Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 258! No. 20, Issue of October 25, pp. 12265-12268.1983 Printed m U.S.A.
Subcellular Location of Chicken Brain Glutamine Synthetase and Comparison with Chicken Liver Mitochondrial Glutamine Synthetase*
(Received for publication, May 20,1983)
Darwin D. Smith, Jr. and James W. Campbell From the Department of Biology, Rice Uniuersity, Houston, Texas 77251
Chicken brain glutamine synthetase has been found to be localized in the cytosolic fraction of this tissue in contrast to its mitochondrial location in chicken liver. Despite this difference in subcellular distribution, the enzyme from brain exhibits the same molecular weight during sodium dodecyl sulfate-polyacrylamide gel electrophoresis, has the same isoelectric point, and is immunochemically cross-reactive. In addition, the im- munochemically isolated primary translation products for glutamine synthetase from brain and liver mRNA are the same size. These data suggest that cytosolic brain and mitochondrial liver glutamine synthetase may be derived from the same gene.
Ammonia detoxification and glutamate recycling in neural tissue is mediated primarily by the enzyme glutamine synthe- tase (EC 6.3.1.2) (1). Immunohistochemical studies indicate that this enzyme is distributed among the glial cells in rat brain (2, 3) and rat and chick retina (4, 5 ) . A cytosolic subcellular location of glutamine synthetase has been found in rat brain (6) and rat liver (7). However, contrary to the situation in ureoteles, glutamine synthetase is mitochon- drially located in the liver of uricoteles such as the chicken and snake (8). Recent studies in this laboratory have dem- onstrated that antibodies raised against the purified chicken mitochondrial enzyme are cross-reactive with both mitochon- drial and cytosolic glutamine synthetases spanning the phy- logenetic tree from shark to human (9). In addition, theoret- ical calculations using the amino acid residue content for the various glutamine synthetases indicate as high as 80% se- quence homology between the cytosolic mammalian enzymes and the mitochondrial chicken enzyme. Since such immuno- logical and sequence relatedness between enzymes located in distinctly different compartments is unusual, studies were initiated to determine if glutamine synthetase exists as a mitochondrial or cytosolic enzyme in the chicken brain. The following is a summary of the subcellular localization of chicken brain glutamine synthetase and a comparison with the purified mitochondrial enzyme.
EXPERIMENTAL PROCEDURES
Adult White Leghorn hens were maintained on a Purina chicken scratch grain diet for brain fractionation and brain mRNA isolation. Chickens were fed a United States Biochemical Corporation chicken diet supplemented with 60% protein for liver mRNA isolation.
Hen brains were prepared as 10% (w/v) homogenates in 5 mM HEPES,' 250 mM sucrose, and 1 mM EDTA near 0 "C and fraction-
* This work was supported by Grant PCM 79-25911 from the National Science Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "aduertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The abbreviations used are: HEPES, 4-(2-hydroxyethyl)-l-piper-
122
ated as described (8) with the modification of sedimenting all mito- chondria at 13,300 X g. Glutamine synthetase, cytochrome oxidase, lactate dehydrogenase, and glucose 6-phosphatase were assayed es- sentially as described (8) with the exception that the glutamine synthetase reaction mixture contained the ATP regeneration system recommended by Vorhaben et al. (10). Glutamate dehydrogenase activity was monitored by the oxidation of NADH at 340 nm as described (11).
One unit of activity is defined as 1 pmol of product produced per min at room (-24 "C) temperature for lactate dehydrogenase, cyto- chrome oxidase, and glutamate dehydrogenase and at 37 "C for glu- cose 6-phosphatase. One unit of glutamine synthetase activity is defined as 1 pmol of product produced per h at 37 "C. Protein was determined by the method of Bradford (12).
Immunoblotting was performed according to the method of Towbin et al. (13) using rabbit anti-chicken liver mitochondrial glutamine synthetase IgG (9), horseradish peroxidase conjugated goat anti- rabbit IgG and 4-chloro-1-napthol (Sigma) (14). Samples were elec- trophoresed in 10% SDS-PAGE slab gels, electrophoretically trans- ferred to nitrocellulose (Schleicher and Schuell, BA-85) and proteins were stained with Amido black. Isoelectric focusing was done in 5% polyacrylamide tube gels containing 2% ampholytes (Bio-Rad) (15). Detection of glutamine synthetase activity in isoelectric focusing gels was as described (16).
