Embed Size (px)
Citation preview

THE JOURNAL Q 1989 by The American Society for Blochemistry and Molecular Biology, Inc.
OF BIOLOGICAL CHEMfSTRV Vol. 264, No , 31, Iesue of November 5, pp. 18589-18597,1989 Printed in U.S.A.
Purification of a Prenyltransferase That Elongates cis-Polyisoprene Rubber from the Latex of Heuea brusiliensis”
(Received for publication, June 2, 1989)
David R. Light$ and Mark S . Dennis From the Department of Medicinal and Bwmolecular Chemistry, Genentech, Inc., South San Francisco, California 94080
We have purified “rubber transferase* from latex of the commercial rubber tree Hevea brasiliemis and find that it is a dimer with a monomeric molecular mass of 38,000 Da, requires M8+, and is stabilized by thiols in agreement with studies of a partially purified prepa- ration previously described (Archer, B. L., and Cock- bain, E. G. (1969) Methods Enzymol. 15, 476-480). Greater than 90% of the [l-’4C]isopentenyl pyrophos- phate which is incorporated into deproteinated rubber particles by the purified prenyltransferase is added to high molecular mass polyisoprene (>20,000 Da). Pu- rified prenyltransferase and deproteinated rubber par- ticles reconstitute 40-60% of the biosynthetic activity of whole latex in samples matched for rubber content. Incorporation is linear with added rubber particles up to at least 10 mg/ml rubber or 20 I .~M rubber molecules (based on a number average molecular mass of 500,000 Da).
Prenyltransferase concentrations estimated in whole latex (0.37% or 160 nM) are sufficient to saturate all elongation sites in whole latex, and addition of purified prenyltransferase does not increase [ l-’4C]isopentenyl pyrophosphate incorporation. Deproteinated rubber particles can be titrated with the pure enzyme (Ka = 9 nM) demonstrating that the fraction of rubber mole- cules available for addition is low (approximately 0.01%). An estimated 7,000 isoprene units are added per complex at a rate of l/s in a typical assay.
Hevea prenyltransferase catalyzes the formation of cis-isoprene in the presence of rubber particles. How- ever, in the absence of rubber particles and in the presence of dimethylallyl pyrophosphate, the purified prenyltransferase catalyzes the formation of geranyl pyrophosphate and all tram-farnesyl pyrophosphate as demonstrated by thin layer chromatography, gas chromatography, and molecular exclusion chromatog- raphy.
Rubber transferase (1-5) is a prenyltransferase that cata- lyzes the polymerization of isopentenyl pyrophosphate (IPP)’ into rubber. Like other prenyltransferases this enzyme re-
* This paper is the first in a series on the biochemistry of cis- polyisoprene rubber synthesis. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed. The abbreviations used are: IPP, isopentenyl pyrophosphate;
FPP, farnesyl pyrophosphate; t,t-FPP, all-trans-FPP; DMAPP, di- methylallyl pyrophosphate; GPP, geranyl pyrophosphate; DTT, di- thiothreitol; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; FPLC, fast protein liquid chromatography; HPLC, high performance liquid chromatography; GC/MS, gas chromatogra- phy/mass spectrometry; M,,, number average molecular mass.
quires a divalent cation ( M e ) to catalyze thousands of 1-4 cis condensations (Z-oligomerization) of IPP, the prenyl ac- ceptor, to rubber, the prenyl donor, before random termina- tion occurs. Other prenyltransferases terminate after a char- acteristic and comparatively low number of additions of IPP to the allylic pyrophosphate substrate. FPP synthase (from porcine and avian liver, Bacillus subtilis, plants and yeast) catalyzes two trans additions to DMAPP (6-10); geranylger- any1 pyrophosphate synthase catalyzes three trans additions to DMAPP (11,12); hexaprenyl pyrophosphate synthase cat- alyzes three trans additions to t,t-FPP (13); heptaprenyl py- rophosphate synthase catalyzes four trans additions to t,t- FPP (14); solanesyl (nonaprenyl) pyrophosphate synthase catalyzes six to seven trans additions to GPP (15); un- decaprenyl pyrophosphate synthase catalyzes seven to eight cis additions to t,t-FPP (12, 16-18); and dolichol pyrophos- phate synthase catalyzes 13-18 cis additions to t, t-FPP (19).
Product analysis shows loss of termination precision with increasing chain length and even FPP synthase is observed to make a small amount of all-trans geranylgeranyl pyrophos- phate (Cz0) at less than 1% of the rate of FPP formation (6, 20). The nature of the allylic prenyl donor (21) or the M e concentration (22) affects the chain length made by other prenyltransferases by a few isoprene units. These fluctuations in termination precision are modest compared with the pren- yltransferase from the rubber tree Heuea brasiliensis. Its prod- uct varies between 100,000 and 4,000,000 Da (3), representing between 1,500 and 60,000 isoprene additions per molecule.
Whole latex obtained by tapping the laticifers of H . brasi- liensis contains rubber transferase and one of its substrates, rubber particles. Centrifugation of whole latex produces well defined fractions for use in studies of rubber biosynthesis (3, 23, 24). Rubber particles between 50 and 1500 nm diameter (5, 23, 25-28), microsomes, microfibrils (29), mitochondria, and organelles of incompletely understood function called lutoids and Frey-Wyssling particles are suspended in C-serum. Studies of the biogenesis of laticifers reveal that these vessels result from the resorption of cell walls between longitudinally contiguous rows of cells (2). Thus, C-serum, a complex mixture of soluble proteins, enzymes, metabolites, and salts is derived from the soluble component of the cytosol from these specialized cells. Guayule rubber biosynthesis oc- curs in the cytoplasm of leaf mesophyll and stem parenchyma cells (30-32); however, these cells do not fuse to form laticifers found in H . brasiliensis.
Archer et al. (1) and Madhavan and Benedict (30) partially purified rubber transferase from H. brasiliensis and the gua- yule Parthenium argentatum. There is no report of a homo- geneous preparation of rubber transferase. The partially pure enzymes from both species show farnesyl pyrophosphate syn- thase (FPP synthase) activity. We present a method for purifying rubber transferase from the latex of H. brasiliensis
18589

18590 cis-Polyisoprene Rubber Prenyltransferase
to greater than 95% homogeneity and describe some of its properties.
MATERIALS AND METHODS
Reagent~"[l-~~C]I~opentenyl pyrophosphate (-57 Ci/mol) was purchased from Amersham Corp. [4-"C]Isopentenyl pyrophosphate (50.4 Ci/mol) was purchased from Du Pont-New England Nuclear and used in a limited number of initial studies. Unlabeled isopentenyl pyrophosphate (IPP) was synthesized by published procedures of Davisson et al. (47). The ammonium salt was a gift from Dr. C. Dale Poulter, University of Utah, and Brian Froehler, Genentech, Inc. The ammonium salt of dimethylallyl pyrophosphate (DMAPP) was syn- thesized by the published procedures of Davisson et al. (47) and was a gift from Dr. D. A. Widdowson and Dr. C. Pemberton, Imperial College. Affi-Gel 501, a sulfhydryl-specific organomercuric affinity column and Triton X-100 were purchased form Bio-Rad. Hydropho- bic chromatography kits I and I1 (alkyl and aminoalkyl series; aga- rose-(CHz),H or (CH').NHZ, n = 0, 2, 4, 6, 8, 10) were purchased from Miles-Yeda, Ltd. Sephacryl S-300, Sepharose 6B, a 1-ml Mono Q column (quarternary amine ion exchanger), and QAE-Sephadex were purchased from Pharmacia LKB Biotechnology Inc.; Hexyl- agarose and Sephacryl S-200 were purchased from Sigma; DE52 cellulose was purchased from Whatman Chemical Separation, Inc.; and a 7.5 X 600-mm Ultro Pac TSK-G3000 SW high performance molecular exclusion column was purchased from LKB-Producer AB; Dolichol (C56), dolichol mixture (Cm-C1O5), solanesol (C~F,), and ger- aniol (Clo cis) were purchased from Sigma. Nerol (Clo, trans) was purchased from Alfa Products, Thiokol/Ventron Division. Mixtures
or purchased from Fluka AG, Buchs SG, or Aldrich. Isopentenol(3- of varying ratios of isomers of farnesol (CIS) were a gift from Givaudan
methyl-2-buten-l-o1), N-chlorosuccinamide, methyl sulfide, di- methylphosphonate, and n-butyl lithium were purchased from Ald- rich. All other chemicals were of analytical grade.
Shipping Latex-Freshly tapped latex from uncultivated H. brasi- liensis in Costa Rica was immediately diluted with 300 ml of 300 mM Tris-HCl, pH 7.5, 0.03% sodium azide/7OO ml latex, quickly frozen in 1-liter bottles, and shipped to this laboratory on dry ice. Dilution with buffered azide attenuates the loss of latex due to coagulation during the tapping, freezing, shipping, and thawing procedures. In general, latex was collected from trees which had never been tapped or had not been tapped for several months. However, in some cases latex samples were collected from trees tapped every 3 days for at least 1 month. Reliable methods of latex collection and shipment were the result of the efforts of Charles Hunter, Granjas Tropicales S. A., San Jose, Costa Rica. In preliminary studies small volumes of latex (5-10 ml) were shipped overnight on wet ice by Dr. Paul K. Soderholm, Subtropical Horticultural Research Station, Miami, FL.
