Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology. Inc.
Vol. 263, No. 14, Issue of May 15, pp. 667&X&?, 1988 Printed m U.S.A.
Biosynthesis of Heme in Immature Erythroid Cells THE REGULATORY STEP FOR HEME FORMATION IN THE HUMAN ERYTHRON*
(Received for publication, May 12, 1987)
Lynne C. Gardner and Timothy M. Cox$ From the Royal Postgraduate Medical School, Hammersmith Hospital, London W12 OHS, United Kingdom
Heme formation in reticulocytes from rabbits and rodents is subject to end product negative feedback regulation: intracellular “free” heme has been shown to control acquisition of transferrin iron for heme syn- thesis. To identify the site of control of heme biosyn- thesis in the human erythron, immature erythroid cells were obtained from peripheral blood and aspirated bone marrow. After incubation with human “Fe trans- ferrin, 2-[‘4C]glycine, or 4-[‘4C]G-aminolevulinate, iso- topic incorporation into extracted heme was deter- mined. Addition of cycloheximide to increase endoge- nous free heme, reduced incorporation of labeled gly- tine and iron but not d-aminolevulinate into cell heme. Incorporation of glycine and iron was also sensitive to inhibition by exogenous hematin (Ki, 30 and 45 pM, respectively) i.e. at concentrations in the range which affect cell-free protein synthesis in reticulocyte ly- sates. Hematin treatment rapidly diminished incorpo- ration of intracellular “Fe into heme by human eryth- roid cells but assimilation of 4-[‘4C]6-a~inolevulinate into heme was insensitive to inhibition by hematin (Ki > 100 PM). In human reticulocytes (unlike those from rabbits), addition of ferric salicylaldehyde isonicoti- noylhydrazone, to increase the pre-heme iron pool in- dependently of the transferrin cycle, failed to promote heme synthesis or modify feedback inhibition induced by hematin. In human erythroid cells (but not rabbit reticulocytes) pre-incubation with unlabeled a-amino- levulinate or protoporphyrin IX greatly stimulated uti- lization of cell “Fe for heme synthesis and also atten- uated end product inhibition.
In human erythroid cells heme biosynthesis is thus primarily regulated by feedback inhibition at one or more steps which lead to d-aminolevulinate formation. Hence in man the regulatory process affects generation of the first committed precursor of porphyrin biosyn- thesis by b-aminolevulinate synthetase, whereas in the rabbit separate regulatory mechanisms exist which control the incorporation of iron into protoporphyrin TV
The erythron is the major organ of heme formation in vertebrates. Heme is a component of the blood pigment but also has a regulatory function in erythroid cells, where it coordinates the pathways of hemoglobin biosynthesis (1). In
* This work was supported by a grant from The Wellcome Trust. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 USC. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed: Dept. of Haema- tology and Medicine, Royal Postgraduate Medical School, Du Cane Road, London, W12 OHS, United Kingdom.
these cells, adequate concentrations of heme are required to maintain active initiation of globin chain synthesis on ribo- somes (2, 3). Excess heme inhibits protein synthesis and the balanced formation of heme de nouo may itself be controlled by end product negative feedback (1).
Although heme biosynthesis in immature erythroid cells may be subject to negative feedback regulation, the mecha- nism of control and point of regulation is unclear. In cultures of developing avian blastoderm, addition of b-aminolevulinate promotes the rapid formation of heme and hemoglobin (4). It was therefore proposed that 6-aminolevulinate synthesis was rate-limiting for hemoglobin formation. However, identifica- tion of the cellular mechanism which controls heme synthesis in long-term cultures of nucleated erythroid progenitors is complicated by concomitant effects of tissue differentiation. Studies using intact and lysed preparations of reticulocytes from rabbits suggested the existence of feedback control at one or more steps leading to the formation of &aminolevuli- nate (1, 5). In rabbit reticulocyte stroma (5, 6) and in prepa- rations of &aminolevulinate synthetase purified from rabbit reticulocytes (7), exogenous hemin unexpectedly had only weak inhibitory effects on formation of aminolevulinate. Many experiments on negative feedback control in reticulo- cytes were carried out under conditions where availability of substrates may have been limiting for hemoglobin synthesis (especially amino acids and transferrin iron (5)). Under these circumstances, the disturbance of cellular metabolism would be expected to desensitize the mechanism which normally regulates heme formation.
Observations made with reticulocytes from experimental animals have shown that changes in intracellular free heme induced by treatment with inhibitors of heme and globin synthesis, as well as hematin, also affect delivery of transfer- rin iron destined for heme formation (8, 9). Raised intracel- lular heme appears, at least in the rabbit, to inhibit internal- ization of the iron transferrin-receptor complex from the reticulocyte surface membrane (10, 11). Further experiments showing that protoporphyrin synthesis is unaffected by hemin treatment and that addition of b-aminolevulinate or free protoporphyrin IX does not stimulate heme synthesis, sug- gested that uptake of iron from transferrin, intracellular iron delivery, or feedback inhibition of ferrochelatase regulates heme formation (12). Further support for these findings was provided by elegant experiments with iron chelates of acylhy- drazones, which donate iron for heme synthesis independently of transferrin (13). These compounds stimulate synthesis of heme from labeled glycine precursor in rabbit reticulocytes and were found to overcome feedback inhibition induced by exogenous hemin (12, 14).
Although it has long been known that synthesis of heme may be detected readily in peripheral blood samples obtained from individuals with sickle cell anemia (15, 16), there are few reported investigations into the control of heme formation
6676
Regulation of Heme Synthesis in Human Erythroid Cells 6677
in the human erythron. Operation of a feedback mechanism affecting delivery of iron from transferrin to erythroid tissues has importance for understanding the genesis of sideroblastic anemia and diseases of iron storage. Likewise, the possibility that infusions of hematin might be used to suppress porphyrin formation in congenital erythropoietic porphyria has been raised by a short-term study (17) but further studies are needed to clarify the site of action of hematin on heme synthesis in human erythroid cells before its general use to inhibit the pathway in man can be accepted.
Accordingly, we have investigated whether feedback control of heme biosynthesis in human reticulocytes and bone marrow tissue occurs, and, if so, how it is regulated. We have also carried out parallel studies using immature erythroid cells obtained from the rabbit.
