Embed Size (px)
Citation preview

Plasmid xxx (2013) xxx–xxx
Contents lists available at SciVerse ScienceDirect
Plasmid
journal homepage: www.elsevier .com/ locate/yplas
Short Communication
The pJan25 vector series: An enhancement of theGateway-compatible vector pGWB533 for broaderpromoter testing applications
0147-619X/$ - see front matter Published by Elsevier Inc.http://dx.doi.org/10.1016/j.plasmid.2013.01.005
⇑ Corresponding author. Address: USDA-ARS PSI, Soybean Genomicsand Improvement Laboratory, 10300 Beltsville Ave., Bldg., 006, Beltsville,MD 20705, United States. Tel.: +1 (301) 504 5730; fax: +1 (301) 504 5728.
E-mail addresses: [email protected], [email protected](B.F. Matthews).
Please cite this article in press as: Alzohairy, A.M., et al. The pJan25 vector series: An enhancement of the Gateway-compatiblepGWB533 for broader promoter testing applications. Plasmid (2013), http://dx.doi.org/10.1016/j.plasmid.2013.01.005
Ahmed Mansour Alzohairy a,b, Margaret H. MacDonald b, Benjamin F. Matthews b,⇑a Genetics Department, Faculty of Agriculture, Zagazig University, Egyptb USDA-ARS Soybean Genomics and Improvement Laboratory, PSI, Beltsville, MD 20705, United States
a r t i c l e i n f o a b s t r a c t
Article history:Received 31 August 2012Accepted 14 January 2013Available online xxxxCommunicated by Dr. G.J. Phillips
Keywords:PlasmidpJan25GatewayeGFPGUSPromoter
Agrobacterium-mediated transformation of plants has enhanced our ability to progressmore rapidly in plant genetic engineering. Development of binary vectors for Agrobacte-rium has played a major role in advancing plant biology. Here, we report new featuresadded to the Gateway-compatible vector pGWB533 for promoter testing with the reportergene encoding b-glucuronidase (GUS). The original vector contains the spectinomycin/streptomycin adenylyltransferase (aadA) gene for bacterial selection and the hygromycinphosphotransferase gene (hpt) for transformed plant selection. However, some bacterialstrains used to transform plants, such as Agrobacterium rhizogenes strain K599, have ele-vated tolerance to spectinomycin and streptomycin, thus making bacterial selection ofpGWB533 inefficient. Although pGWB533 confers chemical selection for transgenic plantsusing hygromycin resistance, the plasmid has no visual marker that enables visual selec-tion of transformed plants or transgenic tissue. In this regard, adding a gene to constitu-tively express green fluorescent protein (eGFP) makes it easier to visually select thetransformed tissue and trim out the non-transformed.
In this report we describe a series of vectors, pJan25S (NCBI: KC416200), pJan25T (NCBI:KC416201) and pJan25X (NCBI: KC416202), that are enhancements of pGWB533 for pro-moter testing. All three vectors contain the gene encoding eGFP as a visual marker fortransformed tissue. However, in pJan25S and pJan25T, eGFP is controlled by the rolD pro-moter for root-specific expression, while in pJan25X it is controlled by the CaMV35S pro-moter for constitutive expression in all plant tissues. Spectinomycin and streptomycinresistance remains in pJan25S for bacterial selection; however, pJan25T and pJan25X con-tain the gene encoding tetracycline resistance (tet) for bacterial selection. These changesresulted in enhanced vectors with better visual and chemical selection that should havebroad application in promoter studies.
Published by Elsevier Inc.
