Embed Size (px)
Citation preview

0 1994 by The American Society for Biochemistry and Molecular Biology, Inc. T H E JOURNAL OF BloLocxcAL CHEMISTRY Vol. 269, No. 50, Issue of December 16, pp. 31383-31389, 1994
Printed in U.S.A.
ERGlO from Saccharomyces cerevisiae Encodes Acetoacetyl-CoA Thiolase*
(Received for publication, July 11, 1994, and in revised form, September 15, 1994)
Laree HiserS, Michael E. BassonO, and Jasper Rine From the Department of Molecular and Cell Biology, University of California, Berkeley, California 94720
Two recessive alleles of ERGlO and three temperature- sensitive recessive alleles of HMGl (3-hydroxy-3-methyl- glutaryl-CoA reductase isozyme 1) were isolated in a screen for mevalonate auxotrophs in Saccharomyces cerevisiae. The essential, single-copy ERGlO gene was cloned by complementation of the temperature-sensi- tive phenotype of erglo-21. The 1,194-base pair continu- ous open reading frame, encoding a 398-amino acid polypeptide with a calculated molecular mass of 41,681 daltons, was demonstrated to encode cytoplasmic aceto- acetyl-coA thiolase. Acetoacetyl-CoA thiolase activity corresponded to the number of copies of ERGlO present in cell extracts, and null alleles of ERGlO produced no detectable acetoacetyl-CoA thiolase enzyme activity. The deduced amino acid sequence was 40-95% identical to acetoacetyl-CoA thiolases from other organisms. This identity included the active site cysteines located at amino acids 91 and 384 in the Erg10 protein.
The mevalonate pathway is essential for life. The products of this pathway include all isoprene-containing compounds such as sterols, quinones, dolichol, and isopentenylated adenosine in tRNAs. In addition, several proteins are post-translationally modified by the isoprenoid groups farnesyl and geranylgeranyl. Mevalonate, the precursor of all isoprenoids, is formed in three steps as shown in Fig. 1.
There are two classes of thiolases which are distinguished by the chain length of their substrates for the thiolysis reaction. Acetoacetyl-CoA thiolases (EC 2.3.1.9) are specific for aceto- acetyl-coA, whereas 3-ketoacyl-CoA thiolases (EC 2.3.1.16) ex- hibit broad chain-length specificities for various P-ketoacyl- CoA substrates. These two classes may be further subdivided based on their subcellular localization. Acetoacetyl-CoA thio- lases are generally found in the cytoplasm, where they partici- pate in the mevalonate pathway, and in mitochondria, where they function in ketone body metabolism. However, a peroxiso- mal acetoacetyl-CoA thiolase has been reported (1). In contrast, 3-ketoacyl-CoA thiolases are localized to mitochondria and per- oxisomes and are involved in fatty acid 0-oxidation.
Two distinct acetoacetyl-CoA thiolase isozymes, cytoplasmic and mitochondrial, have been reported for Saccharomyces cer-
* This work was supported by National Institutes of Health Grant GM 35827, National Science Foundation predoctoral fellowships (to L. H. and M. E. B.), and a National Institutes of Environmental Health Sciences Mutagenesis Center Grant for core support. The costs of pub- lication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequencefs) reported in this paper has been submitted to the GenBankTMIEMBL Data Bank with accession number(s) L20428.
$ To whom correspondence should be addressed: Dept. of Molecular and Cell Biology, 401 Barker Hall, University of California, Berkeley, CA 94720. Tel.: 510-642-7041; Fax: 510-642-6420.
of Technology, Cambridge, MA 02139. 5 Present address: Dept. of Biology 68-422, Massachusetts Institute
evisiae (2). The cytoplasmic acetoacetyl-CoA thiolase is believed to act anabolically in the mevalonate pathway, whereas the mitochondrial acetoacetyl-CoA thiolase is available to regulate the pool sizes of acetoacetyl-CoA originating from fatty acid catabolism and of acetyl-coA entering the tricarboxylic acid cycle. The sizes of the two enzymes, as estimated by gel filtra- tion, are 140,000 daltons for the cytoplasmic isozyme and 65,000 daltons for the mitochondrial isozyme. The isozymes are further distinguished by differing abilities to utilize DTT’ as a thiol donor in vitro (3). No immunological or protein sequence information on the relatedness of the two isozymes exists.
Acetoacetyl-CoA thiolase genes have been isolated from sev- eral organisms. Acetoacetyl-CoA thiolase in bacteria functions in the biosynthesis of poly-0-hydroxybutyrate, a storage mole- cule. Bacterial genes encoding acetoacetyl-CoA thiolases have been isolated from Zoogloea ramigera (4) and Thiocystis viola- cea (5). Genes encoding mitochondrial acetoacetyl-CoA thio- lases have been identified in human (6) and rat (7). The prod- ucts of identified acetoacetyl-CoA thiolase genes from the yeasts Candida tropicalis (8) and Saccharomyces uvarum (9) are believed to be localized to the peroxisomes and cytoplasm, respectively.
Ergosterol is the major sterol product of the mevalonate pathway in the yeast S. cereuisiae, and ergosterol auxotrophy may result from loss-of-function mutations in more than a dozen genes. Previous work (10) identified erg10 mutations that are deficient in acetoacetyl-CoA thiolase activity. These mutants are viable when supplied with ergosterol through the growth medium. Because the synthesis of other essential com- pounds requires isoprene subunits, these mutations must not be null alleles. Thus, the null phenotype of the ERGlO gene was unknown. In addition, these mutations exhibited an unusual pattern of complementation that brought into question the number of genes at the ERGlO locus. We report the isolation of additional erg10 mutations using a more specific screen for mevalonate auxotrophs and establish that the ERGlO locus contains a single gene that is the structural gene for aceto- acetyl-coA thiolase.
EXPERIMENTAL PROCEDURES Media and Genetic Methods-Yeast rich medium, yeast minimal me-
dium (YM), and sporulation medium were prepared as described pre- viously (11). DL-Mevalonolactone (mevalonate; Sigma) was used in me- dia at 5 mg/ml as needed. Standard genetic methods (11) and molecular techniques (12) were performed essentially as described. Plasmids were isolated from yeast by the method of Hoffman and Winston (13).
