Embed Size (px)
Citation preview

Medical Hypotheses 1
Uedrc ol Hvpotheses (1994) 43. I-5 0 Longman Gmup Ltd 1994
I
The Pathogenesis of Osteochondrosis - A Hypothesis
T. NAKANO and F. X. AHERNE
Department of Animal Science, University of Alberta, Edmonton, Alberta, Canada T6G 2P5 (Reprint requests to FXA).
Abstract - Osteochondrosis is a non-infectious syndrome manifested as failure of endochondral ossification that occurs in joint cartilage (atticular cartilage-subatiicular growth cartilage complex) and physis of long bones of growing animals and humans. The pathogenesis of osteochondrosis is poorly understood. It was hypothesized: 1) that the subarticular growth cartilage, where early lesions occur, is a biomechanically weaker tissue than the articular cartilage; and 2) that excessive mechanical stress added to the subarticular growth cartilage impairs normal function of matrix or chondrocytes of the tissue, and results in abnormal matrix destruction which involves proteinases.
Introduction
Osteochondrosis is a syndrome characterized as non- infectious disturbance of endochondral ossification that occurs in joint cartilage (articular cartilage- subarticular growth cartilage complex) and physis of long bones of growing animals and humans (l- 12). Osteochondrosis in domestic species (e.g. swine) is a problem of economic importance as well as an animal welfare issue. Causes of osteochondrosis are poorly understood. Advanced degree of osteo- chondrosis leads to osteochondrosis dissecans or os- teoarthrosis. The medial femoral condyle is the most commonly affected site (4,10,12,13). Because of the similarities of lesions, and because of the difficulty of detection of early lesions in humans, the pig is considered to be an appropriate experimental model for the study of osteochondrosis in humans (14,159.
It has been suggested that osteochondrosis in hu- mans is caused by abnormality of either cartilage or subchondral bone (10,12). However, studies of swine joints, in which relatively early lesions ,could be ex- amined, demonstrated that cartilage is first affected. It
Date received 4 August 1992 Date accepted 27 August 1992
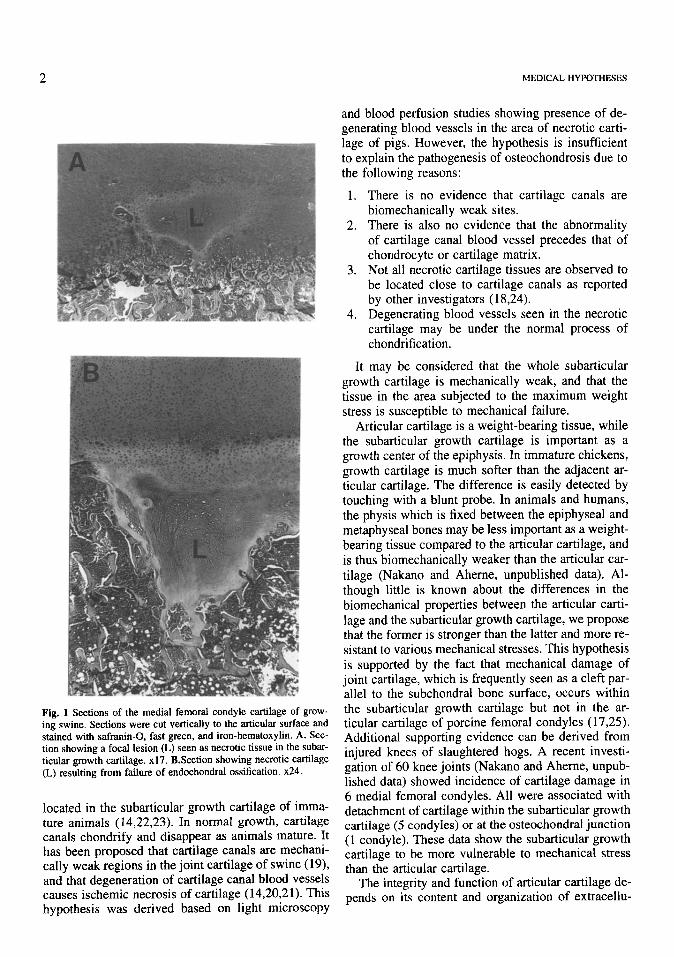
is now accepted by many investigators (1,2,5,14-18) that an early focal lesion occurs in the subarticular growth cartilage (Fig. 1A). The lesion persists and results in failure of endochondral ossification when the ossification front reaches the site of the lesion (Fig. 1B). It is interesting to speculate as to why or how lesions are more frequently developed in the sub- articular growth cartilage than in the articular carti- lage. Since osteochondrotic lesions are consistently observed in the weight-bearing surface of the medial femoral condyle (4,13), mechanical stress appears to be an important etiological factor. Whether the sub- articular growth cartilage is biomechanically weaker than the articmar cartilage is unknown. The aim of this paper is to present a hypothesis to explain the pathogenesis of early cartilage lesions that may result in failure of endochondral ossification.
