Embed Size (px)
Citation preview

Brain (2001), 124, 1619–1634
The neural representation of nouns and verbs:PET studiesLorraine K. Tyler, Richard Russell, Jalal Fadili and Helen E. Moss
Department of Experimental Psychology, University of Correspondence to: Professor L. K. Tyler, Centre forCambridge, UK Speech and Language, Department of Experimental
Psychology, University of Cambridge, CambridgeCB2 3EB, UKE-mail: [email protected]
SummaryNeuropsychological studies of patients with selectivedeficits for nouns or verbs have been taken as evidencefor the neural specialization of different word classes.Noun deficits are associated with lesions in anteriortemporal regions while verb deficits arise from left inferiorfrontal lesions. However, neuroimaging studies do notunequivocally support this account, with only some studiessupporting claims for regional specialization. We carriedout two PET studies to determine whether there is anyregional specialization for the processing of nouns and
Keywords: PET; nouns and verbs
Abbreviations: fMRI � functional MRI; MNI � Montreal Neurological Institute; rCBF � regional cerebral blood flow;ROC � reviewer operating characteristic; SPM � statistical parametric mapping
IntroductionFunctional neuroimaging studies, using PET and functionalMRI (fMRI), suggest that conceptual knowledge isrepresented within an extensive network involving the leftlateral temporal lobe, left posterior parietal lobe and leftinferior frontal gyrus, possibly including some homologousareas in the right hemisphere (Vandenberghe et al., 1996;Mummery et al., 1998). Neuropsychological studies ofpatients with category-specific deficits suggest that thissemantic network might be organized further as a functionof factors such as word class. For example, patients havebeen reported with selective deficits for nouns or verbs, withlesions to anterior temporal regions associated with noundeficits, and left frontal lesions with verb deficits (e.g.Goodglass, 1966; Damasio and Tranel, 1993; Daniele et al.,1994). This neural differentiation is compatible withbehavioural studies showing that children and adults appearto treat nouns and verbs differently. For example, they areacquired at different rates, with nouns being learned earlierthan verbs (Gentner, 1981; Gleitman, 1994), and adultsgenerally exhibit poorer performance with verbs than nounson a variety of tests. These behavioural patterns are, in turn,
© Oxford University Press 2001
verbs. One study used the lexical decision task and theother used a more semantically demanding task, i.e.semantic categorization. We found robust activation of asemantic network extending from left inferior frontalcortex into the inferior temporal lobe, but no differencesas a function of word class. We interpret these data withinthe framework of cognitive accounts in which conceptualknowledge is represented within a non-differentiateddistributed system.
consistent with linguistic analyses of nouns and verbs whichclaim that they differ in their structure and organization.Gentner, for example, has claimed that nouns have coreinvariant meanings, associated with denser connectionsbetween properties (Gentner, 1981), while Miller andFellbaum argue that verbs have looser internal structure andgreater polysemy (Miller and Fellbaum, 1991). Nouns maybe organized into hierarchies with many shared correlatedproperties, while verbs may form a matrix-like structure,where many semantic properties are orthogonally relatedrather than correlated (Huttenlocher and Lui, 1979; Graesseret al., 1987).
However, not all of the data support the claim for neuralspecialization of conceptual knowledge as a function of formclass. For example, there are several reports of patients withverb deficits whose lesions lie outside the left frontal regions(see Daniele et al., 1994) and some whose lesions primarilycompromise the left temporal lobe (Damasio and Tranel,1993). Another problem is that most of the availableneuropsychological data come from naming and other lexicalretrieval tasks, rather than from tasks which probe semantic

1620 L. K. Tyler et al.
representations per se (McCarthy and Warrington, 1985;Caramazza and Hillis, 1991; Breedin et al., 1998). It ispossible that these patients suffer essentially from a namingdeficit in which their major problem is an inability to producethe phonological form of the word, rather than from animpairment in the representation of semantic knowledge.Moreover, behavioural differences do not necessitate neuralspecialization. A number of recent cognitive models ofconceptual knowledge claim that what appear to be selectivedeficits for specific categories/domains of conceptualknowledge can derive from random damage to a distributedsemantic system. Category and domain structure emerges outof this system as a function of the structure and content ofindividual concepts (Gonnerman et al., 1997; Tyler et al.,2000).
Recent functional neuroimaging studies (PET and fMRI)investigating the neural representation of nouns and verbshave been interpreted as supporting the claim for regionalspecialization, with the left inferior prefrontal cortex beingspecialized for verbs and left temporal cortex specialized fornouns (Petersen et al., 1988; Perani et al., 1999). However,the results are, in fact, very mixed and do not support thisstraightforward dissociation unequivocally. Almost all of theimaging studies use a verb generation task in which subjectsproduce a verb in response to a target noun. One of the fewimaging studies which did not use the verb generationtask is the lexical decision study by Perani and colleaguescomparing nouns and verbs (Perani et al., 1999). Thisprovides the clearest evidence for cortical specialization forverbs. When subjects were asked to make lexical decisionsto written words and non-words, the perisylvian languageareas were activated (Wise et al., 1991) for both nouns andverbs. However, verbs produced additional activation overnouns in the left dorsolateral frontal cortex, superior parietaland anterior and middle temporal lobes. Other studies,primarily using the verb generation task (e.g. Petersen et al.,1988; Martin et al., 1995), do not replicate this finding.Although the initial study by Petersen and colleagues(Petersen et al., 1988) reported additional activation for verbsprimarily in the left prefrontal cortex, this region could notbe claimed to be specialized for verb processing since, in asubsequent study, the same pattern was obtained in a namegeneration task (Petersen et al., 1989). Similarly, even thoughMartin and colleagues found greater activation in leftprefrontal cortex when subjects generated action words(verbs) compared with object names, this same region wasalso activated when colour words (adjectives) were generated,although to a lesser extent (Martin et al., 1995). Other studiesreport no differences between nouns and verbs (Warburtonet al., 1996; Buckner et al., 2000). In the study of Warburtonand colleagues, a large left lateralized network was activatedfor nouns and verbs, including temporal, parietal andprefrontal regions (Warburton et al., 1996). This networkwas activated more highly for verbs than nouns, but therewas no regional specificity for one type of word comparedwith another. Similarly, Buckner and colleagues show that
verb generation and word stem completion (involving nouns)activate the same cortical regions (Buckner et al., 2000).They found left prefrontal activation extending from boththe inferior and dorsal parts of the inferior frontal gyrus intomore anterior prefrontal regions and including left inferiortemporal cortex.
Variation in the pattern of activations in functional imagingstudies may be due partly to problems in the selection ofexperimental stimuli, and to the limitations of the verbgeneration task. We know from numerous behaviouralcognitive studies that stimulus factors such as word length,familiarity, frequency and imageability play a crucial role inword recognition and language comprehension (Rubin, 1980;Gernsbacher, 1984). Since these are important variables inbehavioural studies, it is likely that they will also affectperformance in imaging studies and thus sets of stimuli needto be matched on these properties. However, since stimuluscharacteristics are rarely specified in imaging papers on verb–noun differences, it is impossible to determine whethermaterials were in fact matched in the appropriate ways,although there is no indication in most published reports thatthey are.
Another set of problems is raised by the use of the verbgeneration task. This task is used in most of the studiesexploring noun–verb differences, where activation resultingfrom verb generation is often compared with noun generationor stem completion (e.g. Warburton et al., 1996; Bruckneret al., 2000). In the verb generation task, subjects initiallyare given a noun cue and asked to produce a related verb.Thus, in a single trial, they encounter both nouns and verbs,and both will contribute towards activation, so the task cannotbe thought of as selectively tapping verb processing. Secondly,the relationship between the noun and verb is left unspecified,as is the means by which verb–noun pairs are selected;consequently, the sets of word pairs may not be sufficientlyhomogeneous. Finally, form–class ambiguity, i.e. the fact thatmany verbs also function as nouns and vice versa, is ignored.These methodological issues cast doubt on the reliability ofthe findings from previous studies, and provided themotivation for the two PET studies presented here.
