Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY C) 1991 by The American Society for Biochemistry and Molecular Biology, Inc.
Val. 266, No. 22, Issue of August 5, pp. 14367-14370,1991 Printed in U S A .
Expression, Purification, and Crystallization of the Adipocyte Lipid Binding Protein*
(Received for publication, March 18, 1991)
Zhaohui Xu$, Melissa K. Buelts, Leonard J. BanaszakS, and David A. BernlohrsT From the Departments of Biochemistry, $Medical School, University of Minnesota, Minneapolis, Minnesota 55455 and the §College of Biological Sciences, University of Minnesota, St. Paul. Minnesota 55108
The murine adipocyte lipid binding protein (ALBP/ aP2) has been cloned and expressed in Escherichia coli, purified to homogeneity, biochemically characterized, and crystallized for x-ray diffraction study. In the cloning, the ALBP coding region was placed under control of the recA promoter and downstream of the phage T7 g-10 translation enhancer sequence. Nali- dixic acid (50 pg/ml) induced the expression of ALBP 20-fold over that attained using the pT7 system pre- viously reported (Chinander, L. L., and Bernlohr, D. A. (1989) J. Biol. Chem. 264, 19564-19572). Recom- binant ALBP was purified to homogeneity using a combination of pH fractionation, gel filtration, and immobilized metal affinity chromatography. The flu- orescent affinity ligand 12-(9-anthroyloxy)oleic acid bound to homogeneous ALBP with an apparent Kd of 0.5 p ~ . rALBP was devoid of endogenous fatty acid, and oleic acid inhibited cysteine 117 modification by 5,5’-dithiobis-(2-nitrobenzoic acid) indicating integ- rity of the binding domain. Recombinant ALBP was phosphorylated by the soluble kinase domain of the insulin receptor with a VmaX of 11 nmol*min*mg of kinase and an apparent K,,, of 270 p ~ . Purified protein was crystallized using the hanging drop method with seeding. Crystalline ALBP was orthqrhombic with cqll dimensions of a = 34.4 A, b = 54.8 A, and c = 76.3 A. The space group was P212121, and there was one mol- ecule per asymmetric unit.
Intracellular hydrophobic ligand binding proteins are thought to be responsible for the uptake, transport, and metabolism of lipids. Different cell types appear to have different, but related, lipid binding proteins presumably ful- filling cell-specific functions. Fatty acid binding proteins are found in a variety of tissues including intestine, liver, heart, mammary, Schwann cell, and adipose (1-6). The adipocyte protein termed adipocyte lipid binding protein, abbreviated ALBP’ or aP2, is a fat cell specific protein that is considered
* This work was supported by National Institutes of Health Grant GM13925 (to L. J. B.), a grant from the American Diabetes Associa- tion, and a Presidential Young Investigator Award (to D. A. B.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1 To whom correspondence and reprint requests should be ad- dressed Dept. of Biochemistry, University of Minnesota, 1479 Gort- ner Ave., St. Paul, MN 55108. Tel.: 612-624-2712; Fax: 612-625-5780.
’ The abbreviations used are: ALBP, adipocyte lipid binding pro- tein; FABP, fatty acid binding protein; IFABP, intestinal FABP; rALBP, recombinant ALBP; IDA, iminodiacetic acid; SDS, sodium dodecyl sulfate; HEPES, 4-(2-hydroxyethyl)-l-piperazineethanesul- fonic acid; HPLC, high performance liquid chromatography; PAGE, polyacrylamide gel electrophoresis; DTNB, 5,5’-dithiobis-(2-nitro- benzoic acid); 12-AO, 12-(9-anthroyloxy)oleic acid.
to facilitate the bidirectional flux of fatty acids into and out of the adipocyte in response to insulin and epinephrine, respectively (6, 7) .