MATERIALS AND METHODS
L-Amino acids, 6-aminolewlinate, hemin hydrochloride, cyclohex- imide, 4,6-&oxoheptanoic acid, and rabbit transferrin were obtained from Sigma. Free protoporphyrin IX was supplied by Behring Diag- nostics. Ethyl acetate, isonicotinic acid hydrazide, and salicylaldehyde were purchased from Aldrich. Radiochemicals were supplied by Amer- sham Int. Solutions of heme arginate (Medica Pharmaceutical Co., Finland) used in studies of feedback control were diluted in distilled water and were a generous gift from Dr. R. Oakes, Tillotts Labora- tories, United Kingdom. Transferrin was prepared from human serum as described previously (18) and used in the iron-saturated form, unless otherwise stated, a t 1 mg of transferrin/ml(25 pM with respect to iron). Other reagents were of analytical grade and from established suppliers.
Cell Preparations-Immature erythroid cells were freshly obtained from human subjects, seven male and seven female, aged 10-64, with hematologic disorders. Nine subjects had sickle cell anemia; one, recently treated pernicious anemia; one, paroxysmal nocturnal he- moglobinuria with iron deficiency, and one, suffered autoimmune hemolysis. The reticulocyte count, as determined by supravital stain- ing with 1% (w/v) new methylene blue, was 10-25% in all subjects studied. Bone marrow aspirates were obtained before, and 1 month after, iron therapy in a man with mild sideropenic anemia due to chronic hemorrhage. Marrow was also obtained from a man with erythroid hyperplasia due to autoimmune hemolysis.
Samples of blood and aspirated marrow were collected into retic- ulocyte saline buffer (RS): 0.13 M NaC1, 0.0074 M MgCl,, 0.005 M KC1 buffered to pH 7.35 with 0.01 M Na-Hepes’ and containing 50 units/ml preservative-free heparin. The cells were separated from plasma and washed three times in RS buffer by centrifugation. The buffy coat was carefully aspirated after each cycle.
Rabbit reticulocytes for cell-free studies of translation in lysates were obtained after phenylhydrazine treatment to induce hemolysis (18). Reticulocyte-rich blood was also obtained from rabbits which had been bled to maintain the hematocrit a t about 30% and induce a 15-30% peripheral blood reticulocytosis (19).
Studies of Globin and Heme Biosynthesis-Cells were incubated within 1 h of removal as 15-20% suspensions in RS buffer a t 37 “C in the presence of 1 mg/ml iron-saturated transferrin and 0.1% (w/ v) D-glUCOSe. The mixture contained, except where stated, 20 L-amino acids a t 100 FM each and the incubations were agitated to maintain oxygenation and avoid clumping. In experiments to follow protein synthesis, ~-[~~S]methionine (>IO00 Ci/mmol) was added at 100-200 pCi/ml and the final concentration of methionine was reduced to 20- 50 p ~ . Overall protein synthesis was determined by counting trichlo- roacetic acid-insoluble radioactivity incorporated into total cell pro- tein (19), and duplicate determinations were generally within 5% of the mean value.
The ability of hemin preparations to stimulate translation of endogenous globin mRNA was checked in rabbit reticulocyte lysates using ~-[“C]leucine as the isotopic label, as reported previously (18). Hemin hydrochloride stock solution was prepared and stored in ethylene glycol exactly as described in Ref. 18 and diluted in H,O immediately before use; the 40 mM heme arginate stock solution was diluted similarly. Gel filtration experiments using Sephadex G-25
The abbreviation used is: Hepes, 4-(2-hydroxyethyl)-l-pipera- zineethanesulfonic acid.
chromatography in RS buffer showed that the two preparations consisted of unpolymerized hematin, which was completely retained by the columns.
Incorporation of iron into cell heme was followed in incubations in which autologous transferrin was labeled with ‘Te (11). Synthesis of the porphyrin moiety was determined in extracts of cells incubated with either 20 pCi/ml 2-[’4C]glycine (58 mCi/mmol) or 20 pcilml 4- [“C]b-aminolevulinate (53 mCi/mmol). The final concentrations of glycine and 6-aminolevulinate were 445 and 377 pht, respectively. Incubations for heme biosynthesis were stopped by diluting aliquots containing 10 /*I (10’) of packed cells (3 mg of hemoglobin) to 1 ml with ice-cold RS buffer. The diluted mixtures were then underlayed with 100 pl of dibutyl phthalate in 1.5-ml conical tubes and centri- fuged at 12,000 X g for 60 s. The supernatant fluids were aspirated and the cells were resuspended in 1 ml of ice-cold RS buffer and pelleted again. The final cell pellets were mixed thoroughly in 0.5 ml of ice-cold distilled water and lysed by freeze-thawing. Incorporation of labeled precursors into cellular heme was determined by measuring the radioactivity of heme extracted into ethyl acetate. The aqueous lysate was treated repeatedly with ethyl acetate/glacial acetic acid (3:1, v/v) and the organic heme layer was washed with saturated aqueous sodium acetate solution followed by repeated extraction with 3 M HCI to elute labeled porphyrins (20). To monitor recovery of heme, 59Fe-labeled ferriprotohemin was synthesized chemically (21) and used to trace label in non-radioactive cell lysates, which were extracted in parallel. In a given experiment, determinations of cell uptake and incorporation of 59Fe from transferrin into heme were highly reproducible: coefficients of variation were 8.4 and 8.2%, respectively. The coefficient of variation for assays of 2-[“C]glycine incorporation into reticulocyte heme was 9.4%. Recovery of heme was generally greater than 75% and was consistent within 2%. In experi- ments to stimulate heme synthesis, cells were preincubated at 4 “C with designated unlabeled precursors for 30 min before warming to 37 “C in the presence of labeled substrate. Ferric salicylaldehyde isonicotinoylhydrazone was synthesized by mixing acidified ferric citrate and salicylaldehyde isonicotinoylhydrazone. It was used as reported by others (13, 14) but without preincubation.