1. Introduction at a certain level. This can be achieved through the use of
Many applications in biotechnology require promotersthat provide specific temporal and spatial gene expression
specific promoters to express a gene of interest in the cor-rect amount at the specific place and time where it isneeded. In this regard, testing promoters to evaluate theirtemporal and spatial specificity and level of expression isan important area of study in genomics. Many expressionvectors with different resistance and reporter genes weredeveloped by Nakagawa et al. (2007) using the Gateway�
recombinase cloning system (Invitrogen, Carlsbad, CA).Most of these vectors were derived from pPZP221
vector

2 A.M. Alzohairy et al. / Plasmid xxx (2013) xxx–xxx
(Hajdukiewicz et al., 1994), which contains aadA, the geneencoding adenylyltransferase. This enzyme confers bacte-rial resistance to spectinomycin and streptomycin.Although the Gateway� cloning system has been used toconstruct numerous plant expression vectors, these vec-tors cannot be used for all applications that are requiredfor in-depth promoter studies due to certain limitations(Traore and Zhao, 2011). For instance, most of the Gate-way�-compatible vectors use only chemical selection fortransformed plants, e.g., kanamycin or hygromycin resis-tance (Nakamura et al., 2009). This requires growing thetransformed plants on tissue culture media with the rightconcentration of chemical and requires sterile techniquewith all the costs and risks of microbial contamination. Inthis regard, visual selection of transformed plant tissue ismuch less expensive and easier. For instance, when usingAgrobacterium rhizogenes-mediated root transformation,some root tissue could be transformed while other tissueis not. However, if the transforming vector contains thegene encoding enhanced green fluorescent protein (eGFP;Haseloff et al., 1997) to transform roots, the transformedroots can be readily distinguished from non-transformedroots by eGFP fluorescence and thus untransformed rootscan be easily excised and discarded.
Several research studies have used eGFP fluorescence fordistinguishing transformed from non-transformed plantmaterial. For example, gene silencing constructs wereexamined to determine if they conferred resistance to soy-bean cyst nematodes if expressed in soybean roots of com-posite plants (Klink et al., 2009; Ibrahim et al., 2011).Transformed roots were visualized using eGFP and non-transformed roots were excised before adding nematodesto the transformed roots. Mature female nematodes werecounted 35 days later. In this case, A.rhizogenes was usedfor root transformation and the vector contained the geneencoding eGFP, controlled by the root-specific rolD pro-moter from A. rhizogenes (White et al., 1985; Elmayan andTepfer, 1995). Sterile technique and chemical selectionwas not required to identify and select transformed roots.
In this study, the pGWB533 vector (Fig. 1a) was used asthe DNA backbone to develop the pJan25S, pJan25T andpJan25X vectors to be used with A. rhizogenes and A. tum-efaciens-mediated root and whole plant transformation,respectively (Tepfer, 1984; Ko et al., 2006). The Gateway�
(Invitrogen, Carlsbad, CA) compatible pJan25 destinationvectors were engineered specifically for Agrobacterium-based plant transformation and visualization of trans-formed tissue and plants.
2. Material and methods
2.1. Addition of tetracycline resistance and eGFP
The genes encoding tetracycline resistance (tet) andgreen fluorescent protein (eGFP) were obtained from thepRAP17 vector (Klink et al., 2009). Antibiotic selection ofbacteria transformed with the vector is provided by tet,while the eGFP gene provides a visual marker for identify-ing transformed plant tissue. The gene encoding eGFP pro-tein was controlled by the rolD promoter and the T35s
Please cite this article in press as: Alzohairy, A.M., et al. The pJan25 vpGWB533 for broader promoter testing applications. Plasmid (2013), h