Strains-The S. cereuisiae strains used in this study are listed in Table I. JRY1592 and JRY1595 were isogenic except at the MAT locus and were related to strain S288C. JRY1799 was also related to S288C. JRY1745, JRY1746, and JRY1747 were derived from JRY1595 by EMS
The abbreviations used are: DTT, dithiothreitol; EMS, ethyl meth- anesulfonate; kb, kilobase pair(s); HMG-CoA, 3-hydroxy-3-methylglu- taryl-CoA, Y M , yeast minimal medium; PD, parental ditype; T, tetra- type; NPD, nonparental ditype.
31383

31384 Acetoacetyl-CoA Thiolase from S. cerevisiae
mutagenesis. JRY1748 and my1749 were derived from JRY1592 by EMS mutagenesis. JRY1747 was backcrossed to JRY1799, and one of the segregants was crossed to MS127 to introduce leu2-3,112. JRY2478 and JRY4306 were segregants from this cross. erglo-21 in combination with trpl or tyrl greatly reduced viability and was lethal in most cases. Synthetic lethality between other mutations in the mevalonate path- way and mutations affecting biosynthesis of aromatic amino acids has been previously observed (14). JRY4145 and JRY4146 were segregants of a cross between JRY2478 and W303-1A. JRY4213 was formed by crossing JRY4145 to JRY4306. JRY4212 was created by introducing the URA3 gene at the ERGlO chromosomal locus ofmy4146 by integrative transformation (15) of pJR1374 in order to test allelism oferglo-21 and the gene that complemented erglo-21. Strains with a null allele of ERGlO (erglOA:LEU2) were constructed by replacing most of the ERG10 coding sequence with LEU2 (see Fig. 6). The 3.5-kb SstI-BamHI fragment containing eg1OA:LEUZ was then integrated into JRY4213, JRY4146 and JRY4145 by one-step gene replacement (16) to construct JRY4214, JRY4273, and JRY4274, respectively.
Mutagenesis-Cells were mutagenized as previously described (17). The mutagenized cells were stored at 4 "C in YM medium lacking a carbon source while their viability was determined: diluted aliquots of mutagenized and unmutagenized cells were plated onto YM medium, and the number of single colonies formed was counted after two days. Approximately 26% of the mutagenized cells were viable. The frequency of canavanine resistant mutants was determined by plating diluted aliquots of cells onto YM medium containing 60 pg/ml canavanine. The efficiency of mutagenesis, determined from the frequency of canavanine resistance due to mutation of the CAN1 gene, was approximately 1.3 x
Enzyme Assay-Cell cultures (100 ml) were grown in rich medium at room temperature with vigorous shaking to an absorbance of 2-3 at 600 nm. The cells were pelleted and washed twice with 10 ml of 50 mM Tris-HC1 buffer, pH 7.5. The cells were then resuspended in 1 volume of 50 mM Tris-HC1, pH 7.5. Two volumes of glass beads (0.5 mm; Biospec Products, Bartlesville, OK) and 1 mM phenylmethylsulfonyl fluoride were added, and the cells were broken by vortexing at high speed for five 1-min intervals separated by periods of cooling on ice. After a 1,000 x g centrifugation at 4 "C, the supernatant was subjected to centrifu- gation at 100,000 x g for 1 h at 4 "C. The supernatant fraction was then frozen and stored at -70 "C.
Acetoacetyl-CoA thiolase activity was measured spectrophotometri- cally by a modification of the procedure of Middleton and Tubbs (18). Activity was measured at room temperature in 1-ml assays containing 150 p~ CoA, 150 PM acetoacetyl-CoA, and 50 r m Tris-HC1, pH 8.2. In the indicated reactions 200 PM DTT was substituted for CoA. The reactions were started by adding cell extract, and the decrease in absorbance of acetoacetyl-CoA at 303 nm was followed in 5-s intervals using a Shimadzu UV-160 double beam spectrophotometer. One unit of activity was defined as 1 pmol of acetoacetyl-CoA consumed per min, using 3,600 as the molar extinction coefficient for acetoacetyl-CoA (19). Aque- ous stock solutions of CoA, acetoacetyl-CoA, and DlT were prepared at a concentration of 10 mM and stored at -20 "C. The pH of the CoA and acetoacetyl-CoA stock solutions was adjusted to approximately 4.6 with acetic acid, using bromcresol green as an indicator. Protein determina- tions were conducted using the Bio-Rad protein assay, using bovine serum albumin as the standard.
Lithium Acetate fiansformation-Strains containing the erglo-21 allele were observed to be somewhat resistant to transformation with DNA. This phenotype has been observed for other strains with muta- tions in the mevalonate pathway (14). A modification of the lithium acetate transformation method (20) was used for effcient transforma- tion of DNA. Loss of viability was minimized by using lower speed spins. The critical modifications were omitting the final washes and adding an equal volume of TE (10 mM Tris-HC1, pH 7.5,l mM EDTA) to the mixture of competent cells, DNA, and polyethylene glycol solution (36% poly- ethylene glycol M, 3,350,O.l M lithium acetate in TE) immediately prior to centrifugation at 800 x g for 10 s. Cells did not form a tight pellet following centrifugation, but did collect along one wall of the microcen- trifuge tube. Approximately three-fourths of the diluted polyethylene glycol solution was carefully removed, and the cells were plated in the remaining liquid. All incubations were at room temperature, and com- petent cells were prepared by incubation for 18-24 h in the lithium acetate solution.
Gel-transfer Hybridization-Yeast genomic DNA was prepared as described elsewhere (13). The DNA was digested to completion with BamHI, separated on a 0.8% agarose gel and evaluated by gel-transfer hybridization (21). The digested DNA was transferred to nitrocellulose (Schleicher & Schuell) and hybridized at 65 "C to 32P-labeled probe. The
mutants per viable mutagenized cell.
0 II
2 CH3C-CoA
2 Acetyl-coA
3 Acetoacetyl-CoA thiolase
0 I 1
0 II
CH3C"CH2-C-CoA
Acetoacetyl-CoA
CH3C-CoA HMG-CoA synthase
0 I I OH I II
0
-O-C-CH2-F-CH2-C-COA
CH3 3-Hydroxyd-methylglutalyl-CoA
(HMG-CoA)
1 HMG-CoA reductase
Mevalonate
FIG. 1. The early steps of the mevalonate pathway.
