Embed Size (px)
Citation preview

All
THE MORPHOLOGY OF AZOTOBACTER VINELANDII
GROWN IN DIALYZED SOIL MEDIUM
THESIS
Presented to the Graduate Council of the
University of North Texas in Partial
Fulfillment of the Requirements
For the degree of
MASTER OF SCIENCE
By
Hoda A. Jradi, B.A.
Denton, Texas
August, 1992

Hoda, Jradi A. , The Morphology of Azotobacter
vinelandii Grown in Dialyzed Soil Medium. Master of Science
(Biology), August, 1992, 57 pp, 13 Illustrations, List of
references, 6 Titles.
This research describes the changes in cell
morphology of Azotobacter vinelandii cells cultured in
dialyzed soil medium. This particular culture medium was
assumed to provide the bacteria with an environment similar
to their natural habitat, the soil. Cells were grown in the
medium for 4, 8 and 16 days and fixed with glutaraldehyde
and osmium tetroxide. Sections were cut to a thickness of 60
to 90 nm. Observation of the cells was performed using
electron microscopy. Electron micrographs of cells in young
cultures showed morphological differences from cells grown
in chemically-defined, nitrogen-free media. Electron
micrographs of cells in older cultures revealed the presence
of a cell form not previously described in the literature.
These are cells approximately 0.5 pm in diameter surrounded
by a thick, rigid membrane.

TABLE OF CONTENTS
Page
LIST OF IllUSTRATIONS......................................iv
INTRODUCTION................................................1i
Morphology Pleomorphism and Monomorphism Cyst Formation Conclusion
MATERIALS AND METHODS.....................................13
Cultures and Media Microscopic Observations
Light Microscopy Electron Microscopy Negative Staining
RESULTS.........................18
DISCUSSION...................................................48
LIST OF REFERENCES...........................................54
iii

LIST OF ILLUSTRATIONS
Figure Page
1. Electron micrograph of A. vinelandii grown in dialyzed soil medium for four days showing peritrichous flagellation....................22
2. Light micrograph of A. vinelandii grown in dialyzed soil medium for two days ...........24
3. Electron micrograph of A. vinelandii grown in dialyzed soil medium for four days showing double cell form and peritrichous flagellation-............................26
4. Electron micrograph of A. vinelandii grown in dialyzed soil medium for four days showing long rod shaped cells........................38
5. Electron micrograph of A. vinelandii grown in dialyzed soil medium for four days showing electron dense small cells, ghost cells and large spherical cells-............................30
6. Electron micrograph of A. vinelandii grown in dialyzed soil medium for four days showing rigid limiting membrane......................32
7. Electron micrograph of A.vinelandii grown in dialyzed soil medium for four days showing small cells with thick cell walls ............ 34
8. Electron micrograph of A. vinelandii grown in dialyzed soil medium for four days showing multi-layered cell wall......................36
9. Electron micrograph of A. vinelandii grown in dialyzed soil medium for eight days showing thick limiting edge of the cell, ghost cells and large rod shaped cells............... 38
10. Electron micrograph of A. vinelandii grown in dialyzed soil medium for eight days Showing internal membrane of the cell ................ 40
iv

LIST OF ILLUSTRATIONS--CONTINUED
Figure Page
11. Electron micrograph of A. vinelandii grown in dialyzed soil medium for eight days showing internal structure of the small cell.........42
12. Electron micrograph of A. vinelandii grown in dialyzed soil medium for 16 days showing small spherical cells, large oval cells and bizarre cells....................................44
13. Electron micrograph of A. vinelandii grown in dialyzed soil medium for 16 days showing separation of the cell wall..................46
V

INTRODUCTION
In 1890, Winogradsky's (48) concern with the nitrogen
cycle, specifically the fixation of atmospheric nitrogen,
together with his interest in the existence of
oligonitrophiles, brought him success in isolating
anaerobic, spore-forming bacteria capable of fixing
atmospheric nitrogen which were placed in the genus
Clostridium. The method that he employed depended on the
removal of oxygen from the culture by aerobic organisms,
making it possible for the development of anaerobic ones.
His observations led him to believe that he had also
encountered aerobic oligonitrophiles, but he was unable to
obtain them in pure culture.
In 1901, by applying the techniques used by Winogradsky
in discovering the anaerobic nitrogen-fixing bacteria,
Beijerinck (5) isolated pure cultures of aerobic nitrogen
fixing bacteria that he called azotobacter. These came from
the soils and canal waters of the city of Delft, Holland. He
established the genus Azotobacter with two species,
chroococcum and agile. Following this, the taxonomy and
physiology of these bacteria became a popular subject of
1

2
intensive study that led to an extensive literature and
several lasting controversies. Beijerinck (5) noted the
similarity of characteristics between the two species of the
genus Azotobacter and the blue-green alga which he had
previously studied. Because of this resemblance, Beijerinck
(5) named A. chroococcum after the cyanophytan of the
family Chroococcacea. Jensen (19) and Kyle and Eisentark
(23) disagreed with much of the work reported up to 1950 and
considered Azotobacter a non-pigmented, blue-green alga.
Imshenetski (18) also noted such similarities as
nitrogen-fixation, cell dimensions and structures, division
pattern, and capsule formation between Azotobacter and the
blue-green alga, now cyanobacteria. By 1930, considerable
disagreement surrounded the taxonomic position of these
bacteria, their morphology, role in nature, ecology, and
relationship to the plants.
The description of the genus Azotobacter was surrounded
by confusion as mentioned above, and the list of pleomorphic
types was so lengthy that it was difficult to distinguish
among different forms and also to understand existing
terminology, especially when authors failed to publish
photographic evidence for some of the descriptions (5, 14,
18, 19, 23).
More than sixteen species of Azotobacter have been
proposed by various authors, including Beijerinck's initial
designations. This includes the following: Azotobacter