The aim of the two PET experiments reported here wasto determine whether there is neural specialization for nounsand verbs. We attempted to overcome some of the limitationsof earlier studies by matching nouns and verbs on the relevantvariables of letter length, familiarity and imageability. Inaddition, we used two different behavioural paradigms. Inthe first study, we used visual lexical decision since this taskis used widely in activation studies of lexical and semanticprocessing, and is clearly sensitive to semantic processing(Price et al., 1996; Perani et al., 1999). In a second experiment,we used a semantic categorization task which requires moreeffortful control of semantic information and is thus predictedto activate the left inferior prefrontal cortex (Wagner et al.,1997; Buckner et al., 2000), the area that has been implicatedmost strongly in verb deficits and verb processing. In addition,the task demands of the semantic categorization task are

PET studies of noun and verb processing 1621
more comparable with the verb generation task, whileavoiding some of the pitfalls associated with verb generation.In the semantic categorization task, three written cue wordsare presented sequentially, and subjects make decision asquickly as possible about whether a fourth (target) wordbelongs to the same category as the cue words (Devlin et al.,2000). Both the semantic categorization and verb generationtasks require effortful semantic processing. In the case ofverb generation, response selection is required, while forsemantic categorization subjects have to form a conceptualrepresentation of the three cue words, hold this in memoryand compare it with a target word. Since the semanticcategorization task should activate left inferior prefrontalcortex more strongly than the lexical decision task, wepredicted that we would then be in a position to see whetherthere was more activation in this area for verbs than nouns.
MethodsSubjectsSubjects were all native English speakers aged between 18and 40 years. There were nine subjects in Experiment 1,ranging in age between 21 and 34 years (mean � SD,26 � 5), and eight subjects ranging in age between 21 and30 years (25 � 3) in Experiment 2. All participants in theimaging studies were right-handed, neurologically normalmales. Subjects had no prior knowledge of the study, andwritten informed consent was obtained from all subjectsaccording to protocols approved by the Cambridge LocalEthical Research Committee.
Imaging methodsIn both experiments, there were 12 scans per subject, eighttest scans (two for each of the four conditions) and fourbaseline scans. Stimuli were presented using DMDX (Forsterand Forster, 1990) running on a Dell PC (Pentium II, 400MHz) running Windows 98. Items were presented in black26 point Arial font against a white background on a videomonitor ~1 m from the subjects’ heads. The room was dimlylit. In all tasks, subjects responded with a right hand two-choice button press. Accuracy and reaction times for bothtypes of task were recorded to the nearest millisecond.
Scans were performed at the Wolfson Brain Imaging Centrein Cambridge, UK on a GE Advance PET Scanner (GeneralElectric Medical Systems, Milwaukee, Wisc., USA). Itcomprises 18 rings of crystals, which results in 35 imageplanes, each 4.25 mm thick. The axial field-of-view was 15.3cm, thus allowing for whole brain acquisition. Each subjectreceived a bolus of 300 MBq before each scan for a totalradiation exposure of 4.2 mSv. Stimuli were presented duringthe first 45 s, to coincide with the critical period of traceruptake (Silbersweig et al., 1993). The emission data wereacquired with the septa retracted (3D mode) and reconstructedusing the PROMIS algorithm (Kinahan and Rogers, 1989)
with an unapodized Colsher filter. Corrections were appliedfor randoms, scatter, attenuation and dead time. The voxelsizes were 2.34 mm � 2.34 mm � 4.25 mm.
Experiment 1: lexical decision taskMethodsTwo tasks were used in this experiment, one for test conditionsand another for baseline conditions. The test conditions usedlexical decision, where subjects decide whether a string ofletters forms a word in English. The baseline conditions useda letter detection task in which subjects decided whether astring of letters (which did not form a word in English)contained the letter ‘x’. This task required subjects to makea timed response without any semantic processing.
Stimuli for lexical decision conditions consisted of wordsand orthographically legal, pronounceable non-word letterstrings (e.g. hicton, blape). There were a total of 240 testwords. These were divided into four equal groups: highimageability nouns, high imageability verbs, low imageabilitynouns and low imageability verbs. We included imageabilityas a factor in this study since verbs tend to be more abstractthan nouns, and many behavioural studies have showndifferences between abstract and concrete words, withconcrete words showing an advantage relative to abstractwords (we use the terms imageability and concretenessinterchangeably here since they are highly correlated). Also,concrete and abstract words often dissociate following braindamage (Franklin, 1989), suggesting that they form distinctdomains of knowledge, and imaging studies suggest thatabstract words may impose greater processing demands onthe semantic system (Kiehl et al., 1999). By ensuring thatnouns and verbs are matched on imageability, we avoid anypotential confounding effects of this variable, and can alsoevaluate its contribution to levels of activation.
Words in the four conditions were between three and eightletters long (inclusive). Items within the two sets of concretewords were matched for imageability, as were items withinthe two sets of abstract words. Imageability ratings wereobtained either from the Coltheart database (Coltheart, 1981)or from norms collected from 15 young normal controls fromthe Cambridge University Centre for Speech and Languagesubject pool who rated the 250 words from 1 (not imageable)to 7 (highly imageable). Because most of our pre-test wordswere of low imageability, an additional 38 items with ratingsof 600 or higher from the Coltheart database were includedin order to encourage the subjects to use the entire range ofscores. The items had an average Coltheart rating of 628,and were given an average rating of 658 in the pre-test,suggesting rating consistency between the two sets. In total, 94items from this locally collected set were used as experimentalstimuli in our imaging study, 41 concrete verbs and 53abstract verbs. We also matched the sets on their frequencyof occurrence as a member of a specific word class (i.e. asnouns or verbs). Many words in English assume different
1622 L. K. Tyler et al.
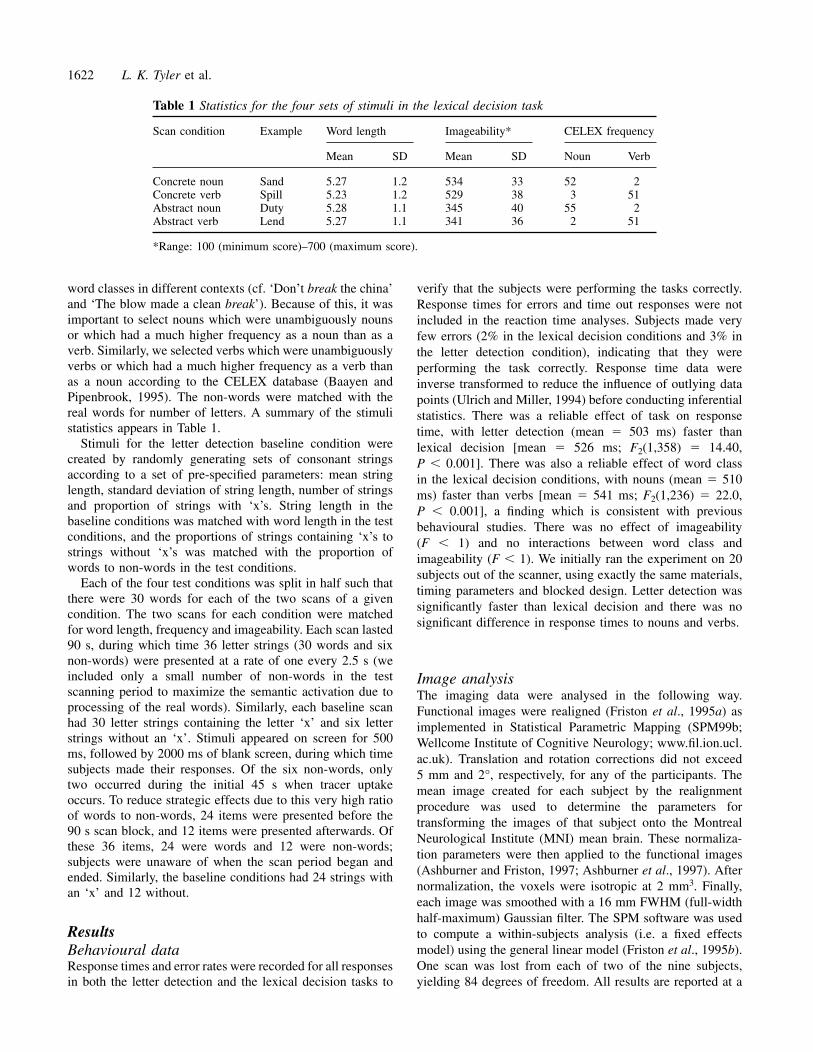
Table 1 Statistics for the four sets of stimuli in the lexical decision task
Scan condition Example Word length Imageability* CELEX frequency
Mean SD Mean SD Noun Verb
Concrete noun Sand 5.27 1.2 534 33 52 2Concrete verb Spill 5.23 1.2 529 38 3 51Abstract noun Duty 5.28 1.1 345 40 55 2Abstract verb Lend 5.27 1.1 341 36 2 51
*Range: 100 (minimum score)–700 (maximum score).
word classes in different contexts (cf. ‘Don’t break the china’and ‘The blow made a clean break’). Because of this, it wasimportant to select nouns which were unambiguously nounsor which had a much higher frequency as a noun than as averb. Similarly, we selected verbs which were unambiguouslyverbs or which had a much higher frequency as a verb thanas a noun according to the CELEX database (Baayen andPipenbrook, 1995). The non-words were matched with thereal words for number of letters. A summary of the stimulistatistics appears in Table 1.
