Embed Size (px)
Citation preview

I96 BIOCHIMICA ET BI(}PHYSI( 'A A('TA
BBA 35436
THE LARGEST KNOWN MONOMERIC GLOBULAR PROTEINS
A. H. R E I S N E R , J A N E T I U ) \ V E AND H E L E N 31. M A ( ' I N I ) ( ) E
C.S I.R.O., Dzz,tston of Anzmal (;enehcs, P (~ Box 90, l:'ppzng, N 5 IV 2zez (Ausiraha)
( R e c e i v e d M a y t 2th, I009)
SUMMARY
The quaternary structures of three of the water-soluble surface proteins (immo- bilization or serotypic antigens 5IA, 5IB and 5zD) of P a r a m e c i u m altrelia stock 5 I (syngen 4) were studied by examining the behavior of the reduced proteins under a variety of conditions. Molecular weight determinations by sedimentation equilibrium of the native and the reduced proteins revealed no significant differences between the two forms while intrinsic viscosities of these molecules, under conditions known to produce random coils, were z33.4 ml/g (5zA), 115.2 ml/g (5zB) and 123.o mt/g (5ID). These data support the conclusion that these proteins (tool. wt. 3.o. IO a, 2.6- IO 5 and 2. 7-zo 5 g/mole) are composed of single polypeptides. In addition neither gel-per- meation chromatography nor sedimentation-velocity analysis indicated the presence of associated small peptides and the reduced antigens produced one predominant zone upon starch-gel electrophoresis. The possible significance of the large size of the~e monomers is discussed.
INTRODUCTION
The soluble surface proteins of the ciliated protozoan, Paranmcimn (immo- bilization antigens, serotypic antigens), are a closely related family of lnolecules whose syntheses are governed by a system of mutual exclusion. An individual may p(~sses.~ the genic potential for synthesizing over a dozen of these proteins but normally only one can be detected at a given time. No more than one structural gene has ever been demonstrated to be involved in the synthesis of an inunobilization antigen, and where breeding analyses have been carried out these genes have been shown to be unlinked (see PREER 1 for the most recent review of the subject and citations of the earlier literature). Clearly, the system is one involving nmlti-cistronic regulation in an eukaryote, and its understanding mayT provide valuable insight into ontological regulation in higher forms.
The study of this regulatory system poses three principal question.~: what is being regulated (structure of the immobilization antigen), what are the mechanisms which preclude the synthesis of all but one member of the family at a time, and how
Btochun Bzophys. Acta, L~8 (~909) 1@~-2o0

LARGEST MONOMERIC GLOBULINS 197
does a cell shut down synthesis of one immobilization antigen and begin the synthesis of another (serotypic transformation) ? We have initially undertaken a study on the structure of these proteins in the belief that the information obtained would help in the analysis of the control systems.
Contrary to previous reports 2-4, we have found no evidence of subunit structure in the three proteins reported on here, i.e. they appear to consist of a single polypeptide and are, therefore, four to five times larger than any previously reported monomeric globular protein.
MATERIALS AND METHODS
Paramecium aurelia stock 51 (syngen 4) free from kappa were grown in mono- xenic mass culture and harvested as previously described 5.
Extraction and purification of the three serotypic antigens (5IA, 5IB and 5ID) were performed using published methodsS, ~, and antigenic activity was determined by adsorption titration ~3. The initial (NH4)2SO 4 precipitation of 5ID was at 67% saturation, while reprecipitations were done at 5 ° % saturation.
Physical determinations. Molecular weights were determined by meniscus- depletion-equilibrium 8 in a Spinco Model E ultracentrifuge ; measurements of photo- graphic plates were made using a Nikon Model 6C comparator equipped with 2/,/di- vision Mitotuyo micrometers. The partial specific volume (~) of native 5IA was de- termined using the H~O-ZH20 system 9, and ~'s for native 5IB (0.703) and 5ID (0.706) were calculated from their amino acid compositions l°,n when it was found that the observed and calculated ~'s for 5IA were in agreement (0.708 ml/g and 0.702 ml/g, respectively)• For all calculations reported in this study, ~ is taken as 0.705 ml/g. The ~'s of these proteins in solutions of urea and guanidine • HC1 were taken as the same as those of the native molecules 12.
Intrinsic viscosities [~l were determined as described previously 6 using either Cannon-Fenske or Cannon-Manning (semi-micro) capillary viscometers. Water flow times were between 175 and 200 sec.
Reagents. Iodoacetic acid and iodoacetamide were twice recrystallized before use. Urea was recrystallized from hot ethanol, and guanidine.HC1 was prepared as already described 12. All other reagents were analytical reagent grade or the purest commercially obtainable and were used without additional purification.
Electrophoresis. Horizontal starch-gel electrophoreses were performed in IO cm × 22 cm slabs. The urea buffer systems principally employed were those of SMITHIES et al.13,14 and gels were run either at 4 ° or at room temperature as indicated in RESULTS.
• 0 / Isoelectric focusing 15 17 was done in a 33 density-step i io-ml column using I / o Ampholine buffer (pH 3-5, LKB, Stockholm). The column was run at 800 V for 65 h with the column temperature maintained at 20 ± 0.2 °. o.i ml of a mixture of the three antigens was mixed with each of Steps 5 to 25. Positions of the antigens were de- termined by pumping the contents of the column through a flow analyzer and by recording the absorbance at 280 m/~. 2-ml fractions were collected and analyzed for antigenicity 5, and the pH of the fractions was determined using a Beckman expanded scale pH meter. In order to obtain the differential absorbance plot shown in Fig. 2, a blank column was run under conditions identical to those used for the experimental column.
