Embed Size (px)
Citation preview

AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_1File # 01cc
Copyright q American Museum of Natural History 2000 ISSN 0003-0082 / Price $4.40
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY
CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024
Number 3288, 38 pp., 18 figures, 12 tables February 2, 2000
The Os Navicular of Humans, Great Apes, OH 8,Hadar, and Oreopithecus: Function, Phylogeny,
and Multivariate Analyses
ESTEBAN E. SARMIENTO1 AND LESLIE F. MARCUS2
CONTENTS
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2Materials and Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
Canonical Analyses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
Relative Orientations of Navicular Facets . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7Curvature and Relative Size of Navicular Facets . . . . . . . . . . . . . . . . . . . . . . . . 12Variation in Relative Size and Set of Navicular Facets . . . . . . . . . . . . . . . . . . . 17Foot Use and Inherited vs. Epigenetic Characters . . . . . . . . . . . . . . . . . . . . . . . 19Body Weight, Lower-limb size, and Navicular Facet Area . . . . . . . . . . . . . . . . 21Navicular Function and Implied Locomotor Behaviors . . . . . . . . . . . . . . . . . . . 25Fossil Foot Use . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
OH 8 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28Hadar . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29Oreopithecus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
Phylogeny Implicit in Navicular Measurements . . . . . . . . . . . . . . . . . . . . . . . . . 32Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
1 Research Associate, Division of Vertebrate Zoology, American Museum of Natural History.2 Research Associate, Division of Paleontology, American Museum of Natural History; Professor of Biology,
Queens College, Flushing NY, 11367.

2 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_2File # 01cc
ABSTRACT
To clarify fossil hominid behavior and phylogeny, and to test the accuracy of basing thesestudies on single bones, navicular measurements of Olduvai and Hadar hominids, Oreopithe-cus, and a representative sample of humans and great apes were compared. The measurementschosen for comparison quantify the relative orientation, articular area, and curvature of thenavicular facets. The measurements demonstrate that the OH 8 navicular belongs to a rigidfoot with an adducted hallux and a strong commitment to terrestriality. The Hadar navicularsbelong to a foot which lacked a fixed longitudinal plantar arch and had at least a degree ofhallucal opposability comparable to that of mountain gorillas. The Oreopithecus navicularbelongs to a mobile foot with a widely divergent hallux committed to arboreal behaviors.Multiple discriminant and canonical variate analyses of navicular measurements emphasizethe uniqueness of Oreopithecus and the similarities between OH 8 and humans, and betweenHadar and African apes. The African apelike morphology of the Hadar naviculars contradictsthe alleged humanlike morphology of the Hadar pelvis and knee joints. This contradictionunderscores the fallacies inherent in constructing phylogenies on the basis of single bonesand/or fragmentary remains, and of reconstructing locomotor behaviors on the basis of local-ized anatomy.
INTRODUCTION
The hands and feet are those parts of theintegumentary and musculoskeletal structurewhich come directly into contact and interactwith the physical variables of the environ-ment. As such, their morphology closely cor-responds to these variables, responding di-rectly both ontogenetically and phylogeneti-cally to changes in use (i.e., changes in theirinteractions with the physical variables of theenvironment) over time (Sarmiento, 1985,1988, 1994). Because hand and foot anatomyreflect the animal’s environmental interac-tions, they are excellent indicators of behav-ior. Composed of a large number of anatom-ical elements that combine to produce the ap-propriate morphology, hands and feet mayalso provide insights into past behaviors andare useful for reconstructing phylogenies(Schaeffer,1947; Beigert, 1963; Lewis, 1969,1974, 1980a, b, c; Szalay and Decker, 1974;Sarmiento, 1983, 1985, 1988, 1994; Beard etal., 1988; Gebo and Dagosto, 1988). This es-pecially applies to the carpus and tarsuswhere many anatomical elements interact toprovide the appropriate movements and dis-tribute loads. The anatomical complexity ofthe carpus and tarsus dictates that changes inhand or foot use are achieved with relativelyminimal changes in structure, i.e., the organ-ism makes the best use of its inherited anat-omy (Sarmiento, 1985, 1988). Thus, the car-pus and tarsus provide an anatomical record
of those past behaviors which have beenstrongly selected for.
Despite their usefulness, carpal and tarsalbones have been largely ignored by most pa-leoanthropologists when reconstructing earlyhominid behaviors (Lovejoy, 1974, 1978,1988; Robinson, 1972; Zihlman and Bunker,1979; Stern and Susman, 1983; Latimer,1991; McHenry, 1991; Susman and Stern,1991; Fleagle, 1998) or phylogenies (Leakeyet al., 1964; Robinson, 1965; Wolpoff, 1974;Skeleton et al., 1986; Olson, 1978, 1981,1985; White et al., 1981; Chamberlain andWood 1987; Edelstein, 1987; Verhaegen,1990, 1994; Tobias, 1991a, b; McHenry,1996; Strait et al., 1997). This is all the moresurprising considering that carpal and tarsalbones are often the best preserved and mostcomplete skeletal elements found at earlyhominid fossil sites, and are well representedin fossil collections. It is the object of thisstudy, therefore, to analyze the navicular ofhumans, great apes, and some early ‘‘homi-nid’’ and hominoid fossils in both a func-tional and phylogenetic context. Because thegreat ape navicular articulates variably withall of the tarsal bones (Lewis, 1980b, c; Sar-miento, 1994), it should reflect functionaldifferences throughout the tarsus. The pres-ence of complete naviculars at many of theearly hominid fossil sites (Day and Napier,1964; Latimer et al., 1982; Clarke and To-bias, 1995) should provide a phylogenetic

2000 3SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_3File # 01cc
perspective of changes in foot use over time.In addition, analyses and comparisons of asingle bone should serve to test the accuracyof reconstructing behavior or phylogenybased on localized anatomy.
MATERIALS AND METHODS
Naviculars of Olduvai (OH 8) and Hadar(AL 333–47, 333–36) hominids, Oreopithe-cus (Basel #39), and a sample of navicularsfrom each of the great ape species and ofhumans were measured and compared for di-mensions, curvature and relative orientationof facets. Differences in the frequency of ter-restrial behaviors in Virunga gorillas (Gorillab. beringei) relative to western gorillas (G.g. gorilla) justified considering the two asseparate, although a specific designation forVirunga gorillas is arguable (Sarmiento,1994; Sarmiento and Butynski, 1996; Sar-miento et al., 1996). Great ape naviculars arefrom the skeletal collections at the followinginstitutions: American Museum of NaturalHistory, New York; Museum of ComparativeZoology at Harvard University, Cambridge;Field Museum of Natural History, Chicago;National Museum of Natural History, Wash-ington D.C.; Philadelphia Academy of Sci-ences, Philadelphia; Powell Cotton Museum,Birchington; Royal African Museum, Ter-vuren; and Swedish Museum of Natural His-tory, Stockholm. Human naviculars are fromthe Dart collection in the Department ofAnatomy, University of the Witwatersrand,and the Anthropology collections at theAmerican Museum of Natural History. Fossilmaterial is housed in the collections of theNational Museum of Tanzania, Dar es Sa-laam (OH 8) and National Museum of Ethi-opia, Addis Ababa (Hadar). Oreopithecusfossil material was studied at the Museum ofNatural History, Basel; Istituto di GeologiaUniversita de Firenze; and Instituto de Pa-leontologia, Sabadell, Spain.
Lengths were measured to the nearest 0.1mm using a digital caliper connected to acomputer and collected in a Lotus spread-sheet. A carpenter’s angle accurate to 0.58was applied to the articular surfaces to mea-sure the cuboectocuneiform angle and hori-zontal mesoectocuneiform angle. The re-maining angles were arrived at by aligning
wires to the articular surfaces and using thecarpenter angle to measure the angles madeby the wires. All measurements are of theright navicular except for some (fossils) inwhich only the left specimen was available.
Comparisons to body size relied on navic-ulars of specimens with reported bodyweights. For those specimens without re-ported body weight, the sums of the midshaftcross-sectional area of the femur and tibiawere used for comparisons. The length of thefemur (mm) times its midshaft cross-section-al area plus the length of the tibia (mm) timesits midshaft cross-sectional area were used tocompare lower-limb volume to correspond-ing navicular measurements for each individ-ual specimen. Measurements for lower-limblong bone lengths are after Sarmiento (1985)and Sarmiento et al. (1996). Cross-sectionalareas of lower-limb long bones were arrivedat by squaring the midshaft circumferenceand dividing it by 4p. Measurements and theindices formulated from them were chosen toreflect mechanical concerns. Figure 1 sum-marizes the length and angular measurementstaken on the navicular and the indices for-mulated from these measurements. Tables 1–10 summarize comparisons of the linear andangular measurements. Figures 2–11 are bi-variate plots of the proportions considered.
Functional interpretation of the navicularof fossil forms relied on nearly 20 years ofstudying foot use in humans and in free rang-ing and captive great apes (Sarmiento, 1983,1985, 1994; Sarmiento and Butynski, inprep.). Articulation of human, great ape, andfossil foot bones in close-packed positionswere used to gauge the orientation of thepedal segments imparted by the set (i.e., rel-ative orientation) of the navicular facets. Lig-amentous preparations of the foot of humans,western gorillas, common chimpanzees, andorangutans were used to further verify therelative orientations of articulated tarsals andmetatarsals. Notes on OH 8, Hadar, and Or-eopithecus foot bones were used to corrob-orate functional interpretations of fossil footuse based on the navicular study.
To complement results from the functionalanalysis and provide insights into phylogeny,a multiple discriminant and canonical variateanalysis was run employing 13 linear mea-
4 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_4File # 01cc
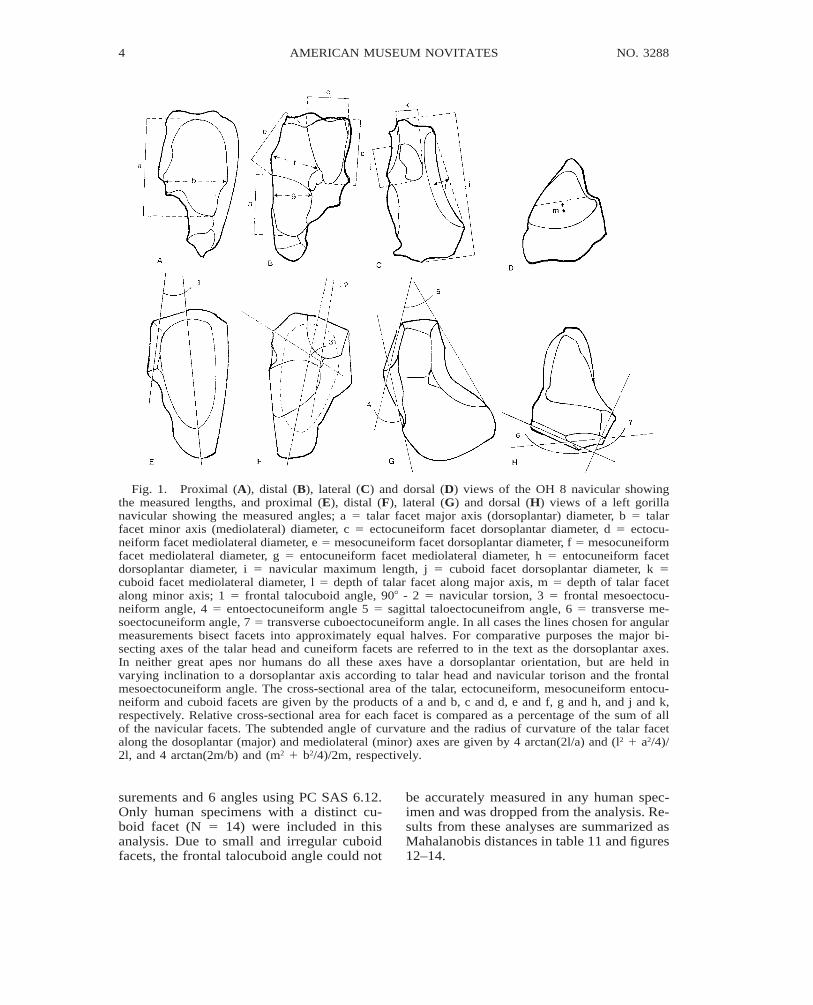
Fig. 1. Proximal (A), distal (B), lateral (C) and dorsal (D) views of the OH 8 navicular showingthe measured lengths, and proximal (E), distal (F), lateral (G) and dorsal (H) views of a left gorillanavicular showing the measured angles; a 5 talar facet major axis (dorsoplantar) diameter, b 5 talarfacet minor axis (mediolateral) diameter, c 5 ectocuneiform facet dorsoplantar diameter, d 5 ectocu-neiform facet mediolateral diameter, e 5 mesocuneiform facet dorsoplantar diameter, f 5 mesocuneiformfacet mediolateral diameter, g 5 entocuneiform facet mediolateral diameter, h 5 entocuneiform facetdorsoplantar diameter, i 5 navicular maximum length, j 5 cuboid facet dorsoplantar diameter, k 5cuboid facet mediolateral diameter, l 5 depth of talar facet along major axis, m 5 depth of talar facetalong minor axis; 1 5 frontal talocuboid angle, 908 - 2 5 navicular torsion, 3 5 frontal mesoectocu-neiform angle, 4 5 entoectocuneiform angle 5 5 sagittal taloectocuneifrom angle, 6 5 transverse me-soectocuneiform angle, 7 5 transverse cuboectocuneiform angle. In all cases the lines chosen for angularmeasurements bisect facets into approximately equal halves. For comparative purposes the major bi-secting axes of the talar head and cuneiform facets are referred to in the text as the dorsoplantar axes.In neither great apes nor humans do all these axes have a dorsoplantar orientation, but are held invarying inclination to a dorsoplantar axis according to talar head and navicular torison and the frontalmesoectocuneiform angle. The cross-sectional area of the talar, ectocuneiform, mesocuneiform entocu-neiform and cuboid facets are given by the products of a and b, c and d, e and f, g and h, and j and k,respectively. Relative cross-sectional area for each facet is compared as a percentage of the sum of allof the navicular facets. The subtended angle of curvature and the radius of curvature of the talar facetalong the dosoplantar (major) and mediolateral (minor) axes are given by 4 arctan(2l/a) and (l2 1 a2/4)/2l, and 4 arctan(2m/b) and (m2 1 b2/4)/2m, respectively.
surements and 6 angles using PC SAS 6.12.Only human specimens with a distinct cu-boid facet (N 5 14) were included in thisanalysis. Due to small and irregular cuboidfacets, the frontal talocuboid angle could not
be accurately measured in any human spec-imen and was dropped from the analysis. Re-sults from these analyses are summarized asMahalanobis distances in table 11 and figures12–14.

