Embed Size (px)
Citation preview

Research Article
The Promotion of Breast Cancer Metastasis Caused byInhibition of CSF-1R/CSF-1 Signaling Is Blocked by Targetingthe G-CSF Receptor
Agnieszka Swierczak1,3, Andrew D. Cook3, Jason C. Lenzo3, Christina M. Restall1, Judy P. Doherty1,Robin L. Anderson1,2,4, and John A. Hamilton3
AbstractTreatment options are limited for patients with breast cancer presenting with metastatic disease. Targeting
of tumor-associated macrophages through the inhibition of colony-stimulating factor-1 receptor (CSF-1R), akey macrophage signaling pathway, has been reported to reduce tumor growth and metastasis, and thesetreatments are now in clinical trials. Here, we report that, surprisingly, treatment with neutralizing anti–CSF-1Rand anti–CSF-1 antibodies, or with two different small-molecule inhibitors of CSF-1R, could actually increasespontaneous metastasis without altering primary tumor growth in mice bearing two independently derivedmammary tumors. The blockade of CSF-1R or CSF-1 led to increased levels of serumG-CSF, increased frequencyof neutrophils in the primary tumor and in the metastasis-associated lung, as well as increased numbers ofneutrophils and Ly6Chi monocytes in the peripheral blood. Neutralizing antibody against the G-CSF receptor,which regulates neutrophil development and function, reduced the enhanced metastasis and neutrophilnumbers that resulted fromCSF-1R blockade. These results indicate that the role of the CSF-1R/CSF-1 system inbreast cancer is far more complex than originally proposed, and requires further investigation as a therapeutictarget. Cancer Immunol Res; 2(8); 765–76. �2014 AACR.
IntroductionThe link between chronic inflammation and the onset and
progression of many cancers is well established (1). Tumor-associated macrophages (TAM), which have been subjected toextensive study, can execute several key roles required fortumor growth and progression (1, 2). Colony-stimulating fac-tor-1 (CSF-1/M-CSF) regulates the recruitment, differentia-tion, survival, and proliferation of macrophages (3). CSF-1activity is mediated through its interaction with the receptortyrosine kinase, CSF-1 receptor (CSF-1R/c-Fms/CD115; ref. 3),which is expressed primarily on mononuclear phagocyticcells and their precursors (4). Interleukin-34 (IL34) is a secondligand for CSF-1R; it controls the development of specificmacrophage lineages, such as microglia and Langerhans cells(5). IL34 and CSF-1 have distinct expression patterns (6).
The association between CSF-1/CSF-1R expression andprognosis in breast cancer is still unclear. A study of invasivebreast adenocarcinomas (grades 1–3) revealed that, althoughnuclear staining of CSF-1 in tumor cells was associated withpoor outcome, it was more strongly predictive of increasednumbers of infiltratingmonocytes (7). In a study of noninvasiveand invasive breast adenocarcinomas (grades 1–3), elevatedexpression of CSF-1 was found predominantly in invasivetumor cells (8). In a study of early-stage 1/2 breast cancer, theauthors reported a correlation between high levels of CSF-1Rin tumors and local recurrence (9). Distant metastasis andsurvival were not assessed in the latter two reports. Assessmentof node-negative and node-positive breast cancer tissuesrevealed that high CSF-1R expression was associated with ashorter overall survival for node-negative, but not node-pos-itive patients (10). However, this study also found that CSF-1Rexpression alone was not an independent predictor of survival(10). Beck and colleagues (11) investigated a "CSF-1 responsegene signature" in whole breast tumor datasets and found thatthe signature was associated with survival in only one of fivedatasets. The authors concluded that the CSF-1 responsesignature has a complex relationship with patient survival (11).
CSF-1 is a potent macrophage chemoattractant, and it pol-arizes macrophages in the tumor microenvironment towardtumor-promoting TAMs (12). Preclinical studies have reporteda role for CSF-1 in metastasis via the regulation of TAMs. Inmacrophage-deficient MMTV-PyMT mice carrying a null mu-tation in the Csf1 gene, TAMs were unable to accumulate inprimary tumors, and metastasis to the lung was reduced (13).In breast cancer xenografts in immunocompromised mice,
Authors' Affiliations: 1Research Division, Peter MacCallum Cancer Centre,East Melbourne; 2Sir Peter MacCallum Department of Oncology; 3Depart-ment of Medicine, Arthritis and Inflammation Research Centre; and 4Depart-ment of Pathology, The University of Melbourne, Parkville, Victoria, Australia
Note: Supplementary data for this article are available at Cancer Immu-nology Research Online (http://cancerimmunolres.aacrjournals.org/).
R.L. Anderson and J.A. Hamilton share senior authorship of this article.
Corresponding Author: Robin L. Anderson, Peter MacCallum CancerCentre, Locked Bag 1, A'Beckett Street, Melbourne, VIC 8006, Australia.Phone: 61-3-9656-1284; Fax: 61-3-9656-1411; E-mail:[email protected]
doi: 10.1158/2326-6066.CIR-13-0190
�2014 American Association for Cancer Research.
CancerImmunology
Research
www.aacrjournals.org 765
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190

depletion of TAMs by CSF-1 antisense oligonucleotides orneutralizing antibody resulted in reduced primary tumorgrowth (14, 15). On the basis of the results from these studies,CSF-1R/CSF-1 signaling is considered a potential target foradjuvant therapy of breast cancer. A phase I clinical trial hasbeen initiated using a monoclonal antibody (mAb) againstCSF-1R—IMC-CS4—as a therapy for solid tumors (16).PLX3397, a small-molecule inhibitor of CSF-1R, is also in phaseI trials for solid tumors (17) and for metastatic breast cancer incombination with chemotherapy (18).
Little is known about the role of tissue macrophage pre-cursors, blood monocytes, in metastasis. CD11bþGr-1þ mye-loid cells facilitate tumor growth (19), and their human homo-logs are reported to be elevated in patients with cancer (20).These cells are also found in blood, the spleen, and in thepremetastatic lung of tumor-bearing mice (19). Because Gr-1recognizes both the Ly6C and Ly6G antigens (21), Gr-1þ cellscan be monocytes, macrophages, or neutrophils. Two majorblood monocyte subpopulations have been defined, basedprimarily on surface marker expression. In the mouse, theseare "classical" monocytes (Ly6C(Gr-1)hiCX3CR1
loCCR2hi
CD62Lhi) and "nonclassical" monocytes (Ly6C(Gr-1)lo CX3CR1hiCCR2loCD62Llo). The former, and possibly less maturesubpopulation, generally appears first at sites of inflammation(22, 23). However, the relationship between monocyte subpo-pulations and tissue macrophages is still unclear, particularlyduring inflammatory/immune reactions (24).
Neutrophils are found in breast tumors and have beenimplicated in promoting metastasis (19, 25, 26). Elevated levelsof neutrophils in tumors are associatedwith decreased survivalin a number of cancer types, including pulmonary and gastricadenocarcinomas (27, 28). Whether there is an associationbetween neutrophil numbers and prognosis in human breastcancer is not known, although increased levels of the neutro-phil-specific protease, neutrophil elastase, have been associ-ated with reduced disease-free survival (29). G-CSF regulatesneutrophil production by inducing proliferation and matura-tion of myeloid progenitors and by promoting neutrophilrelease from the bone marrow (30). A recent study intumor-bearing mice found that blocking G-CSF reduced neu-trophil numbers andmetastasis to the lung, whereas treatmentwith rG-CSF had the opposite effect (25).
Given the association between CSF-1, TAMs, and breastcancer metastasis, we anticipated that blockade of CSF-1R orCSF-1 would deplete TAMs and subsequently limit metastasisand possibly primary tumor growth as well. However, unex-pectedly, we found that such neutralization actually increasesmetastasis to the lung and bone in two independentmammarytumor models, and is associated with an increase in neutro-phils at multiple sites. Blockade of G-CSFR overcomes theincrease in metastasis and neutrophil numbers that resultsfrom CSF-1R neutralization, indicating that this enhancedmetastasis is driven by G-CSF.
Materials and MethodsAntibodies, reagents, and cell lines
The 4T1.2, 66cl4, 67NR, and EMT6.5 murine tumor cell lines(31–34) expressing mCherry fluorescent protein were main-
tained in a Minimal Essential Medium (aMEM), penicillin/streptomycin at 100 U/mL (Invitrogen), and 5% fetal calf serum(CSL Biosciences). All cell lines were tested and validated to beMycoplasma-free. No other authentication was completed.The 4T1, 66cl4, and 67NR lines were obtained directly fromDr. F. Miller, Karmanos Cancer Institute, Wayne State Uni-versity, Detroit, MI (32), and the 4T1.2 line was derived in-house from the 4T1 line. There are no universal standardsfrom which authentication of these lines can be obtained.The following were gifts: neutralizing anti-mouse CSF-1R(AFS-98: Dr. S.-I. Nishikawa, RIKEN Center for Developmen-tal Biology, Kobe, Japan), IgG2a isotype control (GL117.41:Dr. J. Abrams, DNAX, Palo Alto, CA), neutralizing anti-mouseCSF-1 and IgG2a isotype control (Dr. F. Dodeller, Mor-phoSys, Munich, Germany), and neutralizing anti-mouseG-CSFR and IgG1 isotype control (Dr. A Andrews, CSLBiosciences, Parkville, VIC, Australia). GW2580 was providedby Cancer Therapeutics CRC, and CYC11645 was providedby Dr. C. Burns (Walter and Eliza Hall Institute, Parkville,VIC, Australia). mAbs were used against the following anti-gens: CD11b (Mac-1 a-chain, M1/70-APC-Cy7), Ly6C (AL-21-PerCP-Cy5.5), and Ly6G (1A8-PE-Cy7) all from BD Pharmin-gen, CD115 (c-Fms: AFS-98-APC) and F4/80 (BM8-FITC)both from eBioscience.
Tumor growthFemale Balb/c mice (6–8 weeks; Walter and Eliza Hall
Institute) were injected into the fourth mammary gland with1� 105 tumor cells in PBS or with saline alone. Tumor volume,(L � W2/2), was measured using electronic calipers. Foranalysis of circulating monocyte and neutrophil numbers,peripheral blood was collected from the tail vein after tumorcell inoculation as indicated. In some experiments, the primarytumor was resected at 0.5 g (700 mm3), and treatment wascommenced 6 days later. Primary tumors (if not resected),lungs, and spines were collected at endpoint. Primary tumorsand lungs were disaggregated (see below), spines were snap-frozen in liquid nitrogen, and homogenized. All proceduresinvolvingmice were conducted in accordance with the Nation-al Health andMedical Research Council of Australia guidelinesand approved by the University of Melbourne Animal EthicsCommittee and the PeterMacCallumAnimal ExperimentationEthics Committee.