3
chroococcum, A. agile, A. vinelandii, A. woodstownii, A.
svrmii, A. nigricans, A. araxi, A. lacticogenes, A. insigne,
A. macrocytogenes, and A. paspali. Most of these species
have been disregarded and Bergey's Manual of Systematic
bacteriolocw (37) lists only: A. chroococcum, A.
vinelandii, A. beierinckii, A. nigricans, A. armeniacus, and
A. paspali.
Green and Wilson (14) showed by biochemical analyses
that major similarities between the two species chroococcum
and beiierinckii existed. Moreover, he discovered that A.
chroococcum and A. beilrinckii were practically identical,
but differed significantly from A. vinelandii and A. agile.
These results were confirmed by the work of De Ley and Parks
(12), who studied deoxyribonucleic acid homologies and base
ratio composition in the Azotobacter, and found antigenic
similarities between the two species chroococcum and
beiierinckii. Many differences between Azotobacter species
have been made , sometimes on the basis of cultural
characteristics (5, 17, 18, 20, 23).
The description and morphological characteristics of
Azotobacter are given in the of Bergey's Manual of
Systematic Bacteriology (37) as follows: "Large ovoid cells
1.5-2.0 pm or more in diameter. Pleomorphic, ranging from
rods to coccoid cells. Occur singly, in pairs or irregular
clumps, and sometimes in chains of varying lengths. Do not
produce endospores, but form cysts. Gram negative. Motile

4
by peritrichous flagella, or non-motile. Aerobic, but can
also grow under decreased oxygen tensions. Water-soluble
and water-insoluble pigments are produced by some strains of
all species. Chemoorganotrophic, using sugars, alcohols and
salts of organic acids for growth. Nitrogen-fixers;
generally fix nonsymbiotically at least 10 mg of atmospheric
nitrogen/g of carbohydrate (usually glucose) consumed.
Molybdenum is required for nitrogen fixation but may be
partially replaced by vanadium. Non-proteolytic. Can
utilize nitrate and ammonium salts (all but one species) and
certain amino acids as sources of nitrogen. Catalase
positive. The pH range for growth in the presence of
combined nitrogen is 4.8-8.5; the optimum pH for growth and
nitrogen-fixation is 7.0-7.5. Occur in soil and water; one
species occurs in association with plant roots. The mol% G
+ C of the DNA is 63.2-67.5 (Tm)-"
Morphology
A variety of morphological forms of the cells of
bacteria in the genus Azotobacter have been reported by many
investigators. This is a clear indication that there are
many morphological variations in Azotobacter cells, and it
is obvious that these were deemed of prime importance by
several investigators including Lohnis and Smith (27) and
Bisset and Hale (6). In 1913, Jones (20, 21) reported the
presence of intracellular granules in the Azotobacter life-

5
cycle. He came to the conclusion that the filtrable
inclusions represented reproductive bodies that were
liberated from the mother cell and that these eventually
gave rise to normal Azotobacter cells. Many authors (6, 16,
21, 26, 27, 28, 44, 49) considered the varied morphology of
Azotobacter to be an expression of a complex life-cycle,
with the variation in form representing stabilized stages of
the cycle. The life-cycle they described involved some
thirteen different morphological stages, which included
gonidia (27), microcysts (27, 49), and fungoid forms(44).
Using cultures of A. chroococcum, Lohnis and Smith (27),
also reported the presence of filterable inclusions capable
of regenerating the normal cells of Azotobacter.
In 1922, Almequist (2) reported the finding of granules
in cultures of Azotobacter. He wrote, " We must expect to
find similar forms in our environment, probably most of them
are difficult to culture."
In 1937, Lewis (26) claimed that the cultures studied
in reports of the life-cycle of Azotobacter consisted of
empty cells and fragments of membranes, as well as fat and
volutin granules. Jones (20, 21, 22) also, presented data
which showed that involutions were present in aging
cultures. Bisset and Hale (6) reported on the similarity
between the involution forms observed by Jones and the
endospores of Bacillus. Mellon (31) also described a
variety of similar forms including bodies of six to seven

6
micrometers found in Bacillus coli.
In 1927 Hadley (15) examined the available data with
regard to occurrence and possible significance of the
various forms reported previously (2, 19, 27, 49) and tried
to make a conclusive statement when he wrote, "Whether as
suggested by Mellon, the zygospores are the mother cells of
the filtrable forms of bacteria, for the existence of which
in many species sufficient evidence may now be said to
exist, or whether the filtrable bodies are the micro-gonidia
or the gonites as suggested by Enderlein and Almequist . . .
cannot at present be stated. . . But, whatever the actual
significance of these minute bacterial forms may eventually
prove to be, we may be certain of three things; they occur
regularly, consistently, and in great numbers of many
bacterial cultures under certain growth conditions and at a
certain stage of development; they do not always long endure
as such, but after a brief development often appear
apparently passing into other developmental stages; and
although they may closely resemble certain artificial
structures on the slide, they are not artifacts."
Hadley's (15) conclusive statement did not end the
debate that began in the early days of bacteriology. The
question of the existence of minute forms in the bacterial
life-cycle has been going on since that time. Gonzalez and
Vela (28), reported in 1980 that the descriptions given by
previous investigators are inadequate in that they refer

7
only to one phase of the organism's morphology, and that as
such, do not adequately describe the true nature of these
bacteria.
Monomorphism and Pleomorphism
As early as 1877, Nageli (in Smith and Conant, 36)
proposed that all bacteria belonged to one species and that
there was only one cell type, a highly variable form capable
of passing from one state to another both morphologically
and biochemically. That concept was designated as the
conservative view of monomorphism. Those cells, especially
in old cultures, which did not conform to the normal were
said to be contaminants. The extent and variety of cell
forms (pleomorphism) were not acknowledged as a common
phenomena in the life-cycle of bacteria (36).
Despite the lack of correlation and consistency among
the findings of various workers, many different
descriptions, as mentioned above, have been published of the
genus Azotobacter (3, 8, 13, 20, 21, 22, 23, 26, 28).
Jensen (19) noted that many morphological variations
existed in pure cultures of Azotobacter. He presented
evidence of the existence of rod-shaped or oval cells
measuring about 2 to 4 Lm; spherical cells about 2 to 3 tm
in diameter; very small rod-shaped cells or spherical cells
of less than 1 pn in diameter; cysts; and large cells.
Jensen (19) also assumed that these forms were affected by