2000 5SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_5File # 01cc
6 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_6File # 01cc
RESULTS
CANONICAL ANALYSES
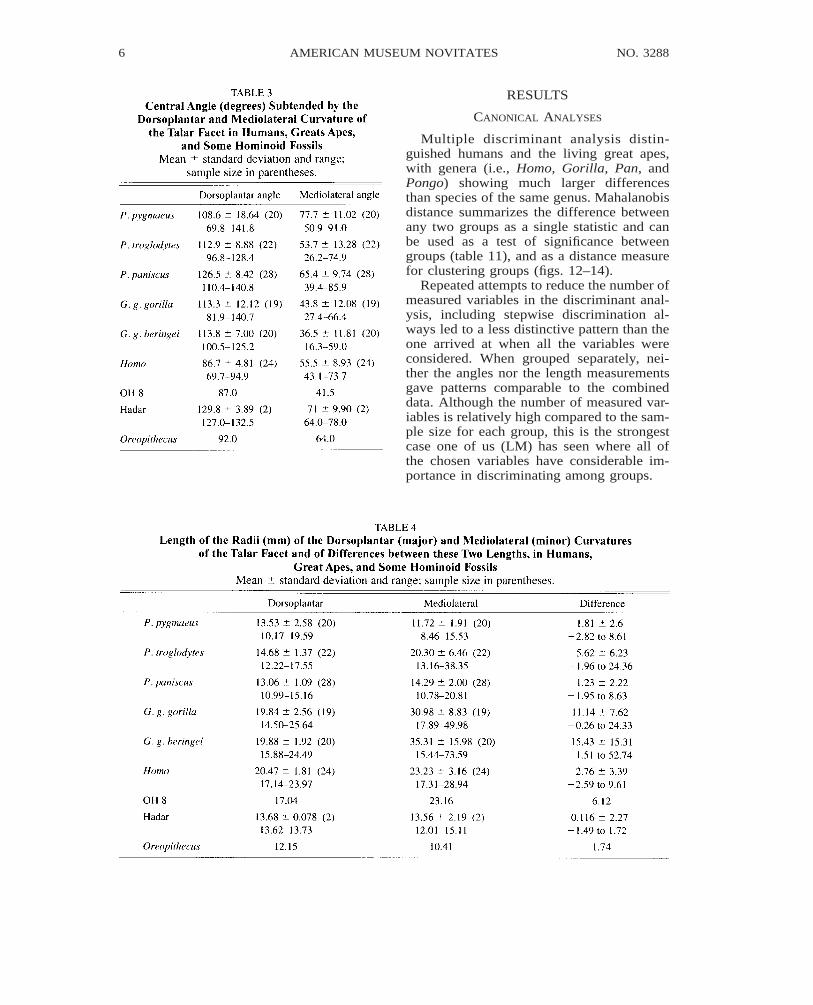
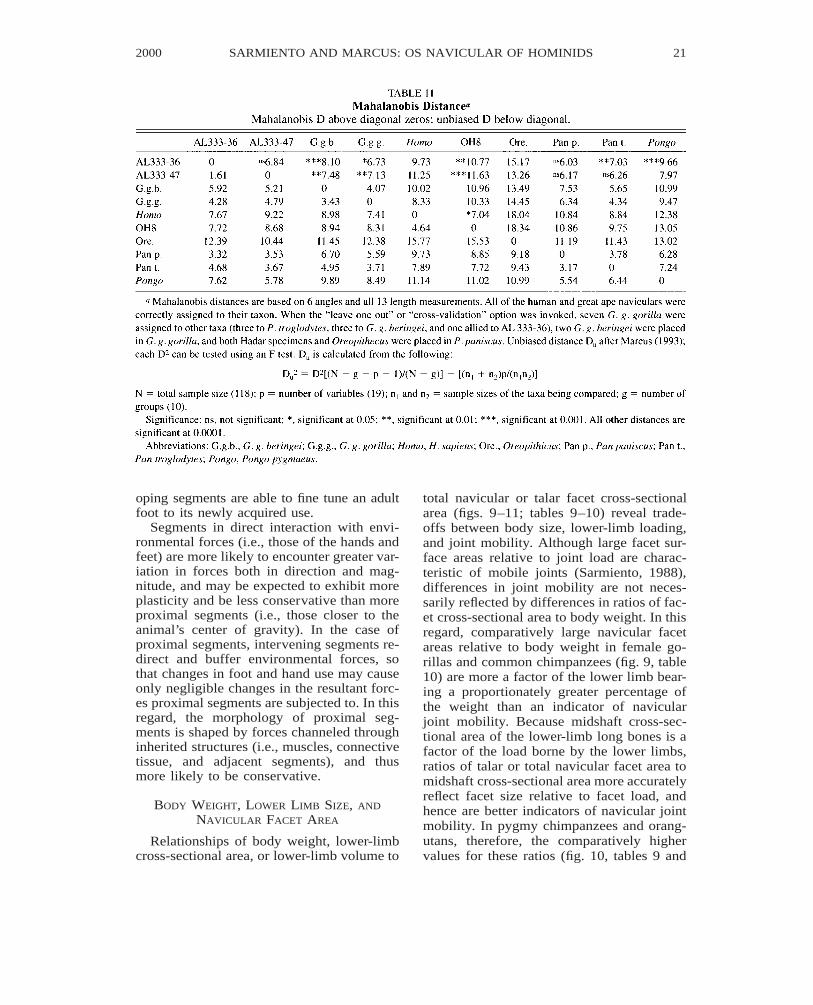
Multiple discriminant analysis distin-guished humans and the living great apes,with genera (i.e., Homo, Gorilla, Pan, andPongo) showing much larger differencesthan species of the same genus. Mahalanobisdistance summarizes the difference betweenany two groups as a single statistic and canbe used as a test of significance betweengroups (table 11), and as a distance measurefor clustering groups (figs. 12–14).
Repeated attempts to reduce the number ofmeasured variables in the discriminant anal-ysis, including stepwise discrimination al-ways led to a less distinctive pattern than theone arrived at when all the variables wereconsidered. When grouped separately, nei-ther the angles nor the length measurementsgave patterns comparable to the combineddata. Although the number of measured var-iables is relatively high compared to the sam-ple size for each group, this is the strongestcase one of us (LM) has seen where all ofthe chosen variables have considerable im-portance in discriminating among groups.
2000 7SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_7File # 01cc
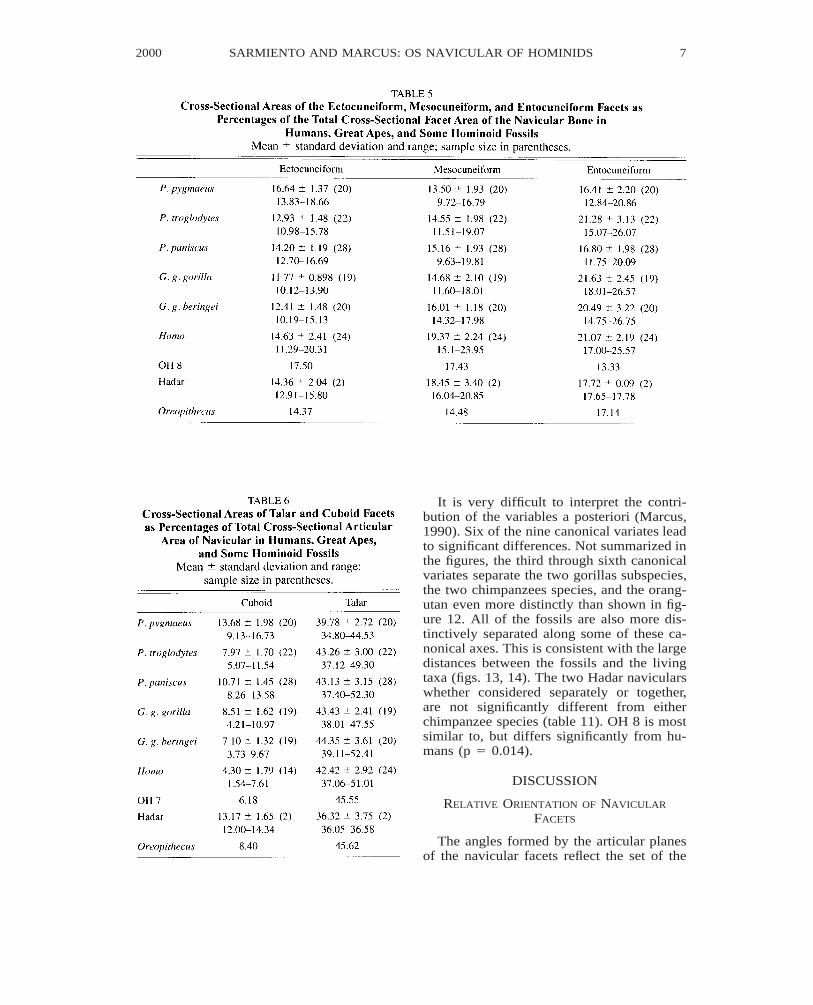
It is very difficult to interpret the contri-bution of the variables a posteriori (Marcus,1990). Six of the nine canonical variates leadto significant differences. Not summarized inthe figures, the third through sixth canonicalvariates separate the two gorillas subspecies,the two chimpanzees species, and the orang-utan even more distinctly than shown in fig-ure 12. All of the fossils are also more dis-tinctively separated along some of these ca-nonical axes. This is consistent with the largedistances between the fossils and the livingtaxa (figs. 13, 14). The two Hadar navicularswhether considered separately or together,are not significantly different from eitherchimpanzee species (table 11). OH 8 is mostsimilar to, but differs significantly from hu-mans (p 5 0.014).
DISCUSSION
RELATIVE ORIENTATION OF NAVICULAR
FACETS
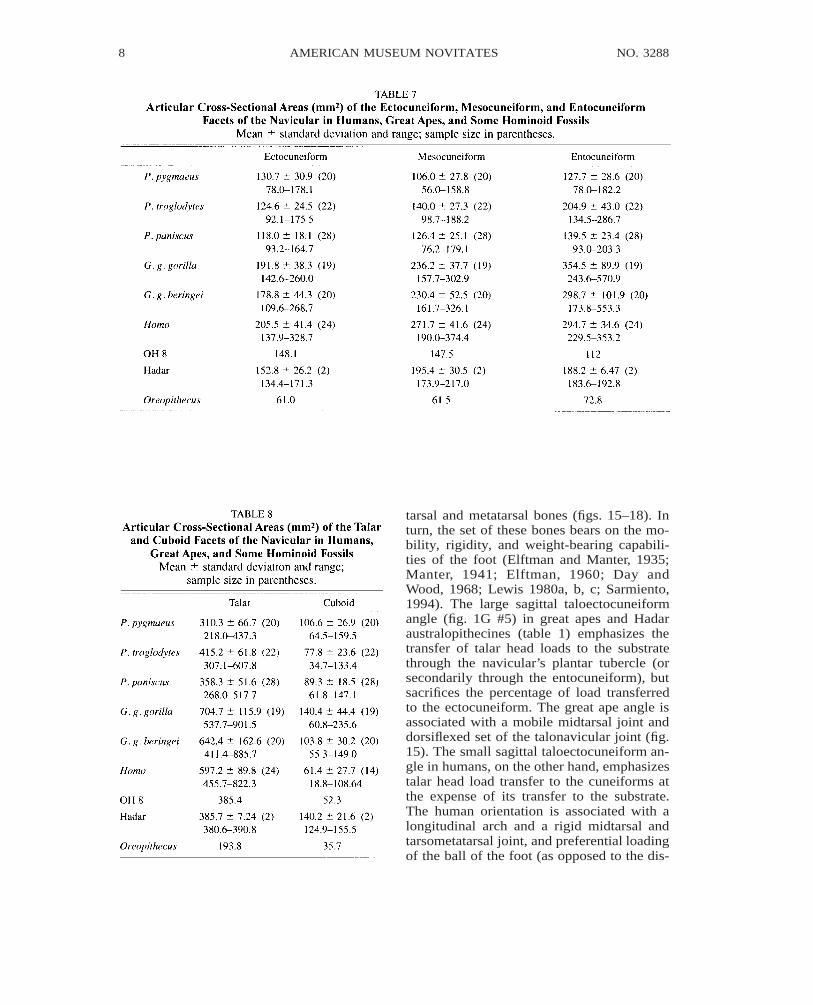
The angles formed by the articular planesof the navicular facets reflect the set of the
8 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_8File # 01cc
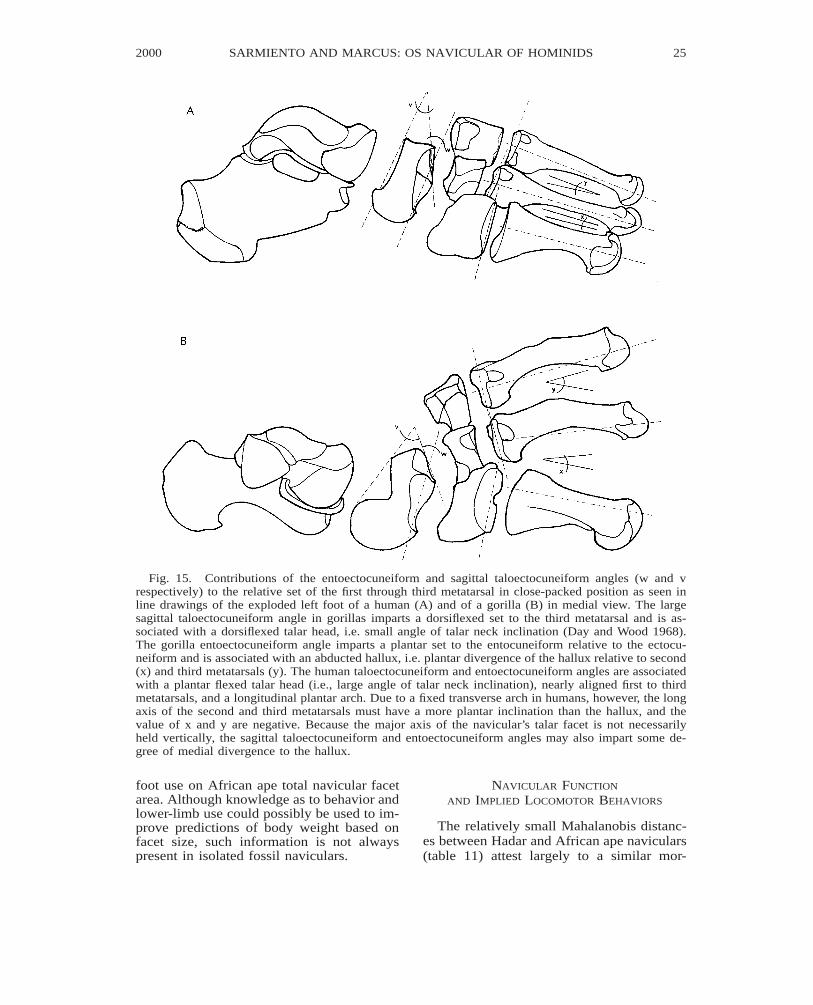
tarsal and metatarsal bones (figs. 15–18). Inturn, the set of these bones bears on the mo-bility, rigidity, and weight-bearing capabili-ties of the foot (Elftman and Manter, 1935;Manter, 1941; Elftman, 1960; Day andWood, 1968; Lewis 1980a, b, c; Sarmiento,1994). The large sagittal taloectocuneiformangle (fig. 1G #5) in great apes and Hadaraustralopithecines (table 1) emphasizes thetransfer of talar head loads to the substratethrough the navicular’s plantar tubercle (orsecondarily through the entocuneiform), butsacrifices the percentage of load transferredto the ectocuneiform. The great ape angle isassociated with a mobile midtarsal joint anddorsiflexed set of the talonavicular joint (fig.15). The small sagittal taloectocuneiform an-gle in humans, on the other hand, emphasizestalar head load transfer to the cuneiforms atthe expense of its transfer to the substrate.The human orientation is associated with alongitudinal arch and a rigid midtarsal andtarsometatarsal joint, and preferential loadingof the ball of the foot (as opposed to the dis-
2000 9SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_9File # 01cc
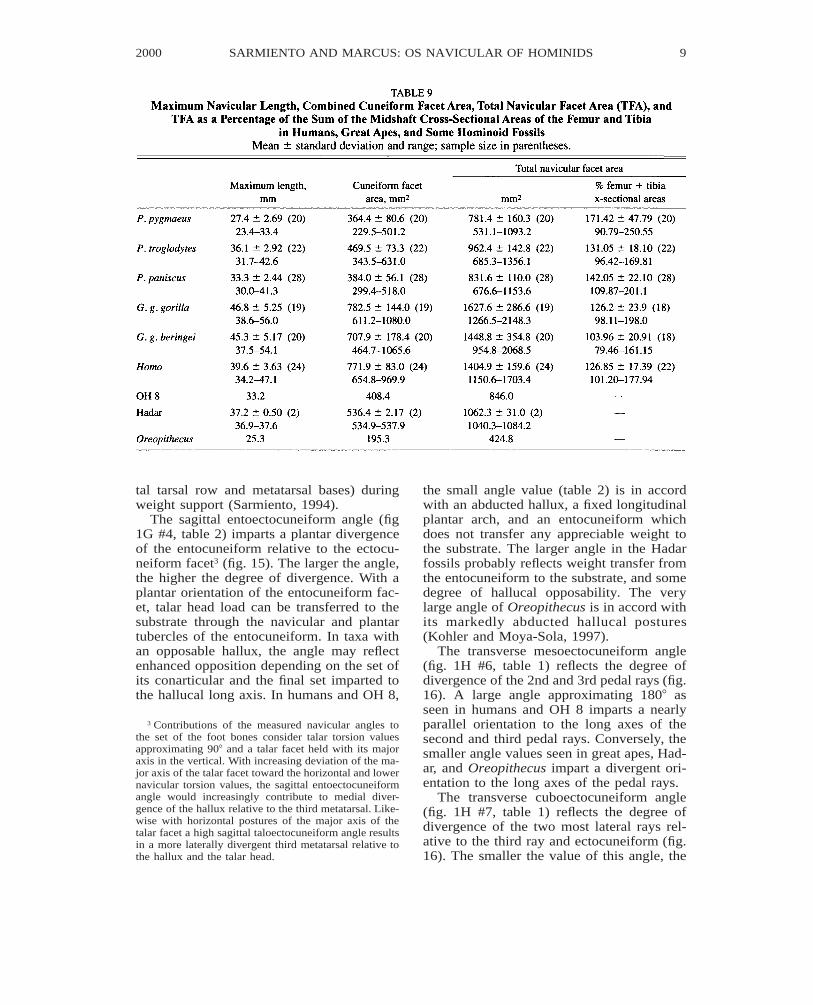
tal tarsal row and metatarsal bases) duringweight support (Sarmiento, 1994).
The sagittal entoectocuneiform angle (fig1G #4, table 2) imparts a plantar divergenceof the entocuneiform relative to the ectocu-neiform facet3 (fig. 15). The larger the angle,the higher the degree of divergence. With aplantar orientation of the entocuneiform fac-et, talar head load can be transferred to thesubstrate through the navicular and plantartubercles of the entocuneiform. In taxa withan opposable hallux, the angle may reflectenhanced opposition depending on the set ofits conarticular and the final set imparted tothe hallucal long axis. In humans and OH 8,
3 Contributions of the measured navicular angles tothe set of the foot bones consider talar torsion valuesapproximating 908 and a talar facet held with its majoraxis in the vertical. With increasing deviation of the ma-jor axis of the talar facet toward the horizontal and lowernavicular torsion values, the sagittal entoectocuneiformangle would increasingly contribute to medial diver-gence of the hallux relative to the third metatarsal. Like-wise with horizontal postures of the major axis of thetalar facet a high sagittal taloectocuneiform angle resultsin a more laterally divergent third metatarsal relative tothe hallux and the talar head.
the small angle value (table 2) is in accordwith an abducted hallux, a fixed longitudinalplantar arch, and an entocuneiform whichdoes not transfer any appreciable weight tothe substrate. The larger angle in the Hadarfossils probably reflects weight transfer fromthe entocuneiform to the substrate, and somedegree of hallucal opposability. The verylarge angle of Oreopithecus is in accord withits markedly abducted hallucal postures(Kohler and Moya-Sola, 1997).
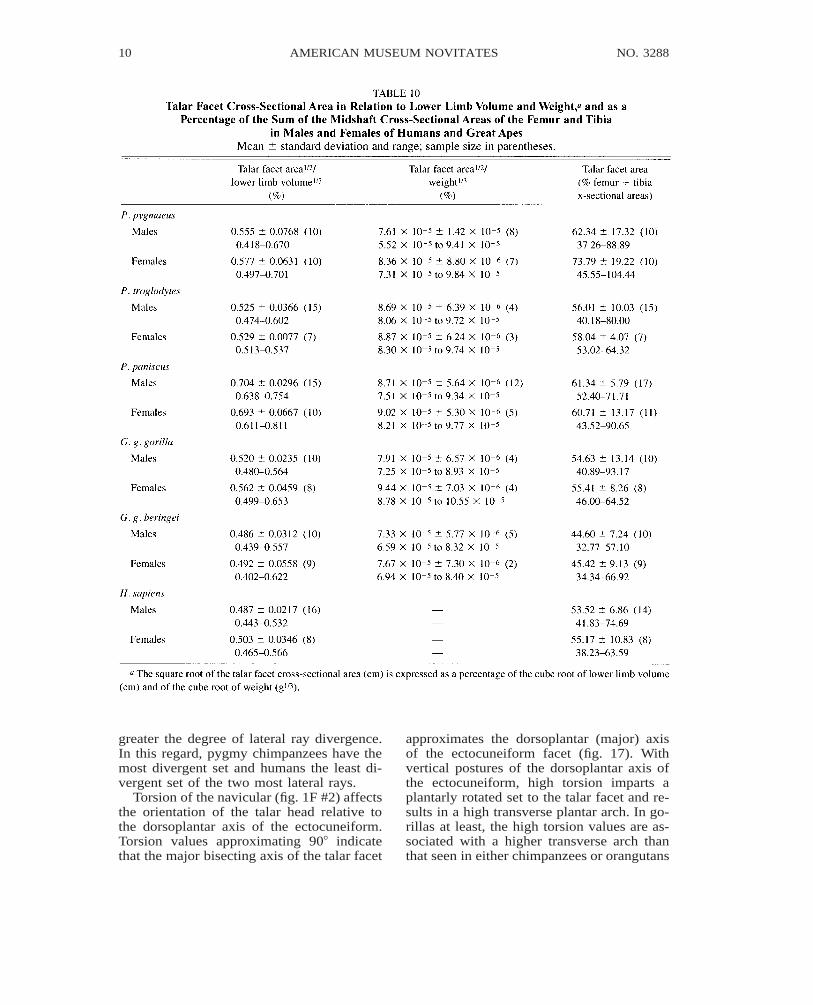
The transverse mesoectocuneiform angle(fig. 1H #6, table 1) reflects the degree ofdivergence of the 2nd and 3rd pedal rays (fig.16). A large angle approximating 1808 asseen in humans and OH 8 imparts a nearlyparallel orientation to the long axes of thesecond and third pedal rays. Conversely, thesmaller angle values seen in great apes, Had-ar, and Oreopithecus impart a divergent ori-entation to the long axes of the pedal rays.
The transverse cuboectocuneiform angle(fig. 1H #7, table 1) reflects the degree ofdivergence of the two most lateral rays rel-ative to the third ray and ectocuneiform (fig.16). The smaller the value of this angle, the
10 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_10File # 01cc
greater the degree of lateral ray divergence.In this regard, pygmy chimpanzees have themost divergent set and humans the least di-vergent set of the two most lateral rays.
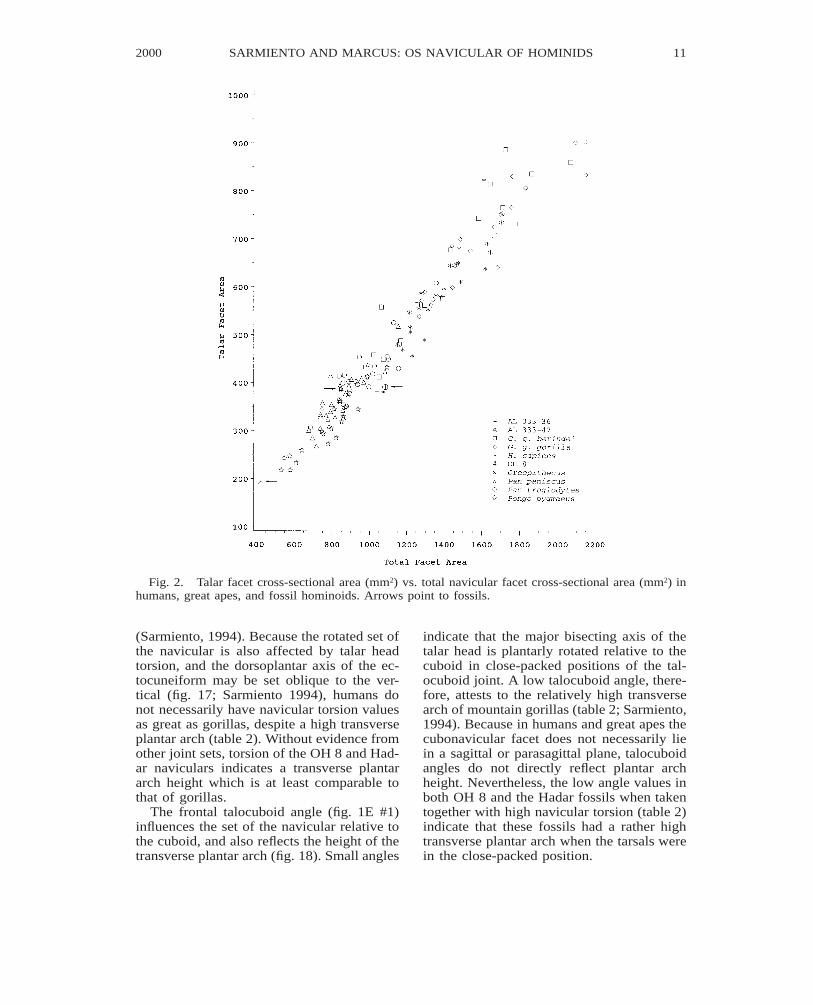
Torsion of the navicular (fig. 1F #2) affectsthe orientation of the talar head relative tothe dorsoplantar axis of the ectocuneiform.Torsion values approximating 908 indicatethat the major bisecting axis of the talar facet
approximates the dorsoplantar (major) axisof the ectocuneiform facet (fig. 17). Withvertical postures of the dorsoplantar axis ofthe ectocuneiform, high torsion imparts aplantarly rotated set to the talar facet and re-sults in a high transverse plantar arch. In go-rillas at least, the high torsion values are as-sociated with a higher transverse arch thanthat seen in either chimpanzees or orangutans
2000 11SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_11File # 01cc
Fig. 2. Talar facet cross-sectional area (mm2) vs. total navicular facet cross-sectional area (mm2) inhumans, great apes, and fossil hominoids. Arrows point to fossils.
(Sarmiento, 1994). Because the rotated set ofthe navicular is also affected by talar headtorsion, and the dorsoplantar axis of the ec-tocuneiform may be set oblique to the ver-tical (fig. 17; Sarmiento 1994), humans donot necessarily have navicular torsion valuesas great as gorillas, despite a high transverseplantar arch (table 2). Without evidence fromother joint sets, torsion of the OH 8 and Had-ar naviculars indicates a transverse plantararch height which is at least comparable tothat of gorillas.
The frontal talocuboid angle (fig. 1E #1)influences the set of the navicular relative tothe cuboid, and also reflects the height of thetransverse plantar arch (fig. 18). Small angles
indicate that the major bisecting axis of thetalar head is plantarly rotated relative to thecuboid in close-packed positions of the tal-ocuboid joint. A low talocuboid angle, there-fore, attests to the relatively high transversearch of mountain gorillas (table 2; Sarmiento,1994). Because in humans and great apes thecubonavicular facet does not necessarily liein a sagittal or parasagittal plane, talocuboidangles do not directly reflect plantar archheight. Nevertheless, the low angle values inboth OH 8 and the Hadar fossils when takentogether with high navicular torsion (table 2)indicate that these fossils had a rather hightransverse plantar arch when the tarsals werein the close-packed position.
12 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_12File # 01cc
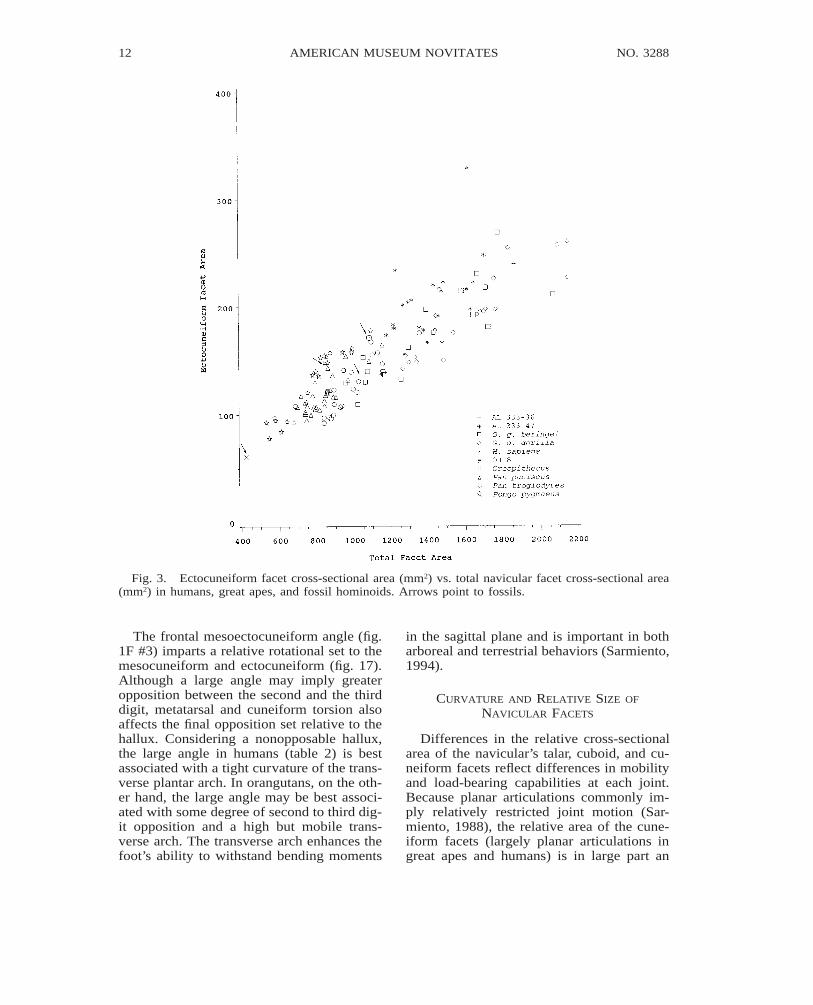
Fig. 3. Ectocuneiform facet cross-sectional area (mm2) vs. total navicular facet cross-sectional area(mm2) in humans, great apes, and fossil hominoids. Arrows point to fossils.
The frontal mesoectocuneiform angle (fig.1F #3) imparts a relative rotational set to themesocuneiform and ectocuneiform (fig. 17).Although a large angle may imply greateropposition between the second and the thirddigit, metatarsal and cuneiform torsion alsoaffects the final opposition set relative to thehallux. Considering a nonopposable hallux,the large angle in humans (table 2) is bestassociated with a tight curvature of the trans-verse plantar arch. In orangutans, on the oth-er hand, the large angle may be best associ-ated with some degree of second to third dig-it opposition and a high but mobile trans-verse arch. The transverse arch enhances thefoot’s ability to withstand bending moments
in the sagittal plane and is important in botharboreal and terrestrial behaviors (Sarmiento,1994).
CURVATURE AND RELATIVE SIZE OF
NAVICULAR FACETS
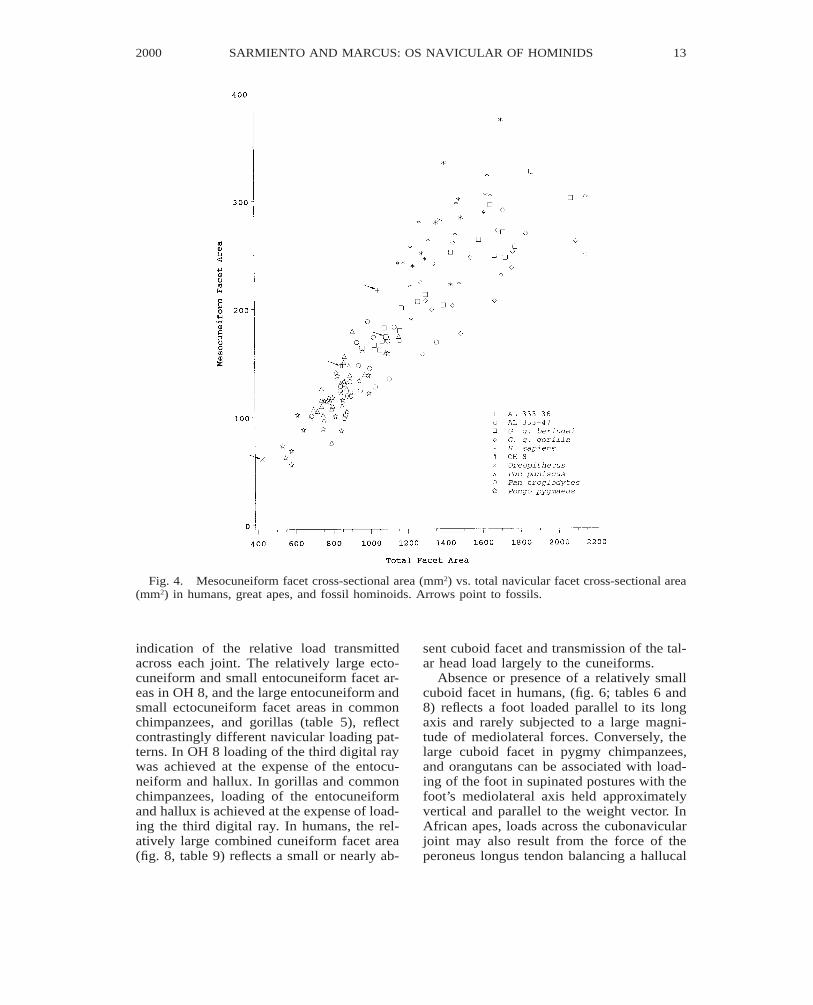
Differences in the relative cross-sectionalarea of the navicular’s talar, cuboid, and cu-neiform facets reflect differences in mobilityand load-bearing capabilities at each joint.Because planar articulations commonly im-ply relatively restricted joint motion (Sar-miento, 1988), the relative area of the cune-iform facets (largely planar articulations ingreat apes and humans) is in large part an
2000 13SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_13File # 01cc
Fig. 4. Mesocuneiform facet cross-sectional area (mm2) vs. total navicular facet cross-sectional area(mm2) in humans, great apes, and fossil hominoids. Arrows point to fossils.
indication of the relative load transmittedacross each joint. The relatively large ecto-cuneiform and small entocuneiform facet ar-eas in OH 8, and the large entocuneiform andsmall ectocuneiform facet areas in commonchimpanzees, and gorillas (table 5), reflectcontrastingly different navicular loading pat-terns. In OH 8 loading of the third digital raywas achieved at the expense of the entocu-neiform and hallux. In gorillas and commonchimpanzees, loading of the entocuneiformand hallux is achieved at the expense of load-ing the third digital ray. In humans, the rel-atively large combined cuneiform facet area(fig. 8, table 9) reflects a small or nearly ab-
sent cuboid facet and transmission of the tal-ar head load largely to the cuneiforms.
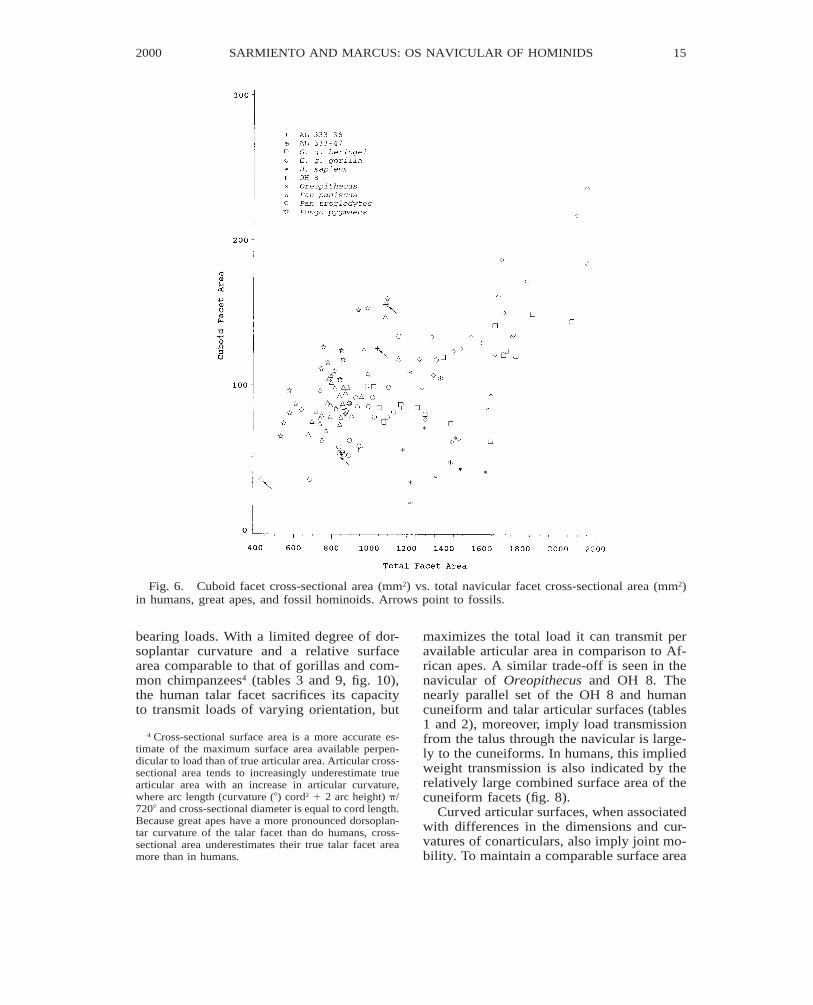
Absence or presence of a relatively smallcuboid facet in humans, (fig. 6; tables 6 and8) reflects a foot loaded parallel to its longaxis and rarely subjected to a large magni-tude of mediolateral forces. Conversely, thelarge cuboid facet in pygmy chimpanzees,and orangutans can be associated with load-ing of the foot in supinated postures with thefoot’s mediolateral axis held approximatelyvertical and parallel to the weight vector. InAfrican apes, loads across the cubonavicularjoint may also result from the force of theperoneus longus tendon balancing a hallucal
14 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_14File # 01cc
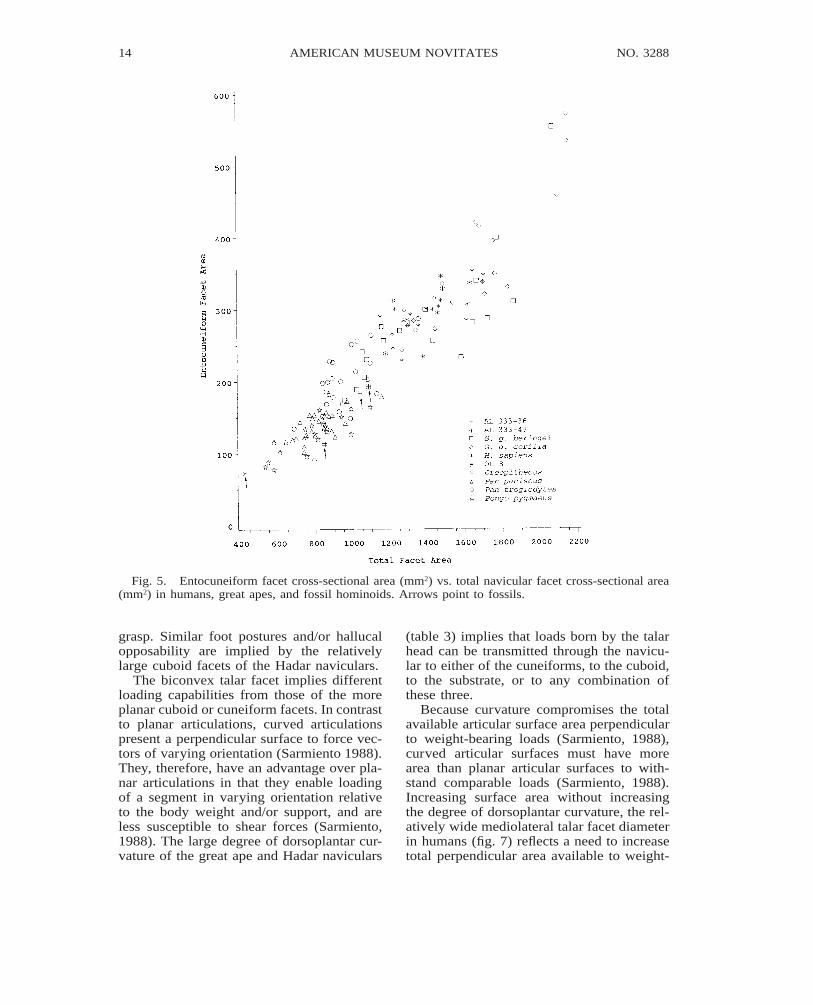
Fig. 5. Entocuneiform facet cross-sectional area (mm2) vs. total navicular facet cross-sectional area(mm2) in humans, great apes, and fossil hominoids. Arrows point to fossils.
grasp. Similar foot postures and/or hallucalopposability are implied by the relativelylarge cuboid facets of the Hadar naviculars.
The biconvex talar facet implies differentloading capabilities from those of the moreplanar cuboid or cuneiform facets. In contrastto planar articulations, curved articulationspresent a perpendicular surface to force vec-tors of varying orientation (Sarmiento 1988).They, therefore, have an advantage over pla-nar articulations in that they enable loadingof a segment in varying orientation relativeto the body weight and/or support, and areless susceptible to shear forces (Sarmiento,1988). The large degree of dorsoplantar cur-vature of the great ape and Hadar naviculars
(table 3) implies that loads born by the talarhead can be transmitted through the navicu-lar to either of the cuneiforms, to the cuboid,to the substrate, or to any combination ofthese three.
Because curvature compromises the totalavailable articular surface area perpendicularto weight-bearing loads (Sarmiento, 1988),curved articular surfaces must have morearea than planar articular surfaces to with-stand comparable loads (Sarmiento, 1988).Increasing surface area without increasingthe degree of dorsoplantar curvature, the rel-atively wide mediolateral talar facet diameterin humans (fig. 7) reflects a need to increasetotal perpendicular area available to weight-
2000 15SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_15File # 01cc
Fig. 6. Cuboid facet cross-sectional area (mm2) vs. total navicular facet cross-sectional area (mm2)in humans, great apes, and fossil hominoids. Arrows point to fossils.
bearing loads. With a limited degree of dor-soplantar curvature and a relative surfacearea comparable to that of gorillas and com-mon chimpanzees4 (tables 3 and 9, fig. 10),the human talar facet sacrifices its capacityto transmit loads of varying orientation, but
4 Cross-sectional surface area is a more accurate es-timate of the maximum surface area available perpen-dicular to load than of true articular area. Articular cross-sectional area tends to increasingly underestimate truearticular area with an increase in articular curvature,where arc length (curvature (8) cord2 1 2 arc height) p/7208 and cross-sectional diameter is equal to cord length.Because great apes have a more pronounced dorsoplan-tar curvature of the talar facet than do humans, cross-sectional area underestimates their true talar facet areamore than in humans.
maximizes the total load it can transmit peravailable articular area in comparison to Af-rican apes. A similar trade-off is seen in thenavicular of Oreopithecus and OH 8. Thenearly parallel set of the OH 8 and humancuneiform and talar articular surfaces (tables1 and 2), moreover, imply load transmissionfrom the talus through the navicular is large-ly to the cuneiforms. In humans, this impliedweight transmission is also indicated by therelatively large combined surface area of thecuneiform facets (fig. 8).
Curved articular surfaces, when associatedwith differences in the dimensions and cur-vatures of conarticulars, also imply joint mo-bility. To maintain a comparable surface area
16 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_16File # 01cc
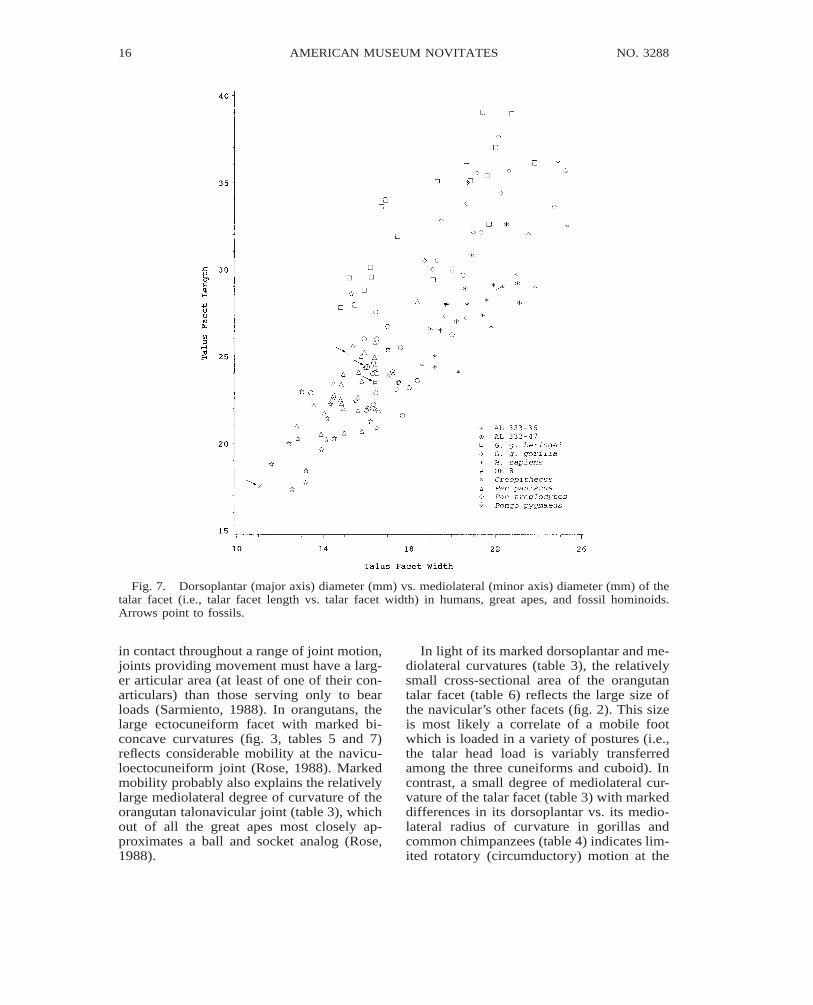
Fig. 7. Dorsoplantar (major axis) diameter (mm) vs. mediolateral (minor axis) diameter (mm) of thetalar facet (i.e., talar facet length vs. talar facet width) in humans, great apes, and fossil hominoids.Arrows point to fossils.
in contact throughout a range of joint motion,joints providing movement must have a larg-er articular area (at least of one of their con-articulars) than those serving only to bearloads (Sarmiento, 1988). In orangutans, thelarge ectocuneiform facet with marked bi-concave curvatures (fig. 3, tables 5 and 7)reflects considerable mobility at the navicu-loectocuneiform joint (Rose, 1988). Markedmobility probably also explains the relativelylarge mediolateral degree of curvature of theorangutan talonavicular joint (table 3), whichout of all the great apes most closely ap-proximates a ball and socket analog (Rose,1988).
In light of its marked dorsoplantar and me-diolateral curvatures (table 3), the relativelysmall cross-sectional area of the orangutantalar facet (table 6) reflects the large size ofthe navicular’s other facets (fig. 2). This sizeis most likely a correlate of a mobile footwhich is loaded in a variety of postures (i.e.,the talar head load is variably transferredamong the three cuneiforms and cuboid). Incontrast, a small degree of mediolateral cur-vature of the talar facet (table 3) with markeddifferences in its dorsoplantar vs. its medio-lateral radius of curvature in gorillas andcommon chimpanzees (table 4) indicates lim-ited rotatory (circumductory) motion at the
2000 17SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_17File # 01cc
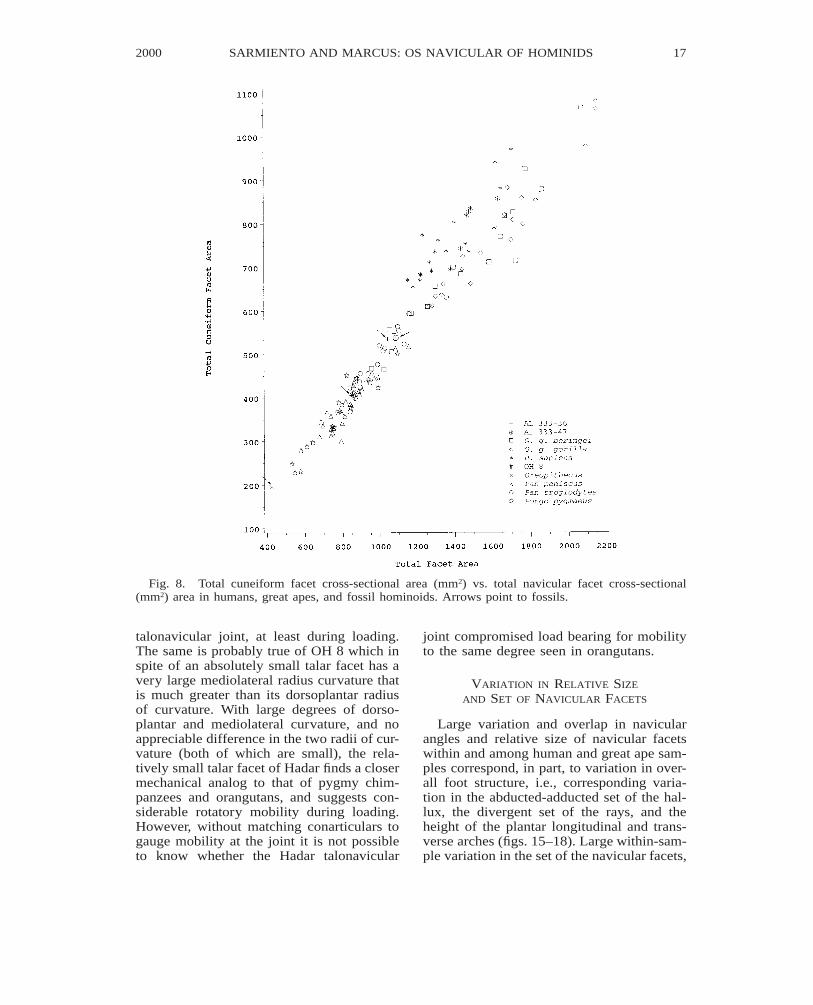
Fig. 8. Total cuneiform facet cross-sectional area (mm2) vs. total navicular facet cross-sectional(mm2) area in humans, great apes, and fossil hominoids. Arrows point to fossils.
talonavicular joint, at least during loading.The same is probably true of OH 8 which inspite of an absolutely small talar facet has avery large mediolateral radius curvature thatis much greater than its dorsoplantar radiusof curvature. With large degrees of dorso-plantar and mediolateral curvature, and noappreciable difference in the two radii of cur-vature (both of which are small), the rela-tively small talar facet of Hadar finds a closermechanical analog to that of pygmy chim-panzees and orangutans, and suggests con-siderable rotatory mobility during loading.However, without matching conarticulars togauge mobility at the joint it is not possibleto know whether the Hadar talonavicular
joint compromised load bearing for mobilityto the same degree seen in orangutans.
VARIATION IN RELATIVE SIZE
AND SET OF NAVICULAR FACETS
Large variation and overlap in navicularangles and relative size of navicular facetswithin and among human and great ape sam-ples correspond, in part, to variation in over-all foot structure, i.e., corresponding varia-tion in the abducted-adducted set of the hal-lux, the divergent set of the rays, and theheight of the plantar longitudinal and trans-verse arches (figs. 15–18). Large within-sam-ple variation in the set of the navicular facets,
18 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_18File # 01cc
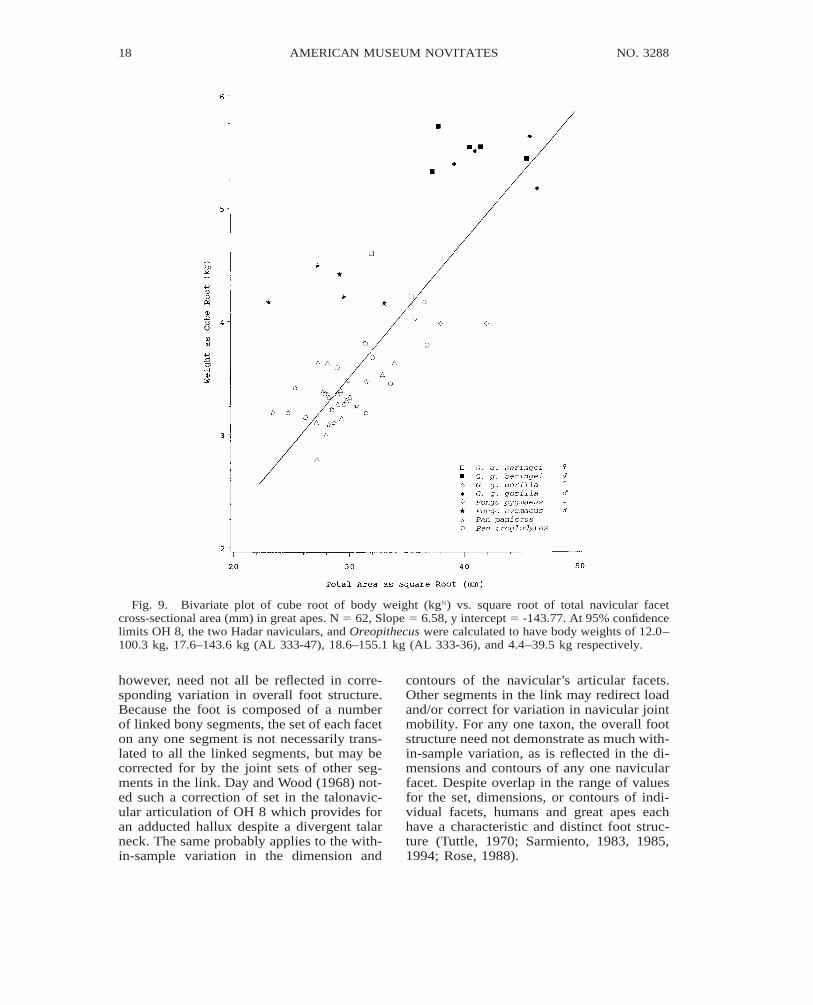
Fig. 9. Bivariate plot of cube root of body weight (kg⅓) vs. square root of total navicular facetcross-sectional area (mm) in great apes. N 5 62, Slope 5 6.58, y intercept 5 -143.77. At 95% confidencelimits OH 8, the two Hadar naviculars, and Oreopithecus were calculated to have body weights of 12.0–100.3 kg, 17.6–143.6 kg (AL 333-47), 18.6–155.1 kg (AL 333-36), and 4.4–39.5 kg respectively.
however, need not all be reflected in corre-sponding variation in overall foot structure.Because the foot is composed of a numberof linked bony segments, the set of each faceton any one segment is not necessarily trans-lated to all the linked segments, but may becorrected for by the joint sets of other seg-ments in the link. Day and Wood (1968) not-ed such a correction of set in the talonavic-ular articulation of OH 8 which provides foran adducted hallux despite a divergent talarneck. The same probably applies to the with-in-sample variation in the dimension and
contours of the navicular’s articular facets.Other segments in the link may redirect loadand/or correct for variation in navicular jointmobility. For any one taxon, the overall footstructure need not demonstrate as much with-in-sample variation, as is reflected in the di-mensions and contours of any one navicularfacet. Despite overlap in the range of valuesfor the set, dimensions, or contours of indi-vidual facets, humans and great apes eachhave a characteristic and distinct foot struc-ture (Tuttle, 1970; Sarmiento, 1983, 1985,1994; Rose, 1988).
2000 19SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_19File # 01cc
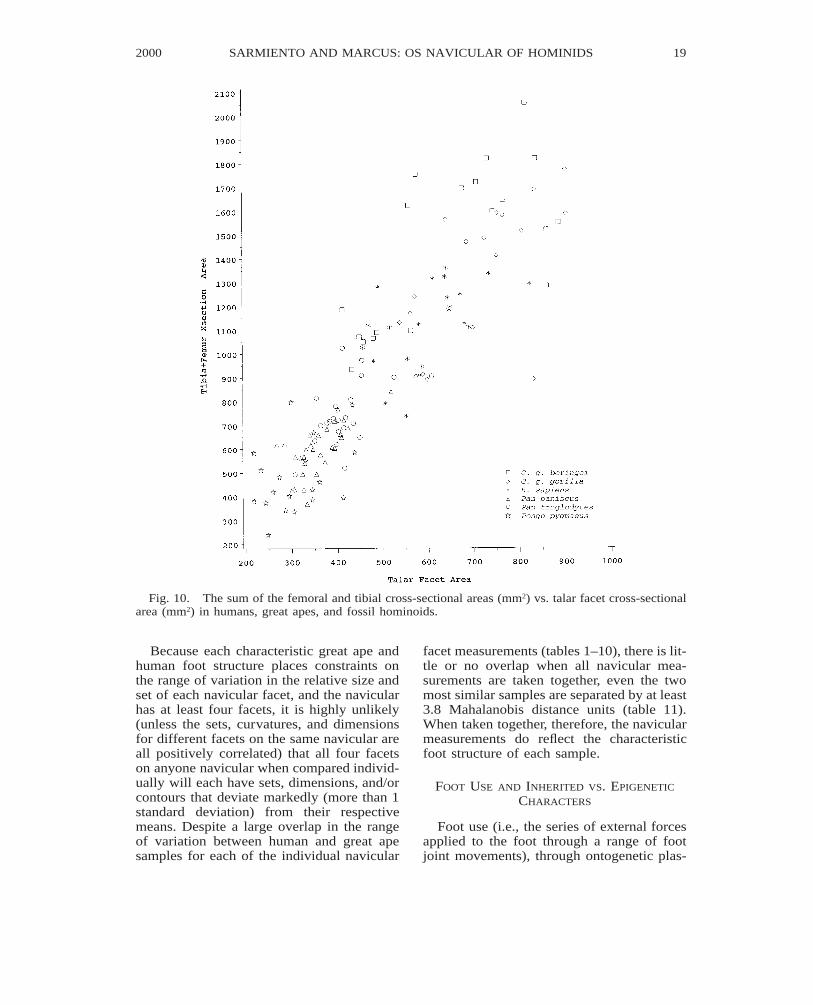
Fig. 10. The sum of the femoral and tibial cross-sectional areas (mm2) vs. talar facet cross-sectionalarea (mm2) in humans, great apes, and fossil hominoids.
Because each characteristic great ape andhuman foot structure places constraints onthe range of variation in the relative size andset of each navicular facet, and the navicularhas at least four facets, it is highly unlikely(unless the sets, curvatures, and dimensionsfor different facets on the same navicular areall positively correlated) that all four facetson anyone navicular when compared individ-ually will each have sets, dimensions, and/orcontours that deviate markedly (more than 1standard deviation) from their respectivemeans. Despite a large overlap in the rangeof variation between human and great apesamples for each of the individual navicular
facet measurements (tables 1–10), there is lit-tle or no overlap when all navicular mea-surements are taken together, even the twomost similar samples are separated by at least3.8 Mahalanobis distance units (table 11).When taken together, therefore, the navicularmeasurements do reflect the characteristicfoot structure of each sample.
FOOT USE AND INHERITED VS. EPIGENETIC
CHARACTERS
Foot use (i.e., the series of external forcesapplied to the foot through a range of footjoint movements), through ontogenetic plas-
20 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_20File # 01cc
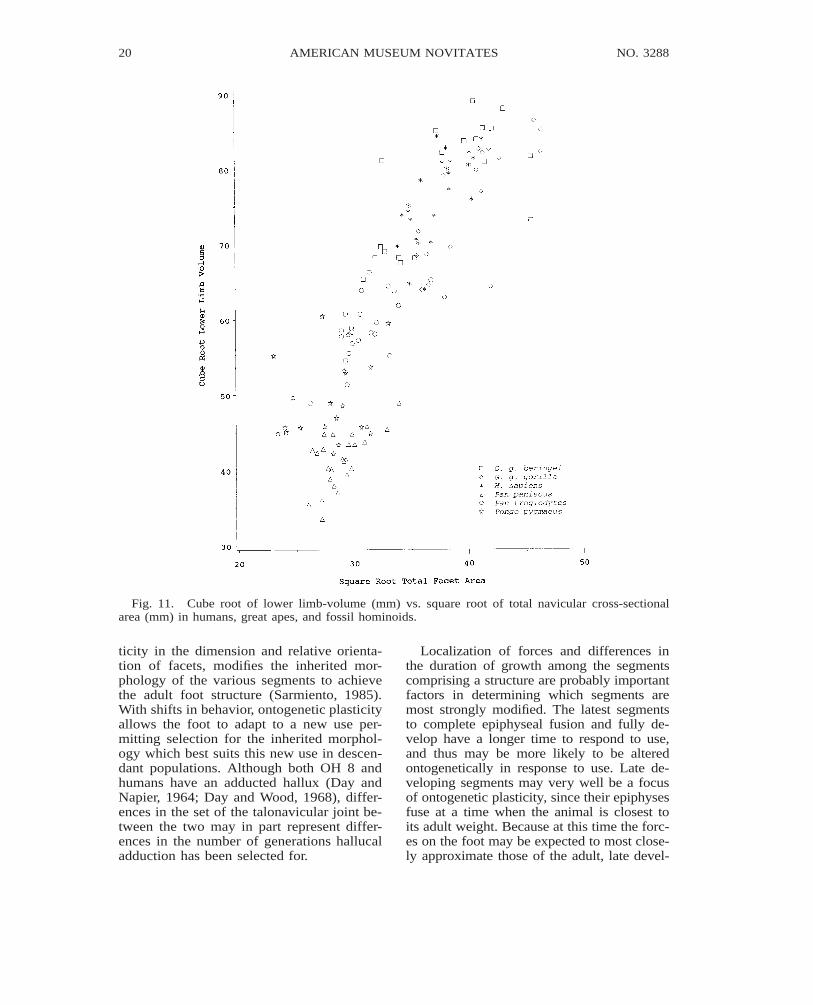
Fig. 11. Cube root of lower limb-volume (mm) vs. square root of total navicular cross-sectionalarea (mm) in humans, great apes, and fossil hominoids.
ticity in the dimension and relative orienta-tion of facets, modifies the inherited mor-phology of the various segments to achievethe adult foot structure (Sarmiento, 1985).With shifts in behavior, ontogenetic plasticityallows the foot to adapt to a new use per-mitting selection for the inherited morphol-ogy which best suits this new use in descen-dant populations. Although both OH 8 andhumans have an adducted hallux (Day andNapier, 1964; Day and Wood, 1968), differ-ences in the set of the talonavicular joint be-tween the two may in part represent differ-ences in the number of generations hallucaladduction has been selected for.
Localization of forces and differences inthe duration of growth among the segmentscomprising a structure are probably importantfactors in determining which segments aremost strongly modified. The latest segmentsto complete epiphyseal fusion and fully de-velop have a longer time to respond to use,and thus may be more likely to be alteredontogenetically in response to use. Late de-veloping segments may very well be a focusof ontogenetic plasticity, since their epiphysesfuse at a time when the animal is closest toits adult weight. Because at this time the forc-es on the foot may be expected to most close-ly approximate those of the adult, late devel-
2000 21SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_21File # 01cc
oping segments are able to fine tune an adultfoot to its newly acquired use.
Segments in direct interaction with envi-ronmental forces (i.e., those of the hands andfeet) are more likely to encounter greater var-iation in forces both in direction and mag-nitude, and may be expected to exhibit moreplasticity and be less conservative than moreproximal segments (i.e., those closer to theanimal’s center of gravity). In the case ofproximal segments, intervening segments re-direct and buffer environmental forces, sothat changes in foot and hand use may causeonly negligible changes in the resultant forc-es proximal segments are subjected to. In thisregard, the morphology of proximal seg-ments is shaped by forces channeled throughinherited structures (i.e., muscles, connectivetissue, and adjacent segments), and thusmore likely to be conservative.
BODY WEIGHT, LOWER LIMB SIZE, AND
NAVICULAR FACET AREA
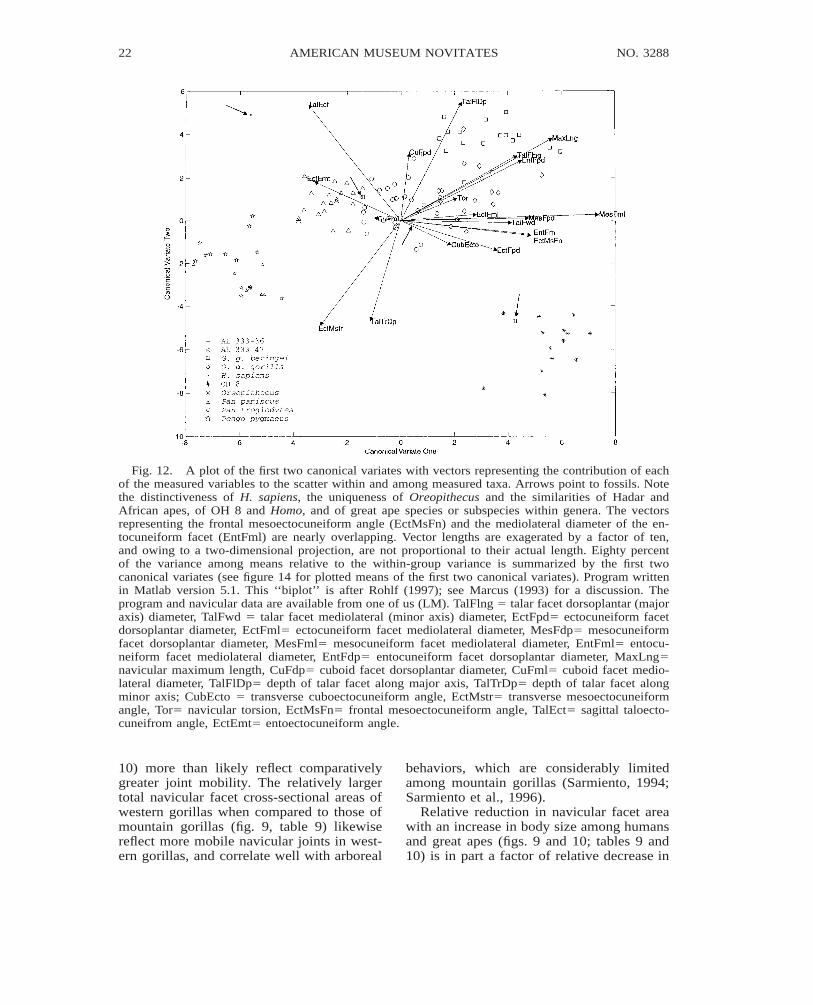
Relationships of body weight, lower-limbcross-sectional area, or lower-limb volume to
total navicular or talar facet cross-sectionalarea (figs. 9–11; tables 9–10) reveal trade-offs between body size, lower-limb loading,and joint mobility. Although large facet sur-face areas relative to joint load are charac-teristic of mobile joints (Sarmiento, 1988),differences in joint mobility are not neces-sarily reflected by differences in ratios of fac-et cross-sectional area to body weight. In thisregard, comparatively large navicular facetareas relative to body weight in female go-rillas and common chimpanzees (fig. 9, table10) are more a factor of the lower limb bear-ing a proportionately greater percentage ofthe weight than an indicator of navicularjoint mobility. Because midshaft cross-sec-tional area of the lower-limb long bones is afactor of the load borne by the lower limbs,ratios of talar or total navicular facet area tomidshaft cross-sectional area more accuratelyreflect facet size relative to facet load, andhence are better indicators of navicular jointmobility. In pygmy chimpanzees and orang-utans, therefore, the comparatively highervalues for these ratios (fig. 10, tables 9 and
22 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_22File # 01cc
Fig. 12. A plot of the first two canonical variates with vectors representing the contribution of eachof the measured variables to the scatter within and among measured taxa. Arrows point to fossils. Notethe distinctiveness of H. sapiens, the uniqueness of Oreopithecus and the similarities of Hadar andAfrican apes, of OH 8 and Homo, and of great ape species or subspecies within genera. The vectorsrepresenting the frontal mesoectocuneiform angle (EctMsFn) and the mediolateral diameter of the en-tocuneiform facet (EntFml) are nearly overlapping. Vector lengths are exagerated by a factor of ten,and owing to a two-dimensional projection, are not proportional to their actual length. Eighty percentof the variance among means relative to the within-group variance is summarized by the first twocanonical variates (see figure 14 for plotted means of the first two canonical variates). Program writtenin Matlab version 5.1. This ‘‘biplot’’ is after Rohlf (1997); see Marcus (1993) for a discussion. Theprogram and navicular data are available from one of us (LM). TalFlng 5 talar facet dorsoplantar (majoraxis) diameter, TalFwd 5 talar facet mediolateral (minor axis) diameter, EctFpd5 ectocuneiform facetdorsoplantar diameter, EctFml5 ectocuneiform facet mediolateral diameter, MesFdp5 mesocuneiformfacet dorsoplantar diameter, MesFml5 mesocuneiform facet mediolateral diameter, EntFml5 entocu-neiform facet mediolateral diameter, EntFdp5 entocuneiform facet dorsoplantar diameter, MaxLng5navicular maximum length, CuFdp5 cuboid facet dorsoplantar diameter, CuFml5 cuboid facet medio-lateral diameter, TalFlDp5 depth of talar facet along major axis, TalTrDp5 depth of talar facet alongminor axis; CubEcto 5 transverse cuboectocuneiform angle, EctMstr5 transverse mesoectocuneiformangle, Tor5 navicular torsion, EctMsFn5 frontal mesoectocuneiform angle, TalEct5 sagittal taloecto-cuneifrom angle, EctEmt5 entoectocuneiform angle.
10) more than likely reflect comparativelygreater joint mobility. The relatively largertotal navicular facet cross-sectional areas ofwestern gorillas when compared to those ofmountain gorillas (fig. 9, table 9) likewisereflect more mobile navicular joints in west-ern gorillas, and correlate well with arboreal
behaviors, which are considerably limitedamong mountain gorillas (Sarmiento, 1994;Sarmiento et al., 1996).
Relative reduction in navicular facet areawith an increase in body size among humansand great apes (figs. 9 and 10; tables 9 and10) is in part a factor of relative decrease in