It has been shown that in 3T3-Ll adipocytes ALBP is phosphorylated on Y19 in response to insulin (8,9). Recently, insulin-stimulated tyrosyl phosphorylation of ALBP has been reconstituted i n vitro using homogeneous ALBP and lectin- Sepharose-enriched insulin receptor (10). The characteristics of ALBP phosphorylation in vitro are very similar to those seen in si tu leading to the consideration that ALBP is a primary substrate of the insulin receptor kinase. Because insulin regulates lipid metabolism in adipocytes, the flux of fatty acids into and out of the fat cell may be regulated by phosphorylation of the lipid carrier, ALBP. This hypothesis suggests some structural relationship between the site of phosphorylation, Y19, and the ligand binding domain.
Crystal structures of two related FABPs have been deter- mined (11, 12). The intestinal fatty acid binding protein (IFABP) has 27% identity and myelin P2 protein 69% identity to ALBP. IFABP and P2 have very similar conformations. They have a single domain with secondary structures largely consisting of two 5-stranded orthogonal antiparallel (3-sheets. The hydrophobic ligand binding site resides in the center of the molecule shielded from the aqueous environment (11, 12). The carboxylate of the bound fatty acid is coordinated with arginine residues in the internal cavity of the protein.
To address the structure-function relationship between ty- rosine phosphorylation and lipid binding and to define the ligand binding region, the structure of ALBP must be deter- mined. We report here the overexpression of recombinant ALBP in Escherichia coli, the characterization of its ligand binding and phosphorylation properties, the preparation of suitable crystalline specimens, and preliminary x-ray data from the resulting crystals.
EXPERIMENTAL PROCEDURES
Materials-”2,51-Labeled protein A (200 Ci/mmol) was obtained from Du Pont-New England Nuclear. [-y-:”PP]ATP was obtained from Amersham. Oleic acid was purchased from Nu Check Prep, Inc. (Elysian, MN). 12-(9-Anthroyloxy)oleic acid was purchased from Molecular Probes Inc. Bacto-tryptone, yeast extract, and casamino acids were purchased from Difco Laboratories. Sephadex G-75, IDA- Sepharose 6B, nalidixic acid, and 5,5’-dithiobis-(2-nitrobenzoic acid) were obtained from Sigma. All components for the polymerase chain reaction were from Perkin-Elmer. Deoxyoligonucleotides were syn- thesized by the University of Minnesota Microcbernical facility. Plas- mid pMON-5840 was generously provided by Dr. Jeffrey Gordon, Washington University (St. Louis, MO) and Dr. Peter Olins, Mon- santo Co. (St. Louis, MO).
Molecular Cloning-Recombinant cDNA clone pT7-5 (IO), encod- ing the full length insert corresponding to murine ALBP, was digested with PuuII, and the coding region was amplified via the polymerase chain reaction. T o facilitate molecular cloning, two oligonucleotides that bracket the coding region were synthesized. A 15-base deoxyoli-
14367
by on August 28, 2007
ww
w.jbc.org
Dow
nloaded from

14368 Crystallization of ALBP gonucleotide (5' CCTCTAGAAAGAAGG 3') corresponding to part of the E. coli ribosome binding site and including an XbaI site was utilized as the 5' primer. A 23-base 3'-deoxyoligonucleotide (5' TCTAGACCATCCAGGCCTCTTCC 3') was synthesized to anneal downstream of the translation stop codon and generated a second XbaI site. Amplification of the ALBP coding region by polymerase chain reaction was performed for 30 cycles. Each cycle utilized a 94 "C denaturation for 1 min, 55 "C annealing for 1 min, 72 "C extension for 1 min, and a final 5-min 72 "C elongation during cycle 30. The final product was extracted once with phenol/chloroform and precipitated with ethanol. The DNA was resolubilized with 10 mM Tris-HC1, 1.0 mM EDTA, pH 8.0, and subjected to restriction endo- nuclease digestion with XbaI.