8
the culture medium or by its composition. In his work, he
recognized the already known and reported shapes: the oval
rods which were subject to great variation in size;
spherical cells; and the cysts, previously called
"arthrospores" by Jones (20, 21, 22). All reported
morphological variants were generally associated with an
udetermined life-cycle of the bacterium.
Many different cell types occur as discrete stabilized
stages in the growth cycle of the organism (2, 6, 13, 20,
22, 26). Frequently encountered forms included large, non
spore-forming rods; coccoid budding cells; Gram negative
rods; and arthrospores (cysts, microcysts, gonidia, and
spores).
Some investigators have suggested that morphological
variability should be attributed to environmental influences
(20, 21, 30, 39). However, Eisentark et al. (13) postulated
that pleomorphic cells could, in fact, be stages in the
bacterium's life-cycle, and that they could be deteriorating
forms resulting from depletion of nutrients in the growth
medium. Also, that they could be contaminants, or cells
unable to divide or synthesize a cell wall. It is well
known from previous investigators (17, 44, 45, 48) that A.
vinelandii grown in the laboratory produces morphological
variants. When grown in Difco peptone by Vela and
Rosenthal (44), A. vinelandii produced fungoid cells, which
were described as having irregular shapes and no structural

9
rigidity. Vela and Rosenthal (46) assumed that glycine
present in a concentration of 38 to 75 mg/l in Difco peptone
induced the formation of those fungoid cells. They were
described as large, lacking in complete cell division, and
sometimes dividing by budding. Eisentark et al. (13)
observed similar giant cells of A. agile grown on media
containing beef extract or soil extracts. Also, van Shreven
(39) has noted transparent forms of A. chroococcum when
grown on mannitol agar with peptone. These autolyze
spontaneously according to van Shreven.
The Cvst Form
In 1938 Winogradsky (48) succeeded in converting the
vegetative cells of Azotobacter chroococcum into the cyst
form. He used organic acids and alcohols as oxidizable
substrates and obtained an entire population of Azotobacter
in the cyst form. He claimed that the formation of those
cysts might be permanently suppressed when the bacteria were
grown on media containing glucose or sucrose as a carbon
source. Wyss et al. (49, 50) observed that encystment
occurred in vegetative cells of A. vinelandii grown on
Burk's medium without glucose. Those cysts had previously
been described by Omelyanski et al. (31) and Batchinskaya
(3) described the formation of two-layered capsules
surrounding the cells of Azotobacter and found those to be
similar to cysts.

10
Not only Azotobacter, but many other bacteria are
characterized by the ability to form cysts. It is reported
that such cysts are generally smaller than the vegetative
cells. Other bacteria capable of producing cysts in special
media belong to the genera Myxobacter, Cystobacter, and
Methanococcus. The cyst is a protective coat which covers
the entire cell, making it resistant to heat and many other
harsh conditions (9, 38, 46, 47). Cysts are structurally
different and easily distinguished from vegetative cells and
from the endospore of other bacteria (38). Each cyst is the
product of a vegetative cell and possess a thick, double
layered cyst coat (38, 50).
In 1964 Vela and Wyss (42), found laboratory cysts to
be more sensitive to gamma rays than were the soil
Azotobacter in situ in the soil. In 1974 Vela et al.(46)
exposed soils to microwaves, both in the laboratory and in
the field, and found that the soil microflora were more
resistant to microwave energy than were laboratory-grown
cells. Also, they found that Azotobacter became susceptible
to microwaves when they were removed from their natural
habitat.
Notwithstanding the numerous reports of the last eighty
years, there is no evidence that natural cyst formation
occurs in the soil, although cyst formation in response to
organic compounds such as n-butanol is common (45, 47).
Many investigators (1, 4, 38, 41, 49, 50) have studied the

I1
physiology and morphology of cysts in order to understanOd
their functions . In 1974, Vela (47) reported that
Azotobacter could survive for 15 years in dry soil in glass
containers. Later, Moreno et al. (30) reported that
Azotobacter could survive in dry soil in glass containers
for periods of time in excess of 24 years. Despite all the
evidence of the resistance of the laboratory grown cysts,
the survival form of Azotobacter in nature has not been yet
identified.
Conclusion
Relying on previous work, it is valid to say that the
morphology and survival form of bacteria of the genus
Azotobacter is quite dependent on the age of the culture and
growth conditions.
Many different descriptions of the bacteria of the
genus Azotobacter have been published, as well as of the
species vinelandii. The morphology of this bacterium is the
object of this investigation. The research presented here
is designed to elucidate the morphology of these very
interesting organisms when they are grown in a medium
resembling their natural habitat.
The question regarding the form of A. vinelandii is
associated with the growth condition. Small rod shapes and
oval cells observed in nitrogen-free media can be, as
mentioned before, a laboratory artifact. Growth in a

12
chemically defined, nitrogen-free medium is probably quite
different from that which occurs in the soil (28, 45). For
this reason, the question concerning naturally occurring
cells cannot be resolved unless Azotobacter can be directly
observed in their natural habitat.
It has been assumed that microorganisms which survive
in the soil have a morphology quite different from that of
microorganisms grown in the laboratory. There is no
evidence to support the assumption that laboratory grown
organisms resemble in any way those found in nature.
The thrust of this work is to describe the cells of A.
vinelandii as they appear in nature. For this purpose, a
medium composed of soil and water is used on the assumption
that this is more like the natural habitat of A. vinelandii
than is a nitrogen-free medium containing 0.5 to 2.0% (w/v)
glucose.
Soil dialysate supports the growth of A. vinelandii,
and it is evident that the nutrients required for the growth
of this organism are found in soil. The medium used for the
growth A. vinelandii is very likely to resemble the natural
habitat. It is composed of soil chemicals and water (28).
It is assumed that the results of this investigation
will yield a clearer view of the morphology of A. vinelandii
as it exists in nature than what can be obtained by looking
at the literature or at cultures grown in laboratory media.

13
MATERIALS AND METHODS
Cultures and Media
The culture of Azotobacter vinelandii used in this
study was obtained from the stock culture collection at the
University of North Texas. It was maintained on slants of
modified Burk's Agar medium, and periodically checked for
purity and strain confirmation by streaking on Burk's medium
and on nutrient agar.
All cultures used in this study had the same origin,
and all experiments were started with inoculum from cultures
grown on Burk's medium plates incubated at 260C. The
composition of Burk's nitrogen-free medium is given below.
Components Concentration Grams/Liter
Sucrose........................................20.0
KH2PO4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .0.20
K2 HPO4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0.80
MgSO4 .7H20.....................................0.20
CaCI2.7H20......................................0.08
FeSO4 .7H20......................................0.005
Na2MoO4 .2H20...................................0.0003
Water..........................................ILit
pH adjusted to 7.0 wiLh either K2 HPO4 or KH2 PO4
Soil Medium Culture
Soil samples were collected from various locations on