Antibody and inhibitor treatmentTumor-bearing mice were treated i.p. every 4 days from
days 7 to 25 after tumor cell inoculation with anti–CSF-1R(250 mg), anti–CSF-1 (200 mg), or IgG2a isotype controlantibody, as indicated. In some experiments, mice weretreated i.p. 3 days per week with 50 mg of anti–G-CSFR orIgG1 isotype control antibody. These doses and treatmentregimens have been determined previously to be optimal formouse studies (ref. 35; and unpublished data). For the anti–CSF-1R and anti–G-CSFR antibody combination treatment,the same protocols were used as described above with the4T1.2 tumor-bearing mice receiving either both experimen-tal antibodies or both isotype controls. For the kinase inhib-itors, mice were treated by oral gavage with 160 mg/kg of
Swierczak et al.
Cancer Immunol Res; 2(8) August 2014 Cancer Immunology Research766
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190
GW2580 or vehicle. This dose has been shown previously tobe optimal for inhibition of CSF-1R (36, 37). CYC11645 isanother potent inhibitor of CSF-1R (38), described previous-ly by Burns and colleagues as compound 15 (38). CYC11645(100 mg/kg) was administered twice daily by oral gavage,from days 1 to 30 after tumor cell inoculation.
Flow cytometryCells or 20 mL of blood were washed, counted, incubated in
mouse serum, washed, and incubated with primary antibodiesor appropriate isotype controls. Cells were analyzed using theBeckman Coulter CyAn ADP Flow Cytometer (Beckman Coul-ter) or the BD LSR II Flow Cytometer (Becton/Dickinson) andFlowJo software (TreeStar; Supplementary Fig. S1). See Sup-plementary Materials and Methods for detailed procedure.
Tumor burden analysisTumor burden for each tissue was measured by qPCR
(StepOne System; Applied Biosystems) and TaqMan chemistry(33). Lung tumor burden was visualized by India ink staining.See Supplementary Materials and Methods for detailedprocedure.
Kaplan–Meier plotter and BreastMark online survivalanalysisKaplan–Meier plotter (39) and BreastMark (40) were used to
assess high and low expression of tumor CSF-1R in relation torelapse-free survival in all breast cancers using default settings.See Supplementary Materials andMethods for a more detaileddescription of these platforms.
Measurement of serum CSF-1 and G-CSFSerum was harvested at endpoint via cardiac puncture.
CSF-1 levels were measured using a murine M-CSF standardELISA development kit (Peprotech), and G-CSF levels weremeasured using a mouse G-CSF DuoSet kit (R&D Systems)as per the manufacturer's instructions.
Statistical analysisTumor growth and peripheral blood data were analyzed
using two-way ANOVA with Bonferroni correction. For allother data, a preliminary statistical analysis (D'Agostino–Pearson omnibus normality test) was used to determinewhether the data followed a normal distribution. If the datawere normally distributed, an unpaired, parametric t test wasused; if not, the Mann–Whitney U test was used. P � 0.05 wasconsidered significant.
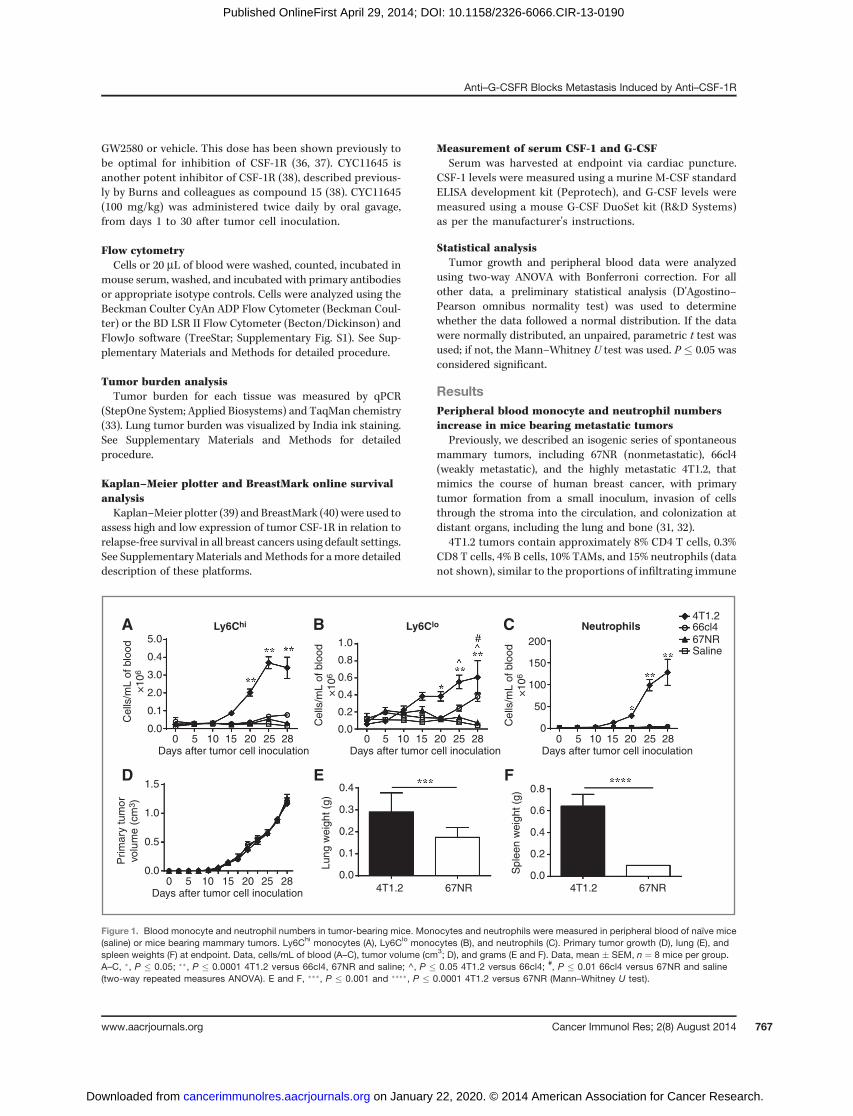
ResultsPeripheral blood monocyte and neutrophil numbersincrease in mice bearing metastatic tumors
Previously, we described an isogenic series of spontaneousmammary tumors, including 67NR (nonmetastatic), 66cl4(weakly metastatic), and the highly metastatic 4T1.2, thatmimics the course of human breast cancer, with primarytumor formation from a small inoculum, invasion of cellsthrough the stroma into the circulation, and colonization atdistant organs, including the lung and bone (31, 32).
4T1.2 tumors contain approximately 8% CD4 T cells, 0.3%CD8 T cells, 4% B cells, 10% TAMs, and 15% neutrophils (datanot shown), similar to the proportions of infiltrating immune
5.0
0.4
3.0
2.0
0.1
0.0
1.0
0.8
0.6
0.4
0.2
0.0
200
150
100
50
0
1.5
1.0
0.5
0.0
0.4
0.3
0.2
0.1
0.0
0.8
0.6
0.4
0.2
0.0
0 5 10
Ly6ChiA B C
D E F
Ly6Clo Neutrophils
15Days after tumor cell inoculation
20 25 28
0 5 10 15Days after tumor cell inoculation
20 25 284T1.2 67NR 4T1.2 67NR
4T1.2
67NRSaline
66cl4
0 5 10 15Days after tumor cell inoculation
20 25 28 0 5 10 15Days after tumor cell inoculation
20 25 28
Cel
ls/m
L of
blo
od×
106
Prim
ary
tum
orvo
lum
e (c
m3 )
Lung
wei
ght (
g)
Spl
een
wei
ght (
g)
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
Figure 1. Blood monocyte and neutrophil numbers in tumor-bearing mice. Monocytes and neutrophils were measured in peripheral blood of naïve mice(saline) or mice bearing mammary tumors. Ly6Chi monocytes (A), Ly6Clo monocytes (B), and neutrophils (C). Primary tumor growth (D), lung (E), andspleen weights (F) at endpoint. Data, cells/mL of blood (A–C), tumor volume (cm3; D), and grams (E and F). Data, mean � SEM, n ¼ 8 mice per group.A–C, �, P � 0.05; ��, P � 0.0001 4T1.2 versus 66cl4, 67NR and saline; ^, P � 0.05 4T1.2 versus 66cl4; #, P � 0.01 66cl4 versus 67NR and saline(two-way repeated measures ANOVA). E and F, ���, P � 0.001 and ����, P � 0.0001 4T1.2 versus 67NR (Mann–Whitney U test).
Anti–G-CSFR Blocks Metastasis Induced by Anti–CSF-1R
www.aacrjournals.org Cancer Immunol Res; 2(8) August 2014 767
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190
cells in human breast tumors (41, 42). Given that TAMs andneutrophils are the predominant immune cells in 4T1.2primary tumors, we assessed whether any changes took placein peripheral blood monocytes and neutrophils duringmammary tumor growth. Monocytes were defined asCD11bþCD115þLy6G�Ly6Chi or CD11bþCD115þLy6G�Ly6Clo
(23, 35, 43), and neutrophils as CD11bþLy6Gþ (21, 25, 35, 43).Compared with 66cl4 or 67NR tumor-bearing mice, those bear-ing highly metastatic 4T1.2 tumors had significant increases incirculating Ly6Chi and Ly6Clo monocytes, as well as neutrophils,fromday 20 onwards (Fig. 1A–C). It is important to note that thislarge increase in myeloid cells is not regularly reported in othertumor models (25, 44, 45). This is due to reporting of thepercentage of monocytes as opposed to the actual number incirculation as shown in Fig. 1. The difference in interpretation ofthe results is revealed in Supplementary Fig. S2, inwhich the twomethods are compared.
Although primary tumor growth was the same in allthree tumor types (Fig. 1D), end-stage lung and spleenweights were increased in 4T1.2 tumor-bearing mice (Fig.1E and F), indicating an enhanced metastatic burden (46).The extensive lung metastatic burden from 4T1.2 com-pared with 67NR tumor-bearing mice is evident in Sup-plementary Fig. S3.