B*och*m. B*ophvs. Acta, 188 (1969) 196-2o6

198 A.H. REISNER el al.
Disulphide cleavage and alkylation. Reduction of the protein was performed either with fl-mercaptoethanol or dithiothreitol (Cleland's reagent). In those cases where reduction was not followed by blocking, the protein was maintained in buffers made o. 15 M to 0.5 M with respect to the reductant. Reduction and S-carboxymethyl- ation was done according to the method of CRESrVlELI~ et al. 18, while reduction and S-carboxamidomethylation was done using the method of SMHmES et al. u.
In preliminary experiments, designed to determine the completeness of disul- phide cleavage, 5IA antigen was subjected to reduction in o.2, o.5, I, 2 and 4 times the fl-mercaptoethanol concentration called for in the standard reduction S-earboxv- methylation procedure TM, and the protein was held under reducing conditions for periods from I to 6 h. Except for the 0.2 times fl-mercaptoethanol concentration which produced a slow moving highly heterogeneous product, all reaction products gave starch-gel patterns identical to those shown in Fig. 6b. Similar experiments using reduction and S-carboxamidomethylation x4 in which reducing conditions were main- tained from Io to 15o rain produced starch-gel patterns identical to Figs. 6a and (~f except for the 0.2 times fl-mercaptoethanol concentration. In all these preparations the amount of iodoacetic acid or iodoacetamide was appropriately adjusted. Finalh' the mobility of the reduced S-carboxamidomethylated product in 7 M urea-formate starch gels (pH 3.o) wa~ t~o °,,"o that of the native antigen.
Gel-permeatiou chromatograpl(v. Bio-Gel P-3oo columns (2.5 cm ~ 37 cm) (Bio- Rad Laboratories, Richnlond, Calif.) were run in the upward flow mode. The buffer system used was 8 M urea-o.2% Na 2 EDTA-o.36 M Tris (pH 8.6) and, in the case of the reduced protein, o. 15 M fl-mercaptoethanol was added. Columns were run at room temperature at a flow rate of 2.o ml/h, and the effluents were monitored at 280 m/z with a flow analyzer.
T A B L E I
S020,w V A L U E S O F 5 I A A N T I G E N IN V A R I O U S S O L V E N T S
T h e T r l s b u f f e r c o n s i s t e d o f o .05 M T n s HC1 ( p H 7.5) , o i M NaC1, o . i M MgC1 a. T h e d i s s o c i a t i n g s o l v e n t s y s t e m s c o n t a i n e d 0 .36 5I T r l s - H C 1 ( p H 8.6) , 0 .2 % (w/v) N a 2 E D T A . S o l v e n t s f o r r e d u c e d p r o t e i n c o n t a i n e d 0 .3 M f l - m e r c a p t o e t h a n o l . C e n t r l f u g a t i o n s w e r e p e r f o r m e d a t 59 780 r e v . / m i n n e a r 20 ° F i g u r e s in p a r e n t h e s e s a r e S%o,u, v a l u e s f o r m u l t i m e r s w h i c h o c c u r r e d in c e r t a i n s o l v e n t s .
Solvc~tt Unreduced 5L4 Reduced Reduced 5zA and S-