Quantitatiue Estimation of Rubber Content-For routine compari- sons of samples containing rubber particles (whole latex and depro- teinated rubber particles), we measure the absorbance at 280 nm of a dilute (1:lOOO) sample. The absorbance is dominated by the light scattering of the rubber particles in the sample and even in whole latex of relatively low rubber content 4 % of the light is absorbed by protein. We determined the rubber content of whole latex and depro- teinated rubber particles in order to convert the A280 to mg/ml of rubber. Samples were filtered and washed as in the standard assay (see below) and dried in a vacuum oven. The weight was determined by weighing the filter before and after filtration of the rubber. Control filters were carried through to correct for changes in the weight of the filter (-5% loss). Samples were filtered in the presence and absence of 1% SDS to remove protein but this precaution did not affect the weight of the rubber deposited on the filter. For whole latex the conversion ratio was determined to be 0.06 * 0.002 mg of rubber/ Azso which compares favorably with the ratio for deproteinated rubber particles of 0.04 f 0.002 mg of rubber/Am.
Elemental Analysis-Deproteinated rubber particles purified on the Sephacryl S-300 column containing -17 mg of total rubber (1.7 ml of 200 Asm) were dried under vacuum and submitted for elemental analysis (Dr. Tashinian, Chemical Analytical Services, University of California, Berkeley).
Affinity Column Synthesis-A nerylmethyl phosphonophosphate affinity column was synthesized by the same procedure described for the synthesis of an analogous geranylmethyl phosphonophosphate column described by Bartlett et at. (38). The synthesis of 046-amino- 1-hexyl)-P-nerylmethyl phosphonophosphate from tetra-n-butylam- monium monohydrogen nerylmethyl phosphonate and N-trifluor- oacetyl-6-amino-1-hexyl phosphate was performed by Dr. Todd Cap-
son and Dr. C. Dale Poulter, University of Utah. Comparison of elemental analysis (Dr. Tashinian, Chemical Analytical Services, University of California Berkeley) of the derivatized and underiva- tized Sepharose 6B for phosphorus (0.099% uersw <O.OOl%) allows calculation of the ligand concentration in the column (1 mM) and the coupling efficiency (5%).
Protein Determination-The Bio-Rad Protein Assay (Bradford assay (48), Bio-Rad Laboratories) with lipid-free bovine serum albu- min (Sigma) as a standard was used for all protein determinations. With the purified prenyltransferase, this method gave very good agreement with amino acid analysis after hydrolysis ( l l O T , 24 h, 6 N HCl) using norleucine as a standard performed with the assistance of William Henzel, Genentech.
SDS-Polyacrylamide Gels-SDS-polyacrylamide gels (10 and 15%) were run by standard methods and stained with Coomassie Blue (Bio- Rad) or silver stained by published methods (49).
Thin Layer Chromatography-Separation of cis- or trans-polypren- 01s between C5 and C I ~ ~ essentially according to molecular weight was achieved on glass-backed Cl&lica reverse phase plates (5 X 20 cm; Baker Chemical) using low percentage HzO:acetone (mineral oil) solvent systems as described by Mankowski et al. (40). Gas Chromatography/Mass Spectrometry-The separation of all
isomers of Cl0 and C16 polyprenols was achieved by gas chromatog- raphy (J&W DB5,30 m X 0.25-mm capillary column, 185 'C, Perkin- Elmer, Sigma 115) and the following order of elution observed nerol, geraniol, c,c-farnesol, c,t-farnesol, and tpfarnesol (poorly resolved), and t,t-farnesol (Dr. Randy Marcinko, Dynamic Services, Menlo Park).' Samples of enzymatically prepared GPP and t,t-FPP were converted to the appropriate alcohol by addition of 1.2 mg of calf alkaline phosphatase to 0.75 ml of the standard FPP assay mixture after incubating with purified prenyltransferase. After incubating an additional 15 min at 37 "C, the alcohols were extracted with 3 X 1-ml aliquots of petroleum ether (Baker Chemical). The combined ether extracts were dried to 50 ml and analyzed. Verification of t,t-farnesol was performed by GC/MS (Hewlett-Packard 5985) with reference to a library spectrum with the assistance of Dr. John Chakel and Dr. Robert Lazarus, Genentech, Inc.
Gel Permeation Chromatography-Separation of soluble rubber from long chain polyprenols up to c105 (1445 Da) was achieved on a 500 8, p-Styragel (Waters Associates, Inc.) with an exclusion limit of 20,000 Da (tetrahydrofuran or toluene, 1 ml/min, 25"C, refractive index detector, Waters Associates, Inc.). In some cases, a t higher sample loads, precipitation of rubber on the column was encountered, and this could be avoided by running the column at 100°C. Samples for injection were prepared from rubber trapped on 0.22-p filters. A sample containing 5.5 mg/ml deproteinated rubber particles, 2.2 mg/ ml purified prenyltransferase (57 nM), 0.3 mM [1-14c]IPP (0.21 ci / mol), 1 mM MgS04, 10 mM DTT, and 50 mM Tris, pH 7.5, in a final volume of 10 ml was incubated 180 min and filtered in 0.5-ml aliquots. Filters were washed with 10% HCl and ethanol, and air-dried as in the standard assay described above. Two of the above filters and two control filters prepared without prenyltransferase were dissolved in scintillant. Four of the remaining filters were extracted overnight and rinsed twice with benzene. After the pooled benzene extract (2.5 ml) was extracted with 0.1 ml of 6 N NaOH, the rubber was precipitated from the benzene by the addition of 2.5 ml of ethanol. The precipitated rubber was collected by centrifugation. Two of the samples of ex- tracted rubber were redissolved in benzene for scintillation counting along with the benzene/ethanol supernatant and the NaOH extract. The remaining rubber samples were dissolved in 10% (v/v) farnesol in toluene and injected on the p-Styragel column.
Curve Fitting-The fit of data by computer to equations in the text was achieved by an iterative approach in which parameters are adjusted by the Marquardt-Levenberg method until a least squares solution is reached. All fits were calculated through the use of RS/1 (BBN Research Systems, Bolt Beranek and Newman Inc.) run on a Vax 11780 with a UNIX operating system (4.2 BSD version, Bell Labs).
Amino Acid Analysis-Following carboxymethylation in urea, prenyltransferase was hydrolyzed for 24 h with constant boiling HC1 in evacuated sealed tubes at 110°C. The dried hydrolysates were analyzed on a Beckman 6300 amino acid analyzer equipped with a ninhidrin detector using an automated program. The concentration of tryptophan was not determined.
Isolation of Particles-Deproteinated rubber particles with low intrinsic prenyltransferase activity were prepared by a modification
R. Marcinko, personal communication.

cis-Polyisoprene Rubber Prenyltransferase 18591
ofthe procedure described by Archer e t al. (1,33,34). A 1-liter bottle of diluted whole latex was thawed and centrifuged at 2000 X g for 10 min to remove coagulated rubber (floating pellet), dirt, and intact organelles (bottom fraction). The remaining whole latex was decanted and frozen in 10-ml portions. Fresh deproteinated rubber particles were prepared from 10 ml of whole latex by size exclusion chroma- tography on a Sepharose S-300 column (5 x 50 cm) equilibrated in 50 mM Tris-HC1, pH 7.5, and run at 0.64 ml/min. The latex particles were eluted in the void volume while the serum proteins were retained in the column. Particles from a typical column had an absorbance at 280 nm of 300. These particles were used in the rubber transferase assay and could be stored frozen at -80 "C for several weeks without appreciable coagulation or loss of function. The Sepharose S-300 column was repacked after each use to remove rubber that became trapped in the column. This rubber would float away from the gel if the gel was allowed to settle several times in 5-10 times the volume of buffer after brief stirring.
Rubber E b n g a t w n Assay-Rubber transferase was assayed by measuring its ability to add radiolabeled isopentenyl pyrophosphate to existing deproteinated latex particles. A 0.5-ml incubation mixture contained 0.01-0.45 ml of deproteinated rubber particles or whole latex, 50 mM Tris-HC1, 1 mM MgSO,, 10 mM DTT, and 0.5 mM [1-'4C]IPP (specific activity = 0.4 Ci/mol) and added prenyltransfer- ase if appropriate. The reaction was incubated at 25 "C for 1-4 h and then terminated by the addition of 50 pl of 0.25 M EDTA, pH 8.0. Each sample was then filtered with an Amicon vacuum filtration manifold and 0.22-p cellulose acetate/cellulose nitrate filters (Milli- pore GSWP 025 00) which provided sufficient flow but still retained the latex particles. The moist cake of latex particles on the filter was coagulated under a heat lamp and washed for 2-15 min in 1 M HC1 followed by two 5-15 min washes with ethanol. Radioactivity on the filters was determined in a toluene-based scintillation mixture (Quan- tiflor, Mallinkrodt). If the filters remained in scintillant overnight the rubber dissolved, the filters themselves became transparent, the 14C counts increased, and the channel ratio observed reached that measured for a I4C standard. The efficiency of 14C counting was 0.7 cpm/dpm. The original intent of the acid and ethanol washes was to protonate and remove, respectively, adventitious ["CIIPP. However, we note that Archer and Audley (2) recently reported that in contrast to pretreatment with base or protease, an acid pretreatment of newly synthesized rubber dramatically increased the solubility of the final product.
DMAPP, 0.5 mM ['4C]IPP (0.4 Ci/mol), 1 mM MgS04, 10 mM DTT, FPP Synthase Assay-A typical assay mixture contained 0.5 mM
50 mM Tris, pH 7.5, and prenyltransferase in 0.5 ml and was incubated 15-60 min at 30 "C. Two methods for the separation of [14C]IPP from ["CIFPP were employed. The first was standard (7, 10) and was based upon the selective hydrolysis of the allylic pyrophosphate product in acid (1 M HCl, 75 "C, 30 min) to products that may be extracted from [14C]IPP in hexane after increasing the pH (2 M NaOH). The radioactivity in a portion of the hexane extract was determined.