Discussion
Recent studies have been focused on cartilage canals in growing pigs (14,18-20). Cartilage canals are channels containing blood vessels and nerves, and are
2 MEDICAL HYPOTHESES
Fig. 1 Sections of the medial femoral condyle cartilage of grow- ing swine. Sections were cut vertically to the articular surface and stained with safranin-0, fast green, and iron-hematoxylin. A. Sec- tion showing a focal lesion (L) seen as necrotic tissue in the subar- titular growth cartilage. x17. B.Section showing necrotic cartilage (L) resulting from failure of endochondral ossification. x24.
located in the subarticular growth cartilage of imma- ture animals (14,22,23). In normal growth, cartilage canals chondrify and disappear as animals mature. It has been proposed that cartilage canals are mechani- cally weak regions in the joint cartilage of swine (19), and that degeneration of cartilage canal blood vessels causes ischemic necrosis of cartilage (14,20,21). This hypothesis was derived based on light microscopy
and blood perfusion studies showing presence of de- generating blood vessels in the area of necrotic carti- lage of pigs. However, the hypothesis is insufficient to explain the pathogenesis of osteochondrosis due to the following reasons:
There is no evidence that cartilage canals are biomechanically weak sites. There is also no evidence that the abnormality of cartilage canal blood vessel precedes that of chondrocyte or cartilage matrix. Not all necrotic cartilage tissues are observed to be located close to cartilage canals as reported by other investigators (18,24). Degenerating blood vessels seen in the necrotic cartilage may be under the normal process of chondrification.
It may be considered that the whole subarticular growth cartilage is mechanically weak, and that the tissue in the area subjected to the maximum weight stress is susceptible to mechanical failure.
Articular cartilage is a weight-bearing tissue, while the subarticular growth cartilage is important as a growth center of the epiphysis. In immature chickens, growth cartilage is much softer than the adjacent ar- titular cartilage. The difference is easily detected by touching with a blunt probe. In animals and humans, the physis which is fixed between the epiphyseal and metaphyseal bones may be less important as a weight- bearing tissue compared to the articular cartilage, and is thus biomechanically weaker than the articular car- tilage (Nakano and Aherne, unpublished data). Al- though little is known about the differences in the biomechanical properties between the articular carti- lage and the subarticular growth cartilage, we propose that the former is stronger than the latter and more re- sistant to various mechanical stresses. This hypothesis is supported by the fact that mechanical damage of joint cartilage, which is frequently seen as a cleft par- allel to the subchondral bone surface, occurs within the subarticular growth cartilage but not in the ar- titular cartilage of porcine femoral condyles (17,25). Additional supporting evidence can be derived from injured knees of slaughtered hogs. A recent investi- gation of 60 knee joints (Nakano and Aherne, unpub- lished data) showed incidence of cartilage damage in 6 medial femoral condyles. All were associated with detachment of cartilage within the subarticular growth cartilage (5 condyles) or at the osteochondral junction (1 condyle). These data show the subarticular growth cartilage to be more vulnerable to mechanical stress than the articular cartilage.
The integrity and function of articular cartilage de- pends on its content and organization of extracellu-
PATHOGENESIS OF OSTEOCHONDROSIS
lar matrix macromolecules, collagen and proteogly- can, which provide the tissue with tensile and com- pressive strength, respectively (26-28). Chondrocytes are in turn responsible for the growth and mainte- nance of their adjacent matrix, including the regu- lation of both biosynthesis and turnover of the ma- trix molecules. Chondrocytes produce enzymes to degrade collagen and proteoglycan, which are re- placed by newly formed molecules. Neutral metallo- proteinases (matrixins) including collagenase, gelati- nase and stromelysin (proteoglycanase) are thought to be major enzymes involved in cartilage matrix degra- dation (29-32). These enzymes are secreted extra- cellularly as zymogens that later require activation by other proteinases (30). All matrixins are virtu- ally inhibited by tissue inhibitor of metalloproteinases (TIMP), which is believed to have a role in regulating the activity of matrixins and thereby preventing inap- propriate destruction of the matrix (29-32). Unfortu- nately, there is no information available on the differ- ence in chemical composition and metabolic activities between the articular cartilage and the subarticular growth cartilage.
