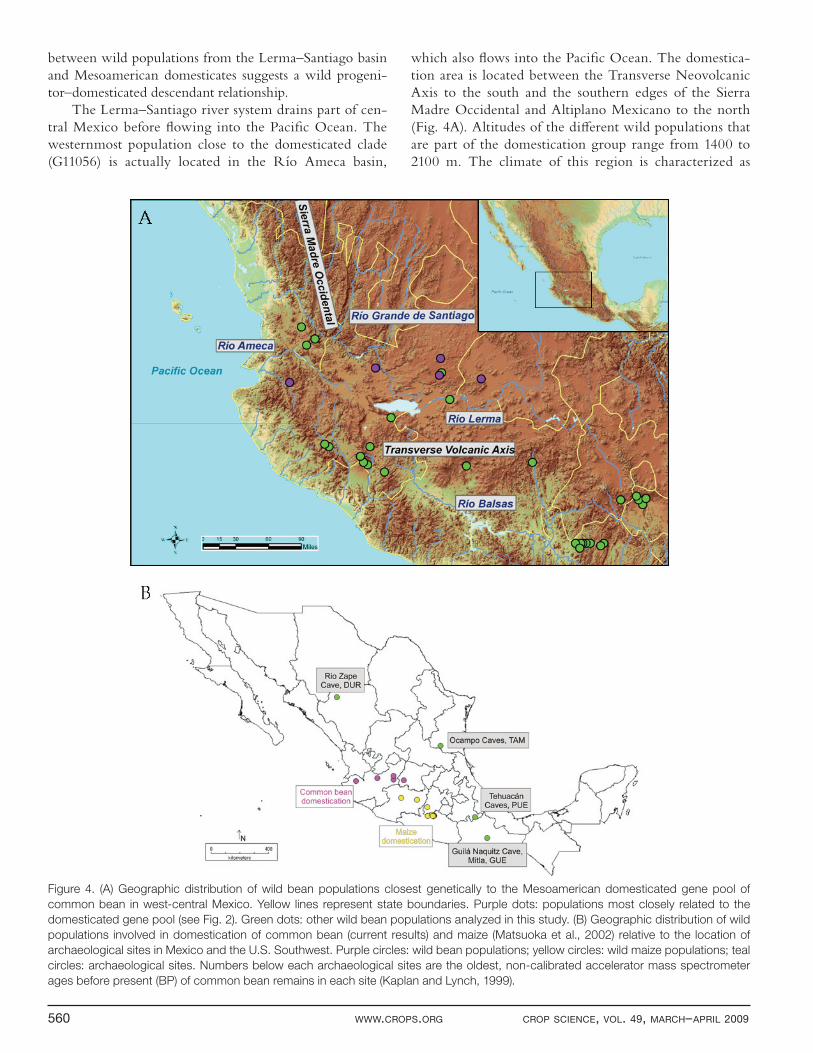
A principal coordinate analysis (Fig. 3) provides a similar picture as the NJ tree. The fi rst axis, which accounts for 34% of the variation, separates wild from domesticated types. The second axis separates the two domesticated groups: race Mesoamerica and races Jalisco and Durango. Among wild types, the separations among diff erent STRUCTURE groups are not as clear. How-ever, the wild groups from Ecuador and northern Peru and Colombia tend to separate from the three wild Mex-ican groups, consistent with their geographic origins. In turn, wild types from Ecuador and northern Peru are separated from the wild Colombian types along the third axis (not shown).
558 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, MARCH–APRIL 2009
Wild Beans Most Closely Related to the Mesoamerican Domesticated CladeA group of seven wild bean populations most closely related to the domesticated clade (G11051, G11056, G12910, G12949, G 12952, G12953, G12957; Fig. 2 and 3) is distributed in a narrow region oriented east–west across the state of Jalisco and into the western part of the neighboring state of Guanajuato. This region is centered in the Río Lerma–Río Grande de Santiago River basin (hereafter called the Lerma–Santiago basin; Fig. 4A). It was also identifi ed earlier using electrophoretic patterns of phaseo-lin seed protein coded by the Phs locus (Gepts, 1988). Three wild bean populations
Figure 1. STRUCTURE analysis (Pritchard et al., 2000) of a Mesoamerican sample of
common bean. The optimal clustering number (K = 8) was determined based on the ad
hoc statistic ΔK calculated using the STRUCTURE-sum program (Evanno et al., 2005).