carboxv- methy~led 5rA
Tr l s b u f f e r 8. 5 ,S M u r e a 7.5 5 .0 " 6 M g u a m d l n e . H C 1 7 . 3 ( I ° . ° , t 3 . 8 , 17.9 ) 4 .8 -- 8 M g u a n i d i n e . H C I 7 9 (1I . 7, 14. 7) 4-7 4 M u r e a + 6. 5 M
g u a n i d i n e - HCI 7 9 (IO-5) - - 8 M u r e a + 5 . I M
g u a n i d i n e • HC1 7.2 --- 0 .02 M ( N H 4 ) H C O a +
o . I M N a C l ( p H 8.8) 5 o
B~,,,h~m. Blophys. Acla, IS8 (1969) 1 9 6 - 2 o O
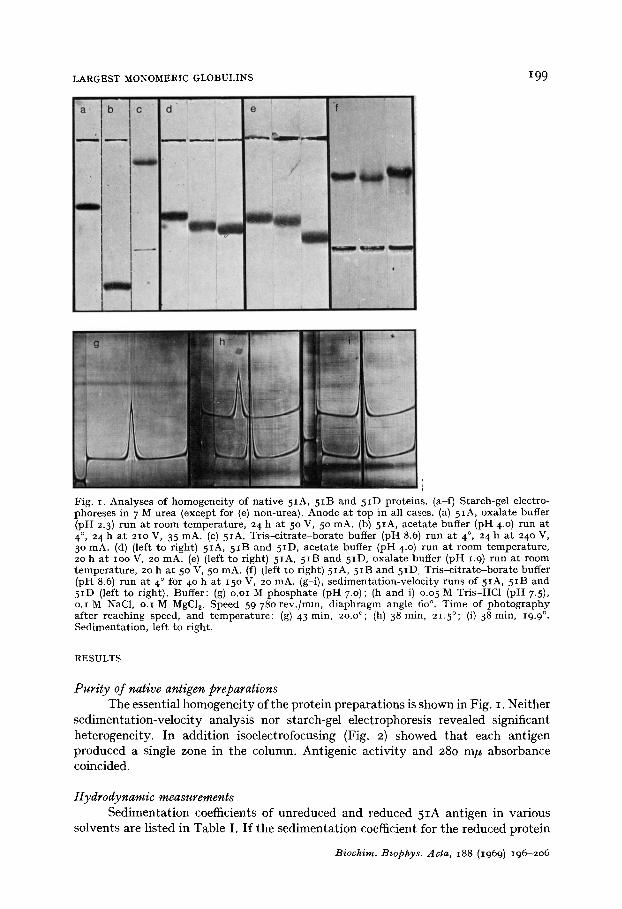
LARGEST MONOMERIC GLOBULINS 199
Fig. I. Analyses of homogene i ty of nat ive 5IA, 5IB and 5ID proteins. (a-f) Starch-gel electro- phoreses in 7 M urea (except for (e) non-urea). Anode at top in all cases. (a) 5IA, oxalate buffer (pH 2.3) run at room tempera ture , 24 h a t 5 ° V, 5 ° mA. (b) 5IA, aceta te buffer (pH 4.0) run at 4 °, 24 h a t 21o V, 35 mA. (c) 5IA, Tr i s -c i t r a t e -bora te buffer (pH 8.6) run at 4 °, 24 h a t 24o V, 3 ° mA. (d) (left to right) 5IA, 51B and 5ID, aceta te buffer (pH 4.0) run at room tempera ture , 20 h a t IOO V, 2o mA. (e) (left to right) 5IA, 5IB and 5ID, oxalate buffer (pH 1.9) run at room tempera ture , 2o h a t 5o V, 5 ° mA. (f) (left to right) 5IA, 5IB and 5 I D Tr i s -c i t r a t e -bora te buffer (pH 8.6) run at 4 ° for 4 ° h a t 15o V, 20 mA. (g-i), sedimenta t ion-veloc i ty runs of 5IA, 5 IB and 5ID (left to right). Buffer: (g) o.oi M phospha te (pH 7.0) ; (h and i) o.o 5 M Tris-HC1 (pH 7.5), o.I M NaC1, o.I M MgC1 v Speed 59 780 rev.]min, d iaphragm angle 60 °. Time of pho tog raphy af ter reaching speed, and t empera tu re : (g) 43 rain, 2o.o°; (h) 38 min, 21.5°; (i) 38 min, 19.9 °. Sedimenta t ion, left to right.
RESULTS
Purity of native antigen preparations The essential homogeneity of the protein preparations is shown in Fig. I. Neither
sedimentation-velocity analysis nor starch-gel electrophoresis revealed significant heterogeneity. In addition isoelectrofocusing (Fig. 2) showed that each antigen produced a single zone in the column. Antigenic activity and 280 m/z absorbance coincided.
Hydrodynamic measurements Sedimentation coefficients of unreduced and reduced 5IA antigen in various
solvents are listed in Table I. If the sedimentation coefficient for the reduced protein
Biochim. Bzophys. Acta, I88 (1969) I96-2o6
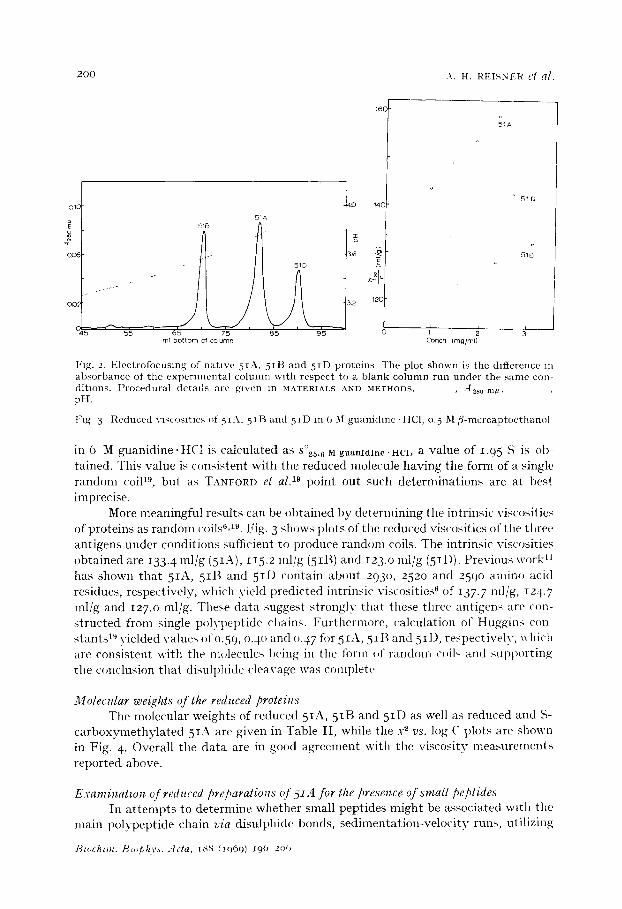
200 A . H . REISNER cl a].
01C
OP(
OD."
t ~ 75 mt bot tom of column
51A
51D
85 95
;6(
4.0 140
3.2 120
11 i 2
Conch (mg/ml)
O 51A
51D
51B
J Fag. 2. Electrofocusmg of native 5IA, 5 IB and 5ID proteins The plot shown is the difference m absorbance of the expermlental column w~th respect to a blank column run under the same con- ditions. Procedural d e t a i l s a r e ~ l v e n i n M A T E R I A L S A N D M E T H O D S . - - , A 2 8 0 m l ~ , - "
pH.