It has been suggested that a certain amount of me- chanical pressure is essential for synthesis of cartilage matrix proteins (33-37). However, tissues have a lim- ited ability to withstand mechanical load, and an ex- cessive stress causes damage to joint tissues (38-40). For example, osteoarthrosis results from mechanical failure of articular cartilage. In such a condition, the activity of extracellular matrix proteinases, including collagenase and proteoglycanase, are abnormally el- evated (41-44) and disorganization of collagen ul- trastructure (45) and reduced cartilage proteoglycan content (46,47) are observed.
Proteoglycan is also affected in osteochondric joints (4). Histochemical examination of porcine femoral condyle cartilage indicated a loss of safranin- 0 staining in the osteochondrotic cartilage tissues (17,48) including the necrotic cartilage found in the subarticular growth cartilage (17) (Fig. IA). Safranin- 0 is a cationic dye which stains proteoglycan (49). Advanced lesions were also reported to contain lower than normal levels of both collagen and proteoglycan
(17). Degradation of cartilage matrix proteins is most
commonly associated with the action of enzymes. We propose that the loss of proteoglycan observed in the early osteochondrotic cartilage (Fig. 2A) is due to the action of proteoglycan degrading enzyme(s). This hypothesis is partially supported by the fcllowing ex- perimental results, which demonstrate the presence of proteoglycan degrading activity in the zones of chon-
3
drocyte proliferation and maturation in the subarticu- lar growth cartilage of swine.
T x .?z
ti al E .- !z .-
s cn
A
I
I II III II I I
i
6 I II III
II I I 1
Region Region
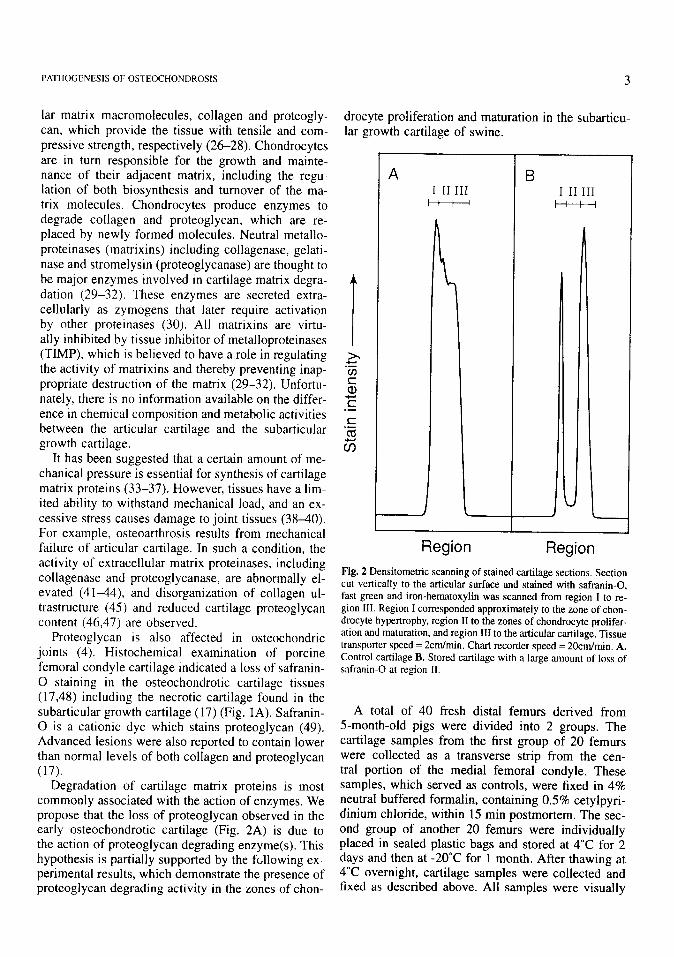
Fig. 2 Densitometric scanning of stained cartilage sections. Section cut vertically to the articular surface and stained with safranin-0, fast green and iron-hematoxylin was scanned from region I to re- gion III. Region I corresponded approximately to the zone of chon- drocyte hypertrophy, region II to the zones of chondrocyte prolifer- ation and maturation, and region III to the articular cartilage. Tissue transporter speed = 2cm/min. Chart recorder speed = 20cm/min. A. Control cartilage B. Stored cartilage with a large amount of loss of safranin-0 at region II.
