Embed Size (px)
Citation preview

The Postnatal Development of Neocortical - Neurons in the Dog
MICHAEL W. FOX, OTTILIE R. INMAN AND WILLIAMINA A. HIMWICH Thudichum Psychiatric Research Laboratory, Galesbwg State Research Hospital, Galesburg, Illinois
ABSTRACT Examinations were made by means of the Golgi-Cox method of the developing neuronal elements of the cortex of the dog during the first ten weeks of life. The neuronal elements undergo gradual morphological differentiation during the first week. From 2 4 weeks of age the most dramatic changes in the development of dendritic arborizations occur. After this time the changes are slower with terminal maturation of cellular elements, cessation of neuronal growth and final organization of apical and basilar dendrites and their collaterals. Comparison with the development of the kitten shows close similarity in the development of canine and feline neocortex. In dogs, however, an essentially mature neocortex is not reached until after four weeks of age. The relation of these data to biochemical and electrophysiological development and behavioral patterns in the young dog is discussed.
The development of dendritic structure in young animals has been thoroughly in- vestigated in the mouse (Kobayashi, '63), the cat (Noback and Purpura, '61) and the rabbit (Schad6, '60). In all three of these species the histological changes have been linked to electrophysiological matura- tion in both the spontaneous and evoked EEG. To the best of our knowledge, al- though data have been published on elec- trophysiological and biochemical develop- ment in the dog brain, no histological material appears to be available. In view of the increasing use of this species in developmental studies, we have investi- gated the development of dendritic struc- ture of the developing cortex in the dog.
METHODS
Tissue samples from the gyrus postcru- ciatus were taken from one mongrel pup at each of the following ages: newborn (seven hours), two, four, four and one- haIf, six, seven, eight, ten, eleven, tweIve and thirteen days, and two, four, five and one-half, nine and ten weeks after birth respectively. Although the animals were of non-isogenic origin they were reared under similar standards of mangement and nu- trition. The Golgi-Cox method used in this work was essentially that of Sholl ('53).
For purposes of the description of the de- velopment of the elements of the canine neocortex our material will be divided in- to age groups as follows: birth to seven days, 14-28 days and three and one-half to ten weeks.
The criteria for determining the develop- ment of the neurons included :
1. Comparison of neurons (pyramidal, stellate and horizontal Retzius-Cajal) at different ages in the various layers of the neocortex from the area studied. The layers were divided as follows : the molecu- lar or plexiform layer (layer I); the ex- ternal granular layer of small pyramidal neurons (layer 11) ; the external pyramidal (layer 111); the internal granular (layer IV) ; the deep pyramidal or internal pyram- idal (layer V); and the inner-most multi- form layer of irregularly arranged cells (layer VI). Layers I, 11-IV in toto and layer V were selected for comparative stud- ies. Layer VI, the deepest layer, was not included in the present investigation be- cause the irregularity of cell arrangement and great variability in cell type did not permit accurate age comparisons.
2. The development of dendrites; that is, number and density; presence of dendritic spines (or pedunculate bulbs) and arbori- zation.
J. COMP. NEUR., 127: 199-206. 199

200 MICHAEL W. FOX, OTTILIE R. INMAN AND WILLIAMINA A. HIMWICH
RESULTS
Cortex at birth to seven days During this initial postnatal period little
differentiation of the cortical neurons occurs.