Fig 3 Reduced VlqCOSltles of 5 I A . 5II~ and 5 tD in (, 3I guanidmc-HC1, 0. 5 M/3-mercaptocthanol
in 6 M guanidine.HC1 is calculated as s°~.6 M guanidine'HC1, a value of 1.95 S is ob- tained. Thi, value is consistent with the reduced molecule having the form of a single random coil tg, but as TANFORD et al. 19 point out such determinations are at best imprecise.
More meaningful results can be obtained by determining the intrinsic viscosities of proteins as random ('oils 6,19. Fig. 3 shows plots of the reduced viscosities of the three antigens under conditions sufficient to produce random coils. The intrinsic viscosities obtained are 133. 4 ml/g (5IA), i I5.2 l n l /g ( 5 I B ) and 123.o lul/g (5ID). Previous work II has shown that 5IA, 5IB and 5ID c,ntain about 293 o, 252o and 259o amino acid residues, respectively, which yield predicted intrinsic viscosities ~ of 137.7 rot/g, 124.7 ml/g and 127.o ml/g. Thebe data suggest strongly that these three antigens are con- structed from single polypeptide chain,~. Furthermore, calculation of Huggins con- stants 1" yielded value~ of o.59 , o.4o and o.47 for 5IA, 5IB and 5ID, respectively, which are consistent with the molecules being in the form of random coil, and supt}orting the conclusion that disulphide cleavage was comI)lete
Molecular weights of the reduced proteins The molecular weights of reduced 5IA, 5IB and 5ID as well as reduced and S-
carboxymethylated 5IA are given in Table II, while the x 2 vs. log C plots are shown in Fig. 4. Overall the data are in good agreement with the viscosity measurements reported above.
Exam inatzon of reduced preparations of 5zA for the presence of small peptides In attempts to determine whether small peptides might be associated with the
main polypeptide chain via disulphide bonds, sedimentation-velocity run~, utilizing
Bu,ch~m. Bmphys..4cta, t•S (lq60) I9(,-2ol,
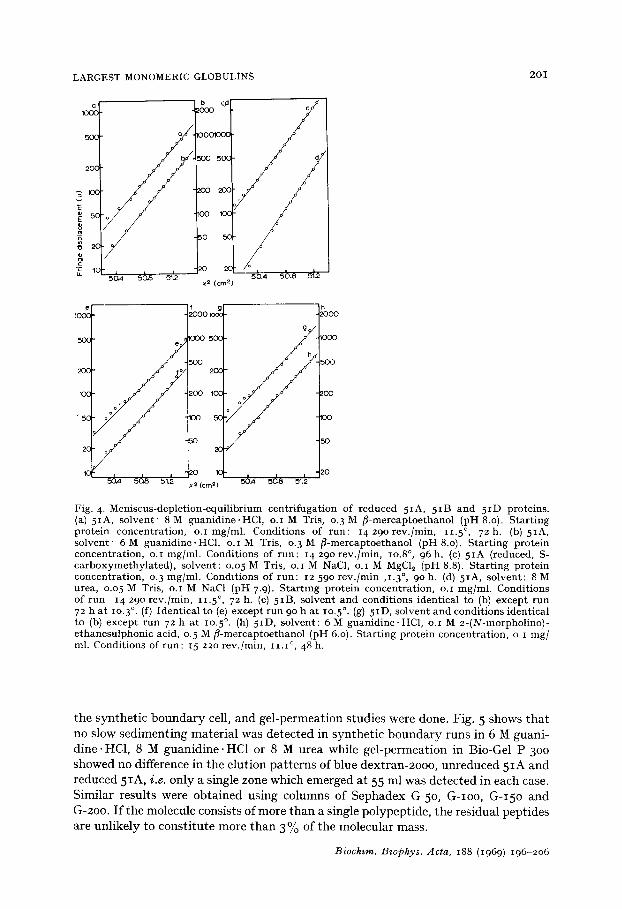
LARGEST MONOMERIC GLOBULINS 2OI
o
50c
20(
1oo
E 5o
2C
~ ~c
Q
56A s&8 ~
b 0 c# oo
9001000
~oo 5oc
_'(30 2oc
0(3 10C
50 5(
20 2( " °5L4u 5O8 512 x2 (crn2)
e IOOC
5C)C
20C
lOC
• 5C
2C
1C
e
5(~4 5~8 51.~
ioo
50(
2C~
~OC r~f
50 2of
20 1C~-
x 2 ( ¢ m 2 ) 5~,4 5C~ 51.~
_•oo
000
500
O0
50
Fig. 4 -Men i scus -dep le t ion -equ i l ib r ium cen t r i fuga t ion of r educed 5IA, 5 IB and 5 ID proteins . (a) 5IA, so lven t ' 8 M g u a n i d i n e . H C l , o.I M Tris, 0. 3 M f i -mercap toe thano l (pH 8.o). S ta r t ing pro te in concent ra t ion , o. i mg/ml . Condi t ions of r un : I 4 2 9 o r e v . / m i n , i i . 5 °, 7 2 h . ( b ) 5 I A , so lven t ' 6 M g u a n i d i n e . H C ] , o. i M Tris, 0. 3 M /5-mercaptoethanol (pH 8.0). S ta r t ing pro te in concent ra t ion , o.I mg/ml . Condi t ions of run : 14 290 rev . /min , lO.8 °, 96 h. (c) 5 IA (reduced, S- c a r b o x y m e t h y l a t e d ) , so lvent : o.05 M Tris, o.I M NaC1, o.I M MgC12 (pH 8.8). S ta r t ing pro te in concent ra t ion , 0. 