Embed Size (px)
Citation preview

The Plant Cell, October 2015 © 2015The American Society of Plant Biologists
10/26/2015
www.plantcell.org/cgi/doi/10.1105/tpc111.tt0411 1
© 2015 American Society of Plant Biologists
STRIGOLACTONES
www.plantcell.org/cgi/doi/10.1105/tpc.109.tt0411
© 2015 American Society of Plant Biologists
Strigolactones contribute to a devastating form of plant parasitism
Striga are parasitic plants that are the single largest biotic cause of reduced crop yields throughout Africa(> $10 billion per year in yield losses)
StrigaHost
Root parasites
Striga germination is induced by
strigolactones produced by host plant roots
Striga hermonthica
Image source: USDA APHIS PPQ Archive
© 2015 American Society of Plant Biologists
Strigolactones (SLs) regulate seemingly unrelated events
Striga
Host
Root
AM fungi
SLs inhibit shoot branching
SLs promote associations with arbuscular mycorrhizal
(AM) fungi
SLs promote germination of parasitic Striga plants
© 2015 American Society of Plant Biologists
Strigolactones inhibit shoot branching
Image courtesy RIKEN
Wild type SL-deficient
In mutant plants unable to make SLs, many more shoot branches grow out
© 2015 American Society of Plant Biologists
Strigolactones promote beneficial symbiotic interactions
Image courtesy of Mark Brundrett
Symbiotic AM fungi
SLs promotes the symbiotic association with AM fungi. This symbiosis occurs in 80% of land plants and helps them assimilate nutrients from the soil
Arbuscular is derived from Latin for tree (arbor). Mycorrhiza means “fungus root”
© 2015 American Society of Plant Biologists
Strigolatones promote germination of parasitic Striga seeds
Striga-infested maize field
Their common name is witchweed because the plants appear to be cursed. Typically Striga infestation causes reductions in crop yields of 50 – 100%
Image source USDA APHIS PPQ Archive
© 2015 American Society of Plant Biologists
Image courtesy RIKEN
What is the
connection between:
•Shoot branching
•Root parasitism, and
•Root symbiosis?
© 2015 American Society of Plant Biologists
Three seemingly independent topics converged on SLs
Evolution of plant parasitism
Origins of plant / mycorrhizal symbiosis
> 460 mya
Search for branching factor...
1960s – 1970s Purification and
characterization of strigol from roots
1900s - Role of auxin in shoot branching described
1990s – 2000s Branching mutants
described in petunia, pea, Arabidopsis and rice
2008: Strigolactonesinhibit shoot branching
1800s - Recognition of AM fungus / plant
symbiosis
1800s - Host plant factor required for parasitic seed
germination
2005: Strigolactones promote hyphal branching
© 2015 American Society of Plant Biologists
Strigolactones have been co-opted for various functions
O O
O O OOH
SL’s ancestral function may have
been communication between individuals of the same species (as
pheromones or quormones)
AM fungi
Root parasite
O
O O OH
O
SLs now contribute to communication within an individual (as hormones), and between
individuals of different species (and kingdoms) (as allelochemicals)
See for example Tsuchiya, Y. and McCourt, P. (2012). Strigolactones as small molecule communicators. Mol. BioSyst. 8: 464-469; Delaux, P.-M., et al., (2012). Origin of strigolactones in the green lineage. New Phytol. 195: 857-871; Proust, H., et al., (2011). Strigolactones regulate protonema branching and act as a quorum sensing-like signal in the moss Physcomitrella patens. Development. 138: 1531-1539.

The Plant Cell, October 2015 © 2015The American Society of Plant Biologists
10/26/2015
www.plantcell.org/cgi/doi/10.1105/tpc111.tt0411 2
© 2015 American Society of Plant Biologists
Lecture Outline
• Synthesis
• Perception and signaling
• Strigolactones in whole-plant processes:– Shoot branching
– Other developmental effects
– Moss colony growth
– Symbiosis
– Germination
• Towards the elimination of Striga parasitism
Image source USDA APHIS PPQ Archive
© 2015 American Society of Plant Biologists
Synthesis and transport
In a search for stimulators of Strigagermination, strigolactones were purified from cotton roots in 1966 and the chemical structure determined in 1972
Cook, C.E., Whichard, L.P., Turner, B., Wall, M.E., and Egley, G.H. (1966). Germination of witchweed (Striga lutea Lour.): Isolation and properties of a potent stimulant. Science 154: 1189-1190; Reprinted with permission from Cook, C.E., Whichard, L.P., Wall, M., Egley, G.H., Coggon, P., Luhan, P.A., and McPhail, A.T. (1972). Germination stimulants. II. Structure of strigol, a potent seed germination stimulant for witchweed (Striga lutea). J. Am. Chem. Soc. 94: 6198-6199.
© 2015 American Society of Plant Biologists
Strigolactones (SLs) are a small family of compounds
Reprinted from Humphrey, A.J., and Beale, M.H. (2006). Strigol: Biogenesis and physiological activity. Phytochemistry 67: 636-640 with permission from Elsevier; See also Boyer, F.D., et al.. and Rameau, C. (2012). Structure-activity relationship studies of strigolactone-related molecules for branching inhibition in garden pea: molecule design for shoot branching. Plant Physiol. 159: 1524-1544.
5-Deoxystrigol
SYNTHESIS
There are many naturally occurring SLs, derived from 5-deoxystrigol
© 2015 American Society of Plant Biologists
The stimulator of Striga germination derives from the carotenoid pathway
fluridone
MEP pathway
GGPP
phytoene
-carotene
5-deoxystrigol
Carotenoid-deficient mutants do not make germination stimulator
WT WT
mutant mutant
Through the use of maize mutants and enzyme inhibitors, carotenoids were demonstrated to be the precursors of SLs
Matusova, R., Rani, K., Verstappen, F.W.A., Franssen, M.C.R., Beale, M.H., and Bouwmeester, H.J. (2005). The strigolactone germination stimulants of the plant-parasitic Striga and Orobanche spp. are derived from the carotenoid pathway. Plant Physiol. 139: 920-934.