2.3-kb KpnI-XbaI ERGlO fragment was isolated from an agarose gel using the Geneclean kit (BIO 101, Inc., La Jolla, CA). This fragment served as the template for preparing the probe using the multiprime DNA labeling kit (Amersham International plc, Amersham, UK). The following washes were at 65 "C for 40 min each: three washes with 2 x SSC (20 x SSC was 3 M NaC1, 0.3 M sodium citrate, pH 7.01, 0.5% SDS and three washes with 1 x SSC, 0.5% SDS.
Plasmids and Plasmid Construction-pJR1368 was isolated from an S. cereuisiae genomic library (22) as described below. The 4.8-kb BamHI and 1.7-kb BglII fragments from the 10-kb pJR1368 insert (see Fig. 3) were placed into the BamHI site of pRS316 (CENGARSH.1 URA3) (23) to create pJR1369 and pJR1370, respectively. The 1.7-kb BgZII frag- ment was deleted from pJR1369 to create pJR1371. pJR1374, pJR1375, and pJR1377 were constructed by inserting the 2.3-kb KpnI-XbaI frag- ment from pJR1368 into the corresponding sites of pRS306 (URA3) (231, pRS316, and pEMBL19 (24), respectively. pJR1378 contained the 4.8-kb BamHI fragment of pJR1368 cloned into the BamHI site of YEp352 ( 2 - p origin URA3) (25). The 2.1-kb PuuII-BamHI fragment carrying LEU2 from pJJ252 (26) was inserted in place of the 0.8-kb Eco47III-BglII fragment of pJR1377, removing 109 base pairs upstream of the start codon and about 60% of the ERGlO coding sequence. The resulting plasmid carrying a null allele of ERG10 (erg1OA::LEUZ) was designated pJR1373 (see Fig. 6).
DNA Sequencing-Double-stranded and single-stranded templates were sequenced using the dideoxy chain-termination method as recom- mended in the Sequenase kit (U. S. Biochemicals, Cleveland, OH). Oligonucleotide primers were synthesized by Operon Technologies, Inc. (Alameda, CA). Sequence analysis was performed using Geneworks (IntelliGenetics, Inc., Mountain View, CA).
RESULTS
Isolation of Alleles of the Structural Gene for Acetoacetyl-CoA Thiolase-Mutations in the genes encoding or regulating the early enzymes of the mevalonate pathway (Fig. I) were isolated from a screen for mevalonate auxotrophs. Because strains lack- ing both isozymes of HMG-CoA reductase in s. cereuisiae (hmgl hmg2 double mutants) require supplementation of the growth medium with mevalonate for growth (271, it was ex- pected that mutations in the genes encoding acetoacetyl-CoA thiolase, HMG-CoA synthase or potential regulators of these

Acetoacetyl-CoA Thiolase from S. cerevisiae 3 1385 TABLE I
Strain list
Strain
JRY1592 JRY1595 JRY1745 JRY1746 JRY1747 JRY1748 JRY1749 JRY1799 JRY2478 JRY4145 JRY4146 JRY4212
JRY4213
JRY4214
JRY4273 JRY4274 JRY4306
MS127" W303-1A
Genotype
MATa ade2-101 his3A200 HMGl hmg2::HIS3 lys2-801 met2 ura3-52 MATa ade2-101 his3h200 HMGl hmg2::HIS3 lys2-801 met2 ura3-52 MATa ade2-101 his3A200 HMGl hmg2::HIS3 lys2-801 met2 ura3-52 erglo-20 MATa ade2-101 his3A200 hmg2::HIS3 lys2-801 met2 ura3-52 hmgl-6 MATa ade2-101 his3A200 HMGl hmg2::HIS3 lys2-801 met2 ura3-52 erglo-21 MATa ade2-101 his3A200 hmg2::HIS3 lys2-801 met2 ura3-52 hmgl-7 MATa ade2-101 his3h200 hmg2::HIS3 lys2-801 met2 ura3-52 hmgl-8 MATa ade2-101 his3A200 HMGl hmg2::HIS3 lys2-801 tyrl ura3-52 MATa ade2-101 leu2-3,112 lys2-801 met2 ergl0-21 MATa ade2 his3-11,15 leu2-3,112 ura3-1 MATa ade2 his3-11,15 leu2-3,112 lys2-801 met2 ura3-1 MATa ade2 his3-11,15 leu2-3,112 lys2-801 met2 ura3-l ERGlO(URA3) MATa ade2 his3-11,15 leu2-3,112 lys2-801 ura3-1 ERGlO MATa ade2 his3-11,15 leu2-3,112 LYS2 URA3 ERGIO MATa a d 2 his3-11,15 leu24112 lys2-801 ura3-1 erglOA::LEU2 MATa ade2 his3-11.15 leu2-3.112 LYS2 URA3 ERGlO MATa ade2 his3-11,15 leu2-3,112 lys2-801 met2 ura3-1 erglOA::LEU2 MATa ade2 his3-11,15 leu2-3,112 ura3-1 erglOA::LEU2 MATa ade2 his3-11,15 leu2-3,112 lys2-801 erglo-21 MATa ade2-IO1 leu2-3,112 trpl-A1 MATa ade2-1 his3-11,15 leu2-3,112 trpl-1 ura3-1
MS127 was provided by M. Rose, and W303-1A was provided by R. Rothstein. All other strains were constructed in the laboratory.
genes would cause the same phenotype. Haploid strains carry- ing a disrupted allele of HMG2 (JRY1592 and JRY1595) were mutagenized with EMS and screened for mevalonate auxotro- phy. Of approximately 60,000 colonies, one (JRY1745) required mevalonate for growth at both 23 and 34 "C and four (JRY1746, JRY1747, JRY1748, and JRY1749) were temperature-sensitive for mevalonate auxotrophy, requiring mevalonate for growth at 34 "C, but not at 23 "C.