Stimuli for the letter detection baseline condition werecreated by randomly generating sets of consonant stringsaccording to a set of pre-specified parameters: mean stringlength, standard deviation of string length, number of stringsand proportion of strings with ‘x’s. String length in thebaseline conditions was matched with word length in the testconditions, and the proportions of strings containing ‘x’s tostrings without ‘x’s was matched with the proportion ofwords to non-words in the test conditions.
Each of the four test conditions was split in half such thatthere were 30 words for each of the two scans of a givencondition. The two scans for each condition were matchedfor word length, frequency and imageability. Each scan lasted90 s, during which time 36 letter strings (30 words and sixnon-words) were presented at a rate of one every 2.5 s (weincluded only a small number of non-words in the testscanning period to maximize the semantic activation due toprocessing of the real words). Similarly, each baseline scanhad 30 letter strings containing the letter ‘x’ and six letterstrings without an ‘x’. Stimuli appeared on screen for 500ms, followed by 2000 ms of blank screen, during which timesubjects made their responses. Of the six non-words, onlytwo occurred during the initial 45 s when tracer uptakeoccurs. To reduce strategic effects due to this very high ratioof words to non-words, 24 items were presented before the90 s scan block, and 12 items were presented afterwards. Ofthese 36 items, 24 were words and 12 were non-words;subjects were unaware of when the scan period began andended. Similarly, the baseline conditions had 24 strings withan ‘x’ and 12 without.
ResultsBehavioural dataResponse times and error rates were recorded for all responsesin both the letter detection and the lexical decision tasks to
verify that the subjects were performing the tasks correctly.Response times for errors and time out responses were notincluded in the reaction time analyses. Subjects made veryfew errors (2% in the lexical decision conditions and 3% inthe letter detection condition), indicating that they wereperforming the task correctly. Response time data wereinverse transformed to reduce the influence of outlying datapoints (Ulrich and Miller, 1994) before conducting inferentialstatistics. There was a reliable effect of task on responsetime, with letter detection (mean � 503 ms) faster thanlexical decision [mean � 526 ms; F2(1,358) � 14.40,P � 0.001]. There was also a reliable effect of word classin the lexical decision conditions, with nouns (mean � 510ms) faster than verbs [mean � 541 ms; F2(1,236) � 22.0,P � 0.001], a finding which is consistent with previousbehavioural studies. There was no effect of imageability(F � 1) and no interactions between word class andimageability (F � 1). We initially ran the experiment on 20subjects out of the scanner, using exactly the same materials,timing parameters and blocked design. Letter detection wassignificantly faster than lexical decision and there was nosignificant difference in response times to nouns and verbs.
Image analysisThe imaging data were analysed in the following way.Functional images were realigned (Friston et al., 1995a) asimplemented in Statistical Parametric Mapping (SPM99b;Wellcome Institute of Cognitive Neurology; www.fil.ion.ucl.ac.uk). Translation and rotation corrections did not exceed5 mm and 2°, respectively, for any of the participants. Themean image created for each subject by the realignmentprocedure was used to determine the parameters fortransforming the images of that subject onto the MontrealNeurological Institute (MNI) mean brain. These normaliza-tion parameters were then applied to the functional images(Ashburner and Friston, 1997; Ashburner et al., 1997). Afternormalization, the voxels were isotropic at 2 mm3. Finally,each image was smoothed with a 16 mm FWHM (full-widthhalf-maximum) Gaussian filter. The SPM software was usedto compute a within-subjects analysis (i.e. a fixed effectsmodel) using the general linear model (Friston et al., 1995b).One scan was lost from each of two of the nine subjects,yielding 84 degrees of freedom. All results are reported at a

PET studies of noun and verb processing 1623
P � 0.05 level after correcting for multiple comparisons(Worsley, 1996), in order to reduce the possibility of falsepositives. (The issue of whether it is more appropriate to usecorrected or uncorrected levels of significance is currently ahotly debated topic in neuroimaging research. We prefer touse corrected levels since this reduces the probability of falsepositives due to large numbers of comparisons over largedata sets and gives a better estimate of robust effects. Thiskind of approach is more compatible with statistical analysisprocedures in experimental psychology, and thus ensures thatthe cognitive behavioural studies and the cognitive activationstudies are subject to similarly rigorous statistical analysisprocedures.) The following contrasts were used in the dataanalysis.
(i) To determine regions common to all four lexicaldecision conditions compared with letter decision conditions,we masked the main effect of lexical conditions versusbaseline with each of the lexical conditions individuallyversus baseline. This contrast is: (CV � AV � CN � AN) –B masked with (CV – B), (AV – B), (CN – B), (AN – B),where regions of the main effect are only considered if theysurpass P � 0.05 in each of the masking contrast images(CV � concrete verbs; AV � abstract verbs; CN � concretenouns; AN � abstract nouns; B � baseline). This enablesus to determine which regions are commonly activated in allconditions. A less conservative approach is to analyse all ofthe conditions minus the baseline without masking. Wecarried out such analyses for the semantics�baseline and theform class and imageability contrasts and found no areas ofsignificant activation that were not also present in the maskedanalyses. The only difference between the two types ofanalysis was that the unmasked analysis produced moresignificant activations with larger extents than in the maskedanalyses. However, we have only reported the maskedanalyses in the text since the potential problem with unmaskedcontrasts is that they can mis-identify regions of commonactivation which, in fact, are only activated by a singlecondition.
(ii) Main effect of word class: to determine whether therewere any regions which were only activated in response toverbs, we contrasted the two verb categories (CV � AV)with the two noun categories (CN � AN) and masked thiscontrast with the four simple contrasts (CV–CN, CV–AN,AV–CN, AV–AN) and the two baseline contrasts (CV–B,AV–B). We carried out comparable analyses to look forregions specific to nouns.
(iii) Main effect of imageability: to assess the effects ofabstract relative to concrete words, we contrasted the twoabstract categories (AV � AN) with the two concretecategories (CV � CN) masking with the four simple contrastsas for the word class effects described above. Comparableanalyses were carried out to investigate the effects of concretewords relative to abstract words.
(iv) Cross-over interactions between word class andimageability: (CV � AN) – (CN � AV) and the reverse.
(v) Other interactions between word class and imageability:CV – (AV � CN � AN), AV – (CV � CN � AN), CN –(CV � AV � AN) and AN – (CV � AV � CN). Thiscollection of contrasts allowed us to consider overall effectsof the experimental conditions versus baseline, main effectsof word class and imageability, and interactions between them.
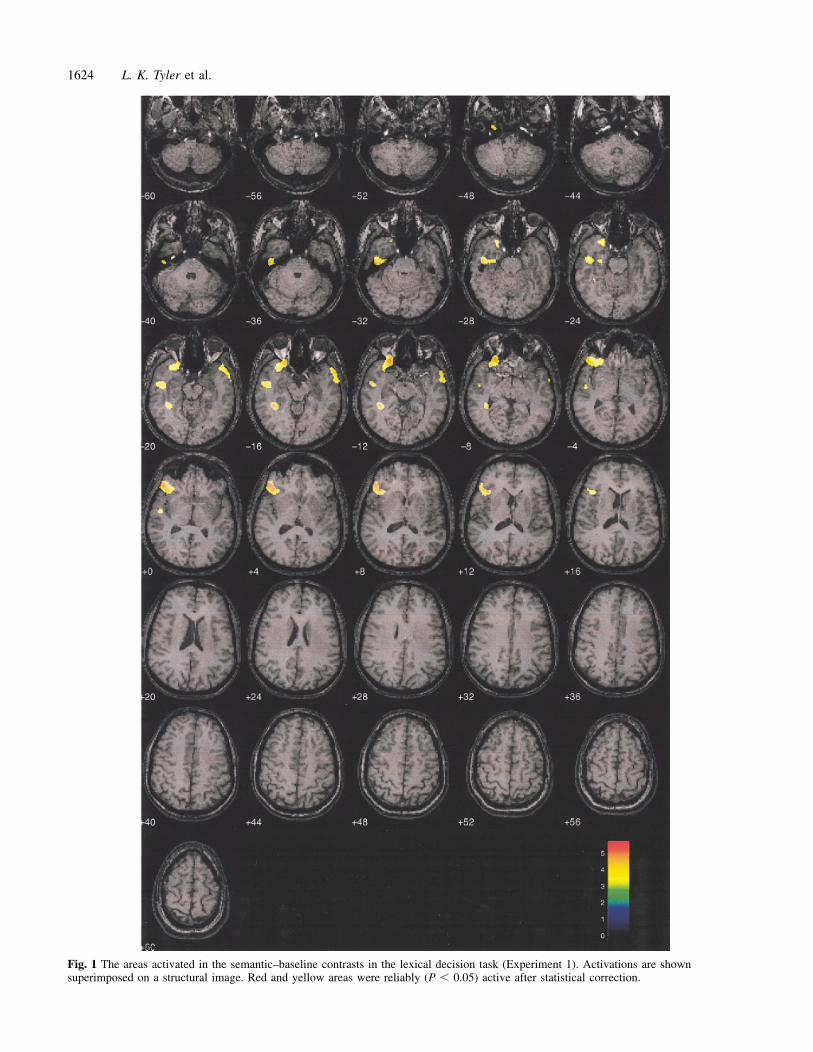
The lexical decision task relative to the baseline showedfour areas of activity. These areas (described in Table 2 andshown in Fig. 1) were in the left inferior frontal gyrus, leftfusiform gyrus, left middle and inferior temporal gyri andright middle temporal gyrus. [It is interesting to contemplatewhy the left fusiform is insignificant at the cluster level butsignificant at the voxel level. The multiple comparisoncorrection is based on statistical threshold based on theprobability of observing a cluster of height s and extent k bychance. This is directly related to random fields theory. Analmost isolated voxel (very small cluster) with a relativelyhigh signal height is likely to be an incorrectly activated areaand thus reflect a false positive. Conversely, large clusterswith low signal height are likely to be due to chance and arethen rejected at the voxel level. This may explain whathappened to the left fusiform which is rejected at the clusterlevel but is still significant at the voxel level.] After correctionfor multiple comparisons (Worsley et al., 1992; Friston et al.,1994), only the inferior frontal gyrus and fusiform gyrusactivations were significant at the voxel and cluster level.The other two regions of activation were significant only atthe cluster level. The areas of cortex activated by the lexicalconditions in contrast to the baseline were quite robust andoverlap with areas found in other studies (Mummery et al.,1996, 1998; Devlin et al., 2000).