14
the campus of the University of North Texas when the ground
was dry. This was done by removing litter from the soil
surface and taking the exposed soil to a depth of one or two
centimeters. Several samples were collected and combined in
polyethylene bags. In the laboratory, the soils were broken
into fine dust with a mortar and pestle and passed through a
100 mesh sieve. Dialyzed soil medium was prepared by
pouring 15 g of the finely powdered garden soil into pre
boiled and washed dialysis tubing. Dialysis tubes were
closed by tieing the ends into secure knots leaving
sufficient empty space for air expansion when autoclaved.
Tubes with soil were then placed in 25 ml of distilled water
in Erlenmeyer flasks and left to sit for two hours. After
this time, the flasks were sterilized in the autoclave at
1210C for 20 minutes and left to sit for 24 hours.
Dialysis tubes containing the soil where then
discarded, bacterial inoculum from the cultures grown on
Burk's Agar plates was added to the clear dialysate.
Azotobacter vinelandii grown in this clear soil dialysate
was used to study the morphology as the cultures aged.
All cultures were grown on the shaker at 260C. Samples for
microscopic observation were obtained from these cultures as
needed. Before each preparation was made for electron
microscopy, samples of the growing cultures were streaked on
plates of Burk's agar and nutrient agar.

15
Microscopic Observation
Light Microscopy Microscopic observation of A. vinelandii
was performed with a light microscope at 100OX (oil
immersion) in order to confirm the presence of cells in the
culture. These were prepared for oil immersion light
microscopy by obtaining a very thin film of culture between
the slide and the coverslip.
Transmission Electron Microscopy Chemical fixation is the
most extensively used method for preparation of biological
tissues and cells for both transmission and scanning
electron microscopy. Its main purpose is to preserve the
structure of the cell with minimum alteration from the
living state and to prepare the cells for subsequent
treatment, including staining and exposure to the electron
beam.
Employing the recommended method in Introduction to
Biological Electron Microscopy: Theory and Techniques(11),
the cells were collected from the soil dialysate medium,
centrifuged at 3000 rpm for 3 minutes, and pre-fixed with 2
percent glutaraldehyde in 0.2 M cacodylate buffer for 15
minutes. The strength of the fixative was set to minimize
extraction and shrinkage of cells. Also, the use of 0.2 M
cacodylate buffer was mainly to avoid fluctuations in the pH
of the cells and the formation of acidic waves as the

16
fixative entered the cell. Following pre-fixation, the
cells were centrifuged at 3000 rpm for one minute, the
supernatant decanted and the pellet washed three times with
the same buffer. The cells were then embedded in 1.5
percent agar. The resulting agar block was cut into small
pieces with a sharp blade and transferred into a test tube
for post-fixation. A 2 percent concentration of osmium
tetroxide was used as the post-fixative. This preparation
was washed in buffer then dehydrated by passing through 30,
50, 75, 85, 95 and 100 percent aqueous acetone before
embedding in Epon 812.
Epon 812 resin was used in this experiment because it
is more electron transparent and offers a higher contrast
for embedded specimens than other embedding agents. Two
solutions, A and B, were prepared and kept in the
refrigerator until needed:
A B
Epon resin 812 62 ml 100 ml
Hardener DDSA 100 ml (dodecenyl succinic anhydride)
Hardener NMA - 98 ml (nadic methyl anhydride)
Accelerator DMP-30 1.5 to 3% [2,4, 6, tris (dimethyl aminomethyl phenol)
The recommended mixture (solution A and B) is as follows:
Solution A 7 ml

17
Solution B 3 ml
DMP-30 0.15 ml
Sections were cut with an MTC-6000 Ultramicrotome
(Sorvall Instruments, Norwak, Conn.) equipped with a glass
knife. All sections were stained with uranyl acetate (2 g
of uranyl acetate in 20 ml of 50% ethanol) for 15 minutes
and with lead citrate (0.01 to 0.04 g of lead citrate in 10
ml of distilled water and 0.1 ml of ION NaOH) for another 15
minutes and examined with the electron microscope (JOEL
100CX, Tokyo, Japan). Pictures of the bacteria were taken,
using Kodak SO-163 film.
Negative Staining
Negative staining was also used in this study because
it produces a dark background that delimits particulate
specimens. Cells were harvested, fixed, and washed as
described above. The bacteria, which do not interact with
electrons were mixed with phosphotungstic acid which
scatters electrons very effectively. A 2 percent aqueous
solution of phosphotungstic acid (PTA) was adjusted to the
desired pH (4.5 to 7.2) with 5N KOH. The solution was then
filtered (Whatman#1 paper) and stored in the refrigerator.
Prior to its use, the PTA stock was mixed with 0.1 percent
aqueous bovine serum albumin (BSA), to avoid accumulation of
stain and in order to obtain maxamal dispersion of

18
particulate matter including bacterial cells. A thin layer
of the cell suspension with the electron-scattering
substance was mixed and dried on a collodion carbon-coated
grid. Such grids were examined with the electron microscope
(JOEL 100CX, Tokyo, Japan) and pictures of the bacteria were
taken. Kodak SO-163 film was used in making prints.
RESULTS
The data obtained from these studies suggest the
existence of a cell form of Azotobacter vinelandii not
previously recognized. Bergey's Manual of Systematic
Bacteriology describes these bacteria as large, rod shaped
organisms which become extremely pleomorphic in aged
cultures (37). Studies carried out in this laboratory
(Cagle, Chang, Hartnett, Vela) agree in every particular
with the descriptions given in Bergey's Manual. These, and
other published and unpublished studies on the morphology of
A. vinelandii cells were based on cultures grown in
chemically defined, nitrogen-free media.
The objective of this research was to show that cells
of A. vinlandii grown in chemically defined, nitrogen-free
media are probably not representative of this organism as it
exists in nature. Since soil dialyzate is more like the
natural habitat of these organisms (28), it is assumed that
the cells grown in it more closely resemble the bacteria in