A 450-base pair XbaI fragment was purified by electroelution from 1% agarose gel. The recovered DNA was ligated with dephosphoryl- ated XbaI-linearized pMON 5840 and transformed into E. coli JM101. The transformants were selected at 37 "C on the basis of ampicillin resistance, and productive transformants were identified by small scale digestions of selected colonies (13). All transformants harboring inserts were verified by restriction digest, and the insert-amplified DNA was sequenced to ensure identity.
Expression-E. coli strains harboring plasmid pMON containing the lipid binding protein insert were inoculated into 5 ml of LB medium containing 100 pg/ml ampicillin and grown overnight. The overnight culture was diluted 100-fold into 20 ml of M9 minimal salt medium supplemented with the following: 0.8% glucose, 1% casamino acids, 0.0005% thiamine, and 100 pg/ml ampicillin and grown a t 37 "C. At an Asso = 0.5, 50 pg/ml nalidixic acid was added, and, 4 h later, the cells were harvested by centrifugation. The pelleted cells were suspended in 200 pl of 0.125 M Tris-HC1, pH 6.8, and sonicated. The supernatant was obtained by centrifugation, and the proteins were separated by 5-15% SDS-polyacrylamide gradient gel electro- phoresis. The separated proteins were transferred to nitrocellulose membrane, and immunoblotting was as described by Chinander et al. (10). Murine ALBP was used as a positive control for ALBP expres- sion. The colony identified by immunochemical analysis with the greatest ALBP expression was termed pMON-rALBP.
Purification-Large scale purification of ALBP was accomplished from 5-liter cultures of E. coli JMlOl harboring pMON-rALBP. The cells were grown as described above on minimal medium and har- vested by centrifugation a t 4 "C for 5 min a t 8,000 rpm in a Beckman JA-10 rotor. Typically, 20 g of cell paste was recovered from 5 liters of culture. 20 ml of buffer A, containing 25 mM imidazole HC1, pH 7.0, 50 mM NaCl, 1 mM 2-mercaptoethanol, 5 mM EDTA, and 0.1 mM phenylmethylsulfonyl fluoride was added to the cell paste. The cell slurry was sonicated, and the cell debris was removed by centrif- ugation at 4 "C for 30 min at 12,000 rpm. The soluble extract was recovered, and protamine sulfate was added over a 20-min period to a final concentration of 1% from a 5% stock prepared in buffer A. The mixture was stirred for another 20 min a t 4 "C and then subjected to centrifugation a t 4 "C for 20 min at 8,000 rpm. The protamine sulfate supernatant was titrated to pH = 5.0 with 2.0 M NaAc, pH 5.0, and allowed to stir overnight. The insoluble material was removed by centrifugation at 4 "C for 20 min a t 8,000 rpm. The soluble extract was concentrated a t room temperature using an Amicon ultrafiltra- tion device equipped with a YM5 filter. The concentrated extract, about 15 ml, was applied to a Sephadex G-75 column (5 X 100 cm) equilibrated with 12.5 mM HEPES, pH 7.5, 0.25 M NaCl (buffer B). Recombinant ALBP was identified by SDS-PAGE. Those fractions containing ALBP were pooled (about 150 ml) and concentrated as described above. Concentrated protein solution was applied to a 10- ml Cu2+-IDA Sepharose 6B column which was equilibrated with Buffer B. The column was discharged with excess Buffer B, and the fractions were pooled. After each use, the column was washed with 0.5 M EDTA. Homogeneous ALBP identified by its migration on SDS-PAGE was found in the breakthrough fractions. 20-50 mg of homogeneous ALBP were obtained from a 5-liter fermenter run.