A total of 40 fresh distal femurs derived from Smonth-old pigs were divided into 2 groups. The cartilage samples from the first group of 20 femurs were collected as a transverse strip from the cen- tral portion of the media1 femoral condyle. These samples, which served as controls, were fixed in 4% neutral buffered formalin, containing 0.5% cetylpyri- dinium chloride, within 15 min postmortem. The sec- ond group of another 20 femurs were individually placed in sealed plastic bags and stored at 4°C for 2 days and then at -20°C for 1 month. After thawing at 4°C overnight, cartilage samples were collected and fixed as described above. All samples were visually

4
normal. They were routinely processed and embedded in paraffin. Sections (7pm thick) were then cut verti- cally to the articular surface and stained with safranin- 0, fast green and iron-hematoxylin (49). Sections were then densitometrically scanned at 530nm. All control samples appeared to be stained intensely with safranin-0 through the whole thickness. However, the densitometric scanning of safranin-0 stained tissues indicated that the intensity was greatest at region I corresponding to the zone of chondrocyte hypertro- phy, and gradually decreased toward the articular sur- face (Fig. 2A). 6 (30%) stored samples showed loss of safranin-0 staining in region II including the zones of chondrocyte proliferation and maturation (Fig. 2B). The other portions of joint cartilage including articu- lar cartilage (region III) and the zone of chondrocyte hypertrophy were apparently not affected (Fig. 2B). Staining patterns were consistent between sections cut vertically and tangentially to the articular surface. The loss of safranin-0 staining in the stored tissues was much more extensive than that seen in pathological cartilage associated with focal degeneration (Fig. 1). The remaining 14 stored samples showed safranin-0 staining patterns apparently similar to those of control samples. Proteoglycan contents of these tissues were not determined. These data suggest that the degra- dation of proteoglycan occurred in the abnormally stained cartilage during the period of storage. Di- gested proteoglycan fragments were probably washed away’ from the cut sections of stored cartilage during the period of fixing to staining. The results further suggest a higher potential for proteoglycan (and prob- ably collagen) destruction in the zones of chondrocyte proliferation and maturation than in the remaining portions of joint cartilage. The reason for this differ- ence is unknown.
Hypothesis
The data discussed above led to the following hy- pothesis concerning the pathogenesis of early lesions in osteochondrosis. The subarticular growth cartilage, where early lesions occur, is a biomechanically weak tissue in the joint cartilage of growing animals. Ex- cessive mechanical stress on the subarticular growth cartilage (e.g. stress associated with rapid increase in body weight) may cause alterations in the metabolism of the matrix or chondrocytes. It is uncertain which is affected first. Under a stress situation, proteinases, including proteoglycanase and collagenase, and their inhibitors and activators escape the normal mecha- nisms of control, which results in abnormally elevated activities of these enzymes. Such enzymatic activity leads to degeneration of the cartilage matrix instead of resulting in normal tissue turnover.
MEDICAL HYPOTHESES
Conclusion
The hypothesis was developed as to the pathogenesis of early osteochondrotic lesions in the joint cartilage: It was proposed that mechanical stress induces de- rangements of cartilage metabolism which results in tissue destruction involving proteinases. To substan- tiate this hypothesis, it will be necessary to exam- ine the biomechanical properties of the two types of cartilaginous tissues, articular cartilage and subarticu- lar growth cartilage. Enzymes from these tissues and their action must be analyzed. Such studies may sug- gest new approaches in the prevention and treatment of osteochondrosis.
References
1.
2.
3.
4.
Grondalen T. Osteochondrosis and arthrosis in pigs. I. Inci- dence in animals up to 120 kg liveweight. Acta Vet Stand 1974; 15: l-25. Reiland S. Pathology of so-called leg weakness in the pig, Acta Radio1 1978: 358 (Suppl): 23-44. Pedersen N C, Pool R R, O’Brien T R. Naturally occurring arthropathies of animals. Diagnosis of Bone and Joint Disor- ders, Vol 1. Philadelphia: W B Saunders, 1981: 221-251. Nakano T, Brennan J J, Aheme F X. Leg weakness and os- teochondrosis in swine: a review. Can J Anim Sci 1987; 67: 883-901.
