Embed Size (px)
Citation preview

THE MECHANISM OF BACTERIAL DEHYDROGENASE ACTIVITY IN VIVO
I. ANAEROBIC DEHYDROGENASE ACTIVITY OF ESCHERICHIA COLI AS A FUNCTION OF TEMPERATURE*
BY BERNARD S. GOULD AND IRWIN W. SIZER
(From the Laboratory of Phjisiology and Biochemistry, Massachusetts Institute of Technology, Cambridge)
(Received for publication, February 23, 1938)
The velocity of chemical reactions increases exponentially with temperature according to the Arrhenius equation k = ze-@IRT where 7c is the velocity constant, x is a constant, e is 2.718, R is the gas constant, T is the absolute temperature, and P represents the energy of activation of the reaction in calories per gm. molecule In inorganic catalysis the activation energy characterizes the catalyst and is independent of the substrate when related sub- strates are used (Hoagland, 1936). Work on enzymes (yeast and malt invertases) indicates that the p value is characteristic of the specific enzyme, but independent of the substrate employed (Sizer, 1937, 1938). The value of p is constant over a wide range of temperature until the inactivation temperature of the enzyme is reached (Craig, 1936).
A possible method of differentiating between the many proposed dehydrogenases is to study the activation energy of dehydrogena- tion. From a study of reaction rate as a function of temperature the activation energy (P) may be calculated. Constant P values would be expected over the temperature range where enzyme in- activation does not occur, provided that a single dehydrogenase plays the predominant role in the reaction. If a single dehydro- genase can activate several substrates, the same temperature characteristic might be obtained for them. If, on the other
* Contribution No. 123 from the Department of Biology and Public Health, Massachusetts Institute of Technology.
269
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

270 Dehydrogenase Activity. I
hand, there is a separate enzyme for each substrate, the P value for each should characterize the specific enzyme involved.
The temperature characteristic of the succinic dehydrogenase of Escherichiu coli was calculated by Crozier (1924) from Quastel’s data and found to be 16,700. Laki (1937), using an enzyme prep- aration from pigeon breast muscle and studying the effects of temperature upon anaerobic hydrogenation of oxalacetic acid and pyruvic acid, observed no change in temperature characteristic over the temperature range from 20-40” and considers it the same for both substrates. Calculations made from his data indicate that for pyruvic acid hydrogenation p = 5900, and for oxalacetic acid I.L = 5000. Since data were obtained at only three different temperatures and the data extrapolated from the published dia- gram, it cannot be stated whether or not these two values are the same within experimental error.
A large number of substrates have been found suitable for the action of bacterial dehydrogenases (Quastel, 1932). In general, a separate enzyme has been postulated for each substrate attacked, although it has been impossible to isolate any single one of these, and teleologically it seems unlikely that specific enzymes would be found for substrates which never normally occur within the organism.
In this study of the mechanism of enzyme action the effect of temperature upon the rate of dehydrogenation was measured. The enzyme preparation was a suspension of washed “resting cells” of Escherichia coli, which was shown to have active dehydro- genating activity toward a variety of common substrates. Since dehydrogenation plays a fundamental role in cellular respiration, it was hoped to obtain information concerning the mechanism of respiration, as well as to test the validity of the Arrhenius equa- tion and its implications with respect to a large number of bac- terial dehydrogenases.
EXPERIMENTAL
Enzyme Preparation-A well identified culture of Escherichia coli was employed for the preparation of active enzyme sus- pensions. The organisms were kept on nutrient agar and transplanted into 1 per cent tryptone (Difco) broth. Liter flasks each containing about 350 cc. of sterile tryptone broth were
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