Figure 2. Unrooted neighbor-joining tree based on the C.S. Chord distance for 26 microsatellites among Mesoamerican wild and
domesticated beans. The colors represent the K = 8 populations identifi ed in the STRUCTURE (Pritchard et al., 2000) analysis in Fig.
1. Filled circles indicate domesticated accessions. Filled diamonds indicate wild populations closest to the principal Mesoamerican
domesticated branch. Abbreviations at the end of wild population branches indicate Mexican state of origin—CHH: Chihuahua; COL:
Colima; DUR: Durango; GTO: Guanajuato; JAL: Jalisco; MICH: Michoacán; MOR: Morelos; NAY: Nayarit; Guatemala: GUA.
CROP SCIENCE, VOL. 49, MARCH–APRIL 2009 WWW.CROPS.ORG 559
combine the closeness to domesticated types based on phase-olin and microsatellites (from west to east: G11056 from Mas-cota, Jalisco; G12957 from Santa María del Valle, Jalisco; and G12910 from Pénjamo, Guanajuato). Thus, both phaseolin and microsatellite data confi rm independently that the main area of domestication of common bean in Mesoamerica may be centered in western Mexico (states of Jalisco and western Guanajuato). Conversely, wild bean accessions from the Bal-sas River basin—the putative maize domestication center—were not as closely related to the domesticated group as those from Jalisco and western Guanajuato (see Fig. 2: accessions from Guerrero and Michoacán).
DISCUSSIONThe close genetic relationship between wild and domesti-cated common bean in the Lerma–Santiago basin observed in this study confi rms earlier results obtained with phaseo-lin seed protein electrophoretic data. Phaseolin, however, is a trait under selection during domestication because of its role in seed weight ( Johnson et al., 1996; Koinange et al., 1996). Thus, the similarity suggested by phaseolin could have been the result not only of a potential ances-tor–descendant relationship but of alternative causes as well, such as selection for seed weight. In contrast, mic-rosatellite markers are most likely neutral and therefore provide a better refl ection of actual genetic relationships. In our case, the corroboration of earlier phaseolin results by microsatellite data markedly strengthens the case for a domestication center in the Lerma–Santiago basin.
The signifi cance with regard to bean domestica-tion of the close relationship between the group of seven wild beans and the domesticated gene pool can be inter-preted in three ways. It may refl ect the location of the fi rst cultivation and domestication of common bean in the Mesoamerican area. Predomestication cultivation of wild common bean, a necessary condition for domestication, would have taken place in this region, as would have selec-tion for the diff erent traits of the domestication syndrome (Koinange et al., 1996) before wild bean populations gave way to bean domesticates. Alternatively, it refl ects the geographic origin of the most successful domesticate that came to predominate the Mesoamerican domesticated gene pool of common bean. This second hypothesis takes into account the possibility that predomestication cultiva-tion of common bean may have been initiated elsewhere, in lieu of or in addition to the Lerma–Santiago basin. This discussion assumes that the overall distribution of wild beans has not changed in a major way since domestication. Because of uncertainties regarding this question (Buckler et al., 1998), the domestication area identifi ed here can only be considered as putative. Nevertheless, the clear single origin in the Mesoamerican domestication pro-cess of common bean (Gepts et al., 1986; Papa and Gepts, 2003; current results) suggests that the divergence among
domesticated ecogeographic races (Durango, Jalisco, and Mesoamerica; Singh et al., 1991a) must have appeared fol-lowing the domestication process instead of resulting from separate domestications. Divergence within the domesti-cated Mesoamerican gene pool cannot be interpreted in this case as an indication of multiple domestications.