3 mg/ml . Condi t ions of run : 12 59o r ev . /min ,I .3 °, 9 o h . ( d ) 5 I A , so lvent : 8 M urea, o.o 5 M Tris, o.I M NaC1 (pH 7.9). S ta r t ing pro te in concent ra t ion , o. i mg/ml . Condi t ions of r un 14 290 rev . /min , 11.5 °, 72 h. (e) 5IB, so lven t and condi t ions ident ical to (b) excep t r u n 72 h a t lO.3 °. (f) Ident ica l to (e) except r u n 90 h a t lO.5 °. (g) 5ID, so lven t and condi t ions ident ical to (b) excep t r u n 72 h a t lO.5 °. (h) 5ID, so lvent : 6 M guan id ine .HC1, o.I M 2-(N-morphol ino)- e thanesu lphon ic acid, o. 5 M ~ -mercap toe thano l (pH 6.0). S ta r t ing pro te in concen t ra t ion , o. i mg/ ml. Condi t ions of run : 15 220 rev . /min , i i . i °, 48 h.
the synthetic boundary cell, and gel-permeation studies were done. Fig. 5 shows that no slow sedimenting material was detected in synthetic boundary runs in 6 M guani- dine.HC1, 8 M guanidine-HCl or 8 M urea while gel-permeation in Bio-Gel P-3oo showed no difference in the elution patterns of blue dextran-2ooo, unreduced 5IA and reduced 5IA, i .e. only a single zone which emerged at 55 ml was detected in each case. Similar results were obtained using columns of Sephadex G-5o, G-Ioo, G-I5O and G-2oo. I f the molecule consists of more than a single polypeptide, the residual peptides are unlikely to constitute more than 3 % of the molecular mass.
Bioch,m. Bzophys. Acta, 188 (1969) I96-2o6

202 A . H . REISNER el a[.
T A B L F 1 l
MOLECULAR \VEIGHTS OF REDUCED SEROTYP]C ANTIGENS 51.\, 511~ AND 31[)
Details of the ultracentrffugal runs are given in Fig. 4. Molecular wmghts of the na twc proteins n . 3Ol 5oo (51A), 259 ooo (5IB), 271 ooo (5H)).
Solvent .'t nttgt'n Reduced N-carbo.D,-
5 ~ A 5114 5 z 1) methy!alcd 5L4
8 M u r e a + 0. 3 M /~-mercaptoethanol 308 ooo
8 M g u a m d m e . HCI -t o. 3 M ~-mereaptoethanol 33 ° ooo
6 M guamdlne .HCI t o 3 M p'-mercaptoethanol 294 ooo
0.05 MTrls k o . I M NaC1 t- o.I M MgC12
251 OOO 204 OOO 268 OOO 239 oOO
- 246 ooo
Fig. 5. Ultracentr ifugat lon of reduced 5IA protein using the cup-type synthetic boundary cell. All runs at 59 78o rev./min, d iaphragm angle, 65 °. (a, b) Solvent: 6 M guanldme. HC1, o I h'i Tns , o. 3 M /3-mercaptoethanol (pH 8.5). Temperature , I8.7 °. Tmle of photograph?" after boundary format ion: (a) 91 nnn; (b) 283 man. (c, d) Solvent 8 M guamdme-HC1, o I M Tris, 0. 3 M /)-mer- captoethanol (pH 8 5)- Temperature , 2o.o". Time of photograph?" after bounda ry formatton (c) 57 min, (d) 377 m m (e, f) Solvent" 8 M urea, o.36 M Tns, o.2% Na, EIYfA, o 15 M/%mercapto- ethanol (pH 8.6). Tempera ture 2o.o °. Time of pho tog raphy after boundary formation (e) 7 {) ram; (f) 268 min. Sedimentatmn, left to right. The apparen t change in the positron of the bo t tom of the cell during the course of the runs is caused by the deposition of the solute
Starch-gel electrophoresis of reduced antigen Antigen 5IA was analyzed using starch-gel electrophoresis under a variety of
conditions. The reduced unblocked protein was examined as well as material treated with iodoacetic acid, iodoacetamide or ethylenimine 2t. In addition reduced but un- blocked antigens 5IB and 5ID were analyzed. Fig. 6 shows the results of running such