Ligand Binding-Fatty acid binding to recombinant ALBP was assessed by employing the fluorescent probe 12-(9-anthroyloxy)oleic acid (12-AO). The concentration of 12-A0 was determined using t:iX:,nm = 6,400 M" in absolute ethanol and diluted into buffer with stirring. ALBP (0 to 5 p ~ ) in 25 mM HEPES, pH 7.5, was incubated with 50 nM 12-AO, and probe fluorescence was recorded using a Perkin-Elmer 650-10s fluorescence spectrophotometer. Fluorescence was measured at 25 "C using an excitation wavelength of 383 nm and an emission wavelength of 460 nm, with slit widths at 4 nm. All manipulations were done in dim light.
Sulfhydryl Modification-ALBP (5 p ~ ) in 50 mM Tris-HC1, pH
8.0, 100 mM NaCl was reacted with 40 p~ DTNB in the presence or absence of 50 p~ oleic acid as described. Sulfhydryl titration was monitored spectrophotometrically, calculating the extent of modifi- cation from the difference in absorbance a t 412 nm ( e = 13,600 M-') between a protein-containing sample and a buffer blank (14).
I n Vitro Phosphorylation of ALBP-Phosphorylation of ALBP was examined using the 48-kDa soluble cytoplasmic domain of the human insulin receptor (15,16) kindly provided by Dr. Leland Ellis, Howard Hughes Medical Institute, Dallas, TX. Kinase (0.25 pg) was auto- phosphorylated in 25 mM HEPES, pH 7.5, containing 5 mM MnC12 and 100 p M [-y-32P]ATP. Autophosphorylation was shown to be complete within 30 min. The kinase was then added to tubes contain- ing varying concentrations of ALBP (50 to 500 p~ final) and incu- bated for the specified times. Reactions were stopped with Laemmli sample buffer (17), and the proteins were separated by SDS-PAGE. Proteins were visualized by staining with Coomassie brilliant blue, and the phosphoproteins were located by autoradiography a t -70 "C using Kodak XAR5 film. To determine the amount of 32P incorpo- rated, the protein bands were excised from the dried gel, and radio- activity was quantitated using a Beckman 3810 liquid scintillation counter.
Crystallization-Small crystals (0.05 X 0.1 X 0.1 mm) were obtained using the hanging drop/vapor equilibration method (18). 10-pl drops of 2.5 mg/ml ALBP in a 0.05 M Tris, 60% ammonium sulfate, 1 mM EDTA, 1 mM dithiothreitol, 0.05% sodium azide buffer with a pH of 7.0 (crystallization buffer) were suspended over wells containing the same buffer with varying concentrations of ammonium sulfate, from 75 to 85% saturation. Small, well shaped crystals were formed within a month at an 80% saturation and 19 "C. These crystals were isolated, washed with mother liquid, and used as seeds by transferring them into a 10-pl drop of 4 mg/ml fresh ALBP in the 80% saturation crystallization buffer over a well containing the same buffer. Large crystals, 0.3 X 0.4 X 0.4 mm, grew in 2 days at a constant temperature of 19 "C.
Data Collection and Processing-Crystals were analyzed with the area detector diffractometer from Siemens/Nicolet. A 0.8-mm colli- mator was used, and the crystal to detector distance was set at 12 cm with the detector midpoint a t 28 = 15". One I$ scan totaling 90" and three w scans of 68" with x a t 45 were collected with the Rigaku Ru200 operating at 50 kV and 180 mA. Each frame consisted of a 0.25" rotation taken for 120 s. The diffractometer data were analyzed with the Xengen package of programs (19). Raw data within 50 frames were searched to find about 100 strong reflections which were then indexed, and the cell dimensions were refined by least square methods. Data from different scans were integrated separately and then merged together.
RESULTS AND DISCUSSION
Murine ALBP cDNA was initially cloned and expressed in E. coli using the T7 system of Tabor and Richardson de- scribed in Ref. 10. However, the yield from this system was far below that necessary for crystallization trials. Thus, we turned our attention to the newly developed "g-10 leader" expression system. This expression system utilizes the phage T7 gene-10 product translation enhancer (20). The g-10 leader was placed upstream of the conventional E. coli ribosome binding site. Using this upstream translational enhancer un- der control of the recA promoter, ALBP was overexpressed to a much larger extent than using the E. coli ribosome binding site alone. Induction of the recA promoter by nalidixic acid resulted in the cytoplasmic expression of ALBP. This was verified by SDS-PAGE and immunoblotting (results not shown).