© 2015 American Society of Plant Biologists
Genes involved in SL biosynthesis were identified by genetic methods
Reprinted from Booker, J., et al. (2004). MAX3/CCD7 is a carotenoid cleavage dioxygenase required for the synthesis of a novel plant signaling molecule. Curr. Biol. 14: 1232-1238with permission from Elsevier; Morris, S.E., et al. (2001). Mutational analysis of branching in pea. Evidence that Rms1 and Rms5 regulate the same novel signal. Plant Physiol. 126: 1205-1213; Ishikawa, S., et al. (2005). Suppression of tiller bud activity in tillering dwarf mutants of rice. Plant Cell Physiol. 46: 79-86 by permission of the Japanese Society of Plant Physiologists. Simons, J.L., et al. (2007). Analysis of the DECREASED APICAL DOMINANCE genes of petunia in the control of axillary branching. Plant Physiol. 143: 697-706.
WT max3 WT rms5
Strigolactone-deficient mutants in Arabidopsis,
pea, rice and petunia show similar short, branchy
phenotypes
The MORE AXILLARY GROWTH3 (MAX3), RAMOSUS5 (RMS5) , DWARF17 (D17) and DECREASED APICAL DOMINANCE3 (DAD3) genes all encode a carotenoid cleavage dioxygenase (CCD7)
WT dad3
© 2015 American Society of Plant Biologists
SL biosynthesis pathway
carlactone
MAX3, D17, RMS5, DAD3:
MAX4, D10 ,RMS1, DAD1:
MAX1
STRIGOLACTONES
CCD7
CCD8
(P450)MAX; ArabidopsisD; riceRMS; peaDAD; petunia
These reactions occur
in the plastid
D27 (β-carotene-9-isomerase)
Umehara, M., Hanada, A., Yoshida, S., Akiyama, K., Arite, T., Takeda-Kamiya, N., Magome, H., Kamiya, Y., Shirasu, K., Yoneyama, K., Kyozuka, J., and Yamaguchi, S. (2008). Inhibition of shoot branching by new terpenoid plant hormones. Nature 455: 195-200.; Seto Y, Kameoka H, Yamaguchi S, Kyozuka J. (2012) Recent advances in strigolactone research: chemical and biological aspects. (in press). Alder, A., Jamil, M., Marzorati, M., Bruno, M., Vermathen, M., Bigler, P., Ghisla, S., Bouwmeester, H., Beyer, P., and Al-Babili, S. (2012). The path from β-carotene to carlactone, a strigolactone-like plant hormone. Science. 335: 1348-1351.
© 2015 American Society of Plant Biologists
D27 encodes an Fe-binding enzyme necessary for SL synthesis
Lin, H., Wang, R., Qian, Q., Yan, M., Meng, X., Fu, Z., Yan, C., Jiang, B., Su, Z., Li, J. and Wang, Y. (2009). DWARF27, an iron-containing protein required for the biosynthesis of strigolactones, regulates rice tiller bud outgrowth. Plant Cell. 21: 1512-1525.
Wild-type (Shiokari) d27
Strigolactones are detected in exudates of wild-type but not d27 roots
Standard
Wild-type exudate
d27 exudate
The rice D27 is a β-carotene-9-isomerase also
found in other plants
© 2015 American Society of Plant Biologists
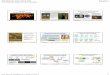
Rice SL biosynthesis mutants are rescued by exogenous SL
Umehara, M., Hanada, A., Yoshida, S., Akiyama, K., Arite, T., Takeda-Kamiya, N., Magome, H., Kamiya, Y., Shirasu, K., Yoneyama, K., Kyozuka, J., and Yamaguchi, S. (2008). Inhibition of shoot branching by new terpenoid plant hormones. Nature 455: 195-200;
Control
2nd leaf tiller1st leaf tiller
1 cm
1 cm
WT d10 d17
GR24(1 µM)
d10 and d17 are rescued by exogenous SL
(GR24 is a synthetic SL)WT d10 d17
D17
D10
© 2015 American Society of Plant Biologists
WT max1 max3 max4
Control
GR24 (5 µM)
Arabidopsis SL biosynthesis mutants are rescued by SL
Umehara, M., Hanada, A., Yoshida, S., Akiyama, K., Arite, T., Takeda-Kamiya, N., Magome, H., Kamiya, Y., Shirasu, K., Yoneyama, K., Kyozuka, J., and Yamaguchi, S. (2008). Inhibition of shoot branching by new terpenoid plant hormones. Nature 455: 195-200; Seto Y, Kameoka H, Yamaguchi S, Kyozuka J. (2012) Recent advances in strigolactone research: chemical and biological aspects. (in press).
MAX1
MAX4
MAX3

The Plant Cell, October 2015 © 2015The American Society of Plant Biologists
10/26/2015
www.plantcell.org/cgi/doi/10.1105/tpc111.tt0411 3
© 2015 American Society of Plant Biologists
MAX1 encodes a P450 enzyme involved in shoot branching
Reprinted from Booker, J., Sieberer, T., Wright, W., Williamson, L., Willett, B., Stirnberg, P., Turnbull, C., Srinivasan, M., Goddard, P., and Leyser, O. (2005). MAX1 encodes a cytochrome P450 family member that acts downstream of MAX3/4 to produce a carotenoid-derived branch-inhibiting hormone. Developmental Cell 8: 443-449 with permission from Elsevier.
max1WT
MAX1 is expressed throughout the plant, primarily in association with vascular tissuesMAX1
© 2015 American Society of Plant Biologists
SL synthesis in root or shoot is sufficient to control shoot branching
Booker, J., Sieberer, T., Wright, W., Williamson, L., Willett, B., Stirnberg, P., Turnbull, C., Srinivasan, M., Goddard, P., and Leyser, O. (2005). MAX1 encodes a cytochrome P450 family member that acts downstream of MAX3/4 to produce a carotenoid-derived branch-inhibiting hormone. Developmental Cell 8: 443-449.
max3WT
max3 ScionRoot
WTmax3WT
Grafts
Reciprocal grafts, in which wild-
type tissue is either the root or
scion, have normal phenotypes;
this says that the branch-
controlling signal can be made in
either tissue, and can move from
root to shoot
© 2015 American Society of Plant Biologists
Booker, J., Sieberer, T., Wright, W., Williamson, L., Willett, B., Stirnberg, P., Turnbull, C., Srinivasan, M., Goddard, P., and Leyser, O. (2005). MAX1 encodes a cytochrome P450 family member that acts downstream of MAX3/4 to produce a carotenoid-derived branch-inhibiting hormone. Developmental Cell 8: 443-449; Alder, A., Jamil, M., Marzorati, M., Bruno, M., Vermathen, M., Bigler, P., Ghisla, S., Bouwmeester, H., Beyer, P., and Al-Babili, S. (2012). The path from β-carotene to carlactone, a strigolactone-like plant hormone. Science. 335: 1348-1351.