RNA was isolated from chicken liver and brain via the guanidine HC1 extraction technique of Deeley et al. (17) and mRNA was affinity purified from this fraction by oligo(dT) chromatography (18). Trans- lation of the mRNA utilized a cell-free rabbit reticulocyte system (19) containing 0.5 mM spermidine, 8 mM creatine phosphate, 25 p~ amino acids minus methionine, 2 mM dithiothreitol, 20 mM HEPES, pH 7.4, 0.65 mM magnesium acetate, 80 mM potassium chloride, 20 p1 of micrococcal nuclease-treated reticulocyte lysate, 25 p~ hemin, 15 pg of creatine phosphokinase, 50 pCi of [35S]methionine (Amer- sham Corp.), 10 pg/ml of each of the protease inhibitors leupeptin, bestatin, antipain, chymostatin, and pepstatin (Sigma) and 2-4 pg of mRNA in a final volume of 50 pl. Incubation was at 30 "C for 1 h.
Glutamine synthetase was immuno-isolated (20) from the trans- lation mixtures containing at least 1 X lo6 cpm by dilution to 1 ml final volume with 10 mM Tris-HC1, pH 7.4, 0.1% SDS, 0.5% Triton X-100, 150 mM NaC1, 10 mM EDTA, and 0.1 mM phenylmethylsul- fonyl fluoride. This solution was then precleared with 20 p1 of 10% (w/v) Staphylococcus aureus cells (The Enzyme Center) incubated at 4 "C for 15 min followed by removal of the cells by centrifugation. Anti-glutamine synthetase antiserum (5-10 p l ) was then added to the supernatant solution and incubated for 1 h at room temperature and at 4 "C overnight. Fifty p1 of 10% S. aureus cells were then added and the mixture was incubated at room temperature for 1 h after which the cells were washed with the dilution buffer 3 times and the cells were then prepared for SDS-PAGE. The radiolabeled proteins were visualized by impregnation of the cells with Enhance (New England Nuclear) and fluorography (21).
RESULTS AND DISCUSSION Table I illustrates the subcellular distribution of glutamine
synthetase in chicken brain. The majority of the activity (76%) is localized in the cytosolic fraction which is character- ized by the soluble marker enzyme lactate dehydrogenase; 64% of the lactate dehydrogenase was Dresent in this fraction.
~
azineethanesulfonic acid SDS-PAGE, sodium dodecyl sulfate-poly- acrylamide gel electrophoresis.
165
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from
12266 Chicken Brain Glutamine Synthetase TABLE I
Subcellular distribution of chicken brain glutamine synthetase
Fraction I,rotein Glutamine synthe- Lactate dehydrogen- Cytochrome oxidase Glutamate dehydro- Glucose 6-phospha- ~~~~ -
tase ase genase tase
rngfy tissue unitslg unitsfy
Homogenate 74.0 384.7 141.9 Nuclear 18.2 49.0 (13)” 33.7 (20) Mitochondrial 17.0 20.1 ( 5 ) 22.9 (13) Microsomal 7.0 22.1 (6) 5.4 (3) Soluble 33.1 287.6 (76) 107.9 (64) Total recovered 75.3 :m.8 (100) 169.9 (100)
unitsfg 38.3
7.2 (24) 21.4 (72)
1.0 (3) 0.0 (0)
29.6 (100) ” .~
unitsfg
6.3 (43) 8.3 (56) 0.1 ( 1 ) 0.0 (0)
14.7 (100)
14.5 unitsfg
3.0 (39) 2.5 (32) 2.2 (29) 0 (0) 7.7 1100)
14.7
Numbers in parentheses are the percentages of recovered activity.
mation may also trap cytosolic enzymes which could contam- inate mitochondrial and nuclear fractions.
