Embed Size (px)
Citation preview

THE PATHOGENESIS OF EHV1 IN HORSES: NOVEL
INSIGHTS FROM EXPERIMENTAL INOCULATIONS AND
FIELD SITUATIONS
Gryspeerdt Annick
Thesis submitted in fulfilment of the requirements for the degree of doctor (Ph.D.) in
Veterinary Sciences, Faculty of Veterinary Medicine, Ghent University, 2011
Promotors:
Prof. Dr. H. Nauwynck
Laboratory of Virology
Department of Virology, Parasitology and Immunology
Prof. Dr. G. Van de Walle
Department of Comparative Physiology and Biometrics
Faculty of Veterinary Medicine


3
ISBN: 9789058642554 EAN: 9789058642554
© 2011 by Laboratory of Virology, Ghent University, Salisburylaan 133, 9820
Merelbeke, Belgium
The author and promoter give the authorization to consult and copy parts of this work for
personal use only. Every other use is subject to the copyright laws. Permission to
reproduce any material contained in this work should be obtained from the author.
Annick Gryspeerdt was supported by a doctoral grant from the Institute for the promotion
of Innovation through Science an Technology Flanders (IWT-Vlaanderen).


Table of contents
5
TABLE OF CONTENTS
LIST OF ABBREVIATIONS
I INTRODUCTION
1. EQUINE HERPESVIRUS 1 (EHV1): general characteristics
1.1 History
1.2 General characteristics
2. EQUINE HERPESVIRUS 1 and the cell
2.1 Virus structure
2.2 Replication cycle
3. EQUINE HERPESVIRUS 1 and the horse
3.1 Epidemiology and prevalence
3.2 Pathogenesis and disease
3.3 Diagnosis
3.4 Disease control
II AIMS OF THE THESIS
III DIFFERENCES IN REPLICATION KINETICS AND CELL TROPISM
BETWEEN NEUROVIRULENT AND NON-NEUROVIRULENT EHV1
STRAINS DURING THE ACUTE PHASE OF INFECTION IN HORSES
IV EXPRESSION OF LATE VIRAL PROTEINS IS HAMPERED IN
INFECTED NASAL MUCOSAL LEUKOCYTES BUT NOT IN
EPITHELIAL CELLS DURING EARLY PATHOGENESIS OF EQUINE
HERPESVIRUS TYPE 1 (EHV1) INFECTION
V DESCRIPTION OF AN UNUSUALLY LARGE OUTBREAK OF
NERVOUS SYSTEM DISORDERS CAUSED BY EQUINE HERPESVIRUS
1 (EHV1) IN 2009 IN BELGIUM
VI GENERAL DISCUSSION
SUMMARY
SAMENVATTING
CURRICULUM VITAE
BIBLIOGRAPHY
DANKWOORD


7
LIST OF ABBREVIATIONS
Ab anitbody
ACV acyclovir
AHV asinine herpesvirus
BM basement membrane
C complement
CD cluster of differentiation
CML cell of the monocytic lineage
CNS central nervous system
CO2 carbon dioxide
ConA concanavalin A
CSF cerebrospinal fluid
CTL cytotoxic T-lymphocyte
D aspartic acid
DC dendritic cell
DMEM Dulbecco’s modified eagle medium
DMSO dimethyl sulfoxide
DNA deoxyribonucleic acid
dpi days post inoculation
E early
EEL equine embryonic lung
EHM equine herpes myelitis
EHV equine herpesvirus
ELISA enzyme-linked immunosorbent assay
ER endoplasmatic reticulum
FCS fetal calf serum
FITC fluoresceine isothiocyanate
g glycoprotein
HCMV human cytomegalovirus
hpi hours post inoculation
IE1 immediate early polypeptide

8
IFN interferon
Ig immunoglobulin
IL interleukin
IONO ionomycin
IPMA immunoperoxidase monolayer assay
IR1 immediate early gene
IV intravenous
L late
mAb monoclonal antibody
MEM minimum essential medium
MHC major histocompatibility complex
MLV modified-live vaccine
mRNA messenger ribonucleic acid
ORF open reading frame
PBMC peripheral blood mononuclear cells
PBS phosphate buffered saline
PCR polymerase chain reaction
PDB phorbol dibutyrate
PHA phytohaemagglutinin
PMW pokeweed mitogen
PRV pseudo rabies virus
RK13 rabbit kidney 13
RLFP restriction length polymorphism
ROCK Rho-associated coiled-coil kinase
RPMI Roswell Park Memorial Institute
RSD arginine-serine-aspartic acid
SD standard deviation
SN seroneutralization
SNP single nucleotide polymorphism
siRNA small interfering ribonucleic acid
TAP transporter associated with antigen processing
TGN trans-Golgi network
TCID tissue culture infectious dose

9
TK thymidine kinase
TR Texas Red
URT upper respiratory tract
USDA United States Department of Agriculture
VACV valacyclovir
VN virus neutralizing
WAHV wild ass herpesvirus
ZHV zebra herpesvirus


Review of the literature
11
CHAPTER I
REVIEW OF THE LITERATURE
1. EQUINE HERPESVIRUS 1 (EHV1): general characteristics
1.1 History
1.2 General characteristics
2. EQUINE HERPESVIRUS 1 and the cell
2.1 Virus structure
2.2 Replication cycle
3. EQUINE HERPESVIRUS 1 and the horse
3.1 Epidemiology and prevalence
3.2 Pathogenesis and disease
3.2.1 Pathogenesis
3.2.2 Symptoms
3.2.3 Treatment
3.3 Diagnosis
3.4 Disease control


Review of the literature
13
1. EQUINE HERPESVIRUS 1 (EHV1): general characteristics
1.1 History
Equine herpesvirus 1 (EHV1) was first described in the early 1930s after a
necropsy examination of an aborted fetus (Dimock & Edwards, 1933), but virus
isolation from such cases succeeded only decades later (Jackson & Kendrick, 1971;
Saxegaard, 1966). The first recorded observation to indicate that major antigenic
differences may exist among clinical isolates of EHV1, was made in 1959 (Shimizu et
al., 1959). Thirteen years elapsed before a high correlation between antigenic subtype
and anatomic origin of the virus isolates was described (Burrows & Goodridge, 1972).
Until 1981, EHV1 and EHV4 were considered as two subtypes of the same virus,
namely ‘EHV1’, causing febrile rhinopneumonitis, ataxia, abortions and neonatal foal
disease in horses (Allen & Bryans, 1986). Unambiguous differentiation into EHV1
and EHV4 came from viral genomic fingerprints or so-called restriction length
polymorphisms (RLFP), but official recognition of the distinction between those two
viruses came almost one decade later (Roizman et al., 1992; Sabine et al., 1981;
Studdert et al., 1981; Turtinen et al., 1981).
1.2 General characteristics
1.2.1 Classification
The revised family Herpesviridae retains the viruses of mammals, birds and
reptiles. Members are highly disseminated in nature, with isolations from most animal
species of at least one and frequently several diverse herpesviruses (Davison et al.,
2009; Roizman & Pellett, 2001). Assignment to the Herpesviridae family is made by
the criterion of the characteristic herpes virion architecture. The known herpesviruses
appear to share four significant biological properties: (1) although the exact array of
enzymes may vary from one herpesvirus to another, all herpesviruses specify a large
array of enzymes involved in nucleic acid metabolism, DNA synthesis and processing
of proteins, (2) synthesis of viral DNA and capsid assembly occurs in the nucleus, (3)
production of infectious progeny virus is always accompanied by the destruction of

Review of the literature
14
the infected cell and (4) herpesviruses are able to remain latent in their natural hosts
(Roizman & Pellett, 2001).
Herpesviruses differ with respect to their biologic properties and are classified into
three different subfamilies based on host range, length of the reproduction cycle,
spread in cell culture, destruction of infected cells and capacity to establish latent
infections primarily but not exclusively in sensory ganglia (Roizman & Pellett, 2001):
(i) members of the Alphaherpesvirinae show a variable host range, short reproduction
cycle, rapid spread in cell culture, efficient destruction of infected cells and capacity
to establish latent infections primarily, but not exclusively, in sensory ganglia; (ii) a
restrictive host range is rather characteristic for the members of the subfamily
Betaherpesvirinae. They have a long reproduction cycle, with a slow infection
progress in cell culture. Cytomegalia is frequently seen in infected cells and latency
can be found in secretory glands, lymphoreticular cells, kidneys and other tissues. (iii)
Finally, host range of the members of the Gammaherpesvirinae is limited to the
family or order to which the natural host belongs. Replication is followed by a lytic
infection and latency is frequently demonstrated in lymphoid tissue.
Equid herpesvirus 1 (EHV1) or equine abortion virus is a member of the order
Herpesvirales, Family Herpesviridae, subfamily Alphaherpesvirinae, genus
Varicellovirus (Davison et al., 2009).
1.2.2 Herpesviruses in equids
In equids, fourteen herpesviruses have been identified so far: six belong to the
subfamily Alphaherpesvirinae and eight to the Gammaherpesvirinae (Table 1). The
horse is the natural host to alphaherpesvirus types 1 (EHV1), 3 (EHV3, coital
exanthema virus), 4 (EHV4) and gammaherpesvirus types 2 (EHV2) and 5 (EHV5).
The donkey is the host of EHV1-homologue asinine herpesvirus type 3 (AHV3),
EHV3-homologue asinine herpesvirus type 1 (AHV1) and asinine gammaherpesvirus
type 2 (EHV2) (Browning et al., 1988; Crabb & Studdert, 1990; 1995; Ficorilli et al.,
1995). In 2002, two new herpesviruses (AHV4 and AHV5) were isolated from the
lungs of donkeys suffering from respiratory disease (Kleiboeker et al., 2002). The
isolation of a new EHV1-related neurotropic virus from a gazelle (Gazelle
herpesvirus- 1, EHV9) and an unknown herpesvirus from a donkey with neurological

Review of the literature
15
disease, indicates that the list of presently known equid herpesviruses is likely to grow
(Fukushi et al., 1997; Hartley et al., 1999; Vengust et al., 2008).
Table 1. Known herpesviruses of equids
Virus
Abbreviation Subfamily Host Clinical signs
Equine herpesvirus
1
EHV1 α horse respiratory disease, abortion, neonatal
foal death, neurological disorders
Equine herpesvirus
2
EHV2 γ horse rhinitis, conjunctivitis,
immunodepression
Equine herpesvirus
3
EHV3 α horse coital exanthema
Equine herpesvirus
4
EHV4 α horse respiratory disorders, (abortion)
Equine herpesvirus
5
EHV5 γ horse respiratory tract disease, multinodular
pulmonary fibrosis
Asinine herpesvirus
1
AHV1
(EHV6)
α donkey lesions on external genitalia and udder
Asinine herpesvirus
2
AHV2
(EHV7)
γ donkey ?
Asinine herpesvirus
3
AHV3
(EHV8)
α donkey rhinitis?
Asinine herpesvirus
4
AHV4 γ donkey pneumonia
Asinine herpesvirus
5
AHV5 γ donkey pneumonia
?
? γ donkey neurological disease
Zebra herpesvirus 1
ZHV1 γ Zebra ?
Wild ass
herpesvirus
WAHV γ wild
ass
?
Gazelle herpesvirus
9
EHV9 α gazelle encephalitis

Review of the literature
16
Of all equid herpesviruses, EHV1 and EHV4 are clinically, economically and
epidemiologically the most relevant pathogens. EHV1 infections can result in
respiratory disease, abortion, fatal neonatal illness and neurologic syndromes (Allen
& Bryans, 1986). In contrast to EHV1 infections, EHV4 infection is mostly limited to
signs of upper respiratory tract disease, while abortion and nervous system disorders
are extremely rare (Ostlund, 1993). EHV2 is suggested to be ubiquitous in the equine
population. EHV2 has been implicated in upper respiratory disease, pyrexia,
inappetance, lymphoadenopathy, immuno-suppression, general malaise and poor
performance, and a high prevalence of the virus could be found in horses with upper
respiratory tract disease, abortion and severe ataxia. However, a correlation between
clinical signs and EHV2 as a causative agent could not be determined (Borchers et al.,
1997; Borchers et al., 1998; Kershaw et al., 2001; Schlocker et al., 1995). EHV3
causes an acute and self-limiting venereal disease characterized by the formation of
papules, vesicles, ulcers, and pustules on genital mucosa and skin (Blanchard et al.,
1992; Pascoe et al., 1968). Little is known about the distribution and natural genesis
of EHV5. A relationship with respiratory tract disease is suspected and two recent
reports suggest an association between the development of equine multinodular
pulmonary fibrosis and EHV5 infection (Verryken et al., 2010; Williams et al., 2007;
Wong et al., 2008).

Review of the literature
17
2. EQUINE HERPESVIRUS 1 and the cell
2.1 Virus structure
Morphologically, herpesviruses are distinct from all other viruses. They contain a
linear double-stranded DNA genome of 125-290 kbp in the form of a torus (Furlong
et al., 1972). The DNA is contained within a icosadeltahedral capsid, which is
surrounded by a proteinaceous matrix, designated the tegument. The latter is
surrounded by a lipid envelope, containing membrane-associated proteins (Davison et
al., 2009; Roizman & Knipe, 2001; Roizman & Pellett, 2001) (Figure 1).
Figure 1. Electronmicroscopic photomicrograph (left) and schematic drawing (right) of an EHV1
virion (picture from University of Wisconsin)
Up till now, 12 different glycoproteins have been described for EHV1: gB, gC,
gD, gE, gG, gH, gI, gK, gL, gM, gN and gp2 (Turtinen & Allen, 1982). These
envelope proteins are involved in virus attachment, penetration, egress and cell-to-cell
spread.
Glycoprotein B is an essential glycoprotein of EHV1. Functional analysis of gB
revealed that this glycoprotein is clearly involved in the penetration process. In
common with gB homologues of herpes simplex virus and other herpesviruses, gB
also plays a role in the cell-cell fusion process (Wellington et al., 1996b). However,
gB-negative EHV1 retained some infectivity and the absolute requirement of gB for
Envelope glycoproteins
gB, gC, gD, gE, gG, gH,
gI, gK, gL, gM,
gN and gp2
Envelope
Tegument
Capsid
Double
stranded DNA

Review of the literature
18
fusion process in EHV1 is not entirely clear (Neubauer et al., 1997). Glycoprotein D
is the second essential protein of EHV1. Monoclonal antibodies against gD strongly
neutralize virus infectivity and inhibit the penetration of the virus into the cell,
suggesting an important and essential function of gD in viral cell penetration (Csellner
et al., 2000; Van de Walle et al., 2008a; Whittaker et al., 1992). Glycoprotein D is
also associated with spontaneous cell-cell fusion, which is consistent with a role in
virus spread and is one of the functions attributed to gD homologues in
alphaherpesviruses (Csellner et al., 2000; Wellington et al., 1996a).
Glycoprotein C was shown to play an important role in the early steps of infection
and in release of virions (Osterrieder, 1999). It also acts as a complement receptor on
EHV1-infected cells, capable of binding the third component of complement (C3)
(Huemer et al., 1995). Glycoprotein G is a viral chemokine-binding protein, capable
of binding a broad range of chemokines, including CCL3 and IL8, thus modulating
pulmonary inflammation and virus replication during an EHV1 infection (Bryant et
al., 2003; Van de Walle et al., 2007; Van de Walle et al., 2008b).
EHV1 glycoprotein M is involved in both virus entry and direct cell-to-cell spread
of virus (Osterrieder et al., 1996). Processing of gM inside the infected cell depends
on the expression of gN (Rudolph et al., 2002). Studies with an EHV1 recombinant
virus containing deletions of glycoprotein E and I showed that these glycoproteins
facilitate cell-to-cell spread and may be important factors in EHV1 virulence
(Matsumura et al., 1998). Glycoprotein K is also critically involved in direct cell-to-
cell spread, as well as in secondary envelopment and/or egress. It facilitates virus
penetration and is at least partially responsible for syncytium formation at later time
points post infection (Neubauer & Osterrieder, 2004).
Although the presence of gH and gL has been demonstrated in infected cells, their
functions in the replication cycle of EHV1 remain unclear (Robertson et al., 1991;
Stokes et al., 1996).
Last, gp2 is a unique glycoprotein that has no positional or structural counterpart
in herpesviruses of other species. Although its function is not fully known, it was
shown to function in virus attachment (Sun et al., 1996) and in vivo, gp2 is proposed
to function as an immuno-modulatory protein (Smith et al., 2005). The simultaneous
absence of gM and gp2 had an additive negative effect on egress but not on secondary
envelopment or cell-to-cell spread (Rudolph & Osterrieder, 2002).

Review of the literature
19
2.2 Replication cycle
The replication cycle of EHV1 is divided into several steps. A schematic
representation of the various steps is shown in Figure 2.
EHV1 attachment to cells is mediated via an interaction between glycoproteins
gB, gC and cellular heparan sulphate (Osterrieder, 1999). After attachment, an
interaction between gD and a putative entry receptor is required to complete the
fusion between the viral envelope and the cellular membrane (Csellner et al., 2000;
Frampton et al., 2005). Recently, it has been shown that Major Histocompatibility
Complex Class I acts as a functional entry receptor for this glycoprotein (Kurtz et al.,
2010; Sasaki et al., 2011). EHV1 can enter cells via fusion at the plasma membrane in
equine endothelial cells or by endocytosis/phagocytosis in peripheral blood
mononuclear cells (Van de Walle et al., 2008a). Successful infection through either
mechanism requires the activation of Rho-associated coiled-coil kinase 1 (ROCK1)
(Frampton et al., 2007). Recently, it has been shown that EHV1 entry via endocytosis
is triggered by the interaction between cellular integrins and the RSD motif present in
gD (Van de Walle et al., 2008a). Subsequent to fusion between the two lipid
membranes, the nucleocapsid is released into the cytoplasm of the infected cell, where
it is transported along microtubules to the nuclear pore of the cell. Transport is both
dependent on the integrity of the microtubule network and the minus-end microtubule
motor protein, dynein. Integrity is in part promoted by EHV1, as it actively induces
the acetylation of tubulin, a marker of microtubule stabilization. Also the cellular
kinase ROCK1 has an important role in virus trafficking to the nucleus (Frampton et
al., 2010). At the pore, the nucleocapsid releases its DNA into the nucleus, leaving an
empty capsid at the cytoplasmic side of the complex (Roizman & Pellett, 2001).
Transcription of the viral genome, replication of viral DNA and assembly of new
capsids take place in the nucleus, but all viral proteins are synthesized in the
cytoplasm. Viral DNA is transcribed throughout productive infection by host RNA
polymerase II, but with the participation of viral factors at all stages of infection.
EHV1 transcription is co-ordinately regulated into immediate-early (IE), early (E) and
late (L) phases in a cascade fashion (Caughman et al., 1985). Several of the gene
products are enzymes and DNA-binding proteins involved in viral DNA replication.
First, the immediate early gene, IR1, is expressed (Gray et al., 1987a; Gray et al.,

Review of the literature
20
1987b). This IE gene region is transcribed into a single mRNA, but multiple species
of the IE protein exist (Robertson et al., 1988). The major IE polypeptide (IE1)
functions as a regulatory protein that is capable of modulating gene expression from
both early and late promotors and also of trans-repressing its own promoter
(Caughman et al., 1985; Gray et al., 1987a). Early genes are expressed before DNA
replication and encode proteins required for DNA replication, e.g. DNA polymerase
and other enzymes such as thymidine kinase. These viral proteins promote viral DNA
replication, and in a late phase, expression of late genes is stimulated. The late genes
are expressed after viral DNA replication has started and their expression is enhanced
by other viral proteins. Late genes mostly encode viral structural components like
capsid proteins and envelope glycoproteins.
Like transcription, viral DNA replication takes place in the nucleus, resulting in
multiple copies of the EHV1 genome. These newly synthesized DNA copies have to
assemble with capsid proteins to form nucleocapsids. Upon translation and expression
in the cytoplasm, capsid proteins enter the nucleus, after which assembly and
incorporation of viral DNA molecules occur, a process called encapsidation.
Assembly occurs in several stages. DNA is packaged into preassembled capsids, after
which the virus matures and nucleocapsids bud at the inner leaflet of the nuclear
membrane, resulting in enveloped particles in the perinuclear space. Egress from the
perinuclear cisterna primarily occurs by fusion of the primary envelope with the outer
leaflet of the nuclear membrane, resulting in intracytoplasmic naked nucleocapsids
(Granzow et al., 2001; Mettenleiter, 2004; Mettenleiter et al., 2006). Inside the
cytoplasm, the majority of the tegument is added to the nucleocapsids (Mettenleiter,
2002; Mettenleiter et al., 2009). Unlike capsid proteins, viral envelope proteins are
co-translationally transported into the endoplasmatic reticulum (ER) and subsequently
incorporated into various cellular membranes while passing through the secretory
pathway (Granzow et al., 2001). The intracytoplasmic naked nucleocapsids are
enveloped at membranes of the trans-Golgi network (TGN) (Granzow et al., 2001).
The enveloped particles are then released through secretory vesicles. Little is known
about viral proteins involved in egress, i.e. transport of vesicles containing enveloped
virions to the plasma membrane and subsequent virus release by fusion of vesicle and
plasma membrane (Mettenleiter et al., 2009).

Review of the literature
21
Figure 2. The EHV1 replication cycle (adapted from Garré, 2008). The first step in the replication of EHV1
is the attachment of free virions to the surface of the target cell (1). After endocytosis (2a) and/or fusion of
the viral envelope with the plasma membrane (2b), the nucleocapsid is released into the cytoplasm (3) and
transported into the nucleus (4). DNA replication (5) and transcription (6) occur and RNA molecules are
transported to the cytoplasm and translated to proteins (7, 9). Capsid proteins are redirected into the nucleus
where encapsidation occurs (8). Viral envelope proteins are co-translationally transported into the
endoplasmatic reticulum (10) and subsequently incorporated into various cellular membranes, while passing
through the secretory pathway. The nucleocapsid leaves the nucleus via budding through the inner leaflet of
the nuclear membrane (11) and subsequent fusion with the outer leaflet of the nuclear membrane, resulting
in entry of naked nucleocapsids in the cytoplasm (12). The nucleocapsid acquires its secondary envelope at
the Golgi apparatus (13) and then leaves the cell via vesicle-mediated exocytosis (14, 15, 16)
2a
2b
3
4
2a
2b
3
4

Review of the literature
22
3. EQUINE HERPESVIRUS 1 and the horse
3.1 Epidemiology and prevalence
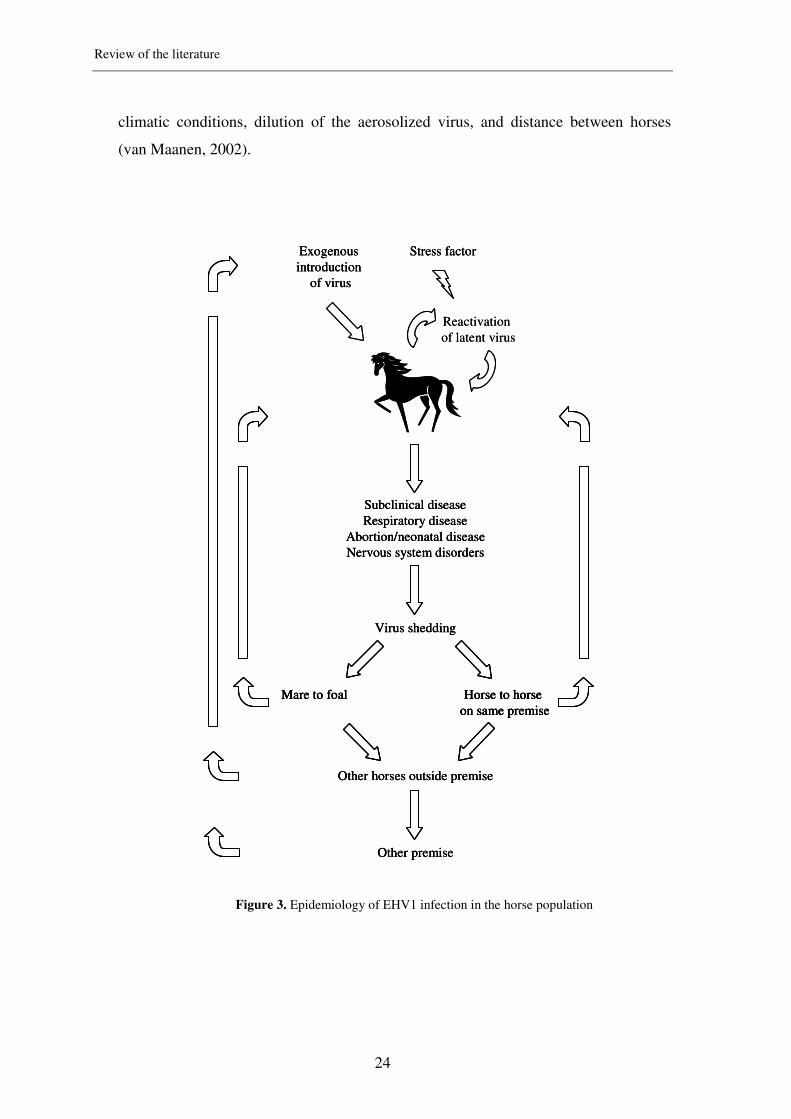
A schematic overview of the epidemiology of EHV1 is depicted in Figure 3
Equine herpesvirus 1 infections are endemic in horse populations worldwide, and
most animals are probably infected during the first year of life (Allen & Bryans, 1986;
Allen et al., 1999; Borchers et al., 2006). However, due to antigenic similarity
between EHV1 and EHV4, the interpretation of data from serological surveys was
complicated by the lack of a type-specific antibody test until early 1990 (Crabb &
Studdert, 1993). It should also be indicated that sero-epidemiological studies are
complicated in horse populations in which EHV1 and/or EHV4 vaccination is
practised. Retrospective testing of sera from Thoroughbred horses collected between
1967-1974 and 1993 gave values of 9-28% for EHV1-specific antibodies (Crabb &
Studdert, 1995). Another study found an incidence of EHV1 antibodies in mares and
foals of 26,2% and 11,4% respectively (Gilkerson et al., 1999).
Latency by alphaherpesviruses is an important epidemiological strategy ensuring
survival and spread within the natural host population. When horses are first infected,
latency is established in the lymph nodes of the respiratory tract, the trigeminal
ganglion and CD5+/CD8
+ leukocytes (Baxi et al., 1995; Chesters et al., 1997;
Edington et al., 1994; Slater et al., 1994). Months or years after primary infection, the
latent virus may again become manifest with renewed replication and with the
potential for initiating new outbreaks of disease in its host as well as in susceptible
cohorts. As EHV1 infection already occurs in the first weeks or months of life, it is
assumed that viral reactivation in latently infected mares leads to this early foal
infection (Gilkerson et al., 1999; Marenzoni et al., 2008). Therefore, lactating mares
may be the primary source of EHV1 infection of foals and the latter may further
transmit the virus to other mares and foals (Gilkerson et al., 1997). However, a recent
study could not demonstrate infection of sentinel horses with EHV1 after direct
contact with horses undergoing experimental corticosteroid-induced recrudescence of
latent infection (Pusterla et al., 2010). Failure to effectively transmit EHV1 to

Review of the literature
23
susceptible horses may have resulted from the low level and short period of viral
shedding in these latently infected horses.
Estimates of the prevalence of EHV1 infection based on viral detection
technologies vary strongly. Latent EHV1 could be found by polymerase chain
reaction (PCR) in 87.5% of examined bronchial lymph nodes and a similar level in the
trigeminal ganglia of 40 horses examined at slaughter (Edington et al., 1994). In pools
of tissue with peripheral blood leukocytes, nasal swabs and abattoir specimens from
116 animals, EHV1 DNA was found in 88% of the analysed samples in Brazil
(Carvalho et al., 2000). In a recent study, the prevalence of latent EHV1 in
submandibular lymph nodes was 54% (Allen et al., 2008). Current estimates of the
prevalence of latent EHV1 suggest a rate in excess of 60%, which is probably more
limited by detection technology than by the actual infection rate. We should presume
that the majority of horses is latently infected with EHV1 (Lunn et al., 2009). The
establishment of latency provides a permanent reservoir for the virus and makes total
disease eradication highly unlikely. It also diminishes the effectiveness of antiviral
therapy or immunotherapy and complicates vaccination protocols for control of EHV1
infections (Allen & Bryans, 1986).
It is thought that subclinical EHV1 infections are common in horses and play an
important role in EHV1 epidemiology, as they result in frequent spread and a high
risk of exposure particularly in open horse populations subjected to stress and
introduction of new animals. However, recent reports in both horses exposed to stress
and healthy horses demonstrated that subclinical shedding of EHV1 is infrequent and
at a very low level (Brown et al., 2007; Wang et al., 2007). These observations
suggest that spread of EHV1 among adult horses is typically accompanied by clinical
disease, either respiratory disorders, abortion or equine herpes myelitis (EHM) (Lunn
et al., 2009).
Either after recrudescence from latency or after exogenous introduction, EHV1
infections can spread rapidly within groups of horses. Evidence of widespread
horizontal infection by direct or indirect contact has been reported repeatedly for
outbreaks of EHV1 induced abortion and neurological disease (Drummer et al., 1995;
Friday et al., 2000; McCartan et al., 1995). Aerosol transmission cannot be excluded
and putatively depends on the amount of infectious virus released by the source,
Review of the literature
24
climatic conditions, dilution of the aerosolized virus, and distance between horses
(van Maanen, 2002).
Figure 3. Epidemiology of EHV1 infection in the horse population
Reactivation
of latent virus
Subclinical disease
Respiratory disease
Abortion/neonatal disease
Nervous system disorders
Virus shedding
Mare to foal Horse to horse
on same premise
Other horses outside premise
Other premise
Exogenous
introduction
of virus
Stress factor
Reactivation
of latent virus
Subclinical disease
Respiratory disease
Abortion/neonatal disease
Nervous system disorders
Virus shedding
Mare to foal Horse to horse
on same premise
Other horses outside premise
Other premise
Subclinical disease
Respiratory disease
Abortion/neonatal disease
Nervous system disorders
Virus shedding
Mare to foal Horse to horse
on same premise
Mare to foal Horse to horse
on same premise
Other horses outside premise
Other premise
Exogenous
introduction
of virus
Stress factorExogenous
introduction
of virus
Stress factor

Review of the literature
25
3.2 Pathogenesis and disease
3.2.1 Pathogenesis
A schematic overview of the complete pathogenesis is depicted in Figure 4.
Primary replication. Horses acquire an EHV1-infection through direct contact or
inhalation of infectious secretions (Allen & Bryans, 1986). After inhalation, EHV1
binds to and penetrates the epithelial cells of the upper and lower respiratory tract
(Kydd et al., 1994b; Patel et al., 1982). Following a rapid replication, EHV1
disseminates into the connective tissue, infecting local blood vessels and regional
lymph nodes (Kydd et al., 1994b; van Maanen, 2002). How the virus penetrates the
basement membrane and reaches the underlying connective tissue and lymphoid
tissue remains unsolved to date.
Viremia. Via a still unknown pathway, EHV1 infects mononuclear cells which
can enter the blood circulation resulting in a cell-associated viremia lasting for 8 to 18
days post infection (Edington et al., 1986; Gibson et al., 1992; Gleeson & Coggins,
1980; Patel et al., 1982). The exact identity of these carrier cells still remains
uncertain. A study from 1983 showed that T-lymphocytes, B-lymphocytes and
monocytes are all able to harbour EHV1, but the predominant mononuclear cells
harbouring virus were found to be T-lymphocytes (Scott et al., 1983). Remarkably,
after disruption of these cells, no infectious EHV1 could be found during virus
isolation, suggesting that EHV1 exists in mononuclear cells in a non-infective form,
or is blocked at a certain replication level until activation occurs (Gleeson & Coggins,
1980; Scott et al., 1983; van der Meulen et al., 2006). A later study confirmed the
latter hypothesis. The majority of infected PBMC do not show viral envelope proteins
on their surface during viremia (van der Meulen et al., 2003b). During a recent study
using PBMC isolated from experimentally infected horses, viral genomic DNA was
found by means of PCR techniques in all PBMC subpopulations, with CD8+
lymphocytes and B-lymphocytes as most frequently infected cells (Wilsterman et al.,
2010). These results indicate that different PBMC subpopulations may play different
roles in EHV1 viremia.

Review of the literature
26
Secondary replication. Transfer of EHV1 from viremic cells to endothelial cells
of target organs causes infection, followed by replication of EHV1 and subsequent
damage to these organs. Smith et al. showed that activation of adhesion molecules in
endothelial cells of the nasal mucosa and the reproductive tract is a key step in
transferring virus from infected leukocytes to endothelial cells. Indeed, an in vitro
flow system using equine veins and arteries showed that EHV1 was only transferred
to endothelial cells if both leukocytes and endothelial cells expressed these surface
molecules (Smith et al., 2002; Smith et al., 2001). An infection model with EHV1-
infected leukocytes on the one hand and carotid artery and brain endothelial cells on
the other hand has recently been established (Goehring et al., 2010a). In this model,
neutralizing antibodies could only prevent infection of endothelial cells when an
indirect contact model was used. In the case of a direct contact model between the two
cell types, neutralizing antibodies could not prevent transfer of virus to endothelial
cells. This further confirms the importance of the direct transfer mechanism of EHV1
from infected leukocytes to endothelial cells.
Transfer of EHV1 from infected leukocytes to placental maternal endothelial cells
plays a major role in the pathogenesis of abortion. Infection of endothelial cells in the
umbilical cord seems to be the primary route of dissemination of virus in the fetus.
These infected cells may then act as centres for the development of focal lesions of
parenchymal necrosis. Further spread of the virus with widespread vasculitis,
thrombosis, cotyledonary infections and ischemic damage to the endometrium with
replication of EHV1 in foetal tissues leads to subsequent abortion (Edington et al.,
1991).

Review of the literature
27
Figure 4. Schematic overview of the pathogenesis of EHV1. 1) Attachment/penetration and replication
of virus in epithelial surfaces of the upper respiratory tract; 2) spread of virus through the basement
membrane; 3) spread to connective tissue (3a) and draining lymph nodes (3b); 4) infection of
leukocytes; 5) leukocyte associated viremia; 6) transfer of virus from infected leukocytes to endothelial
cells of target organs; 7) replication and spread of virus in target organs. Full arrow: known
mechanism; striped arrow: unknown mechanism.
Secondary replication of EHV1 in the nervous system of horses causes EHM.
Different independent researchers agreed that lesions accompanying central nervous
system disorders were characterized by a focal vasculitis with associated oedema,
haemorrhage and thrombosis. Axonal and myelin degeneration occurred focally and
secondary to these vascular changes (Jackson et al., 1977; Mumford & Edington,
1980; Platt et al., 1980). As actual isolation of EHV1 from the nervous system is very
rare, it was postulated that lesions could be induced by circulation immune
complexes. However, in 1986, Edington et al. showed that although recovery of virus
1
3a
45
2
3b
67

Review of the literature
28
from the central nervous system (CNS) was low, immunofluorescence indicated that
endothelial tropism of the virus was the initiating and persisting feature. The action of
immune complexes in EHV1 infections seems to be secondary and localized
(Edington et al., 1986). In contrast to several other alphaherpesviruses, which can
cause encephalitis through primary neurotropism, EHV1 seems to be non-neurotropic
and its propensity to induce myeloencephalopathy reflects a marked
endotheliotropism (Edington et al., 1986; Jackson et al., 1977; Whitwell & Blunden,
1992; Wilson, 1997). Vascular lesions result in secondary hypoxic degeneration of
nervous tissue, causing nervous system disorders due to the impairment of blood flow
and metabolic exchange.
Abortion versus nervous system disorders. For many years, it has been
suspected that distinct isolates of EHV1 might be responsible for outbreaks of either
abortion and neonatal foal death or neurological disease. The difference in pathogenic
potential of EHV1 strains to induce abortion or neurological damage appears to be
correlated with differences in the ability to establish infection at vascular endothelial
sites, in particular within the endometrium and central nervous system (Edington et
al., 1986; Mumford et al., 1994; Patel et al., 1982; Platt et al., 1980). Infection with
neurovirulent strains may result in severe clinical disease, including a high frequency
of abortion and a less frequent occurrence of paralysis, while infection with non-
neurovirulent strains is restricted to mild respiratory signs, with infrequent induction
of abortion and no neurological disease (Mumford et al., 1994; Smith et al., 1992;
Smith et al., 2000). It was noticed that horses inoculated with neurovirulent strains of
EHV1 developed both a greater magnitude and duration of PBMC-associated viremia
than animals inoculated with non-neurovirulent strains of the virus, which contributes
to the risk for development of neurological signs (Allen & Breathnach, 2006).
Recently, it was demonstrated that a variation of a single amino acid is strongly
associated with neurological versus non-neurological disease outbreaks (Nugent et al.,
2006). This study compared different isolates recovered from neurological and non-
neurological outbreaks over the course of 30 years. It was found that a single
nucleotide polymorphism (SNP) in the catalytic subunit of the viral DNA polymerase
(pol), causing a substitution of asparagine (N) by aspartic acid (D) at amino acid
position 752 at the ORF30 position of the DNA polymerase, was a major contributory

Review of the literature
29
factor in determining whether a disease outbreak will include animals presenting
neurological signs. To strengthen the finding that the single amino acid exchange by
itself influences neurovirulence, several studies have been performed, using
recombinant viruses of non-neurological EHV1 strains with an inserted point
mutation for the neurological variant and vice versa, further confirming the previous
finding (Goodman et al., 2007; Van de Walle et al., 2009). However, horses with
neurological disease upon infection with a non-neurovirulent EHV1 isolate, as well as
a high prevalence of neurovirulent strains in equine abortions have been reported,
suggesting that other factors such as host and/or environmental factors will also
contribute to the onset of neurological disease (Fritsche & Borchers, 2010; Goehring
et al., 2006; Perkins et al., 2009; Pronost et al., 2010; Smith et al., 2010).
In recent years, outbreaks of neurological disease due to EHV1 infections have
been reported with increasing frequency and severity (Friday et al., 2000; Lunn et al.,
2009; McCartan et al., 1995; USDA, 2007; van Maanen et al., 2001). Unfortunately,
despite 40 years of worldwide research, EHV1-induced neurological disease cannot
be adequately prevented nor fully explained.
3.2.2 Symptoms
An overview of different symptoms following EHV1 infection, is shown in Figure 5.
Respiratory disorders. Primary replication of EHV1 in tissues of the upper
respiratory tract is mostly accompanied with signs of a mild, self-limiting upper
respiratory tract infection (Gibson et al., 1992). Infected horses spread large amounts
of virus via nasal secretions, allowing the virus to spread to other susceptible animals.
Infectious virus can be detected from day 1 till 7 to 14 days post infection (Gibson et
al., 1992; Heldens et al., 2001; van der Meulen et al., 2006) and in some cases even
till 21 days post infection (Garré et al., 2009; Patel et al., 1982).
Young horses usually develop fever, serous to mucopurulent nasal discharge and
swelling of draining lymph nodes, while older horses mainly show mild or subclinical
disease (Coggins, 1979). Conjunctivitis, depression and anorexia can be observed in
some cases (Edington et al., 1986; Kydd et al., 1994a; Patel et al., 1982).