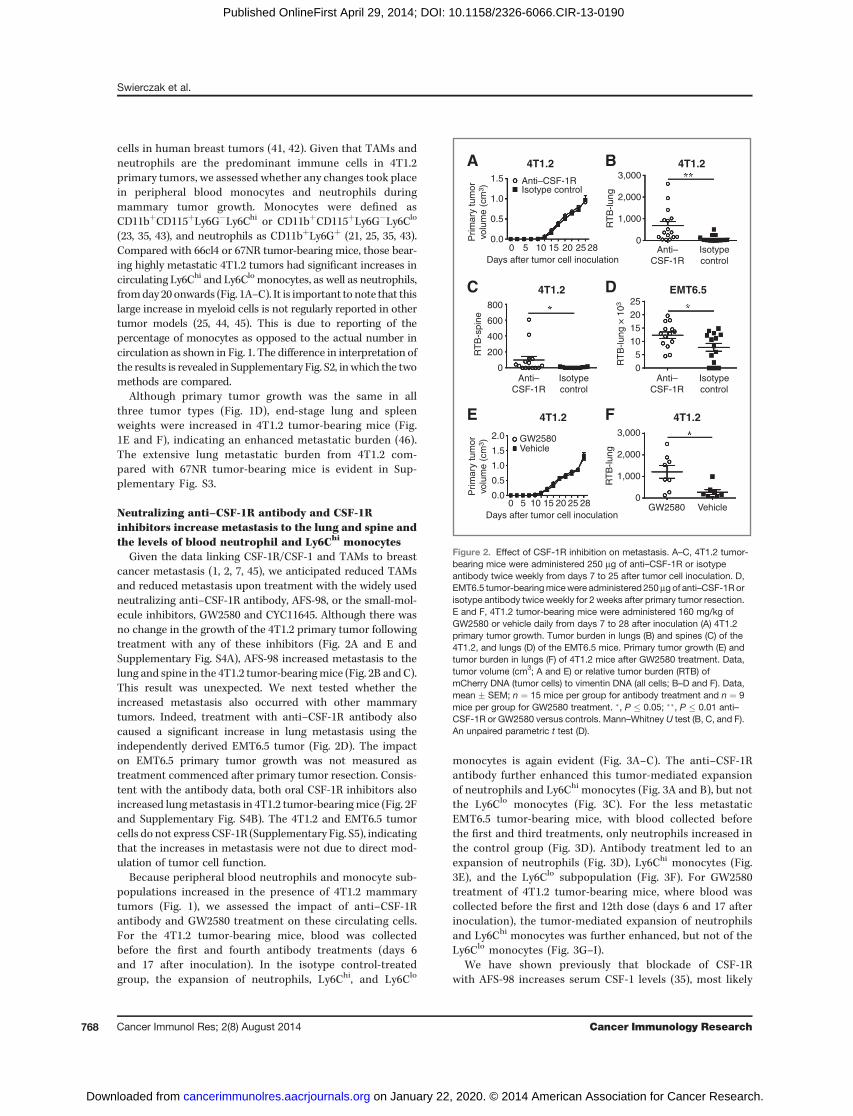
Neutralizing anti–CSF-1R antibody and CSF-1Rinhibitors increase metastasis to the lung and spine andthe levels of blood neutrophil and Ly6Chi monocytes
Given the data linking CSF-1R/CSF-1 and TAMs to breastcancer metastasis (1, 2, 7, 45), we anticipated reduced TAMsand reduced metastasis upon treatment with the widely usedneutralizing anti–CSF-1R antibody, AFS-98, or the small-mol-ecule inhibitors, GW2580 and CYC11645. Although there wasno change in the growth of the 4T1.2 primary tumor followingtreatment with any of these inhibitors (Fig. 2A and E andSupplementary Fig. S4A), AFS-98 increased metastasis to thelung and spine in the 4T1.2 tumor-bearingmice (Fig. 2B andC).This result was unexpected. We next tested whether theincreased metastasis also occurred with other mammarytumors. Indeed, treatment with anti–CSF-1R antibody alsocaused a significant increase in lung metastasis using theindependently derived EMT6.5 tumor (Fig. 2D). The impacton EMT6.5 primary tumor growth was not measured astreatment commenced after primary tumor resection. Consis-tent with the antibody data, both oral CSF-1R inhibitors alsoincreased lungmetastasis in 4T1.2 tumor-bearingmice (Fig. 2Fand Supplementary Fig. S4B). The 4T1.2 and EMT6.5 tumorcells do not express CSF-1R (Supplementary Fig. S5), indicatingthat the increases in metastasis were not due to direct mod-ulation of tumor cell function.
Because peripheral blood neutrophils and monocyte sub-populations increased in the presence of 4T1.2 mammarytumors (Fig. 1), we assessed the impact of anti–CSF-1Rantibody and GW2580 treatment on these circulating cells.For the 4T1.2 tumor-bearing mice, blood was collectedbefore the first and fourth antibody treatments (days 6and 17 after inoculation). In the isotype control-treatedgroup, the expansion of neutrophils, Ly6Chi, and Ly6Clo
monocytes is again evident (Fig. 3A–C). The anti–CSF-1Rantibody further enhanced this tumor-mediated expansionof neutrophils and Ly6Chi monocytes (Fig. 3A and B), but notthe Ly6Clo monocytes (Fig. 3C). For the less metastaticEMT6.5 tumor-bearing mice, with blood collected beforethe first and third treatments, only neutrophils increased inthe control group (Fig. 3D). Antibody treatment led to anexpansion of neutrophils (Fig. 3D), Ly6Chi monocytes (Fig.3E), and the Ly6Clo subpopulation (Fig. 3F). For GW2580treatment of 4T1.2 tumor-bearing mice, where blood wascollected before the first and 12th dose (days 6 and 17 afterinoculation), the tumor-mediated expansion of neutrophilsand Ly6Chi monocytes was further enhanced, but not of theLy6Clo monocytes (Fig. 3G–I).
We have shown previously that blockade of CSF-1Rwith AFS-98 increases serum CSF-1 levels (35), most likely
1.5
1.0
0.5
0.0
800
600
400
200
0
2520151050
3,000
2,000
1,000
0
Anti–CSF-1R
Anti–CSF-1R
Isotype control
Isotypecontrol
Anti–CSF-1R
Isotypecontrol
Anti–CSF-1R
GW2580 Vehicle
GW2580Vehicle
Isotypecontrol
A B
C D
E F
0 5 10 15Days after tumor cell inoculation
20 25 28
0 5 10 15Days after tumor cell inoculation
20 25 28
4T1.2 EMT6.5
4T1.24T1.2
4T1.2 4T1.2
Prim
ary
tum
orvo
lum
e (c
m3 )
2.0
1.5
1.0
0.5
0.0Prim
ary
tum
orvo
lum
e (c
m3 )
RT
B-lu
ng
3,000
2,000
1,000
0
RT
B-lu
ng
RT
B-s
pine
RT
B-lu
ng ×
103
Figure 2. Effect of CSF-1R inhibition on metastasis. A–C, 4T1.2 tumor-bearing mice were administered 250 mg of anti–CSF-1R or isotypeantibody twice weekly from days 7 to 25 after tumor cell inoculation. D,EMT6.5 tumor-bearingmicewere administered 250mgof anti–CSF-1Rorisotype antibody twice weekly for 2 weeks after primary tumor resection.E and F, 4T1.2 tumor-bearing mice were administered 160 mg/kg ofGW2580 or vehicle daily from days 7 to 28 after inoculation (A) 4T1.2primary tumor growth. Tumor burden in lungs (B) and spines (C) of the4T1.2, and lungs (D) of the EMT6.5 mice. Primary tumor growth (E) andtumor burden in lungs (F) of 4T1.2 mice after GW2580 treatment. Data,tumor volume (cm3; A and E) or relative tumor burden (RTB) ofmCherry DNA (tumor cells) to vimentin DNA (all cells; B–D and F). Data,mean � SEM; n ¼ 15 mice per group for antibody treatment and n ¼ 9mice per group for GW2580 treatment. �, P � 0.05; ��, P � 0.01 anti–CSF-1R or GW2580 versus controls. Mann–Whitney U test (B, C, and F).An unpaired parametric t test (D).
Swierczak et al.
Cancer Immunol Res; 2(8) August 2014 Cancer Immunology Research768
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190

due to a reduction in CSF-1R-mediated CSF-1 internaliza-tion by macrophage lineage cells (47, 48). Treatment of 4T1.2and EMT6.5 tumor-bearing mice with AFS-98 significantlyelevated CSF-1 levels in the serum (Fig. 3J and K), indicatingthat the antibody was also blocking CSF-1R in theseexperiments.
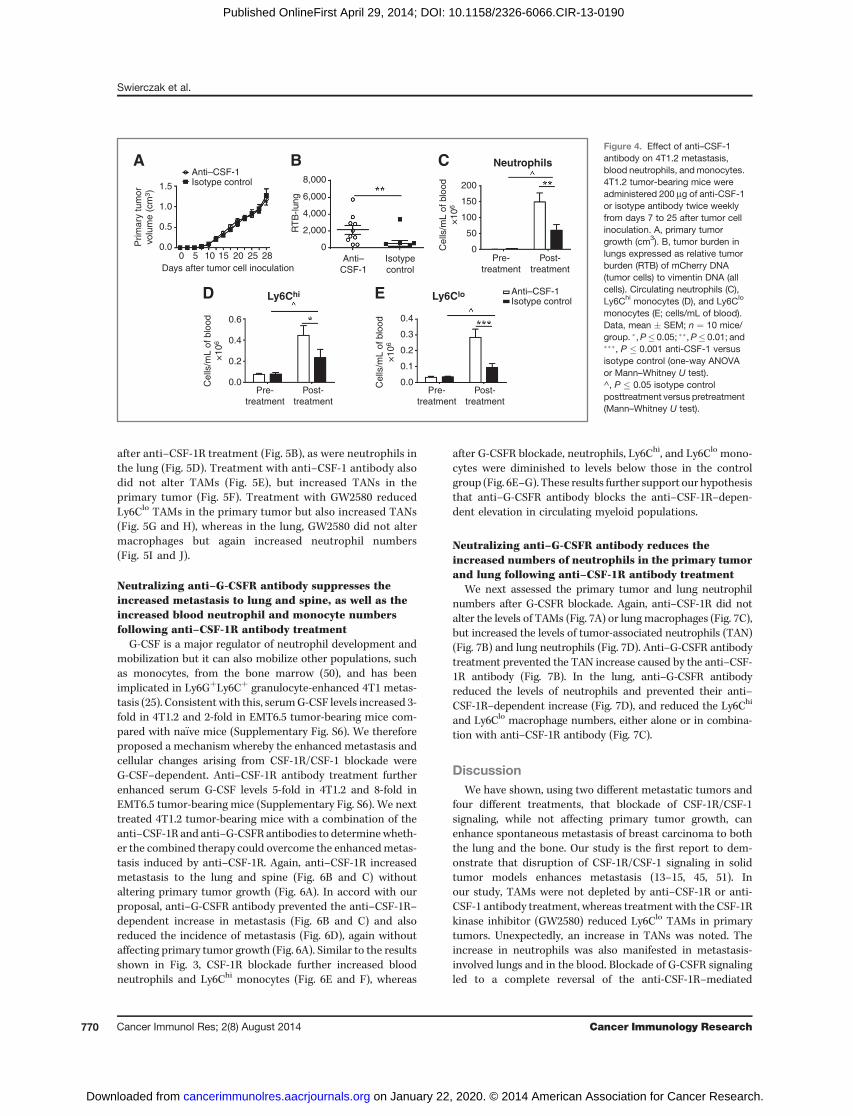
Neutralizing anti–CSF-1 antibody also increasesmetastasis to the lung and spine and further increasesblood neutrophil and monocyte numbersRecently, a second CSF-1R ligand, IL34, was identified (5,
6, 49). We therefore examined the impact of an anti–CSF-1antibody on metastasis. As with the anti–CSF-1R antibody,there was no change in the growth of the 4T1.2 primarytumor (Fig. 4A). Metastasis was increased to the lung (Fig.4B), but was not altered in the spine (data not shown). Bloodwas collected before the first and fourth antibody treat-ments. Expansion of neutrophils, Ly6Chi, and Ly6Clo mono-cytes was seen in the isotype control-treated 4T1.2 tumor-
bearing mice. Treatment with the anti–CSF-1 antibody signi-ficantly enhanced the tumor-mediated expansion of all threepopulations (Fig. 4C–E).