The mutations causing mevalonate auxotrophy were ex- pected to affect one of the three enzymes responsible for the production of mevalonate. Because the mutagenized parent strains carried a disrupted copy of HMG2, the mutant strains were expected to be deficient in acetoacetyl-CoA thiolase, HMG-CoA synthase or HMG-CoA reductase isozyme 1 (Hmglp). Three of the temperature-sensitive candidates (JRY1746, JRY1748, and JRY1749) failed to complement a null allele of HMGl. These were assigned the allele designations shown in Table I. Previously identified alleles of HMGl exhibit intragenic complementation (28). All three hmgl mutations identified in this study were in the hmgl-3 complementation group. The mutations in JRY1745 and JRY1747 defined a sepa- rate complementation group. No acetoacetyl-CoA thiolase ac- tivity was detected in either JRY1745 or JRY1747; and neither strain was deficient for HMG-CoA synthase. These results sug- gested that JRY1745 and JRY1747 contained mutations in the acetoacetyl-CoA thiolase structural gene or in other genes re- sponsible for acetoacetyl-CoA thiolase synthesis or activity. Consistent with this conclusion, plasmids carrying a functional copy of either HMG-CoA synthase (ERG131 or HMG-CoA re- ductase (HMGl) failed to complement the mevalonate auxo- trophy of JRY1747. The alleles isolated in JRY1745 and JRY1747 were designated erglO-20 and erglo-21, respectively (see below), with erglo-21 being the temperature-sensitive allele.
The mutant phenotypes of JRY1745 and JRY1747 were re- cessive and the result of a mutation in a single nuclear gene. A poor growth phenotype co-segregated with erglo-20 and ergl0- 21. The temperature-sensitive mevalonate auxotrophy of ergl0-21 was partially suppressed by high concentrations of salt (100 mM CaC1, or 100 mM MgC1,).
Isolation of ERGlO from a Genomic Library-The wild-type ERGlO gene was isolated from an S. cerevisiae genomic library in a centromere-containing vector carrying the selectable LEU2
gene (22) by screening for clones that complemented the tem- perature-sensitive mevalonate auxotrophy of a strain with the erglo-21 mutation. One of the approximately 7,000 transfor- mants screened grew well at both 34 and 37 "C in the absence of mevalonate. Spontaneous loss of the complementing plas- mid, designated pJR1368, restored the temperature-sensitive phenotype. Strains with the ergl0-21 allele (e.g. JRY2478) be- came temperature-resistant in the absence of mevalonate when retransformed with the purified pJR1368 plasmid.
The cloned DNA fragment in pJR1368 was shown to contain the bona fide ERGlO gene by its ability to direct integration by homologous recombination at the chromosomal ERGlO locus. pJR1374, an integrating plasmid carrying ERGlO, was trans- formed into wild-type strains to produce an allele, designated ERGlO(URA3), which consisted of two intact copies of ERGlO separated by URA3 and vector sequence. DNA gel-transfer hy- bridization (see Fig. 7, lane 2 ) confirmed that the expected integration event had occurred. One ERGlO(URA3) transfor- mant (JRY4212) was crossed to an ergl0-21 ura3 strain (JRY4143). Co-segregation of the Ura- phenotype with the tem- perature-sensitive phenotype in a parental ditype pattern (2 Ts' Ura+:2 Ts- Ura-) was observed in all but one segregant from 73 tetrads with four viable spores. The tight linkage between erglo-21 and ERGlO(URA3) indicated that integration of the cloned DNA occurred at the ERGlO locus and demonstrated that the ERGlO gene had been isolated. In one tetrad, temper- ature-resistance segregated 3:l and Ura' segregated 2:2. The inferred genotype of the one unusual segregant was ERGlO ura3, as expected from gene conversion of ergl0-21.
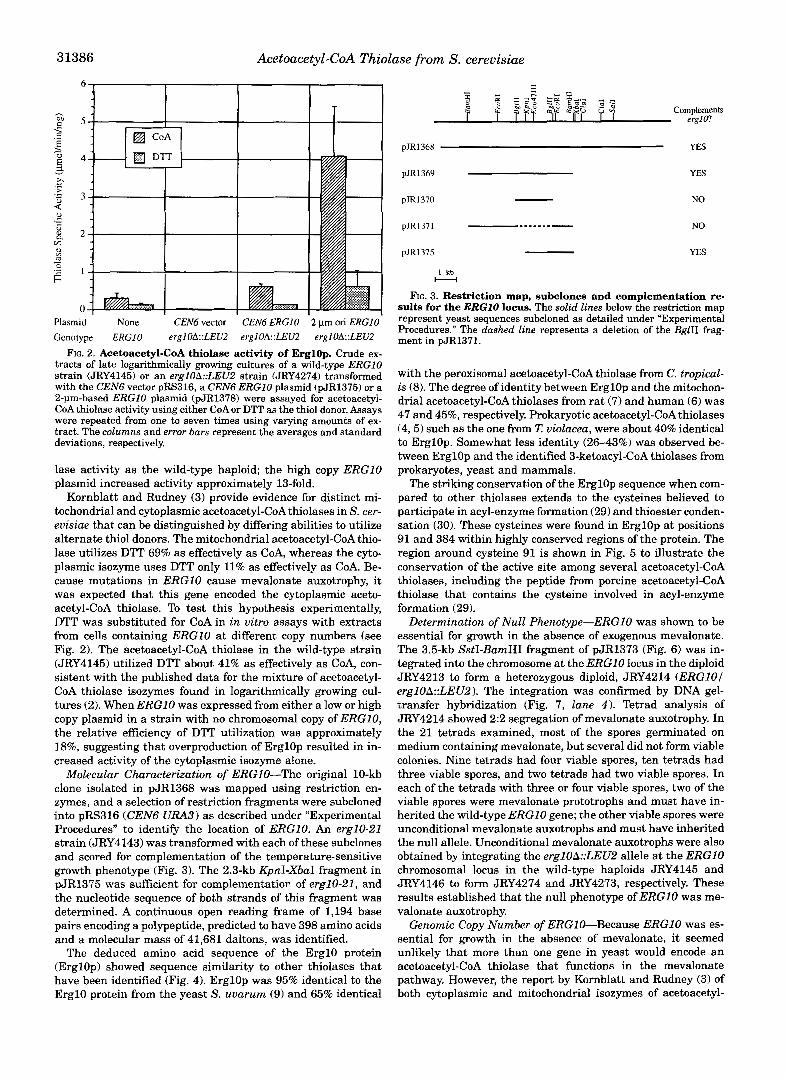
Correspondence of ERGlO Copy Number to Acetoacetyl-CoA Thiolase Actiuity-One indication that ERGlO was the struc- tural gene for acetoacetyl-CoA thiolase was that the level of enzyme activity corresponded to the copy number of the ERGlO gene. Astrain with a deletion of ERGlO (JRY4274; see Fig. 6 for a description of the allele) was transformed with pRS316 (CENG), pJR1375 (CEN6 ERG101 or pJR1378 (2-pm origin ERGlO). Crude extracts from these transformants and an iso- genic wild-type strain (JRY4145) exhibited levels of thiolase activity in vitro that correlated with the copy number of the ERGlO gene (see Fig. 2). No acetoacetyl-CoA thiolase activity was detected in the mevalonate auxotroph (erglOA::LEU2) transformed with the vector. ERGlO on a low copy plasmid produced approximately twice as much acetoacetyl-CoA thio-
31386 Acetoacetyl-CoA Thiolase from S. cerevisiae
Plasmid None . CEN6 vector CEN6 ERG10 -2 Km ori ERGlO
Genotype ERG10 erg1OA::LEUZ erglOA::LEU2 erglOA.:LEUt
FIG. 2. Acetoacetyl-CoA thiolase activity of ErglOp. Crude ex- tracts of late logarithmically growing cultures of a wild-type ERGlO strain (JRY4145) or an erglOA::LEU2 strain (JRY4274) transformed with the CENS vector pRS316, a CENS ERGlO plasmid (pJR1375) or a 2-pm-based ERGlO plasmid (pJR1378) were assayed for acetoacetyl- CoA thiolase activity using either CoA or DTT as the thiol donor. Assays were repeated from one to seven times using varying amounts of ex- tract. The columns and error bars represent the averages and standard deviations, respectively.