The second method was rapid and was based on a method used to extract differentially long chain allylic pyrophosphates from ['4C]IPP (18). At neutral pH, IPP was not partitioned into the 1-butanol phase (18), whereas FPP was partitioned nearly equally in the aqueous and the organic phases. Thus, the reaction was quenched by adding 0.05 ml of 0.25 M EDTA, H20 was added to 1 ml, the mixture was extracted with 2 ml of I-butanol (the butanol phase expands to 2.5 ml owing to Hz0 uptake), 1.5 ml of the butanol phase was back-extracted with 0.5 ml of H20, and 1.0 ml of the butanol phase was counted. The overall extraction yield was 25% with this method. It was easier to use with samples that had a high concentration of protein since these samples form an intractable emulsion when extracted with hexane. The efficiency of "C counting was 0.7 cpm/dpm.
Prenyltransferase Purification-The rubber fraction of whole latex was removed by centrifugation to obtain C-serum (24). For a typical purification, the contents of 11 1-liter bottles of frozen whole latex were cut into chunks and quickly thawed in a container warmed by a waterbath at room temperature. Upon thawing, 2-mercaptoethanol was added to 0.5%. The latex was centrifuged at 13,680 x g (GS3, 9000 rpm) for 2 h causing most of the rubber to coagulate at the top of the centrifuge bottle. The supernatant fluid was then centrifuged at 27,500 X g (GSA, 13,000 rpm) for at least 2.5 h or overnight to remove the remaining rubber. At this point we routinely inspected the C-serum for turbidity, since the presence of rubber adversely effected the ammonium sulfate fractionation that followed. If neces- sary, a third centrifugation step was included to remove latex particles
which could be inadvertently resuspended during decanting. The protein fraction precipitating between 25-65% of saturation with ammonium sulfate (pH 7.6 with Tris base) from the clear C-serum was resuspended in 0.5 liter of 50 mM potassium phosphate, pH 8.0, 1 mM MgSO,, and 0.25% 2-mercaptoethanol and dialyzed at 4°C against 2 X 40 liters of the same buffer containing 0.5% sodium azide. A considerable amount of material could not be resuspended after dialysis and was removed by centrifugation at 27,500 X g (GSA, Sorvall; 13,000 rpm) for 1 h. Although not characterized, inspection of the physical properties of this insoluble material suggested that it contained a large amount of rubber. This observation indicates that even clear C-serum contains rubber particles that are very small and do not cause turbidity. Archer et al. (33) has reported the presence of particles smaller than 100 A in serum. Additional clarification and a partial purification resulted by passing the sample through a DE52 column (5 X 20 cm) in 50 mM potassium phosphate, pH 8.0, 1 mM MgS04, 0.10% dithiothreitol, and 0.01% sodium azide (3 ml/min). The column was washed with the same buffer until no more protein was eluted and then discarded. Active fractions of the unbound protein were concentrated to 50 ml with an Amicon hollow fiber cartridge (HlP10-8) and loaded on a Sepharose S-200 column (5 X 90 cm) equilibrated in 50 mM Tris-HC1, pH 7.5, 1 mM MgSOr,O.IO% dithiothreitol, and 0.10% sodium azide (0.7 ml/min). A single broad band of rubber transferase activity was eluted after the void volume. The broadness of this band resulted from the nonlinearity of the assay with respect to enzyme concentration and from aggregation of a small percentage of the enzyme that could be detected in the void volume by SDS-PAGE (data not shown).
To make purification by hexyl agarose chromatography most effi- cient, isocratic conditions were chosen in which rubber transferase did not bind to the column material, but was slightly retarded. The active pool from the Sepharose S-200 column was concentrated via Amicon filtration (YM30) to 10 ml and dialyzed against 2 liters of 50 mM Tris-HC1, pH 7.5,40 mM NaC1,l mM MgSO,, 0.1% dithiothreitol, and 0.1% sodium azide, and loaded on the hexyl agarose column (2.5 X 100 cm; 0.9 ml/min) equilibrated in the same buffer. The first protein band containing a majority of the protein did not interact with the hexyl agarose column; this band was followed by several smaller and broader protein bands. A single band of rubber transfer- ase activity was observed just after the main band of protein. This activity was pooled, concentrated to 9 ml via Amicon filtration (YM30), and dialyzed against 1 liter of 10 mM Tris-HC1, pH 8.0, 1 mM MgSO,, 0.1% dithiothreitol, and 0.01% sodium azide. This prep- aration was divided in half and each half chromatographed separately on a Pharmacia Mono Q column equilibrated in 25 mM Tris-HC1, pH 8.0, 1 mM MgSO,, 0.1% sodium azide. After eluting unabsorbed protein with the same buffer, the prenyltransferase was eluted with a linear salt gradient from 0 to 125 mM NaCl (Fig. 2B). Dithiothreitol (10 mM) which interfered with the UV detection was added to each fraction immediately after elution. Rubber transferase could be stored without detectable loss of activity for at least 6 months at -80 "C in 25 mM Tris-HC1, pH 8, 10 mM dithiothreitol, 1 mM MgSO,, 30% glycerol, and 0.1% sodium azide.
RESULTS
Characterization of Whole Latex-Much of the published data on rubber transferase was obtained using freshly tapped latex in a laboratory with an adjoining greenhouse (5), so it was useful to compare the properties of our frozen and shipped material with those reported for the freshly tapped material. Archer et al. (35) characterized the composition of freshly tapped H, brasiliensis latex and found that it is 33% (w/v) rubber and that serum is 0.5% (w/v) protein. We monitored these two values over the course of many different shipments and prenyltransferase purifications.
Rubber content was calculated from the measured absorb- ance at 280 nm and converted to milligrams of rubber per ml with the factor 0.06 mg/A,,. Some latex samples used contain as little as 4% rubber owing to extensive coagulation, but typical contents vary between 10 and 30% (36). A small sample of uncoagulated latex rushed overnight on wet ice from Florida had an A,, of 5500 or 33% rubber in agreement with Archer et al. (35). Two frozen and thawed samples from Costa Rica had values as high as 5100 (31%) and 4100
18592 cis-Polyisoprene Rubber Prenyltransferase
(25%), as measured after correcting for dilution by the buffer added after tapping.
In contrast, much less variation was observed in the protein content of the C-serum fraction derived from whole latex by ultracentrifugation. Regardless of the rubber content, the protein content of serum was constant (0.47 -+ 0.13%, n = ll), again in agreement with Archer et al. (35). Thus, the effect of coagulation was to reduce the rubber content of whole latex but not the protein content in C-serum.
Archer and Cockbain (1) reported that rubber transferase is a soluble protein in the C-serum. The data in Table I confirm this and demonstrate that all of the prenyltransferase remained in the C-serum after the rubber fraction was re- moved by ultracentrifugation. We used the FPP synthetase activity of the prenyltransferase to monitor its concentration, since it can be measured in the presence of the rubber particles in whole latex. There was slightly higher prenyltransferase activity in C-serum than in an equivalent volume of whole latex as shown in Table I. This discrepancy could be ac- counted for by correcting for the volume of C-serum in whole latex.
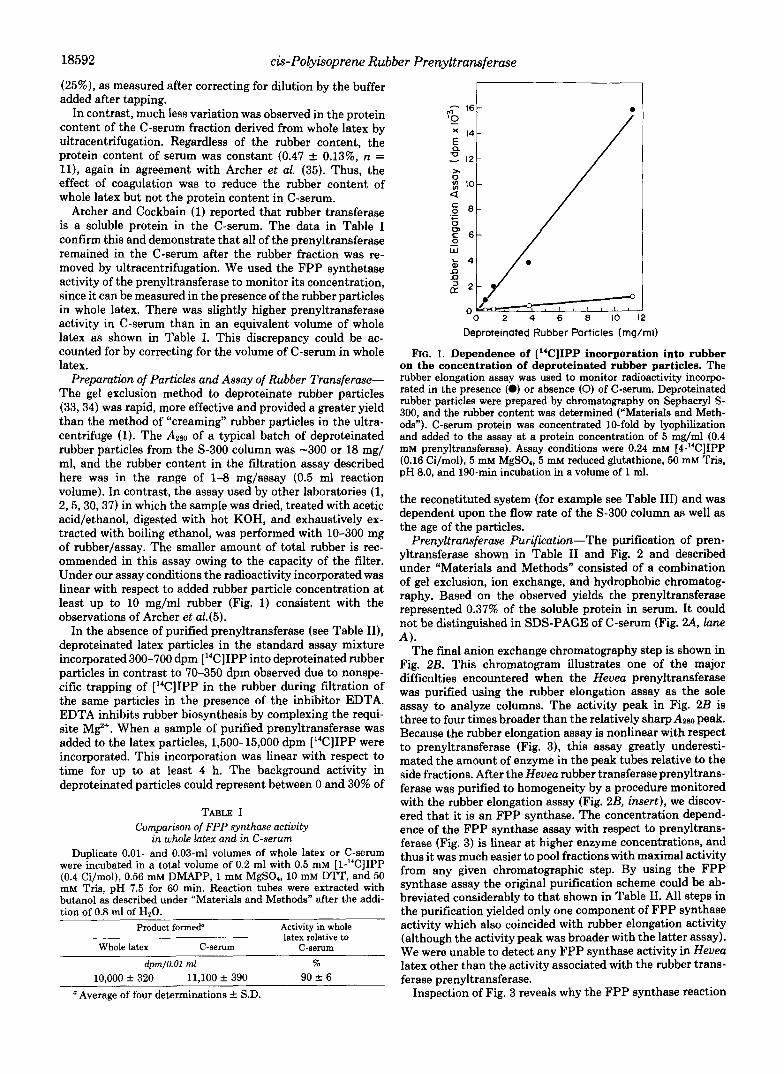
Preparation of Particles and Assay of Rubber Transferase- The gel exclusion method to deproteinate rubber particles (33,34) was rapid, more effective and provided a greater yield than the method of “creaming” rubber particles in the ultra- centrifuge (1). The Azm of a typical batch of deproteinated rubber particles from the S-300 column was -300 or 18 mg/ ml, and the rubber content in the filtration assay described here was in the range of 1-8 mg/assay (0.5 ml reaction volume). In contrast, the assay used by other laboratories (1, 2,5,30,37) in which the sample was dried, treated with acetic acid/ethanol, digested with hot KOH, and exhaustively ex- tracted with boiling ethanol, was performed with 10-300 mg of rubber/assay. The smaller amount of total rubber is rec- ommended in this assay owing to the capacity of the filter. Under our assay conditions the radioactivity incorporated was linear with respect to added rubber particle concentration at least up to 10 mg/ml rubber (Fig. 1) consistent with the observations of Archer et aL(5).