Table 2 shows the results of the form class and imageabilitycontrasts. There were no areas of activity that were significantat either the voxel or the cluster level after corrections formultiple comparisons. The areas of maximal change in signalintensity, although not significant, are also shown in Table 2.Thus, there is no evidence for cortical regions specific to theprocessing of nouns or verbs, or to abstract or concretewords. This was the case in spite of the behavioural datawhich showed a response time advantage for nouns oververbs, a result which is consistent with other behaviouralexperiments.
Power analysesWe carried out further analyses to ensure that the experimenthad sufficient power to detect small differences in activation,if they were present in the data. In contrast to an introspectiveapproach that uses the voxel scores (possibly outliers) astypical observations to calculate the power of the analysis(Van Horn, 1998), we used a more stringent retrospectiveapproach using the whole data set to estimate the effect sizeand the proportion of active voxels. We applied a finitemixture-based model to fit a receiver operating characteristic(ROC) curve to the observed data (Gustard et al., 2000).
1624 L. K. Tyler et al.
Fig. 1 The areas activated in the semantic–baseline contrasts in the lexical decision task (Experiment 1). Activations are shownsuperimposed on a structural image. Red and yellow areas were reliably (P � 0.05) active after statistical correction.

PET studies of noun and verb processing 1625
Table 2 Brain areas of activity in the lexical decision task
Region Coordinates Voxel level Cluster level
x y z Pcorrected t Pcorrected Extent
Contrast (i), test conditions�baseline masked by each condition – baseline contrastFrontal
L inf. gyrus –52 32 4 0.002 5.68 0.000 1453(BA 45, 47) –28 32 –10 0.026 5.01
–40 16 –14 0.117 4.53
TemporalL fusiform gyrus –46 –48 –14 0.008 5.34 0.112 252(BA 37)R mid. temp. gyrus 58 6 –14 0.135 4.49 0.063 315(BA 21) 62 –6 –14 0.286 4.21L mid. and inf. temp gyrus –60 –8 –10 0.144 4.46 0.004 670(BA 21, 20) –52 –14 –28 0.185 4.38
–58 –10 –2 0.408 4.06
Contrast (ii), main effect of word classVerbs–nouns
R substantia nigra 12 –22 –6 0.808 3.64 0.958 10
No effect of nouns–verbs
Contrast (iii), main effect of imageabilityConcrete–abstract
Superior frontal gyrus –16 –64 –2 0.989 3.22 0.988 1(BA 10)
Abstract–concreteCingulate gyrus –18 –42 28 0.598 3.87 0.585 79(BA 31)
Results presented for both the voxel and cluster levels of significance were based on a height thresholdof 0.001. The coordinates are in MNI (Montreal Neurological Institute) space, and the correctedP values, the SPM{t} and the extent of the activation are presented. Multiple peaks within an extent areshown on subsequent lines. L � left; R � right; temp. � temporal; mid. � middle; inf. � inferior. Weinclude Brodmann areas (BAs) of the active regions to assist the reader in quickly determining thelocations of these activations. They are not intended as commentaries on the underlying cytoarchitectureof the active areas.
This model overcomes the binormal assumption of theclassical parametric methods and can then be generalized toany statistical score, and it is more flexible than non-parametric models (Genovese et al., 1997) since it does notrequire a minimal number of replications. Therefore, thismodel is directly applicable in our context using either the tor the F SPMs. Additional details are given in the Appendix I.
The data (provided by the t maps given in SPM) werefitted with a mixture of two t distributions representing thet values of individual voxels under the null hypothesis (H0)and under an active hypothesis (H1). The algorithm providesus with the mixing proportion (the estimated proportion ofactivated voxels) and effect size, which best fit the data.These were estimated using a maximum likelihood proceduredescribed by Gustard and colleagues (Gustard et al., 2000).
For the effect of words relative to the baseline, wefound λ � 0.0545 and δ � 2.6466, i.e. the mean effectsize elicited an ~2.65% change in regional cerebral bloodflow (rCBF) and 5.5% of voxels were activated in thiscondition. (For each analysis, we fit the mixture model tothe SPM output relevant for that contrast. For example,
for the analysis of word class contrasts, the data enteredinto the model were the masked analyses reported earlier.)The detection power was of 60% at a false positivesfraction of 0.008. This point was detected automaticallyin order to maximize the weighted sum of true positivesand true negatives in the SPM. For the verb–noun contrast,we obtained values of λ � 0.1 and δ � 1.9. Consequently,the power deduced using the same criterion as above waslower (42%) with a higher false positives fraction (0.02).Interestingly, when this technique was applied to the nounsversus verbs contrast, the minimization algorithm failed tofind a mixture which fitted the histogram. In other words,no significant alternative distribution was found, thusconfirming that there are no significant differences inneural activation as a function of word class. One potentialproblem with these analyses is that we might not havesufficient statistical power to detect small differences. Wethink that it is unlikely that lack of power can explainour findings since the number of degrees of freedom(subjects, sessions) was sufficiently high to ensure reliableperformance of our signal detection algorithm.

1626 L. K. Tyler et al.
Table 3 Sample stimuli for the semantic categorization task
Cue 1 Cue 2 Cue 3 Target Response
Semantic categorizationbucket basket bin TUB Samewrithe slither wriggle SQUIRM Sameaisle alley lane BADGE Differentradiate glisten dazzle SPIN Different
Letter categorizationaaaaaa aaaaa aaaa AAAAAA Samesssss ssss sssssss SSS Samelllll llllll lll YYYYY Differentddd ddddddd dddddd RRRR Different
Experiment 2: semantic categorization taskMethodsTwo tasks were used in this experiment, one for test conditionsand the other for baseline conditions. For the test conditions,we used a semantic categorization task in which subjectsread three cue words presented one after another on acomputer screen and then made a decision as quickly aspossible about whether a fourth (target) word belonged tothe same category as the cue words. Cue words were inlower case and the target words were in upper case to signalto the participants when to make a response. For instance,subjects made a ‘same’ response to the target OTTER whenit followed the three cue words ‘dolphin, seal, walrus’ bypressing the left mouse button, and a ‘different’ response toCUP when it followed ‘moccasin, sandal, boot’ by pressingthe right mouse button (see Table 3). The mouse was alwaysheld in the subject’s right (dominant) hand and both reactiontimes and accuracy were recorded.
For the baseline task, we used a letter categorization taskwhich shared the same stimulus and response characteristicsas the test task, but had no lexical or semantic component.Instead, subjects were presented with three strings of letters,matched in length to the word stimuli, and were askedwhether a fourth string, in upper case, was made up of thesame letters. For example, ‘fffff, fff, fffffffff, FFFFFF’constituted a ‘same’ trial and ‘ttttt, ttttttt, tttt, HHHH’ was a‘different’ trial. Again, subjects signalled their response bypressing either the left or the right mouse button.