19
in nature. Cell morphology could not be determined in cultures
less than four days old since cell densities were not
sufficiently great to make usable pellet. Figure 1 shows
flagellated cells typical of those observed in soil dialyzate
cultures during the first 2 days of growth. The cells in Figure
1 are large, flagellated, and rod shaped. Peritrichous flagella
is evident in the majority of cells and wet mount observation
shows them to be motile. Figure 2 is a light photomicrograph of
cells grown for two days in soil dialysate. Figure 3 also shows
a flagellated cell. This cell is in the double cell form typical
of A.vinelandii. No internal structures are evident in the cells
in figure 1 and 3 because these are negatively stained
preparations. The morphology of four day old cells of A.
vinelandii in soil dialyzate is similar from that described in
Bergev's Manual of Systematic Bacteriology (37).
By the fourth day of culture in soil dialysate, cells become
rounded and non-motile. Those in Figure 4 are seen only rarely
but are shown here to illustrate the transition from long, rod
shaped cells to the spherical form.
Figure 5 shows the typical morphology of cells grown in
dialyzed soil for four days. Arrow number 1 points to the
predominant cell form seen in four day old cultures of A.
vinelandii in soil medium. Arrow number 2 shows a "hole" in the
section. These holes are quite common in preparations of four
day old cultures. The cell indicated by arrow number 3 is
similar to the rounded cell form of A. vinelandii found in soil

20
dialysate and also in chemically defined nitrogen-free media.
The same cells are shown in Figure 6 for the purpose of
illustrating the cell walls, or limiting membranes of these small
cells. At different post-staining times, interior details of the
small cells (Fig.7) becomes evident. A thick, rigid cell wall or
outer barrier was observed. The internal structure of the small
cells was fully revealed making it obvious that these small cells
of 0.25 to 0.75 pm in diameter are fully formed and organized as
are the other cells in these cultures.
The cell in Figure 8 shows a multi-layered cell wall. This
cell wall is characteristic of Gram negative cells and is the
same as seen in cells grown in Burk's chemically defined,
nitrogen-free medium although the morphology of the cell is quite
different.
Figure 9 shows the morphology of A.vinelandii cells grown on
dialyzed soil for eight days. Arrow number 1 points to the
predominant cell form seen in eight day old cultures, it must be
noted that the arrow points directly at the limiting edge of the
cell. This gives the appearance of a thick rigid cell wall.
Other cells in this picture show the same structure. Arrow
number 2 points to "ghost" cells or possibly holes left in the
sections when the cells that were there first fell off or were
pulled off by the microtome knife. The third arrow points to the
cell form not frequently encountered in the eight day old
cultures. Figure 10 illustrates the interior details of cells in
eight day old culture. The arrows point to the internal membrane

21
of the small cells. Figure 11 shows more detailed internal
structures and also the difference between the external membrane
and the inner one. The latter is plainly separated and pulled
away from the outer one (arrow number 3).
By the sixteenth day of culture in soil dialysate, cells of
A. vinelandii tended to be different from those observed during
the first eight days. Cells of widely varied morphology
including spherical and oval shapes (Fig. 12) appeared. Arrow
number 1 in Figure 12 points to an oval cell representative of a
large part of cells in older cultures. Arrow number 2 points to
a small cell, similar to the ones observed in young cultures. It
was observed that as the culture ages, the number of small
spherical cells decreases and the number of oval cells increases
proportionately. The third arrow points to a bizarre cell. This
bizarre cell is typical of A. vinelandii grown in dialyzed soil
medium for 16 days.
Figure 13 shows cells of A.vinelandii in 16 day old culture.
These cells are considerably smaller than those seen in Burk's
medium or in dialyzed soil medium during incubation periods of
less than four days. The morphology of these cells appears to be
the same as that of cells grown in Burk's medium. The arrow in
this picture points to a separation between the cell wall and the
cell membrane.

22
FIG. 1. Electron micrograph of negatively stained four day old cells of Azotobacter vinelandii grown in dialyzed soil medium. The micrograph shows peritrichous flagellation. Bar= 0.5 pm.

23
.4
- op
.*....
.

24
FIG. 2. Light microscopy of A. vinelandii cells grown for two days in dialyzed soil medium.

25

26
FIG. 3. Electron micrograph of negatively stained four day old cells of A. vinelandii grown in dialyzed soil medium. Micrograph shows double cell form and flagellation. Bar=0.5 Em.

27
4

28
FIG. 4. Electron micrograph of a thin section (60 to 90 nm) of four day old cells of A. vinelandii grown in dialyzed soil medium. The micrograph shows long rod shaped cells. Bar=0.5 gm.

29
0
*
b w

30
FIG. 5. Electron micrograph of a thin section (60 to 90 nm) of four day old cells of A. vinelandii grown in dialyzed soil medium. Arrow number 1 points to an electron dense small cell of 0.25 pm diameter. Arrow number 2 points to a "hole" in the supporting medium. Arrow number 3 points to a cell of 1.2 pm diameter. Bar=1.0 gm.

31
*.1'
p 3 r

32
FIG. 6. Electron micrograph of four day old cells of A. vinelandii grown in dialyzed soil medium. Arrow number 1 points to the thick cell wall of the small cell. Arrows number 2 and number 3 point to the remains of the limiting membrane left behind when the cell was removed by the microtome knife. Bar=1.0 p m.

33
1
{t

34
FIG. 7. Electron micrograph of a thin section (60 to 90 nm) of four day old cells of A. vinelandii grown in dialyzed soil medium. Arrows point to the thick rigid cell wall. Bar=1.0 pim.

35
2
)4..
4 4$
I
* sri
tr
ej.
s

36
FIG. 8. Electron micrograph of a thin section of a four day old cell of A. vinelandii grown in dialyzed soil medium. Arrow points to the multi-layered cell wall. Bar=1.0 m.

37

38
FIG. 9. Electron micrograph of a thin section (60 to 90 nm) of eight day old A. vinelandii cells grown in dialyzed soil medium. Arrows number 1 point to the thick limiting edge of the predominant cell type. Arrows number 2 point to the "ghost" cell. Arrow number 3 points to a cell form not frequently encountered. Bar=1.0 pm

39
p .70,
U 4

40
FIG. 10. Electron micrograph of a thin section (60 to 90 nm) of eight day old cells of A. vinelandii grown in dialyzed soil medium. Micrograph shows internal structures of the cells. Arrows point the internal membrane of the cell. Bar=l.0 Lm.

41
0wo

42
FIG. 11. Electron micrograph of a thin section (60 to 90 nm) of eight day old cells of A. vinelandii grown in dialyzed soil medium. Micrograph shows internal structures of the cells. Arrow number I points to the external membrane of the cell while arrow number 2 points to the internal membrane of the cell. Arrow number 3 points to a separation space between the two membranes. Bar=1.Q gpm.