B. S. Gould and I. W. Sizer 271
inoculated with a 24 hour culture and incubated at 37” for from 40 to 48 hours. After incubation the growth was removed by centrifugation at high speed and washed repeatedly with distilled water to remove all traces of medium and soluble metabolites. Control runs indicated that three to four separate washings with 150 to 200 cc. of water removed all of the reducible material, as indicated by the failure of the preparation to reduce methylene blue in the absence of added substrate even after 6 to 8 hours. Each preparation is made up of the growth from three separate liter flasks. The preparations were kept at 2-4’ until required for use; then proper dilutions were made by adding distilled water. Dilutions were adjusted according to the substrate being investi- gated, concentrated suspensions being employed for those sub- strates which are most difficultly activated. In each case the suspension was so adjusted that the reduction time at 35” was ap- proximately 5 minutes. No attempt was made to count the total number of cells or the relative number of dead and live cells.
Technique Employed-Thunberg tubes (15 cc.) equipped with hollow bulb stoppers were employed throughout. In each tube were placed 1.0 cc. of phosphate buffer at pH 7.4, 1 cc. of 0.05 M
substrate, 0.5 cc. of distilled water, 1.0 cc. of enzyme suspension, and, finally, 0.5 cc. of 1:5000 methylene blue in the bulb of the side arm. The tubes were evacuated at the water pump for about 30 seconds, with shaking, and then partly immersed so that the contents were submerged in the constant temperature bath, con- trolled to f0.1”. The tubes were adjusted to the temperature of the water bath by allowing them to stand for 3 to 5 minutes. The reaction was initiated by introducing the contents of the side arm into the tube. The time to reach 75 per cent decolorization (as measured against an aerobic control containing all the react- ants but 0.5 cc. of 1: 20,000 methylene blue instead of the usual 1: 5000) was taken as the reduction time. Quastel and Whetham (1924) and Yudkin (1934) have shown that bacterial reduction of methylene blue in the concentrations used is linear until 80 to 90 per cent of the dye is reduced, after which the rate is retarded. In each test quadruplicate tubes were usually employed and the average reduction time was recorded. At any single temperature the four reduction times usually agreed within 5 per cent and almost always within 10 per cent. The error was larger when
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

272 Dehydrogenase Activity. I
reduction was very rapid. Determinations were carried out over
that temperature range which showed no enzyme inactivation due either to heat or to long incubation with methylene blue.
32 33 34 35 36
FIG. 1. Log of rate of reduction (calculated from the time required for 75 per cent reduction of methylene blue) in presence of various substrates by a suspension of Escherichia coli plotted against l/T. p in the Arrhenius equation = 19,400. 0 refers to the series of determinations made with a first bacterial preparation; X to determinations made with second separate bacterial preparations. For lactate X refers to determinations made with a preparation consisting of autolyzed cells.
The activating mechanism for the dehydrogenation of sodium lactate was investigated by means of a washed bacterial suspen-
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

B. S. Gould and I. W. Sizer 273
sion prepared as above. When the log of rate of reduction of methylene blue is plotted against l/T (Fig. l), the points fall along a straight line over the temperature range from 2.245”. Inactivation became apparent at 45” and above, as indicated by a falling off of the points from the curve. In the case of all sub-
TABLE I
Summary of Dehydrogenase Activity of Escherichia coli As a Function oj Temperature As Determined by Thunberg Methylene Blue
Substrate, 0.05 M
Formic acid .............. Acetic ” .............. Glycine. ................. Lactic acid ............. Pyruvic “ ............
Succinic “ ............ Glutamic “ ............. Z-Xylose ................. Glucose .................. d-Galactose .............. Mannitol. ................ Fructose .................
I-Sorbose. ................
Sucrose .................. Maltose ..................
-
--
.
-
Technique
Preparation No. (Escherichia coli)
V x, XII xv IV, XI, VIIi XV active XV weak I, II VI XIII III V III XI, XV active XV weak XV active XV weak VIII VI, VII
‘1
-
remperature range
“C
2 -40 6 -37.5
15 -35 2.2-45 0.9-35 0.9-35 4.5-65 8 45
15 -35 12 -55 15 -45 15 -55 10 -45 10 42.5 15 -32.5 20 -35 15 -37.5
1 -42.5
* The energy of activation in the Arrhenius equation
/.4 = 4.58 log kz - log k,
l/T1 - l/T,
where k is the rate and T the absolute temperature. t Preparation of autolyzed cells.
nactiva- ion tem- perature
“C
37.5 37.5
45
45 37.5
45 40 47.5 37.5 37.5
37.5 40
15,000 19,400 19,400 19,400 25,000 25,000 19,400 19,400 19,400 19,400 19,400 19,400 25,000 19,400 25,000 19,400 19,400 21,200
strates the data obtained for dehydrogenation at the inactivation temperature and above have been omitted from the figures. Data on the inactivation temperatures are presented in Table I. Since the points fall along a straight line in the plot, it is apparent that reaction rate increases with temperature in accordance with the
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