Since Experiment 1 established that imageability did notsignificantly affect activations, in this study we decided tomanipulate word frequency rather than imageability. Wordfrequency is another major variable reputed to affect lexicalprocessing in a wide range of psycholinguistic tasks (e.g.Rubenstein et al., 1970). We constructed four sets of stimuli:high frequency nouns, high frequency verbs, low frequencynouns and low frequency verbs. The test stimuli were matchedacross conditions for imageability and letter length. As inExperiment 1, verbs in the two verb conditions had a muchhigher frequency as verbs than as nouns, and stimuli for thetwo noun conditions were much more frequent as nouns thanverbs. (We pre-tested the quadruplets to ensure that the
component words were equally semantically related acrossthe four conditions. In the pre-test, 15 subjects were presentedwith each quadruplet and asked to rate them for their semanticrelatedness on a scale of 1–7, where 1 was very unrelatedand 7 was highly related. Subjects were also asked to circleany word which they thought did not fit into the set.) Thus,we used a 2 � 2 design with word class and frequency asthe main variables. Stimuli statistics are described in Table 4.The items in the baseline letter categorization task werematched on letter length with those in the semanticcategorization task.
The experiment was first run in a pilot study outside thescanner in order to determine presentation rates and durationssuch that the task was challenging for participants but allowedfast and accurate responses. As a result of this pre-testing,in the final version of the experiment each cue word (orletter string) was displayed on a computer screen usingDMDX software (Forster and Forster, 1999) for 200 ms witha 400 ms inter-stimulus period. The target word (or letterstring) was also presented for 200 ms and response timeswere measured from target onset. There was a 1750 ms delayfollowing the target word to allow time for participants tomake a response. Thus each trial lasted 3750 ms.
The eight subjects participated in twelve 90-s scans, eightof the semantic categorization condition and four of the lettercategorization condition. To coincide with the critical periodof tracer uptake (Silbersweig et al., 1993), subjects received45 s of stimuli (12 trials) followed by a blank screen for theremaining 45 s of the scan, during which they were askedto relax and clear their mind. The semantic and letterconditions were presented systematically such that no subjectsaw the conditions in the same order.
ResultsBehavioural dataResponse times were recorded and analysed in the same wayas in Experiment 1. Subjects’ responses to ‘same’ (meanresponse time � 581 ms) and ‘different’ trials (mean responsetime � 579 ms) were not statistically different (F � 1) andfew errors were made in either type of trial (4.5 and4% errors, respectively). Analyses of inverse transformedresponse times were performed on all stimuli (both same anddifferent trials). There was a reliable effect of task onresponse time, with letter decision (mean � 527 ms) fasterthan semantic decision [mean � 703 ms; F2(1,140) � 309.63,P � 0.001]. There was also a reliable effect of word classin the semantic decision task, with nouns (mean � 662 ms)faster than verbs [mean � 747 ms; F2(1,94) � 11.84,P � 0.01]. There was no effect of frequency (F � 1) orinteraction between word class and frequency (F � 1). Theseresults indicate that the letter decision task was easier toperform than the semantic decision task, consistent withsubject reports and previously reported data (Devlin et al.,2001). Consistent with the first experiment, the verb items

PET studies of noun and verb processing 1627
Table 4 Statistics for the four sets of stimuli in the semantic categorization task
Scan condition SR Word length Imageability CELEX frequency
Mean SD Mean SD Noun Verb
High frequency nouns 5.6 5.5 0.9 523 28 111Low frequency nouns 5.4 5.4 0.7 531 32 26High frequency verbs 5.6 5.3 0.9 516 17 89Low frequency verbs 5.8 5.6 0.9 520 28 21
SR � semantic relatedness.
were slightly more difficult than the noun items, even thoughthe conditions were carefully matched.
Imaging dataThe imaging data were analysed in the same way as for theprevious study. Translation and rotation corrections did notexceed 3 mm and 1.5°, respectively for any of the participants.No scans were lost from the eight subjects, yielding 76degrees of freedom. All results are reported at a P � 0.05level after correcting for multiple comparisons (Worsley,1996). The following contrasts were used in the data analysis.
(i) Regions common to all semantic categorizationconditions in comparison with letter categorizationconditions. This was effected by masking the main effect ofthe semantic conditions versus baseline with each of thesemantic conditions individually versus baseline. Thiscontrast is: (HN � LN � HV � LV) – B masked with(HN – B), (LN – B), (HV – B), (LV – B), where regions ofthe main effect are only considered if they surpass P � 0.05in each of the masking contrast images (H � high frequency;L � low frequency; N � noun; V � verb; and B � baseline).
(ii) Main effect of word class: to look for effects of verbsrelative to nouns, we contrasted the two verb categories(HV � LV) with the two noun categories (HN � LN) andmasked this contrast with the four simple contrasts (HV–HN, HV–LN, LV–HN, LV–LN) and the two baseline contrasts(HV–B, LV–B). We carried out comparable analyses to lookfor effects of nouns relative to verbs.
(iii) Main effect of frequency: to assess the effects of highfrequency relative to low frequency words, we contrasted thetwo high frequency categories (HV � HN) with the two lowfrequency categories (LV � LN) masking with the foursimple contrasts as for the word-class effects described above.Comparable analyses were carried out to investigate therelative effects of low frequency words relative to highfrequency words.
(iv) Cross-over interactions between word class andfrequency: (HV � LN) – (HN � LV) and the reverse.
(v) Other interactions between word class and frequency:HV – (LV � HN � LN), LV – (HV � HN � LN), HN –(HV � LV � LN), LN – (HV � LV � HN). This collectionof contrasts allowed us to consider overall effects of the
experimental conditions versus baseline, main effects of wordclass and frequency, and interactions between them.
The semantic categorization task relative to baselineproduced three areas of reliable activity, one in the rightinferior frontal gyrus, another in the left middle temporalgyrus and a massive left perisylvian region extending fromthe inferior frontal gyrus to the inferior temporal gyrus (seeTable 5 and Fig. 2). Within this large region, there weredistinct peaks in the left inferior frontal gyrus and inferiortemporal gyrus. All of these regions of activity were reliableat both voxel and cluster levels after correction for multiplecomparisons.
In the verb–noun contrast, there was a single area ofactivation in the right inferior temporal gyrus. However, itdisappeared when masked by the appropriate contrasts (seeImage analysis above), as the area was not active in all theverb conditions. Similarly, there were no areas more activefor all nouns than verbs. In the frequency analyses, therewere also no areas more active for all high versus all lowfrequency words, or vice versa. In the high frequency nounsminus other test conditions contrast, there was a trend towardsignificant activation in a region of the left middle temporalgyrus. This activity remained after masking the contrast witheach of the high frequency minus other conditions contrasts.Thus the region shows an interaction where effects of wordclass are modulated by frequency.
As with the first experiment, we carried out a poweranalysis to determine whether the task was sufficientlysensitive to detect differences in activation. For the maincontrast of words relative to the baseline, we found λ �0.102 and δ � 3.17. Again, this means that the averageeffect size was a ~3.17% change in rCBF and 10% of voxelswere activated in this condition. The detection power washigh (80%) at a false positives fraction of 0.01. For theverbs–nouns contrast, the algorithm converged with λ �0.0944 and δ � 2.97. The power deduced using the samecriterion as before was slightly lower (76.5%), with a falsepositives fraction of 0.012. When the algorithm was appliedto the nouns–verbs contrast, the minimization proceduresuccessfully converged, this time with a mixing proportionof 0.0664 and an effect size of 2.5. The power correspondingto the chosen reliability criterion was of 58% at a falsepositives fraction of 0.01. These results confirm that the

1628 L. K. Tyler et al.
Fig. 2 The areas activated in the semantic–baseline contrasts in the semantic categorization task (Experiment 2). Activations are shownsuperimposed on a structural image. Red and yellow areas were reliably (P � 0.05) active after statistical correction.