lase activity as the wild-type haploid; the high copy ERGlO plasmid increased activity approximately 13-fold.
Kornblatt and Rudney (3) provide evidence for distinct mi- tochondrial and cytoplasmic acetoacetyl-CoAthiolases in S. cer- evisiae that can be distinguished by differing abilities to utilize alternate thiol donors. The mitochondrial acetoacetyl-CoA thio- lase utilizes DTT 69% as effectively as CoA, whereas the cyto- plasmic isozyme uses DTT only 11% as effectively as CoA. Be- cause mutations in ERGlO cause mevalonate auxotrophy, it was expected that this gene encoded the cytoplasmic aceto- acetyl-coA thiolase. To test this hypothesis experimentally, DTT was substituted for CoA in in vitro assays with extracts from cells containing ERGlO at different copy numbers (see Fig. 2). The acetoacetyl-CoA thiolase in the wild-type strain (JRY4145) utilized DTT about 41% as effectively as CoA, con- sistent with the published data for the mixture of acetoacetyl- CoA thiolase isozymes found in logarithmically growing cul- tures (2). When ERGlO was expressed from either a low or high copy plasmid in a strain with no chromosomal copy of ERG10, the relative efficiency of DTT utilization was approximately 18%, suggesting that overproduction of ErglOp resulted in in- creased activity of the cytoplasmic isozyme alone.
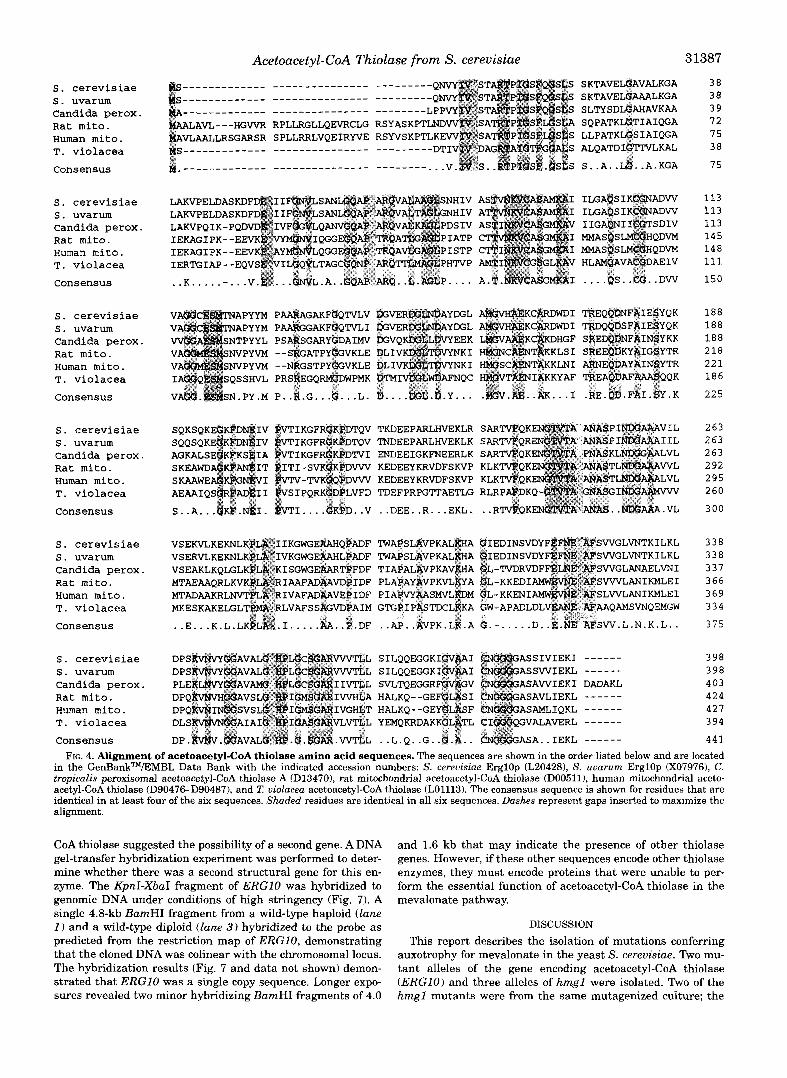
Molecular Characterization of ERGlO-The original 10-kb clone isolated in pJR1368 was mapped using restriction en- zymes, and a selection of restriction fragments were subcloned into pRS316 (CENG URA3) as described under “Experimental Procedures” to identify the location of ERGlO. An ergl0-21 strain (JRY4143) was transformed with each of these subclones and scored for complementation of the temperature-sensitive growth phenotype (Fig. 3). The 2.3-kb KpnI-XbaI fragment in pJR1375 was sufficient for complementation of ergl0-21, and the nucleotide sequence of both strands of this fragment was determined. A continuous open reading frame of 1,194 base pairs encoding a polypeptide, predicted to have 398 amino acids and a molecular mass of 41,681 daltons, was identified.