2000 23SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_23File # 01cc
Fig. 13. Dendrograms of the studied taxa constructed using unweighted pair group method (Rohlf,1997). Inset shows portion of dendrogram which differs when both Hadar naviculars are considered asa single sample.
strength (proportional to muscle cross-sec-tional area) with increase in body size (a cor-ollary of volume), as predicted by the squarecube law. With proportionately weaker mus-cles, large-sized animals are unable to sta-bilize rotational forces of the same relativemagnitude as small-sized animals, and mustlimit their range of joint motion (around apoint of load equilibrium) to limit the mag-nitude of rotational forces. The relativelylarger facet areas of orangutan and gorilla fe-males when compared to their male counter-parts reflect such a trade-off between bodysize and joint mobility (fig. 9, table 9). Interms of behavior, these differences are re-vealed in a greater commitment to arborealbehaviors (Sarmiento, 1985, 1994; Sarmientoet al., 1996) and probably more variable foot
use among females. Additionally, orangutanand gorilla females may be expected to showquicker foot segment and body movements,given proportionately larger joint surface ar-eas to dissipate kinetic energy than their cor-responding adult males. The relatively smallnavicular joint surface areas of humans de-spite a considerably smaller body size thangorillas (tables 9 and 10) is the result of amarked commitment to bipedality and theadditional loading on the feet associated withbipedal behaviors.
Despite marked sexual dimorphism inbody weight, orangutans and gorillas of cor-responding sexes have similar-sized navicu-lar facet areas (fig. 9). This phenomenon isa correlate of sexual dimorphism and a pu-bescent growth spurt which doubles body