PET studies of noun and verb processing 1629
Table 5 Brain areas of activity in the semantic categorization task
Region Coordinates Voxel level Cluster level
x y z Pcorrected t Pcorrected Extent
Contrast (i), test conditions�baseline masked by each condition�baseline contrastFrontal and temporal
L inf. frontal gyrus –42 16 12 0.000 7.30 0.000 6171(BA 44, 45, 47)
L inf. temp. gyrus –44 –24 –30 0.000 7.22(BA 20) –46 30 4 0.000 6.98
L mid. temp. gyrus –62 –38 –2 0.008 5.42 0.006 560(BA 21)R inf. frontal gyrus 60 26 20 0.008 5.43 0.160 202(BA 45)
Contrast (ii), main effect of word classVerbs–nouns
BA 20/37 –48 –44 –26 0.649 3.87 0.803 42
Nouns–verbs: no effects
Contrast (iii), looking at the main effect of frequency†
Contrast (iv), non-cross-over interactionsHigh frequency nouns – all others, masked with high frequency nouns – each other conditionTemporal
L mid. temp. gyrus –62 –22 –10 0.098 4.66 0.056 308(BA 21)
Results presented for both the voxel and cluster levels of significance were based on a height thresholdof 0.001. The coordinates are in MNI space, and the corrected P values, the SPM{t} and the extent ofthe activation are presented. Multiple peaks within an extent are shown on subsequent lines. L � left;R � right; temp. � temporal; mid. � middle; inf. � inferior. †There were no significant effects of highversus low frequency or vice versa.
second experiment provided more robust activation and wasthus more powerful than the first experiment for all theanalysed contrasts.
Analyses comparing Experiments 1 and 2Comparing Tables 2 and 5, we see that the semanticcategorization task generally produced more robustactivations with larger extents than the lexical decisiontask. To investigate statistically areas of overlap betweenExperiments 1 and 2, we analysed both data sets together.When the lexical decision–baseline contrast of Experiment 1and the semantic decision–baseline contrast of Experiment 2are analysed together, there are five reliable areas of activity.All of these areas are in the left hemisphere and are reliableat the voxel level (cluster analyses are not possible becausethe data from the two experiments do not have the sameintrinsic smoothness) after correction for multiple com-parisons. These regions are in the fusiform gyrus, inferiortemporal gyrus, temporal pole, uncus and inferior frontalgyrus (see Table 6). These regions are largely the intersectionof those of Experiments 1 and 2. Notably, there are noreliable effects of word class across both experiments.
DiscussionIn the two PET studies reported here, we found robustactivation in the left prefrontal cortex and inferior temporallobe, associated with semantic processing of written words.These areas have been reported in previous studies of semanticprocessing and are thought to constitute a network which isactivated when subjects are engaged in a variety of tasksinvolving semantics (Price et al., 1994; Rumsey et al., 1997).
Effects of lexical variablesWe found no significant differential activation as a functionof the two lexical variables—imageability and frequency—when we corrected for multiple comparisons. This suggeststhat the neural representation of conceptual knowledge is notspecialized as a function of these variables; for example,there is no cortical region which is specialized for abstractwords or high frequency words. This makes sense when oneconsiders that the variables of frequency/familiarity andimageability are continuous rather than discrete and thus donot lend themselves to principles of categorical organization.While it might be simple to categorize a highly abstract orconcrete word, many words are of medium imageabilityaccording to the norms that are used typically. It is more

1630 L. K. Tyler et al.
Table 6 A conjunction of the lexical decision–baseline contrast of Experiment 1 and thesemantic decision–baseline contrast of Experiment 2
Region Coordinates Voxel level Cluster extent
x y z Pcorrected t
Contrast (i): a conjunction of the lexical decision–baseline contrast of Experiment 1 and the semanticdecision–baseline contrast of Experiment 2, masked by each of the test condition–respective baselinecondition contrasts
Frontal and temporalL inf. frontal gyrus –50 30 4 0.000 5.80 1373(BA 45, 47) –30 30 –10 0.000 4.76
–38 20 –16 0.000 4.15L fusiform gyrus –44 –46 –18 0.000 4.39 65(BA 37)L. inf. temp. gyrus –48 –18 –34 0.000 3.91 204(BA 20) –36 –14 –32 0.001 3.79
–48 –8 –22 0.093 2.92L temp. pole –34 0 –46 0.001 3.66 31(BA 20, 38)
Contrast (ii): main effect of word class*
The results are presented for the voxel level of significance based on a height threshold of 0.001. Thecoordinates are in MNI space, and the corrected P values, the SPM{t} and the extent of the activationare presented. Multiple peaks within an extent are shown on subsequent lines. *No significant effectswere found.
difficult to see how these words would be categorized ifthe neural representation of conceptual knowledge wereorganized by imageability.
Previous studies have reported additional activation inright inferior frontal gyrus for abstract words (Beauregardet al., 1997; Kiehl et al., 1999), challenging earlier cognitiveclaims for the differential representation of abstract andconcrete words based largely on split visual field experiments,which propose that abstract words are only represented inthe left hemisphere while concrete words are representedbilaterally (e.g. Paivio, 1991). However, left hemisphereactivation (in inferior prefrontal cortex) found in imagingstudies typically is associated with increased processingdemands (Buckner et al., 2000), and there is evidence fromcognitive studies that abstract words may be more difficultto process than concrete words. For example, in lexicaldecision tasks outside the scanner, abstract words generateslower latencies and more errors than concrete words (James,1975; Kroll and Merves, 1986). Thus, the additionalprocessing in the right prefrontal cortex associated withabstract words may be due to the extra demands involved inprocessing these words. This is consistent with thebehavioural data from the imaging study of Kiehl andcolleagues which showed longer latencies for abstractcompared with concrete words (Kiehl et al., 1999). In ourlexical decision PET study, there was no difference in theresponse latencies of abstract and concrete words, and noadditional activation in right prefrontal cortex, suggestingthat when abstract and concrete words are matched forprocessing difficulty, there is no differential activationassociated with processing one or other type of stimulus.
This in turn suggests that differential activation found inright inferior frontal gyrus is associated with processingdifficulty.
Our data suggest that there is no neural specialization forabstract and concrete words. How does this comport withthe claim that deep dyslexic patients, who typically havedamage to left inferior frontal cortex, have greaterimpairments for abstract words compared with concretewords (e.g. Franklin, 1989)? We have argued (Tyler et al.,1995) that abstract word deficits in deep dyslexic patientstend to be found only when patients are required to make averbal response. When tasks are used which do not requirelanguage production, the disadvantage for abstract wordstends to disappear. For example, we tested two classic deepdyslexic patients, with extensive left inferior frontal damage,who showed the typical pattern of greater problems in readingaloud abstract than concrete words (Tyler et al., 1995). Thesesame patients were then tested on a semantic priming taskwhere we varied the imageability of the word pairs. Theword pairs were either concrete (street–road) or abstract(luck–chance) synonyms. The patients showed significantsemantic priming for both the abstract and concrete words,supporting the claim that their difficulties with abstract wordswere restricted to tasks requiring a verbal output. Theassociation of phonological output deficits with damage toleft inferior frontal cortex is consistent with claims for therole of this region in phonological processing.
Effects of word classThe main aim of our imaging studies was to determinewhether processing the meaning of nouns and verbs is

PET studies of noun and verb processing 1631
associated with activation in different cortical regions. Wefound no consistent evidence for the differential effects ofword class; both nouns and verbs activated areas in leftinferior frontal and temporal cortex. Thus, although ourbehavioural data and a number of cognitive studies suggestthat nouns are processed more readily than verbs, thisprocessing advantage does not appear to be correlated withdifferential neural activation. That is, differences in theacquisition and processing of nouns and verbs do not seemto be correlated with differential neural representation.Moreover, the power analyses showed that our experimentalparadigms, especially the semantic categorization task, weresufficiently sensitive to have detected small differences inactivation if they had been present.
This set of results is consistent with those cognitiveaccounts which postulate that the conceptual system isrepresented within an undifferentiated distributed networkwith no explicit category or domain structure. Indeed, ourrecent conceptual structure model (Durrant-Peatfield et al.,1997; Tyler et al., 2000) makes the explicit prediction that,to the extent that the cognitive architecture is reflected in theneural architecture, there will be no regional specializationfor different categories/domains of conceptual knowledge.This prediction is supported by the results of both the currentstudies and other recent PET studies in which we looked forcortical specialization for the domains of living and non-living things (Devlin et al., 2001). In these studies, we found,much as we do in the present studies, an extensive semanticnetwork which was activated by all concepts and no regionalspecialization as a function of domain. These findings arealso compatible with Damasio and Tranel’s claims thatconcepts are not represented in a permanent form in oneneural site, but ‘they depend on many interacting networksthat hold the potential for reactivation of components ofconcepts . . .’ (Damasio and Tranel, 1993).
How does the lack of regional specialization for verbs andnouns relate to the neuropsychological findings that noundeficits are associated with damage to left temporal lobewhereas verb deficits are associated with damage to leftinferior frontal cortex? First, as discussed in the Introduction,there is considerable lack of consistency in these reports.Secondly, many neuropsychological dissociations arerestricted to naming deficits and do not involve impairmentsof conceptual knowledge (e.g. Damasio and Tranel, 1993),thus they do not directly address the issue of how differentcategories of knowledge are represented in the neuralsubstrate. Instead, these impairments may reflect theconsequences of damage to phonological processes (Miceliet al., 1984) or to the processes involved in the mappingbetween phonology and semantics (e.g. a mediation system;Damasio and Tranel, 1993). Thirdly, not all studies of noun–verb dissociations stimuli matched on relevant variables.Thus, the neuropsychological data do not provide unequivocalevidence that the semantic representations of nouns and verbsare located in distinct cortical regions.