Review of the literature
30
In sporadically reported cases, EHV1 targets the pulmonary endothelium in young
adults, which causes a severe pulmonary oedema (Del Piero & Wilkins, 2001; Del
Piero et al., 2000). Such severely affected horses die in acute respiratory distress.
Abortion and neonatal foal death. Secondary replication of EHV1 in endothelial
cells of the pregnant uterus plays a major role in the occurrence of EHV1-induced
abortion or neonatal death (Edington et al., 1991). The incubation period for abortion
after respiratory infection with EHV1 varies from 6 days (Gleeson & Coggins, 1980)
to 4 months (Mumford et al., 1987). Virologically positive foetuses, including
placenta, are often fresh and abortion occurs suddenly and spontaneously. However,
abortions with virological negative foetuses are also described (Gleeson & Coggins,
1980; Smith et al., 1992).
EHV1-induced abortion can be preceded by mild upper respiratory tract disease in
the mare but usually no signs of disease or warning signs of impending abortion are
noticed. Following abortion, virus is cleared rapidly from the genital tract, and future
breeding capacity is not impaired unless some uterine damage has occurred due to
dystocia (Smith, 1997). Although mares are not immune to subsequent infection,
EHV1-induced abortions in successive pregnancies are rare (Doll et al., 1955;
Ostlund, 1993). EHV1-induced abortions usually involve only 1 or 2 mares in a herd,
although sometimes abortion storms can occur in which more than 50 % of the foals
are lost (Mumford et al., 1987; Van Maanen et al., 2000).
If infection occurs close to term, the mare may deliver an apparently normal or
weak living foal. These foals usually develop jaundice and respiratory distress and
mostly die within 7 days post partum despite of intensive treatment, due to extensive
viral damage and secondary bacterial infection (Murray et al., 1998; Perkins et al.,
1999).
Field studies have shown that 95% of EHV1-induced abortions occur during the
last 4 months of gestation (Allen & Bryans, 1986; Doll, 1952; Doll & Bryans, 1963).
EHV1-abortion has been recorded on rare occasions as early as the fourth month of
gestation (Prickett, 1970). This apparent resistance of the early pregnant mare to
abortion is not completely understood.

Review of the literature
31
Nervous system disorders.
Secondary replication of EHV1 in the endothelial cells of blood vessels of the
nervous system is the first step in the development of nervous system disorders
(Edington et al., 1986). Neurological disease may affect either single horses or groups
of animals within a herd. An incubation period of 6 to 10 days between initial
infection and the occurrence of nervous system disorders has been described (Jackson
et al., 1977; van Maanen et al., 2001).
Although the disorders may be preceded by signs of respiratory disease such as
fever or nasal discharge one or two weeks earlier, they can also occur without any
previous signs (Carroll & Westbury, 1985; Greenwood & Simson, 1980; Stierstorfer
et al., 2002; van Maanen et al., 2001). Neurological disease is usually acute in onset
and can display a variable set of clinical signs. Most commonly, signs vary from mild
ataxia to complete paralysis, accompanied by bladder and/or rectal paralysis (Jackson
et al., 1977; Stierstorfer et al., 2002). In less common cases, oedema of the hind legs,
swollen testis, urticaria and central nervous disorders such as blindness and torticollis
can be observed (Friday et al., 2000; Greenwood & Simson, 1980; van der Meulen et
al., 2003a). The prognosis for horses with ataxia due to EHV1 myeloencephalitis is
favourable, with the possibility to regain their normal performance level. However, if
the horse becomes recumbent for more than 24 hours, the prognosis is grave and
euthanasia is indicated (McCartan et al., 1995; van Maanen et al., 2001).
Although animals of various age, gender and breed can be affected, a retrospective
study of Goehring et al. (2006) indicated that certain pony breeds and females were
more commonly affected. Although neurological problems have been reported in
weanlings, the clinical picture in young horses seems to be less severe than in adults,
suggesting age to be an important risk factor as well (Goehring et al., 2006;
Greenwood & Simson, 1980).

Review of the literature
32
Figure 5. Different EHV1-related symptoms. A: respiratory disorders; B: abortion; C: neonatal foal
disease; D: nervous system disorders
3.2.3 Treatment
Antiviral therapy. Treatment of EHV1 infections is limited to symptomatic care
since no specific antiviral drugs are available for treatment of EHV1 infections in
horses to date. Experimental antiviral drugs have yet to prove clinical efficacy and
several of them have been tested in field as well as under experimental conditions.
During several natural outbreaks of EHV1, treatment with the antiviral drug
acyclovir has been reported with conflicting results (Friday et al., 2000; Henninger et
al., 2007; van der Meulen et al., 2003a). However, since therapeutic evaluations in
case-control studies under field conditions are difficult due to a lack of good control
animals, great care should be taken in interpreting these results. Acyclovir was also
shown to have serious limitations due to poor oral bioavailability. Therefore, high
therapeutic concentrations are needed in order to reach the desired effect, making this
drug a costly medication with questionable efficacy (Garré et al., 2009; Garré et al.,
A
C
B
D

Review of the literature
33
2007; Maxwell et al., 2008a). When using the pro-drug valacyclovir, sufficiently high
acyclovir levels could be reached in plasma and nasal mucus after oral treatment, but
no effect of this treatment was seen on clinical signs, viral shedding and viremia of
EHV1-infected ponies (Garré et al., 2009). The latter was in contrast to the results of
another study, showing that oral treatment with valacyclovir significantly decreased
signs of EHV1 disease (Maxwell et al., 2008b). The protective effect was greatest
when therapy was initiated before viral infection and continued for two weeks.
However, initiation of therapy at the onset of fever also decreased sings of disease as
compared to control horses. These encouraging results should be viewed cautiously in
light of the small number of horses studied.
Recently, synthetic siRNAs against envelope glycoprotein B and the origin-
binding protein helicase were designed, and their efficacy to limit and prevent EHV1
infection was tested in vitro and in a murine model. The results obtained during this
study, indicated that siRNA treatment could be of great importance during an
outbreak of EHV1 in a horse population, as not only the severity of clinical disease
but also the number of affected animals and the magnitude of nasal shedding could be
reduced (Fulton et al., 2009). Although there was no significant difference in viral
shedding and viremia between treated and control groups during an in vivo
experiment with EHV1-infected horses, euthanasia necessitated in case of
neurological disease was significantly reduced after application of EHV1 specific
siRNA (Brosnahan et al., 2009).
Supportive therapy. Despite many experimental approaches, no systemic
antiviral therapeutic agents are approved for the use in horses or have a proven
efficacy. Therefore, supportive therapy is of paramount importance in the field.
Treatment strategies are aimed at easing clinical signs and preventing secondary
complications. The level of care necessary is dependent on the severity of clinical
signs.
Respiratory infections of immunocompetent animals are generally self-limiting
and require only good nursing care. Broad-spectrum antibiotics can be administered to
assist in combating secondary bacterial infections and high fever can be treated with
antipyretics. Stress avoidance is paramount and horses in training should receive
adequate rest before resuming work (Ostlund, 1993; van Maanen, 2002).

Review of the literature
34
Abortion occurs suddenly with complete expulsion of fetus and placenta and
complications are seldom. Therefore, after an uncomplicated partus, usually no
therapy for the aborting mare is required (Ostlund, 1993; van Maanen, 2002).
However, it is advisable to wash tail and hind quarters with disinfectants to avoid
further viral contamination of the environment and/or other animals (Goulden et al.,
1989).
Weak, but live born affected foals or, less frequently, foals that initially appear
healthy but rapidly deteriorate due to intractable bacterial infections should be treated
with intensive therapy like provision of warmth, oxygen, cardiovascular and
nutritional support and appropriate drugs (Murray et al., 1998; van Maanen, 2002).
However, these measures are almost always unsuccessful, as these foals are almost
always bound to die within a few days due to extensive damage in parenchymal and
lymphoreticular organs and subsequent bacterial infections (Allen & Bryans, 1986;
Goulden et al., 1989).
Animals showing nervous system disorders should receive appropriate intensive
care. Non-recumbent animals should be housed in a deeply bedded box, until
coordination improves. Severely ataxic horses may be temporarily assisted by a sling
to prevent secondary complications associated with recumbency. Regular urinary
catheterization and rectum evacuation may be required in case of bladder or rectum
paralysis (Friday et al., 2000; McCartan et al., 1995). Horses which are able to stand,
should be turned out or hand walked as soon as possible (Greenwood & Simson,
1980; McCartan et al., 1995; van Maanen et al., 2001). Recumbent animals not able
to be supported by a sling should be maintained in a sternal position in a well-bedded
stable if possible. They should have easy access to food and water. Intravenous
rehydratation may be necessary for horses unable to remain in sternal position. Horses
in lateral recumbency should be manually rotated every few hours. Antibiotics are
indicated because of the higher risk of developing aspiration pneumonia and/or
decubital ulcers. Horses can also be treated with corticosteroids in order to reduce the
inflammatory response in the nervous system. However, considerable care should be
taken when treating horses with such drugs during an outbreak, as they induce
immunosuppression and may stimulate a longer and higher spread of virus which can
make them even contra-indicated (Edington et al., 1985; Goehring & Sloet van
Oldruitenborgh-Oosterbaan, 2001; Slater et al., 1994). Therefore, only short-acting

Review of the literature
35
corticosteroids, if at all, should be used, for a limited period (McCartan et al., 1995;
Reed & Toribio, 2004). Other anti-inflammatory drugs such as dimethyl sulfoxide
(DMSO) and non-steroidal anti-inflammatory drugs can be used, although their
capacity to inhibit the development of the lesions of EHM is unknown. Recovery of
non-recumbent animals is usually complete, although improvement is gradual and
several months may elapse before maximal improvement (Ostlund, 1993). However,
when a horse is recumbent for more then 24 hours, the prognosis is grave and
euthanasia may be indicated (van Maanen et al., 2001).
3.3 Diagnosis
Virus culture and isolation are considered the gold standard test for the laboratory
diagnosis of EHV1 (Lunn et al., 2009). Virus culture, isolation and identification of
EHV1 is strongly supportive of a diagnosis of EHV1-induced disease in a horse with
compatible clinical signs. The choice of sample depends on the disease syndrome
encountered (Ostlund, 1993). Both nasal and nasopharyngeal swabs can be collected
during the first phase of infection or in case of respiratory symptoms. A recent study
showed that nasal swabbing represents a viable alternative to the less well tolerated
nasopharyngeal swabbing technique for the molecular detection of EHV1 in horses
(Pusterla et al., 2008a). Heparinized whole blood can also be collected at this stage, as
viremia, associated with the buffy coat fraction, often coincides with clinical signs of
respiratory disease, abortion and neurologic disease. In case of abortion, virus can be
isolated from different foetal tissues. Viral culture and positive identification can be
accomplished in as little as 2-3 days in a laboratory when the sample contains a high
viral load. However, virus isolation is often unsuccessful during outbreaks, because
the peak of virus shedding may already have passed and local antibodies may interfere
(Mumford, 1984; van Maanen et al., 2001). The likelihood of detecting EHV1 during
outbreaks of disease can be increased by testing in-contact horses, especially during
episodes of fever.
Recently, PCR techniques have become more and more common as a diagnostic
test because of their high analytical sensitivity and specificity in case of low viral load
(Sharma et al., 1992; van Maanen et al., 2001; Varrasso et al., 2001). Conventional
PCR, however, is limited in its sensitivity, because no distinction between the

Review of the literature
36
presence of infectious virus and latent infection can be made. The random testing or
screening of healthy horses for EHV1 by conventional PCR should therefore be
avoided (Lunn et al., 2009). The use of novel EHV1 PCR platforms, such as real time
PCR, allows for a more sensitive detection, a greater specificity and calculation of
viral loads (Elia et al., 2006; Hussey et al., 2006). Determination of viral load can
offer important advantages as it can allow for better characterization of disease stage,
assessment of risk of exposure to other horses and monitoring of response to treatment
(Pusterla et al., 2009b; Pusterla et al., 2008b; Pusterla et al., 2009c). Concerning the
circulation of neurovirulent and non-neurovirulent strains of EHV1 in the field, real-
time PCR tests have been developed that distinguish between these two biovars
(Allen, 2007; Pusterla et al., 2009c).
Serologic testing demonstrating a 4-fold or greater increase in serum antibody titre
by serum-neutralizing (SN) or complement-fixation tests on acute and convalescent
samples collected 3 weeks apart, provides presumptive evidence of EHV1 infection
(Kydd et al., 2006). Although serologic testing has limitations in confirming a
diagnosis of EHV1 infection in an individual horse, testing of paired serum samples
from in-contact horses is recommended. However, serologic testing is limited due to
cross-reactivity between EHV1 and EHV4, and most laboratories cannot distinguish
these two viruses (Hartley et al., 2005). Therefore, a specific ELISA test based on the
C-terminal portion of glycoprotein G of both viruses has been developed (Drummer et
al., 1995).
Cerebrospinal fluid analysis with the presence of xanthochromia, a yellowish
appearance of cerebrospinal fluid, is often supportive of an EHM diagnosis. In
addition, increased protein concentrations with or without a monocytic pleocytosis are
typical findings. Histopathology on brain and spinal cord, with typical vasculitis and
thrombosis of small blood vessels, is an essential method for confirming EHV1
infection in a horse with suspected EHM post mortem (Schultheiss et al., 1997).
3.4 Disease control
Vaccination. EHV1 remains a leading cause of upper respiratory tract infection,
viral abortion and nervous system disorders in horses worldwide, despite the
customary use of commercial vaccines (Allen et al., 1999; Kydd et al., 2006).

Review of the literature
37
Following natural infection, EHV1 efficiently induces a local mucosal humoral
response, which offers the potential for immune exclusion of virus at the respiratory
tract epithelium portal of entry. However, neither inactivated, nor attenuated vaccines
are able to induce detectable mucosal antibodies (Breathnach et al., 2001). The
currently available vaccines are not able to induce a significant level of protection
against EHV1-induced viremia. However, several vaccines are able to induce a
significant level of protection against either abortion or nervous system disorders (van
der Meulen et al., 2007).
The purpose of vaccination is twofold. First, vaccination is meant to minimize
virus replication in the respiratory tract upon infection, thus limiting nasal shedding
and the occurrence of respiratory disorders. Second, vaccination should prevent the
occurrence of abortion and/or nervous system disorders. The first purpose of
vaccination seems to be fulfilled by the available vaccines, as demonstrated by
reduced nasal virus titres and the reduced severity of respiratory disease upon
challenge infection of vaccinated horses (Burrows et al., 1984; Goodman et al., 2006;
Heldens et al., 2001). However, the second purpose to protect horses against EHV1-
induced viremia and subsequent abortion or neurological disorders is highly variable
and none of them can guarantee 100% protection. In view of this, we will briefly
review several vaccination/challenge studies performed with commercially available
vaccines.
Many studies have addressed the potential of inactivated vaccines in the battle
against EHV1. Inactivated virus vaccines can stimulate high titres of serum virus
neutralizing (VN) antibody, which cannot only reduce the amount and duration of
virus shedding but also prime the mucosal compartment (Breathnach et al., 2001).
However, they fail to induce cytotoxic T-leukocyte (CTL) responses which have
proven to be of paramount importance to limit EHV1 infection (Kydd et al., 2003;
Minke et al., 2004). Several case-control studies have been performed with the
commercially available vaccine Duvaxyn EHV1,4® (Foote et al., 2002; Heldens et al.,
2001). This inactivated vaccine clearly reduced clinical symptoms, the duration of
virus shedding and the quantity of virus shed. Additionally, a significant reduction in
the occurrence of abortion was noticed. However, vaccination under field conditions
could not prevent continuous circulation of both EHV1 and EHV4 in vaccinated
populations (Foote et al., 2006). Two other studies were performed with another

Review of the literature
38
inactivated and oil-adjuvanted vaccine Pneumabort K®. Neither of these studies could
demonstrate a difference in febrile responses, clinical signs and subsequent abortion
rates between vaccinated and control mares (Bürki et al., 1990; Burrows et al., 1984).
However, a recent study with the latter vaccine did not only show a significant
reduction of clinical signs and nasal shedding after three vaccinations, but also
demonstrated a significant reduction in the number of days of viremia (Goehring et
al., 2010c). A possible influence on abortion or nervous system disorders was not
evaluated during this study.
Modified-live vaccines (MLV) probably stimulate CTL responses and prime the
animals for mucosal antibodies, making them valuable candidates to limit EHV1
infections. They induce a rapid onset of immunity which is both broad and long
lasting. However, care should be taken when using these vaccines as reversion to
virulence remains a possible risk (Minke et al., 2004). A study of Bürki et al. (1990)
showed that all horses became infected and developed viremia upon challenge after
vaccination with the MLV Prevaccinol® (Bürki et al., 1990). Two out of 4 mares
aborted, but the study lacked a control group of non-vaccinated mares. Another study
compared a commercially available inactivated vaccine (Flu-vac Innovator 6®) and a
commercially available modified-live vaccine (Rhinomune®) (Goodman et al., 2006).
The modified-live vaccine induced significantly lower VN antibodies, suggesting a
bias towards a cytotoxic immune response and virus shedding was significantly lower.
Although the lymphocyte viral gene copies were similar, viremia lasted for a shorter
period after vaccination with Rhinomune®. Upon challenge, none of the horses
showed nervous system disorders, in contrast to the horses vaccinated with Flu-vac
Innovator 6® (3/5) and the control group (3/5). Also a recent study showed a great
decrease in clinical symptoms and nasal shedding after vaccination with Rhinomune®.
However a decrease in duration of viremia could not be noticed (Goehring et al.,
2010c).
Other alternative approaches for potential vaccination against EHV1-induced
disease such as DNA vaccines, live-vectored vaccines and gene-deleted mutants are
not yet commercially available, but have undergone pilot studies. A study of Soboll et
al. (2006) could demonstrate a limited immune response and protection following
DNA vaccination with plasmids encoding gB, gC, gD, IE and early proteins of EHV1
(Soboll et al., 2006). A recent study showed that a vaccine with the IE gene of EHV1

Review of the literature
39
in a recombinant modified-live vaccinia vector provides not only a reduction in
clinical disease, but also a reduction of cell-associated viremia (Soboll et al., 2010).
Unfortunately, until now, the currently available vaccines do not provide complete
protection as they especially stimulate high titres of circulating antibodies and are
unlikely to stimulate cytotoxic effector lymphocytes. As EHV1-induced protective
immunity is only short-lived, vaccination has to be repeated every 6 months (Kydd et
al., 2006). Future vaccination strategies should be aimed at stimulating both CD8+ and
CD4+ elements of cell-mediated immunity, in particular CTLs, but at the same time
maximise stimulation of mucosal and plasma VN antibody (Kydd et al., 2006).
Routine management. The overall goals in management of EHV1 infections are
to minimize exposure of horses to exogenous virus, to maximize immune
preparedness to forestall disease in the event of exposure, and to decrease the
likelyhood of latent virus recrudescence under influence of stress (Ostlund, 1993; van
der Meulen et al., 2007). Management has to be concentrated mainly on prevention of
the more serious sequelae of an EHV1 infection like abortion storms and outbreaks of
neurological disease (van Maanen, 2002).
Retrospective analysis of equine herpesviral disease outbreaks often can be linked
to a source of exogenous virus. Age segregation of horses on a farm is one of the best
tactics to limit spread of herpesviruses. In particular, pregnant mares should be
separated from young stock and transient horses. Ideally, the pregnant mare
population should be routinely subdivided into small groups according to expected
delivery dates (Ostlund, 1993). Personnel should handle pregnant mares before
contacting other horses on the premises. New horses should be isolated for a period of
3 weeks, with daily monitoring of rectal temperature and general health, before
mixing with the resident population (Allen & Bryans, 1986; Ostlund, 1993).
Vaccination programs should be instituted on a farm-wide basis and should
include adult horses and non-breeding animals as well as young stock and pregnant
mares (Ostlund, 1993) (see chapter 3.4 Disease control, vaccination).
Although some stress is unavoidable, efforts can be made to avoid unnecessary
stress and to be diligent in routine disease surveillance among animals known to be
recently subjected to stress (Ostlund, 1993). Stress factors such as poor feeding

Review of the literature
40
conditions, overcrowding, travelling in late pregnancy and separation from mates
should be minimized (van Maanen, 2002).
Management in case of an outbreak. The three priorities for management in case
of an outbreak of EHV1 are an early diagnosis, prevention of further spread and
thorough management of clinical cases (Allen, 2002; Pusterla et al., 2009a).
The importance of an early diagnosis of EHV1 is crucial, as several specific
interventions may be implemented in case of an EHV1-induced outbreak. Measures
designed to contain potential EHV1 spread are necessary until EHV1 is either
confirmed or excluded (Lunn et al., 2009). A good understanding of the appropriate
diagnostical methods and understanding what samples to take is vital for outbreak
management (see chapter 3.3 Diagnosis).
To prevent further spread of virus to other horses or facilities, quarantine
measurements of the whole facility are necessary for at least 4 weeks after the last
clinical case. A recent paper even describes the necessity of isolating actively EHV1-
shedding horses in a separate airspace under strict biosecurity and isolation
procedures (Goehring et al., 2010b). All affected animals should be kept isolated and
no horses should be allowed to enter or leave the property until quarantine is lifted
(Allen & Bryans, 1986; Pusterla et al., 2009a; van Maanen, 2002). Areas
contaminated by virus should be carefully cleaned with detergents. In the face of an
EHV1 outbreak, vaccination can be used for horses at increased risk of exposure and
to reduce the spread of infectious virus. There is some controversy associated with
this practice, because of the concern that EHM might be associated with a history of
frequent vaccination (Henninger et al., 2007). However, the latter statement should be
interpreted with care, as the majority of horses that was vaccinated 3-4 times a year
were older than 5 years. The latter has been described as a risk factor for the
development of neurologic disease (Goehring et al., 2006).
Clinical cases should be handled as described in chapter 3.2.3 Treatment.

Review of the literature
41
References
Allen, G. P. (2002). Epidemic disease caused by equine herpesvirus-1: Recommendations
for prevention and control. Equine Vet Educ 4, 177-183.
Allen, G. P. (2007). Development of a real-time polymerase chain reaction assay for
rapid diagnosis of neuropathogenic strains of equine herpesvirus-1. J Vet Diagn
Invest 19, 69-72.
Allen, G. P., Bolin, D. C., Bryant, U., Carter, C. N., Giles, R. C., Harrison, L. R.,
Hong, C. B., Jackson, C. B., Poonacha, K., Wharton, R. & Williams, N. M. (2008). Prevalence of latent, neuropathogenic equine herpesvirus-1 in the
Thoroughbred broodmare population of central Kentucky. Equine Vet J 40, 105-
110.
Allen, G. P. & Breathnach, C. C. (2006). Quantification by real-time PCR of the
magnitude and duration of leucocyte-associated viraemia in horses infected with
neuropathogenic vs. non-neuropathogenic strains of EHV-1. Equine Vet J 38, 252-
257.
Allen, G. P. & Bryans, J. T. (1986). Molecular epizootiology, pathogenesis, and
prophylaxis of equine herpesvirus-1 infections. Prog Vet Microbiol Immunol 2,
78-144.
Allen, G. P., Kydd, J. H., Slater, J. D. & Smith, K. C. (1999). Recent advances in
understanding the pathogenesis, epidemiology and immunological control of
equid herpesvirus-1 (EHV-1) abortion. Proceedings of the Eight International
Conference on Equine Infectious Diseases, 129-146.
Baxi, M. K., Efstathiou, S., Lawrence, G., Whalley, J. M., Slater, J. D. & Field, H. J. (1995). The detection of latency-associated transcripts of equine herpesvirus 1 in
ganglionic neurons. J Gen Virol 76, 3113-3118.
Blanchard, T. L., Kenney, R. M. & Timoney, P. J. (1992). Venereal disease. Vet Clin
North Am Equine Pract 8, 191-203.
Borchers, K., Thein, P. & Sterner-Kock, A. (2006). Pathogenesis of equine herpes-
associated neurological disease: a revised explanation. Equine Vet J 38, 283-287.
Borchers, K., Wolfinger, U., Goltz, M., Broll, H. & Ludwig, H. (1997). Distribution
and relevance of equine herpesvirus type 2 (EHV-2) infections. Arch Virol 142,
917-928.
Borchers, K., Wolfinger, U., Ludwig, H., Thein, P., Baxi, S., Field, H. J. & Slater, J. D. (1998). Virological and molecular biological investigations into equine herpes
virus type 2 (EHV-2) experimental infections. Virus Res 55, 101-106.
Breathnach, C. C., Yeargan, M. R., Sheoran, A. S. & Allen, G. P. (2001). The
mucosal humoral immune response of the horse to infective challenge and
vaccination with Equine herpesvirus-1 antigens. Equine Vet J 33, 651-657.
Brosnahan, M. M., Damiani, A., van de Walle, G., Erb, H., Perkins, G. A. & Osterrieder, N. (2009). The effect of siRNA treatment on experimental equine
herpesvirus type 1 (EHV-1) infection in horses. Virus research,
doi:10.1016/j.virusres.2099.1010.1017.
Brown, J. A., Mapes, S., Ball, B. A., Hodder, A. D., Liu, I. K. & Pusterla, N. (2007). Prevalence of equine herpesvirus-1 infection among Thoroughbreds residing on a
farm on which the virus was endemic. J Am Vet Med Assoc 231, 577-580.
Browning, G. F., Ficorilli, N. & Studdert, M. J. (1988). Asinine herpesvirus genomes:
comparison with those of the equine herpesviruses. Arch Virol 101, 183-190.

Review of the literature
42
Bryant, N. A., Davis-Poynter, N., Vanderplasschen, A. & Alcami, A. (2003). Glycoprotein G isoforms from some alphaherpesviruses function as broad-
spectrum chemokine binding proteins. EMBO J 22, 833-846.
Bürki, F., Rossmanith, W., Nowotny, N., Pallan, C., Mostl, K. & Lussy, H. (1990). Viremia and abortions are not prevented by two commercial equine herpesvirus-1
vaccines after experimental challenge of horses. Veterinary Quaterly 12, 80-86.
Burrows, R. & Goodridge, D. (1972). In vivo and in vitro studies of equine
rhinopneumonitis strains. Proc 3rd Int Conf of Equine Infect Dis, Paris, 306-321.
Burrows, R., Goodridge, D. & Denyer, M. S. (1984). Trials of an inactivated equid
herpesvirus 1 vaccine: challenge with a subtype 1 virus. Vet Rec 114, 369-374.
Carroll, C. L. & Westbury, H. A. (1985). Isolation of equine herpesvirus 1 from the
brain of a horse affected with paresis. Australian Veterinary Journal 62, 345-346.
Carvalho, R., Oliveira, A. M., Souza, A. M., Passos, L. M. & Martins, A. S. (2000). Prevalence of equine herpesvirus type 1 latency detected by polymerase chain
reaction. Arch Virol 145, 1773-1787.
Caughman, G. B., Staczek, J. & O'Callaghan, D. J. (1985). Equine herpesvirus type 1
infected cell polypeptides: evidence for immediate early/early/late regulation of
viral gene expression. Virology 145, 49-61.
Chesters, P. M., Allsop, R., Purewal, A. & Edington, N. (1997). Detection of latency-
associated transcripts of equid herpesvirus 1 in equine leukocytes but not in
trigeminal ganglia. J Virol 71, 3437-3443.
Coggins, L. (1979). Viral respiratory disease of horses. Vet Clin North Am, large Anim
Pract I, 59-72.
Crabb, B. S. & Studdert, M. J. (1990). Comparative studies of the proteins of equine
herpesviruses 4 and 1 and asinine herpesvirus 3: antibody response of the natural
hosts. J Gen Virol 71 ( Pt 9), 2033-2041.
Crabb, B. S. & Studdert, M. J. (1993). Epitopes of glycoprotein G of equine
herpesviruses 4 and 1 located near the C termini elicit type-specific antibody
responses in the natural host. J Virol 67, 6332-6338.
Crabb, B. S. & Studdert, M. J. (1995). Equine herpesvirus 4 (equine rhinopneumonitis
virus) and 1 (equine abortion virus). Advances in Virus Research 45, 153-190.
Csellner, H., Walker, C., Wellington, J. E., McLure, L. E., Love, D. N. & Whalley, J. M. (2000). EHV-1 glycoprotein D (EHV-1 gD) is required for virus entry and
cell-cell fusion, and an EHV-1 gD deletion mutant induces a protective immune
response in mice. Arch Virol 145, 2371-2385.
Davison, A. J., Eberle, R., Ehlers, B., Hayward, G. S., McGeoch, D. J., Minson, A. C., Pellett, P. E., Roizman, B., Studdert, M. J. & Thiry, E. (2009). The order
Herpesvirales. Arch Virol 154, 171-177.
Del Piero, F. & Wilkins, P. A. (2001). Pulmonary vasculotropic EHV-1 infection in
equids. Veterinary Pathology 38, 474.
Del Piero, F., Wilkins, P. A., Timoney, P. J., Kadushin, J., Vogelbacker, H., Lee, J. W., Berkowitz, S. J. & La Perle, K. M. D. (2000). Fatal nonneurological EHV-1
infection in a yearling filly. Veterinary Pathology 37, 672-676.
Dimock, W. W. & Edwards, P. R. (1933). Is there a filterable virus of abortion in
mares? Kentucky Agricultural Experiment Station Bulletin 333, 297-301.
Doll, E. R. (1952). Seasonal incidence and fetal age in equine viral abortion. Cornell
Veterinarian 42, 505-509.
Doll, E. R. & Bryans, J. T. (1963). Epizootiology of equine viral rhinopneumonitis. J
Am Vet Med Assoc 142, 31-37.

Review of the literature
43
Doll, E. R., Crowe, M. E. W., Bryans, J. T. & McCollum, W. H. (1955). Infection
immunity in equine virus abortion. Cornell Veterinarian 45, 387-410.
Drummer, H. E., Reynolds, A., Studdert, M. J., MacPherson, C. M. & Crabb, B. S. (1995). Application of an equine herpesvirus 1 (EHV1) type-specific ELISA to
the management of an outbreak of EHV1 abortion. Vet Rec 136, 579-581.
Edington, N., Bridges, C. G. & Huckle, A. (1985). Experimental reactivation of equid
herpesvirus 1 (EHV1) following the administration of corticosteroids. Equine Vet
J 17, 369-372.
Edington, N., Bridges, C. G. & Patel, J. R. (1986). Endothelial cell infection and
thrombosis in paralysis caused by equid herpesvirus-1: equine stroke. Arch Virol
90, 111-124.
Edington, N., Smith, B. & Griffiths, L. (1991). The role of endothelial cell infection in
the endometrium, placenta and foetus of equid herpesvirus 1 (EHV-1) abortions.
Journal of Comparative Pathology 104, 379-387.
Edington, N., Welch, H. M. & Griffiths, L. (1994). The prevalence of latent Equid
herpesviruses in the tissues of 40 abattoir horses. Equine Vet J 26, 140-142.
Elia, G., Decaro, N., Martella, V., Campolo, M., Desario, C., Lorusso, E., Cirone, F. & Buonavoglia, C. (2006). Detection of equine herpesvirus type 1 by real time
PCR. J Virol Methods 133, 70-75.
Ficorilli, N., Studdert, M. J. & Crabb, B. S. (1995). The nucleotide sequence of asinine
herpesvirus 3 glycoprotein G indicates that the donkey virus is closely related to
equine herpesvirus 1. Arch Virol 140, 1653-1662.
Foote, C. E., Love, D. N., Gilkerson, J. R., Wellington, J. E. & Whalley, J. M. (2006). EHV-1 and EHV-4 infection in vaccinated mares and their foals. Vet Immunol
Immunopathol 111, 41-46.
Foote, C. E., Love, D. N., Gilkerson, J. R. & Whalley, J. M. (2002). Serological
responses of mares and weanlings following vaccination with an inactivated
whole virus equine herpesvirus 1 and equine herpesvirus 4 vaccine. Vet Microbiol
88, 13-25.
Frampton, A. R., Jr., Goins, W. F., Cohen, J. B., von Einem, J., Osterrieder, N., O'Callaghan, D. J. & Glorioso, J. C. (2005). Equine herpesvirus 1 utilizes a
novel herpesvirus entry receptor. J Virol 79, 3169-3173.
Frampton, A. R., Jr., Stolz, D. B., Uchida, H., Goins, W. F., Cohen, J. B. & Glorioso, J. C. (2007). Equine herpesvirus 1 enters cells by two different pathways, and
infection requires the activation of the cellular kinase ROCK1. J Virol 81, 10879-
10889.
Frampton, A. R., Jr., Uchida, H., von Einem, J., Goins, W. F., Grandi, P., Cohen, J. B., Osterrieder, N. & Glorioso, J. C. (2010). Equine herpesvirus type 1 (EHV-1)
utilizes microtubules, dynein, and ROCK1 to productively infect cells. Vet
Microbiol 141, 12-21.
Friday, P. A., Scarratt, W. K., Elvinger, F., Timoney, P. J. & Bonda, A. (2000). Ataxia and paresis with equine herpesvirus type 1 infection in a herd of riding
school horses. J Vet Intern Med 14, 197-201.
Fritsche, A. K. & Borchers, K. (2010). Detection of neuropathogenic strains of Equid
Herpesvirus 1 (EHV-1) associated with abortions in Germany. Vet Microbiol 147,
176-180.
Fukushi, H., Tomita, T., Taniguchi, A., Ochiai, Y., Kirisawa, R., Matsumura, T., Yanai, T., Masegi, T., Yamaguchi, T. & Hirai, K. (1997). Gazelle herpesvirus

Review of the literature
44
1: a new neurotropic herpesvirus immunologically related to equine herpesvirus 1.
Virology 227, 34-44.
Fulton, A., Peters, S. T., Perkins, G. A., Jarosinski, K. W., Damiani, A., Brosnahan, M., Buckles, E. L., Osterrieder, N. & Van de Walle, G. R. (2009). Effective
treatment of respiratory alphaherpesvirus infection using RNA interference. PLoS
One 4, e4118.
Furlong, D., Swift, H. & Roizman, B. (1972). Arrangement of herpesvirus
deoxyribonucleic acid in the core. J Virol 10, 1071-1074.
Garré, B., Gryspeerdt, A., Croubels, S., De Backer, P. & Nauwynck, H. (2009). Evaluation of orally administered valacyclovir in experimentally EHV1-infected
ponies. Vet Microbiol 135, 214-221.
Garré, B., Shebany, K., Gryspeerdt, A., Baert, K., van der Meulen, K., Nauwynck, H., Deprez, P., De Backer, P. & Croubels, S. (2007). Pharmacokinetics of
acyclovir after intravenous infusion of acyclovir and after oral administration of
acyclovir and its prodrug valacyclovir in healthy adult horses. Antimicrob Agents
Chemother 51, 4308-4314.
Gibson, J. S., Slater, J. D., Awan, A. R. & Field, H. J. (1992). Pathogenesis of equine
herpesvirus-1 in specific pathogen-free foals: primary and secondary infections
and reactivation. Arch Virol 123, 351-366.
Gilkerson, J., Love, D. & Whalley, J. M. (1997). Serological evidence of equine
herpesvirus (EHV-1) infection in Thoroughbred foals 30-120 days of age.
Australian Equine Veterinarian 15, 128-134.
Gilkerson, J. R., Whalley, J. M., Drummer, H. E., Studdert, M. J. & Love, D. N. (1999). Epidemiology of EHV-1 and EHV-4 in the mare and foal populations on a
Hunter Valley stud farm: are mares the source of EHV-1 for unweaned foals. Vet
Microbiol 68, 27-34.
Gleeson, L. J. & Coggins, L. (1980). Response of pregnant mares to equine herpesvirus
1 (EHV1). Cornell Veterinarian 70, 391-400.
Goehring, L. S., Hussey, G. S., Ashton, L. V., Schenkel, A. R. & Lunn, D. P. (2010a). Infection of central nervous system endothelial cells by cell-associated EHV-1.
Vet Microbiol doi:10.1016/j.vetmic.2010.08.030.
Goehring, L. S., Landolt, G. A. & Morley, P. S. (2010b). Detection and management of
an outbreak of equine herpesvirus type 1 infection and associated neurological
disease in a veterinary teaching hospital. J Vet Intern Med 24, 1176-1183.
Goehring, L. S. & Sloet van Oldruitenborgh-Oosterbaan, M. M. (2001). The mystery
of equine herpes myeloencephalopathy. Equine Veterinary Education 13, 36-42.
Goehring, L. S., van Winden, S. C., van Maanen, C. & Sloet van Oldruitenborgh-Oosterbaan, M. M. (2006). Equine herpesvirus type 1-associated
myeloencephalopathy in The Netherlands: a four-year retrospective study (1999-
2003). J Vet Intern Med 20, 601-607.
Goehring, L. S., Wagner, B., Bigbie, R., Hussey, S. B., Rao, S., Morley, P. S. & Lunn, D. P. (2010c). Control of EHV-1 viremia and nasal shedding by commercial
vaccines. Vaccine 28, 5203-5211.
Goodman, L. B., Loregian, A., Perkins, G. A., Nugent, J., Buckles, E. L., Mercorelli,
B., Kydd, J. H., Palu, G., Smith, K. C., Osterrieder, N. & Davis-Poynter, N. (2007). A point mutation in a herpesvirus polymerase determines
neuropathogenicity. PLoS Pathog 3, 1583-1592.
Goodman, L. B., Wagner, B., Flaminio, M. J., Sussman, K. H., Metzger, S. M., Holland, R. & Osterrieder, N. (2006). Comparison of the efficacy of inactivated