terminator. The rolD-eGFP-T35s cassette was amplifiedusing Platinum� Taq DNA Polymerase High Fidelity (Invit-rogen, Carlsbad, CA) from pRAP17 as template (Klink et al.,2009) with cassette-specific primers including HindIII sitesfor cloning into pGWB533 (Table 1). Amplification reac-tions were performed according to Williams et al. (1990).Briefly, each reaction mixture contained l� of 10� HighFidelity PCR Buffer (600 mM Tris–SO4 (pH 8.9), 180 mMammonium sulfate), 0.2 mM dNTPs (Pharmacia), 2 mMMgSO4, 0.2 lM of each primer, 50 ng of pRAP17 DNA,and 1.0 unit of Platinum� Taq DNA Polymerase High Fidel-ity in a final volume of 50 ll. The PCR program had an ini-tial denaturation step at 94 �C for 2 min followed by 35cycles of: denature at 94 �C for 30 s, anneal at 55 �C for30 s, and extend at 68 �C for 2 min, followed by a finalextension step at 68 �C for 10 min. Ten microliters of PCRproduct stained with 2 ll E-Z Vision� One DNA LoadingBuffer (Amresco, USA) was separated by agarose (1) gelelectrophoresis run at 100 V in 1X SB (10 mM NaOH solu-tion with boric acid, pH 8.5) and photographed on a UVtransilluminator (Pharmacia) by a Canon S5 digital camerawith a UV filter adaptor. A negative control that containedall the necessary PCR components except template DNAwas included for PCRs. The 40 ll remainder of the rolD-eGFP-T35s cassette amplicon was digested with HindIIIand purified using the MinElute PCR Purification Kit (Qia-gen, Santa Clarita, CA) following the manufacturer’sinstructions. The rolD-eGFP-T35s HindIII cassette was li-gated to the HindIII site of pGWB533 following the manu-facturer’s instructions, resulting in pJan25S. The HindIIIsite lies inside the left and right T-DNA borders that deli-mit the region that is transferred into plant tissue upontransformation (Fig. 1a and b). The 50-phosphate fromthe pGWB533 linear plasmid was removed using shrimpalkaline phosphatase (SAP) (Novagen, Madison, WI) at37 �C for 1 h to prevent self-ligation of the plasmid. TheSAP was inactivated by incubation at 65 �C for 15 min.The rolD-eGFP-T35s cassette was then ligated to thepGWB533-phosphatased vector with T4 DNA ligase (Invit-rogen, Carlsbad, CA) overnight at 14 �C. One microliter ofthe ligation was then transformed into One Shot 2T1R ccdBSurvival Cells (Invitrogen), plated on LB plates supple-mented with 100 mg L�1 spectinomycin and incubatedovernight at 37 �C.
The cassette containing tet was isolated from pRAP17by digesting with BstEII (Invitogen, Carlsbad, CA) fol-lowed by gel purification using a QIAquick� GelExtraction Kit (Qiagen, Santa Clarita, CA) according tothe manufacturer’s instructions. The quality and concen-tration of the purified fragments were measured using aNanoDrop (Thermo Fisher, Wilmington, DE) spectropho-tometer. The tet cassette was ligated into the BstEII siteof the Gateway�-compatible vector pJan25S, which wasdigested by the same restriction enzyme andphosphatased before ligation. The BstEII site lies outsidethe left and right T-DNA borders and so is not trans-ferred to plant tissue (Fig. 2b). When placed at this site,tet also interrupts the addA gene, rendering thespectinomycin and streptomycin resistance nonfunc-tional. Adding the tet cassette to pJan25S resulted inthe formation of pJan25T. Replacing the rolD-eGFP-T35s
ector series: An enhancement of the Gateway-compatible vectorttp://dx.doi.org/10.1016/j.plasmid.2013.01.005
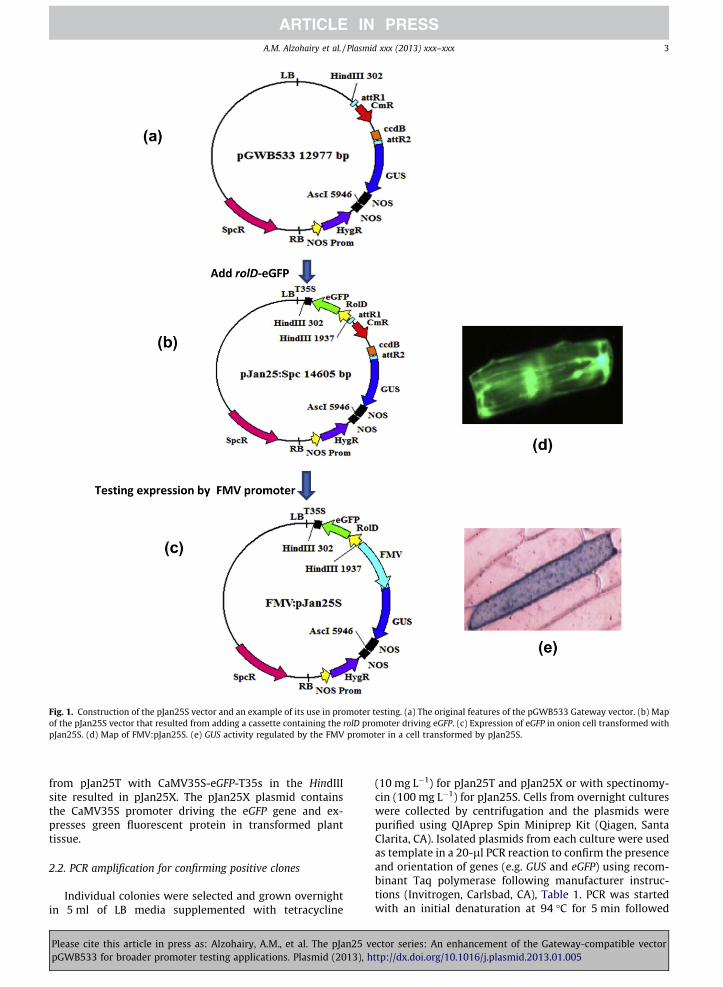
Fig. 1. Construction of the pJan25S vector and an example of its use in promoter testing. (a) The original features of the pGWB533 Gateway vector. (b) Mapof the pJan25S vector that resulted from adding a cassette containing the rolD promoter driving eGFP. (c) Expression of eGFP in onion cell transformed withpJan25S. (d) Map of FMV:pJan25S. (e) GUS activity regulated by the FMV promoter in a cell transformed by pJan25S.