An intermediate between MAX4 and MAX1 action can move in the plant
max4max1
max1max4
MAX1
STRIGOLACTONES
MAX4
In this experiment mobile, graft-transmissible intermediate in SL synthesis is produced in max1roots, and converted into SL in max4 shoots
Carlactone is a good candidate as the SL mobile
signalling molecule
© 2015 American Society of Plant Biologists
Kretzschmar, T., Kohlen, W., Sasse, J., Borghi, L., Schlegel, M., Bachelier, J.B., Reinhardt, D., Bours, R., Bouwmeester, H.J. and Martinoia, E. (2012). A petunia ABC protein controls strigolactone-dependent symbiotic signalling and branching. Nature. 483: 341-344.
WT dad1 (=max4)
PDR1 has been identified as a strigolactone exporter
pdr1
SLs are present in pdr1 root extracts, but not prd1 exudates, supporting its role as a transporter
pdr1 mutants are less colonized by
AM fungi, and stimulate less parasitic seedgermination
Loss-of-function pdr1 mutant show:• increased shoot branching• decreased exudation of SLs • decreased symbiotic interactions
© 2015 American Society of Plant Biologists
The distribution of PDR1 is consistent with its roles in transport
Reprinted from Sasse, J., Simon, S., Gübeli, C., Liu, G.-W., Cheng, X., Friml, J., Bouwmeester, H., Martinoia, E. and Borghi, L. (2015). Asymmetric localizations of the ABC transporter PaPDR1 trace paths of directional strigolactone transport. Curr. Biol. 25: 647-655 and Ruyter-Spira, C., Al-Babili, S., van der Krol, S. and Bouwmeester, H. (2013). The biology of strigolactones. Trends Plant Sci. 18: 72-83 with permission from Elsevier.
SLs are transported shootward and outward from the root
© 2015 American Society of Plant Biologists
Synthesis and transport - summary
Alder, A., Jamil, M., Marzorati, M., Bruno, M., Vermathen, M., Bigler, P., Ghisla, S., Bouwmeester, H., Beyer, P., and Al-Babili, S. (2012). The path from β-carotene to carlactone, a strigolactone-like plant hormone. Science. 335: 1348-1351.
SLs are derived from carotenoids;Early steps occur in the plastids of root and shoot
carlactone
MAX3, RMS5, D17, DAD3
MAX4, RMS1, D10, DAD1
MAX1
STRIGOLACTONES
(CCD7)
(CCD8)
(P450)MAX; ArabidopsisRMS; peaD; riceDAD; petunia
D27 (β-carotene-9-isomerase)
Carlactone is an SL intermediate (and may be a mobile signal)
© 2015 American Society of Plant Biologists
Perception and signaling
Reprinted by permission from Macmillan Publishers Ltd: Zhou, F., et al and Wan, J. (2013). D14-SCFD3-dependent degradation of D53 regulates strigolactone signalling. Nature 504: 406-410.
1. D14 is an α/β-fold hydrolase that
binds SLs
2. D3 is an F-box protein that promotes interaction with the proteasome
3. The interaction between SLs, D14 and D3 leads to degradation of target
proteins including D53 (SMAX)
© 2015 American Society of Plant Biologists
1. D14 is an α/β-fold hydrolase that binds SLs
\
Loss-of-function d14 mutants are SL insensitive and show increased shoot branching; the orthologous gene in petunia is dad2
Arite, T., Umehara, M., Ishikawa, S., Hanada, A., Maekawa, M., Yamaguchi, S. and Kyozuka, J. (2009). d14, a strigolactone-insensitive mutant of rice, shows an accelerated outgrowth of tillers. Plant Cell Physiol. 50: 1416-1424; Hamiaux, C., Drummond, R.S., Janssen, B.J., Ledger, S.E., Cooney, J.M., Newcomb, R.D. and Snowden, K.C. (2012). DAD2 is an α/β hydrolase likely to be involved in the perception of the plant branching hormone, strigolactone. Curr Biol. 22: 2032–2036
Genetic studies Biochemical studies
D14 and DAD2 are α/β-fold hydrolases similar to the GID1 protein involved in gibberellin perception
© 2015 American Society of Plant Biologists
The α/β-fold hydrolase D14/DAD2 cleaves SL
Nakamura, H., Xue, Y.L., Miyakawa, T., Hou, F., Qin, H.M., Fukui, K., Shi, X., Ito, E., Ito, S., Park, S.H., Miyauchi, Y., Asano, A., Totsuka, N., Ueda, T., Tanokura, M., and Asami, T. (2013). Molecular mechanism of strigolactone perception by DWARF14. Nat Commun. 4: 2613. Seto, Y., and Yamaguchi, S. (2014). Strigolactone biosynthesis and perception. Curr. Opin. Plant Biol. 21: 1-6 with permission from Elsevier.
Hydrolysis

The Plant Cell, October 2015 © 2015The American Society of Plant Biologists
10/26/2015
www.plantcell.org/cgi/doi/10.1105/tpc111.tt0411 4
© 2015 American Society of Plant Biologists
SL receptors D14/DAD2 are related to the KAI2 karrikin receptors
Conn, C.E., Bythell-Douglas, R., Neumann, D., Yoshida, S., Whittington, B., Westwood, J.H., Shirasu, K., Bond, C.S., Dyer, K.A., and Nelson, D.C. (2015). Convergent evolution of strigolactone perception enabled host detection in parasitic plants. Science 349: 540-543. Reprinted with permission from AAAS.
Karrikins are small molecules present in
smoke that are structurally similar to SLs and can induce seed germination
D14 probably arose from a duplication of KAI2 before the evolution of seed plants
© 2015 American Society of Plant Biologists
Parasitic Striga make highly-sensitive SL receptors
From Toh, S., Holbrook-Smith, D., Stogios, P.J., Onopriyenko, O., Lumba, S., Tsuchiya, Y., Savchenko, A. and McCourt, P. (2015). Structure-function analysis identifies highly sensitive strigolactone receptors in Striga. Science. 350: 203-207. Reprinted with permission from AAAS.