5.
6.
I.
8.
9.
10.
11.
12.
13.
Hill M A, Ruth G R, Hilley H D et al. Dyschondroplasias, including osteochondrosis, in boars between 25 and 169 days of age: histologic changes. Am J Vet Res 1984; 45: 903-916. Kato M, Onodera T. Spontaneous osteochondrosis in rats. Lab Anim 1984; 18: 1799187. Rejno S, Stromberg B. Osteochondrosis in the horse. II. Pathology. Acta Radio1 1978; 358 (Suppl): 153-178. Reiland S, Stromberg B, Olsson S-E et al. Osteochondrosis in growing bulls: pathology, frequency and severity on different feedings. Acta Radio1 1978; 358 (Suppl): 179-196. Riddel C. Skeletal deformities in poultry. Adv Vet Sci Comp Med 1981; 25: 277-310. Siffert R S. Classification of the osteochondroses. Clin Orthop 1981; 158: 10-18. Omer G E. Primary articular osteochondroses. Clin Orthop 1981; 158: 3340. Pappas A M. Osteochondrosis dissecans. Clin Orthop 1981; 158: 59-69.
14.
15.
16.
17.
Brennan J J, Aheme F X. Effect of dietary calcium and phos- phorous levels on performance, bone bending moment and the severity of osteochondrosis and lameness in boars and gilts slaughtered at 100 or 130 kg body weight. Can J Anim Sci 1986; 66: 177-790. Carlson C S, Meuten D J, Richardson D C. Ischemic necrosis of cartilage in spontaneous and experimental lesions of osteo- chondrosis. J Orthop Res 1991; 9: 317-329. Olsson S-E, Reiland S. The nature of osteochondrosis in ani- mals. Acta Radio1 1978; 358 (Suppl): 299-306. Nakano T. Joint abnormality and leg weakness in swine. The- sis, University of Alberta, Edmonton, Canada, 1980. Nakano T, Aheme F X, Thompson J R. Effects of feed re- striction, sex and diethylstilbestrol on the occurrence of joint lesions with some histological and biochemical studies of the articular cartilage of growing-finishing swine. Can J Anim Sci 1979; 59: 491-502.
18. Ekman S, Rodriguez-Martinez H. Ploen L. Morphology of

PATHOGENESIS OF OSTEOCHONDROSIS 5
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
normal and osteochondrotic porcine articular-epiphyseal car- tilage. Acta Anat 1990; 139: 239-253. Kincaid S A, Allhands R V, Pijanowski G J. Chondrolysis as- sociated with cartilage canals of the epiphyseal cartilage of the distal humerus of growing pigs. Am J Vet Res 1985; 46: 726-132. Carlson C S, Hilley H D, Meuten D J. Degeneration of car- tilage canal vessels associated with lesion of osteochondrosis in swine. Vet Pathol 1989; 26: 47-54. Woodard J C, Becker H N, Poulos P W Jr. Articular cartilage blood vessels in swine osteochondrosis. Vet Path01 1987; 24: IIS-123. Stockwell R A. Biology of cartilage cells. Cambridge, Lon- don: Cambridge University Press, 1979. Kuettner K E, Pauli B U. Vascularity of cartilage. In: Hall B K, ed. Cartilage, Structure, Function, and Biochemistry, Vol 1. New York: Academic Press. 1983: 281-312. Visco D M, Hill M A, Van Sickle D C et al. Cartilage canals and lesions typical of osteochondrosis in growth cartilage from the distal part of the humerus of new born pigs. Vet Ret 1991; 128: 221-228. Walker B, Aheme F X. The effect of mixing stress on the incidence of osteochondrosis in pigs: scanning electron mi- croscopy. Can J Anim Sci 1988; 68: 591602. Kempson G E, Muir H, Swanson S A V et al. Correlations be- tween the compressive stiffness and chemical constituents of human articular cartilage. Biochim Biophys Acta 1970; 215: 70-77. Kempson G E, Muir H, Pollard C et al. The tensile properties of the cartilage of human femoral condyles related to the con- tent of collagen and glycosaminoglycans. Biochim Biophys Acta 1973; 297: 456-472. Myers E R, Mow V C. Biomechanics of cartilage and its re- sponse to biomechanical stimuli. In: Hall B