43
6*
0

44
FIG. 12. Electron micrograph of a thin section (60 to 90 nm) of 16 day old cells of A. vinelandii grown in dialyzed soil medium. Arrow number 1 points to an oval cell, representative of a large population of cells in the culture medium. Arrow number 2 points to a small spherical cell. Arrow number 3 points to a "bizarre" cell. Bar=1.0 pm.

45
.4p4 LMr to401
i) *AI6
Mun0i.

46
FIG. 13. Electron micrograph of a thin section (60 to 90 nm) of 16 day cells of A. vinelandii grown in dialyzed soil medium. Arrow points to the separation of the cell wall.Bar=0.25 gm.

47
t
id

48
DISCUSSION
This study was designed to observe the cell morphology
of Azotobacter vinelandii grown in dialyzed soil medium.
Observations of the cells were performed using electron
microscopy. Cells of A. vinelandii were grown in the soil
medium for a period of 4, 8 and 16 days. Glutaraldehyde and
osmium tetroxide were used as fixatives following the
procedure described in the Materials and Methods. Sections
were cut to a thickness of 60 to 90 nm and observed under a
transmission electron microscope.
Extensive investigation has been focused on the
morphology.of A. vinelandii cells. Despite this fact, only
very few studies were concerned with the cell morphology of
this organism in its natural habitat, the soil. Since soil
dialysate is more like the natural habitat of the organism
(28), it is assumed that the cells grown in it are similar
to those of the bacteria in nature.
The micrographs presented here show morphological
differences between the cells grown in chemically defined
nitrogen-free media and those grown in the soil medium. It
must be noted however, that no other known type of
microscopy has ever led to reports of cells like the ones
encountered in this study.
Negative staining of cells from a four day old culture
of A. vinelandii, showed peritrichous flagellation and

49
morphology similar to that of cells grown in chemically
defined media (Figs. 1, 3). These cells also fit the
description of A. vinelandii given in Bergey's Manual of
Systematic Bacteriology (37). Thin sectioning of the cells
revealed morphological characteristics also consistent with
those of cells grown in chemically defined nitrogen-free
medium and described in Bergev's Manual.
Figure 5 shows that the majority of cells were small
and spherical measuring some 0.25 to 0.75 .m in diameter.
These are light refractile (16) and also refractile to
electrons in the electron transmission microscope. They are
the predominant cell form in four day old cultures and the
size is fairly constant in the range from 0.25 to 0.35 pn.
Many electron micrographs show a separation between the
outer wall (cell wall) and inner limiting membrane (cell
membrane). It appears that the outer wall is hard (Figs. 6,
7, 9, 10, 11), and probably complex in both chemical and
structural composition (Fig. 13). It is not difficult to
imagine that these are the survival forms of the Azotobacter
in nature since they meet the criteria described by Vela in
1964 (40), Vela and Wyss in 1965 (42), and Lopez-Gonzalez
and Vela in 1981 (28). The small round cells seen at four
days in soil dialysate cultures continue to grow and change
as previously described (Figs. 4, 5, 6, 7, 9, 10, 11, 13).
By the eighth day of culture, the average cell size ranges
from 0.5 to 0.75 pm and the cells appear elongated and

50
pleomorphic (Figs. 9, 10, 11).
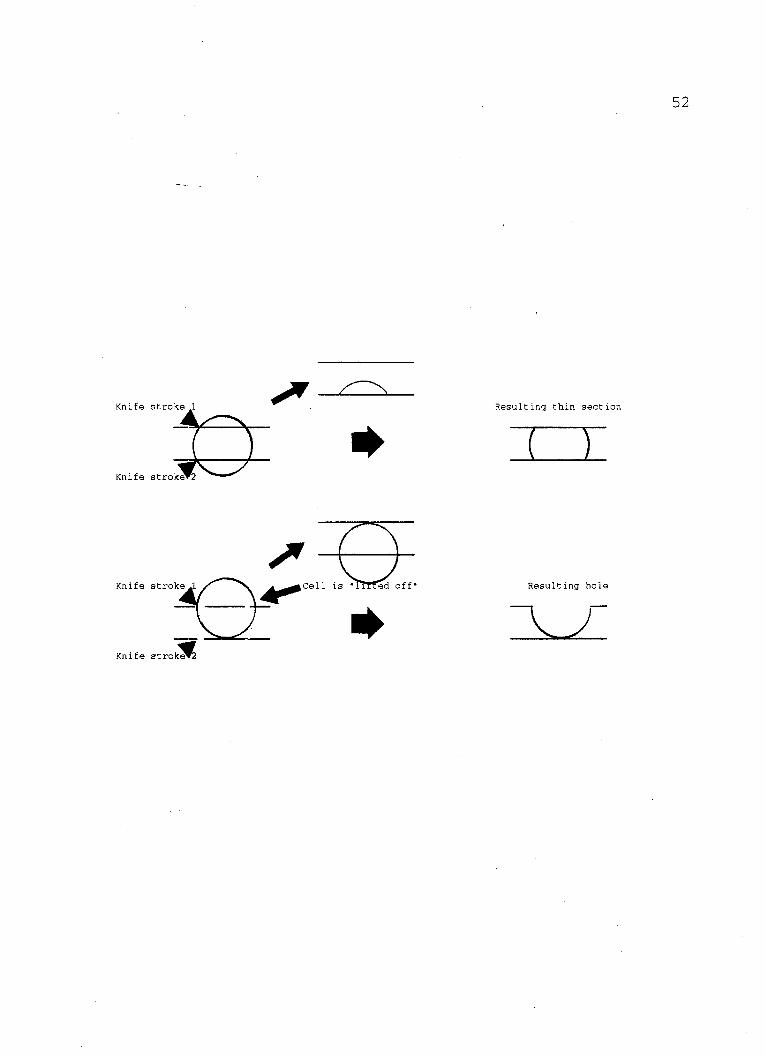
Holes (Figs. 5, 6, 9) in the sections were frequently
encountered in the four day old cultures as well as in the
eight day old cultures. These are cell-shaped holes which
often contain fragments of cell material attached to their
periphery (Fig. 9). The constancy of this observation
strongly suggests that such holes are the result of cells
"lifted off" by the microtome knife and removed from the
embedding material leaving a hole in the section. All the
holes observed were the exact shape of the cells present in
the micrographs examined; an observation which supports the
idea that the cells were "bumped" out of the supporting
medium because they were too hard for the glass knife.
Since some cells were sectioned, it must be assumed that the
glass knife could cut them into sections but only under
certain conditions. The following sketch helps to explain
this:
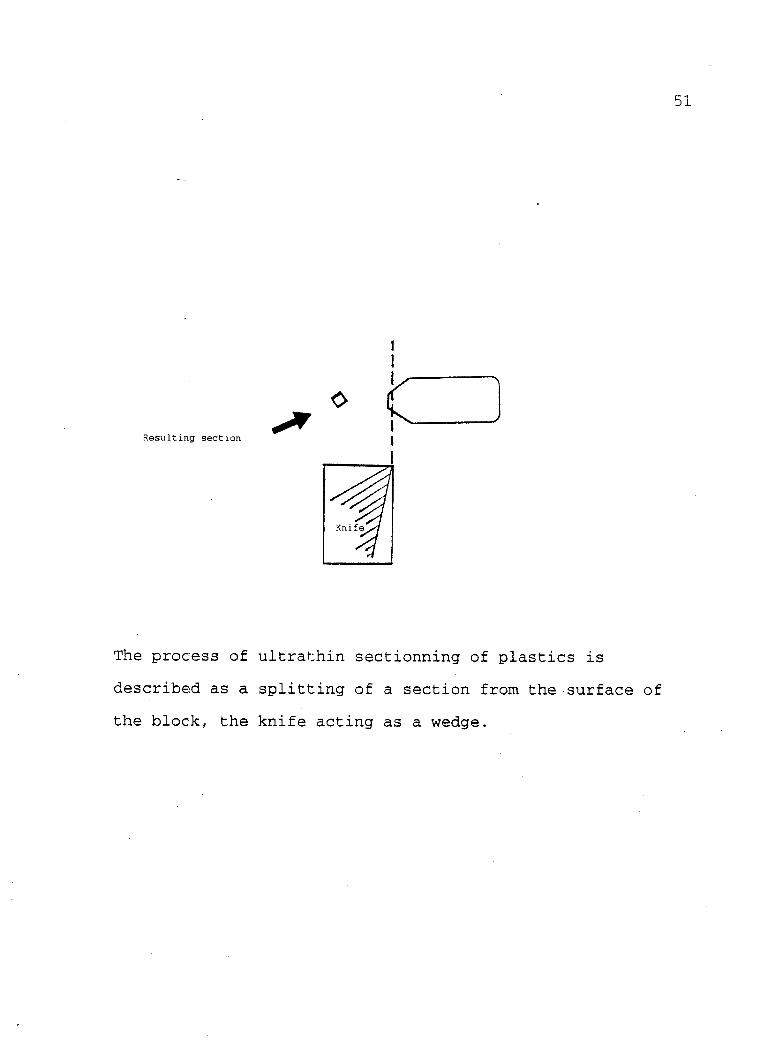
51
Resulting section
The process of ultrathin sectionning of plastics is
described as a splitting of a section from the surface of
the block, the knife acting as a wedge.
Knife
Knife stroke 1
Knife stroke 2
Knife stroke 1 Cell is I ed off*
Knife stroke
Resulting thin section
Resulting hole
52