The SL receptor gene family is amplified in Striga
ShHTL7 encodes a highly sensitive receptor protein that confers germination sensitivity to SLs in vivo
© 2015 American Society of Plant Biologists
2. D3, an F-box protein, promotes interaction with the proteasome
GR24(1 µM)
Control
d3WT
MAX2, D3 and RMS4 encode F-box proteins related to those involved in auxin and jasmonate signaling
Auxin receptor
Jasmonate co-receptor
Umehara, M., Hanada, A., Yoshida, S., Akiyama, K., Arite, T., Takeda-Kamiya, N., Magome, H., Kamiya, Y., Shirasu, K., Yoneyama, K., Kyozuka, J., and Yamaguchi, S. (2008). Inhibition of shoot branching by new terpenoid plant hormones. Nature 455: 195-200. Johnson, X., Brcich, T., Dun, E.A., Goussot, M., Haurogne, K., Beveridge, C.A., and Rameau, C. (2006). Branching genes are conserved across species. Genes controlling a novel signal in pea are coregulated by other long-distance signals. Plant Physiol. 142: 1014-1026.
© 2015 American Society of Plant Biologists
In the presence of SLs, DAD2/D14interacts with D3/MAX2
Hamiaux, C., Drummond, R.S., Janssen, B.J., Ledger, S.E., Cooney, J.M., Newcomb, R.D. and Snowden, K.C. (2012). DAD2 is an α/β hydrolase likely to be involved in the perceptionof the plant branching hormone, strigolactone. Curr Biol. 22: 2032–2036 ;Nelson, D.C., Scaffidi, A., Dun, E.A., Waters, M.T., Flematti, G.R., Dixon, K.W., Beveridge, C.A.,Ghisalberti, E.L. and Smith, S.M. (2011). F-box protein MAX2 has dual roles in karrikin and strigolactone signaling in Arabidopsis thaliana. Proc Natl Acad Sci USA. 108: 8897-8902.
SL concentration-dependent interaction between DAD2 (D14)
and MAX2 (D3)
Pro
tein
inte
ract
ion
Model of SL signal transduction
Note the similarity of this model with those for auxin, jasmonate, salicylate and gibberellin signalling
A current model is that SLs promote the interaction between DAD2/D14 receptor and MAX2/D3,
leading to degradation of a signaling repressor
(D3)
© 2015 American Society of Plant Biologists
Karrikin signals are transduced in a similar manner as SLs
Nelson, D.C., Scaffidi, A., Dun, E.A., Waters, M.T., Flematti, G.R., Dixon, K.W., Beveridge, C.A., Ghisalberti, E.L. and Smith, S.M. (2011). F-box protein MAX2 has dual roles in karrikin and strigolactone signaling in Arabidopsis thaliana. Proc Natl Acad Sci USA. 108: 8897-8902.; Waters, M.T., Nelson, D.C., Scaffidi, A., Flematti, G.R., Sun, Y.K., Dixon, K.W. and Smith, S.M. (2012). Specialisation within the DWARF14 protein family confers distinct responses to karrikins and strigolactones in Arabidopsis. Development. 139: 1285-1295.
By analogy to SLs, karrikin-binding to KAI2 could promote its interaction with MAX2 and the degradation of repressor proteins
MAX2 is needed for SL
and karrikin signaling
© 2015 American Society of Plant Biologists
3. Genetic approaches identified D53/SMXLs in SL signalling
Stanga, J.P., Smith, S.M., Briggs, W.R. and Nelson, D.C. (2013). SUPPRESSOR OF MORE AXILLARY GROWTH2 1 controls seed germination and seedling development in Arabidopsis. Plant Physiol. 163: 318-330. Jiang, L., and Li, J. (2013). DWARF 53 acts as a repressor of strigolactone signalling in rice. Nature 504: 401-405. Reprinted by permission from Macmillan Publishers Ltd: Zhou, F. et al.,, and Wan, J. (2013). D14-SCFD3-dependent degradation of D53 regulates strigolactone signalling. Nature 504: 406-410.
SMAX = Suppressor of MAX2
Loss-of-function smax1mutants suppress SL mutant phenotypes
Gain-of-function dominant d53 mutants show a SL-resistant, branchy phenotype
© 2015 American Society of Plant Biologists
Zhou, F., Lin, Q., Zhu, L., Ren, Y., Zhou, K., Shabek, N., Wu, F., Mao, H., Dong, W., Gan, L., Ma, W., Gao, H., Chen, J., Yang, C., Wang, D., Tan, J., Zhang, X., Guo, X., Wang, J., Jiang, L., Liu, X., Chen, W., Chu, J., Yan, C., Ueno, K., Ito, S., Asami, T., Cheng, Z., Wang, J., Lei, C., Zhai, H., Wu, C., Wang, H., Zheng, N., and Wan, J. (2013). Reprinted by permission from Macmillan Publishers Ltd: D14-SCFD3-dependent degradation of D53 regulates strigolactone signalling. Nature 504: 406-410.
The D53 protein is a SL signaling repressor. D53 degradation by the proteasome depends on interaction with the D14/DAD2 receptor bound to SL.
D14-SL and D3/MAX2 intermediate interaction between the D53 repressor and the SCF complex.
In Arabidopsis, D14 degradation is induced by SL via a MAX2-dependent proteasome mechanism.
Model: Strigolactone promotes D14-SCFD3-mediated degradation of the
repressor D53
© 2015 American Society of Plant Biologists
Reprinted by permission from Macmillan Publishers Ltd: Jiang, L., Liu, X., Xiong, G., Liu, H., Chen, F., Wang, L., Meng, X., Liu, G., Yu, H., Yuan, Y., Yi, W., Zhao, L., Ma, H., He, Y., Wu, Z., Melcher, K., Qian, Q., Xu, H.E., Wang, Y., and Li, J. (2013). DWARF 53 acts as a repressor of strigolactone signalling in rice. Nature 504: 401-405
• D14/DAD2 is an / hydrolyze that binds SL, allowing interactions with the F-box protein D3/MAX2
• D3/MAX2 brings the SCF complex and E2 ubiquitin ligase to D53 for polyubiquitination, and further degradation by the proteasome
• D53 degradation leads to transcriptional activation of SL-target genes
Signaling summary
© 2015 American Society of Plant Biologists
Strigolactones and whole-plant processes
Seto, Y., Kameoka, H., Yamaguchi, S., and Kyozuka, J. (2012). Recent advances in strigolactone research: chemical and biological aspects. Plant Cell Physiol. 53: 1843-1853 by permission of Oxford University Press.