A purification protocol for recombinant ALBP had been established by Chinander and Bernlohr (10). An initial pH 5.0 fractionation was followed by Sephadex G-75 chromatog- raphy and CM-3SW cation exchange HPLC column. The purification was monitored by SDS-polyacrylamide gel elec- trophoresis and immunochemical analysis. However, the pro- cedure was time-consuming and difficult to scale up given the use of HPLC ion exchange chromatography. As an alternative to the existing protocol, we used immobilized metal affinity chromatography as the last step in the purification process.
by on August 28, 2007
ww
w.jbc.org
Dow
nloaded from
Crystallization of ALBP 14369
In immobilized metal affinity chromatography, proteins are fractionated according to their affinity for transition metal ions, largely determined by their surface histidine content (21, 22). In the case of murine ALBP, the protein has no histidine residues suggesting that purification could be achieved by adsorption of contaminants onto a Cu2+-based affinity matrix.
Pooled protein fractions from the G-75 column were loaded onto the charged metal chelated column. ALBP flowed through the column, eluting during the washing of the resin, while most of the contaminant proteins did not elute until after the column was stripped with 0.5 M EDTA. SDS-gel electrophoresis showed homogeneous ALBP in the flow- through fraction. Compared to the pT7-5 vector and coupled with the improved purification scheme, a 20-fold improved yield was achieved by using the new pMON-rALBP expres- sion system. These results are summarized in Fig. 1.
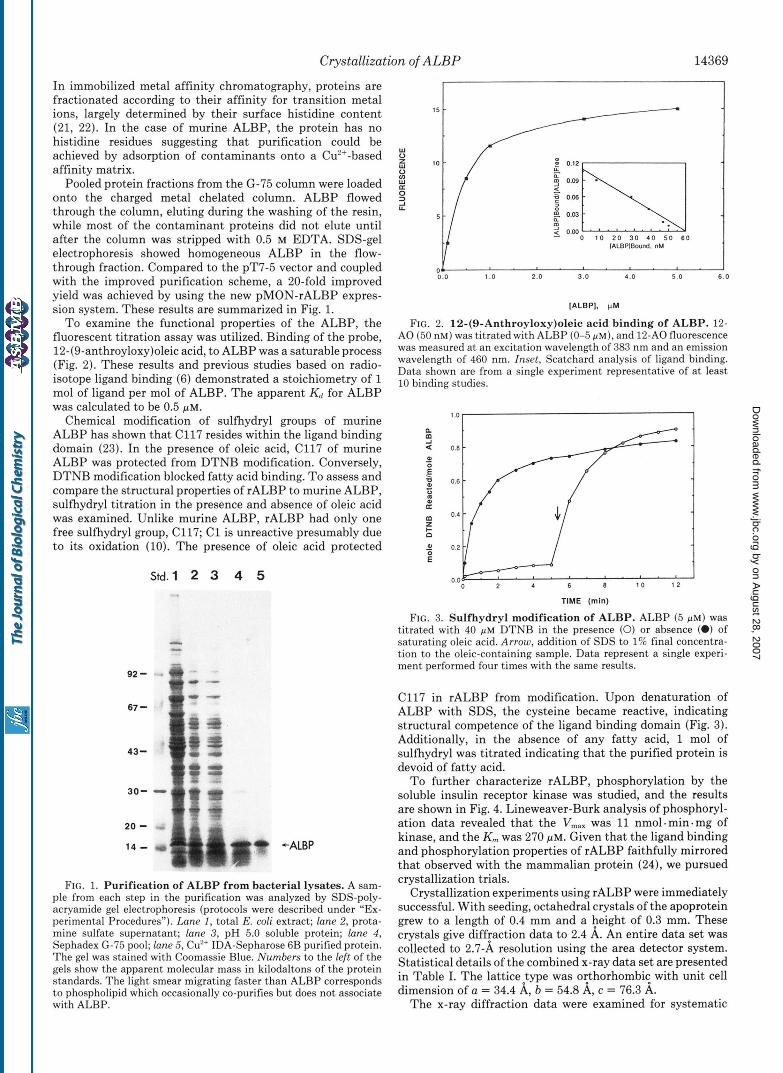
To examine the functional properties of the ALBP, the fluorescent titration assay was utilized. Binding of the probe, 12-(9-anthroyloxy)oleic acid, to ALBP was a saturable process (Fig. 2). These results and previous studies based on radio- isotope ligand binding (6) demonstrated a stoichiometry of 1 mol of ligand per mol of ALBP. The apparent Kd for ALBP was calculated to be 0.5 p ~ .