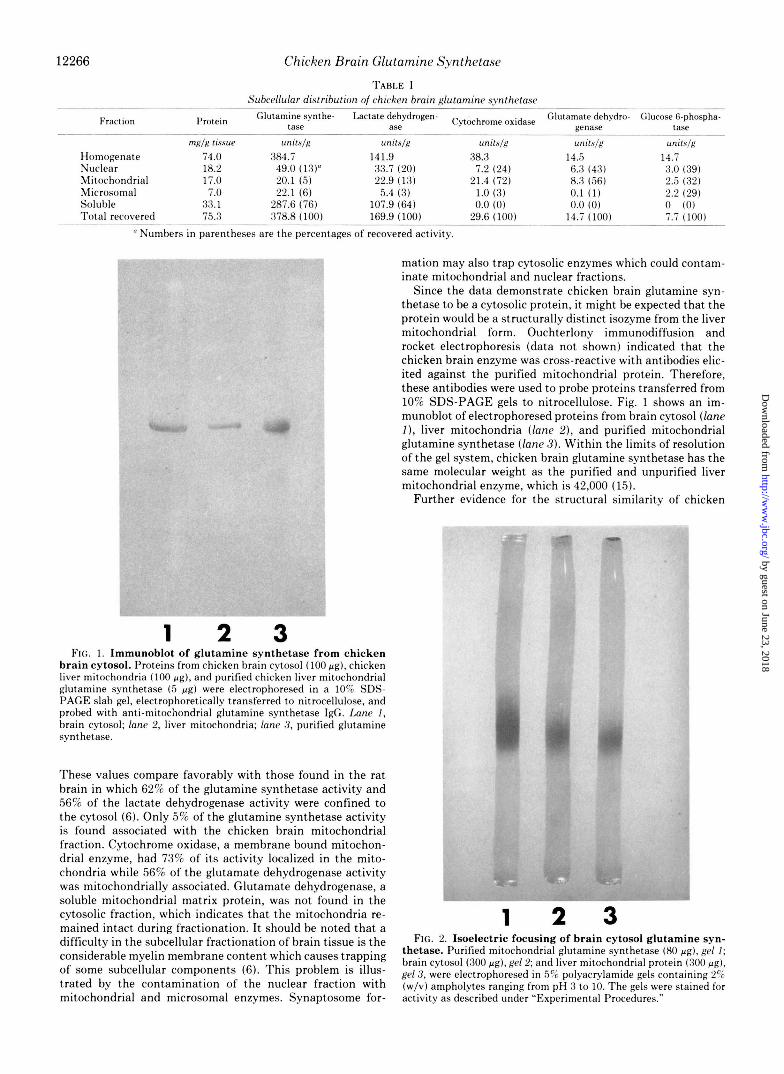
Since the data demonstrate chicken brain glutamine syn- thetase to be a cytosolic protein, it might be expected that the protein would be a structurally distinct isozyme from the liver mitochondrial form. Ouchterlony immunodiffusion and rocket electrophoresis (data not shown) indicated that the chicken brain enzyme was cross-reactive with antibodies elic- ited against the purified mitochondrial protein. Therefore, these antibodies were used to probe proteins transferred from 10% SDS-PAGE gels to nitrocellulose. Fig. 1 shows an im- munoblot of electrophoresed proteins from brain cytosol (lane I), liver mitochondria (lane Z), and purified mitochondrial glutamine synthetase (lane 3) . Within the limits of resolution of the gel system, chicken brain glutamine synthetase has the same molecular weight as the purified and unpurified liver mitochondrial enzyme, which is 42,000 (15).
Further evidence for the structural similarity of chicken
1 2 3 FIG. 1. Immunoblot of glutamine synthetase from chicken
brain cytosol. Proteins from chicken brain cytosol (100 pg) , chicken live: mitochondria (100 pa) , and purified chicken liver mitochondrial glutamine synthetase ( 5 p g ) were electrophoresed in a 10% SDS- PAGE slab gel, electrophoretically transferred to nitrocellulose, and probed with anti-mitochondrial glutamine synthetase IgG. Lane 1, brain cytosol; lane 2, liver mitochondria; lane 3, purified glutamine synthetase.
These values compare favorably with those found in the rat brain in which 62% of the glutamine synthetase activity and 56% of the lactate dehydrogenase activity were confined to the cytosol (6). Only 5% of the glutamine synthetase activity is found associated with the chicken brain mitochondrial fraction. Cytochrome oxidase, a membrane bound mitochon- drial enzyme, had 73% of its activity localized in the mito- chondria while 56% of the glutamate dehydrogenase activity was mitochondrially associated. Glutamate dehydrogenase, a soluble mitochondrial matrix protein, was not found in the cytosolic fraction, which indicates that the mitochondria re- mained intact during fractionation. It should be noted that a 1 2 3 difficulty in the subcellular fractionation of brain tissue is the FIG. 2. Isoektr ic focusing of brain cytosol glutamine SYn-
membrane ‘Ontent which trapping brain cytosol (300 pg) , gel 2; and liver mitochondrial protein (300 pg) , thetase. Purified mitochondrial glutamine synthetase (80 pg) , gel 1;
of subcellular components (6)- This problem is illus- gel 3, were electrophoresed in polyacrylamide gels containing 25; trated by the contamination of the nuclear fraction with (w/v) ampholytes ranging from pH 3 to 10. The gels were stained for mitochondrial and microsomal enzymes. Synaptosome for- activity as described under “Experimental Procedures.”