In the absence of purified prenyltransferase (see Table 11), deproteinated latex particles in the standard assay mixture incorporated 300-700 dpm [14C]IPP into deproteinated rubber particles in contrast to 70-350 dpm observed due to nonspe- cific trapping of [14C]IPP in the rubber during filtration of the same particles in the presence of the inhibitor EDTA. EDTA inhibits rubber biosynthesis by complexing the requi- site M F . When a sample of purified prenyltransferase was added to the latex particles, 1,500-15,000 dpm [14C]IPP were incorporated. This incorporation was linear with respect to time for up to at least 4 h. The background activity in deproteinated particles could represent between 0 and 30% of
TABLE I Comparison of FPP synthase activity
in whole latex and in C-serum Duplicate 0.01- and 0.03-ml volumes of whole latex or C-serum
were incubated in a total volume of 0.2 ml with 0.5 mM [1-’4c]IPP (0.4 Ci/mol), 0.56 mM DMAPP, 1 mM MgSOI, 10 mM DTT, and 50 mM Tris, pH 7.5 for 60 min. Reaction tubes were extracted with butanol as described under “Materials and Methods” after the addi- tion of 0.8 ml of HzO.
Product formed” Activity in whole latex relative to
Whole latex C-serum C-serum
dpm/O.Ol ml % 10,000 * 320 11,100 & 390 90 2 6
a Average of four determinations f S.D.
7 0
x 14- E Q 73 - 12-
Deproteinoted Rubber Particles (mg/rnl)
FIG. 1. Dependence of [“CIIPP incorporation into rubber on the concentration of deproteinated rubber particles. The rubber elongation assay was used to monitor radioactivity incorpo- rated in the presence (0) or absence (0) of C-serum. Deproteinated rubber particles were prepared by chromatography on Sephacryl S- 300, and the rubber content was determined (”Materials and Meth- ods”). C-serum protein was concentrated 10-fold by lyophilization and added to the assay at a protein concentration of 5 mg/ml (0.4 mM prenyltransferase). Assay conditions were 0.24 mM [4-“C]IPP (0.16 Ci/mol), 5 mM MgSO., 5 mM reduced glutathione, 50 mM Tris, pH 8.0, and 190-min incubation in a volume of 1 ml.
the reconstituted system (for example see Table 111) and was dependent upon the flow rate of the S-300 column as well as the age of the particles.
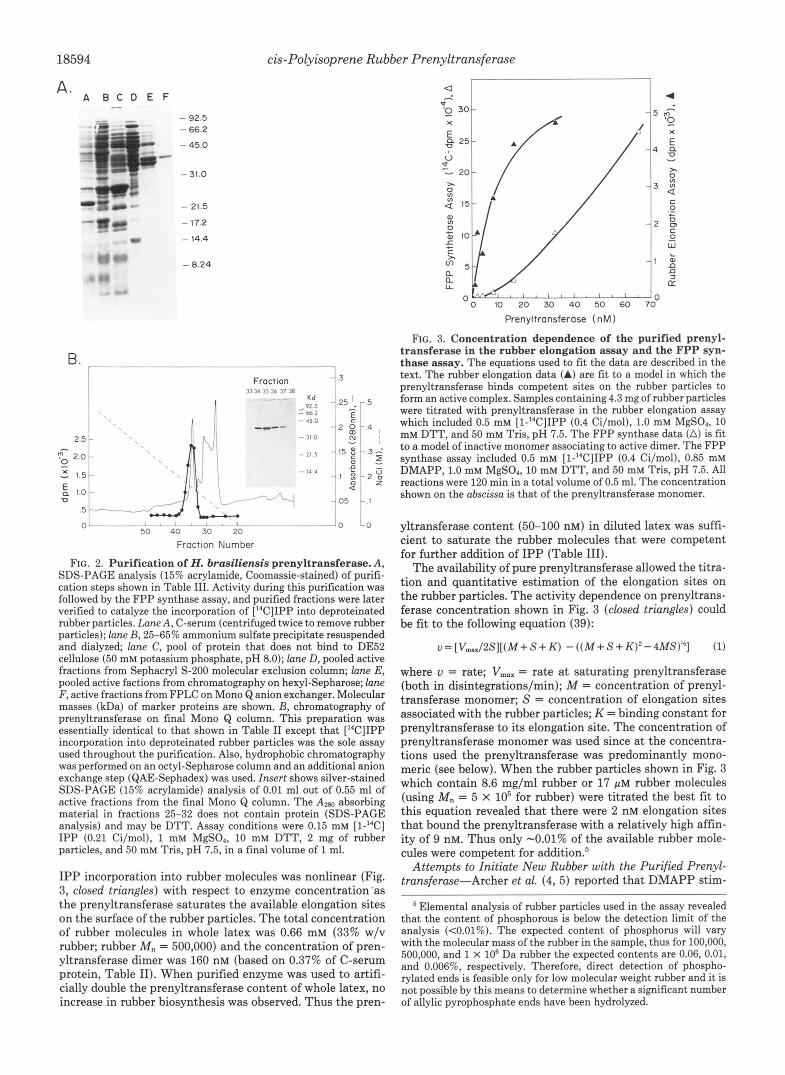
Prenyltransferase Purification-The purification of pren- yltransferase shown in Table I1 and Fig. 2 and described under “Materials and Methods” consisted of a combination of gel exclusion, ion exchange, and hydrophobic chromatog- raphy. Based on the observed yields the prenyltransferase represented 0.37% of the soluble protein in serum. It could not be distinguished in SDS-PAGE of C-serum (Fig. 2 A , lane A).
The final anion exchange chromatography step is shown in Fig. 2B. This chromatogram illustrates one of the major difficulties encountered when the Heuea prenyltransferase was purified using the rubber elongation assay as the sole assay to analyze columns. The activity peak in Fig. 2B is three to four times broader than the relatively sharp Azw peak. Because the rubber elongation assay is nonlinear with respect to prenyltransferase (Fig. 3), this assay greatly underesti- mated the amount of enzyme in the peak tubes relative to the side fractions. After the Heuea rubber transferase prenyltrans- ferase was purified to homogeneity by a procedure monitored with the rubber elongation assay (Fig. ZB, insert), we discov- ered that it is an FPP synthase. The concentration depend- ence of the FPP synthase assay with respect to prenyltrans- ferase (Fig. 3) is linear at higher enzyme concentrations, and thus it was much easier to pool fractions with maximal activity from any given chromatographic step. By using the FPP synthase assay the original purification scheme could be ab- breviated considerably to that shown in Table 11. All steps in the purification yielded only one component of FPP synthase activity which also coincided with rubber elongation activity (although the activity peak was broader with the latter assay). We were unable to detect any FPP synthase activity in Heuea latex other than the activity associated with the rubber trans- ferase prenyltransferase.
Inspection of Fig. 3 reveals why the FPP synthase reaction

cis-Polyisoprene Rubber Prenyltransferase 18593 TABLE I1
Purification of H. brasiliensis FPP synthase (rubber transferase)
step Protein Total activity Specific activity Purification
rng (9%) units" (%) unitsfmg -fold C-serum (spun twice*) 7,910 (100) 25-65% ammonium sulfate precipitation 6,340 (80) 31,700 (100) 5.0 1
DE52 cellulose flow-through pool 1,350 (17) [ 9001 e
Sephacryl S-200 peak 400 (5.0) 34,700 (110) 87 17.4 Hexyl-Sepharose peak 21.3 (0.27) 14,000 (44) 657 131 Mono Q FPLC anion exchange
Peak 3.1 (0.039) 4,000 (13) 1,290 258 Sides 2.2 (0.028) 1,630 (5) 740 150 Total 5.3 (0.067) 4,530 (18)
(dialyzed and spun)
' Nanomoles of ['*CC]IPP incorporated per min, obtained using the FPP synthase assay with analysis by acid
'Diluted whole latex (11 liters) was centrifuged 2 h at 13,700 X g, and the turbid supernatant fluid was
'The total activity recovered in this step is consistently low, but full activity is recovered after the pooled
hydrolysis of allylic pyrophosphate products (1 M HCI, 75 "C, 30 min) and extraction of products into hexane.
centrifuged 15 h at 27,500 X g to obtain 6.1 liters of clarified C-serum.
fractions are chromatographed on SephacryiS-200.
was the more useful of the two for rapidly monitoring columns during the purification of Heuea prenyltransferase. The FPP synthase assay, when diluted to 10 nM prenyltransferase or less, resulted in the underestimation of the activity in the side fractions from a column, but the peak fractions coincided exactly with the peak of enzyme. However, if the rubber elongation assay was used, all fractions greater than 15-20 nM prenyltransferase showed nearly the same response in the assay. In practice this was the most common problem; peaks with the latter assay tended to be broad and flat making it difficult to pool the collected fractions.