Neutralizing anti–CSF-1R and anti–CSF-1 antibodies, aswell as GW2580, increase neutrophil numbers in theprimary tumor and lung
We have reported previously that anti–CSF-1R antibody(AFS-98) reduces tissue macrophages in the steady state andduring inflammation (35). We therefore measured macro-phage and neutrophil numbers in 4T1.2 primary tumorsand lungs, following anti–CSF-1R (AFS-98), anti–CSF-1 anti-body, or GW2580 administration. Macrophages/TAMs werecharacterized as CD11bþCD115þLy6G�Ly6Chi or CD11bþ
CD115þLy6G�Ly6Clo (23), and neutrophils/tumor-associat-ed neutrophils (TAN) as CD11bþLy6Gþ (21, 25). At endpoint,treatment with anti–CSF-1R antibody did not reduce TAMsin primary tumors (Fig. 5A) or macrophages in the lung (Fig.5C). Surprisingly, TANs in primary tumors were elevated
200
100
0
150
100
50
0
1,500
1,000
500
0
1,000800600400200
0
8
6
4
2
0
15
10
5
0
2
1
0Pre-
treatmentPost-
treatmentPre-
treatmentPost-
treatmentPre-
treatmentPost-
treatment
Pre-treatment
Post-treatment
Pre-treatment
Post-treatment
Pre-treatment
Post-treatment
Pre-treatment
Post-treatment
Pre-treatment
Post-treatment
Pre-treatment
Post-treatment
0.80.60.40.20.0
0.8
0.6
0.4
0.2
0.0
Ly6ChiA B C
D E F
G H
J K
I
Ly6CloNeutrophils
Ly6Chi Ly6CloNeutrophils
Ly6Chi Ly6CloNeutrophils
4T1.2
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
1.5
1.0
0.5
0.0
Anti–CSF-1RIsotype control
Anti–CSF-1R
CS
F-1
(pg
/mL)
CS
F-1
(pg
/mL)
Isotype control
Anti–CSF-1R
GW2580Vehicle
NS
Isotypecontrol
Anti–CSF-1R
Isotypecontrol
4T1.2
4T1.2
EMT6.5
EMT6.5
2.0
1.5
1.0
0.5
0.0
Figure 3. Effect ofCSF-1R inhibitionon blood neutrophils andmonocytes in mice bearingmetastatic tumors. 4T1.2 orEMT6.5 tumor-bearing mice wereadministered anti–CSF-1Rantibody or GW2580 as describedin Fig. 2. Peripheral blood wascollected 24 hours before the first(pretreatment) and 24 hours beforethe third (EMT6.5) or fourth (4T1.2)antibody treatment(posttreatment), andbefore the firstand 12th GW2580 treatment. A, D,and G, neutrophils; B, E, and H,Ly6Chi monocytes; C, F, and I,Ly6Clo monocytes in 4T1.2 (A–C),EMT6.5 (D–F) tumor-bearing miceafter antibody treatment and in4T1.2 (G–I) tumor-bearing miceafter GW2580 treatment. Endpointserum CSF-1 levels from 4T1.2 (J)or EMT6.5 (K) tumor-bearing mice.Data, cells/mL of blood (A–I) or aspg/mL (J and K). Data, mean �SEM; n ¼ 15 mice/group forantibody treatment, n ¼ 9 mice/group for GW2580 treatment, andn ¼ 7 mice/group for ELISA.�, P � 0.05; ��, P � 0.01;���, P � 0.001 antibody or inhibitorversus controls (one-way ANOVAor Mann–Whitney U test).^, P � 0.05; ^^, P � 0.01 controlposttreatment versus pretreatment(Mann–Whitney U test).
Anti–G-CSFR Blocks Metastasis Induced by Anti–CSF-1R
www.aacrjournals.org Cancer Immunol Res; 2(8) August 2014 769
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190
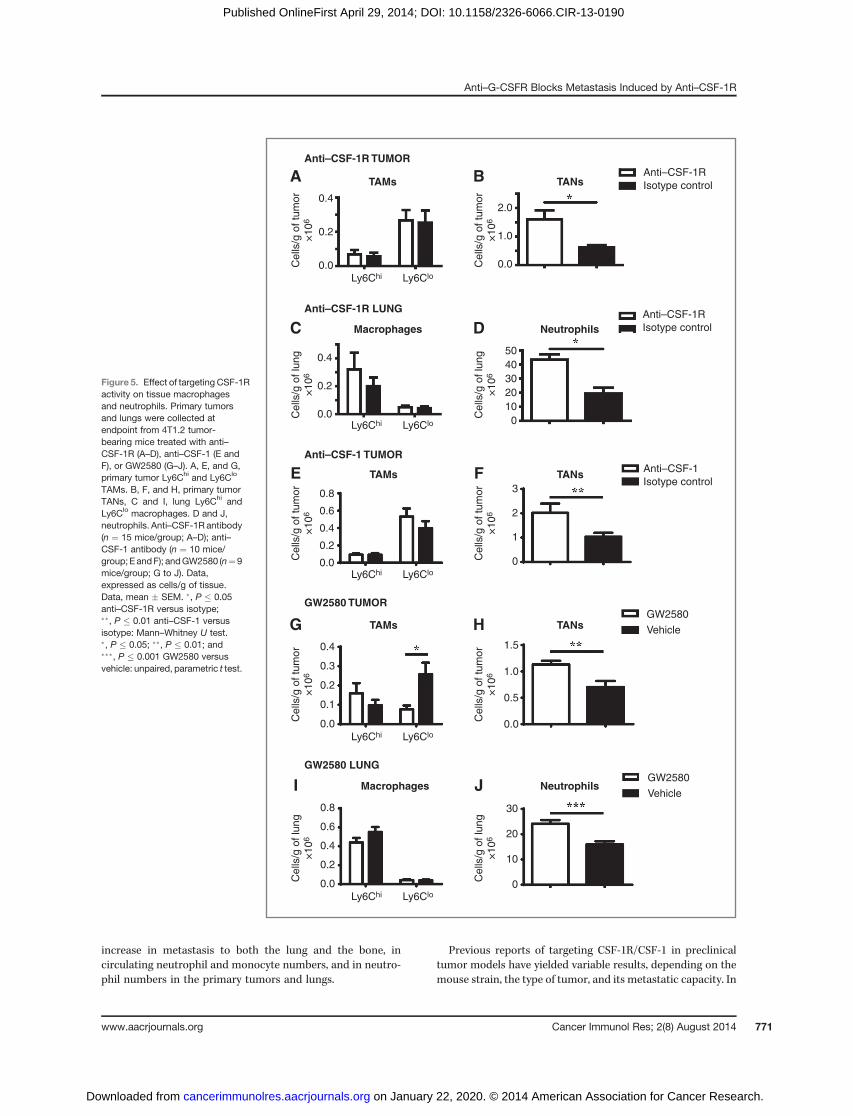
after anti–CSF-1R treatment (Fig. 5B), as were neutrophils inthe lung (Fig. 5D). Treatment with anti–CSF-1 antibody alsodid not alter TAMs (Fig. 5E), but increased TANs in theprimary tumor (Fig. 5F). Treatment with GW2580 reducedLy6Clo TAMs in the primary tumor but also increased TANs(Fig. 5G and H), whereas in the lung, GW2580 did not altermacrophages but again increased neutrophil numbers(Fig. 5I and J).