RESULTS AND DISCUSSION
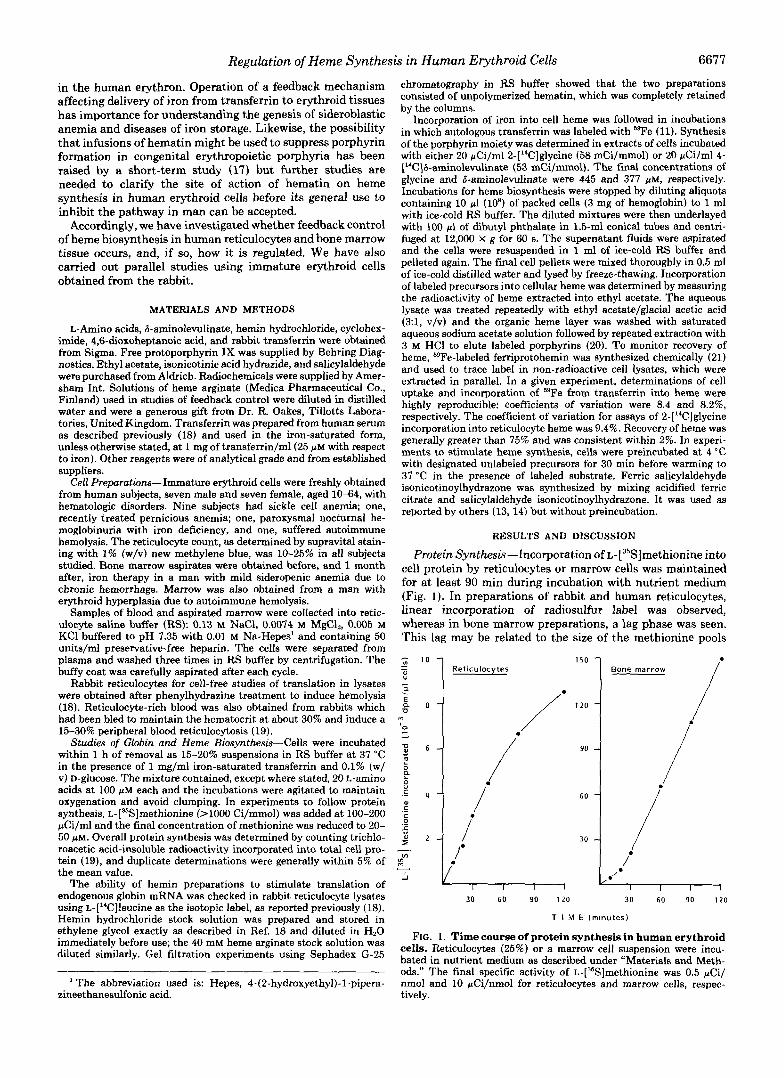
Protein Synthesis-Incorporation of ~-[~~S]methionine into cell protein by reticulocytes or marrow cells was maintained for at least 90 min during incubation with nutrient medium (Fig. 1). In preparations of rabbit and human reticulocytes, linear incorporation of radiosulfur label was observed, whereas in bone marrow preparations, a lag phase was seen. This lag may be related to the size of the methionine pools
l o 1 Reticulocytes 150 1 Bone marrow U i
* I
30 I / 30 60 90 1 2 0 30 60 90 120
T I M E [minutes)
FIG. 1. Time course of protein synthesis in human erythroid cells. Reticulocytes (25%) or a marrow cell suspension were incu- bated in nutrient medium as described under “Materials and Meth- ods.” The final specific activity of ~-[~~S]methionine was 0.5 FCi/ nmol and 10 pCi/nmol for reticulocytes and marrow cells, respec- tively.
6678 Regulation of Heme Synthesis in Human Erythroid Cells
within different cell types. Unlike suspensions of reticulo- cytes, marrow aspirates contain a large proportion of non- erythroid cells, which might thus explain these differences.
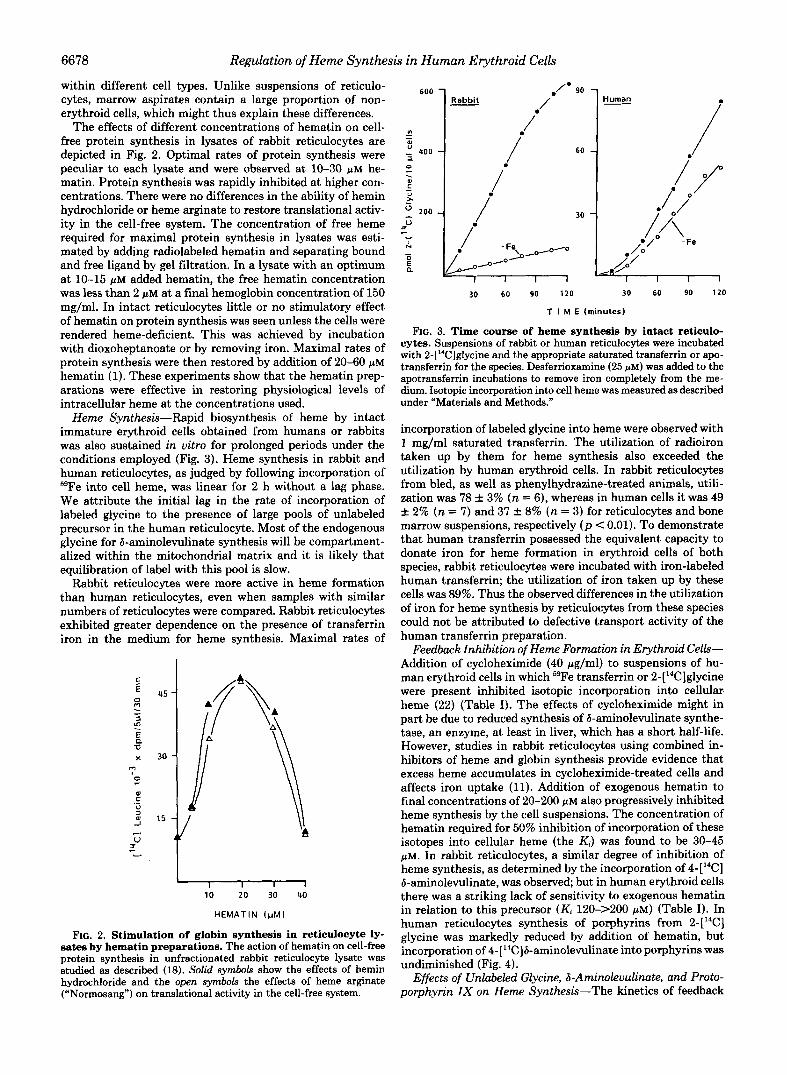
The effects of different concentrations of hematin on cell- free protein synthesis in lysates of rabbit reticulocytes are depicted in Fig. 2. Optimal rates of protein synthesis were peculiar to each lysate and were observed at lo-30 pM he- matin. Protein synthesis was rapidly inhibited at higher con- centrations. There were no differences in the ability of hemin hydrochloride or heme arginate to restore translational activ- ity in the cell-free system. The concentration of free heme required for maximal protein synthesis in lysates was esti- mated by adding radiolabeled hematin and separating bound and free ligand by gel filtration. In a lysate with an optimum at lo-15 pM added hematin, the free hematin concentration was less than 2 pM at a final hemoglobin concentration of 150 mg/ml. In intact reticulocytes little or no stimulatory effect of hematin on protein synthesis was seen unless the cells were rendered heme-deficient. This was achieved by incubation with dioxoheptanoate or by removing iron. Maximal rates of protein synthesis were then restored by addition of 20-60 FM hematin (1). These experiments show that the hematin prep- arations were effective in restoring physiological levels of intracellular heme at the concentrations used.