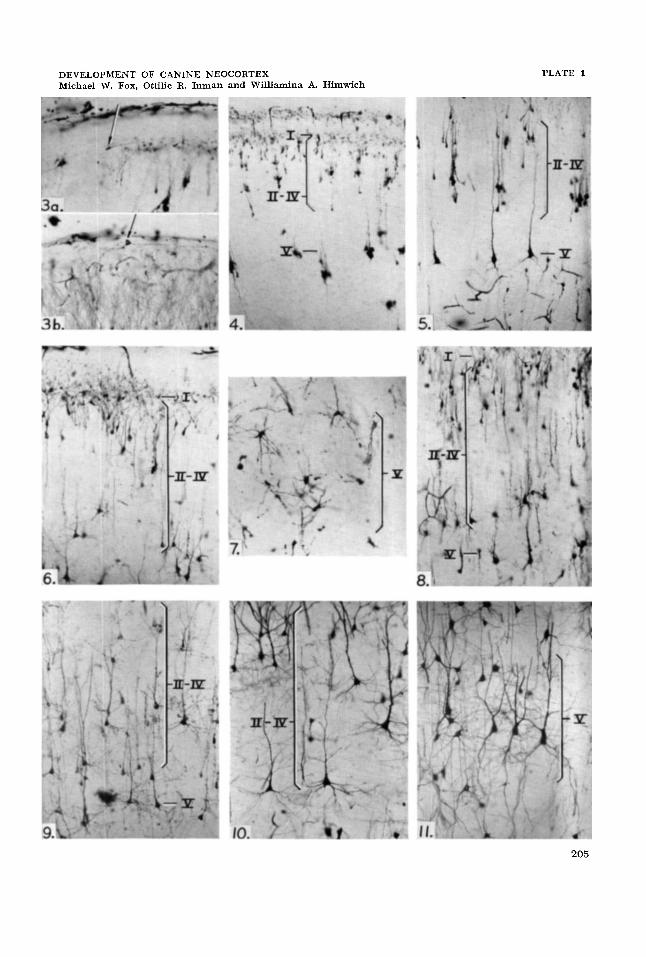
Layer I. The cell bodies of the molecu- lar layer, namely the Retzius-Cajal cells or horizontal cells, are located deeply in the lower third of the layer. In the specimens examined the dendritic processes could be observed to run for over 300 parallel to the surface of the cortex. In many sections, however, these cells were difficult to locate and their staining affinities appeared more variable than those of the neurons of the adjacent cell layers. However, the hori- zontal neurons (Retzius-Cajal), were easily recognized by their characteristic double dendritic processes which are tangentially oriented and run in opposite directions superficially in the molecular layer and give off branches at right angles (fig. 3a) . In many instances the cell bodies were radially oriented giving the appearance of stellate cells with triangular cell bodies (Cajal, '59). Occasionally marginal or pyri- form cells, possibly also representing modi- fied Retzius-Cajal cells, were observed lo- cated just beneath the pial surface of the cortex (fig. 3b).
Layers 11-IV. At birth these layers are most distinct, being comprised of predomi- nantly closely packed, radially oriented, small pyramidal neurons many of which have double apical dendrites. These den- drites pass upward to arborize in the molec- ular layer. However, at birth, many of these small neurons are without basal den- drites (fig. 4). By one week after birth short and unbranched basal dendrites have appeared on most of the neurons as well as on those cells with double apical dendrites. More rarely the cells have in addition a long basal dendrite (fig. 5). Collateral branches from the apical dendrites are present only on cells with basal dendrites and as in the cat (Noback and Purpura, '61) some basilar dendrites appear before apical collaterals form. The bulbous growths which are precursors of spines are visible only on the distal portions of the apical dendrites by one week of age. Stel- late cells occur infrequently in these layers, their dendritic processes extending toward
the molecuIar layer and the deep pyramidal layer.
Layer V. The majority of the apical dendrites of the deep pyramidal cells ar- borize in a few terminal processes near the pial surface and in short apical dendritic collaterals in layers 11-IV which they reach at one week after birth (fig. 5). The lateral spread of these collaterals is minimal, so that the cells give the gross appearance of rod-like radially oriented neurons. The basilar dendrites are, however, poorly dif- ferentiated at birth in these cells many of which resemble bipolar neuroblasts with their apical dendrite directed toward the pin1 surface. By one week after birth the basilar dendrites, although more numer- ous, are still thin and unbranched. No dendritic spines are observed at this time. Stellate cells and double bouquet cells (Noback and Purpura, '61 ) are seen occa- sionally, the former being radially oriented with relatively unbranched apical and basal dendrites. A characteristic feature of these rapidly differentiating deep pyramidal neu- rons is the dense aggregates of cells which they form.
Cortex f r o m 8-13 days Greater differentiation of the neuronal
elements of the cortex takes place during this period with more advanced elaboration of dendritic processes and an increase in neuronal size.