274 Dehydrogenase Activity. I
Arrhenius equation. From the slope of the curve, p in the equa- tion was calculated to be 19,400. With a new bacterial suspen- sion of a different activity, prepared after subculturing several times over a period of a month, the same P value was obtained when a second series of experiments was carried out.
An attempt was made to prepare a cell-free preparation of the activating mechanism for lactate dehydrogenation according to the methods of Stephenson (1928). The cells were autolyzed for the specified time and then the cell debris separated from the digest by centrifugation. The supernatant liquid was filtered through kieselguhr and found to be free of intact cells and cell debris. It had, however, practically no activity, as indicated by only slight reduction of methylene blue in 2 hours. The centri- fugate, which consisted largely of cell debris but contained some intact cells, had the same activity as the intact cells. Data were obtained on the cell debris (Fig. 1) and the analysis was found to be identical with that for the two independent preparations of intact cells. It seems evident that the activating mechanism is similar in dead and living cells, or at least in cells in widely dif- ferent met.abolic states.
To determine whether the same activation mechanism is in- volved in the dehydrogenation of other substrates, a series of com- pounds was investigated: fatty acids (formic, acetic), a hydroxy- substituted acid (lactic), a keto acid (pyruvic), a dicarboxylic acid (succinic), amino acids (glycine, glutamic), an aldopentose (Z-xy- lose), aldohexoses (glucose, d-galactose), ketohexoses (fructose, Z-sorbose), a polyhydric alcohol (mannitol), a c&-fermentable disaccharide (maltose), a c&-non-fermentable disaccharide (suc- rose). In the case of the acids the sodium salt was always employed:
From an analysis of the data (Fig. 1) it becomes apparent that the same energy of activation (P) as for lactate is found for the following substrates: acetate, glycine, glutamate, succinate, glucose, mannitol, galactose, xylose, and sucrose. More than one enzyme preparation was used in the study of certain of these substrates, but the results obtained were always independent of the strength of the bacterial preparation. The lower range of temperature was not studied in the case of certain of these sub- strates, where the reduction time at low temperatures was so long
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