Neutralizing anti–G-CSFR antibody suppresses theincreased metastasis to lung and spine, as well as theincreased blood neutrophil and monocyte numbersfollowing anti–CSF-1R antibody treatment
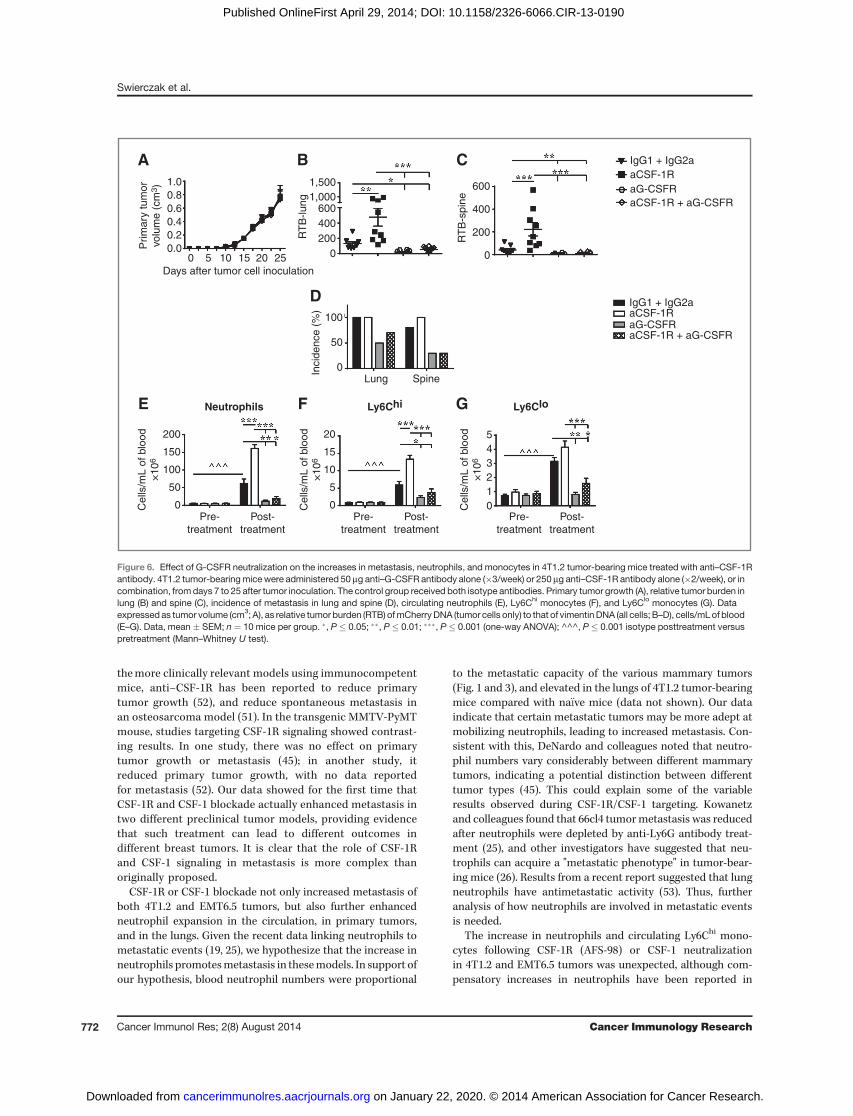
G-CSF is a major regulator of neutrophil development andmobilization but it can also mobilize other populations, suchas monocytes, from the bone marrow (50), and has beenimplicated in Ly6GþLy6Cþ granulocyte-enhanced 4T1 metas-tasis (25). Consistent with this, serumG-CSF levels increased 3-fold in 4T1.2 and 2-fold in EMT6.5 tumor-bearing mice com-pared with na€�ve mice (Supplementary Fig. S6). We thereforeproposed a mechanism whereby the enhanced metastasis andcellular changes arising from CSF-1R/CSF-1 blockade wereG-CSF–dependent. Anti–CSF-1R antibody treatment furtherenhanced serum G-CSF levels 5-fold in 4T1.2 and 8-fold inEMT6.5 tumor-bearing mice (Supplementary Fig. S6). We nexttreated 4T1.2 tumor-bearing mice with a combination of theanti–CSF-1R and anti–G-CSFR antibodies to determinewheth-er the combined therapy could overcome the enhancedmetas-tasis induced by anti–CSF-1R. Again, anti–CSF-1R increasedmetastasis to the lung and spine (Fig. 6B and C) withoutaltering primary tumor growth (Fig. 6A). In accord with ourproposal, anti–G-CSFR antibody prevented the anti–CSF-1R–dependent increase in metastasis (Fig. 6B and C) and alsoreduced the incidence of metastasis (Fig. 6D), again withoutaffecting primary tumor growth (Fig. 6A). Similar to the resultsshown in Fig. 3, CSF-1R blockade further increased bloodneutrophils and Ly6Chi monocytes (Fig. 6E and F), whereas
after G-CSFR blockade, neutrophils, Ly6Chi, and Ly6Clo mono-cytes were diminished to levels below those in the controlgroup (Fig. 6E–G). These results further support our hypothesisthat anti–G-CSFR antibody blocks the anti–CSF-1R–depen-dent elevation in circulating myeloid populations.
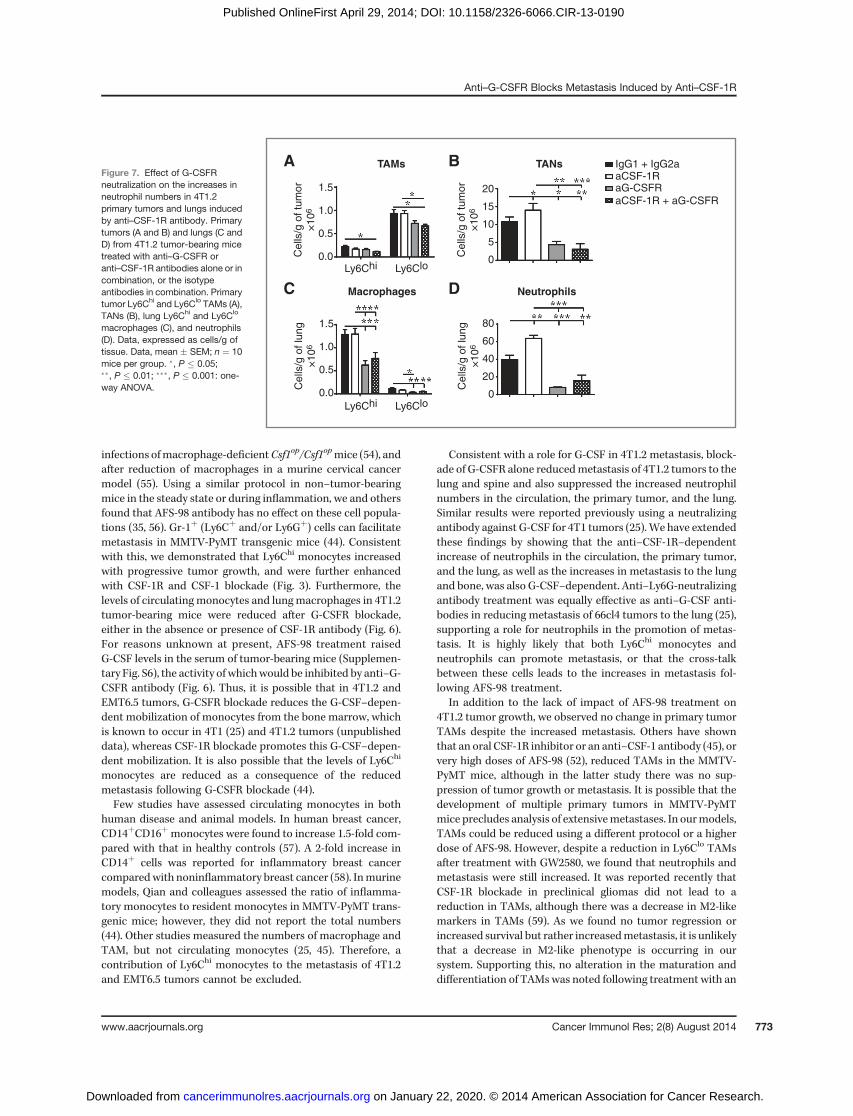
Neutralizing anti–G-CSFR antibody reduces theincreased numbers of neutrophils in the primary tumorand lung following anti–CSF-1R antibody treatment
We next assessed the primary tumor and lung neutrophilnumbers after G-CSFR blockade. Again, anti–CSF-1R did notalter the levels of TAMs (Fig. 7A) or lungmacrophages (Fig. 7C),but increased the levels of tumor-associated neutrophils (TAN)(Fig. 7B) and lung neutrophils (Fig. 7D). Anti–G-CSFR antibodytreatment prevented the TAN increase caused by the anti–CSF-1R antibody (Fig. 7B). In the lung, anti–G-CSFR antibodyreduced the levels of neutrophils and prevented their anti–CSF-1R–dependent increase (Fig. 7D), and reduced the Ly6Chi
and Ly6Clo macrophage numbers, either alone or in combina-tion with anti–CSF-1R antibody (Fig. 7C).
DiscussionWe have shown, using two different metastatic tumors and
four different treatments, that blockade of CSF-1R/CSF-1signaling, while not affecting primary tumor growth, canenhance spontaneous metastasis of breast carcinoma to boththe lung and the bone. Our study is the first report to dem-onstrate that disruption of CSF-1R/CSF-1 signaling in solidtumor models enhances metastasis (13–15, 45, 51). Inour study, TAMs were not depleted by anti–CSF-1R or anti-CSF-1 antibody treatment, whereas treatment with the CSF-1Rkinase inhibitor (GW2580) reduced Ly6Clo TAMs in primarytumors. Unexpectedly, an increase in TANs was noted. Theincrease in neutrophils was also manifested in metastasis-involved lungs and in the blood. Blockade of G-CSFR signalingled to a complete reversal of the anti-CSF-1R–mediated
200
150
100
50
0
Pre-treatment
Post-treatment
Pre-treatment
Post-treatment
Pre-treatment
Post-treatment
0.6
0.4
0.2
0.0
Neutrophils
Ly6Chi Ly6Clo
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
0.4
0.3
0.2
0.1
0.0Cel
ls/m
L of
blo
od×
106
Anti–CSF-1Isotype control
Anti–CSF-1Isotype control
Anti–CSF-1
Isotypecontrol
1.5
1.0
0.5
0.0
8,000
6,000
4,000
2,000
0
A B C
D E
0 5 10 15Days after tumor cell inoculation
20 25 28
Prim
ary
tum
orvo
lum
e (c
m3 )
RT
B-lu
ng
Figure 4. Effect of anti–CSF-1antibody on 4T1.2 metastasis,blood neutrophils, andmonocytes.4T1.2 tumor-bearing mice wereadministered 200 mg of anti-CSF-1or isotype antibody twice weeklyfrom days 7 to 25 after tumor cellinoculation. A, primary tumorgrowth (cm3). B, tumor burden inlungs expressed as relative tumorburden (RTB) of mCherry DNA(tumor cells) to vimentin DNA (allcells). Circulating neutrophils (C),Ly6Chi monocytes (D), and Ly6Clo
monocytes (E; cells/mL of blood).Data, mean � SEM; n ¼ 10 mice/group. �,P� 0.05; ��,P� 0.01; and���, P � 0.001 anti-CSF-1 versusisotype control (one-way ANOVAor Mann–Whitney U test).^, P � 0.05 isotype controlposttreatment versus pretreatment(Mann–Whitney U test).
Swierczak et al.
Cancer Immunol Res; 2(8) August 2014 Cancer Immunology Research770
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190
increase in metastasis to both the lung and the bone, incirculating neutrophil and monocyte numbers, and in neutro-phil numbers in the primary tumors and lungs.