Chemical modification of sulfhydryl groups of murine ALBP has shown that C117 resides within the ligand binding domain (23). In the presence of oleic acid, c117 of murine ALBP was protected from DTNB modification. Conversely, DTNB modification blocked fatty acid binding. To assess and compare the structural properties of rALBP to murine ALBP, sulfhydryl titration in the presence and absence of oleic acid was examined. Unlike murine ALBP, rALBP had only one free sulfhydryl group, C117; C1 is unreactive presumably due to its oxidation (10). The presence of oleic acid protected
Std. 1 2 3 4 5
92 - 67 -
FIG. 1. Purification of ALBP from bacterial lysates. A sam- ple from each step in the purification was analyzed by SDS-poly- acryamide gel electrophoresis (protocols were described under "Ex- perimental Procedures"). Lane I, total E. coli extract; lane 2, prota- mine sulfate supernatant; lune 3, pH 5.0 soluble protein; lune 4, Sephadex G-75 pool; lane 5, Cu2+ IDA-Sepharose 6B purified protein. The gel was stained with Coomassie Blue. Numbers to the left of the gels show the apparent molecular mass in kilodaltons of the protein standards. The light smear migrating faster than ALBP corresponds
with ALBP. to phospholipid which occasionally co-purifies but does not associate
'V a ".r\l 0' h 0.03
2 0.w rn
0 1 0 2 0 3 0 4 0 5 0 6 0 IALBPIBound. nM
O J . ' . ' . ' ' ' . ' " 0 . 0 1 .o 2.0 3.0 4.0 5 .0 6.0
IALBPI. PM
FIG. 2. 12-(9-Anthroyloxy)oleic acid binding of ALBP. 12- A 0 (50 nM) was titrated with ALBP (0-5 PM), and 12-A0 fluorescence was measured at an excitation wavelength of 383 nm and an emission wavelength of 460 nm. Inset, Scatchard analysis of ligand binding. Data shown are from a single experiment representative of a t least 10 binding studies.
=E 0.0 1 0 1 2
TIME (min)
FIG. 3. Sulfhydryl modification of ALBP. ALBP (5 PM) was titrated with 40 PM DTNB in the presence (0) or absence (0) of saturating oleic acid. Arrow, addition of SDS to 1% final concentra- tion to the oleic-containing sample. Data represent a single experi- ment performed four times with the same results.
C117 in rALBP from modification. Upon denaturation of ALBP with SDS, the cysteine became reactive, indicating structural competence of the ligand binding domain (Fig. 3). Additionally, in the absence of any fatty acid, 1 mol of sulfhydryl was titrated indicating that the purified protein is devoid of fatty acid.
To further characterize rALBP, phosphorylation by the soluble insulin receptor kinase was studied, and the results are shown in Fig. 4. Lineweaver-Burk analysis of phosphoryl- ation data revealed that the VmaX was 11 nmol -min. mg of kinase, and the K,,, was 270 pM. Given that the ligand binding and phosphorylation properties of rALBP faithfully mirrored that observed with the mammalian protein (24), we pursued crystallization trials.