A.M. Alzohairy et al. / Plasmid xxx (2013) xxx–xxx 3
from pJan25T with CaMV35S-eGFP-T35s in the HindIIIsite resulted in pJan25X. The pJan25X plasmid containsthe CaMV35S promoter driving the eGFP gene and ex-presses green fluorescent protein in transformed planttissue.
2.2. PCR amplification for confirming positive clones
Individual colonies were selected and grown overnightin 5 ml of LB media supplemented with tetracycline
Please cite this article in press as: Alzohairy, A.M., et al. The pJan25 vpGWB533 for broader promoter testing applications. Plasmid (2013), h
(10 mg L�1) for pJan25T and pJan25X or with spectinomy-cin (100 mg L�1) for pJan25S. Cells from overnight cultureswere collected by centrifugation and the plasmids werepurified using QIAprep Spin Miniprep Kit (Qiagen, SantaClarita, CA). Isolated plasmids from each culture were usedas template in a 20-ll PCR reaction to confirm the presenceand orientation of genes (e.g. GUS and eGFP) using recom-binant Taq polymerase following manufacturer instruc-tions (Invitrogen, Carlsbad, CA), Table 1. PCR was startedwith an initial denaturation at 94 �C for 5 min followed
ector series: An enhancement of the Gateway-compatible vectorttp://dx.doi.org/10.1016/j.plasmid.2013.01.005

Table 1Primer pairs used in the isolation of genes.
Primer name Sequence
rolD HindIII forward ATTTTAAGCTTGCATGCCTGCAGGTTAGT35s HindIII reverse GCCCCAAGCTTGCAGGTCACTGGATTTTGGCAMV35S HindIII
forwardCGTCAAGCTTAACATGGTGGAGCACGACAC
FMV forward plusCACC
CACCTTTACAGTAAGAACTGATAAC
FMV reverse CACTCCCCCTCTCTAAAAAAeGfp complete F TGGTGAGCAAGGGCGAGGAGCeGfp complete R TCGTCCATGCCGAGAGAGATCCCGtet-BF (BstE II) TCGTGGTCACCATCAGCGATCGGCTCGTTtet-BR (BstE II) AGCGGTGACCATCAATCGTCACCCAAAGUS forward GGA CTT TGC AAG TGG TGA ATC CGUS reverse CGA ATC CTT TGC CAC GTA AGT C
4 A.M. Alzohairy et al. / Plasmid xxx (2013) xxx–xxx
by 25 cycles of denaturation at 94 �C for 35 s, annealing atthe primer Tm (Table 1) for 35 s, and extension at 72 �C for2 min per kb of PCR product. Final extension of the ampli-fication was for 10 min at 72 �C. All PCR reactions wereperformed using a DNA Engine Tetrad 2 Peltier ThermalCycler (Bio-Rad, USA).