The deduced amino acid sequence of the ErglO protein (ErglOp) showed sequence similarity to other thiolases that have been identified (Fig. 4). ErglOp was 95% identical to the ErglO protein from the yeast S. uuarum (9) and 65% identical
I - I I
j a e X”
3 0 4 u =& g:: 3 I T 7 T II $ 1 Y I erglo?
I.
91 Complements
pJRl368 YES
pJR1369 YES
pJR1370 - NO
pJR1371 “””” NO
pJR1375 YES
H 1 kb
FIG. 3. Restriction map, subclones and complementation re- sults for the ERGlO locus. The solid lines below the restriction map represent yeast sequences subcloned as detailed under “Experimental Procedures.” The dashed line represents a deletion of the BglII frag- ment in pJR1371.
with the peroxisomal acetoacetyl-CoA thiolase from C. tropicul- is (8). The degree of identity between ErglOp and the mitochon- drial acetoacetyl-CoA thiolases from rat (7) and human (6) was 47 and 45%, respectively. Prokaryotic acetoacetyl-CoA thiolases (4,5) such as the one from T violacea, were about 40% identical to ErglOp. Somewhat less identity (2643%) was observed be- tween ErglOp and the identified 3-ketoacyl-CoA thiolases from prokaryotes, yeast and mammals.
The striking conservation of the ErglOp sequence when com- pared to other thiolases extends to the cysteines believed to participate in acyl-enzyme formation (29) and thioester conden- sation (30). These cysteines were found in ErglOp at positions 91 and 384 within highly conserved regions of the protein. The region around cysteine 91 is shown in Fig. 5 to illustrate the conservation of the active site among several acetoacetyl-CoA thiolases, including the peptide from porcine acetoacetyl-CoA thiolase that contains the cysteine involved in acyl-enzyme formation (29).
Determination of Null Phenotype-ERG10 was shown to be essential for growth in the absence of exogenous mevalonate. The 3.5-kb SstI-BamHI fragment of pJR1373 (Fig. 6) was in- tegrated into the chromosome at the ERGlO locus in the diploid JRY4213 to form a heterozygous diploid, JRY4214 (ERG101 erglOA::LEU2). The integration was confirmed by DNA gel- transfer hybridization (Fig. 7, lune 4). Tetrad analysis of JRY4214 showed 2:2 segregation of mevalonate auxotrophy. In the 21 tetrads examined, most of the spores germinated on medium containing mevalonate, but several did not form viable colonies. Nine tetrads had four viable spores, ten tetrads had three viable spores, and two tetrads had two viable spores. In each of the tetrads with three or four viable spores, two of the viable spores were mevalonate prototrophs and must have in- herited the wild-type ERGlO gene; the other viable spores were unconditional mevalonate auxotrophs and must have inherited the null allele. Unconditional mevalonate auxotrophs were also obtained by integrating the erglOA::LEU2 allele at the ERGlO chromosomal locus in the wild-type haploids JRY4145 and JRY4146 to form JRY4274 and JRY4273, respectively. These results established that the null phenotype of ERGlO was me- valonate auxotrophy.
Genomic Copy Number of ERG10-Because ERGlO was es- sential for growth in the absence of mevalonate, it seemed unlikely that more than one gene in yeast would encode an acetoacetyl-CoA thiolase that functions in the mevalonate pathway. However, the report by Kornblatt and Rudney (3) of both cytoplasmic and mitochondrial isozymes of acetoacetyl-
Acetoacetyl-CoA Thiolase from S . cerevisiae 31387
S. ce rev i s i ae S . uvarum Candida perox. Rat mito. Human mito. T. v io lacea consensus
S . ce rev i s i ae S . uvarum Candida perox. Rat mito. Human mito. T . v io lacea consensus
S. ce rev i s i ae S . uvarum Candida perox. Rat mito. Human mito. T . v io lacea Consensus
S . ce rev i s i ae S . uvarum Candida perox. Rat mito. Human mito. T . v io lacea consensus
S. ce rev i s i ae S . uvarum Candida perox. Rat mito. Human mito. T . v io lacea consensus
S . ce rev i s i ae S . uvarum Candida perox. Rat mito. Human mito. T. v io lacea Consensus
3 8 3 8 3 9 7 2 7 5 3 8
75
1 1 3 1 1 3 1 1 3 1 4 5 1 4 8 111
150
2 6 3 2 6 3 2 6 3 2 9 2 2 9 5 2 6 0
3 0 0
3 3 8 3 3 8 3 3 7 3 6 6 3 6 9 3 3 4
3 7 5
3 9 8 3 9 8 403 4 2 4 4 2 7 3 9 4
4 4 1
in the GenBankm/EMBL Data Bank with the indicated accession numbers: S. cereuisiae ErglOp (L20428), S. uuarum ErglOp (X07976), C. FIG. 4. Alignment of acetoacetyl-CoA thiolase amino acid sequences. The sequences are shown in the order listed below and are located
tropicalis peroxisomal acetoacetyl-CoA thiolase A (D134701, rat mitochondrial acetoacetyl-CoA thiolase (D00511), human mitochondrial aceto- acetyl-coA tholase (D90476-D90487), and T violacea acetoacetyl-CoA thiolase (L01113). The consensus sequence is shown for residues that are identical in at least four of the six sequences. Shaded residues are identical in all six sequences. Dashes represent gaps inserted to maximize the alignment.
CoA thiolase suggested the possibility of a second gene. A DNA gel-transfer hybridization experiment was performed to deter- mine whether there was a second structural gene for this en- zyme. The KpnI-XbaI fragment of ERGlO was hybridized to genomic DNA under conditions of high stringency (Fig. 7). A single 4.8-kb BamHI fragment from a wild-type haploid (lane 1 ) and a wild-type diploid (lane 3 ) hybridized to the probe as predicted from the restriction map of ERG10, demonstrating that the cloned DNA was colinear with the chromosomal locus. The hybridization results (Fig. 7 and data not shown) demon- strated that ERGlO was a single copy sequence. Longer expo- sures revealed two minor hybridizing BamHI fragments of 4.0
and 1.6 kb that may indicate the presence of other thiolase genes. However, if these other sequences encode other thiolase enzymes, they must encode proteins that were unable to per- form the essential function of acetoacetyl-CoA thiolase in the mevalonate pathway.