Review of the literature
45
combination and modified-live virus vaccines against challenge infection with
neuropathogenic equine herpesvirus type 1 (EHV-1). Vaccine 24, 3636-3645.
Goulden, B. C., Wilks, C. & Jones, D. (1989). Equine herpesvirus (EHV1) disease.
Publication of the New-Zealand Equine Research Foundation, 1-32.
Granzow, H., Klupp, B. G., Fuchs, W., Veits, J., Osterrieder, N. & Mettenleiter, T. C. (2001). Egress of alphaherpesviruses: comparative ultrastructural study. J Virol
75, 3675-3684.
Gray, W. L., Baumann, R. P., Robertson, A. T., Caughman, G. B., O'Callaghan, D. J. & Staczek, J. (1987a). Regulation of equine herpesvirus type 1 gene
expression: characterization of immediate early, early, and late transcription.
Virology 158, 79-87.
Gray, W. L., Baumann, R. P., Robertson, A. T., O'Callaghan, D. J. & Staczek, J. (1987b). Characterization and mapping of equine herpesvirus type 1 immediate
early, early, and late transcripts. Virus Res 8, 233-244.
Greenwood, R. E. & Simson, A. R. (1980). Clinical report of a paralytic syndrome
affecting stallions, mares and foals on a thoroughbred studfarm. Equine Vet J 12,
113-117.
Hartley, C. A., Drummer, H. E. & Studdert, M. J. (1999). Gazelle herpesvirus 1
(GHV-1): Closely related to EHV-1 and AHV-3. The nucleotide sequence of the
glycoprotein G homologue of equine herpesvirus 3 (EHV-3) indicates EHV-3 is a
distinct equid hperesvirus. Brief report. Arch Virol 144, 2023-2033.
Hartley, C. A., Wilks, C. R., Studdert, M. J. & Gilkerson, J. R. (2005). Comparison of
antibody detection assays for the diagnosis of equine herpesvirus 1 and 4
infections in horses. Am J Vet Res 66, 921-928.
Heldens, J. G., Hannant, D., Cullinane, A. A., Prendergast, M. J., Mumford, J. A.,
Nelly, M., Kydd, J. H., Weststrate, M. W. & van den Hoven, R. (2001). Clinical and virological evaluation of the efficacy of an inactivated EHV1 and
EHV4 whole virus vaccine (Duvaxyn EHV1,4). Vaccination/challenge
experiments in foals and pregnant mares. Vaccine 19, 4307-4317.
Henninger, R. W., Reed, S. M., Saville, W. J., Allen, G. P., Hass, G. F., Kohn, C. W. & Sofaly, C. (2007). Outbreak of Neurologic disease Caused by Equine
Herpesvirus-1 at a University Equestrian Center. J Vet Intern Med 21, 157-165.
Huemer, H. P., Nowotny, N., Crabb, B. S., Meyer, H. & Hubert, P. H. (1995). gp13
(EHV-gC): a complement receptor induced by equine herpesviruses. Virus
Research 37, 113-126.
Hussey, S. B., Clark, R., Lunn, K. F., Breathnach, C., Soboll, G., Whalley, J. M. & Lunn, D. P. (2006). Detection and quantification of equine herpesvirus-1 viremia
and nasal shedding by real-time polymerase chain reaction. J Vet Diagn Invest 18,
335-342.
Jackson, T. A. & Kendrick, J. W. (1971). Paralysis of horses associated with equine
herpesvirus-1 infection. J Am Vet Med Assoc 159, 1351-1357.
Jackson, T. A., Osburn, B. I., Cordy, D. R. & Kendrick, J. W. (1977). Equine
herpesvirus 1 infection of horses: studies on the experimentally induced
neurologic disease. Am J Vet Res 38, 709-719.
Kershaw, O., von Oppen, T., Glitz, F., Deegen, E., Ludwig, H. & Borchers, K. (2001). Detection of equine herpesvirus type 2 (EHV-2) in horses with
keratoconjunctivitis. Virus Res 80, 93-99.
Kleiboeker, S. B., Schommer, S. K., Johnson, P. J., Ehlers, B., Turnquist, S. E., Boucher, M. & Kreeger, J. M. (2002). Association of two newly recognized

Review of the literature
46
herpesviruses with interstitial pneumonia in donkeys (Equus asinus). J Vet Diagn
Invest 14, 273-280.
Kurtz, B. M., Singletary, L. B., Kelly, S. D. & Frampton, A. R., Jr. (2010). Equus
caballus major histocompatibility complex class I is an entry receptor for equine
herpesvirus type 1. J Virol 84, 9027-9034.
Kydd, J. H., Smith, K. C., Hannant, D., Livesay, G. J. & Mumford, J. A. (1994a). Distribution of equid herpesvirus-1 (EHV-1) in respiratory tract associated
lymphoid tissue: implications for cellular immunity. Equine Vet J 26, 470-473.
Kydd, J. H., Smith, K. C., Hannant, D., Livesay, G. J. & Mumford, J. A. (1994b). Distribution of equid herpesvirus-1 (EHV-1) in the respiratory tract of ponies:
implications for vaccination strategies. Equine Vet J 26, 466-469.
Kydd, J. H., Townsend, H. G. & Hannant, D. (2006). The equine immune response to
equine herpesvirus-1: the virus and its vaccines. Vet Immunol Immunopathol 111,
15-30.
Kydd, J. H., Wattrang, E. & Hannant, D. (2003). Pre-infection frequencies of equine
herpesvirus-1 specific, cytotoxic T lymphocytes correlate with protection against
abortion following experimental infection of pregnant mares. Vet Immunol
Immunopathol 96, 207-217.
Lunn, D. P., Davis-Pointer, N., Flaminio, M. J., Horohov, D. W., Osterrieder, K., Pusterla, N. & Townsend, H. G. (2009). Equine Herpesvirus-1 Consensus
Statement. J Vet Intern Med 23, 450-461.
Marenzoni, M. L., Passamonti, F., Cappelli, K., Veronesi, F., Capomaccio, S., Supplizi, A. V., Valente, C., Autorino, G. & Coletti, M. (2008). Clinical,
serological and molecular investigations of EHV-1 and EHV-4 in 15 unweaned
thoroughbred foals. Vet Rec 162, 337-341.
Matsumura, T., Kondo, T., Sugita, S., Damiani, A. M., O'Callaghan, D. J. & Imagawa, H. (1998). An equine herpesvirus type 1 recombinant with a deletion in
the gE and gI genes is avirulent in young horses. Virology 242, 68-79.
Maxwell, L. K., Bentz, B. G., Bourne, D. W. & Erkert, R. S. (2008a). Pharmacokinetics of valacyclovir in the adult horse. J Vet Pharmacol Ther 31,
312-320.
Maxwell, L. K., Bentz, B. G., Gilliam, L. L., Ritchey, J. W., Holbrook, T. C.,
McFarlane, D., Rezabek, G. B., MacAllister, C. G. & Allen, G. P. (2008b). Efficacy of Valacyclovir Against Clinical Disease After EHV-1 Challenge in
Aged Mares. Proceedings of the Annual Convention of the AAEP 54, 198-199.
McCartan, C. G., Russell, M. M., Wood, J. L. & Mumford, J. A. (1995). Clinical,
serological and virological characteristics of an outbreak of paresis and neonatal
foal disease due to equine herpesvirus-1 on a stud farm. Vet Rec 136, 7-12.
Mettenleiter, T. C. (2002). Herpesvirus assembly and egress. J Virol 76, 1537-1547.
Mettenleiter, T. C. (2004). Budding events in herpesvirus morphogenesis. Virus Res 106,
167-180.
Mettenleiter, T. C., Klupp, B. G. & Granzow, H. (2006). Herpesvirus assembly: a tale
of two membranes. Curr Opin Microbiol 9, 423-429.
Mettenleiter, T. C., Klupp, B. G. & Granzow, H. (2009). Herpesvirus assembly: an
update. Virus Res 143, 222-234.
Minke, J. M., Audonnet, J. C. & Fischer, L. (2004). Equine viral vaccines: the past,
present and future. Vet Res 35, 425-443.
Mumford, J. A. (1984). The development of diagnostic techniques for equine viral
diseases. Veterinary Annals 24, 182-189.

Review of the literature
47
Mumford, J. A. & Edington, N. (1980). EHV1 and equine paresis. Vet Rec 106, 277.
Mumford, J. A., Hannant, D. A., Jesset, D. M., O'Neill, T., Smith, K. C. & Ostlund, E. N. (1994). Abortigenic and neurological disease caused by experimental
infection with equine herpesvirus 1. Proceedings of the 7th International
Conference of Equine Infectious diseases, Newmarket, UK, 261-275.
Mumford, J. A., Rossdale, P. D., Jessett, D. M., Gann, S. J., Ousey, J. & Cook, R. F. (1987). Serological and virological investigations of an equid herpesvirus 1 (EHV-
1) abortion storm on a stud farm in 1985. Journal of Reproduction and Fertility,
supplement 35, 509-518.
Murray, M. J., del Piero, F., Jeffrey, S. C., Davis, M. S., Furr, M. O., Dubovi, E. J. & Mayo, J. A. (1998). Neonatal Equine Herpesvirus Type 1 Infection on a
Thoroughbred Breeding Farm. J Vet Intern Med 12, 36-41.
Neubauer, A., Braun, B., Brandmuller, C., Kaaden, O. R. & Osterrieder, N. (1997). Analysis of the contributions of the equine herpesvirus 1 glycoprotein gB
homolog to virus entry and direct cell-to-cell spread. Virology 227, 281-294.
Neubauer, A. & Osterrieder, N. (2004). Equine herpesvirus type 1 (EHV-1)
glycoprotein K is required for efficient cell-to-cell spread and virus egress.
Virology 329, 18-32.
Nugent, J., Birch-Machin, I., Smith, K. C., Mumford, J. A., Swann, Z., Newton, J. R., Bowden, R. J., Allen, G. P. & Davis-Poynter, N. (2006). Analysis of equid
herpesvirus 1 strain variation reveals a point mutation of the DNA polymerase
strongly associated with neuropathogenic versus non-neuropathogenic disease
outbreaks. J Virol 80, 4047-4060.
Osterrieder, N. (1999). Construction and characterization of an equine herpesvirus 1
glycoprotein C negative mutant. Virus Research 59, 165-177.
Osterrieder, N., Neubauer, A., Brandmuller, C., Braun, B., Kaaden, O. R. & Baines, J. D. (1996). The equine herpesvirus 1 glycoprotein gp21/22a, the herpes simplex
virus type 1 gM homolog, is involved in virus penetration and cell-to-cell spread
of virions. J Virol 70, 4110-4115.
Ostlund, E. N. (1993). The equine herpesviruses. Vet Clin North Am Equine Pract 9,
283-294.
Pascoe, R. R., Spradbrow, P. B. & Bagust, T. J. (1968). Equine coital exanthema. Aust
Vet J 44, 485.
Patel, J. R., Edington, N. & Mumford, J. A. (1982). Variation in cellular tropism
between isolates of equine herpesvirus-1 in foals. Arch Virol 74, 41-51.
Perkins, G. A., Ainsworth, D. M., Erb, H. N., del Piero, F., Miller, M., Wilkins, P. A., Palmer, J. & Frazer, M. (1999). Clinical, haematological and biochemical
findings in foals with neonatal Equine herpesvirus-1 infection compared with
septic and premature foals. Equine Vet J 35, 422-426.
Perkins, G. A., Goodman, L. B., Tsujimura, K., Van de Walle, G. R., Kim, S. G., Dubovi, E. J. & Osterrieder, K. (2009). Investigation of the prevalence of
neurologic equine herpes virus type 1 (EHV-1) in a 23-year retrospective analysis
(1984-2007). Vet Microbiol 139, 375-378.
Platt, H., Singh, H. & Whitwell, K. E. (1980). Pathological observations on an outbreak
of paralysis in brood mares. Equine Vet J 12, 118-126.
Prickett, M. E. (1970). The pathology of disease caused by equine herpesvirus-1.
Proceedings of the 2nd International Conference on Equine Infectious diseases,
Paris, 24-33.

Review of the literature
48
Pronost, S., Leon, A., Legrand, L., Fortier, C., Miszczak, F., Freymuth, F. & Fortier, G. (2010). Neuropathogenic and non-neuropathogenic variants of equine
herpesvirus 1 in France. Vet Microbiol 145, 329-333.
Pusterla, N., David Wilson, W., Madigan, J. E. & Ferraro, G. L. (2009a). Equine
herpesvirus-1 myeloencephalopathy: a review of recent developments. Vet J 180,
279-289.
Pusterla, N., Hussey, S. B., Mapes, S., Johnson, C., Collier, J. R., Hill, J., Lunn, D. P. & Wilson, W. D. (2010). Molecular investigation of the viral kinetics of equine
herpesvirus-1 in blood and nasal secretions of horses after corticosteroid-induced
recrudescence of latent infection. J Vet Intern Med 24, 1153-1157.
Pusterla, N., Hussey, S. B., Mapes, S., Leutenegger, C. M., Madigan, J. E., Ferraro, G. L., Wilson, W. D. & Lunn, D. P. (2009b). Comparison of four methods to
quantify Equid herpesvirus 1 load by real-time polymerase chain reaction in nasal
secretions of experimentally and naturally infected horses. J Vet Diagn Invest 21,
836-840.
Pusterla, N., Mapes, S. & Wilson, W. D. (2008a). Diagnostic sensitivity of
nasopharyngeal and nasal swabs for the molecular detection of EHV-1. Vet Rec
162, 520-521.
Pusterla, N., Mapes, S. & Wilson, W. D. (2008b). Use of viral loads in blood and
nasopharyngeal secretions fot the diagnosis of EHV-1 infection in field cases. Vet
Rec 162, 728-729.
Pusterla, N., Wilson, W. D., Mapes, S., Finno, C., Isbell, D., Arthur, R. M. & Ferraro, G. L. (2009c). Characterization of viral loads, strain and state of equine
herpesvirus-1 using real-time PCR in horses following natural exposure at a
racetrack in California. Vet J 179, 230-239.
Reed, S. M. & Toribio, R. E. (2004). Equine herpesvirus 1 and 4. Vet Clin North Am
Equine Pract 20, 631-642.
Robertson, A. T., Caughman, G. B., Gray, W. L., Baumann, R. P., Staczek, J. & O'Callaghan, D. J. (1988). Analysis of the in vitro translation products of the
equine herpesvirus type 1 immediate early mRNA. Virology 166, 451-462.
Robertson, G. R., Scott, N. A., Miller, J. M., Sabine, M., Zheng, M., Bell, C. W. & Whalley, J. M. (1991). Sequence characteristics of a gene in equine herpesvirus 1
homologous to glycoprotein H of herpes simplex virus. DNA Seq 1, 241-249.
Roizman, B., Desrosiers, R. C., Fleckenstein, B., Lopez, C., Minson, A. C. & Studdert, M. J. (1992). The family Herpesviridae: an update. The Herpesvirus
Study Group of the International Committee on Taxonomy of Viruses. Arch Virol
123, 425-449.
Roizman, B. & Knipe, D. M., (eds) (2001). Herpes Simplex Viruses and Their
Replication. Philadelphia: Lipincott Williams and Wilkins.
Roizman, B. & Pellett, P. E., (eds) (2001). The Family Herpesviridae: A Brief
Introduction. Philadelphia: Lippincott Williams & Wilkins.
Rudolph, J. & Osterrieder, N. (2002). Equine Herpesvirus Type 1 Devoid of gM and
gp2 Is Severely Impaired in Virus Egress but Not Direct Cell-to-Cell Spread.
Virology 293, 356-367.
Rudolph, J., Seyboldt, C., Granzow, H. & Osterrieder, N. (2002). The gene 10
(UL49.5) product of equine herpesvirus 1 is necessary and sufficient for
functional processing of glycoprotein M. J Virol 76, 2952-2963.
Sabine, M., Robertson, G. R. & Whalley, J. M. (1981). Differentiation of sub-types of
equine herpesvirus I by restriction endonuclease analysis. Aust Vet J 57, 148-149.

Review of the literature
49
Sasaki, M., Hasebe, R., Makino, Y., Suzuki, T., Fukushi, H., Okamoto, M., Matsuda, K., Taniyama, H., Sawa, H. & Kimura, T. (2011). Equine major
histocompatibility complex class I molecules act as entry receptors that bind to
equine herpesvirus-1 glycoprotein D. Genes Cells 16, 343-357.
Saxegaard, F. (1966). Isolation and identification of equine rhinopneumonitis virus
(equine abortion virus) from cases of abortion and paralysis. Nordic Veterinary
Medicine 18, 504-510.
Schlocker, N., Gerber-Bretscher, R. & von Fellenberg, R. (1995). Equine herpesvirus
2 in pulmonary macrophages of horses. Am J Vet Res 56, 749-754.
Schultheiss, P. C., Collins, J. K. & Hotaling, S. F. (1997). Immunohistochemical
demonstration of equine herpesvirus-1 antigen in neurons and astrocytes of horses
with acute paralysis. Vet Pathol 34, 52-54.
Scott, J. C., Dutta, S. K. & Myrup, A. C. (1983). In vivo harboring of equine
herpesvirus-1 in leukocyte populations and subpopulations and their quantitation
from experimentally infected ponies. American Journal of Veterinary Research
44, 1344-1348.
Sharma, P. C., Cullinane, A. A., Onions, D. E. & Nicolson, L. (1992). Diagnosis of
equid herpesviruses -1 and -4 by polymerase chain reaction. Equine Vet J 24, 20-
25.
Shimizu, T., Ishizaki, R., Ishii, S., Kawakami, Y., Kaji, R., Sugimura, K. & Matumoto, M. (1959). Isolation of equine abortion virus from natural cases of
equine abortion in horse kidney cell culture. Jap J exp Med 29, 643-649.
Slater, J. D., Borchers, K., Thackray, A. M. & Field, H. J. (1994). The trigeminal
ganglion is a location for equine herpesvirus 1 latency and reactivation in the
horse. J Gen Virol 75, 2007-2016.
Smith, D. J., Hamblin, A. & Edington, N. (2002). Equid herpesvirus 1 infection of
endothelial cells requires activation of putative adhesion molecules: an in vitro
model. Clin Exp Immunol 129, 281-287.
Smith, D. J., Hamblin, A. S. & Edington, N. (2001). Infection of endothelial cells with
equine herpesvirus-1 (EHV-1) occurs where there is activation of putative
adhesion molecules: a mechanism for transfer of virus. Equine Vet J 33, 138-142.
Smith, K. C. (1997). Herpesviral Abortion in Domestic Animals. The Veterinary Journal
153, 253-268.
Smith, K. C., Whitwell, K. E., Binns, M. M., Dolby, C. A., Hannant, D. & Mumford, J. A. (1992). Abortion of virologically negative fetuses following experimental
challenge of pregnant mares with equid herpevirus 1. Equine Vet J 24, 256-259.
Smith, K. C., Whitwell, K. E., Mumford, J. A., Hannant, D., Blunden, A. S. & Tearle, J. P. (2000). Virulence of the V592 isolate of equid herpesvirus-1 in
ponies. Journal of Comparative Pathology 122, 288-297.
Smith, K. L., Allen, G. P., Branscum, A. J., Cook, R. F., Vickers, M. L., Timoney, P. J. & Balasuriya, U. B. R. (2010). The increased prevalence of neuropathogenic
strains of EHV-1 in equine abortions. Vet Microbiol 141, 5-11.
Smith, P. M., Kahan, S. M., Rorex, C. B., von Einem, J., Osterrieder, N. & O'Callaghan, D. J. (2005). Expression of the full-length form of gp2 of equine
herpesvirus 1 (EHV-1) completely restores respiratory virulence to the attenuated
EHV-1 strain KyA in CBA mice. J Virol 79, 5105-5115.
Soboll, G., Breathnach, C. C., Kydd, J. H., Hussey, S. B., Mealey, R. M. & Lunn, D. P. (2010). Vaccination of ponies with the IE gene of EHV-1 in a recombinant

Review of the literature
50
modified live vaccinia vector protects against clinical and virological disease. Vet
Immunol Immunopathol 135, 108-117.
Soboll, G., Hussey, S. B., Whalley, J. M., Allen, G. P., Koen, M. T., Santucci, N., Fraser, D. G., Macklin, M. D., Swain, W. F. & Lunn, D. P. (2006). Antibody
and cellular immune responses following DNA vaccination and EHV-1 infection
of ponies. Vet Immunol Immunopathol 111, 81-95.
Stierstorfer, B., Eichhorn, W., Schmahl, W., Brandmuller, C., Kaaden, O. R. & Neubauer, A. (2002). Equine herpesvirus type 1 (EHV-1) myeloencephalopathy:
a case report. J Vet Med B Infect Dis Vet Public Health 49, 37-41.
Stokes, A., Alber, D. G., Greensill, J., Amellal, B., Carvalho, R., Taylor, L. A., Doel, T. R., Killington, R. A., Halliburton, I. W. & Meredith, D. M. (1996). The
expression of the proteins of equine herpesvirus 1 which share homology with
herpes simplex virus 1 glycoproteins H and L. Virus Res 40, 91-107.
Studdert, M. J., Simpson, T. & Roizman, B. (1981). Differentiation of respiratory and
abortigenic isolates of equine herpesvirus 1 by restriction endonucleases. Science
214, 562-564.
Sun, Y., MacLean, A. R., Aitken, J. D. & Brown, S. M. (1996). The role of the gene 71
product in the life cycle of equine herpesvirus 1. J Gen Virol 77 ( Pt 3), 493-500.
Turtinen, L. W. & Allen, G. P. (1982). Identification of the envelope surface
glycoproteins of equine herpesvirus type 1. J Gen Virol 63, 481-485.
Turtinen, L. W., Allen, G. P., Darlington, R. W. & Bryans, J. T. (1981). Serological
and molecular comparisons of several equine herpesvirus type 1 strains. Am J vet
Res 42, 2099-2104.
USDA (2007). Equine herpesvirus myeloencephalopathy: A potentially emerging disease.
Available at: http://wwwaphisusdagov/vs/nahss/equine/ehv.
Van de Walle, G. R., Goupil, R., Wishon, C., Damiani, A., Perkins, G. A. & Osterrieder, N. (2009). A Single-Nucleotide Polymorphism in a Herpesvirus
DNA Polymerase Is Sufficient to Cause Lethal Neurological disease. J Infect Dis
200, 20-25.
Van de Walle, G. R., May, M. L., Sukhumavasi, W., von Einem, J. & Osterrieder, N. (2007). Herpesvirus chemokine-binding glycoprotein G (gG) efficiently inhibits
neutrophil chemotaxis in vitro and in vivo. J Immunol 179, 4161-4169.
Van de Walle, G. R., Peters, S. T., VanderVen, B. C., O'Callaghan, D. J. & Osterrieder, N. (2008a). Equine herpesvirus 1 entry via endocytosis is facilitated
by alphaV integrins and an RSD motif in glycoprotein D. J Virol 82, 11859-
11868.
Van de Walle, G. R., Sakamoto, K. & Osterrieder, N. (2008b). CCL3 and Viral
Chemokine-Binding Protein gG Modulate Pulmonary Inflammation and Virus
Replication during Equine Herpsvirus 1 Infection. J Virol 82, 1714-1722.
van der Meulen, K., Caij, B., Pensaert, M. & Nauwynck, H. (2006). Absence of viral
envelope proteins in equine herpesvirus 1-infected blood mononuclear cells during
cell-associated viremia. Vet Microbiol 113, 265-273.
van der Meulen, K., Vercauteren, G., Nauwynck, H. & Pensaert, M. (2003a). A local
epidemic of equine herpesvirus 1-induced neurological disorders in Belgium.
Vlaams Diergenskund Tijds 72, 366-372.
van der Meulen, K. M., Gryspeerdt, A. C., Vandekerckhove, A. A., Garre, B. A. & Nauwynck, H. J. (2007). The protective properties of vaccination against equine
herpesvirus 1-induced viremia, abortion and nervous system disorders. Vlaams
Diergenskund Tijds 76, 186-194.

Review of the literature
51
van der Meulen, K. M., Nauwynck, H. J. & Pensaert, M. B. (2003b). Absence of viral
antigens on the surface of equine herpesvirus-1-infected peripheral blood
mononuclear cells: a strategy to avoid complement-mediated lysis. J Gen Virol 84,
93-97.
van Maanen, C. (2002). Equine herpesvirus 1 and 4 infections: an update. Vet Q 24, 58-
78.
van Maanen, C., Sloet van Oldruitenborgh-Oosterbaan, M. M., Damen, E. A. & Derksen, A. G. (2001). Neurological disease associated with EHV-1-infection in
a riding school: clinical and virological characteristics. Equine Vet J 33, 191-196.
Van Maanen, C., Willink, D. L., Smeenk, L. A. J., Brinkhof, J. & Terpstra, C. (2000). An equine herpesvirus 1 (EHV1) abortion storm at a riding school. Vet Q
22, 83-87.
Varrasso, A., Dynon, K., Ficorilli, N., Hartley, C. A., Studdert, M. J. & Drummer, H. E. (2001). Identification of equine herpesviruses 1 and 4 by polymerase chain
reaction. Aust Vet J 79, 563-569.
Vengust, M., Wen, X. & Bienzle, D. (2008). Herpesvirus-associated neurological
disease in a donkey. J Vet Diagn Invest 20, 820-823.
Verryken, K., Saey, V., Maes, S., Borchers, K., Van de Walle, G., Ducatelle, R. & Deprez, P. (2010). First report of multinodular pulmonary fibrosis associated with
equine herpesvirus 5 in Belgium. Vlaams Diergen Tijds 79, 297-301.
Wang, L., Raidal, S. L., Pizzirani, A. & Wilcox, G. E. (2007). Detection of respiratory
herpesviruses in foals and adult horses determined by nested multiplex PCR. Vet
Microbiol 121, 18-28.
Wellington, J. E., Lawrence, G. L., Love, D. N. & Whalley, J. M. (1996a). Expression
and characterization of equine herpesvirus 1 glycoprotein D in mammalian cell
lines. Arch Virol 141, 1785-1793.
Wellington, J. E., Love, D. N. & Whalley, J. M. (1996b). Evidence for involvement of
equine herpesvirus 1 glycoprotein B in cell-cell fusion. Arch Virol 141, 167-175.
Whittaker, G. R., Taylor, L. A., Elton, D. M., Giles, L. E., Bonass, W. A., Halliburton, I. W., Killington, R. A. & Meredith, D. M. (1992). Glycoprotein
60 of equine herpesvirus type 1 is a homologue of herpes simplex virus
glycoprotein D and plays a major role in penetration of cells. J Gen Virol 73 ( Pt
4), 801-809.
Whitwell, K. E. & Blunden, A. S. (1992). Pathological findings in horses dying during
an outbreak of the paralytic form of Equid herpesvirus type 1 (EHV-1) infection.
Equine Vet J 24, 13-19.
Williams, K. J., Maes, R., Del Piero, F., Lim, A., Wise, A., Bolin, D. C., Caswell, J.,
Jackson, C., Robinson, N. E., Derksen, F., Scott, M. A., Uhal, B. D., Li, X., Youssef, S. A. & Bolin, S. R. (2007). Equine multinodular pulmonary fibrosis: a
newly recognized herpesvirus-associated fibrotic lung disease. Vet Pathol 44, 849-
862.
Wilson, W. D. (1997). Equine herpesvirus 1 myeloencephalopathy. Veterinary Clinics of
North America-Equine Practice 13, 53-&.
Wilsterman, S., Soboll-Hussey, G., Lunn, D. P., Ashton, L. V., Callan, R. J., Hussey, S. B., Rao, S. & Goehring, L. S. (2010). Equine herpesvirus-1 infected peripheral
blood mononuclear cell subpopulations during viremia. Vet Microbiol
doi:10.1016/j.vetmic.2010.10.004.
Wong, D. M., Belgrave, R. L., Williams, K. J., Del Piero, F., Alcott, C. J., Bolin, S.
R., Marr, C. M., Nolen-Walston, R., Myers, R. K. & Wilkins, P. A. (2008).

Review of the literature
52
Multinodular pulmonary fibrosis in five horses. J Am Vet Med Assoc 232, 898-
905.

Aims of the thesis
53
CHAPTER II
AIMS OF THE THESIS


Aims of the thesis
55
AIMS OF THE THESIS
Equine herpesvirus 1 (EHV1) is the most important cause of infectious abortion in
horses worldwide and can also lead to severe nervous system disorders with frequent fatal
outcome. Infection occurs via respiratory route and after local replication in tissues of the
upper respiratory tract, the virus spreads via a cell-associated viremia to target internal
organs such as the uterus or the nervous system. Secondary replication of EHV1 in
endothelial cells of these organs can result in late term abortion or nervous system
disorders.
Vaccination with the currently available vaccines cannot prevent clinical symptoms
such as abortion or nervous system disorders. Moreover, treatment of these symptoms is
only supportive and specific medication is not available. Therefore, an improvement of
existing vaccines and/or the development of therapies are necessary. Unfortunately, at the
start of this thesis, several important steps in the pathogenesis of this disease were still not
fully clarified, making it difficult to find new directions in the strategy to develop better
vaccines and to create drugs that inhibit steps of the pathogenesis. In order to achieve this
goal, studies in the natural host of EHV1, the horse, under both experimental and field
conditions, are of paramount importance to obtain relevant and correct information about
the pathogenesis of this disease.
Firstly, a thorough knowledge of the complete pathogenesis from primary replication
and invasion via cell-associated viremia to the target organs is urgent. Therefore, the first
aim of this thesis was to gain a more detailed insight in the invasion of neurologic and
non-neurologic strains of EHV1 from the upper respiratory tract to the carrier cells in the
blood stream by means of experimental infection studies in horses (Chapter III). The
second aim was to examine how individual EHV1-infected cells escape from immune
effector mechanisms at the primary site of replication (Chapter IV).
Secondly, to develop suitable vaccination and treatment strategies during natural
occurring outbreaks of disease, field information about the epidemiology of the virus is
needed. The third and last aim was therefore to gain more information about naturally
occurring outbreaks in Belgium in terms of risk factors, protective properties of
vaccination, the effect of experimental therapies and diagnosis (Chapter V).


Replication kinetics of EHV1
57
CHAPTER III
DIFFERENCES IN REPLICATION KINETICS
AND CELL TROPISM BETWEEN
NEUROVIRULENT AND NON-NEUROVIRULENT
EHV1 STRAINS DURING THE ACUTE PHASE OF
INFECTION IN HORSES
Veterinary microbiology 142 (2010), 242-253
Annick C. Gryspeerdt, A.P. Vandekerckhove, B. Garré, F. Barbé, G.R. Van de Walle, H.J.
Nauwynck


Replication kinetics of EHV1
59
SUMMARY
Equine herpesvirus 1 (EHV1) replicates in the respiratory tract of horses, after which
infected leukocytes transport virus throughout the body, resulting in abortion or nervous
system disorders. Two EHV1 strains circulate in the field: neurovirulent and non-
neurovirulent. To investigate differences in replication in the upper respiratory tract
(URT), an experimental inoculation study in ponies was performed with both strains. Two
groups of six ponies, were inoculated intranasally with 106.5
TCID50 of either strain.
Clinical signs, nasal shedding and viremia were evaluated. At early time points post
inoculation (pi), one pony of each group was euthanized. Tissues were collected for
titration and immunostainings. Number and size of EHV1-induced plaques were
calculated, and individual EHV1-infected cells were quantified and characterized.
Inoculation with either strain resulted in nasal shedding and replication in several tissues
of the URT. Both strains replicated in a plaquewise manner in epithelium of the nasal
mucosa, but replication in epithelium of the nasopharynx was largely limited to non-
neurovirulent EHV1. Plaques were never able to cross the basement membrane, but
individual infected cells were noticed in the connective tissue of all examined tissues for
both strains. The total number of these cells however, was 3-7 times lower with non-
neurovirulent EHV1 compared to neurovirulent EHV1. CD172a+ cells and CD5
+
lymphocytes were important target cells for both strains. Interestingly, in lymph nodes, B-
lymphocytes were also important target cells for EHV1, irrespective of the strain. Viremia
was detected very early pi and infected cells were mainly CD172a+ for both strains. In
summary, these results are valuable for understanding EHV1 pathogenesis at the port of
entry, the URT.

Replication kinetics of EHV1
60
1 INTRODUCTION
Infectious respiratory tract disease has been recognized as the cause of major health
problems in horses. The most important respiratory pathogen is equine herpesvirus 1
(EHV1) and infections with this virus cause serious economic losses in the horse industry
worldwide (Allen & Bryans, 1986; Ostlund, 1993; van Maanen, 2002). The upper
respiratory tract is the first line of defence against respiratory pathogens and is also the
primary replication site of EHV1, where it causes upper respiratory disorders (Kydd et
al., 1994a; b). In addition, EHV1 can spread via infected blood leukocytes to internal
organs, for example the pregnant uterus, causing symptoms such as abortion and neonatal
foal death (Allen & Bryans, 1986). EHV1 can also reach the central nervous system,
where replication in endothelial cells results in severe nervous system disorders with
frequent fatal outcome. Neurological disease in horses caused by infection with certain
isolates of EHV1 is a severe condition which is poorly understood (McCartan et al., 1995;
Stierstorfer et al., 2002; van der Meulen et al., 2003a). The pathogenesis for developing
this devastating condition is largely unknown and full protection against these secondary
and severe symptoms can not be obtained by vaccination with the currently available
vaccines (Goodman et al., 2006; Heldens et al., 2001; van der Meulen et al., 2007).
Therefore, an improvement of the existing vaccines and/or the development of alternative
strategies for the prevention and treatment of EHV1-induced infections are necessary.
Importantly, this implies a good understanding of the pathogenesis of EHV1 during the
acute phase of infection.
It has been suggested that distinct isolates of EHV1, differing in pathogenic capacity,
circulate in the field. Nugent et al. (2006) indicated that a single nucleotide polymorphism
(SNP) in the DNA polymerase is strongly associated with neurological versus non-
neurological disease outbreaks. Studies in naturally infected horses with nervous system
disorders have revealed a more robust cell-associated viremia in horses infected with
neurovirulent isolates of EHV1, in contrast to horses infected with non-neurovirulent
isolates (Allen & Breathnach, 2006). This was further confirmed by reverse genetic
experiments of EHV1 strains with a sole mutation of the DNA polymerase SNP, where
experimental inoculation with such strains altered neurologic disease in horses (Goodman
et al., 2007; Van de Walle et al., 2009). Moreover, in vitro experiments with these strains
revealed differences in leukocyte tropism which could explain the difference in clinical

Replication kinetics of EHV1
61
outcome (Goodman et al., 2007). Still, the exact identity of carrier cells susceptible to
EHV1 infection and hereby responsible for dissemination of infectious virus to sites of
secondary replication, is a matter of debate. Blood samples from experimentally infected
ponies were collected and examined for the presence of neurovirulent EHV1 (Scott et al.,
1983). T-lymphocytes were found to be the most important cell fraction to harbour virus
in vivo. However, no full characterization of these cells was performed. Different studies
with in vitro EHV1-infected leukocytes have also been performed to determine the
identity of these cells. Hereby, mainly monocytes were susceptible to infection with non-
neurovirulent EHV1 (van der Meulen et al., 2000). After stimulation with mitogens, T-
lymphocytes became more susceptible, followed by B-lymphocytes (van der Meulen et
al., 2001). Despite these studies, information on possible differences between
neurovirulent and non-neurovirulent EHV1 strains at the port of entry, and identity of
carrier cells during viremia remains limited.
To investigate this, six Shetland ponies were experimentally inoculated with a
neurovirulent EHV1 strain and six with a non-neurovirulent EHV1 strain. These animals
were subsequently euthanized to collect different tissues of the upper respiratory tract.
Tissues that were positive for EHV1 on titration were stained with markers for different
cell types to determine and quantify the carrier cells of EHV1 in the upper respiratory
tract. In addition, peripheral blood mononuclear cells (PBMC) were also collected to
quantify and identify the infected cell type during cell-associated viremia.
2 MATERIALS AND METHODS
2.1 Horses
Twelve male Shetland ponies, between the age of 6 months and 2 years, were used in
this study. They were housed inside isolated stables. They were fed daily with a
commercial, complete feed. Drinking water and hay were supplied ad libitum.
Prior to the experiment, ponies were monitored for 4 weeks. Rectal temperatures were
measured daily and complement-dependent seroneutralization (SN)-tests and
immunoperoxidase monolayer assays (IPMA) were performed weekly to determine EHV-
specific antibody titres.