Previous reports of targeting CSF-1R/CSF-1 in preclinicaltumor models have yielded variable results, depending on themouse strain, the type of tumor, and its metastatic capacity. In
NeutrophilsMacrophages
TAMs TANs
TAMs TANs
TAMs TANs
NeutrophilsMacrophages
Ly6Chi Ly6Clo
Ly6Chi Ly6Clo
Ly6Chi Ly6Clo
Ly6Chi Ly6Clo
Ly6Chi Ly6Clo
Cel
ls/g
of t
umor
×10
6
Cel
ls/g
of t
umor
×10
6
Cel
ls/g
of t
umor
×10
6
Cel
ls/g
of t
umor
×10
6
Cel
ls/g
of t
umor
×10
6
Cel
ls/g
of t
umor
×10
6
Cel
ls/g
of l
ung
×10
6
Cel
ls/g
of l
ung
×10
6
Cel
ls/g
of l
ung
×10
6
Cel
ls/g
of l
ung
×10
6
Anti–CSF-1RIsotype control
Anti–CSF-1RIsotype control
Anti–CSF-1Isotype control
Anti–CSF-1R TUMOR
Anti–CSF-1 TUMOR
GW2580 TUMORGW2580
Vehicle
GW2580
Vehicle
GW2580 LUNG
Anti–CSF-1R LUNG
0.4
0.2
0.0
0.4
0.2
0.0
0.8
0.6
0.4
0.2
0.0
0.8
0.6
0.4
0.2
0.0
30
20
10
0
0.4
0.3
0.2
0.1
0.0
1.5
1.0
0.5
0.0
3
2
1
0
50403020100
2.0
1.0
0.0
A B
C D
E F
G H
I J
Figure 5. Effect of targeting CSF-1Ractivity on tissue macrophagesand neutrophils. Primary tumorsand lungs were collected atendpoint from 4T1.2 tumor-bearing mice treated with anti–CSF-1R (A–D), anti–CSF-1 (E andF), or GW2580 (G–J). A, E, and G,primary tumor Ly6Chi and Ly6Clo
TAMs. B, F, and H, primary tumorTANs, C and I, lung Ly6Chi andLy6Clo macrophages. D and J,neutrophils. Anti–CSF-1R antibody(n ¼ 15 mice/group; A–D); anti–CSF-1 antibody (n ¼ 10 mice/group; E andF); andGW2580 (n¼9mice/group; G to J). Data,expressed as cells/g of tissue.Data, mean � SEM. �, P � 0.05anti–CSF-1R versus isotype;��, P � 0.01 anti–CSF-1 versusisotype: Mann–Whitney U test.�, P � 0.05; ��, P � 0.01; and���, P � 0.001 GW2580 versusvehicle: unpaired, parametric t test.
Anti–G-CSFR Blocks Metastasis Induced by Anti–CSF-1R
www.aacrjournals.org Cancer Immunol Res; 2(8) August 2014 771
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190
themore clinically relevant models using immunocompetentmice, anti–CSF-1R has been reported to reduce primarytumor growth (52), and reduce spontaneous metastasis inan osteosarcoma model (51). In the transgenic MMTV-PyMTmouse, studies targeting CSF-1R signaling showed contrast-ing results. In one study, there was no effect on primarytumor growth or metastasis (45); in another study, itreduced primary tumor growth, with no data reportedfor metastasis (52). Our data showed for the first time thatCSF-1R and CSF-1 blockade actually enhanced metastasis intwo different preclinical tumor models, providing evidencethat such treatment can lead to different outcomes indifferent breast tumors. It is clear that the role of CSF-1Rand CSF-1 signaling in metastasis is more complex thanoriginally proposed.
CSF-1R or CSF-1 blockade not only increased metastasis ofboth 4T1.2 and EMT6.5 tumors, but also further enhancedneutrophil expansion in the circulation, in primary tumors,and in the lungs. Given the recent data linking neutrophils tometastatic events (19, 25), we hypothesize that the increase inneutrophils promotesmetastasis in thesemodels. In support ofour hypothesis, blood neutrophil numbers were proportional
to the metastatic capacity of the various mammary tumors(Fig. 1 and 3), and elevated in the lungs of 4T1.2 tumor-bearingmice compared with na€�ve mice (data not shown). Our dataindicate that certain metastatic tumors may be more adept atmobilizing neutrophils, leading to increased metastasis. Con-sistent with this, DeNardo and colleagues noted that neutro-phil numbers vary considerably between different mammarytumors, indicating a potential distinction between differenttumor types (45). This could explain some of the variableresults observed during CSF-1R/CSF-1 targeting. Kowanetzand colleagues found that 66cl4 tumormetastasis was reducedafter neutrophils were depleted by anti-Ly6G antibody treat-ment (25), and other investigators have suggested that neu-trophils can acquire a "metastatic phenotype" in tumor-bear-ing mice (26). Results from a recent report suggested that lungneutrophils have antimetastatic activity (53). Thus, furtheranalysis of how neutrophils are involved in metastatic eventsis needed.
The increase in neutrophils and circulating Ly6Chi mono-cytes following CSF-1R (AFS-98) or CSF-1 neutralizationin 4T1.2 and EMT6.5 tumors was unexpected, although com-pensatory increases in neutrophils have been reported in
Pre-treatment
Post-treatment
Pre-treatment
Post-treatment
Pre-treatment
Post-treatment
200
150
100
50
0
20
15
10
5
0
543210
Neutrophils Ly6Chi Ly6Clo
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
Cel
ls/m
L of
blo
od×
106
aCSF-1R + aG-CSFRaG-CSFRaCSF-1RIgG1 + IgG2a
aCSF-1R + aG-CSFRaG-CSFRaCSF-1RIgG1 + IgG2a
1.00.80.60.40.20.0
1,5001,000
600400200
0
600
400
200
0
100
50
0
A B C
E F G
D
0 5 10 15Days after tumor cell inoculation
Inci
denc
e (%
)
20 25
Lung Spine
Prim
ary
tum
orvo
lum
e (c
m3 )
RT
B-lu
ng
RT
B-s
pine
Figure 6. Effect of G-CSFR neutralization on the increases in metastasis, neutrophils, and monocytes in 4T1.2 tumor-bearing mice treated with anti–CSF-1Rantibody. 4T1.2 tumor-bearingmice were administered 50 mg anti–G-CSFR antibody alone (�3/week) or 250 mg anti–CSF-1R antibody alone (�2/week), or incombination, fromdays 7 to 25 after tumor inoculation. The control group received both isotype antibodies. Primary tumor growth (A), relative tumor burden inlung (B) and spine (C), incidence of metastasis in lung and spine (D), circulating neutrophils (E), Ly6Chi monocytes (F), and Ly6Clo monocytes (G). Dataexpressed as tumor volume (cm3; A), as relative tumor burden (RTB) ofmCherryDNA (tumor cells only) to that of vimentinDNA (all cells; B–D), cells/mLof blood(E–G). Data, mean � SEM; n ¼ 10 mice per group. �, P � 0.05; ��, P � 0.01; ���, P � 0.001 (one-way ANOVA); ^^^, P � 0.001 isotype posttreatment versuspretreatment (Mann–Whitney U test).
Swierczak et al.
Cancer Immunol Res; 2(8) August 2014 Cancer Immunology Research772
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190
infections ofmacrophage-deficientCsf1op/Csf1opmice (54), andafter reduction of macrophages in a murine cervical cancermodel (55). Using a similar protocol in non–tumor-bearingmice in the steady state or during inflammation, we and othersfound that AFS-98 antibody has no effect on these cell popula-tions (35, 56). Gr-1þ (Ly6Cþ and/or Ly6Gþ) cells can facilitatemetastasis in MMTV-PyMT transgenic mice (44). Consistentwith this, we demonstrated that Ly6Chi monocytes increasedwith progressive tumor growth, and were further enhancedwith CSF-1R and CSF-1 blockade (Fig. 3). Furthermore, thelevels of circulatingmonocytes and lungmacrophages in 4T1.2tumor-bearing mice were reduced after G-CSFR blockade,either in the absence or presence of CSF-1R antibody (Fig. 6).For reasons unknown at present, AFS-98 treatment raisedG-CSF levels in the serum of tumor-bearing mice (Supplemen-tary Fig. S6), the activity of whichwould be inhibited by anti–G-CSFR antibody (Fig. 6). Thus, it is possible that in 4T1.2 andEMT6.5 tumors, G-CSFR blockade reduces the G-CSF–depen-dent mobilization of monocytes from the bone marrow, whichis known to occur in 4T1 (25) and 4T1.2 tumors (unpublisheddata), whereas CSF-1R blockade promotes this G-CSF–depen-dent mobilization. It is also possible that the levels of Ly6Chi
monocytes are reduced as a consequence of the reducedmetastasis following G-CSFR blockade (44).Few studies have assessed circulating monocytes in both
human disease and animal models. In human breast cancer,CD14þCD16þmonocytes were found to increase 1.5-fold com-pared with that in healthy controls (57). A 2-fold increase inCD14þ cells was reported for inflammatory breast cancercomparedwith noninflammatory breast cancer (58). Inmurinemodels, Qian and colleagues assessed the ratio of inflamma-tory monocytes to resident monocytes in MMTV-PyMT trans-genic mice; however, they did not report the total numbers(44). Other studies measured the numbers of macrophage andTAM, but not circulating monocytes (25, 45). Therefore, acontribution of Ly6Chi monocytes to the metastasis of 4T1.2and EMT6.5 tumors cannot be excluded.
Consistent with a role for G-CSF in 4T1.2 metastasis, block-ade of G-CSFR alone reducedmetastasis of 4T1.2 tumors to thelung and spine and also suppressed the increased neutrophilnumbers in the circulation, the primary tumor, and the lung.Similar results were reported previously using a neutralizingantibody against G-CSF for 4T1 tumors (25). We have extendedthese findings by showing that the anti–CSF-1R–dependentincrease of neutrophils in the circulation, the primary tumor,and the lung, as well as the increases in metastasis to the lungand bone, was also G-CSF–dependent. Anti–Ly6G-neutralizingantibody treatment was equally effective as anti–G-CSF anti-bodies in reducing metastasis of 66cl4 tumors to the lung (25),supporting a role for neutrophils in the promotion of metas-tasis. It is highly likely that both Ly6Chi monocytes andneutrophils can promote metastasis, or that the cross-talkbetween these cells leads to the increases in metastasis fol-lowing AFS-98 treatment.
In addition to the lack of impact of AFS-98 treatment on4T1.2 tumor growth, we observed no change in primary tumorTAMs despite the increased metastasis. Others have shownthat an oral CSF-1R inhibitor or an anti–CSF-1 antibody (45), orvery high doses of AFS-98 (52), reduced TAMs in the MMTV-PyMT mice, although in the latter study there was no sup-pression of tumor growth or metastasis. It is possible that thedevelopment of multiple primary tumors in MMTV-PyMTmice precludes analysis of extensivemetastases. In ourmodels,TAMs could be reduced using a different protocol or a higherdose of AFS-98. However, despite a reduction in Ly6Clo TAMsafter treatment with GW2580, we found that neutrophils andmetastasis were still increased. It was reported recently thatCSF-1R blockade in preclinical gliomas did not lead to areduction in TAMs, although there was a decrease in M2-likemarkers in TAMs (59). As we found no tumor regression orincreased survival but rather increasedmetastasis, it is unlikelythat a decrease in M2-like phenotype is occurring in oursystem. Supporting this, no alteration in the maturation anddifferentiation of TAMswas noted following treatment with an
1.5
1.0
0.5
0.0
20
15
10
5
0
80
60
40
20
0
NeutrophilsMacrophages
TAMs TANs
Ly6Chi Ly6Clo
Ly6Chi Ly6Clo
Cel
ls/g
of t
umor
×10
6
1.5
1.0
0.5
0.0
Cel
ls/g
of l
ung
×10
6
Cel
ls/g
of t
umor
×10
6C
ells
/g o
f lun
g×
106
aCSF-1R + aG-CSFRaG-CSFRaCSF-1RIgG1 + IgG2aA B
C D
Figure 7. Effect of G-CSFRneutralization on the increases inneutrophil numbers in 4T1.2primary tumors and lungs inducedby anti–CSF-1R antibody. Primarytumors (A and B) and lungs (C andD) from 4T1.2 tumor-bearing micetreated with anti–G-CSFR oranti–CSF-1R antibodies alone or incombination, or the isotypeantibodies in combination. Primarytumor Ly6Chi and Ly6Clo TAMs (A),TANs (B), lung Ly6Chi and Ly6Clo
macrophages (C), and neutrophils(D). Data, expressed as cells/g oftissue. Data, mean � SEM; n ¼ 10mice per group. �, P � 0.05;��, P � 0.01; ���, P � 0.001: one-way ANOVA.