24 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_24File # 01cc
Fig. 14. Plot of mean canonical variate scores for studied taxa. All fossils are based on singlesamples. The actual Mahalanobis D for all of the canonical variates separating taxa is given as the valueabove each connecting line (table 11). Connecting lines represent a Minimum Spanning Tree (afterRohlf, 1997). Owing to a two-dimensional projection, the actual lengths of the connecting lines on theplot represent only a fraction of the D values.
size without a corresponding increase in footsize. With nearly adult foot size and navic-ular facet area to body weight ratios closerto their female counterparts, subadult maleorangutans and gorillas are also more arbo-real, and show more variable foot use thanwhen fully adult (Sarmiento, 1985, 1994;personal obs.).
Considering that (1) facet cross-sectionalarea is largely a factor of joint load and mo-bility, and (2) neither of these variables inhumans and great apes have a constant re-lationship with body weight, predictions ofbody weight in fossils from interspecies re-gressions of navicular facet area have a largedegree of error (fig. 9).
Likewise navicular facet size does notseem to have a constant relationship to low-er-limb volume (fig. 11, table 10). With var-ied foot use and navicular mobility and load-ing, including direct transfer of weight by thenavicular to the substrate, African apes havethe relatively largest navicular facet areas (ta-ble 10). A much larger size ratio in Africanapes than in humans is expected given ourhypertrophied lower limbs, limited navicularjoint mobility, and a longitudinal arch thatprevents navicular to substrate contact. How-ever, a comparable or larger ratio in Africanapes than in orangutans despite the orangu-tan’s small lower limbs underscores the ef-fect of terrestrial loading and generalized
2000 25SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_25File # 01cc
Fig. 15. Contributions of the entoectocuneiform and sagittal taloectocuneiform angles (w and vrespectively) to the relative set of the first through third metatarsal in close-packed position as seen inline drawings of the exploded left foot of a human (A) and of a gorilla (B) in medial view. The largesagittal taloectocuneiform angle in gorillas imparts a dorsiflexed set to the third metatarsal and is as-sociated with a dorsiflexed talar head, i.e. small angle of talar neck inclination (Day and Wood 1968).The gorilla entoectocuneiform angle imparts a plantar set to the entocuneiform relative to the ectocu-neiform and is associated with an abducted hallux, i.e. plantar divergence of the hallux relative to second(x) and third metatarsals (y). The human taloectocuneiform and entoectocuneiform angles are associatedwith a plantar flexed talar head (i.e., large angle of talar neck inclination), nearly aligned first to thirdmetatarsals, and a longitudinal plantar arch. Due to a fixed transverse arch in humans, however, the longaxis of the second and third metatarsals must have a more plantar inclination than the hallux, and thevalue of x and y are negative. Because the major axis of the navicular’s talar facet is not necessarilyheld vertically, the sagittal taloectocuneiform and entoectocuneiform angles may also impart some de-gree of medial divergence to the hallux.
foot use on African ape total navicular facetarea. Although knowledge as to behavior andlower-limb use could possibly be used to im-prove predictions of body weight based onfacet size, such information is not alwayspresent in isolated fossil naviculars.
NAVICULAR FUNCTION
AND IMPLIED LOCOMOTOR BEHAVIORS
The relatively small Mahalanobis distanc-es between Hadar and African ape naviculars(table 11) attest largely to a similar mor-