Replication kinetics of EHV1
62
2.2 Virus and inoculation
Ponies were divided into two groups of 6. For the first group the Belgian EHV1 strain
03P37, isolated from the blood of a paralytic horse in 2003, was used for inoculation
(Garré et al., 2009; van der Meulen et al., 2003a). The second group was inoculated with
the Belgian strain 97P70, which was isolated from an aborted fetus (van der Meulen et
al., 2006). The strains were typed in the DNA polymerase as D752 and N752 respectively in
cooperation with the Animal Health Trust in the United Kingdom (Nugent et al., 2006).
Based on their origin and genotyping, these strains will be referred as neurovirulent and
non-neurovirulent in this paper. Virus stocks used for inoculation were at the 6th
passage;
2 passages in rabbit kidney cells (RK13) and 4 subsequent passages in equine embryonic
lung cells (EEL). Both viruses had virtually identical single-step growth properties in
RK13 cells for intra- and extracellular virus yields at all tested time points post infection
(data not shown).
Ponies were inoculated oronasally with a total amount of 20ml virus suspension
containing 106.5
tissue culture infectious dose 50 (TCID50). Ten ml of the virus suspension
was administered intranasally (5 ml per nostril) using a thin probe and 10 ml was
inoculated orally with a syringe. The virus titre was confirmed by titration of the
inoculum on RK13 cells.
2.3 Serological examination
For the seroneutralization (SN)-test, two-fold dilution series of the sera were prepared
in minimum essential medium (MEM). Fifty micro litre of these serial dilutions were
incubated for 23 hours (h) at 37°C with a fixed number of infectious virus (300 TCID50 of
EHV1 strain Arabica in 50 µl). Hereafter, 25 µl of guinea pig complement was added.
After 1 h of incubation, the mixture of serum, virus and complement was added to RK13
cells. Inoculated cultures were further incubated at 37°C in an atmosphere containing 5%
CO2. After 7 days of incubation, the cultures were analysed for the presence of cytopathic
effect. The neutralization titre was calculated as the reciprocal of the highest dilution of
the serum that was able to completely block EHV1 infection in RK13 cells.
For the immunoperoxidase monolayer assay (IPMA), RK13 cells were inoculated with
103 TCID50 of the EHV1 strain 97P70. After 28 h, cells were washed, dried at 37°C for 1

Replication kinetics of EHV1
63
h and stored at -20°C until use. Plates were thawed and subsequently fixed with 4%
paraformaldehyde and a solution containing 1% hydrogen peroxide in methanol.
Following extensive washing, serial 2-fold dilutions of the sera were added and cells were
incubated for 1 h at 37°C. Cells were incubated with peroxidase-labeled goat anti-horse Ig
(Jackson ImmunoResearch Laboratories Inc., PA, USA) and after 1 h, a substrate solution
of 3-amino-9-ethylcarbazole in 0.05 M acetate buffer with 0.05% hydrogen was added to
each well. Following 20 minutes of incubation at 37°C, substrate solution was replaced
with acetate buffer to block the enzymatic staining reaction. The IPMA titre was
calculated as the reciprocal value of the highest serum dilution that induced visual
staining of infected RK13 cells as determined by light microscopy.
2.4 Clinical examination
Horses were monitored daily for clinical signs by physical examination and
measurement of rectal temperatures. Fever was defined as a rectal temperature ≥ 38.5°C.
During clinical examination, nasal discharge (serous, mucous or purulent), lymph node
swelling, tachypnea and coughing were evaluated. In addition, neurological exams were
performed: walking the horse in a straight line, walking on and of a step, walking the
horse in tight circles, pulling the horse’s tail and backing the horse.
2.5 Virological examination
Nasopharyngeal mucus samples were taken daily until euthanasia. Immediately after
collection, swabs were immersed in transport medium containing phosphate-buffered
saline (PBS) supplemented with 10% fetal calf serum (FCS), 1000 U/ml penicillin, 1
mg/ml streptomycin and 0.5 mg/ml kanamycin. EHV1 in nasopharyngeal secretions was
titrated on monolayers of RK13 cells, exactly as described previously (van der Meulen et
al., 2003b).
On 1, 2, 3, 4, 5 and 7 days post inoculation, one pony of each group was euthanized
with an overdose of Natriumpentobarbital® (Kela, Hoogstraten, Belgium). After
euthanasia, different tissue samples were collected from the upper respiratory tract.
Tissues from slaughterhouse horses were collected and served as uninfected control
samples. Per tissue, two equal squares of 1 cm2 were collected from both left and right

Replication kinetics of EHV1
64
side of the head. From each side, one square was frozen immediately with
(immunofluorescence) or without (virus titration) methylcellulose medium
(Methocel®MC, Sigma-Aldrich, St. Louis, MO, USA) at -70°C. To determine viral
replication, 20% suspensions of collected tissues were made and titrated on RK13 cells.
Titres of infected tissues were determined after 7 days by means of Reed and Muench’s
formula (Reed & Muench, 1938).
The experimental design was approved by the local ethical committee of the Faculty
of Veterinary Medicine.
2.6 Quantification and characterization of individual infected cells in tissues of the
upper respiratory tract
Immunofluorescent double stainings were used to quantify and characterize individual
infected cells in tissues that were EHV1-positive after titration on RK13 cells. First,
cryosections (16 µm) were incubated with either monoclonal antibody (mAb) HT23A,
DH59B or 1.9/3.2 (VMRD Inc, Pullman, WA, USA) against CD5 (T-lymphocytes),
CD172a (cells from the monocyte lineage, CML) or IgM (B-lymphocytes) respectively,
and subsequently incubated with Texas Red®-labelled goat anti-mouse Abs (Molecular
Probes, Eugene, OR, USA). Epithelial cells were identified based on the pancytokeratin
marker MNF116 (Abcam, Cambridge, UK) and/or morphological features. Second, viral
EHV1 proteins were stained by incubation with biotinylated polyclonal horse anti-EHV1
IgG (van der Meulen et al., 2000), followed by streptavidin-FITC® (Molecular Probes,
Eugene, OR, USA). As controls, (i) stainings were performed on uninfected tissue and (ii)
isotype control antibodies were included. All cryosections were analyzed using confocal
microscopy (Leica TCS SP2 Laser scanning spectral confocal system, Leica
Microsystems GmbH, Wetzlar, Germany). Number and latitude of EHV1-induced
plaques were calculated using the software program ImageJ. Latitude was used to
evaluate plaque size and therefore the degree of cell-to-cell spread. In order to quantify
infected cells in nasal septum, two main regions were taken into account, on the one hand
regions with and on the other hand regions without plaques in the epithelium. These
regions were then subdivided into three equal regions underneath the epithelium: A, B
and C for regions with epithelial plaques and D, E and F for regions without epithelial
plaques (Fig 4). For nasopharynx, tubal/nasopharyngeal tonsils and lymph nodes,

Replication kinetics of EHV1
65
subdivisions into different zones of functional importance were made (Fig 5, 6 and 7
respectively). All regions were evaluated for the presence of plaques and/or individual
infected cells. In addition, the total number of infected cells (positive for staining with
anti-EHV1 Abs), equine immune cells (positive for staining with the mAbs against
immune cell markers) and infected equine immune cells (double positive) were
determined.
2.7 Quantification and characterization of viremia
Heparinized blood samples were taken daily until euthanasia and peripheral blood
mononuclear cells (PBMC) were isolated by density centrifugation on Ficoll-Paque,
according to manufacturer’s instructions (Pharmacia Biotech AB, Uppsala, Sweden). To
determine the magnitude of viremia, co-cultivation of PBMC on RK13 cells was
performed as previously described (van der Meulen et al., 2003b). Remaining PBMC
were resuspended in RPMI supplemented with 30% FCS and 20% Dimethyl Sulfoxide
(DMSO) and frozen in liquid nitrogen for later examination to identify the nature of
infected cells. Hereby, a double immunofluorescence staining was performed on acetone-
fixed cell smears of PBMC. EHV1 expression was detected using a biotinylated
polyclonal horse anti-EHV1 IgG (van der Meulen et al., 2003b), followed by streptavidin-
FITC®. The identity of infected cells was determined using the mAbs HT23A, DH59B or
1.9/3.2, as described above, followed by the incubation with Texas Red®-labelled goat
anti-mouse Abs. Samples were analyzed using confocal laser scanning microscopy.
3 RESULTS
3.1 EHV-status of the ponies before inoculation
The inclusion criterium for ponies used in this study was a complete absence of EHV-
specific antibodies by either SN-test (<2) or IPMA (<10). In addition, during the 4-week
observation period prior to the experiment, (i) no raise in temperature was noted, (ii) no
EHV-specific antibodies were detected, and (iii) no virus was isolated from
nasopharyngeal swabs. The use of naïve horses is an important tool to minimize variation

Replication kinetics of EHV1
66
in clinical and virological outcome due to differences in immunological status (Cornick et
al., 1990; Heldens et al., 2001; Matsumura et al., 1996).
3.2 Experimental inoculation with either strain induces fever and upper respiratory
tract disease in ponies
All ponies developed fever upon EHV1 inoculation with exception of the two ponies
that were euthanized 24 hours post inoculation (hpi). Fever started at 48 hpi for the
neurovirulent strain (5/5 animals) and at 36 (3/5 animals) or 48 hpi (2/5 animals) for the
non-neurovirulent strain. High temperatures lasted for the complete observation period,
with peak temperatures of 40.7°C for the neurovirulent strain and 40.1°C for the non-
neurovirulent strain. Serous nasal discharge was noted after inoculation with both strains,
starting at 1 day post inoculation (dpi). A transition to purulent nasal discharge was seen
in both groups from 3 dpi. Breathing problems, consisting of tachypnea or coughing, were
observed in two ponies of each group. Swelling of mandibular lymph nodes was observed
in the majority of the ponies and occurred from 1 dpi in 2/6 animals inoculated with a
neurovirulent strain and in 4/6 animals inoculated with a non-neurovirulent strain. Painful
retropharyngeal lymph nodes were noted in 5/6 ponies inoculated with a non-
neurovirulent strain, in contrast to only 2 ponies inoculated with a neurovirulent strain.
None of the ponies showed neurological signs, irrespective of the strain used for
inoculation. The latter is not too unexpected, as the incubation period of EHV1-induced
neurological disorders is normally 6 to 10 days (Edington et al., 1986; Jackson et al.,
1977; Mumford & Edington, 1980). Moreover, nervous system disorders have been
described to occur more frequently in older horses and certain breeds (Garré et al., 2009;
Goehring et al., 2006; Greenwood & Simson, 1980; Jackson et al., 1977; McCartan et al.,
1995).
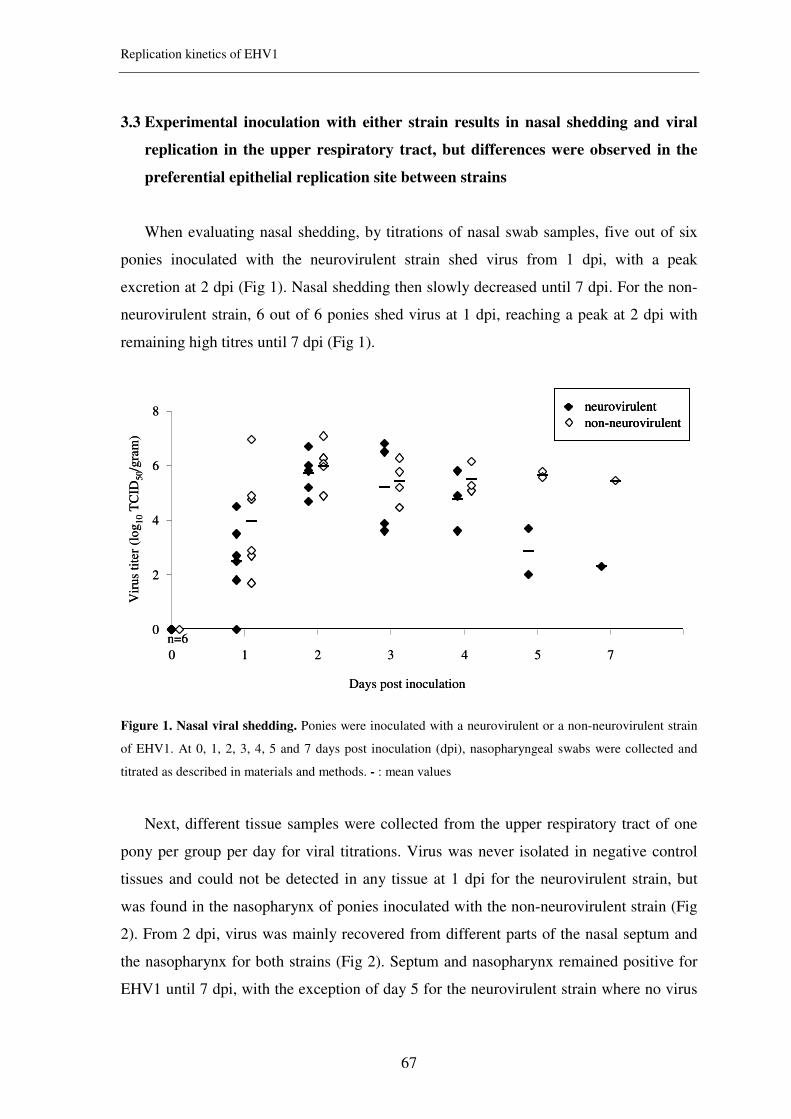
Replication kinetics of EHV1
67
3.3 Experimental inoculation with either strain results in nasal shedding and viral
replication in the upper respiratory tract, but differences were observed in the
preferential epithelial replication site between strains
When evaluating nasal shedding, by titrations of nasal swab samples, five out of six
ponies inoculated with the neurovirulent strain shed virus from 1 dpi, with a peak
excretion at 2 dpi (Fig 1). Nasal shedding then slowly decreased until 7 dpi. For the non-
neurovirulent strain, 6 out of 6 ponies shed virus at 1 dpi, reaching a peak at 2 dpi with
remaining high titres until 7 dpi (Fig 1).
Figure 1. Nasal viral shedding. Ponies were inoculated with a neurovirulent or a non-neurovirulent strain
of EHV1. At 0, 1, 2, 3, 4, 5 and 7 days post inoculation (dpi), nasopharyngeal swabs were collected and
titrated as described in materials and methods. - : mean values
Next, different tissue samples were collected from the upper respiratory tract of one
pony per group per day for viral titrations. Virus was never isolated in negative control
tissues and could not be detected in any tissue at 1 dpi for the neurovirulent strain, but
was found in the nasopharynx of ponies inoculated with the non-neurovirulent strain (Fig
2). From 2 dpi, virus was mainly recovered from different parts of the nasal septum and
the nasopharynx for both strains (Fig 2). Septum and nasopharynx remained positive for
EHV1 until 7 dpi, with the exception of day 5 for the neurovirulent strain where no virus
0
2
4
6
8
0 1 2 3 4 5 7
Days post inoculation
Vir
us
tite
r (l
og
10
TC
ID5
0/g
ram
)
n=6
neurovirulent
non-neurovirulent
0
2
4
6
8
0 1 2 3 4 5 7
Days post inoculation
Vir
us
tite
r (l
og
10
TC
ID5
0/g
ram
)
n=6
neurovirulent
non-neurovirulent
neurovirulent
non-neurovirulent

Replication kinetics of EHV1
68
was detected in the nasopharynx (Fig 2) and day 4 for the non-neurovirulent strain where
the distal nasal septum was negative (Fig 2). No difference in virus titres could be noticed
in the nasal septum, but titres found in the nasopharynx seemed to be higher after
inoculation with the non-neurovirulent strain of EHV1 compared to the neurovirulent
strain (Fig 2). However, because of the limited number of animals used, we should be
careful to draw any conclusions based on these results. Virus was also found in lymphoid
tissues such as the tubal and nasopharyngeal tonsils and mandibular lymph nodes (Fig 2).
In general, titres found in tubal and nasopharyngeal tonsils (Fig 2) were higher for the
non-neurovirulent strain. Titres detected in mandibular lymph nodes started rising from 3
dpi (neurovirulent strain) or 4 dpi (non-neurovirulent strain), and at 7 dpi, mandibular
lymph nodes were positive for the non-neurovirulent strain only (Fig 2). EHV1 could also
be isolated for both strains at different time points pi from the ethmoid, the cranial part of
the trachea and 3 different tonsils (data not shown).

Replication kinetics of EHV1
69
Figure 2. Viral replication in different tissues of the upper respiratory tract. Ponies were inoculated
with either neurovirulent or non-neurovirulent EHV1 and euthanized at 0, 1, 2, 3, 4, 5 and 7 days post
inoculation (dpi). Different tissues were collected and titrated as described in materials and methods.
Finally, replication of EHV1 was evaluated in the epithelium of different tissues of
the upper respiratory tract by immunofluorescence. Hereby, EHV1-induced plaques were
analyzed and plaque latitudes measured. In the epithelium of the entire nasal septum,
EHV1-induced plaques were seen at different time points pi for both strains (Fig 3A).
These plaques were defined as groups of infected epithelial cells. Since results for
proximal, intermedial and distal nasal septum were virtually identical, only results for the
intermedial nasal septum are shown. Plaques were not present in the epithelium at 0 and 1
dpi (Fig 3A). However, individual infected cells were observed at 1 dpi, which were
identified as CD172a+ cells, as determined by the marker DH59B, or epithelial cells, as
determined by the pancytokeratin marker (data not shown). For the neurovirulent strain,
0
2
4
6
0 1 2 3 4 5 7
0
2
4
6
0 1 2 3 4 5 7
Vir
us
tite
r (l
og
10
TC
ID5
0/g
ram
tis
sue)
Days post inoculation
0
2
4
6
0 1 2 3 4 5 7
Intermedial nasal septum
0
2
4
6
0 1 2 3 4 5 7
Intermedial nasal septum
Tubal and nasopharyngeal tonsils
0
2
4
6
0 1 2 3 4 5 7
Distal nasal septum
0
2
4
6
0 1 2 3 4 5 7
Distal nasal septum
0
2
4
6
0 1 2 3 4 5 7
Proximal nasal septum
0
2
4
6
0 1 2 3 4 5 7
Proximal nasal septum
Mandibular lymph nodes
0
2
4
6
0 1 2 3 4 5 7
Nasopharynx
0
2
4
6
0 1 2 3 4 5 7
Nasopharynx
neurovirulent
non-neurovirulent
neurovirulent
non-neurovirulent

Replication kinetics of EHV1
70
EHV1-induced plaques were observed in the epithelium of the nasal mucosa starting from
2 till 7 dpi (Fig 3A). At 2, 3 and 4 dpi, these plaques consisted of epithelial cells and
rarely one or two CD172a+ cells. From 5 dpi, EHV1-positive cells became separated from
the basement membrane (BM) and these cells were exclusively positive for the DH59B
marker which identifies cells of the monocytic lineage (CML) (Fig 3A). This observation
was not seen in ponies inoculated with a non-neurovirulent strain. There, plaques were
present in the epithelium of the nasal mucosa and nasopharynx from 2 till 7 dpi (Fig 3A &
B). These plaques consisted of epithelial cells and rarely some CD172a+ cells, but no
separation of EHV1-positive cells was observed (Fig 3A). No differences were observed
in both number (data not shown) and size (Fig 3A) of EHV1-induced plaques in the
intermedial nasal mucosa at different time points pi for both strains. In the epithelium of
the nasopharynx, plaques could be found for the non-neurovirulent strain from 2 till 7 dpi,
with the exception of 4 dpi (Fig 3B). In contrast, no plaques were found for the
neurovirulent strain in the nasopharyngeal epithelium, with the exception of one small
plaque at 3 dpi (Fig 3B). Interestingly, plaques were not able to cross the BM at any time
point pi, but single EHV1-infected cells could be noticed in the connective tissue from 1
dpi. Plaques or individual cells could never be visualised in uninfected control tissues or
in tissues stained with isotype control antibodies.

Replication kinetics of EHV1
71
Figure 3. Plaque formation in epithelium of intermedial nasal septum and nasopharynx. Tissues of the
intermedial part of the nasal septum (A) and nasopharynx (B) were collected and latitude of plaques were
determined. For each tissue, ten consecutive sections were made and the total number of individual plaques
was determined. This number is presented above the relevant histogram. The largest measurement of
latitude of each individual plaque was used to determine plaque latitude. The sum was used to calculate the
mean plaque latitude ± SEM. Representative immunofluorescence pictures of EHV1-induced plaques in the
intermedial nasal septum are shown for neurovirulent or non-neurovirulent EHV1 for each dpi. BM:
basement membrane, objective 20x, NI: non-infected epithelial cells.
A
neuro-
virulent
B
Days post inoculation
Days post inoculation
Pla
que
lati
tud
e(µ
m)/
0.8
mm
2
NI NI
0
20
40
60
80
100
120
140
160
0 1 2 3 4 5 7
neurovirulent
non-neurovirulentP
laq
ue
lati
tud
e(µ
m)/
0.8
mm
2
neurovirulent
non-neurovirulent
0
20
40
60
80
100
120
140
160
0 1 2 3 4 5 7
non-
neuro-
virulent
BM
BM
7
3
2
9
10
31
12
12
2
48
1
A
neuro-
virulent
B
Days post inoculation
Days post inoculation
Pla
que
lati
tud
e(µ
m)/
0.8
mm
2
NI NI
0
20
40
60
80
100
120
140
160
0 1 2 3 4 5 7
neurovirulent
non-neurovirulentP
laq
ue
lati
tud
e(µ
m)/
0.8
mm
2
neurovirulent
non-neurovirulent
0
20
40
60
80
100
120
140
160
0
20
40
60
80
100
120
140
160
0 1 2 3 4 5 7
non-
neuro-
virulent
BM
BM
7
3
2
9
10
31
12
12
2
48
1

Replication kinetics of EHV1
72
Taken together, these results show that EHV1 infection results in nasal shedding and
replication in several tissues of the upper respiratory tract, irrespective of its neurovirulent
potential. However, whereas both types of EHV1 replicate equally in the epithelium of
the nasal mucosa, replication in the nasopharynx was limited to non-neurovirulent EHV1.
3.4 Neurovirulent EHV1 uses mainly CD172a+ cells as carrier cells in tissues of the
upper respiratory tract, whereas non-neurovirulent EHV1 shows a tropism for
both CD172a+ cells and CD5
+ cells
As mentioned before, EHV1-induced plaques never crossed the BM, irrespective of
the strain used. However, individual infected cells were observed in the underlying
connective tissue and the total number was 3-7 times lower with non-neurovirulent EHV1
compared to neurovirulent EHV1. Individual infected cells were never observed in
uninfected control tissues or in tissues stained with isotype control antibodies.
To identify the nature of these cells, cryosections of tissues were made and double
immunofluorescence stainings were performed.

Replication kinetics of EHV1
73
Nasal septum. In the connective tissue of the nasal septum, no differences were
observed in the total amount of infected cells present beneath epithelium with plaques or
without plaques, with the exception of 5 and 7 dpi for the neurovirulent strain, where a
marked increase in infected cells was observed in region A (Fig 4). Such increase was not
seen in the regions located underneath epithelium without plaques (Fig 4).
Figure 4. Quantification and identification of single EHV1-infected cells in different zones of the
intermedial nasal septum for neurovirulent and non-neurovirulent strains. Every tenth section of a
serially sectioned block of tissue was collected. Ten sections were analysed for each cell marker separately,
giving a total number of 30 sections. The total number of infected cells present in the intermedial nasal
septum per zone and per day, is shown in the upper right table. The results for the identification of single
infected cells are shown in the lower graphs and are represented as the mean of ten sections ± SEM. ROI:
region of interest.
B
A
C F
E
D
a
b
plaque
d
c
d
Strain Roi Total number of single infected cells
present at … dpi/30 sections
0 1 2 3 4 5 7
A - - 34 17 34 137 82
B - - 52 13 62 8 60
Neurovirulent
C - - 24 8 40 4 18
D 0 3 24 34 62 11 9
E 0 0 24 10 58 11 5
F 0 0 28 9 22 8 2
A - - 3 0 4 1 0
B
C
D
E
F
Non-
neurovirulent - - 4 3 15 0 0
- - 0 1 0 0 0
0 0 6 0 19 8 0
0 0 0 0 17 6 0
0 0 0 0 25 3 0
CD5+
CD172a+
IgM+
CD5+CD5+
CD172a+CD172a+
IgM+IgM+
N°
of
infe
cted
cell
s/1
0 s
ecti
on
s
Days post inoculation
Schematic overview of the structure of the nasal
septum and different counting regions
a: epithelium; b: connective tissue; c: glands;
d: blood vessel
CD5+
CD172a+
IgM+
Days post inoculation
0
10
20
30
40
50
60
70
80
90
0 1 2 3 4 5 7
CD5+
CD172a+
IgM+
CD5+CD5+
CD172a+CD172a+
IgM+IgM+
Days post inoculation
0
10
20
30
40
50
60
70
80
90
0 1 2 3 4 5 70
10
20
30
40
50
60
70
80
90
0 1 2 3 4 5 7
Neurovirulent strain Non-neurovirulent strain

Replication kinetics of EHV1
74
Nasopharynx. EHV1-infected cells were present in the connective tissue of the
nasopharynx from 1 till 7 dpi, irrespective of the strain used, however, the localization of
these cells appeared to be strain-dependent (Fig 5). Whereas for the neurovirulent strain,
single infected cells were mainly found in epithelium (region a) and connective tissue
(region b), single infected cells were located in connective tissue (region b) and
lymphocyte aggregates (region c) for the non-neurovirulent strain (Fig 5).
Figure 5. Quantification and identification of single EHV1-infected cells in different zones of the
nasopharynx for neurovirulent and non-neurovirulent strains. Every tenth section of a serially
sectioned block of tissue was collected. Ten sections were analysed for each cell marker separately, giving a
total number of 30 sections. The total number of infected cells present in the nasopharynx per zone and per
day, is shown in the upper right table. The results for the identification of single infected cells are shown in
the lower graphs and are represented as the mean of ten sections ± SEM. ROI: region of interest.
a
b
d
c
e
Strain Roi Total number of single infected cells
present at … dpi/30 sections
0 1 2 3 4 5 7
a 0 3 3 7 2 0 1
b 0 22 24 118 22 18 23
Neurovirulent
c 0 0 0 0 0 0 0
d 0 0 0 0 0 0 0
e 0 0 0 0 0 0 0
a 0 0 0 0 0 0 0
b 0 1 320 15 28 18 1
Non-
neurovirulent
c 0 0 0 0 99 16 1
d 0 0 0 0 0 0 0
e 0 0 0 0 0 1 0
CD5+
CD172a+
IgM+
CD5+CD5+
CD172a+CD172a+
IgM+IgM+
Days post inoculation
CD5+
CD172a+
IgM+
CD5+CD5+
CD172a+CD172a+
IgM+IgM+
Days post inoculation
c
Schematic overview of the structure of the
nasopharynx and different counting regions
a: epithelium; b: connective tissue; c: lymphocyte
aggregate; d: deeper connective tissue; e: blood vessels
Neurovirulent strain Non-neurovirulent strain
0
5
10
15
20
25
30
35
40
0 1 2 3 4 5 70
5
10
15
20
25
30
35
40
0 1 2 3 4 5 7
N°
of
infe
cted
cell
s/10
sec
tion
s

Replication kinetics of EHV1
75
Tubal and nasopharyngeal tonsils. For both EHV1 strains, single infected cells were
observed starting from 2 dpi (Fig 6). They were mostly present in the connective tissue
underneath the epithelium (region b) and lymphocyte aggregates (region c, Fig 6), but
single infected cells could also be observed in epithelium/fundus (region a), deeper
connective tissue (region d), blood vessels (region e) and glands (region f) (Fig 6).
Figure 6. Quantification and identification of single EHV1-infected cells in different zones of the tubal
and nasopharyngeal tonsils for neurovirulent and non-neurovirulent strains. Every tenth section of a
serially sectioned block of tissue was collected. Ten sections were analysed for each cell marker separately,
giving a total number of 30 sections. The total number of infected cells present in the tubal and
nasopharyngeal tonsils per zone and per day, is shown in the upper right table. The results for the
identification of single infected cells are shown in the lower graphs and are represented as the mean of ten
sections ± SEM. ROI: region of interest.
Strain Roi Total number of single infected cells
present at … dpi/30 sections
0 1 2 3 4 5 7
a 0 0 5 77 95 0 14
b 0 0 6 273 538 1 51
Neurovirulent
c 0 0 3 260 241 2 0
d 0 0 0 86 150 2 7
e 0 0 0 133 85 0 2
f 0 0 0 38 117 1 13
a
b
d
c
e
f
a 0 0 7 23 7 18 5
b 0 0 39 173 51 50 2
Non-
neurovirulent
c 0 0 10 34 31 487 8
d 0 0 44 78 14 8 2
e 0 0 0 5 3 0 2
f 0 0 2 0 2 4 13
CD5+
CD172a+
IgM+
CD5+CD5+
CD172a+CD172a+
IgM+IgM+
Days post inoculation
CD5+
CD172a+
IgM+
CD5+CD5+
CD172a+CD172a+
IgM+IgM+
Days post inoculation
Schematic overview of the structure of tubal
and nasopharyngeal tonsils and different
counting regions
a: epithelium; b: connective tissue; c: lymphoid aggregate;
d: deeper connective tissue; e: blood vessel; f: glands
Neurovirulent strain Non-neurovirulent strain
0
50
100
150
200
250
300
350
400
0 1 2 3 4 5 70
50
100
150
200
250
300
350
400
0 1 2 3 4 5 7
N°
of
infe
cted
cell
s/10 s
ect
ions

Replication kinetics of EHV1
76
Mandibular lymph nodes. For the neurovirulent strain, most of the infected cells
were found in the marginal sinus of the mandibular lymph nodes (region c, Fig 7). Their
number increased until 4 dpi and rapidly decreased afterwards, with rarely any infected
cells left at 7 dpi. Furthermore, infected cells were also observed in the capsule and
trabecula (region a), the medulla (region f) and in fewer amounts in vessels of the capsule
and the cortical nodules (regions b and d, Fig 7). In these regions a peak number of
infected cells was observed at 4 and 5 dpi (Fig 7). For the non-neurovirulent strain, single
cells were only found from 4 till 7 dpi and were mostly found in the marginal sinus
(region c) and medulla (region f) (Fig 7).
Figure 7. Quantification and identification of single EHV1-infected cells in different zones of the
mandibular lymph node for neurovirulent and non-neurovirulent strains. Every tenth section of a
serially sectioned block of tissue was collected. Ten sections were analysed for each cell marker separately,
giving a total number of 30 sections. The total number of infected cells present in the mandibular lymph
node per zone and per day, is shown in the upper right table. The results for the identification of single
infected cells are shown in the lower graphs and are represented as the mean of ten sections ± SEM. ROI:
region of interest.
Strain Roi Total number of single infected cells
present at … dpi/30 sections
0 1 2 3 4 5 7
a 0 0 0 40 10 162 1
b 0 12 2 16 40 12 0
Neurovirulent
c 0 17 6 106 846 170 34
d 0 2 2 31 36 4 19
e 0 0 0 3 1 0 0
f 0 0 9 49 70 14 39
a
b
c d
e f
a
b
c d
e f
CD5+
CD172a+
IgM+
CD5+CD5+
CD172a+CD172a+
IgM+IgM+
Days post inoculation
CD5+
CD172a+
IgM+
CD5+CD5+
CD172a+CD172a+
IgM+IgM+
Days post inoculation
a 0 0 0 0 4 5 1
b 0 0 0 0 0 0 0
Non-
neurovirulent
c 0 1 2 4 40 39 1
d 0 0 0 0 5 17 8
e 0 0 0 0 0 0 0
f 0 0 0 0 41 82 1
Schematic overview of the structure of the
mandibular lymph node and different counting
regions
a: capsule and trabecula; b: vessels of the capsule; c:
marginal sinus; d: cortical nodule; e: vessels in medulla;
f: medullaNeurovirulent strain Non-neurovirulent strain
0
50
100
150
200
250
300
350
400
0 1 2 3 4 5 70
50
100
150
200
250
300
350
400
0 1 2 3 4 5 7
N°
of
infe
cted
cell
s/10
sec
tio
ns

Replication kinetics of EHV1
77
Immunofluorescence stainings, using mAbs against immune cell surface markers
showed that in all tissues of the upper respiratory tract, infected cells were largely
CD172a+ CML, followed by a slightly lower amount of infected CD5
+ lymphocytes when
the neurovirulent EHV1 strain was used for inoculation (Table I). These results were very
similar to what was observed with the non-neurovirulent strain, although a more
substantial amount of infected CD5+ lymphocytes was observed in these different tissues
of the upper respiratory tract (Table I). IgM+ lymphocytes were always present, but rarely
infected with the exception of the mandibular (Table I) and retropharyngeal (data not
shown) lymph nodes, were these cells were an important target cell type for both strains.
Table I. Percentage of marker positive EHV1-infected individual cells in different tissues of the upper
respiratory tract
Percentage of marker positive EHV1-infected individual cells
Neurovirulent strain Non-neurovirulent strain
Region of interest
CD172a+ CD5
+ IgM
+ CD172a
+ CD5
+ IgM
+
Nasal septum 53.4 ± 22,8 16.2 ± 11.7 0.3 ± 0.8 43.9 ± 12.8 23.6 ± 8.6 0
Nasopharynx 69.5 ± 20.7 0 8.3 ± 20.4 37.9 ± 35.5 13.6 ± 5.7 1.1 ± 2.7
Tubal and
nasopharyngeal tonsils
66.4 ± 20.7 11.2 ± 12.8 4.8 ± 5.6 28.7 ± 3.6 21.2 ± 5.9 0.5 ± 1.0
Mandibular lymph
nodes
71.3 ± 17.9 7.3 ± 9.5 8.3 ± 8.8 42.6 ± 21 26.7 ± 5.7 7.7 ± 10.8
3.5 Ponies became viremic upon experimental inoculation with either strain, and
CD172a+ cells were the most important carrier cells in the blood
Transmission of EHV1 from PBMC to susceptible RK13 cells, as determined by co-
cultivation, was observed in 5 out of 6 ponies upon experimental inoculation with
neurovirulent EHV1 (Fig 8). No differences in the amount of infected PBMC was noted,
except for one pony, which showed a higher level of viremia at 4 and 5 dpi compared to
the other infected ponies (Fig 8). For the non-neurovirulent strain, viremia was observed
in all six infected ponies, with no differences in magnitude for the first 4 dpi (Fig A).
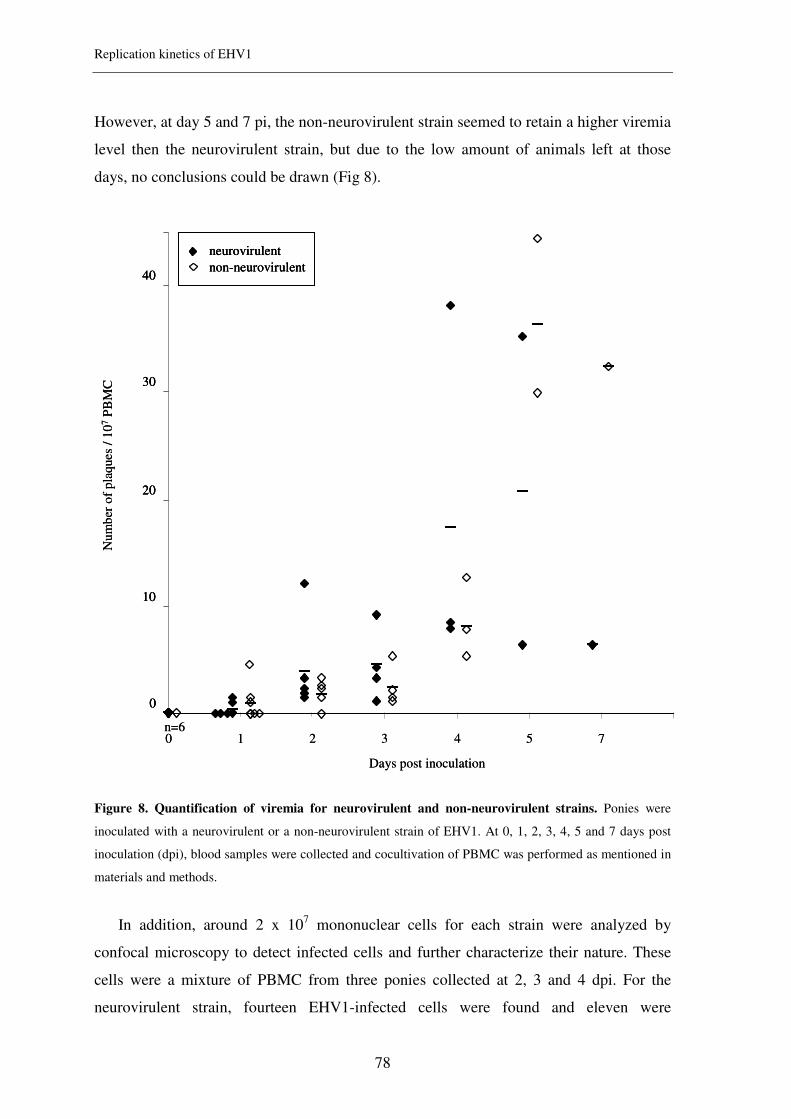
Replication kinetics of EHV1
78
However, at day 5 and 7 pi, the non-neurovirulent strain seemed to retain a higher viremia
level then the neurovirulent strain, but due to the low amount of animals left at those
days, no conclusions could be drawn (Fig 8).
Figure 8. Quantification of viremia for neurovirulent and non-neurovirulent strains. Ponies were
inoculated with a neurovirulent or a non-neurovirulent strain of EHV1. At 0, 1, 2, 3, 4, 5 and 7 days post
inoculation (dpi), blood samples were collected and cocultivation of PBMC was performed as mentioned in
materials and methods.
In addition, around 2 x 107 mononuclear cells for each strain were analyzed by
confocal microscopy to detect infected cells and further characterize their nature. These
cells were a mixture of PBMC from three ponies collected at 2, 3 and 4 dpi. For the
neurovirulent strain, fourteen EHV1-infected cells were found and eleven were
n=6
neurovirulent
non-neurovirulent
0
10
20
30
40
0 1 2 3 4 5 7
Days post inoculation
Nu
mb
ero
f p
laques
/ 1
07
PB
MC
n=6
neurovirulent
non-neurovirulent
neurovirulent
non-neurovirulent
0
10
20
30
40
0
10
20
30
40
0 1 2 3 4 5 7
Days post inoculation
Nu
mb
ero
f p
laques
/ 1
07
PB
MC

Replication kinetics of EHV1
79
characterized as CD172a+ cells (78.5%), whereas three were CD5
+ (21.5%). No infected
B-lymphocytes were found. For the non-neurovirulent strain, nine EHV1-infected cells
were identified, consisting of seven CD172a+ cells (77.8%), one CD5
+ T-lymphocyte
(11.0%) and one cell remained unidentified. Also here, no infected B-lymphocytes were
found.
These data indicate that CD172a+ cells appear to be the main target cell during EHV1-
induced viremia, irrespective of the neurovirulency of EHV1.
4 DISCUSSION
Despite several studies conducted on the pathogenesis of equine herpesvirus 1
(EHV1), it still remains unclear how this virus invades the tissues of the upper respiratory
tract and invades the deeper tissues to initiate a cell-associated viremia. Furthermore, two
strains of EHV1 circulate in the field, namely non-neurovirulent and neurovirulent strains,
the latter causing neurological disease, a severe condition which is poorly understood. In
the present study, we used the two strains of EHV1 to gain more insight into the
pathogenesis of these strains during the acute phase of infection in the upper respiratory
tract in vivo. To this end, Shetland ponies were experimentally inoculated with either a
non-neurovirulent or a neurovirulent EHV1 strain and upon euthanasia, several tissues
from the upper respiratory tract were collected and analyzed.
It was found that EHV1 has different approaches to successfully establish an infection
in the upper respiratory tract. Firstly, we observed that EHV1, irrespective of the strain,
can replicate in the epithelium of the nasal septum and/or nasopharynx by means of virus-
induced plaques. This replication appears to occur in a restricted way, as no increase in
number or size of plaques was observed over time. In addition, a repulsion of EHV1-
induced plaques was observed from 5 dpi onwards for the neurovirulent strain, and a
decrease in plaque latitude was seen for the non-neurovirulent strain starting at day 3 pi.
This reduction in replication might be attributed to the production of interferon (IFN), as
(i) the presence of interferon in nasal secretions after experimental inoculation of horses
with EHV1 has previously been described and (ii) peak concentrations of IFN-α were
noted at day 4 and day 7 pi (Bridges & Edington, 1986; Edington et al., 1989). In the
present study, IFN-α was also found in nasal swabs of experimentally infected ponies
(data not shown), which further points towards the importance of IFN to limit EHV1

Replication kinetics of EHV1
80
replication in the epithelium of the upper respiratory tract. Interestingly, even though the
non-neurovirulent strain was able to replicate in a plaquewise manner in the epithelium of
the nasopharynx, no plaques could be observed at any time point pi in the nasopharynx
after inoculation with the neurovirulent strain. As the epithelial structure of both tissues is
identical, with the exception of the presence of lymphoid aggregates in the nasopharynx,
the reason for this observation remains elusive.
Another interesting observation for both EHV1 strains was that we were unable to
observe EHV1-infected plaques under the basement membrane (BM) at any time point pi,
indicating that EHV1 does not reach the underlying tissue by breaking down the BM.
This is in striking contrast to previous findings with another and closely related
alphaherpesvirus, pseudorabies virus (PRV), which has been shown to cross the BM in a
plaque wise manner between 12 and 24 hpi in a porcine respiratory explant model
(Glorieux et al., 2009). This indicates that different alphaherpesviruses may use different
mechanisms to invade the underlying connective tissue. Indeed, whereas PRV was shown
to break down the BM and replicates further in the underlying tissue in a plaquewise
manner, our study with EHV1 demonstrated only single infected cells below the BM.
This is in agreement with previous studies on EHV1, where the presence of single
infected cells in tissues of the upper respiratory tract has been reported (Edington et al.,
1986; Kydd et al., 1994b). We could further demonstrate that these infected cells in the
underlying tissue were immune cells (as discussed in more detail below), which makes it
tempting to speculate that immune cells in the epithelium become infected with EHV1
and are responsible for transporting the virus to deeper connective tissue. Indeed, we did
found EHV1-infected cells in the epithelium of nasal septum and/or nasopharynx, which
were further characterized to belong to the monocytic lineage (CML) by using the cell
surface maker CD172a.
Not only differences concerning the identity of the EHV1-infected immune cells
present in the underlying tissues were noticed between both strains, moreover, differences
concerning their localization were also observed. When analyzing the identity of the
single EHV1-infected cells in the connective tissue of nasal septum and nasopharynx,
several differences could be observed between the two EHV1 strains. Firstly, the total
number of infected cells in the nasal septum was 10 to 20 times higher for the
neurovirulent strain compared to the non-neurovirulent strain. Secondly, while individual
infected cells in the nasopharynx were mainly present in the connective tissue upon

Replication kinetics of EHV1
81
neurovirulent EHV1 inoculation, individual cells infected with the non-neurovirulent
EHV1 seemed to migrate deeper into the lymphoid follicles of the nasopharynx in a time
wise manner. Thirdly, CD172a+ CML were the most important carrier cells for both
neurovirulent and non-neurovirulent EHV1, although these cells seemed to be more
important for the neurovirulent strain. In addition, both strains were able to infect CD5+
T-lymphocytes, with a higher amount of infected cells for the non-neurovirulent strain
(Table I). When analyzing the data on the identity of infected cells in lymphoid tissues of
the upper respiratory tract, we again observed that the most prominent target cells for
EHV1 were CD172a+ CML, irrespective of the strain used. This is in line with previous
studies, where EHV1-infected macrophages and lymphoblasts were detected in lymph
nodes after inoculation with EHV1 isolates from both an aborted fetus and a paralyzed
mare (Patel et al., 1982). Moreover, another EHV1 experimental inoculation study
showed that lymphocytes, monocytes, macrophages and plasma cells in interstitium and
lymph nodes all expressed EHV1 antigens, as determined by immunohistochemical
staining using anti-EHV1 polyclonal Abs (Kydd et al., 1994b). In the present study, the
majority of EHV1-positive cells was found in the marginal sinus of lymph nodes. At
those sites, transport is possible to extra follicular interdigitating cells in the lymph node
where both mature and virgin lymphocytes have access to antigen, hereby initiating
humoral immune responses (Tew et al., 1990). Surprisingly, and in contrast to what was
observed in nasal septum and nasopharynx, IgM+ B-lymphocytes were also an important
target cell for EHV1. In vitro it has been previously shown that a 6- to 14-fold increase of
EHV1-infected peripheral blood mononuclear cells (PBMC) could be obtained after in
vitro stimulation with proliferating drugs such as pokeweed mitogen (PWM),
concanavalin A (ConA), phytohaemagglutinin (PHA), or ionomycin and phorbol
dibutyrate (IONO/PDB) (van der Meulen et al., 2001). Together with the fact that B-
lymphocytes are known to undergo extensive proliferation in lymphoid follicles (Fu &
Chaplin, 1999), we would like to reason that the elevated percentage of EHV1-infected B-
lymphocytes observed in lymphoid tissues, is due to the proliferation stage of these
immune cells.
The onset of a cell-associated viremia could be detected as early as one day after
experimental inoculation with EHV1, indicating that EHV1-infected cells can enter the
blood stream very rapidly. Moreover, we identified these EHV1-positive cells as mainly
CD172a+ CML. Despite the fact that only 14 infected cells were found, we strongly feel

Replication kinetics of EHV1
82
that this low number is of relevance, as it is known that EHV1-induced viremia is
generally very low with numbers ranging from 1 to 10 positive cells/0.8 x 107 PBMC
(van der Meulen et al., 2006). Combining the results on the identity of EHV1-infected
cells in the connective tissues of the primary port of entry, the upper respiratory tract,
with the results on the identity of the EHV1-infected cells in the blood, we would like to
hypothesize that the CD172a+ cells we observed could be dendritic cells (DCs). The
reasoning behind such hypothesis is as follows. The marker DH59B that was used in the
present study to identify CD172a+ cells is a pan granulocyte/monocyte marker which has
been used in several studies as a DC marker (Ahn et al., 2002; Tumas et al., 1994). In
addition, there is evidence indicating that local tissue DCs are able to re-enter the
bloodstream, which might facilitate the spread of pathogens from tissue to tissue, carried
by DCs serving as Trojan horses (Randolph et al., 2008). Finally, DCs have been shown
to play an important role during the pathogenesis of alphaherpesviruses in general
(Bosnjak et al., 2005; Novak & Peng, 2005; Pollara et al., 2005), and for EHV1 in
specific, both murine and equine blood-derived DCs are susceptible to EHV1 in vitro
(Siedek et al., 1999; Steinbach et al., 1998). Unfortunately, little is known about equine
DCs in general and mucosal DCs in specific in this species, and more research is needed
to elucidate the role of DCs in EHV1 infection and/or other viral infections in equines.
Regardless, the results presented here are in striking contrast to another in vivo study,
where mainly blood T-lymphocytes were found to harbour EHV1 (Scott et al., 1983).
However, in the latter study, leukocyte populations were solely separated by glass-bead
columns and no direct characterization was performed. A possible explanation for the
discrepancy between cell types infected during cell-associated viremia could be the strain
used. In a previous study an American strain, isolated from an aborted fetus, was used
(Scott et al., 1983) whereas in the present study, we used two Belgian EHV1 strains.
Surprisingly, we observed CD172a+ cells as the main carrier cell type for both the
neurovirulent and the non-neurovirulent strain, indicating that the EHV1 strain does not
seem to correlate with a difference in cell tropism during viremia. This is again in contrast
to another study, where a difference in leukocyte tropism between neuro- and non-
neurovirulent EHV1 strains has been demonstrated (Goodman et al., 2007). Hereby it was
observed that the EHV1 strain Ab4, which has the neurovirulent genotype, mainly
infected CD4+ lymphocytes, whereas mutating this strain into the non-neurovirulent
genotype resulted in a preferential infection of monocytes and B-cells. However, these

Replication kinetics of EHV1
83
experiments were performed with in vitro infected PBMC, which most likely explains the
discrepancy with our in vivo data.
Acknowledgements
This research was supported by the Institute for the promotion of Innovation through
Science and Technology in Flanders (IWT-Vlaanderen). The authors like to thank C.
Boone and N. Monteiro for titrations, and C. Bracke for isolation of PBMC and
performing SN-tests.