Anti–G-CSFR Blocks Metastasis Induced by Anti–CSF-1R
www.aacrjournals.org Cancer Immunol Res; 2(8) August 2014 773
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190

anti–CSF-1 antibody or an oral CSF-1R inhibitor in theMMTV-PyMTmodel (45). To account for the lack of reduction in TAMsfollowing CSF-1R blockade, Pyonteck and colleagues suggestedthat TAMs were protected from cell death by survival factorsproduced by the tumor cells (59). A similar mechanismmay beactive in 4T1.2 tumors. In our study, TAM function could alsobe altered such that their influence on neutrophil trafficking ischanged, either through G-CSF elevation or through a com-promised ability to clear neutrophils by phagocytosis (60).
On the basis of our results on metastasis, we assessed theprognostic significance of CSF-1R expression in human breasttumors. In contrast with previous reported data (7–10), lowlevels of the CSF1R gene were found to predict worse overallsurvival using both Kaplan–Meier plotter (39) and BreastMark(40) online survival analyses (Supplementary Fig. S7). Likewise,a recent study by Beck and colleagues found that a "CSF-1response signature" predicted different outcomes for patientswith breast cancer depending on the tumor subtype (11). Theauthors concluded that the relationship between the CSF-1response signature and survival is complex and requires fur-ther investigation. Furthermore, CSF-1 levels in the circulationcan also be prognostic (61). Analysis of CSF-1R and CSF-1 intumors and in the circulation, comparing distant metastasisversus local recurrence in different subtypes of breast cancer,may shed more light on how CSF-1R and CSF-1 are related toprognosis.
The blockade of CSF-1R or CSF-1 signaling leading toincreased metastasis in 4T1.2 and EMT6.5 mammary tumorsraises a cautionary note for the use of this type of therapy, evenin an adjuvant setting, for patients at high risk of metastaticprogression. Although the CSF-1/CSF-1R signaling pathwaymay still prove to be a valid target in cancer, our data indicatethat patients may have variable responses to this treatment. Itmay be prudent to stratify patients based on the properties andfeatures of the primary lesion (45), such asmyeloid cell content,or possibly by assessing circulating myeloid cell numbers, orsystemic CSF-1 levels to identify patients who aremost likely torespond to CSF-1- and CSF-1R–targeted therapies. It is possi-
ble that patients with progressive disease may respond betterto anti–G-CSFR therapies.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: A. Swierczak, R.L. AndersonDevelopment of methodology: A. Swierczak, J.C. Lenzo, J.P. Doherty, R.L.AndersonAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.):A. Swierczak, C.M. Restall, J.P. Doherty, R.L. Anderson,J.A. HamiltonAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): A. Swierczak, A.D. Cook, C.M. Restall, R.L. Anderson,J.A. HamiltonWriting, review, and/or revisionof themanuscript:A. Swierczak, J.C. Lenzo,R.L. AndersonAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): A.D. Cook, C.M. Restall, J.A. HamiltonStudy supervision: A.D. Cook, R.L. Anderson
AcknowledgmentsThe authors thank S.-I. Nishikawa (RIKEN Center for Developmental Biology,
Japan; AFS-98 hybridoma), J. Abrams (DNAX, USA; IgG2a isotype controlhybridoma), A. Andrews (CSL Pty Ltd; anti–G-CSFR and the IgG1 isotypecontrol), and F. Dodeller (MorphoSys AG; anti–CSF-1 and the IgG2a isotypecontrol) for kind gifts of antibodies. Dr. C. Burns, previously at Cytopia, now atWalter and Eliza Hall Institute of Medical Research, provided CYC11645 andvehicle, and L. Phillipson, G. Nikolakopoulos, and J. Alcindor, Cancer Therapeu-tics CRC prepared the GW2580. They also thank Dr. Yuan Cao (Peter MacCallumCancer Centre) for assistance with G-CSF measurements, Ralph Rossi, VikiMilovac, and Sophie Curcio from the Peter MacCallum Flow Cytometry facility,and Lauren Dawes, Katherine Papastratos, and Jacinta Carter from the PeterMacCallum Animal Core facility for their assistance.
Grant SupportThis project was supported by a grant from the National Health & Medical
Research Council (NHMRC) of Australia (APP100880). J.A. Hamilton receivedfellowship from the NHMRC and R.L. Anderson from the National Breast CancerFoundation of Australia. A. Swierczak received postgraduate NHMRCscholarship.
The costs of publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received October 25, 2013; revised March 27, 2014; accepted April 17, 2014;published OnlineFirst April 29, 2014.
References1. Allavena P, Sica A, Solinas G, Porta C, Mantovani A. The inflammatory
micro-environment in tumor progression: the role of tumor-associatedmacrophages. Crit Rev Oncol Hematol 2008;66:1–9.
2. Pollard JW. Tumour-educated macrophages promote tumour pro-gression and metastasis. Nat Rev Cancer 2004;4:71–8.
3. Pixley FJ, Stanley ER.CSF-1 regulation of thewanderingmacrophage:complexity in action. Trends Cell Biol 2004;14:628–38.
4. Byrne PV, Guilbert LJ, Stanley ER. Distribution of cells bearing recep-tors for a colony-stimulating factor (CSF-1) inmurine tissues. JCell Biol1981;91:848–53.
5. WangY, Szretter KJ, VermiW,Gilfillan S, Rossini C,CellaM, et al. IL-34is a tissue-restricted ligand of CSF1R required for the development ofLangerhans cells and microglia. Nat Immunol 2012;13:753–60.
6. Wei S, Nandi S, Chitu V, Yeung YG, Yu W, Huang M, et al.Functional overlap but differential expression of CSF-1 and IL-34in their CSF-1 receptor-mediated regulation of myeloid cells.J Leukoc Biol 2010;88:495–505.
7. Scholl SM, Pallud C, Beuvon F, Hacene K, Stanley ER, RohrschneiderL, et al. Anti-colony-stimulating factor-1 antibody staining in primary
breast adenocarcinomas correlates with marked inflammatory cellinfiltrates and prognosis. J Natl Cancer Inst 1994;86:120–6.
8. Tang R, Beuvon F, Ojeda M, Mosseri V, Pouillart P, Scholl S. M-CSF(monocyte colony stimulating factor) and M-CSF receptor expressionby breast tumour cells: M-CSF mediated recruitment of tumour infil-trating monocytes? J Cell Biochem 1992;50:350–6.
9. Maher MG, Sapi E, Turner B, Gumbs A, Perrotta PL, Carter D, et al.Prognostic significance of colony-stimulating factor receptor expres-sion in ipsilateral breast cancer recurrence. Clin Cancer Res 1998;4:1851–6.
10. Kluger HM, Dolled-Filhart M, Rodov S, Kacinski BM, Camp RL, RimmDL. Macrophage colony-stimulating factor-1 receptor expression isassociated with poor outcome in breast cancer by large cohort tissuemicroarray analysis. Clin Cancer Res 2004;10:173–7.
11. Beck AH, Espinosa I, Edris B, Li R, Montgomery K, Zhu S, et al. Themacrophage colony-stimulating factor 1 response signature in breastcarcinoma. Clin Cancer Res 2009;15:778–87.
12. Mantovani A, Sozzani S, Locati M, Allavena P, Sica A. Macrophagepolarization: tumor-associated macrophages as a paradigm for
Swierczak et al.
Cancer Immunol Res; 2(8) August 2014 Cancer Immunology Research774
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190

polarized M2 mononuclear phagocytes. Trends Immunol 2002;23:549–55.
13. Lin EY, Nguyen AV, Russell RG, Pollard JW. Colony-stimulating factor1 promotes progression of mammary tumors to malignancy. J ExpMed 2001;193:727–40.
14. Aharinejad S, Paulus P, Sioud M, Hofmann M, Zins K, Schafer R, et al.Colony-stimulating factor-1 blockade by antisense oligonucleotidesand small interfering RNAs suppresses growth of human mammarytumor xenografts in mice. Cancer Res 2004;64:5378–84.
15. Paulus P, Stanley ER, Schafer R, Abraham D, Aharinejad S. Colony-stimulating factor-1 antibody reverses chemoresistance inhuman MCF-7 breast cancer xenografts. Cancer Res 2006;66:4349–56.
16. ImClone LLC. Study of IMC-CS4 in subjects with advanced solidtumors; 2011 [cited 2014 May 5]. Available from: http://clinicaltrials.gov/show/NCT01346358.
17. Plexxikon. Safety study of PLX108-01 in patients with solid tumors;2009 [cited 2014 May 5]. Available from: http://clinicaltrials.gov/ct2/show/NCT01004861?term¼PLX3397þandþcancer&rank¼4.
18. Rugo H. Komen, Plexxikon, Eisai Inc. Phase Ib/II study of PLX 3397and eribulin in patients with metastatic breast cancer; 2012 [cited2014 May 5]. Available from: http://clinicaltrials.gov/ct2/show/NCT01596751?term¼PLX3397þandþcancer&rank¼1.
19. Yan HH, Pickup M, Pang Y, Gorska AE, Li Z, Chytil A, et al. Gr-1þCD11bþ myeloid cells tip the balance of immune protection totumor promotion in the premetastatic lung. Cancer Res 2010;70:6139–49.
20. Almand B, Clark JI, Nikitina E, van Beynen J, English NR, Knight SC,et al. Increased production of immature myeloid cells in cancerpatients: a mechanism of immunosuppression in cancer. J Immunol2001;166:678–89.
21. Fleming TJ, Fleming ML, Malek TR. Selective expression of Ly-6G onmyeloid lineage cells in mouse bone marrow. RB6-8C5 mAb togranulocyte-differentiation antigen (Gr-1) detects members of theLy-6 family. J Immunol 1993;151:2399–408.
22. Tacke F, Randolph GJ. Migratory fate and differentiation of bloodmonocyte subsets. Immunobiology 2006;211:609–18.
23. Chan J, Leenen PJ, Bertoncello I, Nishikawa SI, Hamilton JA. Macro-phage lineage cells in inflammation: characterization by colony-stim-ulating factor-1 (CSF-1) receptor (c-Fms), ER-MP58, and ER-MP20(Ly-6C) expression. Blood 1998;92:1423–31.