Heme Synthesis-Rapid biosynthesis of heme by intact immature erythroid cells obtained from humans or rabbits was also sustained in vitro for prolonged periods under the conditions employed (Fig. 3). Heme synthesis in rabbit and human reticulocytes, as judged by following incorporation of 59Fe into cell heme, was linear for 2 h without a lag phase. We attribute the initial lag in the rate of incorporation of labeled glycine to the presence of large pools of unlabeled precursor in the human reticulocyte. Most of the endogenous glycine for 8-aminolevulinate synthesis will be compartment- alized within the mitochondrial matrix and it is likely that equilibration of label with this pool is slow.
Rabbit reticulocytes were more active in heme formation than human reticulocytes, even when samples with similar numbers of reticulocytes were compared. Rabbit reticulocytes exhibited greater dependence on the presence of transferrin iron in the medium for heme synthesis. Maximal rates of
45
30
15
HEMATIN (PM)
FIG. 2. Stimulation of globin synthesis in reticulocyte ly- sates by hematin preparations. The action of hematin on cell-free protein synthesis in unfractionated rabbit reticulocyte lysate was studied as described (18). Solid symbols show the effects of hemin hydrochloride and the open symbols the effects of heme arginate (“Normosang”) on translational activity in the cell-free system.
1. Rabbit /* 90
.’
30 60 90 120
30
1 I M E (minutes1
FIG. 3. Time course of heme synthesis by intact reticulo- cytes. Suspensions of rabbit or human reticulocytes were incubated with 2-[“Clglycine and the appropriate saturated transferrin or apo- transferrin for the species. Desferrioxamine (25 PM) was added to the apotransferrin incubations to remove iron completely from the me- dium. Isotopic incorporation into cell heme was measured as described under “Materials and Methods.”
incorporation of labeled glycine into heme were observed with 1 mg/ml saturated transferrin. The utilization of radioiron taken up by them for heme synthesis also exceeded the utilization by human erythroid cells. In rabbit reticulocytes from bled, as well as phenylhydrazine-treated animals, utili- zation was 78 f 3% (n = 6), whereas in human cells it was 49 + 2% (n = 7) and 37 f 8% (n = 3) for reticulocytes and bone marrow suspensions, respectively (p < 0.01). To demonstrate that human transferrin possessed the equivalent capacity to donate iron for heme formation in erythroid cells of both species, rabbit reticulocytes were incubated with iron-labeled human transferrin; the utilization of iron taken up by these cells was 89%. Thus the observed differences in the utilization of iron for heme synthesis by reticulocytes from these species could not be attributed to defective transport activity of the human transferrin preparation.
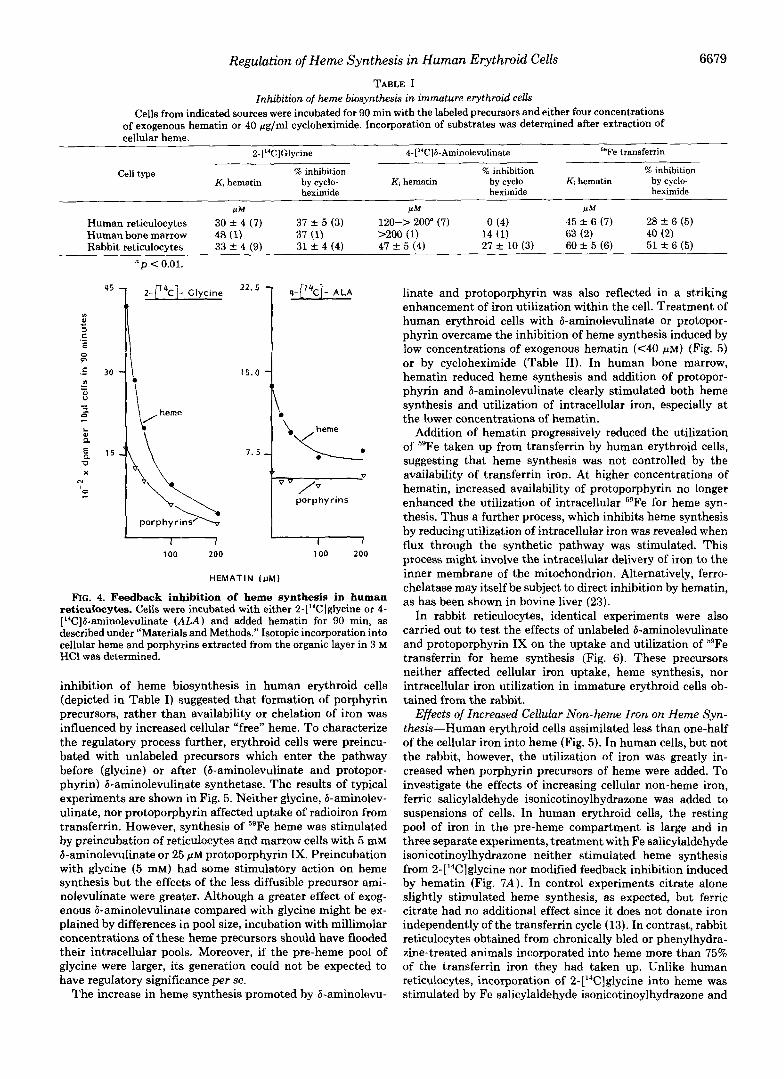
Feedback Inhibition of Heme Formation in Erythroid Cells- Addition of cycloheximide (40 pg/ml) to suspensions of hu- man erythroid cells in which 59Fe transferrin or 2-[14C]glycine were present inhibited isotopic incorporation into cellular heme (22) (Table I). The effects of cycloheximide might in part be due to reduced synthesis of &aminolevulinate synthe- tase, an enzyme, at least in liver, which has a short half-life. However, studies in rabbit reticulocytes using combined in- hibitors of heme and globin synthesis provide evidence that excess heme accumulates in cycloheximide-treated cells and affects iron uptake (11). Addition of exogenous hematin to final concentrations of 20-200 pM also progressively inhibited heme synthesis by the cell suspensions. The concentration of hematin required for 50% inhibition of incorporation of these isotopes into cellular heme (the Ki) was found to be 30-45 PM. In rabbit reticulocytes, a similar degree of inhibition of heme synthesis, as determined by the incorporation of 4-[14C] &aminolevulinate, was observed; but in human erythroid cells there was a striking lack of sensitivity to exogenous hematin in relation to this precursor (Ki 120~>200 FM) (Table I). In human reticulocytes synthesis of porphyrins from 2-[‘“Cl glycine was markedly reduced by addition of hematin, but incorporation of 4-[‘4C]G-aminole~linate into porphyrins was undiminished (Fig. 4).