26 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_26File # 01cc
Fig. 16. Dorsal view of the exploded left tarsus and metatarsus of a pygmy chimpanzee in the close-packed position showing the contribution of the transverse mesoectocuneiform angle (w) and the trans-verse cuboectocuneiform angle (v) to the divergence of the second through fourth metatarsals. Correctionof the talocuboid angle by the facet sets on the cuboid and ectocuneiform results in third and fourthmetatarsals that are nearly alinged (y). A relatively low transverse mesoectocuneiform angle results ina second metatarsal that is divergent from the most lateral three (x) despite a partial correction of thisset by the mesocuneiform.
phology and imply similarities in function.The exclusive association of a localized mor-phology and its implied function to a specificlocomotor behavior, as is routine practiceamong paleoanthropologists (Napier, 1962;Day and Napier, 1964; Robinson, 1972; Con-roy and Fleagle, 1972; Stern and Susman,1983; Latimer et al., 1987; Latimer andLovejoy, 1989, 1990a, b; Lovejoy, 1978,1988; McHenry, 1991; Leakey et al., 1995;Ohman et al., 1997; Ward et al., 1999), may
be used by some to argue that the Hadar fos-sils represent knuckle-walkers. This interpre-tation is at odds with evidence from the pel-vis and knee joint, which although poorlyquantified and never adequately tested, is al-most unanimously taken as indicative of bi-pedality (Lovejoy, 1974; 1978; Stern andSusman, 1983; Susman et al., 1984; McHen-ry, 1991; Susman and Stern, 1991; Ohmanet al., 1977; Fleagle, 1998). The Hadar na-viculars, however, come from a different ho-