Strigolactones have diverse roles in the development of vascular plants and moss
This include positive and negative effects

The Plant Cell, October 2015 © 2015The American Society of Plant Biologists
10/26/2015
www.plantcell.org/cgi/doi/10.1105/tpc111.tt0411 5
© 2015 American Society of Plant Biologists
Strigolactones regulate shoot /root branching and nutrient responses
Reprinted from Brewer, P.B., Koltai, H., and Beveridge, C.A. (2013). Diverse roles of strigolactones in plant development. Mol Plant. 6: 18-28 with permission of Elsevier.
SL mutants produce more shoot and root branches
In nutrient deficiency, elevated SLs repress shoot branching and promote root hair elongation
© 2015 American Society of Plant Biologists
Strigolactones dampen polar auxin transport (PAT)
Reprinted from Bennett, T., Sieberer, T., Willett, B., Booker, J., Luschnig, C., and Leyser, O. (2006). The Arabidopsis MAX pathway controls shoot branching by regulating auxin transport. Curr. Biol. 16: 553-563 with permission from Elsevier; Crawford, S., Shinohara, N., Sieberer, T., Williamson, L., George, G., Hepworth, J., Müller, D., Domagalska, M.A., and Leyser, O. (2010). Strigolactones enhance competition between shoot branches by dampening auxin transport. Development 137: 2905-2913 reproduced with permission.
SL-deficient plants show increased polar auxin transport
Transported auxin
Auxin
+SLPIN
In shoots, strigolactones promote internalization of PIN
auxin efflux transporters, decreasing polar auxin transport
© 2015 American Society of Plant Biologists
How do strigolactones
inhibit bud outgrowth?
Goulet, C. and Klee, H.J. (2010). Climbing the branches of the strigolactones pathway one discovery at a time. Plant Physiol. 154: 493-496.
© 2015 American Society of Plant Biologists
Axillary bud outgrowth is hormonally and environmentally responsive
McSteen, P. (2009). Hormonal regulation of branching in grasses. Plant Physiol. 149: 46-55; Brewer, P.B., Dun, E.A., Ferguson, B.J., Rameau, C., and Beveridge, C.A. (2009). Strigolactone acts downstream of auxin to regulate bud outgrowth in pea and Arabidopsis. Plant Physiol. 150: 482-493.
(Axillla means armpit)
bud• Auxin and SLs
inhibit outgrowth
• Cytokinins (CKs) promote it
© 2015 American Society of Plant Biologists
SL effect on branching can occur via effects on polar auxin transport
Shinohara, N., Taylor, C., and Leyser, O. (2013) Strigolactone can promote or inhibit shoot branching by triggering rapid depletion of the auxin efflux protein PIN1 from the plasma membrane. PLoS Biol 11: e1001474.
© 2015 American Society of Plant Biologists
SL effects on shoot branching can also be auxin-independent
Brewer, P.B., Dun, E.A., Gui, R., Mason, M.G. and Beveridge, C.A. (2015). Strigolactone inhibition of branching independent of polar auxin transport. Plant Physiol. 168: 1820-1829.
Transcriptional targets of SLs include BRC1, a transcription factor that represses bud outgrowth
© 2015 American Society of Plant Biologists
–P+P
Phosphate-starved plants
suppress shoot growth and enhance root growth
Nutrient control of branching
© 2015 American Society of Plant Biologists
Phosphorous deficiency limits plant growth in much of the world
Image courtesy CIMMYT; FAO (2008)
40 million tonnes per year of phosphate
fertilizer is mined, transported, applied to
farmlands, and in many cases run-off to
contaminate lakes and rivers-P
-P-P
-P
© 2015 American Society of Plant Biologists
Strigolactones suppress shoot branching in low phosphorous
No.
of o
utgr
owin
gtil
lers
0
1
2
0
0.2
0.4
0.6
Tillering
1st tiller 2nd tiller3rd tiller
epi-5
DS
(ng
gF
W-1
)
P (µM)
600 300 120 60 30 12 06
+P –P
Strigolactone level
Strigolactone synthesis is high and shoot branching is suppressed when phosphate availability is low
Umehara, M., Hanada, A., Magome, H., Takeda-Kamiya, N., and Yamaguchi, S. (2010). Contribution of strigolactones to the inhibition of tiller bud outgrowth under phosphate deficiency in rice. Plant Cell Physiol. 51: 1118-1126.

The Plant Cell, October 2015 © 2015The American Society of Plant Biologists
10/26/2015
www.plantcell.org/cgi/doi/10.1105/tpc111.tt0411 6
© 2015 American Society of Plant Biologists
In the SL-deficient d10 mutant, branch outgrowth is independent of [P]
(None Detected)0
0.2
0.4
0.6
0
1
2
P (µM)
600 300 120 60 30 12 06
Strigolactone level
3
d10
No.
of o
utgr
owin
g til
lers
0
1
2
0
0.2
0.4
0.6
Tillering
1st tiller 2nd tiller3rd tiller
epi-5
DS
(ng
gF
W-1
)
P (µM)
600 300 120 60 30 12 06
WT
Strigolactone level
Umehara, M., Hanada, A., Magome, H., Takeda-Kamiya, N., and Yamaguchi, S. (2010). Contribution of strigolactones to the inhibition of tiller bud outgrowth under phosphate deficiency in rice. Plant Cell Physiol. 51: 1118-1126.
© 2015 American Society of Plant Biologists
Umehara, M., Hanada, A., Magome, H., Takeda-Kamiya, N., and Yamaguchi, S. (2010). Contribution of strigolactones to the inhibition of tiller bud outgrowth under phosphate deficiency in rice. Plant Cell Physiol. 51: 1118-1126.
carlactone
MAX3/D17 (CCD7)MAX4/D10 (CCD8)
MAX1 (P450)
D27 (β-carotene-9-isomerase)
carotenoid
Low [P] induces SL-related gene expression in shoots and roots
© 2015 American Society of Plant Biologists
Kohlen, W., et al. (2011) Strigolactones are transported through the xylem and play a key role in shoot architectural response to phosphate deficiency in nonarbuscular mycorrhizal host Arabidopsis. Plant Physiol. 155: 974-987.