Effects of Unlabeled Glycine, &Aminolevulinate, and PFOtO-
porphyrin IX on Heme Synthesis-The kinetics of feedback
Regulation of Heme Synthesis in Human Erythroid Cells 6679
TABLE I Inhibition of heme biosynthesis in immature erythroid cells
Cells from indicated sources were incubated for 90 min with the labeled precursors and either four concentrations of exogenous hematin or 40 pg/ml cycloheximide. Incorporation of substrates was determined after extraction of cellular heme.
%[“C]Glycine 4-[’4C]6-Aminolevulinate ’Te transferrin
Cell type % inhibition % inhibition % inhibition K j hematin by CYC~O- K, hematin by CyClO- K; hematin by cyclo-
heximide heximide heximide
P M I.” P M
Human reticulocytes 30 f 4 (7) 37 f 5 (3) 120-> 200” (7) 0 (4) 45 f 6 (7) 28 f 6 (5) Human bone marrow 48 (1) 37 (1) >200 (1) 14 (1) 63 (2) 40 (2) Rabbit reticulocytes 33 f 4 (9) 31 f 4 (4) 47 -t 5 (4) 27 f 10 (3) 60 f 5 (6) 51 f 6 (5)
45
30
1 5
” p c 0.01.
2 - [ “ + ~ ] - Glycine 22.5
porphyrtns’
4-[’4C]- ALA
V
7 v /v porphyrins
1 I 100 200
I I
100 200
HEMATIN (UM)
FIG. 4. Feedback inhibition of heme synthesis in human reticulocytes. Cells were incubated with either 2-[14C]glycine or 4- [“CIB-aminolevulinate (ALA) and added hematin for 90 min, as described under ”Materials and Methods.” Isotopic incorporation into cellular heme and porphyrins extracted from the organic layer in 3 M HC1 was determined.
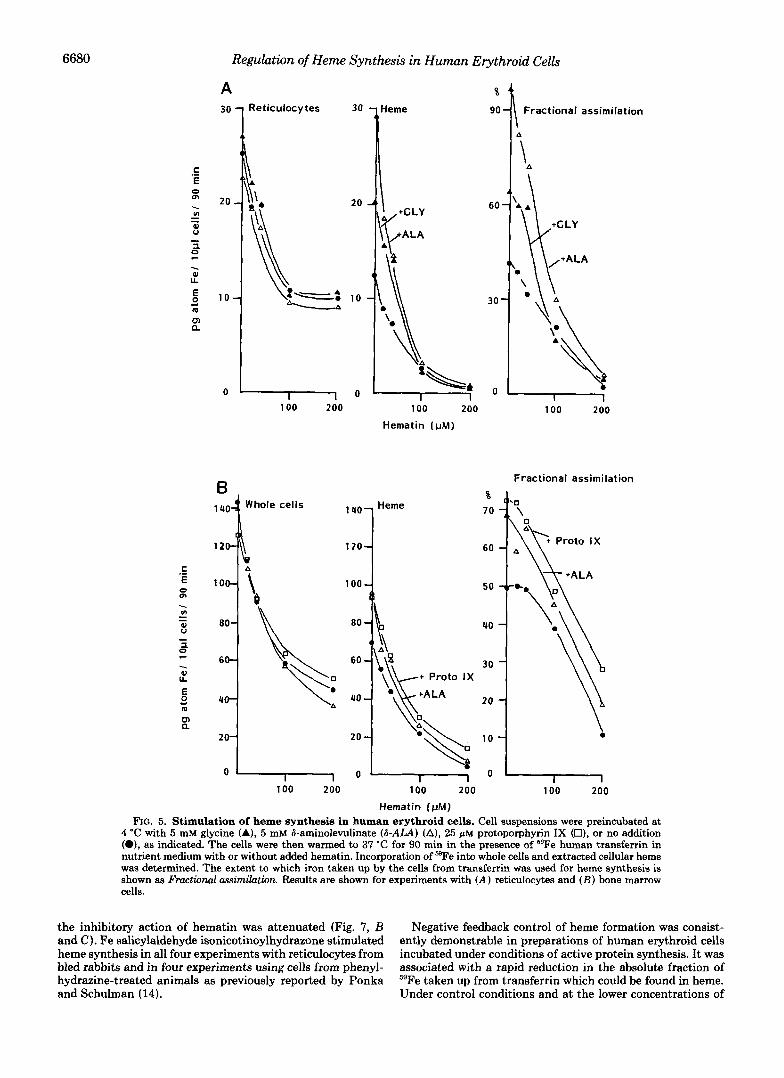
inhibition of heme biosynthesis in human erythroid cells (depicted in Table I) suggested that formation of porphyrin precursors, rather than availability or chelation of iron was influenced by increased cellular “free” heme. To characterize the regulatory process further, erythroid cells were preincu- bated with unlabeled precursors which enter the pathway before (glycine) or after (6-aminolevulinate and protopor- phyrin) 6-aminolevulinate synthetase. The results of typical experiments are shown in Fig. 5. Neither glycine, 6-aminolev- ulinate, nor protoporphyrin affected uptake of radioiron from transferrin. However, synthesis of 59Fe heme was stimulated by preincubation of reticulocytes and marrow cells with 5 mM 6-aminolevulinate or 25 pM protoporphyrin Ix. Preincubation with glycine (5 mM) had some stimulatory action on heme synthesis but the effects of the less diffusible precursor ami- nolevulinate were greater. Although a greater effect of exog- enous 6-aminolevulinate compared with glycine might be ex- plained by differences in pool size, incubation with millimolar concentrations of these heme precursors should have flooded their intracellular pools. Moreover, if the pre-heme pool of glycine were larger, its generation could not be expected to have regulatory significance per se.