Replication kinetics of EHV1
84
References
Ahn, J. S., Konno, A., Gebe, J. A., Aruffo, A., Hamilton, M. J., Park, Y. H. & Davis, W. C. (2002). Scavenger receptor cysteine-rich domains 9 and 11 of WC1 are
receptors for the WC1 counter receptor. J Leukoc Biol 72, 382-390.
Allen, G. P. & Breathnach, C. C. (2006). Quantification by real-time PCR of the
magnitude and duration of leucocyte-associated viraemia in horses infected with
neuropathogenic vs. non-neuropathogenic strains of EHV-1. Equine Vet J 38, 252-
257.
Allen, G. P. & Bryans, J. T. (1986). Molecular epizootiology, pathogenesis, and
prophylaxis of equine herpesvirus-1 infections. Prog Vet Microbiol Immunol 2,
78-144.
Bosnjak, L., Jones, C. A., Abendroth, A. & Cunningham, A. L. (2005). Dendritic cell
biology in herpesvirus infections. Viral Immunol 18, 419-433.
Bridges, C. G. & Edington, N. (1986). Innate immunity during Equid herpesvirus 1
(EHV-1) infection. Clin Exp Immunol 65, 172-181.
Cornick, J., Martens, J., Martens, R., Crandell, R., McConnell, S. & Kit, S. (1990). Safety and efficacy of a thymidine kinase negative equine herpesvirus-1 vaccine
in young horses. Can J Vet Res 54, 260-266.
Edington, N., Bridges, C. G. & Griffiths, L. (1989). Equine interferons following
exposure to equid herpesvirus-1 or -4. J Interferon Res 9, 389-392.
Edington, N., Bridges, C. G. & Patel, J. R. (1986). Endothelial cell infection and
thrombosis in paralysis caused by equid herpesvirus-1: equine stroke. Arch Virol
90, 111-124.
Fu, Y. X. & Chaplin, D. D. (1999). Development and maturation of secondary lymphoid
tissues. Annu Rev Immunol 17, 399-433.
Garré, B., Gryspeerdt, A., Croubels, S., De Backer, P. & Nauwynck, H. (2009). Evaluation of orally administered valacyclovir in experimentally EHV1-infected
ponies. Vet Microbiol 135, 214-221.
Glorieux, S., Favoreel, H. W., Meesen, G., de Vos, W., Van den Broeck, W. & Nauwynck, H. J. (2009). Different replication characteristics of historical
pseudorabies virus strains in porcine respiratory nasal mucosa explants. Vet
Microbiol 136, 341-346.
Goehring, L. S., van Winden, S. C., van Maanen, C. & Sloet van Oldruitenborgh-Oosterbaan, M. M. (2006). Equine herpesvirus type 1-associated
myeloencephalopathy in The Netherlands: a four-year retrospective study (1999-
2003). J Vet Intern Med 20, 601-607.
Goodman, L. B., Loregian, A., Perkins, G. A., Nugent, J., Buckles, E. L., Mercorelli,
B., Kydd, J. H., Palu, G., Smith, K. C., Osterrieder, N. & Davis-Poynter, N. (2007). A point mutation in a herpesvirus polymerase determines
neuropathogenicity. PLoS Pathog 3, 1583-1592.
Goodman, L. B., Wagner, B., Flaminio, M. J., Sussman, K. H., Metzger, S. M., Holland, R. & Osterrieder, N. (2006). Comparison of the efficacy of inactivated
combination and modified-live virus vaccines against challenge infection with
neuropathogenic equine herpesvirus type 1 (EHV-1). Vaccine 24, 3636-3645.
Greenwood, R. E. & Simson, A. R. (1980). Clinical report of a paralytic syndrome
affecting stallions, mares and foals on a thoroughbred studfarm. Equine Vet J 12,
113-117.

Replication kinetics of EHV1
85
Heldens, J. G., Hannant, D., Cullinane, A. A., Prendergast, M. J., Mumford, J. A.,
Nelly, M., Kydd, J. H., Weststrate, M. W. & van den Hoven, R. (2001). Clinical and virological evaluation of the efficacy of an inactivated EHV1 and
EHV4 whole virus vaccine (Duvaxyn EHV1,4). Vaccination/challenge
experiments in foals and pregnant mares. Vaccine 19, 4307-4317.
Jackson, T. A., Osburn, B. I., Cordy, D. R. & Kendrick, J. W. (1977). Equine
herpesvirus 1 infection of horses: studies on the experimentally induced
neurologic disease. Am J Vet Res 38, 709-719.
Kydd, J. H., Smith, K. C., Hannant, D., Livesay, G. J. & Mumford, J. A. (1994a). Distribution of equid herpesvirus-1 (EHV-1) in respiratory tract associated
lymphoid tissue: implications for cellular immunity. Equine Vet J 26, 470-473.
Kydd, J. H., Smith, K. C., Hannant, D., Livesay, G. J. & Mumford, J. A. (1994b). Distribution of equid herpesvirus-1 (EHV-1) in the respiratory tract of ponies:
implications for vaccination strategies. Equine Vet J 26, 466-469.
Matsumura, T., O'Callaghan, D. J., Kondo, T. & Kamada, M. (1996). Lack of
virulence of the murine fibroblast adapted strain, Kentucky A (KyA), of equine
herpesvirus type 1 (EHV-1) in young horses. Vet Microbiol 48, 353-365.
McCartan, C. G., Russell, M. M., Wood, J. L. & Mumford, J. A. (1995). Clinical,
serological and virological characteristics of an outbreak of paresis and neonatal
foal disease due to equine herpesvirus-1 on a stud farm. Vet Rec 136, 7-12.
Mumford, J. A. & Edington, N. (1980). EHV1 and equine paresis. Vet Rec 106, 277.
Novak, N. & Peng, W. M. (2005). Dancing with the enemy: the interplay of herpes
simplex virus with dendritic cells. Clin Exp Immunol 142, 405-410.
Nugent, J., Birch-Machin, I., Smith, K. C., Mumford, J. A., Swann, Z., Newton, J. R., Bowden, R. J., Allen, G. P. & Davis-Poynter, N. (2006). Analysis of equid
herpesvirus 1 strain variation reveals a point mutation of the DNA polymerase
strongly associated with neuropathogenic versus non-neuropathogenic disease
outbreaks. J Virol 80, 4047-4060.
Ostlund, E. N. (1993). The equine herpesviruses. Vet Clin North Am Equine Pract 9,
283-294.
Patel, J. R., Edington, N. & Mumford, J. A. (1982). Variation in cellular tropism
between isolates of equine herpesvirus-1 in foals. Arch Virol 74, 41-51.
Pollara, G., Kwan, A., Newton, P. J., Handley, M. E., Chain, B. M. & Katz, D. R. (2005). Dendritic cells in viral pathogenesis: protective or defective? Int J Exp
Pathol 86, 187-204.
Randolph, G. J., Ochando, J. & Partida-Sanchez, S. (2008). Migration of dendritic cell
subsets and their precursors. Annu Rev Immunol 26, 293-316.
Reed, L. J. & Muench, H. (1938). A simple method of estimating fifty per cent
endpoints. Am J Hyg 27, 493-497.
Scott, J. C., Dutta, S. K. & Myrup, A. C. (1983). In vivo harboring of equine
herpesvirus-1 in leukocyte populations and subpopulations and their quantitation
from experimentally infected ponies. Am J Vet Res 44, 1344-1348.
Siedek, E. M., Whelan, M., Edington, N. & Hamblin, A. (1999). Equine herpesvirus
type 1 infects dendritic cells in vitro: stimulation of T lymphocyte proliferation
and cytotoxicity by infected dendritic cells. Vet Immunol Immunopathol 67, 17-32.
Steinbach, F., Borchers, K., Ricciardi-Castagnoli, P., Ludwig, H., Stingl, G. & Elbe-Burger, A. (1998). Dendritic cells presenting equine herpesvirus-1 antigens
induce protective anti-viral immunity. J Gen Virol 79, 3005-3014.

Replication kinetics of EHV1
86
Stierstorfer, B., Eichhorn, W., Schmahl, W., Brandmuller, C., Kaaden, O. R. & Neubauer, A. (2002). Equine herpesvirus type 1 (EHV-1) myeloencephalopathy:
a case report. J Vet Med B Infect Dis Vet Public Health 49, 37-41.
Tew, J. G., Kosco, M. H., Burton, G. F. & Szakal, A. K. (1990). Follicular dendritic
cells as accessory cells. Immunol Rev 117, 185-211.
Tumas, D. B., Brassfield, A. L., Travenor, A. S., Hines, M. T., Davis, W. C. & McGuire, T. C. (1994). Monoclonal antibodies to the equine CD2 T-lymphocyte
marker, to a pan-granulocyte monocyte marker and to a unique pan-B lymphocyte
marker. Immunobiology 192, 48-64.
Van de Walle, G. R., Goupil, R., Wishon, C., Damiani, A., Perkins, G. A. & Osterrieder, N. (2009). A Single-Nucleotide Polymorphism in a Herpesvirus
DNA Polymerase Is Sufficient to Cause Lethal Neurological disease. J Infect Dis
200, 20-25.
van der Meulen, K., Caij, B., Pensaert, M. & Nauwynck, H. (2006). Absence of viral
envelope proteins in equine herpesvirus 1-infected blood mononuclear cells during
cell-associated viremia. Vet Microbiol 113, 265-273.
van der Meulen, K., Vercauteren, G., Nauwynck, H. & Pensaert, M. (2003a). A local
epidemic of equine herpesvirus 1-induced neurological disorders in Belgium.
Vlaams Diergen Tijds 72, 366-372.
van der Meulen, K. M., Gryspeerdt, A. C., Vandekerckhove, A. A., Garre, B. A. & Nauwynck, H. J. (2007). The protective properties of vaccination against equine
herpesvirus 1-induced viremia, abortion and nervous system disorders. Vlaams
Diergen Tijds 76, 186-194.
van der Meulen, K. M., Nauwynck, H. J., Buddaert, W. & Pensaert, M. B. (2000). Replication of equine herpesvirus type 1 in freshly isolated equine peripheral
blood mononuclear cells and changes in susceptibility following mitogen
stimulation. J Gen Virol 81, 21-25.
van der Meulen, K. M., Nauwynck, H. J. & Pensaert, M. B. (2001). Mitogen
stimulation favours replication of equine herpesvirus-1 in equine blood
mononuclear cells by inducing cell proliferation and formation of close
intercellular contacts. J Gen Virol 82, 1951-1957.
van der Meulen, K. M., Nauwynck, H. J. & Pensaert, M. B. (2003b). Absence of viral
antigens on the surface of equine herpesvirus-1-infected peripheral blood
mononuclear cells: a strategy to avoid complement-mediated lysis. J Gen Virol 84,
93-97.
van Maanen, C. (2002). Equine herpesvirus 1 and 4 infections: an update. Vet Q 24, 58-
78.

Expression of late proteins is hampered
87
CHAPTER IV
EXPRESSION OF LATE VIRAL PROTEINS IS
HAMPERED IN INFECTED NASAL MUCOSAL
LEUKOCYTES BUT NOT IN EPITHELIAL CELLS
DURING EARLY PATHOGENESIS OF EQUINE
HERPESVIRUS TYPE 1 (EHV1) INFECTION
Adapted from:
The Veterinary Journal (conditionally accepted, 2011)
Annick C. Gryspeerdt, A.P. Vandekerckhove, H. Bannazadeh Baghi, G.R. Van de Walle,
H.J. Nauwynck


Expression of late proteins is hampered
89
SUMMARY
Equine herpesvirus 1 (EHV1) starts its replication by infection in the epithelial cells of
the upper respiratory tract (URT), where after the virus invades the lamina propria of the
URT using single mononuclear cells, hereby reaching the bloodstream which allows
further spread of this virus throughout the body. In addition, EHV1 has developed various
strategies to escape from immune effector mechanisms. One of these strategies is to
interfere with antibody-dependent cell lysis by hampering proper viral protein expression
on the cell surface.
Because epithelial and mononuclear cells are crucial cell populations in the early
pathogenesis of EHV1, the present study aimed to evaluate the expression of the late viral
proteins gB, gC, gD and gM in these type of cells, using several experimental systems.
Firstly, epithelial-like rabbit kidney (RK13) cells and peripheral blood mononuclear cells
(PBMC) were infected with EHV1 in vitro. Secondly, the recently established ex vivo
system of equine nasal mucosal explants was used and thirdly, an in vivo evaluation was
performed by means of nasal mucosa tissues of experimentally EHV1-infected ponies.
Our salient findings were that the late proteins gB, gC, gD and gM were clearly expressed
in almost all late-infected epithelial cells, whereas the expression pattern was totally
different in infected mononuclear cells: gB and gM were expressed in 60 to 90% of these
cells, gC and gD in only 20%.
These results show that the expression of late viral proteins during the early
pathogenesis of EHV1 depends upon the cell type infected. Furthermore, this study
underlines the use of nasal mucosal explants as a valuable alternative to experimental in
vivo studies in the horse.

Expression of late proteins is hampered
90
1 INTRODUCTION
Equine herpesvirus 1 (EHV1), a member of the Alphaherpesvirinae, is an ubiquitous
equine respiratory pathogen, affecting horses worldwide (Allen & Bryans, 1986;
Brosnahan & Osterrieder, 2009). After an initial replication in the epithelial cells of the
upper respiratory tract (URT), EHV1 can spread through the basement membrane (BM)
carried by single infected mononuclear immune cells, hereby reaching the underlying
connective tissue (Vandekerckhove et al., 2010). Via a cell-associated viremia in
peripheral blood mononuclear cells (PBMC), EHV1 is then transported to its internal
target organs, the pregnant uterus and the central nervous system, where viral replication
in endothelial cells can cause abortion, neonatal foal death and nervous system disorders
through vascular damage (Smith & Borchers, 2001; Smith et al., 1996). As a result of the
short-lived immunity following an EHV1 infection, viral replication in the respiratory
tract can re-occur as soon as two months and viremia can be detected as early as six
months after primary infection (Edington & Bridges, 1990; Gibson et al., 1992). High
levels of circulating EHV1-specific antibodies are still present when re-infection with
EHV1 occurs, suggesting that EHV1 recognition by the immune system is seriously
hampered (Burrows et al., 1984; Gleeson & Coggins, 1980; Heldens et al., 2001;
Mumford et al., 1987).
The genome of EHV1 encodes at least 12 known glycoproteins, designated
glycoprotein B (gB), gC, gD, gE, gG, gH, gI, gK, gL, gM, gN and gp2. Herpesviral
glycoproteins are present in the viral envelope as distinct, projected spikes and because
they are involved in various steps of the virus infection cycle, such as attachment,
penetration, egress and cell to cell spread, they represent major determinants of
pathogenicity, virulence and host cell tropism (Spear & Longnecker, 2003). For example,
glycoprotein C and gM are non-essential glycoproteins, with gC being involved in the
first unstable binding of the virus to glycosaminoglycan moieties of cell surface heparin
sulphate and in the process of egress (Osterrieder, 1999; Sugahara et al., 1997), and gM
contributing to virus penetration and cell-to-cell spread of the virus (Osterrieder et al.,
1996). The essential protein gD interacts with a putative entry receptor to attain a stable
binding (Csellner et al., 2000; Frampton et al., 2005) and is furthermore also involved in
cell-to-cell spread of the virus (Csellner et al., 2000; Wellington et al., 1996a). The
essential protein gB is involved in the penetration of the virus into the host cell and cell-

Expression of late proteins is hampered
91
associated spread of the virus to uninfected neighbouring cells (Neubauer et al., 1997;
Wellington et al., 1996b).
In general, upon infection of a cell with an alphaherpesvirus, these viral envelope
proteins are synthesised and incorporated into various cellular membranes (Mettenleiter,
2002). The presence of viral proteins in the plasma membrane of infected cells renders
them susceptible to antibody (Ab)-dependent cell lysis, through binding of virus-specific
Abs on viral proteins. Subsequently, infected cells are destructed by phagocytic cells and
natural killer cells or via activation of the complement cascade (Hansch et al., 1997;
Hirsch, 1982; Janeway et al., 2001). Several herpesviruses have developed strategies to
interfere with immune effector mechanisms and EHV1 is no exception in this regard (van
der Meulen et al., 2006b). One strategy of EHV1 is to interfere with Ab-dependent cell
lysis by hampering envelope protein expression on the cell surface. An in vitro study by
van der Meulen et al. (2003b) demonstrated that approximately 70% of all EHV1-infected
PBMC did not express viral envelope proteins on their surface, despite a clear
intracellular expression (van der Meulen et al., 2003b). This absence of viral proteins on
the surface of these infected PBMC was shown to render them insensitive to Ab-
dependent complement-mediated lysis and Ab-dependent cell-mediated cytotoxicity (van
der Meulen et al., 2003b). Importantly, in an in vivo study of experimentally inoculated
ponies, 98% of the infected PBMC collected during cell-associated viremia did not show
viral envelope proteins on their plasma membrane, nor were they found intracellularly,
and only immediate early and at least some early proteins were present (van der Meulen
et al., 2006a). Based on these results, it was hypothesized that EHV1-infected PBMC are
at an early stage of infection during viremia and that further replication steps, including
proper expression of late proteins, only occurs upon adhesion to endothelial cells at the
target organs (van der Meulen et al., 2006b).
The main purpose of the present study was to evaluate the expression of several late
viral envelope proteins, including gB, gC, gD and gM in the different cell types which
become infected in the URT, a first step during the early pathogenesis. Two important
cell types during early pathogenesis were analysed in three different model systems: in
vitro with the standard epithelial RK13 cell line and PBMC, ex vivo by means of the
recently established equine nasal mucosal explant system and finally in vivo, with tissues
collected from experimentally infected ponies.

Expression of late proteins is hampered
92
2 MATERIALS AND METHODS
2.1 Virus
Two different strains were used in this study to take into account a possible influence
of the neurovirulent/non-neurovirulent nature of EHV1. On the one hand the Belgian
EHV1 strain 03P37 was used, isolated from the blood of a paralytic horse (Garré et al.,
2009; van der Meulen et al., 2003a), and on the other hand the Belgian strain 97P70,
isolated from an aborted fetus (van der Meulen et al., 2006a). These strains were typed in
their DNA polymerase in cooperation with the Animal Health Trust in the United
Kingdom (Nugent et al., 2006) and based on their origin and genotyping, these strains are
referred to in this paper as neurovirulent (D752) and non-neurovirulent (N752) respectively.
Virus stocks used for inoculation were at the 6th
passage; 2 passages in rabbit kidney cells
(RK13) and 4 subsequent passages in equine embryonic lung cells (EEL). Both viruses
have previously been shown to display virtually identical in vitro single-step growth
properties (Gryspeerdt et al., 2010).
2.2 Antibodies
For the detection of the late proteins gB, gC, gD and gM, specific antibodies were
used: mouse monoclonal antibodies (mAbs) 4B6 against gB (Osterrieder et al., 1995),
1B6 against gC (Huemer et al., 1995), 107D1D against gD (a kind gift from Merial,
France), and the monospecific anti-gM mouse antiserum F6 against gM (Osterrieder et
al., 1996).
To identify EHV1-infected cells, two antibodies were used: either rabbit polyclonal
antibodies against the immediate early (IE) protein (Caughman et al., 1988), or horse
polyclonal anti-EHV1 antibodies. The latter antibodies were obtained by
hyperimmunisation of a horse (van der Meulen et al., 2003b). The polyclonal antibodies
were purified on a protein G column and subsequently biotinylated (Amersham
International, Buckinghamshire, UK). To ensure that these antibodies could recognise the
late proteins of EHV1, transfection studies were performed. To this end, RK13 cells were
transfected with 1 µg of plasmid DNA expressing gB, gC, gD or gM (a kind gift from
Prof. Osterrieder, Freie Universitaet Berlin), using lipofectamin (Invitrogen). After 48h,

Expression of late proteins is hampered
93
RK13 cells were fixed and stained by double immunofluorescence stainings as described
below. All transfected cells, as identified by a positive staining for each late protein
respectively, were also positive for the anti-EHV1 polyclonal antibodies, indicating that
these EHV1-specific antibodies are able to recognise all late proteins evaluated in the
present study. In addition, kinetic infection studies in RK13 cells showed that these
polyclonal antibodies stain all infected cells from 24 hpi onwards when the late
glycoproteins are fully expressed, indicating that these antibodies are a useful tool to
detect cells in a late stage of infection. These horse polyclonal anti-EHV1 antibodies are
especially advantageous to evaluate infected cells in the nasal mucosa because these cells
are at different stages of infection and they always identify cells in the late stage of
infection. Rabbit polyclonal antibodies against the IE protein cannot discriminate between
cells early in infection from cells late in infection with an abortive infection.
2.3 Expression of late proteins
(i) In RK13 cells and PBMC.
Cells of the epithelial-like rabbit kidney (RK13) cell line were seeded on glass inserts
in 24-well culture plates (Nunc A/S) at a density of 300 000 cells/well and were further
cultured in growth medium (MEM supplemented with 100 U/ml penicillin, 0.1 mg/ml
streptomycin, 0.1 mg/ml kanamycin, 0.3 mg/ml glutamine and 5% foetal calf serum) at
37°C for 24 hours . Then, cultures were infected with either neurovirulent strain 03P37 or
non-neurovirulent strain 97P70 at a multiplicity of infection (MOI) of 10. After 1 hour,
virus was removed by rinsing the cells twice with culture medium and after 0, 4, 8, 13 and
24 subsequent hours of incubation, the cells were fixed with methanol for 20 minutes at -
20°C and stored at -70°C. Infected cells were stained as described below and were
analyzed by confocal laser scanning microscopy (Leica TCS SP2 laser scanning spectral
confocal system, Leica Microsystems GmbH, Wetzlar) and the Leica confocal software.
Hereby, individual cells were counted from their appearance (3 hpi) until the appearance
of plaques (8hpi). Newly infected individual cells present after 13 hpi were not taken into
account as they represent a second replication cycle.
Blood samples were collected from three different, healthy adult horses by jugular
venipuncture and collected on heparin (Leo). PBMC were isolated by density
centrifugation on Ficoll-Paque (Pharmacia Biotech AB), washed 3 times with PBS and

Expression of late proteins is hampered
94
resuspended in complete medium containing 2/1 MEM/RPMI 1640, supplemented with
10% fetal calf serum (Grainer), 0.05 mM 2-mercapto-ethanol (Gibco), 100 U/ml
penicillin (Continental Pharma), 0.1 mg/ml streptomycin (Certa), 0.1 mg/ml kanamycin
(Kela), 0.3 mg/ml glutamin (BDH Biochemical), 1% non-essential amino-acids 100x
(Gibco) and 1 mM sodium pyruvate (Gibco). After 24 hours of cultivation, PBMC were
inoculated at a MOI of 10 with either neurovirulent strain 03P37 or non-neurovirulent
strain 97P70. After 1 hour of incubation at 37 °C, cells were washed thoroughly with
RPMI and cultured in medium supplemented with 10 U/ml heparin (Leo). Cell smear
samples were made 48 hours post inoculation (pi) and fixed in acetone for 20 minutes.
(ii) In ex vivo nasal mucosal explants.
The cultivation and infection of nasal mucosal explants was performed exactly as
previously described (Vandekerckhove et al., 2010). In brief, immediately after slaughter,
the head was removed from the carcass and longitudinally sawn into 2 equal sections.
Tissue from the deep intranasal part of the septum of 3 different horses was collected. The
tissues were transported on ice in phosphate buffered saline (PBS), supplemented with 1
µg/ml gentamycin, 1 mg/ml streptomycin, 1000 U/ml penicillin, 1 mg/ml kanamycin and
5 µg/ml amphotericin, to the laboratory. Mucosal explants were stripped from the surface
of the different tissues by use of surgical blades (Swann-Morton). The stripped mucosa
was divided into equal explants of 25 mm2 and placed epithelium upwards on fine-
meshed gauze for culture at an air-liquid interface. Only a thin film of serum-free medium
DMEM (50%)/RPMI (50%) supplemented with 0.3 mg/ml glutamine (BHD
Biochemical), 1 µg/ml gentamycin (Gibco), 0.1 mg/ml streptomycin (Certa) and 100
U/ml penicillin (Continental Pharma)) covered the explants, thereby mimicking the
air/liquid interface found in the respiratory tract of the living animal. Explants were
maintained at 37°C in an atmosphere containing 5% CO2. Explants were inoculated after
24 h of culture by immersion of the explant in 1 ml of inoculum, containing 106.5
TCID50
of either EHV1 strain for 1 h at 37°C and 5% CO2. After incubation, explants were
washed twice with warm medium and transferred back to their gauze. At 48 hours post
inoculation (pi), explants were collected, embedded in methylcellulose medium
(Methocel® MC, Sigma-Aldrich, St.Louis) and samples were frozen at -70 °C.

Expression of late proteins is hampered
95
(iii) In nasal mucosa of experimentally in vivo infected ponies. Six EHV-negative
ponies (SN titer <2 and IPMA titer <10) were divided in two groups of three and
inoculated either with strain 97P70 or 03P37 as described previously (Gryspeerdt et al.,
2010). In brief, ponies were inoculated oronasally with 20 ml of a virus suspension
containing 106.5
tissue culture infectious dose 50 (TCID50). Ten ml of the virus suspension
was administered intranasally (5 ml per nostril) using a thin probe and 10 ml was
inoculated orally with a syringe. On two dpi, all ponies were euthanized with an overdose
of Natriumpentobarbital® (Kela, Hoogstraten, Belgium), and the intermedial part of the
nasal septum was collected. Tissue samples were immediately frozen with
methylcellulose medium (Methocel®MC, Sigma-Aldrich, St. Louis, MO, USA) at -70°C.
This experimental design was approved by the local ethical committee of the Faculty of
Veterinary Medicine, Ghent University, Belgium.
2.4 Double immunofluorescence stainings
To visualise the presence of gB, gC, gD and gM in infected cells, the specific
antibodies (mAbs) against these glycoproteins were used in a first step, followed by
incubation with secondary Texas Red®-labelled goat anti-mouse antibodies (Molecular
Probes, Eugene, OR, USA). For the identification of EHV1-infected cells, either the
rabbit polyclonal antibodies against the IE protein (RK13 cells), or the biotinylated horse
polyclonal anti-EHV1 antibodies (PBMC, nasal mucosa) were used. Subsequently,
samples were incubated with secondary FITC®-labelled goat anti-rabbit antibodies
(Molecular Probes, Eugene, OR, USA) for the polyclonal rabbit anti-IE antibody or
streptavidin-FITC® (Molecular Probes, Eugene, OR, USA) for the biotinylated polyclonal
anti-EHV1 antibody. Antibodies were incubated for 1 h at 37 °C and 5 % CO2 and
washed thoroughly with phosphate buffered saline (PBS) in between staining steps. As
negative controls, (i) stainings were performed on uninfected RK13, PBMC and nasal
mucosa and (ii) isotype specific control antibodies on infected cells and tissues were
included. All cryosections were analyzed using confocal microscopy (Leica TCS SP2
Laser scanning spectral confocal system, Leica Microsystems GmbH, Wetzlar, Germany).

Expression of late proteins is hampered
96
3 RESULTS
3.1 Late proteins gB, gC, gD and gM are properly expressed in in vitro infected
epithelial-like RK13 cells
In a first experiment, RK13 cells were used to analyse the expression kinetics of the
viral EHV1 glycoproteins gB, gC, gD and gM during an EHV1 infection. To this end, the
immediate early protein (IE) was used to identify EHV1-infected cells and double
immunofluorescent stainings were performed to detect the viral glycoproteins gB, gD, gC
and gM. EHV1-infected cells were observed as early as 1 hpi and their number increased
over time (data not shown). From 8 hpi, all late proteins tested started to be expressed in
individual EHV1-infected cells irrespective of the strains used, with gB and gM being
expressed in a larger percentage of the infected cells than gC and gD (Figure 1).
Expression of these late proteins in infected cells further increased over time and finally
at 24 hpi, the expression of gB, gC, gD and gM was found in all infected cells. This time
point co-incided with the time point when the polyclonal anti-EHV1 antibodies recognise
infected cells and therefore, this Ab was used in all further experimentation to identify
late infected cells.

Expression of late proteins is hampered
97
Figure 1. Kinetic study of the expression of late proteins gB, gC, gD and gM in EHV1-infected RK13
cells inoculated with neurovirulent strain 03P37 (A) and non-neurovirulent strain 97P70 (B). All data
represent means + SD of triplicate independent experiments. EHV1-infected cells were firstly incubated
with mouse monoclonal antibodies against gB, gC, or gD and a monospecific mouse antiserum against gM,
followed by Texas Red®-labelled goat anti-mouse antibodies. Secondly, they were incubated with a
polyclonal rabbit anti-IE antibody, followed by FITC®-labelled goat anti-rabbit antibodies.
3.2 Expression of the late proteins gC and gD is hampered in in vitro EHV1-infected
PBMC
Next, we wanted to evaluate the expression of the late proteins gB, gC, gD and gM in
a cell type which is also relevant during the pathogenesis of EHV1 infection, namely
peripheral blood mononuclear cells (PBMC). PBMC were isolated from the blood and
infected in vitro with the neurovirulent or non-neurovirulent EHV1 strain. At 48 hpi,
A
B
0
20
40
60
80
100
0 5 10 15 20 25
Hours post inoculation
Exp
ress
ion
of
late
pro
tein
sin
IE-p
osi
tiv
eR
K1
3 c
ells
(%)
gB
gC
gD
gM
0
20
40
60
80
100
0 5 10 15 20 25
Hours post inoculation
gB
gC
gD
gM
Exp
ress
ion
of
late
pro
tein
sin
IE-p
osi
tiv
eR
K1
3 c
ells
(%)
A
B
0
20
40
60
80
100
0 5 10 15 20 25
Hours post inoculation
Exp
ress
ion
of
late
pro
tein
sin
IE-p
osi
tiv
eR
K1
3 c
ells
(%)
gB
gC
gD
gM
0
20
40
60
80
100
0 5 10 15 20 25
Hours post inoculation
gB
gC
gD
gM
Exp
ress
ion
of
late
pro
tein
sin
IE-p
osi
tiv
eR
K1
3 c
ells
(%)

Expression of late proteins is hampered
98
infected cells were identified using the polyclonal anti-EHV1 Ab and expression of the
late proteins gB, gC, gD and gM was evaluated in these infected cells. For both strains,
expression of the late proteins gB and gM was almost 100% in infected cells (Figure 2A).
In contrast, expression of gC and gD was only seen in 15 to 25% of EHV1-infected
PBMC (Figure 2A). These results indicate that expression of certain late proteins which
are normally expressed in infected rabbit epithelial cells at 24 hpi, are impaired in their
expression in leukocytes at 48 hpi.
3.3 Expression of late proteins gC and gD is also hampered in EHV1-infected
leukocytes but not epithelial cells from nasal mucosa in explants and experimentally
infected ponies
To assess the relevance of the observations which were made with the epithelial-like
RK13 cells and PBMC, we repeated the double immunofluorescence stainings in
cryosections of (i) infected nasal mucosal explants and (ii) nasal tissue of experimentally
infected ponies. Previous work in our laboratory has shown that in both models, the nasal
epithelial cells become infected with EHV1. In addition, single EHV1-infected cells are
also observed, which are mainly present under the BM in connective tissue and which
were identified as leukocytes (Gryspeerdt et al., 2010; Vandekerckhove et al., 2010).
In nasal explants and in nasal tissues of in vivo inoculated horses, infected epithelial
cells showed a full expression of late proteins tested (Figure 3), which is in line with what
was observed in RK13 cells at 24 hpi. Individual infected leukocytes, located in the
connective tissue below the BM showed the same tendency as in vitro infected PBMC.
Hereby, around 80% to 90% of late infected cells showed expression of the late proteins
gB and gM in nasal explants and in vivo harvested nasal tissues, whereas the expression
of the late proteins gC and gD in these EHV1-infected leukocytes fluctuated around 10 to
15% in in vivo harvested nasal tissues and nasal mucosal explants respectively (Figure 2B
and 2C). No clear differences in expression levels were found in cells infected with either
the neurovirulent 03P37 or the non-neurovirulent 97P70 (Figure 2B and 2C).