Layer 1. The horizontally arranged Ret- zius-Cajal cells may occasionally still be observed in this layer but the bipolar cells with horizontally distributed processes without spines, and the stellate cells are found more frequently observed. The for- mer may be derived from the Retzius-Cajal cells (Cajal, '59).
Layer 11-IV. The apical dendrites of the pyramidal cells are well developed dur- ing this period. Many collateral processes develop some of which are branched and form spines by the fourteenth day. All dendritic processes are much thicker at this stage than in the earlier ones and many cells have well developed basilar den- drites. The axons of the most advanced pyramidal neurons may have collateral branches. By this time the stellate cells have also developed more complex den- dritic processes which, however, are much

20 1 DEVELOPMENT O F CANINE NEOCORTEX
finer than those of the pyramidal cells
Layer V. The stellate cells resemble those of layers 11-IV (fig. 7) while the double dendritic bouquet cells possess char- acteristically long ascending and descend- ing dendrites, which branch infrequently, and many truncated basilar dendrites. The basilar dendrites of the deep pyramidal cells develop more slowly during this pe- riod as compared to the pyramidal cells of layers 11-IV. Considerable maturation of the apical dendrites has taken place since birth. These dendrites have become thicker with more collateral processes on the seg- ments nearest to the cell body and have developed spines on the branches of the apical dendrites (fig. 6 ) .
Cortex from 14-28 days During this period the cortex undergoes
the greatest differentiation with a marked reduction in cell density, and an increased thickness. The neurons become larger with increased thickness and arborization of neuronal processes (figs. 8, 9).
The Retzius-Cajal cells are in- frequently seen at this age but bipolar cells with horizontal axons and occasional mar- ginal or pyriform cells are well developed. All of these cells have thicker dendritic processes but there has been no marked increase in the number of arboTizations.
Layers 11-IV. In contrast to layer I, the neuronal elements of this region show greater differentiation with greater “irregu- larity” between neighboring pyramidal cells than was seen at earlier ages; i.e., rate of development and maturation of some neu- rons greatly exceeds that of others. In the rapidly maturing cells the apical dendrites and the larger collaterals have well devel- oped spines. The collateral processes of the apical dendrites in these cells are in- creased also in number and length. The basilar dendrites show no further increase in number, but have grown longer with more arborization.
Layer V. Similar changes have occurred in the deep pyramidal cells. The basal dendritic processes are thicker as well as longer with more branches (fig. 9). The double bouquet and stellate cells have de- veloped similarly and spinous processes are now evident on the thickest dendrites.
(fig. 6 ) .
Layer I.
Cortex from five and one-half to ten weeks
From the onset of this period there appear to be only minor developmental changes in the neuronal components of the area of dog brain studied as compared with the more dramatic changes which occurred during the second to fourth weeks of life. At five and one-half weeks the neurons appear to have their full complement of spinous dendritic processes (fig. 10 1. Throughout this period there is a slight thickening and lengthening of dendrites as well as an increase in the number of den- dritic spines. One feature of the more ma- ture cortex is the abundance of fine ter- minal dendritic branches.
Very few cells are located in this region and these were seen only in the dogs four weeks old and five and one-half weeks old. The cells were sparsely and irregularly distributed, resembling mar- ginal or inverted Retzius-Cajal cells with their basilar surface oriented toward the pial surface, and with two apical dendrites.
Layers 11-IV. There is greater definition between the smaller pyramidal cells and deeper, larger pyramidal cells of the sub- molecular layer than appeared previously. The larger cells have more elaborate basal dendrites than the smaller ones (fig. lo). The dendrites and collaterals of the pyram- idal neurons and stellate cells, both apical and basilar, grow thicker and longer throughout this period with more numer- ous dendritic spines. There is little further increase in the number of branches of the apical dendrites after five and one-half weeks but their collaterals are thicker and more numerous than those of layer V.
Layer V. The large pyramidal neurons of this region have numerous long branch- ing basilar dendrites that are more elab- orate than those of the submolecular py- ramidal cells and which increase in thickness but not in number during this period. The apical dendrites on these py- ramidal cells have fewer collaterals than those on the submolecular pyramidal cells and develop only a few more collaterals during this period. The collaterals on all the cells have become thicker and hence are more easily traced than at an earlier age (fig. 11) .