2.3. Transfer of the FMV promoter from the pENTR plasmid tothe pJan25 series
To test the new vectors and ensure that the selectionmarkers and promoter site worked properly together, theFMV-sgt promoter fragment, �270 to +31 from the tran-scription start site (Bhattacharyya et al., 2002), was trans-ferred from a FMV: pENTR clone to the pJan25T, pJan25Sand pJan25X plasmids using the Gateway� LR Clonase™II Enzyme Mix kit following the manufacturer’s instruc-tions (Invitrogen, Carlsbad, CA). The LR clonase reaction re-placed the ccdB gene between attR1 and attR2 sites withthe FMV promoter, thus allowing the Escherichia coli cellsto grow. The plasmid was transformed into E. coli (OneShot� Mach1™-T1R chemically competent cells, Invitro-gen, Carlsbad, CA). clones, The transformed cells weregrown overnight at 37 �C on LB selective media containing10 mg L�1 tetracycline for pJan25T and pJan25X, and100 mg L�1 spectinomycin for pJan25S. Plasmids were iso-lated from overnight cultures using the QiaPrep Spin Mini-prep Kit (Qiagen, Santa Clarita, CA). Their sequences wereconfirmed by DNA sequencing.
2.4. Microprojectile bombardment
Microprojectile bombardment (Sanford, 1990) wasconducted using the Biolistic Particle Delivery System(PDS-1000/He, BioRad, Hercules, CA) according to themanufacturer’s instructions. Gold microprojectiles coatedwith 6 ll of plasmid DNA (1 lg/ll) were bombarded intoonion cells at 1100 PSI. Each construct was delivered intothree replicates.
Fluorescence was monitored using a Nikon SMZ 1500stereomicroscope with an eGFP filter (Nikon Corporation;Tokyo, Japan). Stereomicroscope images were capturedwith an Optronics MagnaFire model S99802 CCD camera
Please cite this article in press as: Alzohairy, A.M., et al. The pJan25 vpGWB533 for broader promoter testing applications. Plasmid (2013), h
(Optronics; Goleta, CA). The functionality of the pJan25series plasmids was tested using the FMV promoter drivingexpression of the GUS reporter gene. Bombarded onion tis-sue was incubated with GUS staining solution to determineb-glucuronidase activity after 24 h. GUS staining solutionconsisted of 2 mM 5-bromo-4-chloro-3-indolyl glucuro-nide, 100 mM potassium phosphate buffer pH 7.0, 10 mMEDTA, 0.5 mM potassium ferricyanide, 0.5 mM potassiumferrocyanide, 0.1% Triton X-100 (Jefferson et al., 1987).The colorimetric reaction proceeded by immersion inGUS stain and subsequent vacuum infiltration with500 ll of GUS stain for 10 min. Tissue was subsequentlyincubated at 37 �C overnight to promote development ofthe GUS stain.
2.5. Soybean root transformation with A. rhizogenes
Plasmids in the pJan25 series harboring the FMV pro-moter were used to transform chemically competent A.rhizogenes (strain K599) cells. The transformed cells weregrown overnight at 26 �C on LB selective medium contain-ing 5 mg L�1 tetracycline for pJan25T and pJan25X, and100 mg L�1 spectinomycin for pJan25S. For soybean trans-formation, an A. rhizogenes clone containing a pJan25 plas-mid was grown overnight at 26 �C in TB mediasupplemented with 5 mg L�1 tetracycline or 100 mg L�1
spectinomycin. Cells were resuspended in MS mediumwith 3% sucrose, pH 5.7, to yield a suspension with anOD600 of approximately 0.5. Soybean seedlings 7–9 daysold were cut at the soil line and transferred to 50-ml beak-ers (25 plants/beaker) containing 40 ml of the A. rhizogenessuspension. The cell suspension was vacuum infiltratedinto the base of the stem of the cut plants for 30 min, afterwhich the plants were cocultivated in the same solution at23 �C for 16 h on a rotating platform moving at 65 rpm. Theplants were washed with reverse osmosis water and keptin water in the 50-ml beakers for 2 days before beingplanted in Promix peat-based growing medium in a green-house. Four weeks later, eGFP fluorescence of transformedroots was observed using a Dark Reader Spot lamp (ClareChemical Research, Dolores, CO). Approximately 65–70%of the seedlings had transformed roots.