K, ed. Cartilage, Structure, Function, and Biochemistry, Vol 1. New York: Aca- demic Press, 1983: 313-341. Woessner J F Jr. Matrix metalloproteinases and their inhibitors in connective tissue remodelling. FASEB J 1991; 5: 2145- 2154. Dean D D. Proteinase-mediated cartilage degradation in os- teoarthritis. Semin Arthritis Rheum 1991; 20 (Suppl2): 2-11. Malemud C J. Changes in proteoglycans in osteoarthritis: biochemistry. ultrastructure and biosynthetic processing. J Rheumatol 1991; 18 (Suppl 27): 60-62. Cawston T E. Protein inhibitors or metallo-proteinases. In: Barrett A J, Salvesen G, eds. Proteinase Inhibitors, Amster- dam: Elsevier, 1988: 589-610. Roy S. Ultrastructure of articular cartilage in experimental im- mobilization. Ann Rheum Dis 1970; 29: 637-642. Palmoski M J, Colyer R A, Brandt K D. Joint motion in the absence of normal loading does not maintain normal articular
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
cartilage. Arthritis Rheum 1980; 23: 325-334. Langenskiold A, Michelsson J-E, Viedman T. Osteoarthritis of the knee in the rabbit produced by immobilization. Attempts to achieve a reproducible model for studies on pathogenesis and therapy. Acta Orthop Stand 1979; 50: 1-14. Kiviranta I, Tammi M, Jurvelin J et al. Moderate running exer- cise augments glycosaminoglycans and thickness of articular cartilage in the knee joint of young Beagle dogs. J Orthop Res 1988; 6: 188-195. Saamanen A-M, Tammi M, Kiviranta I et al. Running exercise as a modulator of proteoglycan matrix in the articular cartilage of young rabbits. Int J Sports Med 1988; 9: 127-132. Trias A. Effect of persistent pressure on the articular cartilage. An experimental study. J Bone Joint Surg 1961; 43B: 376 386. Vasan N. Effects of physical stress on the synthesis and degra- dation of cartilage matrix. Connect Tiss Res 1983; 12: 49-58. Brennan J J, Aheme F X, Thompson Jr et al. Effect of in- creased weight stress on joint integrity, synovial fluid com- position and cathepsin B-like activity of articular cartilage in growing swine. Can J Anim Sci 1987; 67: 525-533. Ehrlich M G, Home P A, Vigliani G et al. Correlation be- tween articular cartilage collagenase activity and osteoarthri- tis. Arthritis Rheum 1978; 21: 761-766. Pelletier J-P, Martel-Pelletier J, Howell D S et al. Collagenase and collagenolytic activity in human osteoarthritic cartilage. Arthritis Rheum 1983; 26: 63-68. Martel-Pelletier J, Pelletier J-P, Coutier J-M et al. Neutral proteases capable of proteoglycan digesting activity in os- teoathritic and normal human articular cartilages. Arthritis Rheum 1984; 27: 305-312. Dean D D, Martel-Pelletier J, Pelletier J-P et al. Evidence for metalloproteinase arid metalloproteinase inhibitor imbalance in human osteoarthritis cartilage. J Clin Invest 1989; 84: 678- 685. Wiltberger H, Lust G. Ultrastructure of canine articular cart- lage: comparison of normal and degenerative (osteoarthritic) hip joints. Am J Vet Res 1975; 36: 727-740. Lust G, Pronsky W. Glycosainmoglycan contents of normal and degenerative articular cartilage from dogs. Clin Chim Acta 1972; 39: 281-286. Mankin H J. Lippiello L. Biochemical and metabolic abnor- malities in articular cartilage from osteo-arthritic human hips, J Bone Joint Surg 1970; 52A: 424-434. Nakano T, Aheme F X, Thompson J R. Changes in swine knee articular cartilage during growth. Can J Anim Sci 1979; 59: 167-179. Rosenberg L. Chemical basis for the histologic use of safranin 0 in the study of articular cartilage. J Bone Joint Surg 1971; 53A: 69-82.

























![[Micro] pathogenesis](https://img.pdfslide.us/doc/110x75/55a726df1a28ab7e5e8b45a7/micro-pathogenesis.jpg)