Layer I.

202 MICHAEL W. FOX, OTTILIE R. INMAN AND WILLIAMINA A. HXMWICH
DISCUSSION
These observations show that the neu- ronal elements of the cortex undergo grad- ual morphological differentiation during the first week of life. The layers of the cortex, although fairly distinct at birth be- come more easily recognizable with age. The most dramatic changes in the develop- ment of dendritic components occur from
2-4 weeks of age. After this time, changes are slower involving terminal maturation of cellular elements, cessation of neuronal growth and final organization of apical and basilar dendrites and their collaterals ( f igs . 1 and 12).
During early postnatal life the pyramidal neuron first shows an apical dendrite which develops terminal branches at the same
Fig. 1 Retouched photomicrographs (series in different focus to show dendritic arbori- zatione). A, B and C - upper and lower pyramidal neurons and deep stellate cell respec- tively at six weeks. Compare B with D, a deep pyramidal neuron at ten weeks.

DEVELOPMENT OF CANINE NEOCORTEX 203
time that the basilar dendrites first appear. Collaterals of the apical dendrites nearest to the cell body then develop, followed by further branching of more peripheral col- laterals of apical and basilar dendritic branches. Dendritic spines increase in number and thickness during these later events, being most numerous on the apical dendrites. In accordance with the findings in the kitten by Noback and Purpura ('61) stellate cells were seen infrequently in younger specimens, suggesting that they may be differentiated at a later age than the pyramidal cells.
The deep pyramidal cells, especially by virtue of their size, are more developed than the superficial pyramidal neurons at birth. Of the two groups of pyramidal cells, however, the superficial ones are the most differentiated by two weeks. These latter cells have a greater development of apical dendritic branches than the deep ones and the maturation of basilar dendrites in the two types show similar differences (fig. 2).
The Retzius-Cajal cells were seen only in young animals, and no trace was found at ten weeks of age. One may speculate as to their function for they were rarely found and apparently distributed at random when present in the youngest specimens. In the pup five and one-half weeks old the cells that were thought to be the Retzius-Cajal type had lost their dendritic complexity. This loss may be attributed to regressive changes (Conel, '41). Recent observations in our laboratory have disclosed the pres- ence of well developed Retzius-Cajal cells in dogs at eight and ten weeks of age in the auditory and somato-motor cortex. Older animals are now under investigation to determine the age limit, if one exists, for the occurrence of these cells in the dog.
As reported by Noback and Purpura ('61), more pyramidal and stellate cells were stained in older than in younger ani- mals. The difficulty not only in tracing the dendritic processes but also in obtaining an even impregnation of sections from im- mature subjects indicates an age difference in the staining properties of neural tissue. This difference in the staining affinities of the cortex places some limitations on the use of Golgi-Cox procedure.
A comparison of our figures with those of Noback and Purpura ('61) shows a
I:
I: Fig. 2 Diagrammatic schema of development
of pyramidal neurons in layer V (deep pyramidal cells). 1, birth; 2, one week; 3, two weeks; 4, four weeks; and 5, six weeks. Note marked development from 14-28 days and maturity of basal dendrites thereafter. Untouched.
close similarity in the development of the canine and feline neocortex. The dog, how- ever, although showing the same sequence in the development of the neuronal ele- ments in the different cortical layers, reaches maturity at a later age than the cat. Noback and Purpura ('61) found that the kitten showed an essentially mature neocortex at three weeks but in the dog the equivalent histological picture is not attained until after four weeks of age (fig. 12).
This difference in the rate of maturation of the brain has already been suggested from several other points of view. When strictly comparable EEG studies are con- ducted on the dog and the cat, the sponta- neous EEG showed a slower development to a mature pattern in the puppy than in the kitten (Petersen, Di Perri and Himwich, '64). Biochemically the accumulation of

204 MICHAEL W. FOX, OTTILIE R. INMAN AND WILLIAMINA A. HIMWICH
free amino acids in the cortex of the two species occurs more rapidly in the cat (Berl, '64) than in the puppy (Himwich and Dravid, '65). Lastly, and perhaps of more importance, in reflex and behavior studies of the two species Fox ('64) has shown that the age of adult responsiveness and activity in these species is approximately four weeks for the dog (Fox, '64) and three weeks for the cat (Fox, unpublished observations). Thus all available lines of evidence suggest a slower rate of develop- ment in the dog than in the cat, although in both species, the process of maturation is very similar.