This prenyltransferase did not bind to a nerylmethylphos- phophosphonate affinity column, the cis analog of the geran- ylmethylphosphophosphonate affinity column successfully used to purify FPP synthase from avian and porcine liver (38). It was even more surprising that the prenyltransferase also did not bind to the latter column3 since the purified protein was found to have FPP synthase activity. In both cases over 90% of the activity flowed through and did not bind to either the cis or trans analog containing column. No additional activity was eluted at up to 50 mM pyrophosphate. Thus affinity chromatography of this type has not proven to be a viable approach to purification.
characterization of Prenyltransferase-Prenyltransferase from the Mono Q column was at least 95% pure by SDS gel electrophoresis after silver staining which revealed a single band at 38,000 daltons (Fig. 2B, insert). Characterization of the purified native material on a standardized TSK-G3000 SW column in 0.2 M sodium phosphate, pH 6.5, gave a native molecular mass of 70,000 daltons. Thus, rubber transferase is a dimer of identical monomeric subunits. No sequence greater than 0.5% (5 pmol) was detected when 40 pg (1 nmol) of purified rubber transferase was subjected to NHz-terminal sequence determination. The protein was washed from the analytical filter, digested with CNBr, and subjected to se- quence analysis again. A mixture of four to five sequences at the 30-124 pmol level were detected. Thus, the NHz terminus of the enzyme is blocked, and contaminating proteins are either present in low amounts or are also blocked. The amino acid composition of the purified prenyltransferase is compared to other purified prenyltransferases in Table IV4.
D. L. Bartlett and C. D. Poulter, personal communication. 'We have used HPLC to purify 19 tryptic fragments and several
CNBr fragments from the reduced, carboxymethylated purified pren- yltransferase and have obtained amino acid sequence data from eight tryptic fragments (1, QWVER 2, VLYNXLK; 3, LQGVFTEYENE-
A high level of DTT (10 mM) was required for maximal incorporation of ['4C]IPP into rubber; 2-mercaptoethanol was not as effective at equal concentrations. Free small molecular weight thiols were not required for catalysis in the rubber elongation assay, however, since the same stimulation could be achieved by reducing the prenyltransferase with DTT followed by dialysis to remove the mercaptan before the prenyltransferase was added to assay (data not shown). In addition rubber transferase activity of the purified prenyl- transferase was completely inhibited by thiol alkylating agents such as iodoacetic acid confirming the observation of Archer and Cockbain (1) on the partially pure protein. The divalent cations Zn2+ and Co2+ (0.1-1 mM) completely inhib- ited rubber elongation activity, while both M e and Mn2+ were stimulatory and were maximal at 1 mM; Ca2+ and Pb2+ were nearly as effective at 5 mM. Sodium pyrophosphate and sodium phosphate were also found to be inhibitors of rubber transferase activity with apparent K, values of 5 and 200 mM, respectively (0.3 mM IPP).
Percent Reconstitution of Whole Latex Actiuity-When samples of whole latex and deproteinated rubber particles plus prenyltransferase were matched for rubber content it was possible to reconstitute 40-60% of the activity of whole latex by using the purified system. The addition of purified prenyltransferase to diluted whole latex did not increase its rubber biosynthetic activity even when the final concentration of prenyltransferase was two to four times higher than the endogenous level of the enzyme (Table 111). Thus, the pren- yltransferase concentration in whole latex was saturating even though the molar ratio of rubber molecules to prenyltransfer- ase was 140 in the experiment in Table 111. Saturation was observed in samples in which this ratio was as high as 1500. The lack of 100% reconstitution and the lack of stimulation of whole latex by added prenyltransferase could be due to a slow rate of formation of an active prenyltransferase complex with its rubber substrate. However, preincubation of the pren- yltransferase with rubber particles for up to 5 h did not result in an increase (or decrease) in the measured rate indicating that complex formation is rapid.
Titration of Active Ends-As observed with partially pure preparations from both Heuea and guayule (1, 3, 4, 30), [l4CC]
SYK; 4, GQPCWFR 5, QELLEDPAFEWTPDFA . . . ; 6, VGLIA- ANDGILLR 7, XXQELNEEEIFLADALG . . . ; 8, AYYVDLLDLF- NEVEFQXAXGQVIDLI . . .) and one 25-kDa CNBr fragment (XXLITTLEGEFDLXFYXLXL . . .). None have homology with an active site tryptic fragment from avian liver FPP synthetase (7).
18594 cis-Polyisoprene Rubber Prenyltransferase
A. A B C D E F
L
- 92.5 - 66.2 - - 45.0
- - -jr - - 31.0
" Ikm U
5-l *.>
"1- --". - 21.5
- 17.2
- 14.4
- 8.24
B. P ~~ ~- = -
I
" .
Fraction -y3
Fraction Number
FIG. 2. Purification of H. brasiliensis prenyltransferase. A, SDS-PAGE analysis (15% acrylamide, Coomassie-stained) of purifi- cation steps shown in Table 111. Activity during this purification was followed by the FPP synthase assay, and purified fractions were later verified to catalyze the incorporation of ["CIIPP into deproteinated rubber particles. Lane A, C-serum (centrifuged twice to remove rubber particles); lane B, 25-65% ammonium sulfate precipitate resuspended and dialyzed; lane C, pool of protein that does not bind to DE52 cellulose (50 mM potassium phosphate, pH 8.0); lane D, pooled active fractions from Sephacryl S-200 molecular exclusion column; lane E, pooled active factions from chromatography on hexyl-Sepharose; lane F, active fractions from FPLC on Mono Q anion exchanger. Molecular masses (kDa) of marker proteins are shown. B, chromatography of prenyltransferase on final Mono Q column. This preparation was essentially identical to that shown in Table I1 except that ['4C]IPP incorporation into deproteinated rubber particles was the sole assay used throughout the purification. Also, hydrophobic chromatography was performed on an octyl-Sepharose column and an additional anion exchange step (QAE-Sephadex) was used. Insert shows silver-stained SDS-PAGE (15% acrylamide) analysis of 0.01 ml out of 0.55 ml of active fractions from the final Mono Q column. The AZW absorbing material in fractions 25-32 does not contain protein (SDS-PAGE analysis) and may be DTT. Assay conditions were 0.15 mM [l-"C] IPP (0.21 Ci/mol), 1 mM MgS04, 10 mM DTT, 2 mg of rubber particles, and 50 mM Tris, pH 7.5, in a final volume of 1 ml.
IPP incorporation into rubber molecules was nonlinear (Fig. 3, closed triangles) with respect to enzyme concentration'as the prenyltransferase saturates the available elongation sites on the surface of the rubber particles. The total concentration of rubber molecules in whole latex was 0.66 mM (33% w/v rubber; rubber M , = 500,000) and the concentration of pren- yltransferase dimer was 160 nM (based on 0.37% of C-serum protein, Table 11). When purified enzyme was used to artifi- cially double the prenyltransferase content of whole latex, no increase in rubber biosynthesis was observed. Thus the pren-
Prenyltransferose (nM)
FIG. 3. Concentration dependence of the purified prenyl- transferase in the rubber elongation assay and the FPP syn- thase assay. The equations used to fit the data are described in the text. The rubber elongation data (A) are fit to a model in which the prenyltransferase binds competent sites on the rubber particles to form an active complex. Samples containing 4.3 mg of rubber particles were titrated with prenyltransferase in the rubber elongation assay which included 0.5 mM [1-'4C]IPP (0.4 Ci/mol), 1.0 mM MgSO,, 10 mM DTT, and 50 mM Tris, pH 7.5. The FPP synthase data (A) is fit to a model of inactive monomer associating to active dimer. The FPP synthase assay included 0.5 mM [1-'4C]IPP (0.4 Ci/mol), 0.85 mM DMAPP, 1.0 mM MgSO,, 10 mM DTT, and 50 mM Tris, pH 7.5. All reactions were 120 min in a total volume of 0.5 ml. The concentration shown on the abscissa is that of the prenyltransferase monomer.
yltransferase content (50-100 nM) in diluted latex was suffi- cient to saturate the rubber molecules that were competent for further addition of IPP (Table 111).
The availability of pure prenyltransferase allowed the titra- tion and quantitative estimation of the elongation sites on the rubber particles. The activity dependence on prenyltrans- ferase concentration shown in Fig. 3 (closed triangles) could be fit to the following equation (39):
u=[Vm,/2S][(M+S+K) -((M+S+K)2-4MS)Lh] (1)
where u = rate; Vmax = rate at saturating prenyltransferase (both in disintegrations/min); M = concentration of prenyl- transferase monomer; S = concentration of elongation sites associated with the rubber particles; K = binding constant for prenyltransferase to its elongation site. The concentration of prenyltransferase monomer was used since at the concentra- tions used the prenyltransferase was predominantly mono- meric (see below). When the rubber particles shown in Fig. 3 which contain 8.6 mg/ml rubber or 17 p~ rubber molecules (using M, = 5 x lo5 for rubber) were titrated the best fit to this equation revealed that there were 2 nM elongation sites that bound the prenyltransferase with a relatively high affin- ity of 9 nM. Thus only -0.01% of the available rubber mole- cules were competent for addition?