Expression of late proteins is hampered
99
Figure 2. Expression of late proteins gB, gC, gD and gM in late infected leukocytes at 48 hours post
inoculation with neurovirulent strain 03P37 and non-neurovirulent strain 97P70 in three different
systems: (A) in vitro infected PBMC, (B) nasal mucosal explants and (C) nasal mucosa of experimentally
infected ponies. All data represent means + SD of triplicate independent experiments. Firstly, samples were
incubated with mouse monoclonal antibodies against gB, gC, gD or gM, followed by Texas Red®-labelled
goat anti-mouse antibodies. Secondly, they were incubated with the polyclonal horse anti-EHV1 antibody,
followed by streptavidin-FITC®.
A
B
C
0
10
20
30
40
50
60
70
80
90
100
gB gC gD gM
Pre
sence
of
late
pro
tein
sin
in
vit
ro
infe
cted
PB
MC
(%
)
neurovirulent
non-neurovirulent
neurovirulent
non-neurovirulent
0
10
20
30
40
50
60
70
80
90
100
gB gC gD gM
Pre
sence
of
late
pro
tein
sin
sin
gle
EH
V1
-infe
cte
d
cel
lsin
nas
alex
pla
nts
(%)
0
10
20
30
40
50
60
70
80
90
100
gB gC gD gM
Pre
sence
of
late
pro
tein
sin
sin
gle
EH
V1
-infe
cted
cell
sin
nas
alti
ssue
(%)

Expression of late proteins is hampered
100
Figure 3. Expression of late proteins gB, gC, gD and gM in late infected epithelial cells. Since results were
virtually identical for all three systems and both strains tested, only immunofluorescence pictures of plaques in
in vitro infected nasal mucosal tissue is shown. First, a staining with the biotinylated equine polyclonal anti-
EHV1 IgG, followed by streptavidin-FITC® was performed (A). Second, a staining with specific mouse
antibodies against gB, gC, gD or gM followed by secondary Texas Red®
-labelled goat anti-mouse antibodies
was performed (B).
A
B
A
B

Expression of late proteins is hampered
101
4 DISCUSSION
Equine herpesvirus 1 (EHV1) starts its pathogenesis in the upper respiratory tract
where primary replication occurs and results in upper respiratory tract disease. After
invasion through the basement membrane (BM) barrier, the virus enters blood vessels and
rapidly disseminates in the body via a cell-associated viremia. Recent research has shown
that EHV1 replicates in epithelial cells by means of plaque formation and in migrating
individual cells, allowing EHV1 to cross the BM. The latter cells were identified as cells
of the monocytic lineage, T-lymphocytes and a small amount of B-lymphocytes
(Gryspeerdt et al., 2010; Kydd et al., 1994; Vandekerckhove et al., 2010). Hence, two
main cell populations appear to play a crucial role in the early pathogenesis of EHV1,
namely epithelial cells on the one hand and leukocytes on the other hand.
In the present study, we studied the expression of the late envelope glycoproteins
(g)B, gC, gD and gM in rabbit and horse epithelial cells as well as equine leukocytes, in
three frequently used EHV1 infection models. Hereby, our salient findings were that in
epithelial cells, a complete replication cycle of EHV1 was seen, with expression of all
four tested glycoproteins, gB, gC, gD and gM in 100% of the infected cells. This,
however, was in striking contrast to EHV1-infected leukocytes. In this cell type,
expression of the late proteins gB and gM was observed in 60 to 90% of infected cells,
whereas expression of the late proteins gC and gD never exceeded 20% of EHV1-infected
cells. As these results were consistent in vitro as well as in vivo infected cells, we
concluded that this phenomenon is truly cell-type dependent and not just merely a result
of the experimental set-up used.
A decreased expression of late proteins in certain cell types has already been
described for another herpesvirus, the human cytomegalovirus (HCMV). For this human
herpesvirus, viral gene expression in placental trophoblasts was shown to be restricted to
immediate-early or early gene products during HCMV infection (Sinzger et al., 1993;
Toth et al., 1995). In another study, where a human trophoblast continuous cell line was
used, expression of immediate-early, early and late genes was only delayed when
compared to expression in a fibroblastic continuous cell line (Halwachs-Baumann et al.,
1998). Hence, these studies indicate the importance of the cell type used when evaluating
protein expression after viral infection. For EHV1, quantitative dot blot hybridization
experiments revealed that expression of the late genes gD and gK was significantly lower

Expression of late proteins is hampered
102
in placental trophoblast cells than in foetal epithelial lung tissue (Kimura et al., 2004).
This low level of late gene transcription was linked with a limited production of viral
progeny and the lack of immunoreactivity for viral antigen in trophoblasts infected with
EHV1. However, all these studies have studied EHV1 protein expression in the later
stages of pathogenesis at the secondary replication sites, and therefore, our study is the
first to describe these events of decreased late protein expression in certain cell types
during the early stages of pathogenesis at the upper respiratory tract. The underlying
mechanism responsible for this decreased expression of EHV1 late proteins in certain cell
types, however, remains unclear. These findings could be an explanation why no plaque
formation in the connective tissue of the upper respiratory tract could be found in
previous studies (Gryspeerdt et al., 2010; Vandekerckhove et al., 2010), as gD is an
essential glycoprotein for virus spread and was found to be present in only very low
amounts of individual infected PBMC in the present study.
When combining our present results with data from previously performed studies in
the authors’ laboratory (van der Meulen et al., 2006a; van der Meulen et al., 2003b), it
could be hypothesized that EHV1-infected leukocytes which show a hampered expression
of late proteins, are the cells reaching the blood stream and subsequently causing a cell-
associated viremia. This is supported by the results of van der Meulen et al. (2006),
stating that 98% of EHV1-infected PBMC present in the blood during cell-associated
viremia in experimentally inoculated ponies lacked expression of certain late viral
envelope proteins on their cell surface. They did, however, show expression of the
immediate early protein and at least one early protein, ICP22, indicating that these cells
were truly infected (van der Meulen et al., 2006a). So one could hypothesize that these
cells are in an early stage of infection and that some activation signals need to trigger the
expression of envelope proteins on the cell surface to free the way for transmission of
infectious virus at the secondary replication sites. Such hypothesis has already been put
forward a long time ago by Scott et al. (1983). They described that although virus could
be recovered after cocultivation of intact leukocytes with a permissive cell line, no
infectious virus could be found after disruption of these cells (Scott et al., 1983). All this
indicates that EHV1-infected leukocytes exist in a non-infective or subvirion form.
Indeed, an in vitro study showed that activation of adhesion molecules in endothelial cells
of the nasal mucosa, and the reproductive tract, is a key step in transferring virus from
infected leukocytes to endothelial cells (Smith et al., 2002; Smith et al., 2001).

Expression of late proteins is hampered
103
In conclusion, although this is only a basic study where cells in a later stage of
infection were studied, these results indicate that the expression of certain viral proteins
during the early pathogenesis of EHV1 depends upon the cell-type infected. Furthermore,
this study underlines again the use of nasal mucosal explants as a valuable alternative to
experimental in vivo studies in the horse.
ACKNOWLEDGEMENTS
This research was supported by the Institute for the promotion of Innovation through
Science and Technology in Flanders (IWT-Vlaanderen). We would like to thank Prof. Dr.
N. Osterrieder for kindly providing the plasmids used in transfection studies.

Expression of late proteins is hampered
104
References
Allen, G. P. & Bryans, J. T. (1986). Molecular epizootiology, pathogenesis, and
prophylaxis of equine herpesvirus-1 infections. Prog Vet Microbiol Immunol 2,
78-144.
Brosnahan, A. J. & Osterrieder, K. (2009). Equine herpesvirus-1: a review and update.
Equine Vet J in press.
Burrows, R., Goodridge, D. & Denyer, M. S. (1984). Trials of an inactivated equid
herpesvirus 1 vaccine: challenge with a subtype 1 virus. Vet Rec 114, 369-374.
Caughman, G. B., Robertson, A. T., Gray, W. L., Sullivan, D. C. & O'Callaghan, D. J. (1988). Characterization of equine herpesvirus type 1 immediate early proteins.
Virology 163, 563-571.
Csellner, H., Walker, C., Wellington, J. E., McLure, L. E., Love, D. N. & Whalley, J. M. (2000). EHV-1 glycoprotein D (EHV-1 gD) is required for virus entry and
cell-cell fusion, and an EHV-1 gD deletion mutant induces a protective immune
response in mice. Arch Virol 145, 2371-2385.
Edington, N. & Bridges, C. G. (1990). One way protection between equid herpesvirus 1
and 4 in vivo. Research in veterinary science 48, 235-239.
Frampton, A. R., Jr., Goins, W. F., Cohen, J. B., von Einem, J., Osterrieder, N., O'Callaghan, D. J. & Glorioso, J. C. (2005). Equine herpesvirus 1 utilizes a
novel herpesvirus entry receptor. J Virol 79, 3169-3173.
Garré, B., Gryspeerdt, A., Croubels, S., De Backer, P. & Nauwynck, H. (2009). Evaluation of orally administered valacyclovir in experimentally EHV1-infected
ponies. Vet Microbiol 135, 214-221.
Gibson, J. S., Slater, J. D., Awan, A. R. & Field, H. J. (1992). Pathogenesis of equine
herpesvirus-1 in specific pathogen-free foals: primary and secondary infections
and reactivation. Arch Virol 123, 351-366.
Gleeson, L. J. & Coggins, L. (1980). Response of pregnant mares to equine herpesvirus
1 (EHV1). Cornell Veterinarian 70, 391-400.
Gryspeerdt, A. C., Vandekerckhove, A. P., Garre, B., Barbe, F., Van de Walle, G. R. & Nauwynck, H. J. (2010). Differences in replication kinetics and cell tropism
between neurovirulent and non-neurovirulent EHV1 strains during the acute phase
of infection in horses. Vet Microbiol 142, 242-253.
Halwachs-Baumann, G., Wilders-Truschnig, M., Desoye, G., Hahn, T., Kiesel, L., Klingel, K., Rieger, P., Jahn, G. & Sinzger, C. (1998). Human trophoblast cells
are permissive to the complete replicative cycle of human cytomegalovirus. J
Virol 72, 7598-7602.
Hansch, G. M., Rother, K. O. & Till, G. O., (eds) (1997). The Complement System.
Berlin, Heidelberg: Springer-Verlag.
Heldens, J. G., Hannant, D., Cullinane, A. A., Prendergast, M. J., Mumford, J. A.,
Nelly, M., Kydd, J. H., Weststrate, M. W. & van den Hoven, R. (2001). Clinical and virological evaluation of the efficacy of an inactivated EHV1 and
EHV4 whole virus vaccine (Duvaxyn EHV1,4). Vaccination/challenge
experiments in foals and pregnant mares. Vaccine 19, 4307-4317.
Hirsch, R. L. (1982). The complement system: its importance in the host response to
viral infection. Microbiol Rev 46, 71-85.

Expression of late proteins is hampered
105
Huemer, H. P., Nowotny, N., Crabb, B. S., Meyer, H. & Hubert, P. H. (1995). gp13
(EHV-gC): a complement receptor induced by equine herpesviruses. Virus
Research 37, 113-126.
Janeway, C. A., Travers, P., Walport, M. & Shlomchik, M., (eds) (2001). The
humoral immune response. New York, USA: Garland Publishing.
Kimura, T., Hasebe, R., Mukaiya, R., Ochiai, K., Wada, R. & Umemura, T. (2004). Decreased expression of equine herpesvirus-1 early and late genes in the placenta
of naturally aborted equine fetuses. Journal of Comparative Pathology 130, 41-47.
Kydd, J. H., Smith, K. C., Hannant, D., Livesay, G. J. & Mumford, J. A. (1994). Distribution of equid herpesvirus-1 (EHV-1) in respiratory tract associated
lymphoid tissue: implications for cellular immunity. Equine Vet J 26, 470-473.
Mettenleiter, T. C. (2002). Herpesvirus assembly and egress. J Virol 76, 1537-1547.
Mumford, J. A., Rossdale, P. D., Jessett, D. M., Gann, S. J., Ousey, J. & Cook, R. F. (1987). Serological and virological investigations of an equid herpesvirus 1 (EHV-
1) abortion storm on a stud farm in 1985. Journal of Reproduction and Fertility,
supplement 35, 509-518.
Neubauer, A., Braun, B., Brandmuller, C., Kaaden, O. R. & Osterrieder, N. (1997). Analysis of the contributions of the equine herpesvirus 1 glycoprotein gB
homolog to virus entry and direct cell-to-cell spread. Virology 227, 281-294.
Nugent, J., Birch-Machin, I., Smith, K. C., Mumford, J. A., Swann, Z., Newton, J. R., Bowden, R. J., Allen, G. P. & Davis-Poynter, N. (2006). Analysis of equid
herpesvirus 1 strain variation reveals a point mutation of the DNA polymerase
strongly associated with neuropathogenic versus non-neuropathogenic disease
outbreaks. J Virol 80, 4047-4060.
Osterrieder, N. (1999). Construction and characterization of an equine herpesvirus 1
glycoprotein C negative mutant. Virus Research 59, 165-177.
Osterrieder, N., Neubauer, A., Brandmuller, C., Braun, B., Kaaden, O. R. & Baines, J. D. (1996). The equine herpesvirus 1 glycoprotein gp21/22a, the herpes simplex
virus type 1 gM homolog, is involved in virus penetration and cell-to-cell spread
of virions. J Virol 70, 4110-4115.
Osterrieder, N., Wagner, R., Brandmuller, C., Schmidt, P., Wolf, H. & Kaaden, O. R. (1995). Protection against EHV-1 challenge infection in the murine model after
vaccination with various formulations of recombinant glycoprotein gp14 (gB).
Virology 208, 500-510.
Scott, J. C., Dutta, S. K. & Myrup, A. C. (1983). In vivo harboring of equine
herpesvirus-1 in leukocyte populations and subpopulations and their quantitation
from experimentally infected ponies. American Journal of Veterinary Research
44, 1344-1348.
Sinzger, C., Muntefering, H., Loning, T., Stoss, H., Plachter, B. & Jahn, G. (1993). Cell types infected in human cytomegalovirus placentitis identified by
immunohistochemical double staining. Virchows Archiv A, Pathological Anatomy
and Histopathology 423, 249-256.
Smith, D. J., Hamblin, A. & Edington, N. (2002). Equid herpesvirus 1 infection of
endothelial cells requires activation of putative adhesion molecules: an in vitro
model. Clin Exp Immunol 129, 281-287.
Smith, D. J., Hamblin, A. S. & Edington, N. (2001). Infection of endothelial cells with
equine herpesvirus-1 (EHV-1) occurs where there is activation of putative
adhesion molecules: a mechanism for transfer of virus. Equine Vet J 33, 138-142.

Expression of late proteins is hampered
106
Smith, K. C. & Borchers, K. (2001). A study of the pathogenesis of equid herpesvirus-1
(EHV-1) abortion by DNA in-situ hybridization. Journal of Comparative
Pathology 125, 304-310.
Smith, K. C., Mumford, J. A. & Lakhani, K. (1996). A comparison of equid
herpesvirus-1 (EHV-1) vascular lesions in the early versus late pregnant equine
uterus. Journal of Comparative Pathology 114, 231-247.
Spear, P. G. & Longnecker, R. (2003). Herpesvirus entry: an update. J Virol 77, 10179-
10185.
Sugahara, Y., Matsumura, T., Kono, Y., Honda, E., Kida, H. & Okazaki, K. (1997). Adaptation of equine herpesvirus 1 to unnatural host led to mutation of the gC
resulting in increased susceptibility of the virus to heparin. Arch Virol 142.
Toth, F. D., Mosborg-Petersen, P., Kiss, J., Aboagye-Mathiesen, Hager, G., Juhl, C.
B., Gergely, L., Zdravkovic, M., Aranyosi, J., Lampé, L. & Ebbesen, P. (1995). Interactions between human immunodeficiency virus type 1 and human
cytomegalovirus in human term syncytiotrophoblastcells coinfected with both
viruses. J Virol 69, 2223-2232.
van der Meulen, K., Caij, B., Pensaert, M. & Nauwynck, H. (2006a). Absence of viral
envelope proteins in equine herpesvirus 1-infected blood mononuclear cells during
cell-associated viremia. Vet Microbiol 113, 265-273.
van der Meulen, K., Vercauteren, G., Nauwynck, H. & Pensaert, M. (2003a). A local
epidemic of equine herpesvirus 1-induced neurological disorders in Belgium.
Vlaams Diergenskund Tijds 72, 366-372.
van der Meulen, K. M., Favoreel, H. W., Pensaert, M. B. & Nauwynck, H. J. (2006b). Immune escape of equine herpesvirus 1 and other herpesviruses of
veterinary importance. Vet Immunology and Immunopathology 111, 31-40.
van der Meulen, K. M., Nauwynck, H. J. & Pensaert, M. B. (2003b). Absence of viral
antigens on the surface of equine herpesvirus-1-infected peripheral blood
mononuclear cells: a strategy to avoid complement-mediated lysis. J Gen Virol 84,
93-97.
Vandekerckhove, A. P., Glorieux, S., Gryspeerdt, A. C., Steukers, L., Duchateau, L., Osterrieder, N., Van de Walle, G. R. & Nauwynck, H. J. (2010). Replication
kinetics of neurovirulent versus non-neurovirulent equine herpesvirus type 1
strains in equine nasal mucosal explants. J Gen Virol 91, 2019-2028.
Wellington, J. E., Lawrence, G. L., Love, D. N. & Whalley, J. M. (1996a). Expression
and characterization of equine herpesvirus 1 glycoprotein D in mammalian cell
lines. Arch Virol 141, 1785-1793.
Wellington, J. E., Love, D. N. & Whalley, J. M. (1996b). Evidence for involvement of
equine herpesvirus 1 glycoprotein B in cell-cell fusion. Arch Virol 141, 167-175.

Characteristics of a natural occurring outbreak of EHV1
107
CHAPTER V
DESCRIPTION OF AN UNUSUALLY LARGE
OUTBREAK OF NERVOUS SYSTEM DISORDERS
CAUSED BY EQUINE HERPESVIRUS 1 (EHV1) IN
2009 IN BELGIUM
Adapted from:
Vlaams Diergeneeskundig Tijdschrift 80 (2011), 139-145
Annick C. Gryspeerdt, A.P. Vandekerckhove, J. Van Doorsselaere, G.R. Van de
Walle, H.J. Nauwynck


Characteristics of a natural occurring outbreak of EHV1
109
SUMMARY
Neurological disease in horses caused by neurovirulent strains of EHV1 has been reported
with increasing frequency during the last decade. Therefore, concern has been voiced
within the US horse industry that the neurologic form of EHV1 may be intensifying in
prevalence and/or morbidity and mortality. Full protection against these severe signs
cannot be obtained by vaccination with the currently available vaccines and effective
antiviral therapies are not available. In Belgium, outbreaks of EHV1-induced abortions
are a yearly recurrent phenomenon, but outbreaks of equine herpes myelitis (EHM) are
extremely rare. This report describes an unusually large outbreak of EHV1-induced
neurological disease in Antwerp, Belgium, which involved at least 13 different premises.
Seven of them were characterized in more detail and were included in this research. A
total of 212 horses were exposed to EHV1 and a morbidity of 26% was seen. The
incidence of neurological disorders in the affected horses was 43%. Furthermore, the
outbreak was characterized by the rapid occurrence of ataxia and paralysis immediately
after disappearance of fever. EHV1 was diagnosed as the etiological agent by means of
virus isolation and/or the detection of seroconversion. Isolated virus was classified as
neurovirulent EHV1, with all isolates belonging to group 4 upon sequencing in the
ORF30 and ORF68 region. The latter confirms the suspicion of the outbreak spreading
from the first premise. The extent of this current outbreak and the high percentage of
horses with neurological signs, along with the fact that EHM is only sporadically seen in
Flanders, might indicate that the neurological form of EHV1 is possibly also emerging in
Belgium.

Characteristics of a natural occurring outbreak of EHV1
110
1 INTRODUCTION
Equine herpesvirus 1 (EHV1), a member of the Alphaherpesvirinae, is endemic in the
horse population worldwide. After exposure, virus replicates in tissues of the upper
respiratory tract. Primary replication is followed by a leukocyte-associated viremia which
enables EHV1 to reach internal organs (Gryspeerdt et al., 2010; Kydd et al., 1994a; b). A
second replication in endothelial cells at the site of the pregnant uterus or the nervous
system may result in abortion and neonatal foal death or nervous system disorders
respectively (Allen & Bryans, 1986; Gilkerson & Barrett, 2008). The latter, also
designated equine herpes myelitis (EHM), is a severe condition which is still poorly
understood despite the correlation between a single amino acid variation in the ORF30
region of the viral DNA polymerase and neurological disease (Lunn et al., 2009; Nugent
et al., 2006; Van de Walle et al., 2009).
EHV1 is a ubiquitous pathogen with a worldwide distribution and yearly problems
with respiratory disorders and abortion are common. In contrast, outbreaks of nervous
system disorders caused by infection with EHV1 seem to be more rare. However, during
the last decade, large outbreaks of EHM have been reported in the USA with enhancing
frequency and the fatal outcomes after substantial suffering appeared to increase (Lunn et
al., 2009). In Belgium, EHV1 is also endemic in the horse population and EHV1-induced
abortion is a yearly recurrent event. However, EHM is rare and mostly involves a limited
number of horses on a single premise (van der Meulen et al., 2000). To date, only one
description of EHV1-induced neurological disorders has been reported in Belgium. No
extensive spread of EHM was seen during this episode and severe disease was limited to
the affected premise. The signs in neurologically affected horses consisted mainly of
ataxia and paralysis, but cerebral dysfunctions were also noticed (van der Meulen et al.,
2003).
The current report describes a large outbreak of EHV1-induced neurological disease
in Flanders in 2009, that started on a riding school and spread to at least 12 premises that
had been in direct contact with the riding school. The extent of this outbreak, along with
the higher percentage of horses with neurological signs, without the occurrence of
cerebral dysfunctions, and the rapid occurrence of severe signs raises the intriguing
question if EHM is also emerging in Belgium.

Characteristics of a natural occurring outbreak of EHV1
111
2 MATERIALS AND METHODS
2.1 Description of the premises
During an outbreak of EHV1-induced nervous system disorders in Antwerp, Flanders
(June 2009), fever and neurological disease were observed on at least 13 premises. All
these premises had attended a jumping contest on May 30th
, 2009, at a riding school
where the first cases of neurological signs occurred (premise n° 1). Although 13 premises
reported neurological disease, only the data of 7 premises were included in this report. On
these premises, EHV1 infection was confirmed by virus isolation and/or seroconversion
(Table II). The other 6 affected premises had several horses with neurological problems,
but refused to give any further information and were therefore not included in this
research (Figure 1). Information about vaccination state, management and housing can be
found in Figure 1 and Table I. Each premise was visited for evaluation of clinical signs
and local veterinarians were asked to collect blood samples from horses suspected of
infection with EHV1 for both virus isolation from leukocytes and detection of
seroconversion in paired serum samples.
2.2 Serological examination
The presence of EHV-specific antibodies was determined with a complement-
dependent seroneutralization (SN)-test in acute serum samples and convalescent serum
samples three weeks later. Two-fold dilution series of the sera were prepared in MEM.
Fifty µl of these serial dilutions were incubated for 23 hours (h) at 37°C with a fixed
number of infectious virus (300 TCID50 of EHV1 strain Arabica in 50 µl). Hereafter, 25
µl of guinea pig complement was added. After 1 h of incubation, the mixture of serum,
virus and complement was added to RK13 cells. Inoculated cultures were further
incubated at 37°C in an atmosphere containing 5% CO2. After 7 days of incubation, the
cultures were analyzed for the presence of cytopathic effect. The neutralization titre was
calculated as the reciprocal of the highest dilution of serum that was able to completely
block EHV1 infection in RK13 cells. Seroconversion was present when the antibody titre
of the convalescent sample was at least four times higher than in the acute sample.

Characteristics of a natural occurring outbreak of EHV1
112
2.3 Virological examination
Virus isolation from peripheral blood mononuclear cells (PBMC) was performed on
RK13 cells. Unclotted blood samples (EDTA) were taken from horses with clinical signs
(fever and/or neurological disorders). PBMC were isolated by means of density
centrifugation on Ficoll-Paque (Pharmacia Biotech AB, Uppsala, Sweden) and
subsequently inoculated on monolayers of RK13 cells. Monolayers were incubated at
37°C in an atmosphere containing 5% CO2 and examined daily for 7 consecutive days for
the presence of cytopathic effects.
2.4 PCR amplification and sequencing of ORF30 and ORF68
DNA was extracted from EHV1 isolates using a QiaAmp DNA extraction kit (Qiagen
Benelux, Venlo, The Netherlands). Amplification of ORF30 was performed using the
primers ORF30fw (gctacttctgaaaacggaggc) and ORF30rev (ctatcctcagacacggcaaca) and
amplification of ORF68 with the primers ORF68atg1 (atgggtgtggtcttaattac) or ORF68fw
(aagcattgccaaacagttcc) in combination with ORF68end1 (aacgtgtggatgtccggcc). PCR
products were treated with Exonuclease I and Antarctic Phosphatase (New England
Biolabs, Ipswich, USA) and used directly for cycle sequencing with a Big Dye
Terminator Cycle sequencing kit V1.1
(Applied Biosystems, Foster City, USA).
Additional internal oligonucleotides were used for sequencing of ORF68. Cycle
sequencing reaction products were purified using ethanol precipitation and separated on
an ABI Genetic Analyzer 310 (Applied Biosystems, Foster City, USA). The sequences
were analyzed and compiled using Align, ClustalW and Sixframe in the workbench
(workbench.sdsc.edu) and BlastN and BlastP at www.ncbi.nlm.nih.gov. To validate the
sequencing, previously characterized strains 97P70 (Genbank accession number:
GU271940), 03P37 (# GU271941) and Ab4 (# AY665713) were included (Garré et al.,
2007; Nugent et al., 2006).

Characteristics of a natural occurring outbreak of EHV1
113
2.5 Treatment
All affected animals received supportive therapy consisting of anti-inflammatory
drugs and antibiotics. Horses with severe ataxia and/or bladder paralysis were hung in
slings and the bladder was emptied manually twice a day. Some animals received an
antiviral therapy, consisting of 40 mg of valacyclovir/kg body weight, administered 3
times daily for different consecutive days (Garré et al., 2009). The doses were prepared
by mixing the calculated amount of valacyclovir with water and administered orally.
Because of cost restraints and the limited amount of valacyclovir that was available, only
four affected horses were treated for a different number of consecutive days.
3 RESULTS
3.1 Description of the number, age and sex of affected animals during the outbreak
On the first premise, a morbidity (affected horses/total n° of horses present) of 21%
was seen during the outbreak and this premise reported to vaccinate twice a year with two
different inactivated vaccines (Figure 1). The animals on premise n° 4 had a variable
vaccination status. The morbidity was only 9%. However, clear clinical disease was only
seen in non-vaccinated animals and not all animals were checked for signs of EHV1. All
other premises did not vaccinate against EHV1 and morbidity varied from 25% (premise
n° 5) up to 100% (premise n° 2, 3, 6 and 7). Overall, a morbidity of around 23% was
observed during this outbreak.
The age of the affected animals varied from 3 to 21 years. No correlation between the
severity of clinical signs and age could be noticed during the course of this outbreak.
In total, 43% of the affected horses developed nervous system disorders. Of all
infected females 55% (18/33) developed nervous system disorders. Only 24% (5/21) of
all infected males developed nervous system disorders (Table I). Of the affected horses
with neurological signs 30% (7/23) were eventually euthanized and these were mainly
female (6/7 or 86%) (Table I).

Characteristics of a natural occurring outbreak of EHV1
114
Figure 1. Schematic overview of the course of the outbreak of EHV1-induced neurological disorders
at the different premises. NSD: nervous system disorders; open management: public riding school with
incoming and outgoing horses; half-open management: private premise, but where horses attend
competitions at other premises; housing with direct contact: stables separated from each other by bars;
housing with indirect contact: stables separated from each other by full walls.
Table I. Description of the animals on the premises with EHV1 associated neurological disorders
Total n° of horses present
Total n° of affected horses
N° of horses with neurological
disorders
Premise
N°
Male Female Total Male Female Total (%) Male Female Total (%)
1 53 43 96 8 12 20 (21%) 2 6 8 (40%)
2 1 2 3 1 2 3 (100%) 0 1 1 (33%)
3 2 10 12 0 3 3 (25%) 0 3 3 (100%)
4 42 38 80 2 5 7 (9%) 0 1 1 (14%)
5 3 4 7 3 4 7 (100%) 1 4 5 (71%)
6 2 1 3 2 1 3 (100%) 1 1 2 (67%)
7 5 6 11 5 6 11 (100%) 1 2 3 (27%)
108 104 212 21 33 54 (26%) 5 18 23 (43%)
Premise 1
First case: May 14th
Last case: June 16th
N°of horses present: 96
% morbidity: 21
% of horses with NSD: 40
Management: open
Housing: direct contact
Vaccination state: 2x/year
Premise 2
First case: June 5th
Last case: June 20th
N°of horses present: 3
% morbidity: 100
% of horses with NSD: 33
Management: half open
Housing: indirect contact
Vaccination state: none
Premise 3
First case: June 9th
Last case: June 10th
N°of horses present: 12
% morbidity: 25
% of horses with NSD: 100
Management: half open
Housing: direct, indirect contact
Vaccination state: none
Premise 4
First case: June 9th
Last case: June 19th
N°of horses present: 80
% morbidity: 9
% of horses with NSD: 14
Management: open
Housing: indirect contact
Vaccination state: variable
Premise 5
First case: June 9th
Last case: June 14th
N°of horses present: 7
% morbidity: 100
% of horses with NSD: 71
Management: half open
Housing: indirect contact
Vaccination state: none
Premise 6
First case: June 15th
Last case: June 28th
N°of horses present: 3
% morbidity: 100
% of horses with NSD: 67
Management: half open
Housing: direct, indirect contact
Vaccination state: none
Premise 7
First case: June 17th
Last case: June 17th
N°of horses present: 11
% morbidity: 100
% of horses with NSD: 27
Management: half open
Housing: direct, indirect contact
Vaccination state: none
6 additional premises
with reports of
neurological disorders,
but without any
addtional information,
nor confirmation of
EHV1

Characteristics of a natural occurring outbreak of EHV1
115
3.2 Methodical description of clinical symptoms during the outbreak
The most prominent clinical sign present, was fever (>39.5°C) that lasted for 2-5 days
and swelling of lymph nodes. During pyrexia, most of the horses showed a temporary
weakness of the tail. In the majority of the affected horses, a slight serous nasal discharge
was noted. On premise n° 1 icterus and oedema of the hind legs were also reported. These
symptoms were not regularly observed in the six other affected premises, as only three of
them reported oedema of the hind legs in a few horses. In two horses, urticaria were seen
during the pyretic period and two other horses suffered from muscle trembling after
disappearance of fever. During the outbreak, 43% of the affected horses showed nervous
system disorders (Table I). This occurred in most (65%) cases immediately after
disappearance of pyrexia, but some horses (35%) developed ataxia and paralysis without
preceding elevated body temperatures. Since body temperatures were not checked
regularly in the beginning of the outbreak, a period of fever could have passed unnoticed.
During this outbreak, moderate as well as severe ataxia were noticed. Severe ataxia was
accompanied by bladder paralysis in approximately 50% of the cases. Several horses
became quadriplegic shortly after developing ataxia, but quadriplegia also occurred
without previous neurological symptoms. In total, seven horses were euthanized during
this outbreak. Six of these horses (from premise n° 1) were presented with paralysis
before an etiological diagnosis was made and were euthanized within 24 hours after the
onset of neurological symptoms. The last horse (premise n° 4) was euthanized two weeks
after the occurrence of paralysis since no improvement of the situation was seen. All other
horses with neurological symptoms that survived were clinically normal 4 months later
and regained normal performance levels, with the exception of one mare that is again
suffering from urinary incontinence after a clinically normal period of four months.
3.3 Evaluation of antiviral treatment during outbreak
During the outbreak, four horses were treated orally with the antiviral drug
valacyclovir (Table II). Horse A was a gelding of 4 years of age (premise n° 1). He was
treated when the fever ceased after 5 days and the horse showed muscle tremors. As the
muscle tremors stopped after one day and no other symptoms of EHV1 infection were
observed, the therapy was stopped after 2 days. Horse B was a mare of 7 years (premise

Characteristics of a natural occurring outbreak of EHV1
116
n° 2), and the valacyclovir treatment started at the moment she became quadriplegic. This
horse was unable to stand and was hung up in a sling. Two days later, she was not yet
able to stand up on her own, but she could stand without the help of the sling and her
condition improved rapidly. Treatment was stopped after 5 days. One week later, she was
still slightly ataxic but was able to stand up and regained bladder control. Treatment of
horse C (female, 5 years, premise n° 2) was started when she had difficulties standing up
and was found severely ataxic. She was treated for 3 consecutive days. One week later,
she was still obviously ataxic but this slowly improved over the next couple of weeks.
Bladder paralysis lasted for almost three weeks. Finally, horse D (female, 7 years,
premise n° 3) was treated after she became quadriplegic for more than 36 hours. She was
lifted up in a sling and could barely stand on her own. The first two days after the
treatment was initiated, the horse showed a significant improvement. She was able to
stand on her own in the sling and even tried to walk. After three days, however, treatment
was stopped due to a lack of available valacyclovir. The horse remained stable for several
days, but no further improvement of clinical symptoms was observed. Two weeks after
the onset of symptoms, the horse was able to stand in the sling and started to regain
bladder control, but was not able to lay in sternal position or to stand up on her own.
Because this caused severe anxiety for the horse, she was euthanized.
Table II. Treatment of different animals with valacyclovir
Horse Gender Age Symptoms Duration of treatment Improvement
A ♂ 4 tremor 2 days +++
B ♀ 7 quadriplegia 5 days ++
C ♀ 5 severe ataxia 3 days +
D ♀ 7 quadriplegia 3 days ± †
†: euthanasia
3.4 Diagnosis
Blood samples were collected from seven horses with high fever, six horses with
ataxia and two horses with quadriplegia at the moment of sampling (Table III). EHV1
was isolated by means of PBMC co-cultivation from six of the seven horses with high
fever (86%), from four of the six horses with ataxia (67%) and from the two horses with

Characteristics of a natural occurring outbreak of EHV1
117
quadriplegia (100%) (Table III). Seroconversion of EHV-specific antibodies was
evaluated for thirteen of the fifteen horses, both at the time of clinical signs (acute
sample) as well as three weeks later (convalescent sample). Twelve horses seroconverted
for EHV1 (Table III).
All EHV1 isolates were typed as neurovirulent by sequencing of the D752 allele in the
ORF30 region (e.g. 09P240, # GU271939). In addition, all viruses were sequenced in
their ORF68 region, a genetic marker for grouping isolates into six major geographically
restricted groups (Nugent et al., 2006). All isolates in this outbreak of neurological
disease belonged to group 4. These genetic analyses, along with a direct link between the
confirmed premises and premise n° 1, further confirmed the assumption that all EHV1
infections on the different premises were caused by the same virus spread from premise
n° 1.
Table III. Results from different diagnostic tools to confirm EHV1 infection in discussed horses
Antibody titre in serum Premise Horse Signs
at the
moment of
sampling
Isolation of
EHV1 from
PBMC Acute sample Convalescent
sample
1 1 Fever + 128 1536
2 quadriplegia + 64 NA
2 1 Fever + 48 512
2 Fever + 16 256
3 1 Ataxia + 192 512
2 Ataxia + 96 1024
3 Ataxia - 16 256
4 1 quadriplegia + 96 NA
5 1 Fever + 4 1536
2 Fever + 12 768
3 Fever - 8 1024
4 Fever + 12 2048
6 1 Ataxia + 32 1536
7 1 Ataxia + 4 1536
2 Ataxia - 8 1536
NA: not available due to euthanasia

Characteristics of a natural occurring outbreak of EHV1
118
4 DISCUSSION
The present paper reports a large outbreak of EHV1-induced neurological disease in
Antwerp, Flanders in June 2009. How the present epidemic started on this premise is not
known, however two weeks before the first horse became recumbent and had to be
euthanized, a 5-year old mare was presented with fever, lasting for 4 days. It might be
hypothesized that this mare either reactivated latent EHV1 or got infected with EHV1 due
to many contacts of visiting horses on the premise. Secondary outbreaks on other
premises then occurred as a consequence of direct contact with infected horses from the
first premise. Both the first premise as well as secondary infected premises were put
under quarantine for three weeks and all horses without clinical symptoms in the
neighbourhood were vaccinated. The outbreak was successfully controlled and one month
after the onset of the outbreak, no more cases of EHV1 infection were reported.
An interesting feature of the present outbreak was the rapid occurrence of ataxia and
paralysis immediately after disappearance of fever. Previous reports mention an
incubation period of 7 to 10 days for the development of nervous system disorders after
experimental inoculation (Edington et al., 1986; Jackson et al., 1977). Interestingly,
although sex and age did not seem to influence the incidence of becoming infected, mares
appeared to be predisposed to develop severe ataxia and paralysis. Indeed, 55% of the
infected mares developed nervous system disorders (18/33), whereas this was only 24%
(5/21) for the infected male horses. The higher sensitivity of mares to develop severe
symptoms after EHV1 infection has already been described, however, no explanation for
this exists to date (Goehring et al., 2006; McCartan et al., 1995).
Although the efficacy of current commercial vaccines to protect horses against EHV1-
related symptoms is highly variable and none of them can guarantee 100% protection,
several vaccines have been shown to induce significant protection against either abortion
or nervous system disorders under experimental and field conditions (Goodman et al.,
2006; van der Meulen et al., 2007). The premises which were mainly affected during this
outbreak had an inadequate or no vaccination protocol at all. This observation has also
been reported during other outbreaks of EHV1-induced neurological disease, where most
outbreaks occurred on premises with unvaccinated horses (Goehring et al., 2006; van der
Meulen et al., 2003). During this outbreak, morbidity rates were lower (7 to 21%) in
vaccinated premises (n° 1 and 4) compared to the non-vaccinated premises (25 to 100%)

Characteristics of a natural occurring outbreak of EHV1
119
(Figure 1). On premise n° 4, where both vaccinated and unvaccinated horses were
present, fever and neurological disease was only reported in their unvaccinated horses
whereas the vaccinated horses showed no clinical signs of EHV1 infection. This could
indicate that vaccinated horses have a clinical protection against severe symptoms of
EHV1. Whether this protection is the result of a reduced spread of the virus or due to a
virological effect is not clear. Interestingly, although lower morbidity rates were observed
at the vaccinated premises n° 1 and 4, no difference could be seen in the number of horses
that eventually developed nervous system disorders compared to the non-vaccinated
premises (Table I). In Belgium, only inactivated vaccines are registered for use in horses
which mainly stimulate humoral immunity, whereas it is suggested that cell-mediated
immunity is of paramount importance to limit EHV1 infection (Kydd et al., 2006; Kydd
et al., 2003).
During this outbreak, four horses on three different premises were treated orally with
the antiviral drug valacyclovir. Experimental in vivo studies to evaluate the efficacy of
valacyclovir to reduce viral shedding and provide clinical recovery have recently been
performed, but with contradictory results (Garré et al., 2009; Maxwell et al., 2008).
During this outbreak, 8 horses showed paralysis. Two out of three treated horses (66%)
regained normal prestations, while five out of five (100%) untreated horses had to be
euthanized. Although the treated horses seemed to recover quite fast during treatment
compared to untreated horses with the similar clinical features, it is difficult to evaluate
the benefit of valacyclovir treatment during an outbreak, due to the limited amount of
treated horses versus non-treated horses and the variable treatment periods. Still,
extensive supportive care remains indispensable during an outbreak of EHV1-induced
neurological disorders to increase the likelyhood of recovery.
Both virus isolation from peripheral blood mononuclear cells (PBMC) as well as
seroconversion were used to diagnose EHV1 during this outbreak. EHV1 was
successfully isolated from PBMC during the present outbreak. This is in contrast to
previous published studies, mentioning that virus isolation from PBMC in outbreak
situations can be problematic (Lunn et al., 2009; van Maanen et al., 2001). In addition,
seroconversion has also been reported to have limitations in confirming a diagnosis of
EHV1, especially in horses with neurological signs (Lunn et al., 2009). Interestingly,
during the current outbreak all horses with neurological disorders seroconverted, with the
exception of one horse. An explanation for the successful diagnosis during this outbreak

Characteristics of a natural occurring outbreak of EHV1
120
could be the rapid occurrence of severe signs during the current outbreak which increased
the chance of detecting infectious virus and seroconversion. Low initial EHV1 antibodies
in a largely unvaccinated population could also have contributed to the easy detection of
seroconversion.
Although some interesting findings concerning risk factors, vaccination, treatment and
diagnosis were observed during this outbreak, conclusions drawn from natural occurring
outbreaks should be taken with care, owing to the small numbers involved and variability
in both vaccination and anti-viral treatment.
During the only other reported outbreak of EHV1-induced neurological disease in
Belgium six years ago, a morbidity of 42% was seen with 15% of the horses showing
neurological disorders. Ataxia and paralysis were observed, but also cerebral disorders
such as blindness, torticollis and severe apathy were seen (van der Meulen et al., 2003).
Cerebral disorders were not observed during this recent outbreak, but interestingly, this
outbreak was characterized by the rapid onset of ataxia and paralysis immediately after
the disappearance of fever. In addition, the incidence of neurological signs in the EHV1-
affected horses was much higher in the current outbreak (43%) compared to the outbreak
in 2003 (15%) (van der Meulen et al., 2003). These observations might possibly point
towards a change in neurovirulent potential of naturally circulating EHV1 strains.
In summary, concern has been voiced within the U.S. horse industry that the
neurologic form of EHV1 might be emerging, based on an increased number of EHM
outbreaks with a larger number of seriously affected horses throughout the United States
over the last decade (USDA, 2007). Until now, no such evolution was seen in Belgium
and no reports of EHM outbreaks have been reported with the exception of one limited
outbreak in 2003. The present report describes a recent large outbreak of EHV1 with a
rapid spread and a remarkably high number of animals suffering of EHM. Together with
the fact that also in 2010 two small outbreaks were reported, the question raises whether
the neurological form of EHV1 is also emerging in Belgium and hence Europe. However,
it is not clear if these observations are related to an evolving disease with a changed
virulence and behaviour or if they are the consequence of a greater awareness and the
development of better diagnostic methods (USDA, 2007). Many data gaps exist, and more
investigations need to be conducted to better understand this evolving situation and to
identify other possible factors playing a role. Therefore, a good surveillance system in
Belgium and Europe would be of great importance to monitor outbreaks of EHM, to gain

Characteristics of a natural occurring outbreak of EHV1
121
better insights into the virulence of naturally circulating EHV1 strains and to determine if
EHM is in fact an emerging disease.
Acknowledgements
This research was supported by the Institute for the promotion of Innovation
through Science and Technology in Flanders (IWT-Vlaanderen). The authors like to
thank C. Boone for the help with the processing of all samples and Ine Vanherpe for
performing PCR and sequencing. We would also like to thank the veterinarians Marc
Schelkens, Jacques Corstiens, Wim Vermeiren, Dries Vermeiren, Kamiel Warnants, Jef
Nuytten and all horse owners for providing samples and detailed information.