LITERATURE CITED Berl, S. 1964 Postnatal changes in amino acid
metabolism of kitten brain. The Developing Brain, W. A. Himwich and H. E. Himwich, eds., Progress in Brain Research, Elsevier Pub- lishing Company, Amsterdam, 9: 178-182.
Cajal, S. Ramon y 1929 Studies on vertebrate neurogenesis. Translated by L. Guth, 1959, Charles C Thomas, Springfield, Illinois.
Fox, M. W. 1964 The ontogeny of behaviour and neurologic responses in the dog. Animal Behaviour, 12: 2-3.
Himwich, W. A., and A. R. Dravid 1965 Amino acid content of various brain parts as related to neurophysiological and behavioral matura- tion. Symposium on Regional Maturation of the Nervous System in Early Life. A. Min- kowski, ed., Blackwell, London ( in press).
Kobayashi, T., 0. Inman, W. Bun0 and H. E. Himwich 1963 A multidisciplinary study of changes in mouse brain with age. In: Recent Advances in Biological Phychiatry, J. Wortis, ed., Grune and Stratton, New York, pp. 293- 308.
Noback, C. R., and D. P. Purpura 1961 Post- natal ontogenesis of neurons in cat neocortex. J. Comp. Neur., 117: 291-307.
Petersen, J., R. Di Perri and W. A. Himwich 1964 The comparative development of the EEG in rabbit, cat and dog. Electroenceph. clin. Neurophysiol., 17: 557-563.
SchadB, J. P. 1960 Origin of the spontaneous electrical activity of the cerebral cortex. Re- cent Advances in Biological Psychiatry, J. Wortis, ed., Grune and Stratton, New York, pp. 2 3 4 2 .
Sholl, D. A. 1953 Dendritic organization in the neurons of the visual and motor cortices of the cat. J. Anat., 87: 387407 .
PLATE 1
EXPLANATION OF FIGURES
Figures are untouched microphotographs of Golgi-Cox preparations of cells from the G. pericruciatus of the immature dog. Pial surface is above each figure. All figures are a t the same magnification ( x 220). Black Roman numerals indicate various layers.
3a
3b
4
5
6
7
8
9
10
11
Retzius-Cajal cell in neocortex of seven-day-old dog.
Modified Retzius-Cajal cell (pyriform) in upper molecular layer of the neocortex of six-week-old dog.
Note neurons of layers 11-IV less developed than neurons of layer V of cortex of new- born dog.
Note grcater development of apical and basal dendrites and prococious maturation of neurons in layer V (seven-day-old dog).
Layers 11-IV better differentiated than at earlier ages with marked increase in basal dendritic growth (from ten-day-old dog).
Well €ormed deep stellate cells in layer V (from ten-day-old dog).
Note increased dendritic elaboration in molecular layer and layers 11-V (from two- week-old dog).
Layers 11-V with thicker and more elaborate apical and basal dendrites than in younger specimens (from four-week-old dog).
Superficial pyramidal cells of layers from 11-IV at s i x weeks of age.
Dcep pyramidal cells at ten weeks of age.
DEVELOPMENT OF CANINE NEOCORTEX Michael W. Fox, Ottilie R. Inman and Williamina A. Himwich
PLATE 1
205

DEVELOPMENT OF CANINE NEOCORTEX Michael W. Fox, Ottilie R. Inman and Williamina A. Himwich
PLATE 2
4 weeks 6 weeks 10 weeks
EXPLANATION OF FIGURES
12 Montage of untouched microphotographs of Golgi-Cox preparations of cells from the G. pericruciatus of the immature dog comparing development of neocortical neurons; magnification, x 220. Pial surface is above in each case. Note little change after six weeks of age and relative maturity at four weeks.
206





