Crystallization experiments using rALBP were immediately successful. With seeding, octahedral crystals of the apoprotein grew to a length of 0.4 mm and a 3;leight of 0.3 mm. These crystals give diffraction data to 2.4 A. An entire data set was collected to 2.7-A resolution using the area detector system. Statistical details of the combined x-ray data set are presented in Table I. The 1attice:ype was oehorhombit with unit cell dimension of a = 34.4 A, b = 54.8 A, c = 76.3 A.
The x-ray diffraction data were examined for systematic
by on August 28, 2007
ww
w.jbc.org
Dow
nloaded from

14370 Crystallization of ALBP
I I
[ALBPI, PM
FIG. 4. Phosphorylation of ALBP with the soluble insulin receptor kinase. ALBP was phosphorylated in 25 mM HEPES, pH 7.5, containing 5 mM MnClz and 100 pM [32P]ATP by 0.25 pg of autophosphorylated soluble receptor for 10 and 22 min. The reactions were terminated, and the proteins were subjected to SDS-PAGE and autoradiography. The rate of 32P incorporation into ALBP was meas- ured by excision of the radioactive protein and liquid scintillation counting. Results are presented from one analysis representative of three experiments.
TABLE I X-ray data collection statistics for crystalline ALBP
Merging R factor based on I 0.0426 Resolution limits 2.2 A Number of observations 20,478 Number of unique x-ray reflections collected 5,473 Average number of observations for each reflec- 4.0
% of possible reflections collected to 2.7 4 98 % of possible reflections collected to 2.4 A 36
tion
absences to determine the space group. Such absences were observed along the a*, b*, and c* axes. Only reflections with h, k, or 1 = 2n were observed along the reciprocal axes. This indicated that the space group is P212121 (25). A unit cell with the dimensions described above has a volume of 1.44 x lo5 A3. Assuming that half of the crystal volume is water, the volume of protein is approximately 7.2 x lo4 A3. Considering the space group here, the volume of protein in 1 asymmetric unit would be 1.8 X lo4 &. By averaging the specific volume of constituent amino acids, the specific volume of ALBP is 0.715 ml/g. This led to the conclusion that the molecular mass in one asymmetric unit is 15,155 daltons. Since the molecular mass of ALBP is approximately 15 kDa, there is only 1 molecule of ALBP in an asymmetric unit.
As indicated, ALBP belongs to a family of low molecular weight fatty acid binding proteins (26). The sequences of the proteins in the family have been shown to be very similar and in particular in the amino-terminal domain where Y19 resides. Among them, the structure of myelin P2 and IFABP has been solved. Since the amino acid identity between ALBP and myelin P2 is about 69%, P2 should be a good starting structure to obtain phase information for ALBP using the methods of molecular replacement. Preliminary solutions to the rotation and translation functions have been obtained. Seeding tech- niques will allow us to obtain large crystals for further study of the holo- and phosphorylated protein. By comparing the crystal structures of these different forms, it should be possi-
ble to structurally determine the effects of protein phosphoryl- ation on ligand binding and ligand binding on phosphoryla- tion. These results will provide clues as to the effect of insulin on regulation of lipid trafficking in adipose cells.
Acknowledgments-We would like to thank Anne J. Smith for expert technical assistance for the polymerase chain reaction exper- iments. We are indebted to Drs. Jeffrey Gordon and Peter Olins for providing the pMON-5840 plasmid and Dr. James Fuchs for the E. coli JMlOl strain. We are also very grateful to Dr. Alwyn Jones and his colleagues for the use of the myelin P2 crystal coordinates and Dr. Leland Ellis for providing cloned human insulin receptor soluble kinase domain. In addition, we thank Dr. Eugene Sulkowski for suggestions concerningpurification of ALBP using immobilized metal affinity chromatography.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14. 15.