Low [P] lowers branch outgrowth in Arabidopsis, dependent on SLs
Low P SL synthesis
Col-0 max4-1
© 2015 American Society of Plant Biologists
Furthermore, under low [P], SLs enhance root branching
Ruyter-spira, C., et al. (2011) Physiological effects of the synthetic strigolactone analog GR24 on root system architecture in Arabidopsis: Another below-ground role for strigolactones? Plant Physiology. 155: 721-734.
Low P SL synthesis
In wild-type plants grown with exogenous auxin, outgrowth of lateral roots is enhanced by SL
SLs, in combination with auxin, may
control plant architecture under
nutrient limiting conditions
© 2015 American Society of Plant Biologists
These interlocking networks provide for local and systemic responses
For example, hormone crosstalks coordinate shoot’s development in response to nitrogen and phosphate limitation
Low auxinLow SLHigh CK
High bud outgrowth
Low bud outgrowth
High auxinHigh SLLow CK
de Jong, M., George, G., Ongaro, V., Williamson, L., Willetts, B., Ljung, K., McCulloch, H., and Leyser, O. (2014). Auxin andstrigolactone signaling are required for modulation of Arabidopsis shoot branching by nitrogen supply. Plant Physiol. 166: 384-395
© 2015 American Society of Plant Biologists
Strigolactones suppress development of adventitious roots
Rasmussen, A., Mason, M.G., De Cuyper, C., Brewer, P.B., Herold, S., Agusti, J., Geelen, D., Greb, T., Goormachtig, S., Beeckman, T. and Beveridge, C.A. (2012). Strigolactones suppress adventitious rooting in Arabidopsis and pea. Plant Physiol. 158: 1976-1987.
A. SL mutants of Arabidopsis show enhanced development of adventitious roots. B. Development of adventitious roots is supressed in wild-type and mutant with SL application in a dose-dependent manner.
A. SL mutants of pea also show enhanced development of adventitious
roots. B-C. Cuttings of wild-type (B) show
development of less adventitious roots than the rms5 synthesis mutant (C).
© 2015 American Society of Plant Biologists
Strigolactones stimulate auxin-dependent secondary growth
Agusti, J., Herold, S., Schwarz, M., Sanchez, P., Ljung, K., Dun, E.A., Brewer, P.B., Beveridge, C.A., Sieberer, T., Sehr, E.M., and Greb, T. (2011). Strigolactone signaling is required for auxin-dependent stimulation of secondary growth in plants. Proc. Natl. Acad. Sci. USA. 108: 20242-20247.
SL mutants show decreased lateral and longitudinal extension of interfascicular cambium-derived tissues (ICD)
WT
max1-1
© 2015 American Society of Plant Biologists
Strigolactones regulate rice shoot gravitropism via auxin
Sang, D., Chen, D., Liu, G., Liang, Y., Huang, L., Meng, X., Chu, J., Sun, X., Dong, G., Yuan, Y., Qian, Q., Li, J. and Wang, Y. (2014). Strigolactones regulate rice tiller angle by attenuating shoot gravitropism through inhibiting auxin biosynthesis. Proc. Natl. Acad. Sci. USA. 111: 11199-11204.
The lazy lamutant is deficient in lateral auxin distribution
The sol1 mutant (suppressor of lazy), is a loss-of-function mutant of the F-box protein D3. SL-deficient mutants have enhanced shoot gravitropism, perhaps due to increased auxin synthesis
© 2015 American Society of Plant Biologists
Strigolactones stimulate leaf senescence
Reprinted from Hamiaux, C., Drummond, R.S., Janssen, B.J., Ledger, S.E., Cooney, J.M., Newcomb, R.D. and Snowden, K.C. (2012). DAD2 is an α/β hydrolase likely to be involved in the perception of the plant branching hormone, strigolactone. Curr Biol. 22: 2032–2036, with permission from Elsevier. Snowden, K.C., Simkin, A.J., Janssen, B.J., Templeton, K.R., Loucas, H.M., Simons, J.L., Karunairetnam, S., Gleave, A.P., Clark, D.G., and Klee, H.J. (2005). The Decreased apical dominance1/Petunia hybrida CAROTENOID CLEAVAGE DIOXYGENASE8 gene affects branch production and plays a role in leaf senescence, root growth, and flower development. Plant Cell 17: 746-759; Ueda, H. and Kusaba, M. (2015). Strigolactone regulates leaf senescence in concert with ethylene in Arabidopsis. Plant Physiol. doi:10.1104/pp.15.00325.
SL mutants show delayed leaf
senescence in petunia and Arabidopsis Exogenous
SLs accelerate senescence

The Plant Cell, October 2015 © 2015The American Society of Plant Biologists
10/26/2015
www.plantcell.org/cgi/doi/10.1105/tpc111.tt0411 7
© 2015 American Society of Plant Biologists
Strigolactone mutants are more sensitive to abiotic stress
Ha, C.V., Leyva-González, M.A., Osakabe, Y., Tran, U.T., Nishiyama, R., Watanabe, Y., Tanaka, M., Seki, M., Yamaguchi, S., Dong, N.V., Yamaguchi-Shinozaki, K., Shinozaki, K., Herrera-Estrella, L., and Tran, L.S. (2014). Positive regulatory role of strigolactone in plant responses to drought and salt stress. Proc Natl Acad Sci USA. 111: 851-856
These results suggest that strigolactones play a positive role in abiotic stress tolerance
control drought
© 2015 American Society of Plant Biologists
Strigolactones are present in non-vascular plants and some green algae
See Ruyter-Spira, C. and Bouwmeester, H. (2012). Strigolactones affect development in primitive plants. The missing link between plants and arbuscular mycorrhizal fungi? New Phytologist. 195: 730-733. Delaux, P.M., Xie, X., Timme, R.E., Puech-Pages, V., Dunand, C., Lecompte, E., Delwiche, C.F., Yoneyama, K., Becard, G. and Sejalon-Delmas, N. (2012). Origin of strigolactones in the green lineage. New Phytol. 195: 857-871. Tirithel; Christian Fischer
Bryophytes –mosses, liverworts
Lycopods –club mosses
Ferns Gymnosperms Angiosperms
Charales
Chlorophyceae
Plants
Green
algae
© 2015 American Society of Plant Biologists
Strigolactones regulate moss colony growth
Reproduced with permission from Proust, H., Hoffmann, B., Xie, X., Yoneyama, K., Schaefer, D.G., Yoneyama, K., Nogué, F., and Rameau, C. (2011). Strigolactones regulate protonema branching and act as a quorum sensing-like signal in the moss Physcomitrella patens. Development 138: 1531-1539.