DISCUSSION
This report describes the isolation of mutations conferring auxotrophy for mevalonate in the yeast S. cereuisiae. Two mu- tant alleles of the gene encoding acetoacetyl-CoA thiolase (ERGIO) and three alleles of hmgl were isolated. Two of the hmgl mutants were from the same mutagenized culture; the
31388 Acetoacetyl-CoA Thiolase from S. cerevisiae
Pig active site peptide QAVLGAGLPC NTTTSPIIKV CASGMX S. cerevisiae (71-96) QVALAAGLSN H I V A S W V CASAMK S. uvarum (71-96) QVALTAGLGN HIVATTVNKV CASAMK Candida perox. (71-96) QVALKAGLPD SIVASTINKV CASGMK Rat mito. (103-128) QATLGAGLPI ATPCTTVNKV CASGMX Human mito. (106-131) QAT 'I STPCTTINKV CASGMK T. violacea (69-94) QT 'H TVPAMTINKV CGSGLK
Consensus Q..u.nUYC). ... A.TINKV CASGMK FIG. 5. Comparison of active site peptide sequences. The porcine active site peptide is aligned with the homologous regions of the other
acetoacetyl-CoA thiolases shown in Fig. 4. The numhcrs in parentheses indicate the amino acids shown in this alignment. The consensus sequence is shown for residurs that are identical in at least four of the seven sequences. Shadrd residues are identical in a11 seven sequences.
JLGAGLF I'LMAGLF
I kh
FIG. 6. The erglOAAEU2 allele contained in pJR1373. The Eco47III-R~III fragment of pJR1377 was replaced by thr PwII-HamHI fragment from pJ.12.52 to construct pJR137.7. The hold lines represent yeast sequence and the narrow I i n m represent vector sequence. The arrows show the direction and extent of open reading frame sequences.
1 2 3 4
4- 6.2
FIG. 7. Genomic DNAhyhridizetion to E R G I O . Lane 1 (JRY4146 ERGIO), lane 2 (JRY4212; k : l M ; l O ' ~ ~ l L \ 3 1 1. I nw 3 (JRY4213; ala BRCIOIERCIO), and Ianc 4 I,JI{Y4214; d n E R ~ l O I e r ~ l O ~ : : L E U 2 ) contain genomic DNA digested with RanzHI and hybridized to a :"!"laheled KpnI-XhnI ERG10 probe as drscrihed under "Experimental Procedures."
other three mutanL9 were independent. The frequency of recov- ery of mevalonate auxotrophs from this screen was approxi- mately 8 x IO-', more than an order of magnitude less than expected based upon the frequency ofcanl mutants in the same cultures. Because four of the five mutants required mevalonate only a t high temperature, the poor recovery of mutants may have been due to the poor viability of cells requiring meval- onate even when 5 mg/ml mevalonate was provided in the growth medium. Mevalonate was absent during the mutagen- esis and storage of cells prior to the screen. A reduced viability
of mevalonate auxotrophs under these conditions could also be responsible for the low number of mutants recovered.
Several observations led to the conclusion that ERG10 en- coded acetoacetyl-CoA thiolase. Comparison of the ErglOp se- quence to the sequences of other acetoacetyl-CoA thiolases showed 40-95% sequence identity along the entire length of the open reading frame. The region around the cysteine involved in acyl-enzyme formation (amino acid 91 in ErglOp) revealed greater similarity to acetoacetyl-CoA thiolases than to 3-keto- acyl-CoA thiolases (data not shown). There was no detectable acetoacetyl-CoA thiolase activity in crude extracts from ergl0- 20, erglo-21, or erg109::LEU2 strains. Enzyme activity in vitro correlated with the number of copies ofERGlO present (Fig. 2).
In all other organisms studied to date, the cytoplasmic and mitochondrial acetoacetyl-CoA thiolase isozymes are encoded by different genes. The phenotype of the identified erg10 mu- tants was mevalonate auxotrophy, and the cloned ERGlO gene satisfied the mevalonate requirement. These were the expected results for the gene encoding the cytoplasmic acetoacetyl-CoA thiolase, the isozyme involved in the mevalonate pathway. However, the possibility remained that both isozymes, or sub- units of both isozymes, were encoded by ERGIO. This possibil- ity was addressed by examining several physical and enzy- matic properties of the two isozymes by which they can be distinguished from one another.
In experiments with DTT substituted for CoA in in vitro acetoacetyl-CoA thiolase activity assays (Fig. 2), a wild-type haploid strain used DTT with an efficiency intermediate be- tween the reported values for the cytoplasmic and mitochon- drial isozymes (2). This result indicated that the assay detected activity from a mixture of the isozymes. Therefore, it was sig- nificant that no acetoacetyl-CoA thiolase activity was detected in an erglOA::LEU2 strain although fumarase, another mito- chondrial matrix enzyme, was present in the extract (data not shown). One interpretation of this result is that ERG10 also encodes the mitochondrial isozyme or an essential subunit of the mitochondrial isozyme. Another possible explanation is that ERG10 does not encode any part of the mitochondrial isozyme, but when the pool size of acetyl-coA is large due to a block in the utilization of acetyl-coA caused by mutation of the gene encoding cytoplasmic acetoacetyl-CoA thiolase, the mito- chondrial isozyme is down-regulated. This hypothesis is plau- sible because the cytoplasmic isozyme functions in the direction of acetoacetyl-CoA formation in vivo, whereas the mitochon- drial isozyme functions in the opposite direction, forming acetyl-coA. It is puzzling, however, that moderate overproduc- tion of ErglOp in an erglOA::LEU2 strain resulted in a DTT to CoA utilization ratio more similar to massive overproduction of ErglOp than to wild type. To date, we have been unable to isolate mitochondria with active acetoacetyl-CoA thiolase to examine this issue more directly.