Characteristics of a natural occurring outbreak of EHV1
122
REFERENCES
Allen, G. P. & Bryans, J. T. (1986). Molecular epizootiology, pathogenesis, and
prophylaxis of equine herpesvirus-1 infections. Prog Vet Microbiol Immunol 2,
78-144.
Edington, N., Bridges, C. G. & Patel, J. R. (1986). Endothelial cell infection and
thrombosis in paralysis caused by equid herpesvirus-1: equine stroke. Arch Virol
90, 111-124.
Garré, B., Gryspeerdt, A., Croubels, S., De Backer, P. & Nauwynck, H. (2009). Evaluation of orally administered valacyclovir in experimentally EHV1-infected
ponies. Vet Microbiol 135, 214-221.
Garré, B., van der Meulen, K., Nugent, J., Neyts, J., Croubels, S., De Backer, P. & Nauwynck, H. (2007). In vitro susceptibility of six isolates of equine herpesvirus
1 to acyclovir, ganciclovir, cidofovir, adefovir, PMEDAP and foscarnet. Vet
Microbiol 122, 43-51.
Gilkerson, J. R. & Barrett, E. J. (2008). Equine herpesvirus neurological disease.
Equine Vet J 40, 102-103.
Goehring, L. S., van Winden, S. C., van Maanen, C. & Sloet van Oldruitenborgh-Oosterbaan, M. M. (2006). Equine herpesvirus type 1-associated
myeloencephalopathy in The Netherlands: a four-year retrospective study (1999-
2003). J Vet Intern Med 20, 601-607.
Goodman, L. B., Wagner, B., Flaminio, M. J., Sussman, K. H., Metzger, S. M., Holland, R. & Osterrieder, N. (2006). Comparison of the efficacy of inactivated
combination and modified-live virus vaccines against challenge infection with
neuropathogenic equine herpesvirus type 1 (EHV-1). Vaccine 24, 3636-3645.
Gryspeerdt, A. C., Vandekerckhove, A. P., Garre, B., Barbe, F., Van de Walle, G. R. & Nauwynck, H. J. (2010). Differences in replication kinetics and cell tropism
between neurovirulent and non-neurovirulent EHV1 strains during the acute phase
of infection in horses. Vet Microbiol 142, 242-253.
Jackson, T. A., Osburn, B. I., Cordy, D. R. & Kendrick, J. W. (1977). Equine
herpesvirus 1 infection of horses: studies on the experimentally induced
neurologic disease. Am J Vet Res 38, 709-719.
Kydd, J. H., Smith, K. C., Hannant, D., Livesay, G. J. & Mumford, J. A. (1994a). Distribution of equid herpesvirus-1 (EHV-1) in respiratory tract associated
lymphoid tissue: implications for cellular immunity. Equine Vet J 26, 470-473.
Kydd, J. H., Smith, K. C., Hannant, D., Livesay, G. J. & Mumford, J. A. (1994b). Distribution of equid herpesvirus-1 (EHV-1) in the respiratory tract of ponies:
implications for vaccination strategies. Equine Vet J 26, 466-469.
Kydd, J. H., Townsend, H. G. & Hannant, D. (2006). The equine immune response to
equine herpesvirus-1: the virus and its vaccines. Vet Immunol Immunopathol 111,
15-30.
Kydd, J. H., Wattrang, E. & Hannant, D. (2003). Pre-infection frequencies of equine
herpesvirus-1 specific, cytotoxic T lymphocytes correlate with protection against
abortion following experimental infection of pregnant mares. Vet Immunol
Immunopathol 96, 207-217.
Lunn, D. P., Davis-Pointer, N., Flaminio, M. J., Horohov, D. W., Osterrieder, K., Pusterla, N. & Townsend, H. G. (2009). Equine Herpesvirus-1 Consensus
Statement. J Vet Intern Med 23, 450-461.

Characteristics of a natural occurring outbreak of EHV1
123
Maxwell, L. K., Bentz, B. G., Gilliam, L. L., Ritchey, J. W., Holbrook, T. C.,
McFarlane, D., Rezabek, G. B., MacAllister, C. G. & Allen, G. P. (2008). Efficacy of Valacyclovir Against Clinical Disease After EHV-1 Challenge in
Aged Mares. Proceedings of the Annual Convention of the AAEP 54, 198-199.
McCartan, C. G., Russell, M. M., Wood, J. L. & Mumford, J. A. (1995). Clinical,
serological and virological characteristics of an outbreak of paresis and neonatal
foal disease due to equine herpesvirus-1 on a stud farm. Vet Rec 136, 7-12.
Nugent, J., Birch-Machin, I., Smith, K. C., Mumford, J. A., Swann, Z., Newton, J. R., Bowden, R. J., Allen, G. P. & Davis-Poynter, N. (2006). Analysis of equid
herpesvirus 1 strain variation reveals a point mutation of the DNA polymerase
strongly associated with neuropathogenic versus non-neuropathogenic disease
outbreaks. J Virol 80, 4047-4060.
USDA (2007). Equine herpesvirus myeloencephalopathy: A potentially emerging disease.
Available at: http://wwwaphisusdagov/vs/nahss/equine/ehv.
Van de Walle, G. R., Goupil, R., Wishon, C., Damiani, A., Perkins, G. A. & Osterrieder, N. (2009). A Single-Nucleotide Polymorphism in a Herpesvirus
DNA Polymerase Is Sufficient to Cause Lethal Neurological disease. J Infect Dis
200, 20-25.
van der Meulen, K., Nauwynck, H. & Pensaert, M. (2000). Equine herpesvirus type 1
abortion, neonatal foal death and nervous system disorders diagnosed in Belgium
in 1999. Vlaams Diergenskund Tijds 69, 38-41.
van der Meulen, K., Vercauteren, G., Nauwynck, H. & Pensaert, M. (2003). A local
epidemic of equine herpesvirus 1-induced neurological disorders in Belgium.
Vlaams Diergenskund Tijds 72, 366-372.
van der Meulen, K. M., Gryspeerdt, A. C., Vandekerckhove, A. A., Garre, B. A. & Nauwynck, H. J. (2007). The protective properties of vaccination against equine
herpesvirus 1-induced viremia, abortion and nervous system disorders. Vlaams
Diergenskund Tijds 76, 186-194.
van Maanen, C., Sloet van Oldruitenborgh-Oosterbaan, M. M., Damen, E. A. & Derksen, A. G. (2001). Neurological disease associated with EHV-1-infection in
a riding school: clinical and virological characteristics. Equine Vet J 33, 191-196.


General discussion
125
CHAPTER VI
GENERAL DISCUSSION


General discussion
127
GENERAL DISCUSSION
Equine herpesvirus 1 (EHV1) is a ubiquitous equine alphaherpesvirus. After
replication in tissues of the upper respiratory tract (URT), EHV1 spreads via a leukocyte-
associated viremia to internal organs such as the pregnant uterus and the nervous system
where a second replication cycle is initiated in the endothelial cells of these organs. The
viral replication can lead to upper respiratory tract disease, abortion or neonatal foal death
and equine herpes myelitis (EHM) (Allen & Bryans, 1986; van Maanen, 2002). In
veterinary medicine, vaccination is the most designated weapon against viral infections.
Several vaccines attempting to prevent severe consequences of infection with EHV1 are
currently on the market. Unfortunately, to date, none of these is able to fully prevent the
EHV1-induced severe secondary symptoms (van der Meulen et al., 2007). Therefore,
effective antiviral medicines could be of great importance to limit virus replication and
clinical signs during acute and recurrent viral infections. To date, no antiviral drugs are
registrated for the use in horses, neither are they efficient enough to battle EHV1
infections. One of the major problems in developing efficient vaccines or antiviral drugs
is that the pathogenesis and immune evasion strategies of EHV1 are not fully known,
which necessitates further research on this matter.
In vivo experiments are a necessary tool to study the pathogenesis of specific
pathogens in animals. However, in the case of horses, such experiments are expensive,
labour-intensive and are often a sensitive matter from an ethical point of view. Therefore,
attempts have been made to find appropriate models to replace those horse experiments.
Laboratory animals may be convenient and valuable tools for use in experimental systems
of disease in large animals. However, they are often criticized because of their failure to
provide relevance to the disease in the target species. The most frequently used in vivo
model for the assessment of virological and histological characteristics of EHV1-induced
disease in horses has been the murine model. The extent to which valid comparisons and
extrapolations about pathogenesis can be made from mouse to horse remains highly
questionable (Walker et al., 1999). For example, the EHV1 strain RacL11 causes only
mild disease in horses (Van de Walle et al., personal communication), while in mice,
inoculation with RacL11 results in severe symptoms and even death (von Einem et al.,
2007). Even though the mouse model is suggested to provide a valid method for

General discussion
128
investigation into several aspects of EHV1-induced disease (Walker et al., 1999),
investigating the pathogenesis of EHV1 in its natural host, the horse, remains inevitable.
Therefore, EHV1 pathogenesis studies in the horse were performed in this thesis, which
provides relevant and unambiguous new insights. It was found that EHV1 replicates in a
plaquewise manner in the epithelium of the upper respiratory tract. Plaques were not able
to penetrate the basement membrane, but individual infected cells could be found in the
connective tissue from 1 dpi. These cells were mainly CD172a+ cells and CD5
+
lymphocytes. Viremia was detected from 1 dpi, with again CD172a+ cells as main carrier
cells. Infected epithelial cells and mucosal leukocytes were examined more thoroughly
and expression of viral late glycoproteins gB, gC, gD and gM was evaluated. It was found
that although a complete replication cycle with formation of plaques was seen in
epithelial cells, individual infected cells showed a low expression of late glycoproteins gC
and gD. The latter indicates that the protein expression in EHV1-infected immune cells is
hampered. Interestingly, the results obtained in this thesis were virtually identical as the
results obtained in the recently established in vitro model of equine nasal mucosa
(Vandekerckhove et al., 2010). Therefore, the latter model could be a valuable tool for the
study of EHV1 pathogenesis in vitro.
Based on the observations above, several new insights in the pathogenesis of EHV1
were obtained and some missing links were elucidated. Therefore, an improved model on
the pathogenesis of EHV1 in the upper respiratory tract is put forward in this thesis
(Figure 1). Upon infection, EHV1 replicates in epithelial cells of the nasal septum and
nasopharynx, which results in a productive infection with plaque formation and nasal
virus shedding (2). This nasal shedding allows transmission of virus to susceptible contact
animals and ensures that EHV1 can persist in the equine population. This productive
infection in epithelial cells, however, is self-limiting, most likely due to IFN-α
production. In addition, EHV1 seems to cross the basement membrane through the help of
CD172a+ carrier cells, probably mucosal dendritic cells (3). We postulate that these
infected carrier cells can transport the virus across the basement membranes and through
connective tissues in the direction of vascular endothelial cells (4a) on the one hand and
lymphatic epithelium (4b) on the other hand. As a result, a cell-associated viremia is
originated (5), together with migration of infected immune cells to local lymph nodes,
hereby initiating the start of a specific immune response. During viremia, also CD172a+
cells were the main carrier cells, suggesting that these cells indeed originate during the

General discussion
129
early stages of infection in the upper respiratory tract. Although epithelial cells undergo a
complete replication cycle with the progeny of new infectious virus particles, infected
leukocytes seem to have a hampered expression of some late proteins. The same
observation has been previously demonstrated, as it was seen that during viremia, 98% of
EHV1-infected PBMC present in the blood stream lacked expression of certain late
proteins (van der Meulen et al., 2003b). When we combine the latter results with our
findings, we can conclude that infected leukocytes in the upper respiratory tract could be
the source of the cell-associated viremia. These cells undergo an incomplete replication
cycle even in the early steps of pathogenesis, which might be a mechanism to protect
these cells from destruction by the immune system. Interestingly, the latter could explain
why despite high antibody levels, viremia is still observed in immune animals. Transfer of
EHV1 from viremic cells to endothelial cells of target organs (6) causes infection,
followed by replication of EHV1 and subsequent damage to these organs (7).
Figure 1. Adapted schematic overview of the pathogenesis of EHV1. 1) Attachment/penetration of virus in
epithelial cells of the upper respiratory tract; 2) replication of virus in epithelial cells by means of plaque
formation; 3) spread of virus through the BM by means of individual infected leukocytes; 4) spread of
infected leukocytes to vascular endothelial cells (4a) and draining lymph nodes (4b); 5) leukocyte
associated viremia in CD172a+ carrier cells; 6) transfer of virus from infected leukocytes to endothelial cells
of target organs; 7) replication and spread of virus in target organs. Full arrow: known mechanism; striped
arrow: unknown mechanism.
1
4a
5
67
3
4b
2

General discussion
130
New insights in the pathogenesis and the natural occurrence of EHV1, may provide
new directions in the amelioration of vaccines and the development of new drugs for
treatment. Indeed, at present, the two most important tools in the battle against EHV1 are
vaccination and antiviral therapy.
An efficient vaccine against EHV1 is of great importance to control outbreaks of
EHV1 and prevent the development of serious problems such as abortion and nervous
system disorders. Firstly, during this research, we have observed that the first steps in the
pathogenesis of EHV1 occur very rapidly. During primary replication in tissues of the
upper respiratory tract, single infected cells are already seen at 1 day pi (dpi) and plaque
formation was seen from 2 dpi. Already after 1 or 2 dpi, single infected cells were present
beneath the basement membrane and viremia could be detected. The latter implies that
vaccines can only prevent viremia and subsequent spread to internal organs if an effective
local immunity can be established at the level of the upper respiratory tract, preventing
the origin of individual infected cells beneath the basement membrane and the subsequent
viremia. Because horses have been shown to be protected against viremia up to 6 months
after a natural infection with EHV1 (Breathnach et al., 2001; Edington & Bridges, 1990),
local immunity can prevent viremia at least for some time. This makes a modified life
vaccine (MLV) a good candidate in vaccine development and a suitable recombinant
EHV1 MLV could be a virus which is able to replicate in the epithelium of the upper
respiratory tract but unable to penetrate the basement membrane by the use of individual
infected cells. As it has been shown that the mucosal nasal explant model is an accurate
alternative for in vivo testing, this model can be used to rapidly screen several potential
MLV candidates before starting expensive and labour-intensive in vivo experiments
(Vandekerckhove et al., 2009; Vandekerckhove et al., 2010). Hereby, suitable
recombinant viruses could be selected for the use in in vivo experiments based on their
inability to penetrate the basement membrane by using individual infected immune cells.
Secondly, during viremia, carrier cells of EHV1 are present in a blocked stage as certain
late glycoproteins are not expressed (van der Meulen et al., 2006). In fact, we could
demonstrate that this immune evasion strategy already took place in the early stages of
infection, as soon as individual infected cells appeared in the connective tissues following
penetration of the basement membrane (Gryspeerdt et al., 2011). Such immune evasion
strategy could supposedly protect them against antibody-dependent destruction,
suggesting that humoral antibodies are of no use during this phase of infection (van der

General discussion
131
Meulen et al., 2003b). However, a humoral protection could be of importance in the last
step of the pathogenesis, when virus is transferred from its carrier cells to the endothelium
of the target organs. Here, antibodies can play a role in either (i) preventing virus to be
transmitted from carrier cells to endothelial cells, or (ii) limiting the extent of plaque
formation in target organs. Unfortunately, a recent study demonstrated that virus could be
transferred in vitro to endothelial cells despite the presence of neutralizing antibodies
(Goehring et al., 2010), meaning that humoral immunity will probably be of limited use
in this step of the pathogenesis. However, antibodies may still play an important role in
limiting replication in target organs and thus limiting the damage and the occurrence of
severe symptoms. The latter could be of interest for future research, where the effect of
neutralizing antibodies on plaque formation and virus spread in endothelial cells of target
organs should be evaluated.
A thorough knowledge on the pathogenesis of the virus is of paramount importance
for the development of antivirals. Based on the results of this thesis, we are now able to
predict where an antiviral could exert its action. When we look at the rapid invasion of
EHV1 into the connective tissue, combined with the fact that carrier cells are already in a
blocked stage during early pathogenesis, there are only three places where antivirals could
have an effect on the replication of EHV1 in the horse: (i) the entry and primary
replication of the virus in the mucosa of the URT, (ii) the transmission of virus from
infected PBMC to endothelial cells, and (iii) the secondary replication of the virus in
endothelial cells at the target organs. To have a rapid effect on the primary replication in
the epithelium of the URT, an antiviral with a metaphylactic use, applied locally in the
nose to protect horses from getting a productive infection, would be the most suitable. An
antiviral with an effect on the amount and duration of viremia will be difficult since
during viremia the infected leukocyte seems to be blocked, which limits the possible
targets of antivirals (van der Meulen et al., 2003b). Due to the rapid course of an EHV1
infection, most animals during an outbreak will already be in a late stage of infection,
namely the secondary replication in endothelial cells of the target organs, before a therapy
can be initiated. An antiviral which either inhibits and/or limits the extent of this
secondary replication seems to have great potential in limiting damage at secondary target
organs and therefore severe symptoms. However, until now, the latter is difficult to
evaluate since there is no appropriate in vitro or in vivo model to study the possible effect
of an antiviral at secondary replication sites. Given the fact that after oral valacyclovir

General discussion
132
treatment no influence on nasal shedding or viremia was observed (Garré et al., 2009),
but that a clinical effect and a more rapid recovery is suspected (Maxwell et al., 2008), it
seems that an effect of valacyclovir on the secondary replication of EHV1 cannot be
excluded. Interestingly, during the outbreak of neurological disease studied in this thesis,
a rapid spread of the virus was seen followed by a rapid appearance of neurological
disorders. Some horses did not show previous signs such as nasal discharge, fever or
ataxia and were merely found paralyzed, necessitating euthanasia. As treatment should
preferentially be initiated before severe symptoms such as paralysis become apparent
(Maxwell et al., 2008), any antiviral treatment would have been initiated too late in the
course of disease. As there is no way to predict in which stage of infection the horse is
during a natural outbreak of EHV1, an efficient antiviral drug should have both a
metaphylactic and a curative action. The best way to achieve this goal would be a local
treatment via aerosol, to prevent primary replication in tissues of the upper respiratory
tract. Horses with (severe) symptoms however, should be treated in a way to prevent or
inhibit further extension of endothelial damage and should be treated orally or
intravenously with a curative antiviral drug. Future research should be focussed on how
EHV1 penetrates through the basement membrane and gets into the blood stream. Also
the mechanism of transmission of the virus from infected PBMC to endothelial cells
should be studied. These are two phenomena where future antivirals could possibly
interfere with the pathogenesis of the virus, hereby limiting severe consequences after
infection with EHV1.
In 2009, several outbreaks of abortion and neonatal death were seen during the first
period of the year. Exceptionally, a large outbreak of EHV1-induced neurological disease
was seen in Antwerp. Concern has been voiced within the U.S. horse industry that the
neurologic form of EHV1 is emerging. The latter is based on the detection of an increased
number of EHM outbreaks throughout the United States over the last decade (USDA,
2007). Until now, no such evolution was seen in Belgium and no reports of large EHM
outbreaks have been observed with the exception of one limited outbreak in 2003 (van der
Meulen et al., 2003a). The recent large outbreak of EHM showed a rapid spread and a
remarkably high number of animals with severe neurological symptoms. The latter,
together with the report of two novel outbreaks in 2010, raises the question whether the
neurological form of EHV1 is also emerging in Belgium, and hence, Europe. In 2009, one

General discussion
133
outbreak of EHM was reported in the UK, one in France and three in Ireland. In 2010,
five outbreaks were reported in France, two in the UK and one in Italy. Until now, three
outbreaks of EHM have been reported in France in 2011. Unfortunately, most European
countries do not report outbreaks of EHM, providing insufficient data to decide if EHM is
possibly emerging in Europe. Additionally, it is not clear if the reported observations are
related to an evolving disease with a changed virulence and behaviour or if they are the
consequence of a greater awareness and the development of better diagnostic methods
(USDA, 2007). Many data gaps exist, and more investigations need to be conducted to
better understand this evolving situation and to identify other possible factors playing a
role. Therefore, a good surveillance system in Belgium and Europe would be of great
importance to monitor outbreaks of EHM and to get better insights into the virulence of
naturally circulating EHV1 strains. An efficient communication system and a centralized
diagnostic centre would be of paramount importance to achieve this goal. Diagnostical
methods should be quick and efficient to have a rapid diagnosis during suspected
outbreaks of EHV1 (Lunn et al., 2009). Although virus isolation proved to be a reliable
method to confirm infection within two days, PCR techniques could be of importance as
results can be achieved within a few hours and information could be given about the
neurovirulence of the isolated EHV1 strain and the quantity of virus present (Allen, 2007;
Pusterla et al., 2008; Pusterla et al., 2009b).
Worldwide, several guidelines are available of what needs to be done during an
outbreak of EHV1. These guidelines vary widely both within and between countries. A
protocol on how to manage an EHV1 outbreak, from being prepared to collect samples
appropriately and how to contain the situation would be of great importance (USDA,
2007). Some guidelines found in literature, formed the basis of our approach during the
outbreak of EHM and were checked for their efficiency (Allen, 2002; Lunn et al., 2009;
Pusterla et al., 2009a). Based on our observations, we would like to discuss some of these
guidelines.
In the case of a suspected outbreak of EHV1, immediate measures should be taken to
prevent any possible spread of the virus within the affected premise or to other premises
(Lunn et al., 2009). Affected horses should be put in isolation and contact with other
horses or care takers should be completely avoided. Other horses should be monitored for
the appearance of fever or other symptoms suggesting an early infection (van Maanen,
2002). If possible, all horse movement in affected regions should be avoided until three

General discussion
134
weeks after the last reported case. During the outbreak of EHM in Antwerp, isolation and
quarantine measures proved to be of great importance to prevent further spread of the
virus. Suspected premises were closed until EHV1 infection was either confirmed or
excluded. Quarantine measures in confirmed premises were only lifted three weeks after
the last observed clinical case. Major horse events were cancelled and all horse
movements in affected regions were discouraged.
An early and accurate diagnosis of the cause of the observed symptoms, with virus
culture and isolation as the gold standard test, is of paramount importance to further
determine the strategies to be followed in managing the outbreak (Lunn et al., 2009).
Several samples were collected during the outbreak: unclotted blood samples for isolation
of PBMC, nasal swabs and paired serum samples. Isolation of EHV1 by means of
cocultivation of PBMC proved to be an important and efficient diagnostic tool. Within
two days, virus could be isolated from affected horses with fever, ataxia or paralysis.
Although nasal swabbing could be a good alternative in an even earlier stage of infection,
during this outbreak, no virus could be isolated from collected swabs. An explanation for
this observation could be inappropriate conservation techniques or the long transport
period under non-ideal circumstances. A seroconversion could be noticed in almost all
infected horses, but such methodology takes about 4 weeks before the final results are
known. Therefore, the best samples to collect in an acute stage of infection, at least in our
hands, seemed to be unclotted blood samples. All samples should be immediately put at
4°C and sent as soon as possible to the laboratory.
EHV1 infections in horses are very difficult to treat. The virus has a peculiar
pathogenesis which makes it hard to interfere. Until now, no antivirals have been licensed
in Belgium for the use in horses, limiting treatment to intensive supportive therapy only
(van Maanen, 2002), along with supportive antibiotics and anti-inflammatory drugs.
During several naturally occurring outbreaks of EHV1-induced neurological disease,
attempts were made to prevent infection or treat horses with severe symptoms using
antiviral drugs (Friday et al., 2000; Henninger et al., 2007; van der Meulen et al., 2003a).
Acyclovir and its prodrug valacyclovir are the most popular drugs in this matter. During
the outbreak in Antwerp, a limited number of animals were treated orally with
valacyclovir. Treated horses received an oral dose of valacyclovir of 40 mg/kg body
weight, three times a day. This treatment was initiated as soon as possible. Preferentially a
treatment should be started before paralysis occurs in order to stop this process. However,

General discussion
135
evaluation of a drug in clinical cases will always remain difficult as animals are mostly
not treated under identical conditions and EHV1-negative animals need to be present as
controls to exclude any influence of the immune system. Despite these limitations, it was
observed in this outbreak that valacyclovir-treated animals seemed to recover more
rapidly from clinical symptoms during treatment than non-treated animals. Treatments
with anti-inflammatory drugs and antibiotics to prevent secondary bacterial infections
seemed to contribute to a better recovery. Finally, intensive care greatly improved the
chance of recovery in the severely affected horses, emphasizing the benefit of such
treatment.
The development of a fully protective vaccine against EHV1 would be the most
efficient way to eradicate EHV1-induced problems from the horse population. Until an
efficient vaccine has been found, management procedures are still the most important
measures to prevent or control an outbreak of EHV1 (Lunn et al., 2009; van Maanen,
2002). During the present outbreak, vaccination of horses against EHV1 was highly
recommended. Within an affected premise, horses without clinical symptoms were
vaccinated. Further, owners of large premises and organisers of competitions were
advised to prohibit contact between their horses and visiting horses, and to allow only
horses on their premises which were vaccinated. To date, only inactivated vaccines are
commercially available in Belgium. Although they can not completely prevent abortion or
nervous system disorders, they seemed to protect horses partially from abortion or
nervous system disorders during the outbreak of EHM. It was observed that morbidity
rates during the outbreak of 2009 were the lowest on the first affected premise where a
correct vaccination protocol was used. Moreover, and as far as we know, virus only
spread to premises where horses were not vaccinated against EHV1, suggesting some
clinical protection of vaccinated horses against severe symptoms (Henninger et al., 2007;
Studdert et al., 2003).
In conclusion, given the peculiar and rapid pathogenesis of EHV1, the development of
a curative antiviral will be a very difficult task, as the points of action of such antiviral are
rather limited. Therefore, the best way to protect horses from severe complications due to
an EHV1 infection, is to prevent them from getting infected. This goal can be achieved in
two ways: i) the development of a fully protective vaccine and/or ii) the development of
an antiviral with a metaphylactic action to protect in-contact horses during the course of

General discussion
136
an outbreak. Until further research is able to fulfil the above mentioned goals,
management procedures are still very important measures.

General discussion
137
References
Allen, G. P. (2002). Epidemic disease caused by equine herpesvirus-1: Recommendations
for prevention and control. Equine Vet Educ 4, 177-183.
Allen, G. P. (2007). Development of a real-time polymerase chain reaction assay for
rapid diagnosis of neuropathogenic strains of equine herpesvirus-1. J Vet Diagn
Invest 19, 69-72.
Allen, G. P. & Bryans, J. T. (1986). Molecular epizootiology, pathogenesis, and
prophylaxis of equine herpesvirus-1 infections. Prog Vet Microbiol Immunol 2,
78-144.
Breathnach, C. C., Yeargan, M. R., Sheoran, A. S. & Allen, G. P. (2001). The
mucosal humoral immune response of the horse to infective challenge and
vaccination with Equine herpesvirus-1 antigens. Equine Vet J 33, 651-657.
Edington, N. & Bridges, C. G. (1990). One way protection between equid herpesvirus 1
and 4 in vivo. Research in veterinary science 48, 235-239.
Friday, P. A., Scarratt, W. K., Elvinger, F., Timoney, P. J. & Bonda, A. (2000). Ataxia and paresis with equine herpesvirus type 1 infection in a herd of riding
school horses. J Vet Intern Med 14, 197-201.
Garré, B., Gryspeerdt, A., Croubels, S., De Backer, P. & Nauwynck, H. (2009). Evaluation of orally administered valacyclovir in experimentally EHV1-infected
ponies. Vet Microbiol 135, 214-221.
Goehring, L. S., Hussey, G. S., Ashton, L. V., Schenkel, A. R. & Lunn, D. P. (2010). Infection of central nervous system endothelial cells by cell-associated EHV-1.
Vet Microbiol doi:10.1016/j.vetmic.2010.08.030.
Gryspeerdt, A. C., Vandekerckhove, A. P., Bannazadeh Baghi, H., Van de Walle, G. R. & Nauwynck, H. J. (2011). Expression of late viral proteins is hampered in
infected nasal mucosal leukocytes but not in epithelial cells during early
pathogenesis of equine herpesvirus type 1 (EHV1) infection. The Veterinary
Journal in press.
Henninger, R. W., Reed, S. M., Saville, W. J., Allen, G. P., Hass, G. F., Kohn, C. W. & Sofaly, C. (2007). Outbreak of Neurologic disease Caused by Equine
Herpesvirus-1 at a University Equestrian Center. J Vet Intern Med 21, 157-165.
Lunn, D. P., Davis-Pointer, N., Flaminio, M. J., Horohov, D. W., Osterrieder, K., Pusterla, N. & Townsend, H. G. (2009). Equine Herpesvirus-1 Consensus
Statement. J Vet Intern Med 23, 450-461.
Maxwell, L. K., Bentz, B. G., Gilliam, L. L., Ritchey, J. W., Holbrook, T. C.,
McFarlane, D., Rezabek, G. B., MacAllister, C. G. & Allen, G. P. (2008). Efficacy of Valacyclovir Against Clinical Disease After EHV-1 Challenge in
Aged Mares. Proceedings of the Annual Convention of the AAEP 54, 198-199.
Pusterla, N., David Wilson, W., Madigan, J. E. & Ferraro, G. L. (2009a). Equine
herpesvirus-1 myeloencephalopathy: a review of recent developments. Vet J 180,
279-289.
Pusterla, N., Mapes, S. & Wilson, W. D. (2008). Use of viral loads in blood and
nasopharyngeal secretions fot the diagnosis of EHV-1 infection in field cases. Vet
Rec 162, 728-729.
Pusterla, N., Wilson, W. D., Mapes, S., Finno, C., Isbell, D., Arthur, R. M. & Ferraro, G. L. (2009b). Characterization of viral loads, strain and state of equine

General discussion
138
herpesvirus-1 using real-time PCR in horses following natural exposure at a
racetrack in California. Vet J 179, 230-239.
Studdert, M. J., Hartley, C. A., Dynon, K., Sandy, J. R., Slocombe, R. F., Charles, J. A., Milne, M. E., Clarke, A. F. & El-Hage, C. (2003). Outbreak of equine
herpesvirus type 1 myeloencephalitis: new insights from virus identification by
PCR and the application of an EHV-1-specific antibody detection ELISA.
Veterinary Record 153, 417-423.
USDA (2007). Equine herpesvirus myeloencephalopathy: A potentially emerging disease.
Available at: http://wwwaphisusdagov/vs/nahss/equine/ehv.
van der Meulen, K., Caij, B., Pensaert, M. & Nauwynck, H. (2006). Absence of viral
envelope proteins in equine herpesvirus 1-infected blood mononuclear cells during
cell-associated viremia. Vet Microbiol 113, 265-273.
van der Meulen, K., Vercauteren, G., Nauwynck, H. & Pensaert, M. (2003a). A local
epidemic of equine herpesvirus 1-induced neurological disorders in Belgium.
Vlaams Diergenskund Tijds 72, 366-372.
van der Meulen, K. M., Gryspeerdt, A. C., Vandekerckhove, A. A., Garre, B. A. & Nauwynck, H. J. (2007). The protective properties of vaccination against equine
herpesvirus 1-induced viremia, abortion and nervous system disorders. Vlaams
Diergenskund Tijds 76, 186-194.
van der Meulen, K. M., Nauwynck, H. J. & Pensaert, M. B. (2003b). Absence of viral
antigens on the surface of equine herpesvirus-1-infected peripheral blood
mononuclear cells: a strategy to avoid complement-mediated lysis. J Gen Virol 84,
93-97.
van Maanen, C. (2002). Equine herpesvirus 1 and 4 infections: an update. Vet Q 24, 58-
78.
Vandekerckhove, A., Glorieux, S., Broeck, W. V., Gryspeerdt, A., van der Meulen, K. M. & Nauwynck, H. J. (2009). In vitro culture of equine respiratory mucosa
explants. Vet J 181, 280-287.
Vandekerckhove, A. P., Glorieux, S., Gryspeerdt, A. C., Steukers, L., Duchateau, L., Osterrieder, N., Van de Walle, G. R. & Nauwynck, H. J. (2010). Replication
kinetics of neurovirulent versus non-neurovirulent equine herpesvirus type 1
strains in equine nasal mucosal explants. J Gen Virol 91, 2019-2028.
von Einem, J., Smith, P. M., Van de Walle, G. R., O'Callaghan, D. J. & Osterrieder, N. (2007). In vitro and in vivo characterization of equine herpesvirus type 1
(EHV-1) mutants devoid of the viral chemokine-binding glycoprotein G (gG).
Virology 362, 151-162.
Walker, C., Love, D. N. & Whalley, J. M. (1999). Comparison of the pathogenesis of
acute equine herpesvirus 1 (EHV-1) infection in the horse and the mouse model: a
review. Vet Microbiol 68, 3-13.

Summary
139
SUMMARY


Summary
141
SUMMARY
Equine herpesvirus 1 (EHV1) is the most important infectious cause of abortion
worldwide. Infection can also result in severe nervous system disorders with frequent
fatal outcome. Horses are infected via the respiratory system. After local replication in the
upper respiratory tract, virus spreads via a cell-associated viremia to the uterus and/or the
nervous system. Replication in endothelial cells of these organs can result in abortion or
nervous system disorders. Vaccination with current available vaccines against EHV1 can
not offer full protection against these severe symptoms. Therefore there’s an urgent need
for improvement of existing vaccines or the development of alternative strategies for
prevention or treatment of EHV1 infections. However, the lack of knowledge about the
invasion strategies of EHV1 is an important obstacle. A thorough knowledge about the
pathogenesis of EHV1 is crucial for vaccine development or efficient therapy.
In Chapter 1, an overview of the current knowledge of EHV1 was given. First a short
overview of the common characteristics of EHV1 was given. Further, interactions of the
virus with the cell and with the horse as host were reviewed.
In Chapter 2, the aims of the studies described in the thesis are put forward. The first
aim was to gain a more detailed insight in the invasion strategies of EHV1 strains from
the upper respiratory tract until the origin of a cell-associated viremia. The second aim
was to examine how individual EHV1-infected cells escape from immune effector
mechanisms at the primary site of replication. The third and last aim was to gain more
information about naturally occurring outbreaks in Belgium in terms of risk factors,
protective properties of vaccination, the effect of experimental therapies and diagnosis.
The first aim was addressed in Chapter 3. An experimental inoculation study in
ponies was performed with both a neurovirulent and a non-neurovirulent strain. Two
groups of six ponies were inoculated intranasally with either strain. Clinical signs, nasal
shedding and viremia were evaluated. At early time points post inoculation, one pony of
each group was euthanized. Tissues were collected for titration and immunostainings.
Number and size of EHV1-induced plaques were calculated, and individual EHV1-
infected cells were quantified and characterized. Inoculation with either strain resulted in

Summary
142
nasal shedding and replication in several tissues of the upper respiratory tract. Both
strains replicated in a plaquewise manner in epithelium of the nasal mucosa, but
replication in epithelium of the nasopharynx was largely limited to non-neurovirulent
EHV1. Plaques were never able to cross the basement membrane, but individual infected
cells were noticed in the connective tissue of all examined tissues for both strains. The
total number of these cells however, was 3-7 times lower with non-neurovirulent EHV1
compared to neurovirulent EHV1. CD172a+ cells and CD5
+ lymphocytes were important
target cells for both strains. Interestingly, in lymph nodes, B-lymphocytes were also
important target cells for EHV1, irrespective of the strain. Viremia was detected very
early post inoculation and carrier cells were mainly CD172a+ for both strains.
The second aim was addressed in Chapter 4. Since both epithelial and mononucleair
cells appeared to be crucial cell types during the pathogenesis of EHV1, we studied if
EHV1 uses immuno-evasive techniques in these cell types during the early stages of
pathogenesis. Therefore, the expression of viral late proteins gB, gC, gD and gM in these
target cells was studied in different experimental models. Firstly, RK13 cells and
peripheral blood mononuclear cells were infected with EHV1 in vitro. Secondly, the
recently established equine nasal mucosal explant system was used and thirdly, nasal
mucosal tissues of experimentally EHV1-infected ponies were analyzed. Our salient
findings were that for all three systems tested, the late proteins gB, gC, gD and gM were
expressed on EHV1-infected epithelial cells. On the contrary, gB and gM expression was
observed in 60 to 90% of infected mononuclear cells, whereas expression of gC and gD
on these cells was hampered, because only 20% of EHV1-infeced mononuclear cells
showed gC and gD expression. These results indicate that the expression of certain viral
proteins depends upon the cell type infected and that a hampered expression could be a
mean of EHV1 to stay invisible for the immune system during the early pathogenesis of
EHV1.
The third aim was addressed in Chapter 5. To gain more information about EHV1
problematics in Belgium, a recent outbreak of EHV1-induced neurologic disease in
Flanders which involved at least 13 premises, was both clinically as genetic analyzed. A
total of 212 horses was exposed to EHV1 and a morbidity of 26% was seen. The
incidence of neurological disorders in the affected horses was 43%. Furthermore, the

Summary
143
outbreak was characterized by the rapid occurrence of ataxia and paralysis immediately
after disappearance of fever. Morbidity and virus spread was remarkably higher at
premises were no or an incorrect vaccination schedule was used. Mares seemed to be
more sensible for the development of nervous system disorders with fatal outcome. A
positive influence of valacyclovir treatment during the course of disease was seen if
treatment was initiated immediately after appearance of nervous system disorders. EHV1
was diagnosed in each case by means of virus isolation and/or the detection of
seroconversion. Isolated virus was classified as neurovirulent EHV1, with al isolates
belonging to group 4 upon sequencing in the ORF30 and ORF68 region. The latter
confirms the suspicion of the outbreak spreading from the first premise.
Following general conclusions can be made from the studies performed in the
present thesis:
1. EHV1 has a rapid course after infection and replicates in the epithelium of the
upper respiratory tract in a plaquewise manner. These plaques do not penetrate the
basement membrane, but individual infected cells can be observed in the
connective tissue. These cells are mainly CD172a+ cells and CD5
+ lymphocytes.
At the time point of appearance of these individual infected cells, a cell-associated
viremia could be noticed, harbouring the virus mainly in CD172a+ cells.
2. In the beginning of infection individual infected cells are already in a blocked
stage, allowing them to elope the immune system. Contrary to infected epithelial
cells who expressed al tested glycoproteins, individual infected cells only express
gC and gD in only 20% of the cases.
3. As observed in America, equine herpes myelitis caused by infection with EHV1
seems to be emerging in Belgium. After the large outbreak in 2009, also in 2010
two small outbreaks of EHM were observed, showing a larger number of horses
with nervous system disorders and a fast spread of the virus. Again, this points out
the necessity of a thorough research into the pathogenesis of EHV1, in order to
develop efficient vaccines and antiviral therapy. Vaccinated horses seem to have a
certain clinical protection against the appearance of severe symptoms. Diagnosis
is possible by means of virus isolation and seroconversion. If treatment with

Summary
144
valacyclovir is started at an early time point in infection, horses seem to have a
quicker and more complete recovery.