B. S. Gould and I. W. Sizer 275
that the prolonged incubation with methylene blue inhibited the enzyme action (Yudkin, 1933). This identity of P values sug- gests that the same activation mechanism may be involved in the dehydrogenation of all these substrates.
In the study of the dehydrogenation of sodium formate a dis- tinctly different temperature characteristic of 15,000 was obtained (Fig. 2). This suggests that perhaps a different activation me- chanism is involved for the dehydrogenation of formate. This is consistent with the results reported by Quastel (1932) who states that, unlike most dehydrogenases, formic acid dehydrogen- ase is very resistant to the action of anesthetics but is readily destroyed by oxidizing agents and KCN. It is the only dehydro- genase which can be reversibly inactivated.
A different temperature characteristic was also obtained for the dehydrogenation of maltose (Fig. 2), where p = 21,200. The question arises as to whether maltose is attacked directly or whether it must first be hydrolyzed to glucose before dehydro- genation takes place. If the latter were true, it might be the rate of hydrolysis which was determining rate of reduction and also the temperature characteristic. This seems possible, since in hydrolysis and fermentation experiments the presence of maltase has been demonstrated in the strain of Escherichia coli used. Moreover, with the non-fermentable disaccharide, sucrose, the typical P value of 19,400 was obtained. Since sucrose is dehydrogenated directly, a similar situation may hold true for maltose and p 21,200 may represent a distinct dehydrogenation mechanism for maltose.
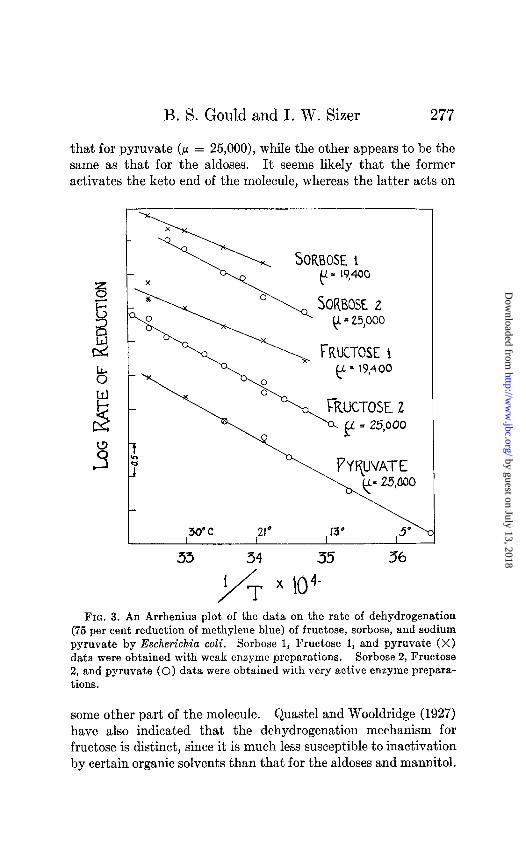
An entirely different energy of activation is indicated in the case of sodium pyruvate (Fig. 3), where a p = 25,000 was ob- tained. The same p is encountered in the dehydrogenation of other compounds containing keto groups, such as fructose and sorbose. Values of P = 25,000 were obtained with these sub- strates when extremely active enzyme preparations were em- ployed (Fig. 3). This new p value was not due to a change in the nature of the bacterial suspension, for when a series of experi- ments was made on lactate with the same active enzyme prepara- tion, the usual p 19,400 was obtained. However, when weak enzyme preparations were used in the dehydrogenation of fructose and sorbose, the typical p 19,400 was obtained. In the case of
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

276 Dehydrogenase Activity. I
pyruvate, however, the value of 25,000 is independent of the activity of the enzyme preparation. It thus appears that two
34
/ i rx104
36
FIG. 2. An Arrhenius plot of the data on the rate of dehydrogenation (75 per cent reduction of methylene blue) of sodium formate, sucrose, and maltose by Escherichia coli. 0 refers to the series of determinations made with a first bacterial preparation; X to determinations with a sec- ond bacterial preparation.
entirely separate activating mechanisms may bring about the dehydrogenation of ketohexoses. One appears to be the same as
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from
B. S. Gould and I. W. Sizer 277
that for pyruvate (P = 25,000), while the other appears to be the same as that for the aldoses. It seems likely that the former activates the keto end of the molecule, whereas the latter acts on
33 34 35 36
FIG. 3. An Arrhenius plot of the data on the rate of dehydrogenation (75 per cent reduction of methylene blue) of fructose, sorbose, and sodium pyruvate by Escherichia coli. Sorbose 1, Fructose 1, and pyruvate (X) data were obtained with weak enzyme preparations. Sorbose 2, Fructose 2, and pyruvate (0) data were obtained with very active enzyme prepara- tions.
some other part of the molecule. Quastel and Wooldridge (1927) have also indicated that the dehydrogenation mechanism for fructose is distinct, since it is much less susceptible to inactivation by certain organic solvents than that for the aldoses and mannitol.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