Attempts to Initiate New Rubber with the Purified Prenyl- transferase-Archer et u1. (4, 5) reported that DMAPP stim-
Elemental analysis of rubber particles used in the assay revealed that the content of phosphorous is below the detection limit of the analysis (<O.Ol%). The expected content of phosphorus will vary with the molecular mass of the rubber in the sample, thus for 100,000, 500,000, and 1 X lo6 Da rubber the expected contents are 0.06, 0.01, and 0.006%, respectively. Therefore, direct detection of phospho- rylated ends is feasible only for low molecular weight rubber and it is not possible by this means to determine whether a significant number of allylic pyrophosphate ends have been hydrolyzed.
cis-Polyisoprene Rubber Prenyltransferase 18595
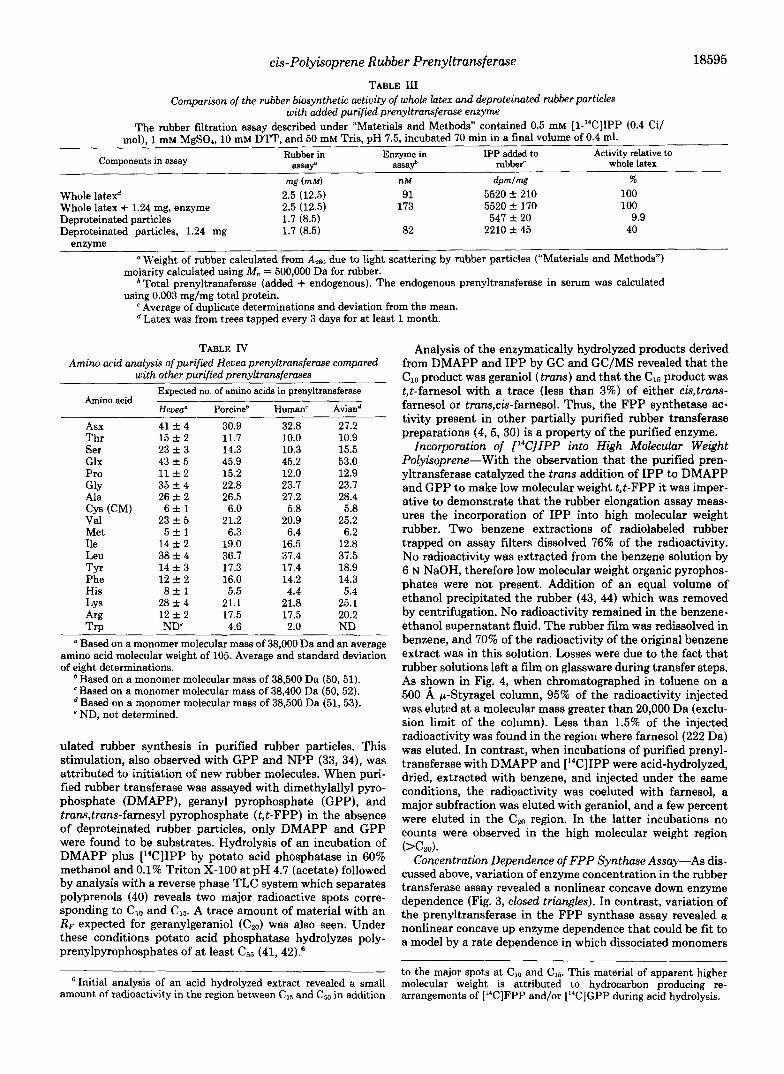
TABLE I11 Comparison of the rubber biosynthetic activity of whole latex and deproteinated rubber particles
with added purified prenyltransferase enzyme The rubber filtration assay described under “Materials and Methods” contained 0.5 mM [l-“C]IPP (0.4 Ci/
mol), 1 mM MgS04, 10 mM DTT, and 50 mM Tris, pH 7.5, incubated 70 min in a final volume of 0.4 ml.
Components in assay Rubber in Enzyme in IPP added to Activity relative to assaf assay“ rubber‘ whole latex
rng (mM) nM d p m l w % Whole latexd 2.5 (12.5) 91 5520 * 210 100 Whole latex + 1.24 mg, enzyme 2.5 (12.5) 173 5520 f 170 100 Deproteinated particles 1.7 (8.5) 547 f 20 9.9 Deproteinated particles, 1.24 mg 1.7 (8.5) 82 2210 f 45 40
enzyme a Weight of rubber calculated from A2m due to light scattering by rubber particles (“Materials and Methods”)
’ Total prenyltransferase (added + endogenous). The endogenous prenyltransferase in serum was calculated
e Average of duplicate determinations and deviation from the mean.
molarity calculated using M, = 500,000 Da for rubber.
using 0.003 mg/mg total protein.
Latex was from trees tapped every 3 days for at least 1 month.
TABLE JV Amino acid analysis of purified Hevea prenyltransferase compared
with other purified prenyltransferases
Amino acid
Asx 41 f 4 30.9 32.8 27.2 Thr 15 f 2 11.7 10.0 10.9 Ser 23 f 3 14.3 10.3 15.5 Glx 43 * 5 45.9 45.2 53.0 Pro 11 f 2 15.2 12.0 12.9 GlY 35 f 4 22.8 23.7 23.7 Ala 26 & 2 26.5 27.2 28.4 Cys (CM) 6 f 1 6.0 5.8 5.8 Val 23 f 5 21.2 20.9 25.2 Met 5 f 1 6.3 6.4 6.2 Ile 14 f 2 19.0 16.5 12.8 Leu 38 f 4 36.7 37.4 37.5 TYr 14 f 3 17.3 17.4 18.9 Phe 12 * 2 16.0 14.2 14.3 His 8 f l 5.5 4.4 5.4 LY 5 28 f 4 21.1 21.8 25.1 Arg 12 f 2 17.5 17.5 20.2 Trp ND’ 4.6 2.0 ND
Based on a monomer molecular mass of 38,000 Da and an average amino acid molecular weight of 105. Average and standard deviation of eight determinations.
Expected no. of amino acids in prenyltransferase
Heuea” Porcine’ Human‘ Aviand
‘ Based on a monomer molecular mass of 38,500 Da (50,51). Based on a monomer molecular mass of 38,400 Da (50,52). Based on a monomer molecular mass of 38,500 Da (51,53).
e ND, not determined.
ulated rubber synthesis in purified rubber particles. This stimulation, also observed with GPP and NPP (33, 34), was attributed to initiation of new rubber molecules. When puri- fied rubber transferase was assayed with dimethylallyl pyro- phosphate (DMAPP), geranyl pyrophosphate (GPP), and trans,tram-farnesyl pyrophosphate (t,t-FPP) in the absence of deproteinated rubber particles, only DMAPP and GPP were found to be substrates. Hydrolysis of an incubation of DMAPP plus [14C]IPP by potato acid phosphatase in 60% methanol and 0.1% Triton X-100 at pH 4.7 (acetate) followed by analysis with a reverse phase TLC system which separates polyprenols (40) reveals two major radioactive spots corre- sponding to Clo and c15. A trace amount of material with an R F expected for geranylgeraniol (C20) was also seen. Under these conditions potato acid phosphatase hydrolyzes poly- prenylpyrophosphates of at least C,, (41, 42).6
Initial analysis of an acid hydrolyzed extract revealed a small amount of radioactivity in the region between CIS and CSo in addition
Analysis of the enzymatically hydrolyzed products derived from DMAPP and IPP by GC and GC/MS revealed that the Clo product was geraniol (trans) and that the C16 product was t,t-farnesol with a trace (less than 3%) of either cis,trans- farnesol or trans,cis-farnesol. Thus, the FPP synthetase ac- tivity present in other partially purified rubber transferase preparations (4,5,30) is a property of the purified enzyme.