Left inferior prefrontal cortexWe found reliable activation in the left inferior frontal cortex,supporting claims that this cortical region is involved insemantic processing (Fiez, 1997; Buchner et al., 2000). Inaddition, the semantic categorization task generated moreactivation in this region than did the lexical decision task.We observed the same pattern in two other PET studies wehave reported recently, comparing activations for wordsdenoting living and non-living things using the lexicaldecision and semantic categorization tasks (Devlin et al.,2001). In the lexical decision task, there was no significantleft inferior frontal activation, in contrast to the semanticcategorization task which generated highly significantactivation across the left inferior frontal cortex. Similarly,using the same semantic categorization task in an fMRIstudy, we also found robust activation in left inferior frontalcortex (Devlin et al., 2000). These findings are consistentwith the proposal that the left inferior frontal cortex is moreactivated in semantic tasks which require effortful, explicitprocessing, or when more semantic information has to beheld in memory while a subject is deciding upon a response(Gabrieli et al., 1998). Our data are less compatible with theview, recently proposed by Thompson-Schill and colleagues,that the left inferior frontal cortex is associated with selectionamongst competing semantic alternatives rather than withretrieval of semantic knowledge (Thompson-Schill et al.,1997). The semantic categorization task, which generatedadditional activation in the left inferior prefrontal cortex, didnot involve selection between alternatives. Instead, subjectshad to decide whether a target word was a member of thesame grouping as three preceding cue words. This involvedaccessing the meaning of each word, constructing anappropriate semantic grouping/category and determiningwhether the target was a member of the same category. Thistask does not involve the activation of multiple competingalternatives followed by a process of selection, but ratherrequires subjects to maintain in memory the meaning of eachword and the category to which they belong. However,instead of the inferior frontal cortex being specialized forone type of cognitive process, it may be the case that it isinvolved in a diverse array of cognitive functions, as hasbeen argued recently (Duncan and Owen, 2000).
ConclusionsThe results of the two PET studies reported here do notsupport the claim that nouns are represented in the temporallobes and verbs in left inferior frontal cortex. Instead, theyare more compatible with the alternative hypothesis that themeanings of nouns and verbs are represented within anundifferentiated cortical network which is not divided bycategory or domain. However, lack of regional specializationdoes not preclude the possibility that specific cortical regionsmay be involved to different degrees in the processing ofsubsets of concepts. These studies focused on the meaning

1632 L. K. Tyler et al.
representations of nouns and verbs. However, nouns andverbs differ most strongly in their syntactic function. Itremains to be seen whether, when the task emphasizes thesyntactic roles of nouns and verbs, we find any evidence ofregional specialization.
AcknowledgementsWe wish to thank Emma Williams at the WBIC, Joe Devlinand Elizabeth Dick for their help with this research. Thisresearch was funded by an MRC programme grant to L.K.T.and W. D. Marslen-Wilson, a McDonnell-Pew grant to L.K.T.and a Wellcome Trust Fellowship to H.E.M.
ReferencesAshburner J, Friston K. Multimodal image coregistration andpartitioning—a unified framework. Neuroimage 1997; 6: 209–17.
Ashburner J, Neelin P, Collins DL, Evans A, Friston K. Incorporatingprior knowledge into image registration. Neuroimage 1997; 6:344–52.
Baayen R, Pipenbrook R. The Celex Lexical Database. Philadelphia:Linguistic Data Consortium, University of Pennsylvania; 1995.
Beauregard M, Chertkow H, Bub D, Murtha S, Dixon R, Evans A.The neural substrate for concrete, abstract, and emotional wordlexica: a positron emission tomography study. J Cogn Neurosci1997; 9: 441–61.
Breedin SD, Saffran EM, Schwartz MF. Semantic factors in verbretrieval. An effect of complexity. Brain Lang 1998; 63: 1–31.
Buckner RL, Koutstaal W, Schacter DL, Rosen BR. FunctionalMRI evidence for a role of frontal and inferior temporal cortex inamodal components of priming. Brain 2000; 123: 620–40.
Caramazza A, Hillis AE. Lexical organization of nouns and verbsin the brain. Nature 1991; 349: 788–90.
Coltheart M. The MRC Psycholinguistic Database. Q J Exp Psychol1981; 33A: 497–505.
Damasio AR, Tranel D. Nouns and verbs are retrieved withdifferently distributed neural systems. Proc Natl Acad Sci USA1993; 90: 4957–60.
Daniele A, Giustolisi L, Silveri MC, Colosimo C, Gainotti G.Evidence for a possible neuroanatomical basis for lexical processingof nouns and verbs. Neuropsychologia 1994; 32: 1325–41.
Devlin JT, Russell RP, Davis MH, Price CJ, Wilson J, Moss HE,et al. Susceptibility-induced loss of signal: comparing PET andfMRI on a semantic task. Neuroimage 2000; 11: 589–600.
Devlin J, Russell R, Davis M, Price C, Moss H, Fadili J, et al. Isthere an anatomical basis for category-specificity? Semantic memorystudies in PET and fMRI. Neuropsychologia. In press 2001.
Dorfman D, Alf E. Maximum likelihood estimation of parametersof signal-detection theory and determination of confidence intervals-rating method data. J Math Psychol 1969; 6: 487–96.
Duncan J, Owen A. Common regions of the human frontal loberecruited by diverse cognitive demands. Trends Neurosci 2000; 23:475–82.
Durrant-Peatfield MR, Tyler LK, Moss HE, Levy JP. The distinctive-ness of form and function in category structure: a connectionistmodel. In: Shafto MG, Langley P, editors. Proceedings of the 19thAnnual Conference of the Cognitive Science Society. Mahwah (NJ):Lawrence Erlbaum; 1997. p. 193–8.
Everitt BS, Bullmore ET. Mixture model mapping of the brainactivation in functional magnetic resonance images. Hum BrainMapp 1999; 7: 1–14.
Fiez JA. Phonology, semantics, and the role of the left inferiorprefrontal cortex [editorial]. [Review]. Hum Brain Mapp 1997; 5:79–83.
Forster KI, Forster JC. The DMASTR display system for mentalchronometry. Tucson (AZ): University of Arizona; 1990.
Franklin S. Dissociations in auditory word comprehension: evidencefrom 9 ‘fluent’ aphasic patients. Aphasiology 1989; 3: 189–207.
Friston KJ, Worsley KJ, Frackowiak RS, Mazziotta JC, Evans AC.Assessing the significance of focal activations using their spatialextent. Hum Brain Mapp 1994; 1: 214–20.
Friston KJ, Ashburner J, Frith CD, Poline J-B, Heather JD,Frackowiak RS. Spatial registration and normalization of images.Hum Brain Mapp 1995a; 3: 165–89.
Friston KJ, Holmes AP, Worsley KJ, Poline J-B, Frith CD,Frackowiak RS. Statistical parametric maps in functional imaging:a general linear approach. Hum Brain Mapp 1995b; 2: 189–210.
Gabrieli JD, Poldrack RA, Desmond JE. The role of left prefrontalcortex in language and memory. [Review]. Proc Natl Acad Sci USA1998; 95: 906–13.
Genovese CR, Noll DC, Eddy WF. Estimating test–retest reliabilityin functional MR imaging. I. Statistical methodology. Magn ResonMed 1997; 38: 497–507.
Gentner D. Some interesting differences between nouns and verbs.Cognit Brain Theory 1981; 4: 161–78.
Gernsbacher MA. Resolving 20 years of inconsistent interactionsbetween lexical familiarity and orthography, concreteness, andpolysemy. J Exp Psychol Gen 1984; 113: 256–81.
Gleitman LR. Words, words, words . . . [Review]. Philos Trans RSoc Lond B Biol Sci 1994; 346: 71–7.
Goodglass H, Klein B, Carey P, Jones K. Specific semantic wordcategories in aphasia. Cortex 1966; 2: 74–89.
Gonnerman LM, Andersen ES, Devlin JT, Kempler D, SeidenbergMS. Double dissociation of semantic categories in Alzheimer’sdisease. Brain Lang 1997; 57: 254–79.
Graesser AC, Hopkinson PL, Schmid C. Differences in interconceptorganization between nouns and verbs. J Mem Lang 1987; 26:242–53..
Gustard S, Fadili MJ, Williams E, Hall LD, Carpenter AT, Bretty M,et al. Effect of slice orientation on reproducibility of fMRI motoractivation at 3T. Magn Reson Imag. In press 2001.