53
Micrographs of the eight day old cultures showed the
small round cells observed in the four day old cultures and
also larger cells similar to those grown in chemically
defined, nitrogen-free media. The outer wall of the small
round cells appeared thinner and less. This implies that he
outer wall is not as hard as at four days.
Micrograph of 16 day old cells showed spherical and
oval cells. These cells did not vary much in size and
ranged between 0.75 and 1.5 ptm in diameter. It was noted
that as the culture aged, the number of small spherical
cells seen in the younger cultures decreased. When sizes
were compared, it was noticed that the small cells with
dense outer walls were approximately 3/4 the size of large
cells. The large cells are similar to those grown in
chemically defined media. It is assumed that the small
cells observed earlier continue to grow in the presence of
water and eventually become full grown, mature cells. One
can assume that in nature, the small cells would remain as
such in the absence of water but would continue growing in
its presence.
The objective of this work, to examine the morphology
of A. vinelandii in a soil medium, was accomplished. A new
cell form was described for the first time although its
existence had been postulated by Vela (45, 47), Vela and
Wyss (41, 42) and Lopez-Gonzalez and Vela (28).

LIST OF REFERENCES
1. Aladegbami, S.L., J.C. Tsai, and G.R. Vela. 1979. Adenylate energy charge of Azotobacter vinelandii during encystment. Curr. Microbiol. 2:327-329.
2. Almquist, E. 1922. Variation and life cycles of pathogenic bacteria. J. Infect. Dis. 31:438-494.
3. Batchinskaya, A.A. 1935. Sur la structure et le development de l'Azotobacter. Bull. Inst. Microbiol. Agr. 6:1-47.
4. Beamen. B.L., C.E. Jackson and D.M. Shankel. 1968. Formation of multiple central bodies in giant cyst of Azotobacter vinelandii. J. Bacteriol. 96:266269.
5. Beijerinck, M.W. 1901. On oligonitrophilic microorganisms. Milestones in Microbiolocfv, edited by T.D. Brock, Englewood Cliffs, New Jersey, Prentice Hall Publishing Co.
6. Bisset, K.A., and C.M.F. Hale. 1964. The cytology and life cycle of Azotobacter chroococcum, J. Gen. Microbiol. 23:442-448.
7. Boylene, C.W., and J.C. Enision. 1970. Long term starvation survival of rod and spherical cells of Arthrobacter crystallopoites. J. Bacteriol. 103: 569-577.
8. Breed, R.S., E.G.D. Murray, and N.S. Smith, editors. 1957. Bergey's Manual of Determinative Bacteriology. Williams and Wilkins. Baltimore, Md.
9. Cagle, G.D., and G.R. Vela. 1971. Giant cysts and cysts with multiple central bodies in Azotobacter vinelandii. J. Bacteriol. 109:1191-1197.
10. Chang, C.C. 1976. Unpublished notes. Serological reactions of azotobacter species.
11. Dawes, C.J. 1990. Introduction to Biological Electron Microscopy: Theory and Techniques. Ladd Research Industries, Inc. Burlington, Vermont.
54