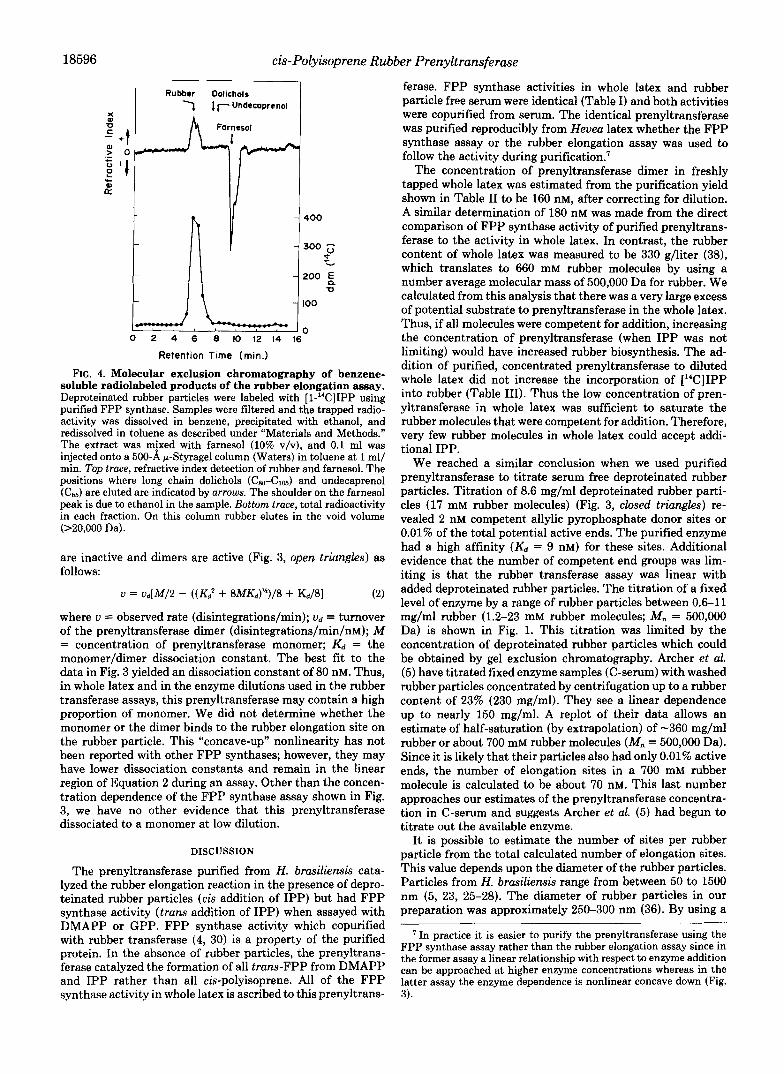
Incorporation of [14C]IPP into High Molecular Weight Polyisoprene-With the observation that the purified pren- yltransferase catalyzed the tram addition of IPP to DMAPP and GPP to make low molecular weight t,t-FPP it was imper- ative to demonstrate that the rubber elongation assay meas- ures the incorporation of IPP into high molecular weight rubber. Two benzene extractions of radiolabeled rubber trapped on assay filters dissolved 76% of the radioactivity. No radioactivity was extracted from the benzene solution by 6 N NaOH, therefore low molecular weight organic pyrophos- phates were not present. Addition of an equal volume of ethanol precipitated the rubber (43, 44) which was removed by centrifugation. No radioactivity remained in the benzene- ethanol supernatant fluid. The rubber film was redissolved in benzene, and 70% of the radioactivity of the original benzene extract was in this solution. Losses were due to the fact that rubber solutions left a film on glassware during transfer steps. As +own in Fig. 4, when chromatographed in toluene on a 500 A F-Styragel column, 95% of the radioactivity injected was eluted at a molecular mass greater than 20,000 Da (exclu- sion limit of the column). Less than 1.5% of the injected radioactivity was found in the region where farnesol(222 Da) was eluted. In contrast, when incubations of purified prenyl- transferase with DMAPP and [14C]IPP were acid-hydrolyzed, dried, extracted with benzene, and injected under the same conditions, the radioactivity was coeluted with farnesol, a major subfraction was eluted with geraniol, and a few percent were eluted in the Czo region. In the latter incubations no counts were observed in the high molecular weight region ( > C d
Concentration Dependence of FPP Synthase Assay-As dis- cussed above, variation of enzyme concentration in the rubber transferase assay revealed a nonlinear concave down enzyme dependence (Fig. 3, closed triangles). In contrast, variation of the prenyltransferase in the FPP synthase assay revealed a nonlinear concave up enzyme dependence that could be fit to a model by a rate dependence in which dissociated monomers
to the major spots at Clo and CIS. This material of apparent higher molecular weight is attributed to hydrocarbon producing re- arrangements of [’4C]FPP and/or [“CIGPP during acid hydrolysis.
18596 cis-Polyisoprene Rubber Prenyltransferase
I
I
'I t t I-
Rubber Dolicholr 7 Undecoprenol
0 I L - 4 2 4 6 !L-+.+. 8 IO 12 14 16 I O
Retention Time (min.)
FIG. 4. Molecular exclusion chromatography of benzene- soluble radiolabeled products of the rubber elongation assay. Deproteinated rubber particles were labeled with [1-"C]IPP using purified FPP synthase. Samples were filtered and the trapped radio- activity was dissolved in benzene, precipitated with ethanol, and redissolved in toluene as described under "Materials and Methods." The extract was mixed with farnesol (10% v/v), and 0.1 ml was injected onto a 500-A F-Styragel column (Waters) in toluene at 1 ml/ min. Top trace, refractive index detection of rubber and farnesol. The positions where long chain dolichols ( C & & , ) and undecaprenol (C66) are eluted are indicated by arrows. The shoulder on the farnesol peak is due to ethanol in the sample. Bottom trace, total radioactivity in each fraction. On this column rubber elutes in the void volume (>20,000 Da).
are inactive and dimers are active (Fig. 3, open triangles) as follows:
U = Ud[M/2 - (( K: i- 8M&)")/8 i- &/8] ( 2 )
where u = observed rate (disintegrations/min); ud = turnover of the prenyltransferase dimer (disintegrations/min/nM); M = concentration of prenyltransferase monomer; K d = the monomer/dimer dissociation constant. The best fit to the data in Fig. 3 yielded an dissociation constant of 80 nM. Thus, in whole latex and in the enzyme dilutions used in the rubber transferase assays, this prenyltransferase may contain a high proportion of monomer. We did not determine whether the monomer or the dimer binds to the rubber elongation site on the rubber particle. This "concave-up" nonlinearity has not been reported with other FPP synthases; however, they may have lower dissociation constants and remain in the linear region of Equation 2 during an assay. Other than the concen- tration dependence of the FPP synthase assay shown in Fig. 3, we have no other evidence that this prenyltransferase dissociated to a monomer at low dilution.
DISCUSSION
The prenyltransferase purified from H. brasiliensis cata- lyzed the rubber elongation reaction in the presence of depro- teinated rubber particles (cis addition of IPP) but had FPP synthase activity (trans addition of IPP) when assayed with DMAPP or GPP. FPP synthase activity which copurified with rubber transferase (4, 30) is a property of the purified protein. In the absence of rubber particles, the prenyltrans- ferase catalyzed the formation of all trans-FPP from DMAPP and IPP rather than all cis-polyisoprene. All of the FPP synthase activity in whole latex is ascribed to this prenyltrans-
ferase. FPP synthase activities in whole latex and rubber particle free serum were identical (Table I) and both activities were copurified from serum. The identical prenyltransferase was purified reproducibly from Heuea latex whether the FPP synthase assay or the rubber elongation assay was used to follow the activity during purification?
The concentration of prenyltransferase dimer in freshly tapped whole latex was estimated from the purification yield shown in Table I1 to be 160 nM, after correcting for dilution. A similar determination of 180 nM was made from the direct comparison of FPP synthase activity of purified prenyltrans- ferase to the activity in whole latex. In contrast, the rubber content of whole latex was measured to be 330 g/liter (38), which translates to 660 mM rubber molecules by using a number average molecular mass of 500,000 Da for rubber. We calculated from this analysis that there was a very large excess of potential substrate to prenyltransferase in the whole latex. Thus, if all molecules were competent for addition, increasing the concentration of prenyltransferase (when IPP was not limiting) would have increased rubber biosynthesis. The ad- dition of purified, concentrated prenyltransferase to diluted whole latex did not increase the incorporation of ['*C]IPP into rubber (Table 111). Thus the low concentration of pren- yltransferase in whole latex was sufficient to saturate the rubber molecules that were competent for addition. Therefore, very few rubber molecules in whole latex could accept addi- tional IPP.
We reached a similar conclusion when we used purified prenyltransferase to titrate serum free deproteinated rubber particles. Titration of 8.6 mg/ml deproteinated rubber parti- cles (17 mM rubber molecules) (Fig. 3, closed triangles) re- vealed 2 nM competent allylic pyrophosphate donor sites or 0.01% of the total potential active ends. The purified enzyme had a high affinity ( & = 9 nM) for these sites. Additional evidence that the number of competent end groups was lim- iting is that the rubber transferase assay was linear with added deproteinated rubber particles. The titration of a fixed level of enzyme by a range of rubber particles between 0.6-11 mg/ml rubber (1.2-23 mM rubber molecules; M, = 500,000 Da) is shown in Fig. 1. This titration was limited by the concentration of deproteinated rubber particles which could be obtained by gel exclusion chromatography. Archer et al. (5) have titrated fixed enzyme samples (C-serum) with washed rubber particles concentrated by centrifugation up to a rubber content of 23% (230 mg/ml). They see a linear dependence up to nearly 150 mg/ml. A replot of their data allows an estimate of half-saturation (by extrapolation) of -360 mg/ml rubber or about 700 mM rubber molecules (M, = 500,000 Da). Since it is likely that their particles also had only 0.01% active ends, the number of elongation sites in a 700 mM rubber molecule is calculated to be about 70 nM. This last number approaches our estimates of the prenyltransferase concentra- tion in C-serum and suggests Archer et al. (5) had begun to titrate out the available enzyme.
It is possible to estimate the number of sites per rubber particle from the total calculated number of elongation sites. This value depends upon the diameter of the rubber particles. Particles from H. brasiliensis range from between 50 to 1500 nm (5, 23, 25-28). The diameter of rubber particles in our preparation was approximately 250-300 nm (36). By using a
In practice it is easier to purify the prenyltransferase using the FPP synthase assay rather than the rubber elongation assay since in the former assay a linear relationship with respect to enzyme addition can be approached at higher enzyme concentrations whereas in the latter assay the enzyme dependence is nonlinear concave down (Fig. 3).

cis-Polyisoprene Rubber Prenyltransferase 18597
density of 0.9 g/ml for natural rubber (45) and a rubber content of 8.6 mg/ml, rubber particle diameters of 100, 300, 1. Archer, B. L., and Cockbain, E. G. (1969) Methods Enzymol. 15,476-480
and 500 nm correspond to 30, 1, and 0.24 nM rubber particles. 3. Archer, B. L., and Audley, B. G. (1967) in Advances in Enzymology (Nord,
low as 0.03 growing ends per rubber particle. Active ends may Research Association Jubilee Conference in Cambridge (Mullins, L., ed)
may have no growing ends whereas smaller particles may have Biochem. J. 89,565-574
diameter washed rubber particles have a disproportionately
were made for their neater surface to volume ratio.