PET studies of noun and verb processing 1633
Huttenlocher J, Lui F. The semantic organisation of some simplenouns and verbs. J Verb Learn Verb Behav 1979; 18: 141–62.
James CT. The role of semantic information in lexical decisions.J Exp Psychol Hum Percept Perform 1975; 104: 130–6.
Kiehl KA, Liddle PF, Smith AM, Mendrek A, Forster BB, HareRD. Neural pathways involved in the processing of concrete andabstract words. Hum Brain Mapp 1999; 7: 225–33.
Kinahan PE, Rogers JG. Analytic 3D image reconstruction usingall detected events. IEEE Trans Nucl Sci 1989; 35: 680–4.
Kroll JF, Merves JS. Lexical access for concrete and abstract words.J Exp Psychol Learn Mem Cognit 1986; 12: 92–107.
Martin A, Haxby JV, Lalonde FM, Wiggs CL, Ungerleider LG.Discrete cortical regions associated with knowledge of colour andknowledge of action. Science 1995; 270: 102–5.
McCarthy R, Warrington EK. Category specificity in an agrammaticpatient: the relative impairment of verb retrieval and comprehension.Neuropsychologia 1985; 23: 709–27.
Metz CE. ROC methodology in radiological imaging. Invest Radiol1986; 21: 720–33.
Miceli G, Silveri MC, Villa G, Caramazza A. On the basis for theagrammatics’ difficulty in producing main verbs. Cortex 1984; 20:207–20.
Miller GA, Fellbaum C. Semantic networks of English. Cognition1991; 41: 197–229.
Mummery CJ, Patterson K, Hodges JR, Wise RJ. Generating ‘tiger’as an animal name or a word beginning with T: differences in brainactivation. Proc R Soc Lond B Biol Sci 1996; 263: 989–95
Mummery CJ, Patterson K, Hodges JR, Price CJ. Functional neuro-anatomy of the semantic system: divisible by what? J Cogn Neurosci1998; 10: 766–77.
Paivio A. Dual coding theory: retrospect and current status. Can JPsychol 1991; 45: 255–87.
Perani D, Cappa SF, Schnur T, Tettamanti M, Collinia S, Rosa M,et al. The neural correlates of verb and noun processing: a PETstudy. Brain 1999; 122: 2337–44.
Petersen SE, Fox PT, Posner MI, Mintun M, Raichle ME. Positronemission tomographic studies of the cortical anatomy of single-word processing. Nature 1988; 331: 585–9.
Petersen SE, Fox PT, Posner MI, Mintun M, Raichle ME. Positronemission tomographic studies of the processing of single words.J Cogn Neurosci 1989; 1: 153–70.
Price CJ, Wise RJ, Watson JD, Patterson K, Howard D, FrackowiakRS. Brain activity during reading: the effects of exposure durationand task. Brain 1994; 117: 1255–69.
Price CJ, Wise RJ, Frackowiak RS. Demonstrating the implicitprocessing of visually presented words and pseudowords. CerebCortex 1996; 6: 62–70.
Rubenstein H, Garfield L, Millikan JA. Homographic entries in theinternal lexicon. J Verb Learn Verb Behav1970; 9: 487–94.
Rubin DC. 51 properties of 125 words: a unit analysis of verbalbehavior. J Verb Learn Verb Behav 1980; 19: 736–55.
Rumsey JM, Horwitz B, Donohue BC, Nace K, Maisog JM,Andreason P. Phonological and orthographic components of wordrecognition: a PET–rCBF study. Brain 1997; 120: 739–59.
Silbersweig DA, Stern E, Frith CD, Cahill C, Schnorr L,Grootoonk S, et al. Detection of thirty-second cognitive activationsin single subjects with positron emission tomography: a newlow-dose H2(15)O regional cerebral blood flow three-dimensionalimaging technique. J Cereb Blood Flow Metab 1993; 13: 617–29.
Skudlarski P, Constable RT, Gore JC. ROC analysis of statisticalmethods used in functional MRI: individual subjects. Neuroimage,1999; 9: 311–29.
Sorensen JA, Wang X. ROC methods for evaluation of fMRItechniques. Magn Reson Med 1996; 36: 737–44.
Thompson-Schill SL, D’Esposito M, Aguirre GK, Farah MJ. Roleof left inferior prefrontal cortex in retrieval of semantic knowledge:a reevaluation. Proc Natl Acad Sci USA 1997; 94:14792–7.
Tyler LK, Moss HE, Jennings F. Abstract word deficits in aphasia:evidence from semantic priming. Neuropsychology 1995; 9: 354–63.
Tyler LK, Moss HE, Durrant-Peatfield M, Levy J. Conceptualstructure and the structure of concepts: a distributed account ofcategory-specific deficits. Brain Lang 2000; 75: 195–231
Ulrich R, Miller J. Effects of truncation on reaction time analysis.J Exp Psychol Gen 1994; 123: 34–80.
Vandenberghe R, Price C, Wise R, Josephs O, Frackowiak RS.Functional anatomy of a common semantic system for words andpictures. Nature 1996; 383: 254–6.
Van Horn JD, Ellmore TM, Esposito G, Berman KF. Mappingvoxel-based statistical power on parametric images. Neuroimage1998; 7: 97–107.
Wagner AD, Desmond JE, Demb JB, Glover GH, Gabrieli JDE.Semantic repetition priming for verbal and pictorial knowledge: afunctional MRI study of left inferior prefrontal cortex. J CognNeurosci 1997; 9: 714–26.
Warburton E, Wise RJ, Price CJ, Weiller C, Hadar U, Ramsay S.Noun and verb retrieval by normal subjects. Studies with PET.[Review]. Brain 1996; 119: 159–79.
Wise R, Chollet F, Hadar U, Friston K, Hoffner E, Frackowiak, R.Distribution of cortical neural networks involved in wordcomprehension and word retrieval. Brain 1991; 114: 1803–17.
Worsley KJ, Evans AC, Marrett S, Neelin P. A three-dimensionalstatistical analysis for CBF activation studies in human brain.J Cereb Blood Flow Metab 1992; 12: 900–18.
Received July 24, 2000. Revised February 2, 2001.Accepted April 5, 2001

1634 L. K. Tyler et al.
Appendix IOverview of ROC methodsEstimation of the receiver operating characteristic (ROC) curve isa well-known methodology for quantifying the detection accuracyof a given statistical test. A ROC curve is a plot of 1 – β versus α,where α and β are, respectively, the type I (false-positive) and typeII (false-negative) error probabilities. The area under the ROC curveis commonly regarded as a good single criterion for characterizingdetection accuracy (Sorenson and Wang, 1996; Skudlarski et al.,1999); the larger the area under the ROC curve, the better thedetection accuracy of the test. ROC curves can be fitted to anobserved data set by assuming certain forms for the probabilitydistributions of the test statistic under the null and alternativehypotheses. The parameters of the distributions in such a finitemixture model can then be estimated by maximum likelihood(Dorfman and Alf, 1969; Metz, 1986, Everitt and Bullmore, 1999).
More formally, the mixture model for the observed test statisticscan be written
f(x) � λfH1 � (1 – λ)fH0
where λ is the mixing proportion (the proportion of truly activatedvoxels); f(x) is the probability of observing a voxel as beingactivated; and fH0 and fH1 are the probability density functions underthe null and the alternative hypotheses, respectively.
If the test statistic is a t statistic, as it is in this study, theseprobability density functions are assumed to be respectively thecentral and non-central t distributions with the same number ofdegrees of freedom; the non-centrality parameter for the probabilitydensity function under the alternative hypothesis is denoted δ.Assuming the voxels are independent, the model parameters λ andδ can be estimated conveniently by maximizing the log likelihoodfunction of the mixture model, for example using a modified Newtonmethod for iterative optimization. For each value of α, we can thencalculate the corresponding power (1 – β) using the alternativeprobability density function parameterized by δ. The ROC curve isthen plotted and the area under the curve is calculated.
Additionally, the ROC curve can be used to identify rationalprobability thresholds for activation mapping. For example, it canbe shown that the threshold which maximizes the fraction of trulyactivated voxels identified as activated (true positive fraction) andthe fraction of truly non-activated voxels identified as non-activated(true negative fraction) is given by the point on the ROC curvewith gradient � (1 – λ)/λ (Gustard et al., 2001). This approach,whereby the key decision concerning which probability thresholdto use for activation mapping is informed by consideration of false-negative as well as false-positive error rates, provides an alternativeto the traditional emphasis in brain mapping on false-positive errorrate as the sole criterion for threshold selection.





