Alternatively, the close genetic relationship could be due to gene fl ow between domesticated and wild types. Although common bean is a predominantly self-polli-nated species, high levels of outcrossing can occasionally be observed (Ibarra-Pérez et al., 1997). Overall, gene fl ow is asymmetric in common bean as it occurs predominantly from domesticated to wild types (Papa and Gepts, 2003). It aff ects the distribution of genetic diversity both among populations (Payró de la Cruz et al., 2005) and within the bean genome (Papa et al., 2005). However, these out-crossing events are rare, as suggested by very high levels of observed homozygosity (>97%; M. Kwak and P. Gepts, unpublished results). Furthermore, it would be very unlikely that separate outcrossing events in the putative domestica-tion region, which stretches over a distance of some 300 km (Fig. 4A), would yield progeny with similar microsatellite profi les. The majority of microsatellites used in this study are unlinked; in the event of an outcross, one would there-fore expect free recombination among these microsatellite loci, unless higher-order epistatic interactions among the genome regions marked by these loci would lead to similar microsatellite profi les. There is no evidence for such interac-tions to our knowledge. Thus, the close genetic relationship
Figure 3. Principal coordinate analysis of microsatellite diversity
among wild and domesticated common bean belonging to the
Mesoamerican gene pool. Colors correspond to populations
inferred from the STRUCTURE analysis (Fig. 1). Pink squares: race
Jalisco and Durango (K6); red circles: race Mesoamerica (K7);
purple diamonds: Arandas hybrid (K5); gray dashes: Mexican
wild (K4); green diamonds: Mexican wild (K8); light blue circles:
Mexican wild (K3); brown asterisks: Colombian wild (K1); blue
triangles: Ancestral Peru & Ecuador wild (K2). Circled symbols:
wild accessions closest to the domesticated group, also identifi ed
in the NJ tree (Fig. 2).
560 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, MARCH–APRIL 2009
between wild populations from the Lerma–Santiago basin and Mesoamerican domesticates suggests a wild progeni-tor–domesticated descendant relationship.
The Lerma–Santiago river system drains part of cen-tral Mexico before fl owing into the Pacifi c Ocean. The westernmost population close to the domesticated clade (G11056) is actually located in the Río Ameca basin,
which also fl ows into the Pacifi c Ocean. The domestica-tion area is located between the Transverse Neovolcanic Axis to the south and the southern edges of the Sierra Madre Occidental and Altiplano Mexicano to the north (Fig. 4A). Altitudes of the diff erent wild populations that are part of the domestication group range from 1400 to 2100 m. The climate of this region is characterized as
Figure 4. (A) Geographic distribution of wild bean populations closest genetically to the Mesoamerican domesticated gene pool of
common bean in west-central Mexico. Yellow lines represent state boundaries. Purple dots: populations most closely related to the
domesticated gene pool (see Fig. 2). Green dots: other wild bean populations analyzed in this study. (B) Geographic distribution of wild
populations involved in domestication of common bean (current results) and maize (Matsuoka et al., 2002) relative to the location of
archaeological sites in Mexico and the U.S. Southwest. Purple circles: wild bean populations; yellow circles: wild maize populations; teal
circles: archaeological sites. Numbers below each archaeological sites are the oldest, non-calibrated accelerator mass spectrometer
ages before present (BP) of common bean remains in each site (Kaplan and Lynch, 1999).

CROP SCIENCE, VOL. 49, MARCH–APRIL 2009 WWW.CROPS.ORG 561
subtropical (temperature of the coldest month between 5 and 18°C), subhumid (between 4 and 6 mo of humidity), and semiwarm (average annual temperature between 18 and 22°C) (López Soto et al., 2005). The original vegeta-tion changes gradually from a conifer–oak forest at the western end through a dry deciduous forest in the central part to a drier thorn forest at the eastern end of this area (Rzedowski, 1981).
The proposed domestication area for common bean does not overlap with the area proposed for maize domes-tication, based on genetic data (Matsuoka et al., 2002), nor does it overlap with any of the three major archaeo-logical locations pertaining to the origin of agriculture in the Mesoamerican area: the Oaxaca Valley, the Tehuacán Caves in Puebla, and the Ocampo Caves in Tamaulipas (Fig. 4B). Buckler et al. (1998) already observed that these archaeological sites were situated in an arid ecological zone that was unimportant in the domestication of most Mesoamerican crops. Thus, the question whether the major food crops in the Mesoamerican center of agricul-tural origins resulted from domestication in a single or multiple locations in Mexico remains to be answered. The oldest archaeological records for common bean are from the southwestern United States (2470 and 2140 BP), the Tehuacán Caves (2285 BP), and the Oaxaca Valley (2098 BP) (Kaplan and Lynch, 1999) (all Accelerator Mass Spec-trometry data, uncalibrated years; Fig. 4B). That these domesticated remains of approximately the same age are distributed over a distance of some 2300 km suggests that they refl ect an established, agriculture-based economy. Older remnants, representing the transition from wild to domesticated beans, remain to be discovered. Further sup-port for the utilization of beans at an earlier stage than suggested by archaeological dating is provided by lin-guistic analyses of ancient indigenous languages such as proto-Mayan, which suggest an origin of at least 3400 BP (Brown, 2006). Our data also suggest that a portion of the Lerma–Santiago basin should be the focus of archaeologi-cal eff orts to identify new sites pertaining to the origin of agricultural origins in Mesoamerica. Furthermore, Jalisco has also been suggested as one of two possible domestica-tion centers for tepary bean (P. acutifolius A. Gray) (Muñoz et al., 2006).