REFERENCES
2. Archer, B. L., and Audley, B. G. (1987) Bot. J. Linn. SOC. 9 4 , 181-196
Thus, nM active ends leads to a calculation Of from to as 4. Archer, B. L. (1964) in Proceedings of the Natural Rubber Producers'
not be distributed among particles- Larger particles 5. Arcger, B. L., Audley, B. G., Cockbain, E. G. and McSweeney, G. P. (1963)
multiple ends. Archer et al. (5) have &own that small 6. Poulter, c. D., and Rilling, H. c. (1981) in Biosynthesis of Isoprenoid Compounds (Spurgeon, S. L., and Porter, J. W., eds) Vol. 1, John Wiley
high rate of [14c]1pp incorporation even when corrections 7. Takahashi, I., and Ogura, K. (1981) J. Biochem. (Tokyo) 89,1581-1587 & Sons, New York
F. F., ed) Vol. 29, pp. 221-257, Interscience, New York
p . 101-112, Maclaren & Sons, Ltd., London
8. Brems, D. N., Bruenger, E., and Rllling, H. C. (1981) Biochemistry 2 0 , 3711-3818 -
A low percentage of rubber molecules in our preparation was competent to add to IPP. We expect the loss of elongation sites in latex occurs as a consequence of the head growth mechanism (46) of rubber polymerization in which the poly- mer rather than the monomer is activated. Three mechanisms for activated allylic pyrophosphate loss from rubber molecules can be considered. Both nonenzymatic hydrolysis, especially at low pH and enzymatic hydrolysis by phosphatases will reduce the number of competent allylic pyrophosphate sites. However, this mechanism is unlikely to be of physiological significance, since latex is compartmentalized and the latex phosphatase and inorganic pyrophosphatase activities are confined inside organelles called lutoid particles (3-5). We postulate a third, less obvious explanation for polymer acti- vation on the basis of the known ability of liver FPP synthase to catalyze the inorganic pyrophosphate-dependent solvolysis of allylic pyrophosphate substrates DMAPP and GPP (6). In the presence of optimal inorganic pyrophosphate this rate is 2% of the normal prenyl transfer rate. In whole latex, rubber molecules are continually exposed to the prenyltransferase whether or not IPP is available for elongation. The amount of enzyme-dependent hydrolysis in laticifers will be dependent upon the efficiency of inorganic pyrophosphate removal by inorganic pyrophosphatase in the lutoid particles and the ratio of prenyltransferase to solvolysis activities for the Heuea prenyltransferase. Rubber particles from trees that were con- tinually tapped and in which rubber was newly synthesized had more competent end groups (-30 times) than rubber particles from untapped trees (36). The contribution of pren- yltransferase-dependent solvolysis to the mechanism of chain termination is yet to be determined.
The titrations in Fig. 3 can be used to calculate the maxi- mum turnover per active site for each of the assays. In a typical assay an elongation complex added approximately 7000 isoprene units at a rate of 60 min-' in the rubber elongation assay (Fig. 3, closed triangles). The most efficient mechanism for the polymerization is for the elongation com- plex, which includes the Heuea prenyltransferase, to remain associated with the individual rubber molecule during these thousands of additions. One active site in the fully associated prenyltransferase dimer incorporated IPP into GPP and t,t- FPP at a rate of 600 min". A 10-fold difference in the rates of prenylation in each reaction indicates they may not have the same rate determining step.
Acknowledgments-We would like to thank Dr. C. Dale Poulter for many helpful suggestions and discussions.
9. Ogura, K., Nishino, T., Shinka, T., and Seto, S. (1985) Methods Enzymol. ~ ~~ ~~~~
10. Eberhardt, N. L., and Rilling, H. C. (1975) J. Biol. Chem. 260,863-866 11. Shinka, T., Ogura, K., and Seto, S. (1975) J . Biochem. (Tokyo) 78, 1177-
12. Takahashi, I., and Ogura, K. (1982) J. Biochem. (Tokyo) 92,1527-1537 13. Fujii, H., Koyama, T., and Ogura, K. (1982) J. Bid . Chem. 267 , 14610-
14. Takahashi, I., Ogura, K., and Seto, S. (1980) J. Biol. Chem. 2 6 6 , 4539-
15. Sagarni, H., Ogura, K., and Seto, S. (1977) Biochemistry 16,4616-4622 16. Baba, T., and Allen, C. M. (1980) Arch. Biochem. Biophys. 200,474-484 17. Muth, J. D., and Allen, C. M. (1984) Arch Biochem. Biophys. 230,49-60 18. Allen, C. M., Jr., Keenan, M. V., and Sack, J. (1976) Arch. Biochem.
19. Adatr. W. L.. Jr.. Cafmever. N.. and Keller. R. K. (1984) J. Biol. Chem.
110,167-170
1181
14612
4543
Bwphys. 175,236-248 - . .
259,444114446 20. Saito, A., and Rilling, H. C. (1979) J. Biol. Chem. 264,8511-8515
22. Fujii, H., Sagami, H., Koyama, T., Ogura, K., Seto, S., Baba, T., and Allen, 21. Baba, T., and Allen, C. M., Jr. (1978) Biochemistry 17,559&5604
C. M. (1980) Biochem. Biophys. Res. Commun. 96,1648-1653 23. McMullen, A. I., and McSweeney, G. P. (1966) Biochem. J. 101,42-47
25. Ho, C. C., and Ng, W. L. (1979) Colloid & Poiym. Sci. 267,406-412 24. Sipat, A. (1985) Methods Enzymol. 110.40-51
26. Lucas, F. F. (1938) Ind. Eng. Chem. 30,146-153 27. Schoon, T. G. F., and van der Bie, G. J. (1955) J. Polym. Sci. 1 6 , 6 3 4 28. Stavely, F. W., Biddison, P. H., Forster, M. J., Dawson, H. G., and Binder,
29. Audley, B. G. (1966) Biochem. J. 98,335-341 30. Madhavan, S., and Benedict, C. R. (1984) Phnt Physiol. 76,908-913 31. Backhaus, R. A., and Walsh, S. (1983) Bot. Gaz. 144,391-400 32. Backhaus, R. A. (1985) Isr. J . Bot. 34,283-293 33. Archer, B. L., Audley, B. G., and Bealing, F. J. (1982) Plast. Rubber Int. 7 ,
34. Archer, B. L., Audley, B. G., and Bealing, F. J. (1981) Rubber Dev. 34,96-
35. Archer, B. L., and Audley, B. G. (1973) in Phytochemistry (Miller, L. P.,
36. Dennis, M. S., and Light, D. R. (1989) J. Bwl. Chem. 264 , 18608-18617 37. Hirschbuehler, K. R. (1979) The Baosynthesrs of Natural Rubber and Other
Hydrocarbon Polymers. Ph.D. thesis, Washington University, Sever In-
38. Bartlett, D. L., King, C.-H. R., and Poulter, C. D. (1985) Methods Enzymol. stitute of Technology
110,171-183 39. Segel, I. H. (1975) Enzyme Kinetics, p. 74, Wiley-Interscience, New York 40. Mankowski, T., Jankowski, W., Chojnacki, T., and Franke, P. (1976)
41. Fujii, H., Koyama, T., and Ogura, K. (1982) Biochim. Biophys. Acta 7 1 2 ,
42. Koyama, T., Fujii, H., and Ogura, K. (1985) Methods Enzymol. 1 1 0 , 153- 155
43. Hager, T., MacArthur, A., McIntyre, D., and Seeger, R. (1979) Rubber Chem. Technol. 52,693-709
44. Archer, B. L., Barnard, D., Cockbain, E. G., Cornforth, J. W., Cornforth, R. H., and Popjak, G. (1966) Proc. R. SOC. L o n d . B Biol. Sci. 163 , 519- 523
J. L. (1961) Rubber Chem. Technol. 34,423-432
109-111
98
ed) Vol. 2, Van Nostrand Reinhold Co., Inc., New York
Biochemistry 16,2125-2130
716-718
45. Kaye, G. W. C., and Laby, T. H. (eds) (1978) Tables of Physical and
46. Kznberg, A. (1980) DNA Replication, p. 94, W. H. Freeman & Co., San Chemical Constants, 14th Ed., Longman, London
47.
49. 48.
50. 51.
Davisson, V. J., Woodside, A. B., and Poulter, C. D. (1985) Methals
Bradford, M. M. (1976) Anal. Biochem. 72,248-254 Morrissey, J. H. (1981) Anal. Biochem. 72,307-310 Barnard, G. F., and Popj&, G. (1981) Biochim. Biophys. Acta 661,87-99 Barnard, G. F., and Popj&, G. (1890) Biochim. Biophys. Acta 6 1 7 , 169-
Francisco
Enzymol. 1 1 0 , 130-144
182 52. Barnard, G. F. (1985) Methods Enzymol. 110 , 155-167 53. Reed, B. C., and Rilling, H. C. (1976) Biochemistry 16,3739-3745
![A Stilbenoid-Specific Prenyltransferase Utilizes ...A Stilbenoid-Specific Prenyltransferase Utilizes Dimethylallyl Pyrophosphate from the Plastidic Terpenoid Pathway1[OPEN] Tianhong](https://img.pdfslide.us/doc/110x75/5eb88723cf80df2b4822048a/a-stilbenoid-specific-prenyltransferase-utilizes-a-stilbenoid-speciic-prenyltransferase.jpg)




