Htochzm B~opky~ Acta, i88 (Iq69) t96-2o()
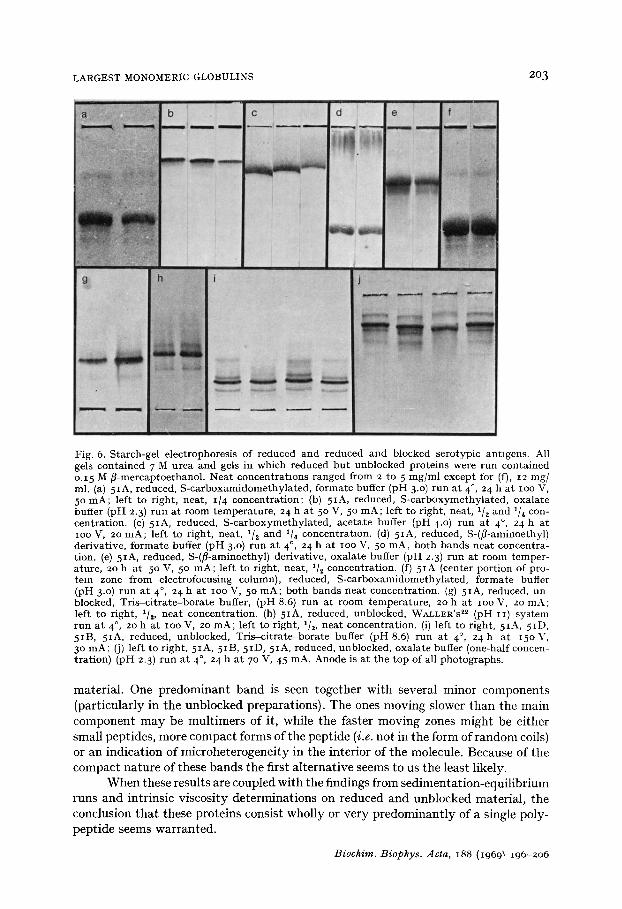
LARGEST MONOMERIC GLOBULINS 203
Fig. 6. S tarch-gel e lectrophoresis of reduced and reduced and blocked serotypic ant igens . All gels con ta ined 7 M urea a n d gels in which reduced b u t unb locked pro te ins were run con ta ined o.15 M f i -mercaptoe thanol . Nea t concen t ra t ions ranged f rom 2 to 5 m g / m l except for (f), 12 mg/ ml. (a) 5IA, reduced, S - c a r b o x a m i d o m e t h y l a t e d , fo rma te buffer (pH 3.0) r u n a t 4 °, 24 h a t IOO V, 5 ° m A ; left to r ight , neat , i /4 concen t ra t ion ; (b) 5IA, reduced, S -ca rboxyme thy l a t ed , oxa la te buffer (pH 2.3) r u n a t room t empera tu re , 24 h a t 5 ° V, 5 ° m A ; left to r ight , neat , 1/2 and 1/4 con- cent ra t ion . (c) 5IA, reduced, S - ca rboxyme t hy l a t ed , ace ta te buffer (pH 4.0) r un a t 4 °, 24 h a t IOO V, 20 m A ; left to r ight , neat , 1/~ and 1/4 concent ra t ion . (d) 5IA, reduced, S-(f i-aminoethyl) der ivat ive , f o rma te buffer (pH 3.0) r u n a t 4 °, 24 h a t ioo V, 5 ° m A , bo th bands nea t concen t ra - t ion. (e) 5IA, reduced, S-(~-aminoethyl) der ivat ive , oxa la te buffer (pH 2.3) r u n a t room t emper - a ture , 20 h a t 5 ° V, 5 ° m A ; left to r ight , neat , x/i concent ra t ion . (f) 5 IA (center por t ion of pro- rein zone f rom elect rofocusing column), reduced, S - ca rboxamidome thy l a t ed , fo rma te buffer (pH 3.0) r un a t 4 °, 24 h a t ioo V, 5 ° m A ; bo t h b a n d s nea t concent ra t ion . (g) 5IA, reduced, un- blocked, T r i s - c i t r a t e - b o r a t e buffer, (pH 8.6) r un a t room t empe ra tu r e , 20 h a t ioo V, zo m A ; left to r ight , l/z, n e a t concent ra t ion . (h) 5IA, reduced, unblocked, WALLER'S ~2 (pH II) s y s t e m r u n a t 4 °, 20 h a t ioo V, 2o m A ; left to r ight , 1[2, n e a t concent ra t ion . (i) left to r ight , 5IA, 5 ID, 5IB, 5IA, reduced, unblocked, T r i s - c i t r a t e - b o r a t e buffer (pH 8.6) r u n a t 4 °, 24 h a t 15o V, 3 ° m A ; (j) left to r ight , 5IA, 5IB, 5ID, 5IA, reduced, unblocked, oxa la te buffer (one-half concen- t ra t ion) (pH 2.3) r u n a t 4 °, 24 h a t 7 ° V, 45 mA. Anode is a t t he top of all pho tographs .
material. One predominant band is seen together with several minor components (particularly in the unblocked preparations). The ones moving slower than the main component may be multimers of it, while the faster moving zones might be either small peptides, more compact forms of the peptide (i.e. not in the form of random coils) or an indication of microheterogeneity in the interior of the molecule. Because of the compact nature of these bands the first alternative seems to us the least likely.
When these results are coupled with the findings from sedimentation-equilibrium runs and intrinsic viscosity determinations on reduced and unblocked material, the conclusion that these proteins consist wholly or very predominantly of a single poly- peptide seems warranted.