Samenvatting
145
SAMENVATTING


Samenvatting
147
SAMENVATTING
Infectie met het equiene herpesvirus 1 (EHV1) is wereldwijd de belangrijkste
infectieuze oorzaak van abortus bij paarden. Infectie kan ook resulteren in ernstige
zenuwstoornissen met veelal fatale afloop. Paarden worden geïnfecteerd via het
ademhalingsstelsel. Na lokale vermeerdering in de bovenste luchtwegen verspreidt het
virus zich via een celgeassocieerde viremie naar baarmoeder en/of zenuwstelsel. Daar kan
vermeerdering van EHV1 ter hoogte van endotheelcellen resulteren in abortus of
zenuwstoornissen. Vaccinatie met de huidig beschikbare vaccins tegen EHV1 kan geen
volledige bescherming bieden tegen deze ernstige symptomen. Er is dus nood aan
verbetering van vaccins of en het ontwikkelen van alternatieve strategieën voor preventie
of behandeling van EHV1 infecties. Een belangrijke hinderpaal hierbij is het gebrek aan
kennis over de invasie van EHV1. Een grondige kennis van de pathogenese van EHV1 is
onontbeerlijk voor de ontwikkeling van efficiënte vaccins of een doelgerichte therapie.
In Hoofdstuk 1 werd een overzicht gegeven van de huidige kennis over EHV1. Eerst
werd een beknopte uitleg gegeven over de algemene eigenschappen van het virus.
Vervolgens werden de interacties van het virus met de cel en met het paard als gastheer
besproken.
In Hoofdstuk 2 werden de doelstellingen van de verschillende studies uiteengezet. De
eerste doelstelling bestond erin om een meer gedetailleerd inzicht te verkrijgen in de
invasie van EHV1 stammen van de bovenste ademhalingswegen tot aan de
celgeassocieerde viremie. De tweede doelstelling was na te gaan hoe individuele EHV1-
geïnfecteerde cellen er in slagen te ontsnappen aan de immuun effector mechanismen op
de plaats van primaire vermeerdering. De derde en laatste doelstelling was om het verloop
van natuurlijke infecties met EHV1 beter in kaart te brengen wat betreft risicofactoren,
mogelijke bescherming door middel van vaccinatie, het effect van experimentele
therapieën en diagnostische middelen.
In Hoofdstuk 3 kwam de eerste doelstelling aan bod. Een experimentele inoculatie
met zowel een neurovirulente als een niet-neurovirulente stam van EHV1 werd
uitgevoerd bij pony’s. Twee groepen van 6 pony’s werden intranasaal geïnoculeerd met

Samenvatting
148
één van beide stammen. Klinische tekenen, uitscheiding via de neus en viremie werden
geëvalueerd. Tijdens de eerste dagen na infectie werd telkens 1 pony van elke groep
geëuthanaseerd. Er werden weefsels verzameld voor titratie en immunofluorescentie
kleuringen. Het aantal en de grootte van EHV1-geïnduceerde plaques werd bepaald en
EHV1-geïnfecteerde individuele cellen werden gekwantificeerd en gekarakteriseerd.
Inoculatie met elke stam resulteerde in virusuitscheiding via de neus en een replicatie in
verschillende weefsels van de bovenste ademhalingswegen. Beide stammen
vermeerderden door middel van plaque formatie in de mucosa van de neus, maar
replicatie in het epitheel van de nasopharynx was grotendeels gelimiteerd tot niet-
neurovirulent virus. Plaques doorbraken op geen enkel moment de basaalmembraan, maar
voor beide stammen werden individueel geïnfecteerde cellen gevonden in het
bindweefsel. Het aantal individuele cellen was echter 3-7 maal lager voor niet-
neurovirulent EHV1 vergeleken met neurovirulent EHV1. CD172a+ cellen en CD5
+
lymfocyten waren de belangrijkste doelwitcellen voor beide stammen. In lymfeknopen
bleken B-lymfocyten ook belangrijke dragercellen te zijn voor EHV1, onafhankelijk van
welke stam gebruikt werd. Viremie werd reeds vroeg na infectie vastgesteld en de
geïnfecteerde cellen bleken voor beide stammen vooral CD172a+ te zijn.
In Hoofdstuk 4 kwam de tweede doelstelling aan bod. Gezien het feit dat zowel
epitheliale als mononucleaire cellen cruciale celtypes bleken te zijn gedurende de
pathogenese van EHV1, werd in dit hoofdstuk nagegaan of EHV1 in deze cellen reeds
vroeg in de pathogenese gebruik maakt van immuno-evasieve technieken. Daarom werd
de expressie van de late virale proteïnen gB, gC, gD en gM in deze doelwitcellen
nagegaan in verschillende experimentele modellen. Ten eerste werden RK13 cellen en
perifere bloed mononucleaire cellen in vitro geïnfecteerd met EHV1. Ten tweede werd
het onlangs gerealiseerde equiene nasale explant systeem gebruikt. Ten derde werd de
expressie nagegaan in nasale mucosa van experimenteel geïnfecteerde pony’s. In elk van
de drie geteste systemen bleken de late eiwitten gB, gC, gD en gM volledig tot expressie
te komen in EHV1-geïnfecteerde epitheliale cellen. Dit was in tegenstelling tot
mononucleaire cellen, waar gB en gM expressie werd gezien bij 60 tot 90% van de
geïnfecteerde cellen, terwijl gC en gD expressie enkel werd gezien bij 20% van de
geïnfecteerde cellen. De expressie van bepaalde virale proteïnen lijkt dus afhankelijk te

Samenvatting
149
zijn van het celtype dat geïnfecteerd wordt en het lijkt er op dat EHV1 op deze manier
kan verborgen blijven voor het immuun systeem in vroege stadia van infectie.
In Hoofdstuk 5 kwam de derde doelstelling aan bod. Om meer informatie te
verkrijgen over de EHV1 problematiek in België, werd een recente uitbraak van EHV1-
geïnduceerde neurologische stoornissen in Vlaanderen, waarbij minstens 13 bedrijven
werden getroffen, zowel klinisch als genetisch grondig onderzocht. In totaal werden 212
paarden blootgesteld aan EHV1 en werd een morbiditeit van 26% gezien. De incidentie
van neurologische stoornissen bij aangetaste paarden bedroeg 43%. Verder werd deze
uitbraak gekarakteriseerd door het snelle optreden van ataxie en paralyse na een korte
periode van koorts. De morbiditeit en spreiding van het virus was merkelijk hoger op
bedrijven waar niet of niet correct werd gevaccineerd. Merries leken gevoeliger voor het
ontwikkelen van zenuwstoornissen met fatale afloop. Indien onmiddellijk na het
verschijnen van zenuwstoornissen gestart werd met een valacyclovir behandeling, werd
een positief effect gezien op het ziekteverloop. Diagnose was in alle gevallen mogelijk
door ofwel virusisolatie uit perifere bloed mononucleaire cellen ofwel bepaling van
seroconversie bij gepaarde serum stalen. Het geïsoleerde virus werd geclassificeerd als
neuropathogeen, behorende tot groep 4 na sequenering in de ORF30 en ORF68 regio. Dit
laatste bevestigt het vermoeden dat de uitbraak verspreid werd vanuit het eerste
aangetaste bedrijf.
Op basis van de studies uitgevoerd in deze thesis kunnen de volgende algemene
conclusies naar voor gebracht worden:
1. EHV1 kent een snel verloop na infectie en vermeerdert in het epitheel van de
bovenste ademhalingswegen door middel van plaque formatie. Deze plaques zijn
niet in staat om de basaalmembraan te doorbreken, maar individueel geïnfecteerde
cellen kunnen worden teruggevonden in het bindweefsel. Deze cellen zijn vooral
CD172a+ cellen en CD5
+ lymfocyten. Op het moment van het verschijnen van
deze individueel geïnfecteerde cellen kon een celgeassocieerde viremie
waargenomen worden, waarbij het virus zich vooral in CD172a+ cellen bevindt.
2. Reeds in het begin van de infectie bevinden individueel geïnfecteerde cellen zich
in een geblokkeerde staat, waarbij ze kunnen ontsnappen aan het immuun
mechanisme. In tegenstelling tot geïnfecteerde epitheelcellen die alle geteste

Samenvatting
150
glycoproteines tot expressie brachten, werden in de individueel geïnfecteerde
leukocyten enkel in 20% van de gevallen gC en gD tot expressie gebracht.
3. Naar analogie met gebeurtenissen in Amerika, lijkt ook in België equiene herpes
myelitis door EHV1 in opmars. Na een grote uitbraak in 2009, waren er ook twee
kleine uitbraken in 2010, waarbij het voorkomen van zenuwstoornissen in een
groter percentage van de gevallen optrad en de spreiding sneller verliep. Dit
benadrukt nogmaals het belang van grondig onderzoek naar de pathogenese van
EHV1, voor gerichte vaccin- en therapie ontwikkeling. Gevaccineerde paarden
lijken een bepaalde graad van klinische bescherming te bezitten tegenover het
optreden van ernstige symptomen. Diagnose is mogelijk door middel van
virusisolatie en seroconversie. Bij het tijdig starten van een behandeling met
valacyclovir lijken paarden sneller en vollediger te herstellen.

Curriculum Vitae
151
CURRICULUM VITAE


Curriculum Vitae
153
CURRICULUM VITAE
Annick Gryspeerdt werd geboren op 3 april 1981 te Roeselare. Na het beëindigen
van het secundair onderwijs, richting Latijn-Wetenschappen aan het Bisschoppelijk
Lyceum der Grauwe Zusters te Roeselare, begon ze in 1999 met de studies
Diergeneeskunde aan de Universiteit Gent. In 2005 behaalde ze het diploma van
dierenarts met onderscheiding.
Onmiddellijk na het beëindigen van haar studies trad zij in dienst bij de vakgroep
Virologie, Parasitologie en Immunologie bij het laboratorium voor Virologie. Ze
verrichtte er een onderzoeksstudie getiteld “De pathogenese van EHV1 bij paarden bij
experimentele inoculaties en veldstudies”. Naast het verrichten van onderzoek, was
Annick ook verantwoordelijk voor het verder uitbreiden van de diagnostiek van virale
ziekten bij het paard en het routinematig uitvoeren van deze diagnostiek. Tevens was ze
betrokken bij het praktische onderricht van laatstejaars studenten in de optierichting
paard.
Annick Gryspeerdt is auteur en mede-auteur van verschillende wetenschappelijke
publicaties in internationale tijdschriften. Ze nam eveneens deel aan meerdere nationale
en internationale congressen en was verantwoordelijk voor het geven van verschillende
lezingen, waaronder een opleidingsnamiddag voor dierenartsen en sensibilisatie van
paardeneigenaren.


Bibliography
155
BIBLIOGRAPHY


Bibliography
157
BIBLIOGRAPHY
Publications in international journals
Gryspeerdt, A. C., Vandekerckhove, A. P., Garré, B., Barbé, F., Van de Walle, G. R. &
Nauwynck, H. J. (2009). Differences in replication kinetics and cell tropism
between neurovirulent and non-neurovirulent EHV1 strains during the acute phase
of infection in horses. Vet Microbiol 142, 242-253.
Gryspeerdt, A. C., Chiers, K., Govaere, J., Vercauteren, G., Ducatelle, R., Van de Walle,
G. R. & Nauwynck, H. J. (2009). Neonatal foal death due to infection with equine
arteritis virus in Belgium. Vlaams Diergen Tijds 78, 189-193.
Gryspeerdt, A. C., Vandekerckhove, A. P., Bannazadeh Baghi, H., Osterrieder, N., Van
de Walle, G. R. & Nauwynck H. J. (2011). Expression of late proteins is hampered
in infected nasal mucosal leukocytes but not in epithelial cells during early
pathogenesis of equine herpesvirus type 1 (EHV1) infection. Vet J (accepted)
Gryspeerdt, A. C., Vandekerckhove A. P., Van Doorsselaere, J., Van de Walle, G. R. &
Nauwynck, H. J. (2011). Description of an unusually large outbreak of nervous
system disorders caused by equine herpesvirus 1 (EHV1) in 2009 in Belgium.
Vlaams Diergen Tijds 80, 139-145.
Van der Meulen, K. M., Gryspeerdt, A. C., Vandekerckhove, A. P., Garré, B. A. &
Nauwynck, H. J. (2008). The protective properties of vaccination against equine
herpesvirus 1-induced viremia, abortion and nervous system disorders. Vlaams
Diergen Tijds 76, 186-194.
Garré, B. A., Gryspeerdt, A. C., Croubels, S., De Backer, P. & Nauwynck, H. J. (2009).
Evaluation of orally administered valacyclovir in experimentally EHV1-infected
ponies. Vet Microbiol 135, 214-221.

Bibliography
158
Garré, B., Shebany, K., Gryspeerdt, A. C., Baert, K., van der Meulen, K., Nauwynck, H.
J., Deprez, P., De Backer, P. & Croubels, S. (2007). Pharmacokinetics of
acyclovir after intravenous infusion of acyclovir and after oral administration of
acyclovir and its prodrug valacyclovir in healthy adult horses. Antimicrob Agents
Chemother 51, 4308-4314.
Vandekerckhove, A. P., Glorieux, S., Gryspeerdt, A. C., Steukers, L., Duchateau, L.,
Osterrieder, N., Van de Walle, G. R. & Nauwynck, H. J. (2010). Replication
kinetics of neurovirulent versus non-neurovirulent equine herpesvirus type 1
strains in equine nasal mucosal explants. J Gen Virol 91, 2019-2028.
Vandekerckhove, A. P., Glorieux, S., Gryspeerdt, A. C., Steukers, L., Van Doorsselaere
J., Osterrieder N., Van de Walle, G. R. & Nauwynck, H. J (2011). Equine
alphaherpesviruses (EHV) differ in their efficiency to infect mononuclear cells
during early steps of infection in nasal mucosa explants. Vet Mic (in press).
Vandekerckhove, A. P., Glorieux, S., Van Den Broeck, W. V., Gryspeerdt, A. C., van
der Meulen, K. M. & Nauwynck, H. J. (2009). In vitro culture of equine
respiratory mucosa explants. Vet J. 181, 280-287.
Oral presentations
Vandekerckhove, A. P., Van den Broeck, W., van der Meulen K. M., Glorieux, S.,
Gryspeerdt, A. C. & Nauwynck, H. J. (2007). Replication of equine herpesvirus
1 in equine respiratory mucosa explants. 32nd International Herpesvirus
Workshop, Asheville, North Carolina, VS.
Gryspeerdt, A. C., Vandekerckhove, A. P. & Nauwynck, H. J. (2008). Replication of
equine herpesvirus 1 in tissues of the upper respiratory tract during the acute phase
of infection in horses. 33rd International Herpesvirus Workshop, Estoril, Portugal.
Gryspeerdt, A. C., Vandekerckhove, A. P., Van de Walle, G. R. & Nauwynck H. J.
(2009). Differences between the replication of neurovirulent en non-neurovirulent

Bibliography
159
EHV1 strains in the upper respiratory tract in vivo. 34th
International Herpesvirus
Workshop, Ithaca, New York, VS.
Vandekerckhove, A. P., Gryspeerdt, A. C., Glorieux, S., Van de Walle, G. R. &
Nauwynck, H. J. (2009). Replication of non-neurovirulent versus neurovirulent
equine herpesvirus type 1 strains: a study in equine respiratory mucosa explants.
34th
International Herpesvirus Workshop, Ithaca, New York, VS.
Gryspeerdt, A. C., Vandekerckhove, A. P., Van de Walle, G. R. & Nauwynck H. J.
(2009). Differences between the replication of neurovirulent en non-neurovirulent
EHV1 strains in the upper respiratory tract in vivo. Summer Course on Infection
and Immunity, 29th
– 30th
September, Ghent, Belgium.
Vandekerckhove, A. P., Gryspeerdt, A. C., Glorieux, S., Van de Walle, G. R. &
Nauwynck, H. J. (2009). Replication of non-neurovirulent versus neurovirulent
equine herpesvirus type 1 strains: a study in equine respiratory mucosa explants.
Symposium of the Belgian Society for Microbiology, 11th
December, Brussels,
Belgium.
Gryspeerdt, A. C. & Nauwynck, H. J. (2009). Prevalence and occurrence of clinical
EHV1 in Flanders, Belgium. Equine Influenza and Herpesvirus – Current Disease
Situation, 16th
October, Berlin, Germany.
Gryspeerdt, A. C. (2010). Voorkomen, behandeling en preventie van rhinopneumonie in
Vlaanderen. Studienamiddag rond virale ziekten bij het paard, Vlaams
Paardenloket, 27th
March, Nazareth, Belgium.
Gryspeerdt, A. C. & Nuytten, J. (2010). Welke lessen kunnen getrokken worden uit de
recente uitbraken van virale ziekten bij het paard? Belgian Equine Practitioners
Society (BEPS), 27ste
studienamiddag, 13th
November, Leuven, Belgium.

Bibliography
160
Gryspeerdt, A. C. (2010). Voorkomen, behandeling en preventie van rhinopneumonie in
Vlaanderen. Studienamiddag rond virale ziekten bij het paard, Vlaams
Paardenloket, 4th
December, Lummen, Belgium.
Gryspeerdt, A. C. (2011). Rhinopneumonie: waarheden en onwaarheden over recente
uitbraken. Baanpraktijk de Morette, 3th
March, Essene, Belgium.
Abstracts
Vandekerckhove, A. P., Van den Broeck, W., van der Meulen K. M., Glorieux, S.,
Gryspeerdt, A. C. & Nauwynck, H. J. (2007). Replication of equine herpesvirus
1 in equine respiratory mucosa explants. Symposium of the Belgian Society for
Microbiology, 23rd
November, Brussels, Belgium.
Gryspeerdt, A. C., Vandekerckhove, A. P. & Nauwynck, H. J. (2008). Replication of
equine herpesvirus 1 in tissues of the upper respiratory tract during the acute phase
of infection in horses. Symposium of the Belgian Society for Microbiology, 12th
December, Brussels, Belgium. BSM award for excellent poster
Gryspeerdt, A. C.*, Vandekerckhove, A. P.*, Van de Walle, G. R. & Nauwynck, H. J.
(2009). In vitro respiratory mucosa explants are useful tools to study respiratory
tract diseases in horses. 3d ESVV Veterinary Herpesvirus Symposium, Insel
Riems, Germany. * equally contributed

Dankwoord
161
DANKWOORD


Dankwoord
163
DANKWOORD
Na ettelijke jaren van hard werken, bloed, zweet, tranen, maar ook veel memorabele
momenten, ligt het hier dan eindelijk: mijn doctoraat. Het is moeilijk te vatten wat een
grote waarde dit kleine boekje heeft. Na al die jaren verdienen heel wat mensen een
woord van dank en dat in het ongetwijfeld meest gelezen hoofdstuk van elk doctoraat…
Eerst en vooral wil ik hier graag mijn promotoren danken. Hans, jou wil ik graag
bedanken om me de kans te geven in jouw laboratorium te werken. Ik denk dat er weinig
werkplaatsen zijn waar de sfeer tussen de werknemers zo goed zit. Ik denk ook dat er
maar weinig manieren zijn om jou te beschrijven. Zelden heb ik zo een gedreven en
hyper-enthousiaste wetenschapper meegemaakt. Je bent echt een uniek persoon! Ik hoop
dat het labo virologie nog lang zo mag groeien en bloeien. Karen, jij was de persoon die
me geïntroduceerd heeft in de wondere wereld van de rhinopneumonie. Met jouw hulp is
de IWT-beurs binnengehaald en kon mijn onderzoek echt starten. Je hebt het werk niet
kunnen afmaken, maar je hebt het startschot gegeven zodat ik op eigen benen kon staan.
Mijn schrijfsels zijn nog steeds niet top, maar ik zal me altijd de verbeterde versies met
kleurcodes herinneren die ik in het begin van jou terugkreeg. Gerlinde, jou ben ik heel
veel dankbaarheid verschuldigd. We werden gewaarschuwd dat er een felle madam in
aantocht was, die de tucht in onze paardengroep moest herstellen. Die felle madam bleek
echter een heel aangename en gedreven onderzoeker te zijn, die er als eerste in slaagde
om een artikel van mij gepubliceerd te krijgen. Honderden schrijfsels heb je verbeterd en
hoewel ik nog steeds geen bestseller auteur kan genoemd worden, is er toch enige
verbetering merkbaar. Verder stond je altijd voor me klaar toen het weer eens moeilijk
ging met mijn doctoraat. Dankzij jouw motivering ligt dit boekje hier vandaag! En ik
denk dat ik zeker mag zeggen dat je goed bezig bent, Professor Van de Walle! Ik ben er
zeker van dat je een prachtig labo zal uitbouwen! Herman, ik wil je graag bedanken voor
je geloof in mijn onderzoek en praktijkgerichte aanpak. Bedankt voor je luisterend oor
tijdens moeilijke momenten en voor de aangename congressen wanneer je van de partij
was!
Ook de overige leden van de begeleidings- en examencommissie zou ik hartelijk
willen bedanken voor de tijd die ze vrijgemaakt hebben en voor hun waardevolle en

Dankwoord
164
opbouwende kritieken: Prof. dr. P. Deprez, Prof. dr. J. Kydd, dr. K. van Maanen, mevr. B.
Caij, dr. K. van der Meulen, dr. J. Nuytten en dr. T. Picavet.
Het agentschap voor Innovatie door Wetenschap en Technologie wil ik bedanken voor
de financiële steun die het mogelijk heeft gemaakt om dit doctoraat tot een goed einde te
brengen.
Eerst wil ik even de moeite nemen om de fantastische laboranten en technische
krachten van het labo te bedanken. Zij zijn het hart en ziel van de dienst en zorgen er
dagelijks voor dat alles op wieltjes loopt, wat met al dat volk geen evidente taak kan
genoemd worden. Chris, ondertussen ben je op pensioen, maar ik zal al het werk dat je
voor mij gedaan hebt nooit vergeten. Je was de onbetwiste kampioen wat betreft het
isoleren van paardenleukocyten! Bovendien was je een zeer aangename persoon en
hebben we samen ook veel plezier gehad. Ik hoop dat je geniet van je pensioen!
Vervolgens hebben we Lieve. Ondanks het feit dat we nooit echt rechtstreeks hebben
samengewerkt, kan ik me nog goed herinneren dat je me in het begin ogenblikkelijk hebt
duidelijk gemaakt dat het een slecht idee was om mijn voorraden slechts op de dag van
mijn experiment te checken. Geregeld ben ik met rode kaken tot bij jou gekomen om te
zeggen dat mijn medium weer eens op was of beschimmeld was. Ik heb dankzij je strenge
aanpak echter zeer snel geleerd om me wat beter te organiseren, waarvoor dank! Chantal,
jou moet ik bedanken voor het opgroeien van ontelbare virusstocks, want zonder virus
geen experimenten! Vervolgens hebben we Carine. Wij hebben heel veel samengewerkt
en het is altijd met het grootste plezier geweest. Je bent een zeer aangename en
hardwerkende vrouw en niets was ooit te veel voor jou. Ik kan zelfs al niet meer tellen
hoeveel weefsels je vakkundig getitreerd hebt. Het enthousiasme waarmee we de
zoveelste diagnose verwerkten was legendarisch! En het plezier dat we hadden als we
weer iets geïsoleerd hadden na veel problemen maakte het de moeite waard ☺. Ik hoop
dat de diagnostiek even goed blijft draaien en ik hoop dat we daar in de toekomst nog veel
contact over hebben. Nele en Melanie hebben me ook geregeld uit de brand geholpen.
Nele door het biotinyleren van ontelbare antistoffen en Melanie met het ingeven van
diagnose uitslagen en het inspringen als het praktische werk mij weer eens boven het
hoofd groeide. Dries heeft de ondankbare taak gekregen om me af en toe moleculair te
begeleiden. Helaas voor hem ben ik er zeker van dat ik op dat vlak vervloekt ben.
Bedankt voor de moeite die je er toch telkens weer instak ondanks de hardnekkige

Dankwoord
165
tegenvallende resultaten. Jan Van Doorsselaere wil ik bedanken voor alle sequeneringen
die hij voor mij heeft uitgevoerd. Verder hebben we de technische krachten nog, waarvan
vooral Geert een eervolle vermelding verdient. Je hebt me op hele vroege uren geholpen
met de paardjes en zulke goede hulp heb ik nooit meer gehad. Ik hoop dat je gelukkig
bent op je nieuwe werk! Fernand is ondertussen ook al aan zijn pensioen begonnen, maar
heeft ook dikwijls zijn steentje bijgedragen bij diverse experimenten, waarvoor mijn
dank! Bart en Zeger, jullie hebben de moeilijke taak gekregen dit duo op te volgen. Ik
hoop dat jullie je draai vinden en genieten van jullie job! Marijke mag ook zeker niet
vergeten worden, de persoon die als moeilijke taak heeft om het labo netjes te houden.
Bedankt voor het onderhouden van mijn bureautje en de vele leuke gesprekken! Mieke,
Ann, Gert en Louise, bedankt voor alle praktische dingen die jullie dag na dag moeten
zien te regelen. Jullie zijn echt de duizendpoten van het labo! Dirk, bedankt voor het
oplossen van vele computergerelateerde problemen, maar toch ben ik nog steeds heel erg
blij met mijn PC’tje ☺. Mocht ik mij in de verre toekomst ooit eens een Mac aanschaffen,
dan zal ik eens aan jou terug denken!
En dan hebben we de collega’s. Er zijn er in die 6 jaar zoveel gekomen en gegaan dat
ik echt hoop dat ik niemand vergeten ben!
Ik ben mijn tijd op het labo gestart in de paardenbureau. Annelies, jij was mijn collega
van het eerste uur, eerst als laatstejaars, vervolgens als Phd student. Ik herinner me vooral
onze vreugdedans in het bureau van Hans toen we vernamen dat we beide onze IWT-
beurs gehaald hadden. Verder heb je me ontelbare keren geholpen met
paardenexperimenten. Ik wil je bedanken voor alle leuke jaren. Ik hoop dat je als “the last
one standing” de eer van de paardengroep hoog kunt houden en dat je het EHV onderzoek
nieuw leven kan inblazen. Ik wens je een prachtig leven toe met je allerliefste Lieven en
hoop dat jullie heel gelukkig worden samen! Filip had de ondankbare taak om als enige
man ons bureautje te bevolken. Ik moet zeggen, ik ken niemand waarbij de spreuk “stille
waters, diepe gronden” zo goed van toepassing was! Je kon je mannetje heel goed staan
tussen al dat vrouwengeweld! Zelfs de Flair werd op de duur opgenomen in je
literatuurlijst ☺. Hartelijk dank voor alle gezellige etentjes, gesprekjes en je hulp met de
paardjes. Ik hoop dat we elkaar in de toekomst nog geregeld eens zien en ik wens je veel
geluk toe met de bouw van je droomhuis!

Dankwoord
166
Na vier jaar werd het tijd voor een nieuw onderkomen, wat werd gevonden in de
beroemde “eerste bureau”. Sarah G., als ancien van deze bureau heb je de nieuwelingen
hartelijk ontvangen! Ik wil je bedanken voor de talloze gezellige etentjes en het
onvergetelijke congres dat we samen in Portugal hebben mogen meemaken. Ik hoop dat
alles vlot loopt met je onderzoek en dat je heel gelukkig mag worden met je toekomstige
man! Amy, you arrived a bit later, but you are really the sweetest girl in the office! I am
really glad that your husband finally arrived in Belgium and I hope you will enjoy your
time here in Belgium. I want to thank you for all the nice Chinese gifts I received from
you and I wish you all the best with your research! Hossein, you got the very difficult task
to find our well-hidden equine dendritic cells. I hope you will find them eventually! Keep
practising with your bike and then you will avoid becoming as fat as I once was ☺.
Sabrina! You are the most energetic person I ever met! It is really a delight to have such
person in the office! With you, nothing will ever be boring! I hope you will succeed in
your Phd, you really deserve it after all the work you’ve put in to it. Enjoy your beautiful
daughter and always remain your cheerful self! Mieke, jij bent stiekem binnengeglipt toen
ik op zwangerschapsverlof was, maar ik heb de indruk dat je je plaatsje al goed veroverd
hebt! Ik wens je veel geluk toe in je onderzoek! Lennert, tja, er zijn zoveel woorden
waarmee jij beschreven kunt worden, maar saai is er in elk geval niet één van! Je brengt
altijd plezier in het bureau en bent altijd in voor een grapje. Ik ben blij dat er nog een
echte veearts in het labo aanwezig is! Veel succes in Amerika en ik ben zeker dat jij er op
professioneel vlak wel zal geraken!
Verder heb ik nog een heel tijdelijk onderkomen gevonden in mijn oude bureau
tijdens het “pimpen” van ons nieuw bureau (kunnen jullie nog volgen?), waar ik hartelijk
werd ontvangen door Sjouke en Annebel. Bedankt voor alle leuke en serieuze gesprekken
en Sjouke, bedankt voor de hulp met het streepje aan de bovenkant van mijn pagina’s! Je
bent een ferme madam en je zal er zeker komen in het leven!
Marc, jij verdient ook zeker een eervolle vermelding! Met jou is er altijd tijd voor
plezier en dat al sinds ons eerste jaar in Gent. Ik vond het super dat je onze ploeg op het
labo kwam vergezellen. De sfeer op het labo is er enkel maar op vooruit gegaan! Bedankt
voor alle amusante gesprekken, maar ook voor de steun die ik altijd van jou heb gekregen.
Werp je maar op het schrijven en haal dat doctoraat binnen! Merijn en Wander, jullie wil
ik bedanken voor de talloze vragen die jullie beantwoord hebben naar aanleiding van mijn
moleculaire problemen! Niettegenstaande het feit dat mijn moleculaire vervloeking overal

Dankwoord
167
stokken in de wielen stak, heb ik wel veel nuttige tips gekregen! Wander, het was fijn om
een compagnon te hebben bij de finishing touch van onze doctoraten. Ik hoop dat je in de
toekomst iets meer relax je deadlines zal halen!
Ik denk dat ons labo ondertussen wel gekend is als één van de grootste van de
faculteit diergeneeskunde. Volgens mij is het ook één van die uitzonderlijke plekken waar
de sfeer tussen de werknemers echt super is. Daarom wil ik graag iedereen bedanken die
nog niet aan bod is gekomen, zowel onze inheemse (Matthias, Tim, Veerle, Ytse, Hannah,
Annelike, Dominique, Ben, Lowiese, Sebastiaan, Hanne, Inge, Karen, Miet, Céline,
Thary en Nina) als uitheemse mensen (Sabine, Ilias, Eva, Dipu, Joao, Tù, Uladzimir,
Xiaoyun, Jun, Lang, Liping, Wenfeng, Yu, Zhongfang and Maria) en natuurlijk iedereen
die zijn/haar carrière op het labo nog maar net gestart is! Elk van jullie draagt zijn steentje
bij tot de tweede thuis die het labo voor veel mensen geworden is.
Nog een woordje voor de collega’s die het labo reeds verlaten hebben, maar die zeker
ook een plaatsje in dit dankwoord verdienen. Barbara, ik weet niet zo goed wat ik hier
moet neerschrijven. Ik herinner me nog de eerste keer dat je me moest wegwijs maken in
het labo en je me heel vreemd aankeek omdat je me maar “raar en luid” vond. Maar heel
snel hebben we een gezamenlijke passie ontdekt, die ons tot heel goede vriendinnen heeft
gemaakt. Je hebt me door de moeilijke periode in het labo geholpen en hebt me zowel
privé als werkgerelateerd heel veel plezier bezorgd. We hebben van in de vroege tot de
late uurtjes samen gewerkt en om het met je eigen woorden te zeggen “bedankt voor heel
veel en nog zoveel meer”… Hopelijk vinden jij en Eric jullie droomhuis en kunnen jullie
een prachtig leven uitbouwen! Sarah C., jij was ook één van de meest opmerkelijke
personen in het labo. Ik vind het super dat we nog af en toe contact hebben en hoop dat
het je op alle vlakken voor de wind blijft gaan! Leslie, ik heb grote bewondering voor het
feit dat je zo goed weet wat je wil in het leven en daar ook helemaal voor gaat. Veel
succes met je nieuwe job en bedankt voor alle leuke gesprekken!
Hoewel het minder gebruikelijk is, wil ik ook alle paarden en pony’s bedanken die
een bijdrage geleverd hebben aan dit doctoraat. Ze hebben me veel plezier bezorgd en een
aantal van hen hebben ondertussen een super thuis gevonden! Een speciale vermelding
wilde ik graag geven aan Cointreau, het veulen dat ons bloed, zweet en tranen gekost
heeft omwille van zijn speelse karakter. Ik denk dat de jacht op Cointreau het meest
memorabele moment was in die 6 jaar. Ik vergeet nooit de hulp van Prof. dr. Gasthuys,
die in volledige jacht outfit, verrekijker en laarzen inclusief, er in slaagde onze ontsnapte

Dankwoord
168
dwarsligger na twee dolle dagen van zijn touw te verlossen… Helaas bleek onze
ontsnapkampioen twee weken later nog steeds niet bereid om in zijn lekker warme
stalletje te komen staan, waardoor er weer een paar dolle jachturen moesten gespendeerd
worden, deze keer met dank aan de sterke mannen van verloskunde. Ondertussen is
Cointreau een flinke vijfjarige geworden en doet hij zijn uiterste best voor zijn nieuwe
baasje! Ik hoop dat hij een lang en gelukkig leven tegemoet gaat!
In de loop van mijn doctoraatsjaren heb ik ook enorm veel steun mogen ontvangen
van mijn collega dierenartsen in de praktijk. Ik wil hen graag bedanken voor alle hulp en
suggesties die ze mij geboden hebben. Ze hielden me met mijn voeten op de grond en
zorgden ervoor dat ik niet vergat waarom virologisch onderzoek voor paarden zo
belangrijk is. Hierbij moet zeker een speciale plaats gegeven worden aan mijn ”mentor”
Jef. Het is door jou dat ik 6 jaar geleden de stap genomen heb om in het onderzoek te
beginnen. Jij was het spreekwoordelijke duwtje in de rug dat ik toen nodig had om er
helemaal in te vliegen. Gedurende al die jaren heb je echter mijn interesse in de praktijk
levendig gehouden en heb je er op gehamerd dat de virologie voor paarden dringend
verder moest onderzocht worden. Ook ons eigen onderzoekje met de honderden swabs en
bloedjes mag daarbij zeker niet vergeten worden. Evenals het ongeduld waarmee je belde
om de uitslag van de zoveelste diagnose. Hierbij wil ik je nog eens uitgebreid danken
voor alles wat je me bijgebracht hebt aan ervaring en “West-Vlaamse levenswijsheden”
en voor de vele uren ontspanning en plezier als het onderzoek mij weer eens te zwaar
werd. Ik hoop dat we in de toekomst nog evenveel praktijkplezier mogen meemaken en
dat er af en toe nog eens een glas champagne geleegd mag worden ☺.
Ook buiten het labo zijn er zoveel vrienden die me geholpen en gesteund hebben.
Lieve Leen, je woont misschien wel ver weg, maar het contact is er na meer dan tien jaar
nog steeds en onze schaarse ontspannende uitstapjes en de deugddoende gesprekken zal
ik nooit vergeten! Julie, in al die jaren zijn er honderden en honderden mails over en weer
gevlogen! Super dat we nog altijd zo een leuk contact hebben en ik hoop dat dat nog jaren
mag blijven duren. Bedankt voor al je steun en begrip! Je bent een super meid en
supermama! Sophie, jij bent er maar later bijgekomen, maar op die korte tijd ben je echt
een goede vriendin geworden. Bedankt voor je altijd luisterende oor en voor alle leuke
etentjes die er al geweest zijn. Veel geluk met je prachtige gezinnetje! En last, but not
least: de familie Demeyer en hun bonte beestenboel. Ik kan mij al lang niet meer

Dankwoord
169
herinneren wanneer ik bij jullie ben binnen gewaaid. Ik weet enkel nog dat het begon met
een brommertje zonder benzine en 100 frank. Zoals jullie het zelf zeggen ben ik bij jullie
ontvangen als vijfde dochter en ik denk dat jullie maar al te goed beseffen wat jullie
daarmee in huis gehaald hebben ☺. Jullie staan stuk voor stuk altijd voor me klaar en
hebben gezorgd voor jaren ontspanning, plezier en serieuze momenten. Dank vanuit de
grond van mijn hart! Miss Domino moet ik nog danken voor de hersenschudding die ze
me bezorgde vlak voor ik mijn project moest indienen… Gelukkig heeft haar zoon dat
ruimschoots goed gemaakt met de ontspannen momenten gedurende de laatste loodjes
van mijn doctoraat!
Wie hier zeker niet mogen ontbreken zijn mijn allerliefste ouders. Zonder jullie steun,
of het nu op financieel vlak was, of door jullie rotsvaste geloof dat ik er wel zou komen in
mijn leven, zou ik nooit staan waar ik nu sta. Jullie hebben me altijd gemotiveerd om
datgene te doen wat ik wilde en hebben ervoor gezorgd dat ik mijn grote droom als
veearts kon waarmaken. Ik ben jullie oneindig dankbaar hiervoor en besef dat er velen
zijn die die steun moeten missen. Mijn zus Charlotte en broertjes Filip en Frederic moet
ik natuurlijk ook danken, want ze hebben heel wat geklaag mogen aanhoren! Ook de
aanhangsels zijn hier vandaag aanwezig en wil ik danken omdat ze onze familie nog
groter en gezelliger gemaakt hebben!
Liefste Jan-Bart, jij hebt het gedurende al die jaren het zwaarst te verduren gekregen.
Je moest het regelmatig ontgelden als de stress weer eens te hoog opflakkerde. Gelukkig
kan je wel tegen een stootje. Ons leven is op korte tijd grondig omgegooid met ons eigen
huisje op een ver afgelegen plaats en onze prachtige, lieve dochter. Nu komt er weer een
verandering met een nieuwe werkuitdaging bij. Ik wil je danken voor al de steun die ik al
gekregen heb en die ik nog steeds krijg, want jij begrijpt wat een carrière vrouw is en
vooral wat een gestresseerde carrièrevrouw betekent! Lieve Aline, sinds jij er bent, weet
ik wat belangrijk is in het leven, weet ik waarom ik het doe en kan ik alles wat beter
relativeren. Met je stralende glimlach kon je me altijd weer doen smelten. Ik hoop dat je
mag opgroeien tot een gelukkige vrouw en dat je evenveel passies mag kennen als je
mama en papa.
Dit dankwoord is zeer uitgebreid geworden en dat maakt me blij, want het betekent
dat er veel mensen zijn die het de moeite waard vonden om me te steunen. Ik hoop vanuit

Dankwoord
170
de grond van mijn hart dat ik niemand ben vergeten, daarom nogmaals: dank aan
iedereen, dit is jullie doctoraat!





