The increase in heme synthesis promoted by b-aminolevu-
linate and protoporphyrin was also reflected in a striking enhancement of iron utilization within the cell. Treatment of human erythroid cells with 6-aminolevulinate or protopor- phyrin overcame the inhibition of heme synthesis induced by low concentrations of exogenous hematin (<40 pM) (Fig. 5) or by cycloheximide (Table 11). In human bone marrow, hematin reduced heme synthesis and addition of protopor- phyrin and 6-aminolevulinate clearly stimulated both heme synthesis and utilization of intracellular iron, especially at the lower concentrations of hematin.
Addition of hematin progressively reduced the utilization of 59Fe taken up from transferrin by human erythroid cells, suggesting that heme synthesis was not controlled by the availability of transferrin iron. At higher concentrations of hematin, increased availability of protoporphyrin no longer enhanced the utilization of intracellular 59Fe for heme syn- thesis. Thus a further process, which inhibits heme synthesis by reducing utilization of intracellular iron was revealed when flux through the synthetic pathway was stimulated. This process might involve the intracellular delivery of iron to the inner membrane of the mitochondrion. Alternatively, ferro- chelatase may itself be subject to direct inhibition by hematin, as has been shown in bovine liver (23).
In rabbit reticulocytes, identical experiments were also carried out to test the effects of unlabeled 6-aminolevulinate and protoporphyrin IX on the uptake and utilization of 59Fe transferrin for heme synthesis (Fig. 6). These precursors neither affected cellular iron uptake, heme synthesis, nor intracellular iron utilization in immature erythroid cells ob- tained from the rabbit.
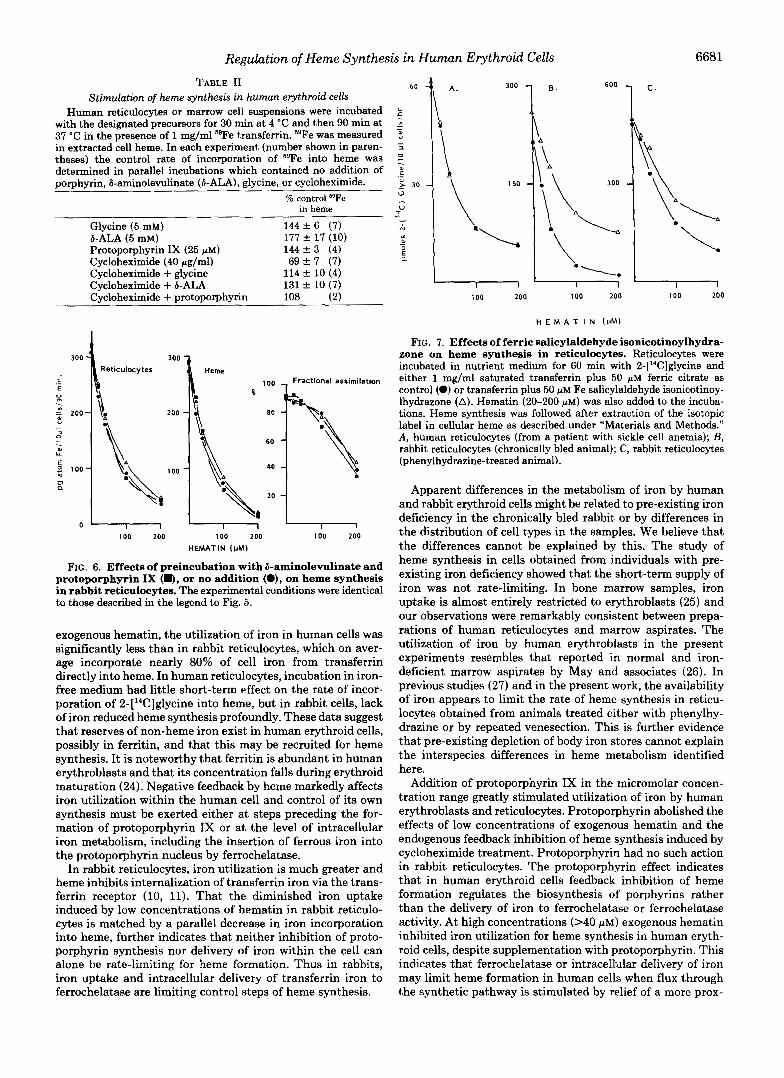
Effects of Increased Cellular Non-heme Iron on Heme Syn- thesis-Human erythroid cells assimilated less than one-half of the cellular iron into heme (Fig. 5 ) . In human cells, but not the rabbit, however, the utilization of iron was greatly in- creased when porphyrin precursors of heme were added. To investigate the effects of increasing cellular non-heme iron, ferric salicylaldehyde isonicotinoylhydrazone was added to suspensions of cells. In human erythroid cells, the resting pool of iron in the pre-heme compartment is large and in three separate experiments, treatment with Fe salicylaldehyde isonicotinoylhydrazone neither stimulated heme synthesis from 2-[’4C]glycine nor modified feedback inhibition induced by hematin (Fig. 7 A ) . In control experiments citrate alone slightly stimulated heme synthesis, as expected, but ferric citrate had no additional effect since it does not donate iron independently of the transferrin cycle (13). In contrast, rabbit reticulocytes obtained from chronically bled or phenylhydra- zine-treated animals incorporated into heme more than 75% of the transferrin iron they had taken up. Unlike human reticulocytes, incorporation of 2-[’4C]glycine into heme was stimulated by Fe salicylaldehyde isonicotinoylhydrazone and
6680 Regulation of Heme Synthesis in Human Erythroid Cells
A 30 1 Reticulocytes 30
20
10
0 100 200 100 200
Hematin (DM)
100 ::;I-
”c Fractional assimilation
/+ALA
30
0
% 70
60
50
40
30
20
10
0
100 200
Fractional assimilation
100 200 100 200 100 200
Hematin (DM) FIG. 5. Stimulation of heme synthesis in human erythroid cells. Cell suspensions were preincubated at
4 ‘c with 5 mM glycine (A), 5 mM 6-aminolevulinate (&ALA) (A), 25 pM protoporphyrin IX (O), or no addition (O), as indicated. The cells were then warmed to 37 “C for 90 min in the presence of ‘9Fe human transferrin in nutrient medium with or without added hematin. Incorporation of ‘BFe into whole cells and extracted cellular heme was determined. The extent to which iron taken up by the cells from transferrin was used for heme synthesis is shown as Fractional assimilation. Results are shown for experiments with ( A ) reticulocytes and ( B ) bone marrow cells.