3. Results and discussion
3.1. Overview of pJan25S, pJan25T and pJan25X
The pJan25 vectors were developed from the pGWB533vector (12,977 bp), which contains the GATEWAY� attR1and attR2 sites for easy cloning of promoters of interest.The attR2 site is adjacent to the gene encoding GUS whichserves as a reporter gene for promoter activity (Fig. 1a).
3.2. Construction of pJan25S
The pJan25S plasmid was designed to add root-specificeGFP expression to the pGWB533 vector (Fig. 1a), thusallowing visual selection of transformed root material viaeGFP fluorescence. Consequently, the rolD promoter, whichis highly active in roots, was used to regulate eGFP gene
ector series: An enhancement of the Gateway-compatible vectorttp://dx.doi.org/10.1016/j.plasmid.2013.01.005

Fig. 2. Construction of the pJan25T vector and an example of its use in promoter testing. Maps of pJan25S before (a) and after (b) adding the tetracyclineresistance (tet) gene, which resulted in pJan25T. (c) Onion cell transformed with pJan25T exhibiting eGFP fluorescence. (d) Map of pJan25T with FMVpromoter. (e) FMV driving GUS activity in onion cell.
A.M. Alzohairy et al. / Plasmid xxx (2013) xxx–xxx 5
expression. A rolD-eGFP-T35s cassette was added topGWB533, resulting in the new vector pJan25S (Fig. 1b).Presence of the eGFP gene in the plasmid confirmed byPCR. As in pGWB533, the pJan25S plasmid provides specti-nomycin resistance for bacterial selection. The functional-ity of pJan25S for promoter studies was tested byinserting the figwort mosaic virus subgenomic transcript(FMV) promoter (Bhattacharyya et al., 2002) into theattR1-attR2 cloning site upstream from the GUS gene(Fig. 1 c) and using this construct to transiently transformonion cells via microprojectile bombardment. Expressionof both eGFP and GUS proteins in transformed onion cellswas confirmed visually (Fig. 1d, e).
Please cite this article in press as: Alzohairy, A.M., et al. The pJan25 vpGWB533 for broader promoter testing applications. Plasmid (2013), h
3.3. Construction of pJan25T
A.rhizogenes (strain K599) has been used for plant trans-formation and in the production of composite plants withtransformed roots (Klink et al., 2009; Ibrahim et al.,2011). However, strain K599 has an elevated tolerance tospectinomycin but is very sensitive to tetracycline.Thus, addition of the tetracycline resistance gene (tet) topJan25S allows more efficient selection of transformed A.rhizogenes cells. Therefore, tetracycline resistance wasengineered into pJan25S (Fig. 2a), outside the left and rightborders, resulting in the new vector pJan25T (Fig. 2b). Pres-ence of the tet gene was confirmed by PCR. The FMV
ector series: An enhancement of the Gateway-compatible vectorttp://dx.doi.org/10.1016/j.plasmid.2013.01.005

Fig. 3. Construction of the pJan25X vector and an example of its use in promoter testing. Maps of pJan25X before (a) and after (b) adding the CaMV35Spromoter. (c) Onion cell transformed with pJan25X exhibiting fluorescence. (d) FMV driving GUS activity in onion cell.
6 A.M. Alzohairy et al. / Plasmid xxx (2013) xxx–xxx
promoter was inserted to express the GUS gene (Fig. 2c) totest the functionality of pJan25T for promoter studies. Thevector was tested in transiently transformed onion cells.Expression of both GUS and eGFP proteins intransformed onion cells was confirmed visually (Fig. 2d, e).