Biochim. Biophys. Acta, 188 (I969~ 196-2o6

204 A. It. I~EISNER et al.
DISCUSSION
The physical studies reported in this paper strongly suggest tha t immobilization antigens 5IA, 5 IB and 5 ID are each composed solely or predominant ly of a single polypeptide chain. I f inter-chain linkages do exist they are composed of other than weak bonds or disulphide bridges. There is to our knowledge no precedent flw postulat- ing such bonds in globular proteins. In addit ion the intrinsic viscosities of the reduced proteins in 6 M guanidine • HC1 fall very close to the values predicted for random coils, an unlikely finding if there were significant cros.~-linkages between nmltiple polypet> tides of more or less equal length.
Prior to the present report three previous publications 2 a have dealt with the qua te rnary s tructure of these molecules. All three concluded that the immobilization antigens are composed of nmltiple subunits and in the case of two of the studies a,4 rather complex arrangements were proposed (STEERS a, a nanomer composed of three different polypeptides, linked through disulphide bridges; FINGER et al?, at least a hexamer composed of three different polypeptides) JoxEs '~, however, concluded that antigens from syngen I were composed of two identical polypeptides, linked via thiol bridges, which might contain further nonidentical subunits (these "sub-subunits" are apparent ly postulated because of the variable and heterogeneous products ~,btained upon carboxymethyla t ion of the reduced protein and the determination ~,f 8 7 ooo daltons as the molecular weight of the reduced protein in 8 M urea). The proposal of two identical subunits (tool. wt. 125 ooo g/mole) stems from fingerprints ,,f the t ryp- tides obtained from several of the antigens. The molecular weight of the reduced protein was obtained using the short colunm equilibrium method at a rather higl, protein concentrat ion and nonideal behavior of the solute may have cau.~ed a con- siderable reduction in the apparent molecular weight (we have fnund thi~ t,~ be the case in meniscus-depletion-equilibrium runs). \Vhile there IS, in our opinion, no con> pelling evidence for ">ub-subumts" in these .%'ngen I molecules, the t rypt idc finger- print~ JONES obtained are in good agreement with the concept of two identical sub- units. On present evidence, therefore, there appears t~ be a real difference in the quaternary' s tructure of the immobilization antigenq studied m .,yngen I and the syngen 4 molecule,~ reported on here.
F I N G E R el al. ~ proposed that antigens from syngen z. were composed of at least six subunits representing three different polypeptides. Their conclusion \va~ based wholly on inmmnological evidence and, while elegant, the approach used wa, rather indirect. I t required the assumptum that the nnxed antigenic activities obtained from heterozygote population~ were due to single hybrid molecules rather than t,~ a mixed population of molecules The evidence justifying this assumption i.~ n~t overly strong.
The work reported by STEH~S a was on the 5IA molecule and can be eolnpared directly with the analyses recorded here. There is htt le doubt tha t the protein examined in the two studies is the .~ame, for the molecular weight determination~ on the native material are in agreement, a> are the amino acid analvseslL STEERS deternfined the s~z0,~, value for the reduced ca rboxymethy la ted derivative to be 2.2 S while we obtain s".,o,,,, 5.0 S. In addit ion we obtained S°2o,u, values for the reduced protein in 8 M urea and 0 M guanidine • HCI of 5.o S and 4.8 S, respectively. Although he does not give the protein concentration, STEERS determined the s,,0,,c values of the reduced antigen in
Htochzm t4~.pkv~ tcta l~S (l~)l~9) t~)t~ -ol~

LARGEST MONOMERIC GLOBULINS 205
Fig. 7. Sedimenta t ion-veloc i ty ul t racentr i fugat ion of reduced, S -ca rboxymethy la ted 51A protein. Solvent 0.02 M (NH~)HCO~, o i M NaC1 (pH 8.8). (a-c) Reduct ion and alkylat ion done according to STEERS 3, (d-f) done according to CRESTFIELD et al. TM. Condit ions of runs : 59 780 rev.]min, dia- phragm angle, 60 °, t empera tu re 18.5 ° (a-c), 19.6 ° (d-f). Pho tographs t aken i i , 27 and 43 min (a-c) and 32, 64 and 96 mm (d-f) after reaching speed. (d-f) .Neat and 1/2 concentra t ion runs. Sedimentat ion, left to right.
these solvents to be 2.1 S and 1. 9 S. In our hands reduced 5IA had an S2o,u, - ~ 2.1 S at a concentration of 9 mg/ml in either solvent system.
STEERS further reported a value of 35 ooo g/mole as the molecular weight of the reduced carboxymethylated product. The method of reduction and alkylation adopted by him was rather different from that used by us a,2°. We made several a t tempts to duplicate his technique but were unsuccessful in producing a homogeneous product. Fig. 7 compares the reduced carboxymethylated products produced by the two techniques. That some heterogeneity exists even in the carboxymethylated material produced by the method of CRESTFIELD et a l . TM is evident. Further evidence on this point comes from the high-speed equilibrium run (Fig. 4c). Point number average molecular weights ranged from 183 ooo to 240 ooo g/mole while point weight average molecular weights ranged from 189 ooo to 254 ooo g/mole (/~rw ---- 246 ooo g/mole and llT/z ~ 268 ooo g/mole.