the inhibitory action of hematin was attenuated (Fig. 7, B Negative feedback control of heme formation was consist- and C). Fe salicylaldehyde isonicotinoylhydrazone stimulated ently demonstrable in preparations of human erythroid cells heme synthesis in all four experiments with reticulocytes from incubated under conditions of active protein synthesis. It was bled rabbits and in four experiments using cells from phenyl- associated with a rapid reduction in the absolute fraction of hydrazine-treated animals as previously reported by Ponka 59Fe taken up from transferrin which could be found in heme. and Schulman (14). Under control conditions and at the lower concentrations of
Regulation of Heme Synthesis in Human Erythroid Cells
TABLE I1 Stimulation of heme synthesis in human erythroid cells
Human reticulocytes or marrow cell suspensions were incubated with the designated precursors for 30 min at 4 "C and then 90 min at 37 "C in the presence of 1 mg/ml "Fe transferrin. 59Fe was measured in extracted cell heme. In each experiment (number shown in paren- theses) the control rate of incorporation of "Fe into heme was determined in parallel incubations which contained no addition of porphyrin, 6-aminolevulinate (&ALA), glycine, or cycloheximide.
% control V e in heme
Glycine (5 mM) 144 f 6 (7) &ALA (5 mM) 177 f 17 (10) Protoporphyrin IX (25 pM) 144 f 3 (4) Cycloheximide (40 pg/ml) 69 f 7 (7) Cycloheximide + glycine 114 f 10 (4) Cycloheximide + &ALA 131 f 10 (7) Cycloheximide + protoporphyrin 108 (2)
1
200
100
100
ao
60
40
100 200 100 200
HEMATIN (I"
100 200
FIG. 6. Effects of preincubation with 6-aminolevulinate and protoporphyrin IX 0, or no addition (O), on heme synthesis in rabbit reticulocytes. The experimental conditions were identical to those described in the legend to Fig. 5.
exogenous hematin, the utilization of iron in human cells was significantly less than in rabbit reticulocytes, which on aver- age incorporate nearly 80% of cell iron from transferrin directly into heme. In human reticulocytes, incubation in iron- free medium had little short-term effect on the rate of incor- poration of 2-[14C]glycine into heme, but in rabbit cells, lack of iron reduced heme synthesis profoundly. These data suggest that reserves of non-heme iron exist in human erythroid cells, possibly in ferritin, and that this may be recruited for heme synthesis. It is noteworthy that ferritin is abundant in human erythroblasts and that its concentration falls during erythroid maturation (24). Negative feedback by heme markedly affects iron utilization within the human cell and control of its own synthesis must be exerted either at steps preceding the for- mation of protoporphyrin IX or at the level of intracellular iron metabolism, including the insertion of ferrous iron into the protoporphyrin nucleus by ferrochelatase.
In rabbit reticulocytes, iron utilization is much greater and heme inhibits internalization of transferrin iron via the trans- ferrin receptor (10, 11). That the diminished iron uptake induced by low concentrations of hematin in rabbit reticulo- cytes is matched by a parallel decrease in iron incorporation into heme, further indicates that neither inhibition of proto- porphyrin synthesis nor delivery of iron within the cell can alone be rate-limiting for heme formation. Thus in rabbits, iron uptake and intracellular delivery of transferrin iron to ferrochelatase are limiting control steps of heme synthesis.
B. 600
fr
\A
100 200 100 200
H E M A T I N (DM)
6681
C . r I
100 I
200
FIG. 7. Effects of ferric salicylaldehyde isonicotinoylhydra- zone on heme synthesis in reticulocytes. Reticulocytes were incubated in nutrient medium for 60 min with 2-['4C]glycine and either 1 mg/ml saturated transferrin plus 50 p~ ferric citrate as control (0) or transferrin plus 50 p M Fe salicylaldehyde isonicotinoy- lhydrazone (A). Hematin (20-200 p M ) was also added to the incuba- tions. Heme synthesis was followed after extraction of the isotopic label in cellular heme as described under "Materials and Methods." A, human reticulocytes (from a patient with sickle cell anemia); B, rabbit reticulocytes (chronically bled animal); C, rabbit reticulocytes (phenylhydrazine-treated animal).
Apparent differences in the metabolism of iron by human and rabbit erythroid cells might be related to pre-existing iron deficiency in the chronically bled rabbit or by differences in the distribution of cell types in the samples. We believe that the differences cannot be explained by this. The study of heme synthesis in cells obtained from individuals with pre- existing iron deficiency showed that the short-term supply of iron was not rate-limiting. In bone marrow samples, iron uptake is almost entirely restricted to erythroblasts (25) and our observations were remarkably consistent between prepa- rations of human reticulocytes and marrow aspirates. The utilization of iron by human erythroblasts in the present experiments resembles that reported in normal and iron- deficient marrow aspirates by May and associates (26 ) . In previous studies (27) and in the present work, the availability of iron appears to limit the rate of heme synthesis in reticu- locytes obtained from animals treated either with phenylhy- drazine or by repeated venesection. This is further evidence that pre-existing depletion of body iron stores cannot explain the interspecies differences in heme metabolism identified here.
Addition of protoporphyrin IX in the micromolar concen- tration range greatly stimulated utilization of iron by human erythroblasts and reticulocytes. Protoporphyrin abolished the effects of low concentrations of exogenous hematin and the endogenous feedback inhibition of heme synthesis induced by cycloheximide treatment. Protoporphyrin had no such action in rabbit reticulocytes. The protoporphyrin effect indicates that in human erythroid cells feedback inhibition of heme formation regulates the biosynthesis of porphyrins rather than the delivery of iron to ferrochelatase or ferrochelatase activity. At high concentrations (>40 PM) exogenous hematin inhibited iron utilization for heme synthesis in human eryth- roid cells, despite supplementation with protoporphyrin. This indicates that ferrochelatase or intracellular delivery of iron may limit heme formation in human cells when flux through the synthetic pathway is stimulated by relief of a more prox-

6682 Regulation of Heme Synthesis in Human Erythroid Cells
imal regulatory step. The failure of Fe-salicylaldehyde iso- nicotinoylhydrazone to affect heme synthesis further points to ferrochelatase as a secondary control step. In human eryth- roid cells delivery of iron and activity of ferrochelatase, how- ever, does not, under normal circumstances, appear to limit heme biosynthesis.