16.
17. 18.
19.
REFERENCES Ockner, R. K., Manning, J. A., and Kane, J. P. (1982) J. Biol.
Chem. 257, 7872-7878 Ockner, R. K., and Manning, J. A. (1974) J. Clin. Znuest. 54,
326-338 Sacchettini, J . C., Said, B., Schulz, H., and Gordon, J . I. (1986)
J. Biol. Chem. 261, 8281-8223 Bohmer, F.-D., Kraft, R., Otto, A., Wernstedt, C., Hellman, U.,
Kurtz, A.. Muller, T., Rohde, K., Etzold, G., Lehmann, W., Langen, P., Heldin, C.-H., and Grosse, R. (1987) J. Biol. Chem. 262, 15137-15143
Suzuki, M., Kitamura, K., Sakamoto, Y., and Uyemura, K. (1982) J. Neurochem. 39, 1759-1762
Matarese, V., and Bernlohr, D. A. (1988) J. Biol. Chem. 263, 14544-14551
Bernlohr, D. A,, Angus, C. W., Lane, M. D., Bolanowski, M. A., and Kelly, T. J., Jr. (1984) Proc. Natl. Acad. Sci. U. S. A. 81, 5468-5472
Bernier, M., Laird, D. M., and Lane, M. D. (1987) Proc. Natl. Acad. Sci. U. S. A. 84, 1844-1848
Hresko, R. C., Bernier, M., Hoffman, R. D., Flores-Riveros, J . R., Liao, K., Laird, D. M., and Lane, M. D. (1988) Proc. Natl. Acad. Sci. U. S. A. 85,8835-8839
Chinander, L. L., and Bernlohr, D. A. (1989) J. Biol. Chem. 264, 19564-19572
Sacchettini, J. C., Gordon, J . I., and Banaszak, L. J . (1989) J. Mol. B i d . 208, 327-339
Jones, T. A., Bergfors, T., Sedzik, J., and Unge, T. (1988) E M B O J. 7,1597-1604
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular C1oning:A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
Ellman, G. L. (1959) Arch. Biochem. Biophys. 82, 70-77 Cobb, M. H., Sang, B.-C., Gonzalez, R., Goldsmith, E., and Ellis,
Herrera, R., Lebwohl, D., de Herreros, A. G., Kallen, R. G., and
Laemmli, U. K. (1970) Nature 227,680-685 McPherson, A. (1982) Preparation and Analysis of Protein Crys-
Howard, A. J., Nielsen, C., and Xuong, N. H. (1985) Methods
L. (1989) J. Biol. Chem. 264, 18701-18706
Rosen, 0. M. (1988) J. Biol. Chem. 263, 5560-5568
tals, John Wiley & Sons, New York
Enzvmol. 114. 452-471 20. Olins,-P. O., and-Rangwala, S. H. (1990) Methods Enzymol. 185,
21. Sulkowski, E. (1985) Trends Biotechnol. 3, 1-7 22. Ostrove, S., and Weiss, S. (1990) Methods Enzymol. 182, 371-
23. Buelt, M. K., and Bernlohr, D. A. (1990) Biochemistry 29, 7408-
24. Buelt, M. K., Shekels, L. L., Jarvis, B. W., and Bernlohr, D. A.
25. Hahn, T. (ed) (1983) International Tables for Crystallography, pp.
26. Matarese, V., Stone, R. L., Waggoner, D. W., and Bernlohr, D.
115-119
379
7413
(1991) J. Biol. Chem. 266, 12266-12271
40-48, D. Reidel Publishing Co., Boston
A. (1990) Prog. Lipid Res. 28, 245-272
by on August 28, 2007
ww
w.jbc.org
Dow
nloaded from





