2000 27SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_27File # 01cc
Fig. 17. Contribution of navicular torsion (v) and the frontal mesoectocuneiform angle (w) to therelative set of the ectocuneiform, mesocuneiform, and the talar facet in the close-packed position asseen in line drawings of an exploded right foot of a human (A) and a gorilla (B) from a dorsodistalview. High values of the frontal mesoectocuneiform angle in humans (w) do not result in markedopposition of the second and third metatarsals given metatarsal, ectocuneiform, and mesocuneiformtorsion values which correct for the imparted set. Despite similar torsion values in humans and gorillas,the metatarsal, ectocuneiform, and mesocuneiform torsion all contribute to causing more marked op-position of the second and third metatarsals in gorillas. Marked talar torsion or large frontal mesoec-tocuneiform angles, are also associated to a high transverse arch (see text).
rizon (AL 333w/333) and an earlier time pe-riod than the Hadar pelvis (AL 288–1an,–ao)(Johanson et al., 1982), and the two remainsmay represent different taxa with differentlocomotor behaviors (Ferguson, 1984, 1986;Senut and Tardieu, 1985; Olson, 1981, 1985;Gommery, 1997).
At Hadar a contradiction in implied be-havior, however, also arises when comparingremains from the same horizon and may notnecessarily be the result of sampling differenttaxa. The alleged bipedal knee joint mor-phology is also seen in fossils from the samehorizon as the naviculars (AL 333w-56, 333-4, 333-111, 333x-26, 333-42), and the di-mensions, curvature, set, and configuration
of the navicular articulation on the AL 288-1 talus suggest an African apelike navicularsimilar to those from the AL 333/333w ho-rizon. The contradictions in behavior implicitin the navicular, pelvic, and knee joint mor-phology actually arise from the practice ofassociating localized anatomy exclusively toany one locomotor behavior and underscoresthe fallacy inherent in such a practice. Be-cause behavioral changes during evolution(i.e., adaptive shifts) require localized struc-tures to satisfy the mechanical requisites oftwo or more behaviors (Darwin, 1859), ex-clusive association of localized morphologyto any one locomotor behavior denies evo-lutionary change. Arguments for bipedality

28 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_28File # 01cc
Fig. 18. Proximal views of the right cuboid and navicular of a human (A) and a common chimpanzee(B) in a close-packed position showing the contribution of the talocuboid angle (v) to the height of thetransverse arch (h). In the close-packed position the small angle shown by chimpanzees results in ahigher transverse arch than normally seen in humans. The human arch, however, unlike that of chim-panzees, exhibits a fixed height. Owing to a small, irregular, and often absent cuboid facet, the humantalocuboid angle could not be accurately measured.
based on localized morphology do not ex-clude the likelihood that these morphologiesand their implied localized functions alsoserved other behaviors (table 12). The pres-ence of many of the alleged bipedal charac-ters in cursorial quadrupeds and the mechan-ical advantages these characters impart toquadrupeds, support the theory that many ofthe allegedly bipedal human characters aroseas a result of selection pressures for cursorialquadrupedality (Sarmiento, 1998). The ab-sence of a humanlike lumbar lordosis, theorientation of the acetabulum’s lunate sur-face, and the relative size of the humeral andfemoral midshaft circumferences and first sa-cral body cross-sectional area, all characterswhich are altered ontogenetically as a resultof use, show that AL 288-1 had a body-to-limb weight distribution and pelvic joint pos-tures similar to those of quadrupeds (Sar-miento, 1998).
Although it may be argued that the pelvisis phylogenetically more conservative thanthe navicular (see above) and thus the latteris more likely to reflect use, while the formeris more likely to reflect heritage, the Hadarpelvic, femoral, and navicular morphology ifassociated must have satisfied the mechanicalrequisites of many of the same behaviors. Inthis regard, it is not possible to fully interpretwhat the range of these behaviors may havebeen without analyzing all of the animal’s
morphology. No matter how bipedal, quadru-pedal or climbing-like a localized morphol-ogy may be, it is usually not possible to pre-dict on the basis of that morphology alonethe overall morphology of an unknown fossiltaxon and the range of behaviors the un-known morphology enabled. In this regard,functional analysis of the navicular alone islimited to the functional role the navicularmay have had in the foot, and in a range ofhypothesized behaviors usually based onthose seen in living taxa.
FOSSIL FOOT USE
OH 8
In OH 8 the comparatively shallow talarfacet, small sagittal taloectocuneiform andentoectocuneiform angles and the large trans-verse mesoectocuneiform angle suggest thepresence of a fixed longitudinal arch. The lat-erally expanded tubercle midway on the lat-eral border of the navicular which serves asthe attachment site for the calcaneonavicular(spring) ligament, and the large articular areafor this ligament on the talar head indicate alongitudinal plantar arch with energy-storingproperties comparable to or approximatingthose of humans (Alexander, 1989). A dor-siflexed talar head and plantar processes onthe navicular and entocuneiform, however,suggest the longitudinal arch height was low-