55
12. Difco Manual of Dehydrated Culture Media and Reagents for Microbiological and Clinical Laboratory Procedures. 1990. Difco Laboratories Inc., Detroit, Mi.
13. Eisentark, A., C.B. Ward, and T.A. Kyle. 1950. A study of large bodies in Azotobacter agile. J. Bacteriol. 60:525-531.
14. Green, M., and P.W. Wilson. 1935. Utilization of nitrogen by the azotobacter. J. Gen. Microbiol.9:89-96.
15. Hadley, P. 1927. Microbic dissociation. Journal of Infect. Dis. 40:1-311.
16. Hartnett, G.H. 1984. Unpublished notes. Morphology of Azotobacter vinelandii grown on nutrient agar.
17. Holt, S.C., and E.R. Leadsetter. 1969. Comparative ultrastructure of selected aerobic spore-forming bacteria. A freeze-etching study. Bacteriol. Rev. 33:346-378.
18. Imshenetski, A.A. 1946. Ofilogenii Azotobacter chroococcum (Phylogeny of Azotobacter chroococcum) Mikrobiologiya. 15:6.
19. Jensen, H.L. 1954. The Azotobacteriaciae. Bacteriol Rev. 18:195-214.
20. Jones, D.H. 1913. A cultural and morphological studies of some Azotobacter. Sci. 30:411-414.
21. Jones, D.H. 1913. A morphological and cultural study of some Azotobacter. Centr. f. Bakt. II Abt., 38:14-24.
22. Jones, D.H. 1937. Further studies on the growth cycle of Azotobacter. J. Bacteriol. 104:191-203.
23. Kyle, T.S., and A. Eisentark. 1951. The genus Azotobacter. Bull. Okla. Agr. Mech. Coll. 48:1-49.
24. Layne, J.S., and E.J. Johnson, 1964. Resistant properties of Azotobacter cysts induced in response to mineral deficiencies. J. Bacteriol. 88:956-959.
25. Lewis, I.M., 1941. The cytology of bacteria. Bacteriol.

56
Rev. 5: 181-230.
26. Lewis, I.M. 1937. Cell inclusions and the life cycle of Azotobacter. J. Bacteroil. 104: 191-203.
27. Lohnis, F., and N.R. Smith. 1916. Life cycles of bacteria. J. Agr. Re3. 6:675-720.
28. Lopez, J. Gonzalez, and Vela, G.R. 1981. True morphology of the Azotobacteraceae: filterable bacteria. Nature 289(5798):588-590.
29. Mellon, R.R.. Studies in microbic heredity. 1925. J. Bacteriol. 10:481-505.
30. Moreno, J., J. Gonzalez Lopez, and G.R. Vela. 1986. Survival of Azotobacter spp. in dry soils. Apple. Environ. Microbiol. 51:123-125.
31. Omelyanski. S.W., V. Btstviky, and K. Maramero. 1966. The occurrence of microbial forms of unusual morphology in European and Asian Soils. Can. J. Microbiol. 12:1291-1292.
32. Page, W.J., and H.L. Socolofsky. 1975. Relationship between calcium and vronic acids in the encystment of Azotobacter vinelandii. J. Bacteriol. 122:145151.
33. Parker, L.T., and M.D. Socolofsky. 1966. Central body of the Azotobacter cyst. J. Bacteriol. 91: 297303.
34. Pochon, J., and Y.T. Tchan. 1948. Precis de Microbiologie du Sol, p.84. Masson et Cie, Paris.
35. Pochon, J., Y.T. Tchan, and T.L. Wang. 1948. Recherche sur le cycle morphologique et l'appareil nucleare des Azotobacter. Ann. Inst. Pasteur. 84:182-188.
36. Smith, D., and N.F. Conant. 1952. Zinsser's Textbook of Bacteriology. 10th edition. Appleton Century Crofts. New York.
37. Sneath, H.A., N.S. Mair, M.E. Sharpe, and J.G. Holt, editors. 1986. Bergey's Manual of Systematic Bacteriology. Williams & Wilkins, Baltimore.
38. Socolofsky, M.D., and O.Wyss. 1962. Resistance of the Azotobacter cyst. J. Bacteriol. 84: 119-124.
39. Van Schreven, D.A. 1962. Effect of the composition of

57
the growth medium on morphology dual reproduction of Azotobacter chroococcum. Antonie Van Leuwenhoeck J. Microbiol Serol. 28: 97-120, 1962.
40. Vela, G.R. 1964. Doctoral Dissertation. Department of microbiology. University of Texas, Austin, Texas.
41. Vela, G.R. and 0. Wyss. 1964. Improved stain for visualization of Azotobacter encystment. J. Bacteriol. 87: 476-477.
42. Vela, G.R. and 0. Wyss. Radiation resistance of soil Azotobacter. J. Bacteriol, 89:1280-1285.
43. Vela, G.R., G.D. Cagle and P.R. Holmgren. 1970. Ultrastructure of Azotobacter vinelandii. J. Bacteriol. 104:933-939.
44. Vela, G.R., and R.S. Rosenthal. 1972. Effect of peptone on Azotobacter. J. Microbiol. 111:260-266.
45. Vela, G.R. 1972. Unpublished notes. Department of biological sciences, University of North Texas, Denton, Texas.
46. Vela, G.R., J.F. Wu, and D. smith, 1974. Effect of 1456 NH2 microwave radiation on some soil microorganisms. In situ. Soil Sci. 121:44-51.
47. Vela, G.R. 1974. Survival of Azotobacter in dry soil. Appl. Microbiol. 28:77-79.
48. Winogradsky, S. 1938. Sur la morphologie et l'ecologie des Azotobacter. Ann. Inst. Pasteur. 60:315-400.
49. Wyss, 0., D.D. Smith, L.M. Pope, and K.E. Olson. 1969. Enodogenous encystment of Azotobacter vinelandii. J. Bacteriol. 101:475-479.
50. Wyss, 0., M.G. Neuman, and M.D. Socolofsky. 1961. Development and germination of the Azotobacter cyst. J. Biochem. Biophys. Cytol. 10:555-565.













![Characterization of the [3Fe–4S]0/1+ cluster from the D14C ...chemgroups.ucdavis.edu/~cramer/Publications_pdf/cramer...Fe protein [4Fe–4S] cluster from Azotobacter vinelandii (Av2),18](https://img.pdfslide.us/doc/110x75/60f745b99af9f75ada100207/characterization-of-the-3fea4s01-cluster-from-the-d14c-cramerpublicationspdfcramer.jpg)