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

Chicken Brain Glutamine Synthetase 12267
brain and liver glutamine synthetase is presented in Fig. 2, in which the isoelectric focusing patterns of the cytosolic and mitochondrial enzymes are compared. The chicken brain en- zyme (gef 2) has the same isoelectric point as the purified ( g e f I ) and unpurified (ge l 3) liver mitochondrial enzyme (PI = 6.1) as shown by activity staining. This has been further substantiated (data not shown) by crossed immunoelectro- phoresis of similar isoelectric focusing gels into agarose gels containing anti-glutamine synthetase IgG in which the activ- ity staining and immunoprecipitin lines corresponded and were the same for the enzyme from both sources.
The high degree of structural and compositional homology of the cytosolic brain glutamine synthetase with its liver mitochondrial counterpart, apparent from the immunological and electrophoretic data, also holds true for the initial in vitro translation products as shown in Fig. 3. Current studies in this laboratory (22) indicate that the chicken liver glutamine synthetase subunit is initially synthesized as its 42,000-dalton mature form, while most mitochondrial matrix proteins are synthesized as higher molecular weight precursors (23, 24). Chicken liver glutamine synthetase is an octameric protein with eight identical 42,000-dalton subunits (15).
Comparison of the immunoprecipitated translation prod- ucts, from both liver mRNA ( fane 6) and brain mRNA ( fane .5) shows that the glutamine synthetase primary translation products are the same size. The controls of nonimmune anti- serum in fane 3 and competition with unlabeled mitochondrial glutamine synthetase in fane 4 confirm the identity and similarity of the brain glutamine synthetase in fane 5.
j_ D_
1 2 3 4 5 6 FIG. 3. Translation in vitro of chicken brain mRNA. Cell-
free translation products labeled with [‘%]methionine were electro- phoresed and fluorographed as described under “Experimental Pro- cedures.” Lane I, no added RNA; lane 2, total translation products from brain mRNA; lane 3 , immunoprecipitation of brain mRNA products using nonimmune antiserum; lane 4 , immunoprecipitation using monospecific anti-liver glutamine synthetase in the presence of competing levels (50 pg) of unlabeled, purified glutamine synthetase; lane 5 , immunoprecipitation as in lanc 4 omitting the unlabeled glutamine synthetase; [an(. 6, immunoprecipitation of glutamine syn- thetase programmed hv chicken liver mRNA.
Glutamine synthetase of adult chicken brain thus appears to be a cytosolic enzyme which is identical to the adult chicken liver mitochondrial enzyme based on the criteria of SDS- PAGE, isoelectric focusing, immunological reactivity, and primary translation products. This extent of homology is considered unusual for isozymes located in different subcel- lular compartments (25). One might expect that a difference would be observed in the primary translation products as in the case of chicken aspartate aminotransferase (26) and chicken phosphoenolpyruvate carboxykinase (27). The mito- chondrial and cytosolic forms of the two proteins are coded for by different genes and are not immunochemically cross- reactive. Although the subcellular fractionation studies for glutamine synthetase in other tissues has not been reported, Soh and Sarkar (28) mention that during the preparation of polysomes from cultured chick embryonic retina the mito- chondrial fraction had little glutamine synthetase activity while the cytosolic and polysomal fractions contained signif- icant activity. Since the purified enzyme from adult retina has been shown to also have a molecular weight of 42,000 (29), it may be possible that glutamine synthetase from retina is cytosolic as shown here for the brain enzyme.
Existence of a cytosolic glutamine synthetase in the chicken with physical properties the same as that of the mitochondrial enzyme suggests that the two are encoded for by the same gene and that their ultimate subcellular destination is tissue- specific and post-translationally determined by either subtle changes in protein structure or modification of mitochondrial competence for sequestering glutamine synthetase. Studies discerning this subcellular targeting problem in the chicken may help explain the molecular evolutionary divergence of glutamine synthetase with regard to ureotelism and uricotel- ism and also delineate some of the features necessary for import of mitochondrial proteins.