24. Geissmann F, Manz MG, Jung S, Sieweke MH, Merad M, Ley K.Development of monocytes, macrophages, and dendritic cells. Sci-ence 2010;327:656–61.
25. Kowanetz M, Wu X, Lee J, Tan M, Hagenbeek T, Qu X, et al. Gran-ulocyte-colony stimulating factor promotes lung metastasis throughmobilization of Ly6GþLy6Cþ granulocytes. Proc Natl Acad Sci U S A2010;107:21248–55.
26. Mantovani A. The yin-yang of tumor-associated neutrophils. CancerCell 2009;16:173–4.
27. Bellocq A, AntoineM, Flahault A, Philippe C, Crestani B, Bernaudin JF,et al. Neutrophil alveolitis in bronchioloalveolar carcinoma: inductionby tumor-derived interleukin-8 and relation to clinical outcome. Am JPathol 1998;152:83–92.
28. Zhao JJ, Pan K, WangW, Chen JG, Wu YH, Lv L, et al. The prognosticvalue of tumor-infiltrating neutrophils in gastric adenocarcinoma afterresection. PLoS ONE 2012;7:e33655.
29. Yamashita J, Ogawa M, Shirakusa T. Free-form neutrophil elastase isan independentmarker predicting recurrence in primary breast cancer.J Leukoc Biol 1995;57:375–8.
30. Hamilton JA, Achuthan A. Colony stimulating factors and myeloid cellbiology in health and disease. Trends Immunol 2013;34:81–9.
31. Lelekakis M, Moseley JM, Martin TJ, Hards D,Williams E, Ho P, et al. Anovel orthotopic model of breast cancer metastasis to bone. Clin ExpMetastasis 1999;17:163–70.
32. Aslakson CJ, Miller FR. Selective events in the metastatic processdefined by analysis of the sequential dissemination of subpopulationsof a mouse mammary tumor. Cancer Res 1992;52:1399–405.
33. Eckhardt BL, Parker BS, van Laar RK, Restall CM, Natoli AL, TavariaMD, et al. Genomic analysis of a spontaneous model of breast cancer
metastasis to bone reveals a role for the extracellular matrix. MolCancer Res 2005;3:1–13.
34. Rockwell S, Kallman RF. Cellular radiosensitivity and tumor radiationresponse in the EMT6 tumor cell system. Radiat Res 1973;53:281–94.
35. Lenzo JC, Turner AL, Cook AD, Vlahos R, Anderson GP, Reynolds EC,et al. Control of macrophage lineage populations by CSF-1 receptorand GM-CSF in homeostasis and inflammation. Immunol Cell Biol2012;90:429–40.
36. Priceman SJ, Sung JL, Shaposhnik Z, Burton JB, Torres-Collado AX,MoughonDL, et al. Targeting distinct tumor-infiltratingmyeloid cells byinhibiting CSF-1 receptor: combating tumor evasion of antiangiogenictherapy. Blood 2010;115:1461–71.
37. Conway JG, McDonald B, Parham J, Keith B, Rusnak DW, Shaw E,et al. Inhibition of colony-stimulating-factor-1 signaling in vivowith theorally bioavailable cFMS kinase inhibitor GW2580. Proc Natl Acad SciU S A 2005;102:16078–83.
38. Burns CJ, Harte MF, Bu X, Fantino E, Giarrusso M, Joffe M, et al.Discovery of 2-(alpha-methylbenzylamino) pyrazines as potent Type IIinhibitors of FMS. Bioorg Med Chem Lett 2009;19:1206–9.
39. Gyorffy B, Lanczky A, Eklund AC, Denkert C, Budczies J, Li Q, et al. Anonline survival analysis tool to rapidly assess the effect of 22,277geneson breast cancer prognosis using microarray data of 1,809 patients.Breast Cancer Res Treat 2010;123:725–31.
40. Madden SF, Clarke C, Aherne ST, Gaule P, O'Donovan N, Crown J,et al. BreastMark: an integrated approach to mining publicly availabletranscriptomic datasets relating to breast cancer outcome. BreastCancer Res 2013;15:R52.
41. Ruffell B, Au A, Rugo HS, Esserman LJ, Hwang ES, Coussens LM.Leukocyte composition of human breast cancer. Proc Natl Acad SciU S A 2012;109:2796–801.
42. Toomey D, Harmey J, Condron C, Kay E, Bouchier-Hayes D. Pheno-typing of immune cell infiltrates in breast and colorectal tumours.Immunol Invest 1999;28:29–41.
43. Rose S, Misharin A, Perlman H. A novel Ly6C/Ly6G-based strategy toanalyze the mouse splenic myeloid compartment. Cytometry A2012;81:343–50.
44. Qian BZ, Li J, Zhang H, Kitamura T, Zhang J, Campion LR, et al. CCL2recruits inflammatory monocytes to facilitate breast-tumour metasta-sis. Nature 2011;475:222–5.
45. DeNardo DG, Brennan DJ, Rexhepaj E, Ruffell B, Shiao SL, MaddenSF, et al. Leukocyte complexity predicts breast cancer survival andfunctionally regulates response to chemotherapy. Cancer Discov2011;1:54–67.
46. Li R, Doherty J, Antonipillai J, Chen S, Devlin M, Visser K, et al. LIMkinase inhibition reduces breast cancer growth and invasiveness butsystemic inhibition does not reduce metastasis in mice. Clin ExpMetastasis 2013;30:483–95.
47. Tushinski RJ, Oliver IT, Guilbert LJ, Tynan PW,Warner JR, Stanley ER.Survival of mononuclear phagocytes depends on a lineage-specificgrowth factor that the differentiated cells selectively destroy. Cell1982;28:71–81.
48. Bartocci A, Mastrogiannis DS, Migliorati G, Stockert RJ, Wolkoff AW,Stanley ER. Macrophages specifically regulate the concentration oftheir own growth factor in the circulation. Proc Natl Acad Sci U S A1987;84:6179–83.
49. Lin H, Lee E, Hestir K, Leo C, Huang M, Bosch E, et al. Discovery of acytokine and its receptor by functional screening of the extracellularproteome. Science 2008;320:807–11.
50. Lord BI, Molineux G, Pojda Z, Souza LM, Mermod JJ, Dexter TM.Myeloid cell kinetics in mice treated with recombinant interleukin-3,granulocyte colony-stimulating factor (CSF), or granulocyte-macro-phage CSF in vivo. Blood 1991;77:2154–9.
51. Kubota Y, Takubo K, Shimizu T, Ohno H, Kishi K, Shibuya M, et al. M-CSF inhibition selectively targets pathological angiogenesis and lym-phangiogenesis. J Exp Med 2009;206:1089–102.
52. Fend L, Accart N, Kintz J, Cochin S, Reymann C, Le Pogam F, et al.Therapeutic effects of anti-CD115 monoclonal antibody in mousecancer models through dual inhibition of tumor-associated macro-phages and osteoclasts. PLoS ONE 2013;8:e73310.
Anti–G-CSFR Blocks Metastasis Induced by Anti–CSF-1R
www.aacrjournals.org Cancer Immunol Res; 2(8) August 2014 775
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190

53. Granot Z, Henke E, Comen EA, King TA, Norton L, Benezra R. Tumorentrained neutrophils inhibit seeding in the premetastatic lung. CancerCell 2011;20:300–14.
54. Guleria I, Pollard JW. Aberrant macrophage and neutrophil populationdynamics and impaired Th1 response to Listeria monocytogenes incolony-stimulating factor 1–deficient mice. Infect Immun 2001;69:1795–807.
55. Pahler JC, Tazzyman S, Erez N, Chen YY,Murdoch C, NozawaH, et al.Plasticity in tumor-promoting inflammation: impairment of macro-phage recruitment evokes a compensatory neutrophil response. Neo-plasia 2008;10:329–40.
56. Greter M, Lelios I, Pelczar P, Hoeffel G, Price J, Leboeuf M, et al.Stroma-derived interleukin-34 controls the development and mainte-nance of Langerhans cells and themaintenance ofmicroglia. Immunity2012;37:1050–60.
57. Goodale D, Phay C, Brown W, Gray-Statchuk L, Furlong P, Lock M,et al. Flow cytometric assessment ofmonocyte activationmarkers and
circulating endothelial cells in patients with localized or metastaticbreast cancer. Cytometry B Clin Cytom 2009;76:107–17.
58. Mohamed MM, El-Ghonaimy EA, Nouh MA, Schneider RJ, SloaneBF, El-Shinawi M. Cytokines secreted by macrophages isolatedfrom tumor microenvironment of inflammatory breast cancerpatients possess chemotactic properties. Int J Biochem Cell Biol2014;46:138–47.
59. Pyonteck SM, Akkari L, Schuhmacher AJ, Bowman RL, Sevenich L,Quail DF, et al. CSF-1R inhibition alters macrophage polarization andblocks glioma progression. Nat Med 2013;19:1264–72.
60. Silva MT. Macrophage phagocytosis of neutrophils at inflammatory/infectious foci: a cooperativemechanism in the control of infection andinfectious inflammation. J Leukoc Biol 2011;89:675–83.
61. Aharinejad S, Salama M, Paulus P, Zins K, Berger A, Singer CF.Elevated CSF1 serum concentration predicts poor overall survival inwomen with early breast cancer. Endocr Relat Cancer 2013;20:777–83.
Cancer Immunol Res; 2(8) August 2014 Cancer Immunology Research776
Swierczak et al.
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190

2014;2:765-776. Published OnlineFirst April 29, 2014.Cancer Immunol Res Agnieszka Swierczak, Andrew D. Cook, Jason C. Lenzo, et al. Receptorof CSF-1R/CSF-1 Signaling Is Blocked by Targeting the G-CSF The Promotion of Breast Cancer Metastasis Caused by Inhibition
Updated version
10.1158/2326-6066.CIR-13-0190doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerimmunolres.aacrjournals.org/content/suppl/2014/05/02/2326-6066.CIR-13-0190.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerimmunolres.aacrjournals.org/content/2/8/765.full#ref-list-1
This article cites 58 articles, 25 of which you can access for free at:
Citing articles
http://cancerimmunolres.aacrjournals.org/content/2/8/765.full#related-urls
This article has been cited by 8 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerimmunolres.aacrjournals.org/content/2/8/765To request permission to re-use all or part of this article, use this link
on January 22, 2020. © 2014 American Association for Cancer Research. cancerimmunolres.aacrjournals.org Downloaded from
Published OnlineFirst April 29, 2014; DOI: 10.1158/2326-6066.CIR-13-0190