2000 29SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_29File # 01cc
er than is characteristic of humans, and thedistal tarsal row contacted the substrate (Dayand Wood, 1968). Given a fixed longitudinalarch, however, only a small percentage of theweight borne by the foot could have beentransferred to the substrate by the navicularand entocuneiform, hence the relativelysmall plantar tubercles.
Although the small frontal talocuboid an-gle (table 2) indicates a high transverse pedalarch, the development of a hamulus andgroove for the peroneus longus tendon on theplantar surface of the ectocuneiform indi-cates this arch was not fixed. Consideringvery little rotatory mobility at the talonavic-ular (as implied by the shallow talar facetwith different mediolateral and dorsoplantarradii of curvature) and calcaneocuboid joints(Lewis 1980c), transverse arch mobility musthave resulted mainly from movement at thesubtalar joint.
The sagittal entoectocuneiform angle andthe set of the talonavicular, naviculoentocu-neiform, and the first tarsometatarsal jointsall indicate an adducted hallux which wasachieved through a different combination ofjoint sets than that seen in humans (Day andWood, 1968; Oxnard and Lisowski, 1980;Oxnard, 1984). Although some degree of op-posability may have been possible, depend-ing on the unknown metatarsophalangealjoint set, this would have been very limited.
The adducted hallux and large transversemesoectocuneiform angle reflect a compactfoot that was strongly committed to terres-triality. The relatively small cuboid facetsuggests the foot was usually loaded in pro-nated postures (with the foot’s mediolateralaxis approximating the horizontal and itslong axis in the plane of forward movement).The relatively large ectocuneiform facet (fig.3, tables 5 and 7) and small mesocuneiformand entocuneiform facets (figs. 4 and 5, ta-bles 5 and 7) indicate a preference for load-ing the lateral side of the foot at the expenseof the hallux. Differences relative to humansin the orientation of the subtalar and talocru-ral joints (Lewis, 1980c) probably reflect dif-ferences in terrestrial foot use in both qua-drupedal and bipedal behaviors, and mayhave also been important for proper foot ori-entation in vertical climbing or walkingalong horizontal branches. Given the rela-
tionship between body size and tree use(Cartmill and Milton, 1977; Sarmiento, 1983,1985, 1988, 1994, 1998; Cartmill, 1985), ananimal corresponding in size to the OH 8foot, regardless of its terrestrial specializa-tions, was no doubt better suited to move ar-borealy than humans and or male gorillas(Sarmiento, 1985, 1994, 1998).
Hadar
When considered in light of the other Al333/333x pedal remains, the Hadar navicu-lars do provide considerable insight into footuse and function. The large sagittal taloec-tocuneiform angle, the large degree of dor-soplantar talar facet curvature (table 1) andthe large inflated navicular tuberosity indi-cate that the Hadar foot lacked the longitu-dinal plantar arch characteristic of modernhumans. The navicular’s large plantar tuber-osity and the overlying horizontal portion ofthe talar facet enabled talar head loads to betransmitted directly to the substrate throughthe navicular. The well-developed plantarprocess on the entocuneiform (AL 333-28)and the plantar set of the naviculoectocunei-form facet suggest the entocuneiform alsoparticipated in transferring weight to the sub-strate. Although the high values of the frontalmesoectocuneiform angle and low values ofthe frontal talocuboid angles (table 2) reflectsome degree of a transverse pedal arch, thepresence of a plantar process flanked distallyby the peroneus longus groove on the AL333-79 ectocuneiform suggests the trans-verse arch was either lower than that of hu-mans or was mobile. Given a high and fixedtransverse arch, the peroneus longus tendonbowstrings and does not groove the plantarsurface of the ectocuneiform (Sarmiento,1994). The implied mobility at the talonavic-ular joint also indicates the Hadar foot didnot have a fixed transverse pedal arch.
The large cuboid facet (fig. 6 and tables 6and 8) and mobility at the talonavicular jointsuggest the Hadar foot was loaded in supi-nated postures as is customary among greatapes when climbing vertical supports of rel-atively large diameter (Sarmiento, 1985.1994). Considering the large, inflated tubercalcaneus (Latimer and Lovejoy, 1989),prominent plantar processes on the anterior
30 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_30File # 01cc

2000 31SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_31File # 01cc
calcaneus, navicular, and entocuneiform (thisstudy), the dorsiflexed set at the metacarpo-phalangeal joints, and the short phalanges(Stern and Susman 1983; Latimer and Love-joy, 1990a,b), the Hadar foot was much bet-ter suited for terrestrial plantigrade posturesthan for arboreal grasps. Substrate contact bythe distal tarsus indicates a human bipedalstride involving heel-to-ball weight transfercould not have been commonly employed.The more divergent set of the second andthird rays as implied from the transverse me-soectocuneiform angle and the mobile trans-verse arch suggests a less compact foot thanin modern humans, one which was not asstrongly committed to terrestrial behaviors.
Mobility at the talonavicular joint and therelative plantar orientation of the entocunei-form facet indicates more abducted postures
of the hallux in Hadar than is charactersticof humans. Because the degree of hallucalabduction is defined by the set of the talo-navicular, naviculoentocuneiform, first tar-sometatarsal and metatarsophalangeal joints(Day and Wood, 1968; Lewis, 1980b, c; Sar-miento, 1994), and these are not all presentamong the Hadar AL 333/333w remains, es-timates as to degree of hallucal abduction areequivocal. Regardless, a degree of hallucalabductability comparable to or greater thanthat seen in mountain gorillas is a reasonableestimate.
The Hadar navicular is best interpreted asbelonging to the foot of a generalized quad-ruped which probably employed plantigradebipedal postures and limited its arboreal be-haviors to supports of relatively large diam-eter (Sarmiento, 1989, 1991, 1998).

32 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_32File # 01cc
Oreopithecus
The large sagittal taloectocuneiform andentoectocuneiform angles and the relativelysmall transverse mesoectocuneiform angle(table 1 and 2) indicate the Oreopithecus foothad markedly divergent rays. The low navic-ular torsion with the strong medial inclina-tion of the talar neck specifically indicate amarkedly divergent hallux. Among the hom-inoids such a marked divergence is unique toOreopithecus and is corroborated when theassociated tarsal and metatarsal bones are ar-ticulated (Kohler and Moya-Sola, 1997). Ashallow talar facet with small and approxi-mately equal radii of curvature (table 4) cor-responds to a loose talonavicular joint withrotatory ability. Such joint laxity is also re-flected in the calcaneo-cuboid joint (Sar-miento, 1987; Szalay and Langdon, 1987)and in the single continuous navicular faceton the talar head. A talar facet which is largerelative to the cuneiform facets (fig. 8) sug-gests an emphasis on mobility at the talona-vicular relative to the naviculocuneiformjoints. The absence of weight-bearing tuber-cles on the navicular and entocuneiform(Sarmiento, 1987) suggest that the footlacked a commitment to terrestrial behaviorsand/or was not often used for walking alonglarge diameter horizontal supports. The largecuboid facet (fig. 6 and table 6) is in accordwith powerful hallucal opposability and afoot loaded in supinated postures.
The Oreopithecus navicular best corre-sponds to the foot of an arboreal verticalclimber. A relatively mobile foot with a wideopposable grasp, this foot could be apposedagainst vertical trunks when climbing, orused along horizontal supports of diameterspermitting hallucal grasps.
PHYLOGENY IMPLICIT IN NAVICULAR
MEASUREMENTS
It is striking how close the phenetic treebased on the Mahalanobis distances betweensamples of human and great ape navicularmeasurements (fig. 13) coincide with homi-noid phylogenies as hypothesized by earlyanatomists (Keith, 1916, 1934, 1940; Morton1927; Schultz, 1936; Le Gros Clark, 1971).Because navicular measurements were cho-sen to reflect largely functional concerns, co-
inciding phylogenies and phenetics may beunexpected, especially when epigenetic andinherited characters are given equal weight.Early anatomists and systematists, however,did not usually test the inferred homologiesused to construct phylogenies for the likeli-hood of parallelisms (Sarmiento, 1998).Thus, these phylogenies are in essence phe-netic trees, i.e., they equate degree of overallmorphological similarity with the degree ofrelationship. Without the morphologicalcomplexity necessary to test for homologies,quantification of overall similarities in frag-mentary fossils may be the only analysis pos-sible. Results from such an analysis, how-ever, do not accurately reflect phylogeneticaffinities, since they conflate shared derivedcharacters (synapomorphies) and parallel-isms (plesiomorphies) and do not distinguishinherited characters from those originatingontogenetically with use.
Through cladistic analysis and knowledgeas to what navicular characters are primitivevs. derived, testing for homologies may bepossible. For instance, a weight-bearing plan-tar tubercle with a variable sustentacular ar-ticulation is a shared derived character of theAfrican ape navicular absent in other catar-rhines and the earliest known fossil homi-noids (i.e. Oreopithecus and Sivapithecus;Sarmiento, 1994, this study). The presence ofa large weight-bearing plantar tubercle and asustentacular facet on the navicular of Hadarforms may be used to argue that many of theangles and metric characters shared by Hadarand African apes are also homologous. Assuch, the Hadar naviculars would seem toshare a special relationship with African apesexclusive of humans and orangutans. As withall cladistic analyses this argument assumesorthoselection with minimal reversals. In thiscase, the assumptions are that the large plan-tar tubercle and sustentacular facet was not(1) independently acquired by chimpanzees,gorillas and/or Hadar fossils, (2) a sharedtrait of humans and African apes which waslater lost in humans, or (3) a shared hominoidtrait that was independently lost in humans,orangutans, and Oreopithecus. With relative-ly few comparative taxa as outgroups and alimited number of shared derived charactersfor comparison, such an analysis may proveequivocal. In most cases more than a bit of

2000 33SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_33File # 01cc
localized morphology is necessary for phy-logenetic resolution (Sarmiento, 1987).
Regardless of its inaccuracy, navicularphenetics and the cladistic analysis that canbe done on the limited number of navicularcharacters does reflect phylogeny to somedegree. In this regard, there must be a limitas to how much the naviculars of closely re-lated forms may differ from each other and/or how similar the navicular of more distallyrelated forms may be. Similarities in inher-ited anatomy predispose the types of prob-lems an organism encounters in its environ-ment and the behaviors used to solve theseproblems. Differences in inherited anatomy,on the other hand, reduce the likelihood ofencountering similar problems in the envi-ronment or arriving at similar solutions. Evenif the same problems are encountered andsimilar solutions arrived at, inherited differ-ences are more apt to result in greater dif-ferences in navicular morphology in moredistally related taxa than in closely relatedones.
Notably navicular phenetics also approxi-mates the currently accepted hominoid phy-logeny (Sarmiento, 1998). The only dis-agreement exists at the point of human di-vergence, and humans have a decidedly spe-cialized foot committed to terrestrialbipedality (Weidenreich, 1922; Morton,1924; Schultz 1963; Sarmiento, 1994, 1998).The close similarities in the navicular of OH8 and humans, and of Hadar australopithe-cines and African apes, and the unique na-vicular of Oreopithecus must reflect to somedegree phylogenetic affinities. This applieswhether or not the measured characters alsoreflect functional concerns. As regards thenavicular, the AL 333/333w remains aremore likely to represent ancestral Africanapes than ancestral humans. The measureddifferences between OH 8, Hadar, and Or-eopithecus naviculars are consistent with ge-neric differences among humans and livinggreat apes.
CONCLUSIONS
The Oreopithecus, Hadar, and OH 8 na-viculars, in order of decreasing geologic age,show decreasing hallucal abduction, increas-ing commitment to terrestrial behaviors, and
a decreasing Mahalanobis distance to humannaviculars. Such a sequence of evolutionarychanges in the foot are in accord with thosepredicted in human phylogeny by currentmodels of hominoid and hominid divergence(Sarmiento, 1995, 1998).
Results from this study inspire caution asto how much of an animal’s phylogeny oroverall behavior can be interpreted from asingle bone. Phylogenies are unlikely to beaccurate without the morphologic complexitynecessary to test for homologies, and withoutat least a limited understanding of characterpolarity. When interpreting either phylogenyand/or function from fossil remains, incom-plete remains are likely to lead to spuriousconclusions. Interpretations of australopithe-cine systematics and behavior, therefore, canonly be credible when all of the morpholog-ical evidence is accounted for. In retrospect,the notion that the australopithecine gaitcould have been predicted based on a singleos coxa (Sts 14) and some nonassociatedfragmentary femora (Lovejoy, 1974; see alsoJohanson et al., 1976) is supercilious.
ACKNOWLEDGMENTS
I thank the following for access to and as-sistance with comparative collections: W.Van Neer, Royal African Museum Tervuren;R. Thorington, National Museum of NaturalHistory, Washington D.C.; O. Grondwall,Swedish Museum of Natural History, Stock-holm; Malcom Harmon, Powell Cotton Mu-seum, Birchington Kent; Maria Rutzmoser,Museum of Comparative Zoology, Cam-bridge Massachusetts; and J. Kerbis-Peter-Hans, Field Museum of Natural History, Chi-cago. For access to fossil material I am grate-ful to: Solomon Woerde Kal of the NationalMuseum of Ethiopia, Addis Ababa; M. L.Mbago of the National Museums of Tanza-nia, Dar es Salaam; M. Mazzinni, Istituto diGeologia, University of Florence; B. Enges-ser and the late J. Hurzeler, Natural HistoryMuseum, Basel. The senior author wouldalso like to extend a special thanks to M.Kohler and S. Moya-Sola for their gracioushopsitality during a visit to Sabadell to ex-amine Oreopithecus remains in their care. M.D. Rose and F. J. Rohlf reviewed an earlier

34 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_34File # 01cc
version of this manuscript and provided help-ful suggestions for its improvement.
REFERENCES
Abitbol, M. M.1988. Evolution of the ischial spine and of the
pelvic floor in the Hominoidea. Am. J.Phys. Antropol. 75: 53–67.
Alexander, R. McN.1989. Elastic mechanisms in animal move-
ment. New York: Cambridge Univ.Press.
Beard, K. A., M. Dagosto, D. Gebo and M. God-inot
1988. Interrelationship among primate highertaxa. Nature 331: 712–714.
Biegert, J.1963. The evaluation of characteristics of the
skull, hands and feet for primate tax-onomy. In S. L. Washburn (ed.), Clas-sification and human evolution: 116–145. Chicago: Aldine.
Biegert, J., and R. Mauer1972. Rumpskelettlange, Allometrien und
Korperproportionen bie catarrhinen Pri-maten. Folia Primatol. 17: 142–156.
Broom, R., and J. T. Robinson1950. Notes on the pelves of the fossil ape-
man, Pleisianthropus. Am. J. Phys. An-thropol. 8: 489–494.
Carrano, M. T.1997. Morphological indicators of foot pos-
ture in mammals, a statistical and bio-mechanical study. Zool. J. Linn. Soc.12: 77–104.
Cartmill, M.1985. Climbing. In M. Hildebrand, D. M.
Bramble, K. F. Leim and D. B. Wake(eds.), Functional vertebrate morphol-ogy: 73–88. Cambridge: Belknap Press.
Cartmill, M., and K. Milton1977. The lorisform wrist joint and the evo-
lution of brachiating adaptions in theHominoidae. Am. J. Phys. Anthropol.47: 249–272.
Chamberlain, A. T. and B. A. Wood1987. Early Hominid phylogeny. J. Hum.
Evol. 16: 119–133.Clarke, R. J., and P. V. Tobias
1995. Sterkfontein member 2 foot bones ofthe oldest South African hominid. Sci-ence 269: 521–524.
Conroy, G. C. and J. G. Fleagle1972. Locomotor behaviors in living and fos-
sil pongids. Nature 237: 103–104.Darwin, C.
1859. The origin of species by means of nat-
ural selection or the preservation of fa-vored races in the struggle of life. NewYork: Modern Library.
Day, M. H., and J. R. Napier1964. Fossil foot bones. Nature 201: 969–
970.Day, M. H., and B. A. Wood
1968. Functional affinities of the Olduvaihominid 8 talus. Man 3: 440–455.
Edelstein, S. J.1987. An alternative paradigm for hominoid
evolution. Hum. Evol. 2: 169–174.Elftman, H.
1960. The transverse tarsal joint and its con-trol. Clin. Orthop. 16: 41–46.
Elftman, H. and J. Manter1935. The evolution of the human foot, with
special reference to the joints. J. Anat.70: 56–67.
Ferguson, W. W.1984. Revision of fossil hominid jaws from
the Plio/pleistocene of Hadar, in Ethi-opia, including a new species of the ge-nus Homo (Hominoidea; Homininae).Primates 25: 519–529.
1986. The taxonomic status of Praeanthropusafricanus (Primates: Pongidae) fromthe late Pliocene of eastern Africa. Ibid.27: 485–492.
Fleagle, J. G.1998. Primate adaptation and evolution. 2nd
ed. New York: Academic Press.Gebo, D. L.
1986. The anatomy of the prosimian foot andits application to the primate fossil re-cord. Ph.D. Diss. Duke Univ., Durham.
Gebo, D. L., and M. Dagosto1988. Foot anatomy, climbing and the origin
of the Indriidae. J. Human Evol. 17:135–154.
Gommery, D.1997. Le atlas et les axis des Hominides du
Plio-pleistocene: morphologie et syste-matique. C.R. Acad. Sci. Paris 325:639–642.
Heiple, K. G., and C. O. Lovejoy1971. The distal femoral anatomy of Austral-
opithecus. Am. J. Phys. Anthropol. 35:75–84.
Hill, W.C.O.1960. Primates: comparative anatomy and
taxonomy. IV. Cebidae, Pt. A. Edin-burgh: Edinburgh Univ. Press.
Johanson, D. C., C. O. Lovejoy, K. G. Heiple, andA. H. Burstein
1976. Biomechanical implications of the Afarknee joint. Am. J. Phys. Anthropol. 44:188.