DNA marker analyses of cereals domesticated in Southwest Asia (the Fertile Crescent), combined with archaeological data, had identifi ed a “core area” for agri-cultural origins within the Fertile Crescent (Abbo et al., 2006). It is from this core area that agriculture rippled out to other areas of the Fertile Crescent and, eventually, to other regions in Asia, Africa, and Europe. (Recently, this centric model has been questioned; Kilian et al., 2007). Our data, showing that the two main components of the milpa cropping system were domesticated in diff erent areas of Mesoamerica, question whether a similar core area
could be identifi ed for this center of agricultural origins. The assembly of the traditional milpa cropping system, consisting of beans, maize, and squash, may have resulted from a more diff use and extensive pre- or postdomestica-tion cultivation and domestication process.
Finally, our data should provide additional impetus for a refocused archaeological eff ort into the river basins of west-central Mexico. The moister areas may provide a more appropriate target to identify sites relevant to the ori-gins of agriculture than the existing sites in arid regions. Recent progress in archaeobotanical approaches has dem-onstrated the possibility of identifying crop remains such as phytoliths or starch grains in moister environments (Holst et al., 2007; Perry et al., 2007). Our results should also encourage additional research on wild relatives and landraces of crops in west-central Mexico, including addi-tional explorations and ecological studies on adaptation to climatic and edaphic factors and interactions with patho-gens and pests.
AcknowledgmentsWe thank D. Debouck and O. Toro (CIAT, Cali, Colombia)
and M. Welsh (Western Regional Plant Introduction Station,
USDA, Prosser, WA) for supplying seed samples. D. Zizumbo,
A. Delgado, D. Velasco, S. Repinski, and K. Kraft provided
comments on an earlier version of this manuscript. This work
was supported by the USDA CSREES NRI Plant Genome
program to PG and a graduate scholarship from the Univ. of
California, Davis, Dep. of Plant Science to MK.
ReferencesAbbo, S., A. Gopher, Z. Peleg, Y. Saranga, T. Fahima, F. Salamini,
and S. Lev-Yadun. 2006. The ripples of “The big (agricul-
tural) bang”: The spread of early wheat cultivation. Genome
49:861–863.
Becerra Velásquez, V.L., and P. Gepts. 1994. RFLP diversity in
common bean (Phaseolus vulgaris L.). Genome 37:256–263.
Beebe, S., P.W. Skroch, J. Tohme, M.C. Duque, F. Pedraza, and
J. Nienhuis. 2000. Structure of genetic diversity among com-
mon bean landraces of Middle American origin based on cor-
respondence analysis of RAPD. Crop Sci. 40:264–273.
Blair, M.W., F. Pedraza, H.F. Buendia, E. Gaitán-Solís, S.E. Beebe,
P. Gepts, and J. Tohme. 2003. Development of a genome-
wide anchored microsatellite map for common bean (Phaseo-
lus vulgaris L.). Theor. Appl. Genet. 107:1362–1374.
Bressani, R. 1983. Research needs to upgrade the nutritional qual-
ity of common beans (Phaseolus vulgaris). Plant Foods Hum.
Nutr. 32:101–110.
Brown, C.H. 2006. Prehistoric chronology of the common bean
in the New World: The linguistic evidence. Am. Anthropol.
108:507–516.
Buckler, E., D. Pearsall, and T. Holtsford. 1998. Climate, plant
ecology, and Central Mexican Archaic subsistence. Curr.
Anthropol. 39:152–164.
Cardoso, E., M.A. Nogueira, and S.M.G. Ferraz. 2007. Biological
N-2 fi xation and mineral N in common bean-maize inter-
cropping or sole cropping in southeastern Brazil. Exp. Agric.
43:319–330.