In wild-type moss, colony size decreases with increasing colony density
In SL-deficient moss, colony size is insensitive to colony density
SLs act like a quorum-sensing molecule
© 2015 American Society of Plant Biologists
Model – SLs are diffusible signals that reveal moss density
WT – signals repress growth ccd8∆ – NO signal, no growth repression WT moss at periphery send
signal, ccd8∆ moss in center perceives it and restricts growth
Reproduced with permission from Proust, H., Hoffmann, B., Xie, X., Yoneyama, K., Schaefer, D.G., Yoneyama, K., Nogué, F., and Rameau, C. (2011). Strigolactones regulate protonema branching and act as a quorum sensing-like signal in the moss Physcomitrella patens. Development 138: 1531-1539.
© 2015 American Society of Plant Biologists
SLs in whole plant responses -summary
Reprinted from Smith, Steven M. and Waters, Mark T. (2012). Strigolactones: Destruction-dependent perception? Curr. Biol. 22: R924-R927, with permission from Elsevier.
SLs have pleiotropic effects on plant
development, mediated in part by auxin and
other hormones
© 2015 American Society of Plant Biologists
Strigolactones promote branching in arbuscular mycorrhizal fungi
Reprinted by permission from Macmillan Publishers Ltd: Akiyama, K., Matsuzaki, K.-i., and Hayashi, H. (2005). Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 435: 824-827 copyright 2005.
SL
SL
SL
SL
Time Zero Time 24 hours
SLs promote hyphal branching
© 2015 American Society of Plant Biologists
When AM fungi perceive SLs, they initiate symbiosis with the host plant
Reprinted by permission from Macmillan Publishers Ltd: Parniske, M. (2008) Arbuscular mycorrhiza: the mother of plant root endosymbiosis. Nat. Rev. Microbiol. 6: 763 – 775 copyright 2008.
© 2015 American Society of Plant Biologists
Both partners benefit from the symbiosis
The fungus gets sugars produced by photosynthesis
The plant gets nitrogen and phosphorus from the soil by way of the symbiotic fungal association
The arbuscule provides a large surface area for nutrient exchange
© 2015 American Society of Plant Biologists
0 500 1000 1500 2000 25005-deoxystrigol (pg/plant/5 days)
Low Mg
Low Ca
Low K
Low P
Low N
control
SL levels are elevated in sorghum root exudates under low P and N
Courtesy of K. Yoneyama and adapted from Yoneyama, K., Xie, X., Kusumoto, D., Sekimoto, H., Sugimoto, Y., Takeuchi, Y., and Yoneyama, K. (2007). Nitrogen deficiency as well as phosphorus deficiency in sorghum promotes the production and exudation of 5-deoxystrigol, the host recognition signal for arbuscular mycorrhizal fungi and root parasites. Planta 227: 125-132.

The Plant Cell, October 2015 © 2015The American Society of Plant Biologists
10/26/2015
www.plantcell.org/cgi/doi/10.1105/tpc111.tt0411 8
© 2015 American Society of Plant Biologists
The nutrient-effect on SL production is plant-specific
Some non-AM host plant
Leguminous plant
Non leguminous,AM host plant (e.g sorghum)
Low P
Low N
Instead, in low N, leguminous plants enter a symbiosis with
nitrogen-fixing bacteria
SL
SL
SL Or SLOr
© 2015 American Society of Plant Biologists
–P+P
AM fungi
In nutrient-poor soils: •SL synthesis increases•Shoot branching decreases and root branching increases •AM symbiosis increases
These responses enhance plant
survival under low nutrient conditions
© 2015 American Society of Plant Biologists
Strigolactones promote germination in parasitic and other plants
Umehara, M., Hanada, A., Yoshida, S., Akiyama, K., Arite, T., Takeda-Kamiya, N., Magome, H., Kamiya, Y., Shirasu, K., Yoneyama, K., Kyozuka, J., and Yamaguchi, S. (2008). Inhibition of shoot branching by new terpenoid plant hormones. Nature 455: 195-200 Dörr, I. (1997). How Striga parasitizes its host: a TEM and SEM study. Annals of Botany. 79: 463-472, by permission of Oxford University Press;
O O
O O OOH
Strigolactones were first characterized as inducers of Strigagermination (1960s)
© 2015 American Society of Plant Biologists
Strigolactones also promote Arabidopsis germination in some conditions
Tsuchiya, Y., Vidaurre, D., Toh, S., Hanada, A., Nambara, E., Kamiya, Y., Yamaguchi, S. and McCourt, P. (2010). A small-molecule screen identifies new functions for the plant hormone strigolactone. Nat .Chem. Biol. 6: 741-749.
Striga seeds show stronger seed dormancy and enhanced germination dependency on SLs
Germination response can be studied in Arabidopsis
© 2015 American Society of Plant Biologists
Striga species (witchweeds) are serious agricultural pests
•Major cereal crops are infested: corn, sorghum, millet and rice
•70 million hectares are infested
•Food productions for 300 million people are affected
•Financial loss is estimated to be approximately 10 billion USD
•No effective control measure has been developed
Adapted from Ejeta, G. and Gressel, J. (eds) (2007) Integrating new technologies for striga control: towards ending the witch-hunt. World Scientific Publishing, Singapore; Image sources: USDA APHIS PPQ Archive, Florida Division of Plant Industry Archive, Dept Agriculture and Consumer Services.
Witchweed infestation
HeavyModerateLight
Striga asiaticaStriga hermonthica
© 2015 American Society of Plant Biologists
Parasitism has evolved recently and several times
Reprinted from Westwood, J.H., Yoder, J.I., Timko, M.P., and dePamphilis, C.W. (2010) The evolution of parasitism in plants. Trends Plant Sci. 15: 227-235 with permission from Elsevier.
Some plants are facultative parasites; others, like Striga, obligate parasites. Strigaroots cannot grow normally. The primary root tip forms a haustorium specialized to penetrate host plant roots.
© 2015 American Society of Plant Biologists
Once attached to the host root, the plant grows and reproduces
Dörr, I. (1997). How Striga parasitizes its host: a TEM and SEM study. Annals of Botany. 79: 463-472, by permission of Oxford University Press; USDA APHIS PPQ Archive; USDA APHIS PPQ Archive
Seedling Flowering Tiny seeds
Mustard
Tobacco
Striga
© 2015 American Society of Plant Biologists
How can we move towards Striga-resistant crops?