2000 35SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_35File # 01cc
Johanson, D. C., M. Taieb, and Y. Coppens1982. Pliocene hominids from the Hadar for-
mation, Ethiopia (1973–1977): strati-graphic, chronologic and paleo-envi-ronmental contexts, with notes on hom-inid morphology and systematics. Am.J. Phys. Anthropol. 57: 373–402.
Jolly, C. J.1965. The origins and specializations of the
long-faced Cercopithecoidea. Ph.D.diss., Univ. of London.
1970. The seed eaters: a new model of hom-inid differentiation based on baboonanalogy. Man 5: 5–26.
Keith, A.1916. Lo schema dell’origine umana. Revista
di Antropologia 20: 3–20.1934. The construction of man’s family tree.
London: Watts.1940. Fifty years ago. Am. J. Phys. Anthro-
pol. 26: 251–267.Kohler, M., and S. J. Moya-Sola
1997. Ape-like or hominid-like? The posi-tional behavior of Oreopithecus bam-bolii reconsidered. Proc. Natl. Acad.Sci. 94: 11747–11750.
Latimer, B.1991. Locomotor adaptations in Australopi-
thecus afarensis: the issue of arboreal-ity. In Y. Coppens and B. Senut (eds.),Origine(s) de la bipedie ches les hom-inides: 169–176. Paris: CNRS.
Latimer, B., and C. O. Lovejoy1989. The calcaneus of Australopithecus
afarensis and its implication for theevolution of bipedality. Am. J. Phys.Anthropol. 78: 369–386.
1990a. Hallucal tarsometatarsal joint in Aus-tralopithecus afarensis. Ibid. 82: 125–133.
1990b. Metatarsophalangeal joints of Austral-opithecus afarensis. Ibid. 83: 13–23.
Latimer, B., C. O. Lovejoy, D. C. Johanson, andY. Coppens
1982. Hominid tarsal, metatarsal and phalan-geal bones recovered from the Hadarformation: 1974–1977 collections. Am.J. Phys. Anthropol. 57: 701–719.
Latimer, B., J. C. Ohman, and C. O. Lovejoy1987. Talocrural joint in African hominoids:
implications for Australopithecus afar-ensis. Am. J. Phys. Anthropol. 74: 155–175.
Leakey, L., P. V. Tobias, and J. Napier1964. A new species of the genus Homo from
Olduvai Gorge. Nature 202: 7–10.
Leakey M. G., C. S. Feibel, I. McDougall, and A.Walker
1995. New four-million-year-old hominidspecies from Kanapoi and Allia bay,Kenya. Nature 376: 565–571.
Le Gros Clark, W. E.1955. The os inominatum of the recent Pon-
gidae with special reference to that ofthe Australopithecinae. Am. J. Phys.Anthropol. 13: 19–27.
1971. The antecedents of man. New York:Quadrangle Press.
Lewis, O. J.1969. The hominoid wrist joint. Am. J. Phys.
Anthropol. 30: 251–267.1974. The wrist articulations of the Anthro-
poidea. In F. A. Jenkins (ed.), Primatelocomotion: 143–169. New York: Ac-ademic Press.
1980a. The joints of the evolving foot. Pt. I.The ankle joint. J. Anat. 130: 527–543.
1980b. The joints of the evolving foot. Pt II.The intrinsic joints. Ibid.: 833–857.
1980c. The joints of the evolving foot. Pt. III.The fossil evidence. Ibid. 131: 275–298.
Lovejoy, C. O.1974. The gait of australopithecines. Yearb.
Phys. Anthropol. 17: 147–161.1978. A biomechanical review of the loco-
motor diversity of early hominids. In C.J. Jolly (ed.), Early Hominids of Africa,403–429. New York: St. Martins Press.
1979. A reconstruction of the pelvis of AL-288 (Hadar Formation, Ethiopia). Am.J. Phys. Anthropol. 50: 460.
1988. Evolution of human walking. Sci. Am.259: 118–125.
Lovejoy, C.O., K.G. Heiple, and A.H. Burstein1973. The gait of Australopithecus. Am. J.
Phys. Anthropol. 32: 33–40.Lovejoy, C.O., T. D. White, M. J. Cohn, and J. C.
Ohman1996. The Maka femur: A case study in the
application of analytical paradigms dic-tated by new developments in morpho-logical ontogeny. Am. J. Phys. Anthro-pol., Suppl. 22: 152.
MacLatchy, L. M.1996. Another look at the Australopithecine
hip. J. Hum. Evol. 31: 455–476.Manter, J. T.
1941. Movements of the subtalar and trans-verse tarsal joints. Anat. Rec. 80: 397–410.
Marcus, L. F.1990. Traditional morphometrics. In F. J.
Rohlf and F. L. Bookstein (eds.). Pro-

36 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_36File # 01cc
ceedings of the Michigan morphomet-rics workshop: 77–112. Univ. MichiganMus. Zool. Spec. Publ.
1993. Some aspects of multivariate statisticsfor morphometrics. In L. F. Marcus, E.Bello, and A. G. Valdecasas, (eds.),Contributions to morphometrics: 99–130. Madrid: Monografias Museo Nac-cional Ciencias Naturales.
McHenry, H. M.1991. First steps? Analyses of the postcran-
ium of early hominids. In Y. Coppensand B. Senut (eds.), Origine(s) de la bi-pedie chez les hominides: 133–141.Paris: CNRS.
1996. Homoplasy, clades and hominid phy-logeny. In W. E. Meikle, F. C. Howell,and N. G. Jablonksi (eds.), Contempo-rary issues in human evolution: 77–91.San Francisco: California Acad. Sci.
McHenry, H. M., R. S. Corruccinni, and F. C.Howell
1975. Analysis of an early hominid ulna fromthe Omo basin, Ethiopia. Am. J. Phys.Anthropol. 44: 295–304.
Mednick, L. W.1955. The evolution of the human ilium. Am.
J. Phys. Anthropol. 13: 203–216.Meldrum, D. J., M. Dagosto, and J. White
1997. Hindlimb suspension and hind foot re-versal in Varecia variegata and otherarboreal mammals. Am. J. Phys. An-thropol. 103: 85–102.
Morton, D. J.1924. Evolution of the human foot. II. Am. J.
Phys. Anthropol. 7: 1–52.1927. Human origin. Am. J. Phys. Anthropol.
10: 173–203.Napier, J. H.
1962. Fossil hand bones from Olduvai Gorge.Nature 196: 409.
Ohman, J. C., T. J. Krochta, C. O. Lovejoy, R. P.Mensforth, and B. Latimer
1997. Cortical bone distribution in the femo-ral neck of hominoids: implications forthe locomotion of Australopithecusafarensis. Am. J. Phys. Anthropol. 104:117–131.
Olson, T. R.1978. Hominid phylogenetics and the exis-
tence of Homo in Member I of theSwartkrans formation, South Africa. J.Hum. Evol. 7: 159–178.
1981. Basicranial morphology of the extanthominoids and Pliocene hominids: thenew material from the Hadar forma-tion, Ethiopia, and its significance inearly human evolution and taxonomy.
In C. B. Stringer (ed.), Aspects of hu-man evolution: 99–128. London: Tay-lor and Francis.
1985. Cranial morphology and systematics ofthe Hadar fossil hominids and A. afri-canus. In E. Delson and I. Tattersall(eds.), Ancestors: the hard evidence:102–119. New York: Alan R. Liss.
Oxnard, C. E.1984. The Order of Man. New Haven, CT:
Yale Univ. Press.Oxnard, C. E., and F. P. Lisowski
1980. Functional articulation of some homi-noid foot bones: implications for theOlduvai (Hominid 8) foot. Am. J. Phys.Anthropol. 52: 107–117.
Rafferty, K. A.1998. Structural design of the femoral neck in
primates. J. Hum. Evol. 34: 361–383.Reynolds, E.
1931. The evolution of the human pelvis inrelation to the mechanics of erect pos-ture. Pap. Peabody Mus. Archaeol. Eth-nol. Harvard Univ. 11: 249–334.
Robinson, J. T.1965. Homo habilis and the australopithe-
cines. Nature 205: 121–124.1972. Early hominid posture and locomotion.
Chicago: Univ. Chicago Press.Rohlf, F. J.
1997. NTSYSpc version 2.02h. Setauket, NY:Exeter Software.
Rose, M. D.1988. Functional anatomy of the cheiridia. In
J. H. Schwartz (ed.), Orangutan biolo-gy: 509–524. London: Plenum Press.
Sarmiento, E. E.1983. The significance of the heel process in
anthropoids. Int. J. Primatol. 4(2): 127–152.
1985. Functional differences in the skeletonof wild and captive orang-utans andtheir adaptive significance. Ph.D. diss.,New York Univ., New York.
1987. The phylogenetic position of Oreopi-thecus and its significance in the originof the Hominoidea. Am. Mus. Novita-tes 2881: 44 pp.
1988. Anatomy of the hominoid wrist joints,its evolutionary and functional impli-cations. Int. J. Primatol. 9: 281–345.
1991. Functional and phylogenetic implica-tions of the differences in the pedalskeleton of Australopithecines. Am. J.Phys. Anthropol., Suppl. 12: 232.
1994. Terrestrial traits in the hands and feetof gorillas. Am. Mus. Novitates 3091:56 pp.

2000 37SARMIENTO AND MARCUS: OS NAVICULAR OF HOMINIDS
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_37File # 01cc
1995. Cautious climbing and folivory: a mod-el of hominoid differentiation. Hum.Evol. 10(4): 289–321.
1998. Generalized quadrupeds, committed bi-peds and the shift to open habitats: anevolutionary model of hominid diver-gence. Am. Mus. Novitates 3250: 78pp.
Sarmiento, E. E., and T. M. Butynski1996. Present problems in gorilla taxonomy.
Gorilla J. 12: 5–7.In prep. The foot musculature of Bwindi go-
rillas: its functional and phylogeneticsignificance.
Sarmiento, E. E., T. M. Butynski, and J. Kalina1996. Gorillas of Bwindi-Impenetrable Forest
and the Virunga Volcanoes: taxonomicimplications of morphological and eco-logical differences. Am. J. Primatol.40: 1–21.
Schaeffer, B.1947. Notes on the origins and function of the
artiodactyl tarsus. Am. Mus. Novitates1356: 24 pp.
Schultz, A. H.1930. The skeleton of the trunks and limbs of
higher primates. Hum. Biol. 2: 303–438.
1936. Characters common to higher primatesand characters specific to man. Q. Rev.Biol. 11: 259–283; 425–455.
1963. Relations between the lengths of themain part of the foot skeleton in pri-mates. Folia Primatol. 1: 150–171.
Senut, B., and C. Tardieu1985. Functional aspects of hominid limb
bones: implication for taxonomy andphylogeny. In E. Delson and I. Tatter-sall (eds.), Ancestors: the Hard Evi-dence: 175–203. New York: Alan R.Liss.
Skelton, R. R., H. M. McHenry, and G. M. Draw-horn
1986. Phylogenetic analysis of early homi-nids. Curr. Anthropol. 27: 21–43.
Stern, J. T., and R. L. Susman1983. The locomotor anatomy of Australopi-
thecus africanus. Am. J. Phys. Anthro-pol. 60: 279–316.
Strait, D.S., F. E. Grine, and M. A. Moniz1997. A reappraisal of early hominid phylog-
eny. J. Hum. Evol. 37: 17–82.Straus, W. L., Jr.
1929. Studies on the primate ilia. Am. J.Anat. 43: 403–460.
Susman, R. L., and J. T. Stern1991. Locomotor behavior of early hominids:
epistomology and fossil evidence. In Y.
Coppens and B. Senut (eds.), Origi-ne(s) de la bipedie chez les hominides:121–131. Paris: CNRS.
Susman, R. L., J. T. Stern, Jr., and W. L. Jungers1984. Arboreality and bipedality in the Hadar
hominids. Folia Primatol. 43: 113–156.Szalay, F. S., and R. L. Decker
1974. Origin, evolution and function of thetarsus in late Cretaceous eutherians andPaleocene primates. In F. Jenkins (ed.),Primate locomotion: 223–259. NewYork: Academic Press.
Szalay, F. S., and J. H. Langdon1987. The foot of Oreopithecus: an evolution-
ary assessment. J. Hum. Evol. 15: 585–621.
Tardieu, C., and H. Preuschoft1996. Ontogeny of the knee joint in humans,
great apes and fossil hominids: Pelvi-femoral relationships during postnatalgrowth in humans. Folia Primatol. 66:68–81.
Tobias, P. V.1991a. Olduvai Gorge, Vol. 4: the skulls en-
docasts and teeth of Homo habilis.Cambridge: Cambridge Univ. Press.
1991b. Relationships between apes and hu-mans. In A. Sahni and R. Gaur (eds.),Perspectives in human evolution: 1–19.Delhi: Renaissance Publishing House.
Tuttle, R. H.1970. Postural propulsive and prehensile ca-
pabilities in the cheiridia of chimpan-zees and other great apes. In G. H.Bourne (ed.), The chimpanzee. 2: 167–253. Basel: Karger.
Verhaegen, M.1990. African ape ancestry. Hum. Evol. 5:
295–297.1994. Australopithecines: ancestors of the Af-
rican apes? Ibid. 7: 63–64.Walker, A.C.
1973. New Australopithecus femora fromEast Rudolf Kenya. J. Hum. Evol. 2:545–555.
Ward, C.V., M.G. Leakey, B. Brown, F. Brown, J.Harris and A. Walker
1999. South Turkwel: A new Pliocene homi-nid site in Kenya. J. Hum. Evol. 36:69–95.
Waterman, H. C.1927. Studies on the evolution of the pelves
of man and other primates. Bull. Am.Mus. Nat. Hist. 11: 585–641.
Weidenreich, F.1922. Der Menschenfuss. Z. Morphol. An-
thropol. 22: 51–282.

38 NO. 3288AMERICAN MUSEUM NOVITATES
AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_38File # 01cc
White, T. D., D. C. Johanson, and W. H. Kimbel1981. Australopithecus africanus: its phylo-
genetic position reconsidered. S. Afr. J.Sci. 77: 445–470.
Wolpoff, M. H.1974. The evidence of two australopithecine
lineages in South Africa. Yearb.Phys.Anthropol. 17: 113–139.
1999. Paleoanthropology. New York: Mc-Graw Hill.
Wood, B. A.1974. Olduvai Bed I postcranial fossils; a re-
assessment. J. Hum. Evol. 3: 373–378.
Zihlman, A. L.1969. Human locomotion: a reappraisal of the
functional and anatomical evidence.Ph.D. diss., Univ. California at Berke-ley.
1978. Interpretations of early hominid loco-motion. In C. J. Jolly (ed.), Early Hom-inids of Africa: 361–377. New York: StMartins Press.
Zihlman, A. L., and L. Bunker1979. Hominid bipedalism: then and now.
Yearb. Phys. Anthropol. 22: 132–162.

AMNH NOVITATESTuesday Dec 11 2001 10:13 AM 2000Allen Press • DTPro System
novi 99146 Mp_40File # 01cc
No RH this page!!
Recent issues of the Novitates may be purchased from the Museum. Lists of back issues of theNovitates and Bulletin published during the last five years are available at World Wide Web sitehttp://nimidi.amnh.org. Or address mail orders to: American Museum of Natural History Library,Central Park West at 79th St., New York, NY 10024. TEL: (212) 769-5545. FAX: (212) 769-5009. E-MAIL: [email protected]
a This paper meets the requirements of ANSI/NISO Z39.48-1992 (Permanence of Paper).








![HADAR[Hadar + Controller] rises Hadar in the real life! Of course, Hadar as an industrial real-time dispatcher has not in the scope. However, Hadar can be in hours used in a Raspberry](https://img.pdfslide.us/doc/110x75/6024261f646a3256b0433968/hadar-hadar-controller-rises-hadar-in-the-real-life-of-course-hadar-as-an.jpg)