562 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, MARCH–APRIL 2009
Cavalli-Sforza, L.L., and A.W.F. Edwards. 1967. Phylogenetic
analysis: Models and estimation procedures. Am. J. Hum.
Genet. 19:233–257.
Chacón, S., M.I., B. Pickersgill, D.G. Debouck, and J.S. Arias.
2007. Phylogeographic analysis of the chloroplast DNA varia-
tion in wild common bean (Phaseolus vulgaris L.) in the Ameri-
cas. Plant Syst. Evol. 10, doi:1007/s00606-007-0536-z.
Delgado Salinas, A., A. Bonet, and P. Gepts. 1988. The wild rela-
tive of Phaseolus vulgaris in Middle America, p. 163–184. In
P. Gepts (ed.) Genetic resources of Phaseolus beans. Kluwer,
Dordrecht, the Netherlands.
Doebley, J. 2004. The genetics of maize evolution. Annu. Rev.
Genet. 38:37–59.
Doyle, J., and J. Doyle. 1987. A rapid DNA isolation procedure for
small quantities of fresh leaf tissue. Phytochem. Bull. 19:11–15.
Ehrich, D. 2006. AFLPDAT: A collection of R functions for conve-
nient handling of AFLP data. Mol. Ecol. Notes 6:603–604.
Evanno, G., S. Regnaut, and J. Goudet. 2005. Detecting the num-
ber of clusters of individuals using the software STRUC-
TURE: A simulation study. Mol. Ecol. 14:2611–2620.
Freytag, G.F., and D.G. Debouck. 2002. Taxonomy, distribution,
and ecology of the genus Phaseolus (Leguminosae–Papilion-
oideae) in North America, Mexico, and Central America.
Botanical Research Inst. of Texas, Forth Worth.
Gaitán-Solís, E., M.C. Duque, K.J. Edwards, and J. Tohme. 2002.
Microsatellite repeats in common bean (Phaseolus vulgaris):
Isolation, characterization, and cross-species amplifi cation in
Phaseolus ssp. Crop Sci. 42:2128–2136.
Gepts, P. 1988. Phaseolin as an evolutionary marker, p. 215–241.
In P. Gepts (ed.) Genetic resources of Phaseolus beans. Kluwer,
Dordrecht, the Netherlands.
Gepts, P. 1998. Origin and evolution of common bean: Past events
and recent trends. HortScience 33:1124–1130.
Gepts, P., and F.A. Bliss. 1985. F1 hybrid weakness in the common
bean: Diff erential geographic origin suggests two gene pools
in cultivated bean germplasm. J. Hered. 76:447–450.
Gepts, P., and F.A. Bliss. 1986. Phaseolin variability among wild
and cultivated common beans (Phaseolus vulgaris) from Colom-
bia. Econ. Bot. 40:469–478.
Gepts, P., T.C. Osborn, K. Rashka, and F.A. Bliss. 1986. Phase-
olin-protein variability in wild forms and landraces of the
common bean (Phaseolus vulgaris): Evidence for multiple cen-
ters of domestication. Econ. Bot. 40:451–468.
Gepts, P., K. Kmiecik, P. Pereira, and F.A. Bliss. 1988. Dissemina-
tion pathways of common bean (Phaseolus vulgaris, Fabaceae)
deduced from phaseolin electrophoretic variability: I. The
Americas. Econ. Bot. 42:73–85.
Guzmán, P., R.L. Gilbertson, R. Nodari, W.C. Johnson, S.R.
Temple, D. Mandala, A.B.C. Mkandawire, and P. Gepts.
1995. Characterization of variability in the fungus Phaeoisa-
riopsis griseola suggests coevolution with the common bean
(Phaseolus vulgaris). Phytopathology 85:600–607.
Hancock, J.F. 2004. Plant evolution and the origin of crop species.
2nd ed. CAB Int., Wallingford, UK.
Holst, I., J.E. Moreno, and D.R. Piperno. 2007. Identifi cation of
teosinte, maize, and Tripsacum in Mesoamerica by using pol-
len, starch grains, and phytoliths. Proc. Natl. Acad. Sci. USA
104:17608–17613.
Ibarra-Pérez, F., B. Ehdaie, and G. Waines. 1997. Estimation of
outcrossing rate in common bean. Crop Sci. 37:60–65.