3.4. Construction of pJan25X
The pJan25X plasmid was designed to add eGFP expres-sion capability throughout the plant by replacing the rolD
Please cite this article in press as: Alzohairy, A.M., et al. The pJan25 vpGWB533 for broader promoter testing applications. Plasmid (2013), h
promoter driving eGFP expression with the CaMV35S pro-moter, which is active throughout the plant, though itsactivity in the root is lower than that of rolD. The rolD-eGFP-T35s fragment of pJan25T (Fig. 3a) was replaced bya DNA fragment containing CaMV35S-eGFP-T35s to pro-duce the pJan25X (Fig. 3b). The pJan25X vector was testedfor tetracycline resistance by growing it in E. coli on LBplates containing 10 mg L�1 tetracycline. Expression ofeGFP (Fig. 3c) and the functionality of pJan25X for pro-moter studies were tested as described above. The FMV
ector series: An enhancement of the Gateway-compatible vectorttp://dx.doi.org/10.1016/j.plasmid.2013.01.005

Fig. 4. Expression of eGFP and GUS genes in soybean roots transformed with pJan25 constructs. (a) Soybean roots exhibiting fluorescence (left) and stainedfor GUS activity (right) after transformation with FMV:pJan25S. (b) Soybean roots exhibiting fluorescence (left) and GUS staining (right) aftertransformation with FMV:pJan25T. (c) Soybean roots exhibiting fluorescence (left) and GUS staining (right) after transformation with FMV:pJan25X. (d)Nontransformed root from a negative control plant.
A.M. Alzohairy et al. / Plasmid xxx (2013) xxx–xxx 7
promoter was used to express the GUS gene in transientlytransformed onion cells (Fig. 3d).
3.5. Transforming plants with pJan25S, pJan25T and pJan25Xplasmids
The pJan25S, pJan25T, and pJan25X constructs wereshuttled into A. rhizogenes K599, and soybean roots weretransformed with each of these vectors individually. Trans-formed roots were apparent from 7 to 14 days after trans-formation. The transformation efficiency ranged from 65%to 70% of the total number of seedlings used in this study.The transformed soybean roots exhibited both eGFP andGUS gene expression (Fig. 4).
4. Conclusions
In summary, genes conferring green fluorescence andtetracycline resistance were added to the pGWB533 vec-tor to enhance its usefulness in promoter testing studies.The pJan25S vector contains a spectinomycin resistancegene for bacterial selection, while the pJan25T and
Please cite this article in press as: Alzohairy, A.M., et al. The pJan25 vpGWB533 for broader promoter testing applications. Plasmid (2013), h
pJan25X vectors contain a tetracycline resistance gene.All three vectors contain the gene encoding eGFP for vi-sual screening of the transformed tissue. The eGFP geneproduct is a visual beacon for screening transformedcells (Collier et al., 2005) and works particularly well inpJan25S and pJan25T to identify transformed roots, sinceits expression is regulated by a root-specific promoter inthose vectors (Klink et al., 2009). In pJan25X, eGFPexpression is regulated by a promoter active throughoutthe plant. Since the eGFP gene and the promoter-GUS fu-sion cloning site are not linked together, the overalltransformation event can be easily selected by eGFP fluo-rescence under Dark Reader lamps, and promoter-spe-cific tissue that may be minute (specific to infectionsites, for example) can be further identified by GUSexpression under microscopic conditions. The pJan25Svectors are available for research use only.
Acknowledgments
The authors would like to acknowledge Prof. Dr Tsuyo-shi Nakagawa, Department of Molecular and Functional
ector series: An enhancement of the Gateway-compatible vectorttp://dx.doi.org/10.1016/j.plasmid.2013.01.005

8 A.M. Alzohairy et al. / Plasmid xxx (2013) xxx–xxx
Genomics, Center for Integrated Research in Science, Shi-mane University, for providing the Gateway vectorpGWB533 for research purposes. We also thank Dr. I. Maiti,University of Kentucky, for providing the figwort mosaicvirus subgenomic transcript (FMV) promoter. The authorsthank Eric Brewer for critical review of the manuscript,and gratefully acknowledge support from United SoybeanBoard and from the Fulbright and Arab Fund fellowshipprograms. Mention of trade name, proprietary product, orvendor does not constitute a guarantee or warranty ofthe product by the U.S. Department of Agriculture or implyits approval to the exclusion of other products or vendorsthat also may be suitable. The names of these new plas-mids were inspired by the Egyptian revolution in January25th 2011 (AMA).
References
Bhattacharyya, S., Dey, N., Maiti, I.B., 2002. Analysis of cis-sequence ofsubgenomic transcript promoter from the figwort mosaic virus andcomparison of promoter activity with the cauliflower mosaic viruspromoters in monocot and dicot cells. Virus Res. 90, 47–62.