The molecular characterization of ERGIO provided further evidence that it encodes the cytoplasmic isozyme. First, the

Acetoacetyl-CoA Thiolase from S. cerevisiae 31389
sequence of ErglOp did not reveal a leader sequence with any resemblance to mitochondrial targeting sequences, a feature of most proteins residing in the mitochondrial matrix. If ERGlO encodes all or part of a mitochondrial acetoacetyl-CoA thiolase, it must employ a different import mechanism than the charac- terized rat and human acetoacetyl-CoA thiolases. Second, the calculated molecular mass of ErglOp (41,681 daltons) was more consistent with reports for the cytoplasmic isozyme. In general, acetoacetyl-CoA thiolase is found as a homotetramer (31-35). If this structure applies to the yeast enzyme, then the predicted mass of the functional acetoacetyl-CoA thiolase encoded by ERGlO would be about 160,000 daltons. Kornblatt and Rudney (3) reported that the yeast cytoplasmic thiolase has a molecular mass of about 140,000 daltons, whereas the mitochondrial isozyme has an estimated molecular mass of 65,000 daltons. Third, ERGlO, like other genes in the mevalonate pathway, was demonstrated to be essential for viability in the absence of exogenous mevalonate.
Taken together, these results indicate that ERGlO encodes the cytoplasmic acetoacetyl-CoA thiolase. Further study will be necessary to determine whether the mitochondrial isozyme is encoded by weakly homologous DNA sequences or by an unre- lated gene, or is a modified form of ErglOp.
Acknowledgment-We thank Randy Hampton for critical reading of the manuscript.
REFERENCES
2. Kornblatt, J. A,, and Rudney, H. (1971) J. Biol. Chem. 246,4424-4430 1. Kurihara, T., Ueda, M., and Tanaka, A. (1988) FEBS Lett. 229,215-218
3. Kornblatt, J. A,, and Rudney, H. (1971) J. Biol. Chem. 246, 4 4 1 7 4 2 3 4. Peoples, 0. P., Masamune, S., Walsh, C. T., and Sinskey, A. J. (1987) J. Biol.
5. Liebergesell, M., and Steinbiichel, A. (1993) Appl. Microbiol. Biotechnol. 38,
6. Fukao, T., Yamaguchi, S., &no, M., Orii, T., Fujiki, Y., Osumi, T., and
7. Fukao, T., Kamijo, It, Osumi, T., Fujiki, Y., Yamaguchi, S., Orii, T., and
8. Kurihara, T., Ueda, M., Kanayama, N., Kondo, J., Teranishi, Y., and Tanaka,
Chem. 262,97-102
493-501
Hashimoto, T. (1990) J. Clin. Inuest. 86,208€&2092
Hashimoto, T. (1989) J. Biochem. (Tokyo) 106, 197-204
A. (1992) Eur. J . Biochem. 210,999-1005
9. Dequin, S., Gloeckler, R., Herbert, C. J., and Boutelet, F. (1988) Curr. Genet.
10. Servouse, M., Mons, N., Baillargeat, J.-L., and Karst, F. (1984) Biochem. Biophys. Res. Commun. 123,424-430
11. Rose, M. D., Winston, F., and Hieter, P. (1989) Laboratory Course Manual for Methods in Yeast Genetics, Cold Spring Harbor Laboratory, Cold Spring
12. Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning: A Harbor, NY
Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
13,471-478
13. Hoffman, C. S., and Winston, F. (1987) Gene (Amst.) 67,267-272 14. Gaber, R. F. Copple, D. M., Kennedy, B. K., Vidal, M., and Bard, M. (1989) Mol.
Cell. Biol. 9; 3447-3456 15. Hinnen, A,, Hicks, J. B., and Fink, G. R. (1978) Proc. Natl. Acad. Sci. U. S. A.
16. Rothstein, R. J. (1983) Methods Enzymol. 101, 202-210 17. Sherman, F., Fink, G. R., and Hicks, J. B. (1986) Laboratory Course Manual for
Methods in Yeast Genetics, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
75,1929-1933
18. Middleton, B., and Tubbs, P. It (1972) Biochem. J. 126,27-34
20. Ito, H., Fukuda, Y., Murata, K., and Kimura, A. (1983) J. Bacteriol. 163, 19. Stern, J. R. (1956) J. Biol. Chem. 221,3344
21. Southern, E. M. (1975) J. Mol. Biol. 98,503-517 22. Kouprina, N., Kroll, E., Bannikov, V., Bliskovsky, V., Gizatullin, R., Kirillov, A.,
Zakharyev, V., Hieter, P., Spencer, F., and Larionov, V. (1992) Mol. Cell. Biol.
163-168
. .
23. Sikorski, R. S., and Hieter, P. (1989) Genetics 122, 19-27 24. Dente, L., Cesareni, G., and Cortese, R. (1983) Nucleic Acids Res. 11, 1645-
12,5736-5747
1655 25. HiG, J E., Myers, A. M., Koerner, T. J., and Tzagoloff, A. (1986) Yeast 2,
26. Jones, J. S., and Prakash, L. (1990) Yeast 6, 363-366 27. Basson, M. E., Thorsness, M., andRine, J. (1986)Proc. Netl. Acad. Sci. U. S. A.
163-167
28. Basson, M. E., Moore, R. L., O'Rear, J., and Rine, J. (1987) Genetics 117, 83,5563-5567
645455 29. Gehring, U., and Hams, J. I. (1970) Eur. J. Biochem. 16,492-498 30. Palmer, M. A. J., Differding, E., Gamboni, R., Williams, S. E , Peoples, 0. P.,
Walsh, C. T., Sinskey, A. J., and Masamune, S. (1991) J. Biol. Chem. 266, 83694375
~~~ ~~.
31. Gehring, U., and Hams, J. I. (1970) Eur. J. Biochem. 16,487491 32. Davis, J. T., Moore, R. N., Imperiali, B., Pratt, A. J. , Kobayashi, K, Masamune,
S., Sinskey, A. J., Walsh, C. T., Fukui, T., and Tomita, K. (1987) J. Biol. Chem. 262,8249
33. Clinkenbeard, K. D., Sugiyama, T., Moss, J., Reed, W. D., and Lane, M. D.
34. Suzuki, F., Zahler, W. L., and Emerich, D. W. (1987) Arch. Biochem. Biophys. (1973) J. Biol. Chem. 248,2275-2284
35. Duncombe, G. R., and Frerman, F. E. (1976) Arch. Biochem. Biophys. 176, 254,272-281
159-170


















![ShopNotes Index [Vol. 1 - Vol. 82] & Synopsis](https://img.pdfslide.us/doc/110x75/55cf9d38550346d033acbbf3/shopnotes-index-vol-1-vol-82-synopsis.jpg)