Johnson, W., C. Menéndez, R. Nodari, E. Koinange, S. Singh, and
P. Gepts. 1996. Association of a seed weight factor with the
phaseolin seed storage protein locus across genotypes, envi-
ronments, and genomes in Phaseolus–Vigna spp.: Sax (1923)
revisited. J. Agric. Genomics ( J. Quant. Trait Loci) 2:5.
Kami, J., B. Becerra Velásquez, D.G. Debouck, and P. Gepts.
1995. Identifi cation of presumed ancestral DNA sequences
of phaseolin in Phaseolus vulgaris. Proc. Natl. Acad. Sci. USA
92:1101–1104.
Kaplan, L., and T. Lynch. 1999. Phaseolus (Fabaceae) in archaeol-
ogy: AMS radiocarbon dates and their signifi cance for pre-
Columbian agriculture. Econ. Bot. 53:261–272.
Kelly, J.D., and V.A. Vallejo. 2004. A comprehensive review of the
major genes conditioning resistance to anthracnose in com-
mon bean. HortScience 39:1196–1207.
Khairallah, M.M., M.W. Adams, and B.B. Sears. 1990. Mitochon-
drial DNA polymorphisms of Malawian bean lines: Further
evidence for two major gene pools. Theor. Appl. Genet.
80:753–761.
Khairallah, M.M., B.B. Sears, and M.W. Adams. 1992. Mito-
chondrial restriction fragment polymorphisms in wild Phaseo-
lus vulgaris: insights in the domestication of common bean.
Theor. Appl. Genet. 84:915–922.
Kilian, B., H. Ozkan, A. Walther, J. Kohl, T. Dagan, F. Salamini,
and W. Martin. 2007. Molecular diversity at 18 loci in 321
wild and 92 domesticate lines reveal no reduction of nucle-
otide diversity during Triticum monococcum (einkorn) domesti-
cation: Implications for the origin of agriculture. Mol. Biol.
Evol. 24:2657–2668.
Koinange, E.M.K., and P. Gepts. 1992. Hybrid weakness in wild
Phaseolus vulgaris L. J. Hered. 83:135–139.
Koinange, E.M.K., S.P. Singh, and P. Gepts. 1996. Genetic control
of the domestication syndrome in common-bean. Crop Sci.
36:1037–1045.
Liu, K., and S.V. Muse. 2005. PowerMarker: Integrated analy-
sis environment for genetic marker data. Bioinformatics
21:2128–2129.
López Soto, J.L., J.A. Ruiz Corral, J.J. Sánchez González, and R.
Lépiz Ildefonso. 2005. Climatic adaptation of 25 wild bean
species (Phaseolus spp) in Mexico. Rev. Fitotecnia Mexicana
28:221–230.
Matsuoka, Y., Y. Vigouroux, M.M. Goodman, G.J. Sanchez, E.
Buckler, and J. Doebley. 2002. A single domestication for
maize shown by multilocus microsatellite genotyping. Proc.
Natl. Acad. Sci. USA 99:6080–6084.
Montes-Hernandez, S., and L.E. Eguiarte. 2002. Genetic struc-
ture and indirect estimates of gene fl ow in three taxa of
Cucurbita (Cucurbitaceae) in western Mexico. Am. J. Bot.
89:1156–1163.
Muñoz, L.C., M.C. Duque, D.G. Debouck, and M.W. Blair. 2006.
Taxonomy of tepary bean and wild relatives as determined
by amplifi ed fragment length polymorphism (AFLP) markers.
Crop Sci. 46:1744–1754.
Pallottini, L., E. Garcia, J. Kami, G. Barcaccia, and P. Gepts. 2004.
The genetic anatomy of a patented yellow bean. Crop Sci.
44:968–977.
Papa, R., J. Acosta, A. Delgado-Salinas, and P. Gepts. 2005. A
genome-wide analysis of diff erentiation between wild and
domesticated Phaseolus vulgaris from Mesoamerica. Theor.
Appl. Genet. 111:1147–1158.
Papa, R., and P. Gepts. 2003. Asymmetry of gene fl ow and dif-
ferential geographical structure of molecular diversity in wild
and domesticated common bean (Phaseolus vulgaris L.) from
Mesoamerica. Theor. Appl. Genet. 106:239–250.