As previously stated, an additional source of error in such determinations may be nonideal behavior of the solute which can cause a significant reduction in the apparent molecular weight. We believe that it is this which is responsible for the low molecular weight values near the bot tom of the cell. Nonetheless, one point is clear, neither the sedimentation-equilibrium data reported in Fig. 4, nor the intrinsic viscosities computed from the data in Fig. 3 are consistent with a nmltisubunit structure for 5IA, 5 IB or 5ID. In addition the starch-gel electrophoretic runs indicate that these antigens contain wholly or predominantly a single type of polypeptide.
Assuming the validity of our conclusion that the syngen 4 antigens are mono- meric, several points can be made concerning their synthetic control. I f serotypic transformation is controlled at the transcriptional level, a serotypic transformation
Bioch~m. B~o#hys. Acta, 188 (1969) 196-2o6

206 a . H . R E I S N E R el al,
requires only one type of cistron to be switched off and one to be switched on, ~.e. there is no requirement for synchronous switching of different types of cistrons (the argument holds as well t'or proteins composed of identidal subunits). In addition, unless some sort of cyclic synthetic pattern exists for producing these proteins, the messenge~ RNA's coding for these molecules would have molecular weights approaching IO ~ daltons and if we assume ribosomal spacing similar to that found in hemoglobin synthesis, a maximum of 8o-Ioo ribosomes would be associated with them. Were such polyribosomes to exist in paramecia, it seems not unlikely that they nlight be associated with and stabolized by the cell membrane. The fact that these proteins re.~ide on the cell surface also suggests that this may be the case, indeed it may be this situation which allows the large size of the monomers.
ACKNOWLEDGMENT~
We thank Misses S. Gibbes and R. McDougall for their able assistance and Miss C. Aylmer for her skillflfl darkroom work.
I I E F E R F . N C F.S
J . R. PREER, JR , m "1" T ('HEN, l~e~caJch in Proto:.olot~,v, Vol 3, l>er~ a n I ° n | ' r c s . , N e w Y o r k , 1968, p. 129
2 1. G. JONES, i3~ochcm. ] , 90 ( t905) I7 3 E STEERS, t3u~chcmlstry, t ( I905) IS00 4 [. FINGER, F. ONORATO, ('. HELLER ~.ND l-[. B. \VILcOX, ] 3lol. l¢v , l , I 7 (l<~W,I ,',f) 5 H I~'IACINDOE AND A 1-[ REISNER, Aztstraha~ ,[ Bzol N(z., 2o (19071 14[ 6 A. H. REISNER AND J. ]{OXVE, Naturc, 222 (1969) 558. 7 A H. ][{EISNER, J RowI~ AND H. M. ~IACINDOE, ,/. 31ol Btol., 32 (IQI)N) 507 8 D. A. YPHANTIS, Btochcmzslry, 3 (IOta4) 207. Q ~. J EDELSTEIN AND H K. ~CHACHMAN, ,[ Bzol. Chem., 242 (IQ07) 3ol).
IO E J. COHN AND J T ]~I)SALL, [~tIItclHs, ADlltH) Aclds a~td Pcplzdes, | <emhoh l , Nc~ Y o r k
1943. I I A. H. REISNER, J t,[OXVE ANI) |{ \ ¥ ~LEIGH, Btochem~strv, s u t m n t t e d to t p u b h c a t m n I2 F. J. CASTELLINO AND ]{ BARKER, tltochemzstry, 7 ( Ig0g) 22o7. 13 () ~MITHIES, (~. E CONNELL AND (; H DIXON, .4~11 .[. Hu~nan (;cm' t , 14 (IQO2I I 4 14 () SMITHIES, (]. E. CONNELL AND (; H. DIXON, ]. Mol I~ol , 2I (1O(>t>) 21 15 H. SVENSSON, A c t a C h e m S c a n d , 15 (I061) 325 16 H SVENSSON, Acta Chem. S c a n d , iO (t9(~2) 450. 17 H SVENSSON, Arch Bzochcm t3zophys., Nuppl. I (IQ62) 132 18 A M. CRESTFIELD, G MOORE AND \\7 I-[. STEIN, .[. Blol Chem., 23b (I(4(,3) *,22. 19 C TANFORD, I( KA~,VAHARA AND S LAPAJE, .] .4ttl Chem. ,~oc., 89 (I967) 72(). 20 C. B. ANFINSEN AND 1'; HABER, ./ Bull Chem., 230 ( t g 0 I ) ~30I. 21 J B CLEGG, ~I A NAU(,HTON ~N1) [) J . WEATHER.XLI_, [ .1Iol 1~,>! , t<l (I0661 ,~[ 22 J. P. WALLER, /. ~1ol 1¢~ol, ~o (IQO 4) 3IQ
Bu)ch*nl B~ophy.~ .-lcta, 1,~ (IO6ql too _,w~


















![PIG - enzymes. What are enzymes? [3] 3 of the following: Biological catalysts Globular proteins Increase the rate of (chemical) reaction Complimentary](https://img.pdfslide.us/doc/110x75/551456535503466d1a8b6238/pig-enzymes-what-are-enzymes-3-3-of-the-following-biological-catalysts-globular-proteins-increase-the-rate-of-chemical-reaction-complimentary.jpg)