Agricultural practices:•Field treatments•Allelopathic approaches
Genetic approaches: •Modify SL structure: encrypt the signal•Suppress branching in SL-deficient plants
Image courtesy of International Institute of Tropical Agriculture (IITA)
© 2015 American Society of Plant Biologists
Orobanche (broomrape)-infested carrot field
Photo credit Shmuel Golan courtesy of Yaakov Goldwasser

The Plant Cell, October 2015 © 2015The American Society of Plant Biologists
10/26/2015
www.plantcell.org/cgi/doi/10.1105/tpc111.tt0411 9
© 2015 American Society of Plant Biologists
Striga asiatica
Image courtesy of Prof. Julie Scholes & Mamadou Cissoko
Rice field infested with Striga
© 2015 American Society of Plant Biologists
Parasitic plants infect 60% of farmlands in sub-Saharan Africa
Image source USDA APHIS PPQ Archive
© 2015 American Society of Plant Biologists
Agricultural practices can reduce crop losses from Striga
Photo courtesy Ken Hammond (USDA)
• Before planting, apply germination stimulants to promote “suicidal germination” (no host = no survival)
• Apply fertilizers to reduce SL production by crop plants.
• These methods are prohibitively expensive in many parts of the developing world.
© 2015 American Society of Plant Biologists
Intercropping with beneficial plants can reduce Striga infestation
Maize intercropped with Desmodium uncinatum
Desmodium is a nitrogen-fixing legume that enriches the soil, and also produces allelopathic chemicals that interfere with Striga parasitism
Hassanali, A., Herren, H., Khan, Z.R., Pickett, J.A. and Woodcock, C.M. (2008). Integrated pest management: the push–pull approach for controlling insect pests and weeds of cereals, and its potential for other agricultural systems including animal husbandry. Phil. Trans. R. Soc. B 363: 611-621 copyright 2008 The Royal Society.
Desmodium uncinatum
© 2015 American Society of Plant Biologists
Do all SLs promote Strigagermination and affect branching?
5-Deoxystrigol Orobanchol 2’-epi-orobanchol
Sorgomol SolanacolOrobachyl acetate
Strigyl acetateStrigolSorgolactone
Fabacyl acetate7-oxoorobanchyl acetate
Orobanchyl acetate
A BC
D
There are many different naturally occurring SLs –how does their structure affect their function?
© 2015 American Society of Plant Biologists
Saturated (3,6'-dihydro-) GR24 isomers do not induce Striga germination
O
O O O
O O
O O O
O O
O O O
O O
O O O
O
Isomer I Isomer II Isomer III Isomer IV
GR24
Enol etherO O
O O O
Isomers of saturated GR24
© 2015 American Society of Plant Biologists
…but an isomer of saturated GR24 promotes interactions with AM fungi
Adapted from Akiyama, K., Ogasawara, H., Ito, S. and Hayashi, H. (2010) Structural requirements of strigolactones for hyphal branching in AM fungi. Plant Cell Physiol., 51: 1104-1117 see also Boyer, F.D., de Saint Germain, A., Pillot, J.P., Pouvreau, J.B., Chen, V.X., Ramos, S., Stevenin, A., Simier, P., Delavault, P., Beau, J.M. and Rameau, C. (2012). Structure-activity relationship studies of strigolactone-related molecules for branching inhibition in garden pea: molecule design for shoot branching. Plant Physiol. 159: 1524-1544.
Can we make synthetic SLs with beneficial but not detrimental effects? YESCan we engineer plants to make these? MAYBE
AM fungi Striga
+++ –O
O O O
O
Saturated GR24
O O
O O O+++ +++GR24
© 2015 American Society of Plant Biologists
Can we engineer Striga resistance? Rice d10 SL-deficient mutant...
(1) Parasitism
(2) Symbiosis
Root parasite
(3) Shoot branching
O
O O OH
O
O O
O OO
(1) No germination of Strigaseeds around the root
(2) Reduced AM fungi symbiosis(but not fully inhibited)
(3) Too many shoot branches
Are these effects separable?
AM fungi
© 2015 American Society of Plant Biologists
Can we engineer Striga resistance? Rice d10 SL-deficient mutant...
(1) Parasitism
(2) Symbiosis
Root parasite
(3) Shoot branching
O
O O OH
O
O O
O OO
Modify downstream component
(e.g. d10 suppressor mutants)
Can we normalize shoot branching in SL-deficient mutants?
These experiments are in progress with a goal to producing Striga-resistant plants
(1) No germination of Strigaseeds around the root
(2) Reduced AM fungi symbiosis(but not fully inhibited)
(3) Too many shoot branches
AM fungi

The Plant Cell, October 2015 © 2015The American Society of Plant Biologists
10/26/2015
www.plantcell.org/cgi/doi/10.1105/tpc111.tt0411 10
© 2015 American Society of Plant Biologists
Summary (1) Perception and signaling
de Saint Germain, A., Bonhomme, S., Boyer, F.D., and Rameau, C. (2013). Novel insights into strigolactone distribution and signalling. Curr Opin Plant Biol. 16: 583-589.
© 2015 American Society of Plant Biologists
Conclusions and future directions
AM fungi
O
O O OH
O
O O
O OO
Strigolactones are synthesized in roots of nutrient-limited plants
SLs enhance AM fungi symbiosis
SLs repress shoot branching and adventitious root formation
SLs stimulate primary root growth, root hair elongation, secondary growth, and leaf senescence
© 2015 American Society of Plant Biologists
AM fungi
O
O O OH
O
O O
O OO
Conclusions and future directions
How do SLs integrate with other hormones and signals to control shoot, root branching and leaf senescence?
Can we modify the SL biosynthesis pathway to alter the types and activities of SLs produced?
What are the evolutionary origins and ancestral functions of SLs?
© 2015 American Society of Plant Biologists
Conclusions and future directions
Photo courtesy CIMMYT
“Striga has a disproportionately large impact on those least equipped to control it, as it thrives in low-fertility soil”- International Maize and Wheat Improvement Center (CIMMYT)
Knowledge of the
production and
effects of SLs gives
us the power to
work towards
eliminating the
devastation of
parasitic Striga