REFERENCES 1. Shank, R. P., and Aprison, M. H. (1981) Life Sci. 28,837-842 2. Martinez-Hernandez, A., Bell, K. P., and Norenberg, M. D. (1977)
3. Norenberg, M. D., and Martinez-Hernandez, A. (1979) Brain Res.
4. Riepe, R. E., and Norenberg, M. D. (1977) Nature (hnd.) 268,
5. Norenberg, M. D., Dutt, K., and Reif-Lehrer, L. (1980) J . Cell
6. Sellinger, 0. Z., and de Balbain Verster, F. (1962) J . Biol. Chem.
7. Wu, C. (1963) Riochim. Biophys. Acta 77,482-493 8. Vorhaben, J. E., and Campbell, d. W. (1972) J . Riol. Chem. 247,
9. Smith, D. D., <Jr., Vorhaben, J . E., and Campbell, J. W. (1983) J .
10. Vorhaben, J. E., Wong, L., and Campbell, J. W . (1973) Biochem.
11. Casey, C. A., Perlman, D. F., Vorhaben, J . E., and Campbell, J.
12. Bradford, M. M. (1976) Anal. Riochem. 72. 248-254 13. Towbin, H., Staehlin, T., and Gordon, J . (1979) Proc. Natl. Acad.
14. Hawkes, R., Niday, E., and Gordon, J. (1982) Anal. Biochem.
15. Vorhaben, J . E., Smith, D. D.. and Campbell, J. W. (1982) Int. J . Biochem. 14, 747-756
16. Vorhaben, J . E., and Campbell, J . W. (1977) J. Cell Bioi. 73,300- 310
17. Deeley, R. G., Gordon, J. I., Burns, A. T. H., Mullinix, K. P., Bina-Stein, M., and Goldberger, R. F. (1977) J. Bioi. Chem.
18. Aviv, H., and Leder, P. (1972) Proc. Natl. Acad. Sci. L! S. A. 69,
19. Pelham, H. R. H., and Jackson, R. ,J. (1976) Bur. J. Biochem. 67,
Science (Wash. D. C . ) 195, 1356-1358
161,303-310
654-655
Bioi. 84,803-807
237,2836-2844
2763-2767
EXP. Zed. 226, 29-35
J . 135,893-896
W. (1983) Mol. Physiol. 3, 107-126
Sci. U. S. A. 76, 4350-4354
119, 142-147
252.8310-8319
1408-1412
247-256
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

12268 Chicken Brain Glutamine Synthetase 20. Kessler, S. W. (1975) J. Immunol. 115, 1617-1624 21. Bonner, W. M., and Laskey, R. A. (1975) Eur. J. Biochem. 4 6 ,
22. Smith, D. D., Jr., and Campbell, J. W. (1983) Fed. Proc. 42 , 914 23. Sabatini, D. D., Kreibich, G., Morimoto, T., and Adesnik, M.
24. Schatz, G., and Butow, R. A. (1983) Cell 32 , 316-318 25. Dixon, M., and Webb, E. (1979) in The Enzymes, 3rd ed., pp.
83-88
(1982) J. Cell Biol. 92 , 1-22
640-646, Academic Press, New York
J. Biol. Chem. 257,13334-13340 26. Junssi, R. Sonderegger, P. Fluckiger, J., and Christen, P. (1982)
27. Hod, Y., Utter, M. F., and Hanson, R. W. (1982) J. Biol. Chem.
28. Soh, B. M., and Sarkar, P. K. (1978) Deu. Biol. 64,316-328 29. Sarkar, P. K., Fischman, D. A,, Goldwasser, E., and Moscona, A.
257,13787-13794
A. (1972) J. Biol. Chem. 247,7743-7749
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from

D D Smith, Jr and J W Campbellchicken liver mitochondrial glutamine synthetase.
Subcellular location of chicken brain glutamine synthetase and comparison with
1983, 258:12265-12268.J. Biol. Chem.
http://www.jbc.org/content/258/20/12265Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/258/20/12265.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 23, 2018http://w
ww
.jbc.org/D
ownloaded from





