278 Dehydrogenase Activity. I
DISCUSSION
A summary of the results is embodied in Table I, an analysis of which indicates that there may be four activating mechanisms involved in the dehydrogenation of the substrates investigated which differ in length of carbon chain, stereo configuration, and nature of substituent groups.
The temperature characteristics obtained on dehydrogenation by a bacterial suspension are extremely suggestive in the light of values obtained for other respiratory and dehydrogenating mech- anisms: rate of reduction of cytochrome (added glucose) p = 19,600 (Stier, 1935) ; oxygen consumption (added glucose) by Axotobacter, p = 19,300 (Lineweaver et al., 1932), by Chlorella, p = 19,000 (Crozier et al., 1934), and yeast, p = 19,500 (Stier, 1933); dehydrogenation of luciferin by luciferase from Cypridina, p = 25,000 (Crozier, 1924).
SUMMARY
The mechanism of anaerobic dehydrogenation of a variety of substrates by a suspension of Escherichia coli was investi- gated as a function of temperature. Rate of reduction was calculated from the time required to bring about 75 per cent de- colorization of the dye, according to the Thunberg methylene blue technique. In the case of all substrates investigated the rate of dehydrogenation increases exponentially with temperature in accordance with the Arrhenius equation. Above 37-47” the plotted points fall off from the curves, indicating temperature inactivation of the enzyme system.
The energy of activation, calculated from the slope of the straight lines obtained when the log of rate is plotted against l/T, is 19,400 calories per gm. molecule for the following substrates: acetate, glycine, glutamate, lactate, succinate, glucose, mannitol, galactose, xylose, and sucrose. This suggests the possibility of a single activating mechanism which brings about the dehydrogena- tion of all these substrates.
P = 25,000 is obtained for the dehydrogenation of pyruvate, and also for fructose and sorbose, with an active enzyme prepara- tion. With a weak enzyme suspension a P value of 19,400 is ob- tained for the ketoses. The explanation is offered that P 25,000 refers to the dehydrogenase system for the keto end of the hexose,
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

B. S. Gould and I. W. Sizer 279
while p = 19,400 represents the system activating the H in another part of the molecule.
For the dehydrogenation of sodium formate p = 15,000, while for maltose it is 21,200. Evidence is reviewed which lends further evidence that formic acid dehydrogenase and fructose dehydrogen- ase are different from the other dehydrogenases. It is possible that hydrolysis is complicating the situation in the case of mal- tose dehydrogenation.
BIBLIOGRAPHY
Craig, F. N., J. Biol. Chem., 114, 727 (1936). Crazier, W. J., J. Gen. Physiol., 7, 189 (1924). Crazier, W. J., Tang, P. S., and French, S., J. Gen. Physiol., 18, 113 (1934). Hoagland, H., in Cold Spring Harbor symposia on quantitative biology,
Cold Spring Harbor, 4, 267 (1936). Laki, K., 2. physiol. Chem., 249, 57 (1937). Lineweaver, H., Burk, D., and Horner, K., J. Gen. Physiol., 16, 497 (1932). Quastel, J. H., in Nord, F. F., and Weidenhagen, R., Ergebnisse der Enzym-
forschung, Leipsic, 1, 209 (1932). Quastel, J. H., and Whetham, M. D., Biochem. J., 18, 519 (1924). Quastel, J. H., and Wooldridge, W. R., Biochem. J., 21, 148 (1927). Sizer, I. W., J. Cell. and Comp. Physiol., 10, 61 (1937); Enxymologia, 4, 215
(1938). Stephenson, M., Biochem. J., 22,605 (1928). Stier, T. J. B., J. Gen. PhysioZ., 16, 815 (1933); 19, 339 (1935). Yudkin, J., Biochem. J., 27, 1849 (1933); 28, 1454 (1934).
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Bernard S. Gould and Irwin W. SizerOF TEMPERATURE
ESCHERICHIA COLI AS A FUNCTIONDEHYDROGENASE ACTIVITY OF
VIVO: I. ANAEROBICDEHYDROGENASE ACTIVITY IN
THE MECHANISM OF BACTERIAL
1938, 124:269-279.J. Biol. Chem.
http://www.jbc.org/content/124/1/269.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/124/1/269.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from