CROP SCIENCE, VOL. 49, MARCH–APRIL 2009 WWW.CROPS.ORG 563
Paredes, O.M., and P. Gepts. 1995. Extensive introgression of
Middle American germplasm into Chilean common bean
cultivars. Genet. Resour. Crop Evol. 42:29–41.
Payró de la Cruz, E., P. Gepts, P. Colunga GarciaMarín, and
D. Zizumbo Villareal. 2005. Spatial distribution of genetic
diversity in wild populations of Phaseolus vulgaris L. from
Guanajuato and Michoacán, México. Genet. Resour. Crop
Evol. 52:589–599.
Peakall, R., and P.E. Smouse. 2006. GENALEX 6: Genetic anal-
ysis in Excel: Population genetic software for teaching and
research. Mol. Ecol. Notes 6:288–295.
Perry, L., R. Dickau, S. Zarrillo, I. Holst, D.M. Pearsall, D.R.
Piperno, M.J. Berman, R.G. Cooke, K. Rademaker, A.J.
Ranere, J.S. Raymond, D.H. Sandweiss, F. Scaramelli, K.
Tarble, and J.A. Zeidler. 2007. Starch fossils and the domesti-
cation and dispersal of chili peppers (Capsicum spp. L.) in the
Americas. Science 315:986–988.
Piperno, D., and K. Flannery. 2001. The earliest archaeological
maize (Zea mays L.) from highland Mexico: New accelerator
mass spectrometry dates and their implications. Proc. Natl.
Acad. Sci. USA 98:2101–2103.
Pritchard, J., M. Stephens, and P. Donnelly. 2000. Inference of
population structure using multilocus genotype data. Genet-
ics 155:945–959.
Rosenberg, N.A., T. Burke, K. Elo, M.W. Feldmann, P.J. Freidlin,
M.A.M. Groenen, J. Hillel, A. Maki-Tanila, M. Tixier-Boich-
ard, A. Vignal, K. Wimmers, and S. Weigend. 2001. Empiri-
cal evaluation of genetic clustering methods using multilocus
genotypes from 20 chicken breeds. Genetics 159:699–713.
Rosenberg, N.A. 2002. Distruct: A program for the graphical
display of Structure results Available at http://rosenberglab.
bioinformatics.med.umich.edu/distruct.html (verifi ed 4 Feb.
2009). Univ. of Michigan, Ann Arbor.
Rzedowski, J. 1981. Vegetación de México. Limusa, Mexico, D.F.
Singh, S.P., P. Gepts, and D.G. Debouck. 1991a. Races of common
bean (Phaseolus vulgaris L., Fabaceae). Econ. Bot. 45:379–396.
Singh, S.P., J.A. Gutiérrez, A. Molina, C. Urrea, and P. Gepts.
1991b. Genetic diversity in cultivated common bean: II.
Marker-based analysis of morphological and agronomic traits.
Crop Sci. 31:23–29.
Singh, S.P., R. Nodari, and P. Gepts. 1991c. Genetic diversity in
cultivated common bean: I. Allozymes. Crop Sci. 31:19–23.
Smith, B.D. 1995. The emergence of agriculture. Scientifi c Amer-
ican Library, New York.
Smith, B.D. 1997. The initial domestication of Cucurbita pepo in
the Americas 10,000 years ago. Science 276:932–934.
Takezaki, N., and M. Nei. 1996. Genetic distances and reconstruc-
tion of phylogenetic trees from microsatellite DNA. Genetics
144:389–399.
Tohme, J., D.O. González, S. Beebe, and M.C. Duque. 1996.
AFLP analysis of gene pools of a wild bean core collection.
Crop Sci. 36:1375–1384.
Wilkes, G. 2004. Corn, strange and marvelous: But is a defi nitive
origin known? p. 3–63, In C. Smith (ed.) Corn: Origin, his-
tory, technology, and production. John Wiley & Sons, New
York.
Yu, K., S. Park, V. Poysa, and P. Gepts. 2000. Integration of simple
sequence repeat (SSR) markers into a molecular linkage map
of common bean (Phaseolus vulgaris L.). J. Hered. 91:429–434.
Zeder, M.A., E. Emshwiller, B.D. Smith, and D.G. Bradley. 2006.
Documenting domestication: The intersection of genetics and
archaeology. Trends Genet. 22:139–155.





