Collier, R., Fuchs, B., Walter, N., Kevin Lutke, W., Taylor, C.G., 2005. Ex vitrocomposite plants: an inexpensive, rapid method for root biology.Plant J. 43, 449–457.
Elmayan, T., Tepfer, M., 1995. Evaluation in tobacco of the organspecificity and strength of the rolD promoter, domain A of the 35Spromoter and the 35S2 promoter. Transgenic Res. 4, 388–396.
Haseloff, J., Siemering, K.R., Prasher, D.C., Hodge, S., 1997. Removal of acryptic intron and subcellular localization of green fluorescentprotein are required to mark transgenic arabidopsis plants brightly.Proc. Natl. Acad. Sci. USA 94, 2122–2127.
Hajdukiewicz, P., Svab, Z., Maliga, P., 1994. The small, versatile pPZPfamily of Agrobacterium binary vectors for plant transformation. PlantMol. Biol. 25 (6), 989–994.
Please cite this article in press as: Alzohairy, A.M., et al. The pJan25 vpGWB533 for broader promoter testing applications. Plasmid (2013), h
Ibrahim, H.M.M., Alkharouf, N.W., Meyer, S.L.F., Aly, M.A.M., El-Din,A.E.K.G., Hussein, E.H.A., Matthews, B.F., 2011. Post-transcriptionalgene silencing of root knot-nematode in transformed soybean roots.Exp. Parasitol. 127, 90–99.
Jefferson, R.A., Kavanagh, T.A., Bevan, M.W., 1987. GUS fusions: b-glucuronidase as a sensitive and versatile gene fusion marker inhigher plants. EMBO J. 6, 3901–3907.
Klink, V.P., Kim, K.H., Martins, V.E., MacDonald, M.H., Beard, H.S.,Alkharouf, N.W., Park, S.C., Matthews, B.F., 2009. A correlationbetween host-mediated expression of parasite genes as tandeminverted repeats and abrogation of the formation of femaleHeterodera glycines cysts during infection of Glycine max. Planta230, 53–71.
Ko, T.S., Korban, S.S., Somers, D.A., 2006. Soybean (glycine max)transformation using immature cotyledon explants. Methods Mol.Biol. (Clifton, NJ) 343, 397–405.
Nakagawa, T., Suzuki, T., Murata, S., Nakamura, S., Hino, T., Maeo, K.,Tabata, R., Kawai, T., Tanaka, K., Niwa, Y., Watanabe, Y., Nakamura, K.,Kimura, T., Ishiguro, S., 2007. Improved gateway binary vectors: high-performance vectors for creation of fusion constructs in transgenicanalysis of plants. Biosci. Biotechnol. Biochem. 71 (8), 2095–2100.
Nakamura, S., Nakao, A., Kawamukai, M., Kimura, T., Ishiguro, S.,Nakagawa, T., 2009. Development of gateway binary vectors,R4L1pGWBs, for promoter analysis in higher plants. Biosci.Biotechnol. Biochem. 73 (11), 2556–2559.
Sanford, J.C., 1990. Biolistic plant transformation. Physiol. Plant. 79, 206–209.
Tepfer, D., 1984. Transformation of several species of higher plants byAgrobacterium rhizogenes: sexual transmission of the transformedgenotype and phenotype. Cell 37, 959–967.
Traore, S.M., Zhao, B., 2011. A novel Gateway�-compatible binary vectorallows direct selection of recombinant clones in Agrobacteriumtumefaciens. Plant Methods 7 (1), 42.
Williams, J.G.K., Kubelik, A.R., Livak, K.J., Rafalski, J.A., Tingey, S.V., 1990.DNA polymorphisns amplified by arbitrary primers are useful asgenetic markers. Nucleic Acids Res. 18, 6531–6535.
White, F.F., Taylor, B.H., Huffman, G.A., Gordon, M.P., Nester, E.W., 1985.Molecular and genetic analysis of the transferred DNA regions of theroot-inducing plasmid of Agrobacterium rhizogenes. J. Bacteriol. 164,33–44.
ector series: An enhancement of the Gateway-compatible vectorttp://dx.doi.org/10.1016/j.plasmid.2013.01.005





