In the rabbit reticulocyte, addition of ferric salicylaldehyde isonicotinoylhydrazone enhanced heme synthesis, as deter- mined by incorporation of 2-['*C]glycine (13, 14). The effect of Fe salicylaldehyde isonicotinoylhydrazone was to reduce the inhibitory action of exogenous hematin on heme synthesis. We concur with Ponka and Schulman (12,14) that in rabbits, acquisition of iron from transferrin regulates heme synthesis by reticulocytes but this view appears to be untenable in relation to the cells of the human erythron.
It has been found that increased free heme inhibits uptake of glycine required for heme biosynthesis in erythroid cells (12). It has also been shown that the plasma membrane of ':uman red cells contains a distinct system for the selective wansport of glycine (28). To determine whether 6-aminolev- ulinate synthesis, and hence porphyrin and heme formation, is subject to control by the availability of intracellular glycine (mediated by the Gly transport system) the effect of prein- cubating erythroid cells with excess unlabeled glycine was . tudied. Glycine slightly stimulated heme biosynthesis in ::lman erythroid cells, an effect which probably resulted from :lass astion on flux through the Shemin reaction. Stimulation
of heme biosynthesis in human erythroid cells by 6-aminolev- ulinate greatly exceeded the glycine effect. Thus formation of this first committed precursor of porphyrin synthesis, rather than the availability of glycine as a substrate, appears to represent the point of metabolic control for biosynthesis of heme.
Erythrocytes and reticulocytes from different species vary in their maturity and in their ability to undertake metabolic reactions (29). However, it is difficult to reconcile inconsist- encies in the reported literature (1, 5, 8, 12, 14, 27), particu- larly those concerning regulation of heme synthesis in rabbit reticulocytes. In several reports the effects of hemin on heme formation at concentrations below 50 pM (which are optimal for protein synthesis when added to intact reticulocytes or reticulocyte lysates (18)) were slight (1, 5 ) and their physio- logical significance is unclear. In the early investigations availability of substrates essential for balanced synthesis of heme and globin was limited and this may account for the dissident conclusions.
Heme regulates its own synthesis in erythroid cells and also stimulates the formation of globin. However, the action of heme on hemoglobin formation differs according to the ma- turity of the erythroid cell in question. In differentiating erythroblasts and other nucleated cells, heme has pleiotropic effects. In erythroblasts from rabbits, heme greatly promotes transcription of globin mRNA and may also stabilize tran- scribed globin RNA (30, 31); but in reticulocytes RNA syn- thesis cannot occur and here heme acts to sustain transla- tional activity on polyribosomes (2, 3). In the short term, feedback inhibition of heme on its own formation appears to operate by a common mechanism in human reticulocytes and erythroblasts.
This action of heme in the human erythron occurs at a step which precedes formation of the porphyrin macrocycle. In a study of the effects of parenteral hematin in a patient with
congenital porphyria, Watson et al. (17) noted a striking reduction in uroporphyrin concentrations in circulating red cells, plasma, and urine shortly after administration of he- matin. Hitherto it has not been clear as to whether the diminished synthesis of porphyrins was related to an action on nucleated red cell precursors or reticulocytes. We show that addition of exogenous hematin to suspensions of human red cell precursors in vitro directly inhibits the synthesis of porphyrins in both types of cell. Hematin might thus be considered for further evaluation in the treatment of congen- ital porphyria.
In summary, our findings indicate that heme biosynthesis in human erythroid cells is subject to end product feedback regulation: this occurs at one or more rate-limiting steps which lead to the formation of 6-aminolevulinate. Subsidiary mechanisms appear also to regulate the uptake of iron from transferrin and its incorporation into heme, presumably en- suring that neither excess iron nor unwanted cellular inter- mediates accumulate.
Acknowledgments-We thank Dr. I. M. London and Prof. L. A. Luzzatto for encouragement and Drs. I. Dokal, D. Huang, A. Bashey, and M. Coupe for providing tissue samples. We also thank Jean Black, who kindly typed the manuscript and A. Nunn who generously provided technical assistance during the early stages of this work.
REFERENCES 1. London, I. M., Bruns, G. P., and Karibian, D. (1964) Medicine (Baltimore)
2. Grayzel, A. I., Horchner, P., and London, I. M. (1966) Pmc. Natl. Acad.
3. Zucker, W. V., and Schulman, H. M. (1968) Proc. Natl. Acad. Sei. U. S. A.
4. Levere, R. D., and Granick, S. (1965) Proc. Natl. Acad. Sci. U. S. A. 54,
5. Ibrahim. N. G.. Gruensuecht. N. R.. and Freedman. M. L. (1978) Biochem.
4 3 , 789-802
Sei. U. S. A. 55,650-655
59 , 582-589
134-137
Biophys. Res. Comm;n. 80,722-128 6. Neuwirt, J., Ponka, P., and Borova, J. (1969) Eur. J. Biochem. 9,36-41 7. Aoki, Y., Wada, O., Urata, G., Takaku, F., Nakao, K. (1971) Biochem.
8. Ponka, P., and, Neuwirt, J. (1969) Blood 33,609-707 9. Idem (1971) Btochim. Biophys. Acta 230,381-392
Biophys. Res. Commun. 42,568-575
10. Iacopetta, B., and Morgan, E. (1984) Biochim. Biophys. Acta 805,211-216 11. Cox, T. M., O'Donnell, M. W., Aisen, P., and London, I. M. (1985) Proc.
12. Ponka, P., and Schulman, H. M. (1985) Blood 65,850-857 13. Ponka, P., Schulman, H. M., and Wilczynska, A. (1982) Biochim. Biophys.
14. Ponka, P., and Schulman, H. M. (1985) J. Biol. Chem. 260,14717-14721 15. London, I. M., Shemin, D., and Rittenberg, D. (1948) J. Biol. Chem. 173 ,
Natl. Acad. Sci. U. S. A . 82,5170-5174
Acta 718,151-156
m 7 - m ~
16. Walsh, R. J., Thomas, E. D., Chow, S. K., Fluarty, R. G., and Finch, C. A.
17. Watson, C. J., Bossenmaier, I., Cardinal, R., and Petryka, 2. J. (1974) Proc.
18. Ranu, R. S., and London, I. M. (1979) Methods Enzymol. 60,459-484 19. Cox, T. M., O'Donnell, M. W., Aisen, P., and London, I. M. (1985) J. Clin.
20. Schwartz, S., and Wikoff, H. M. (1952) J. Bid. Chem. 194,563-573 21. Fuhrop, J.-H., and Smith, K. M. (1975) in Laboratory Methods in Porphyrin
and Metallowmhvrin Research. PO. 47-48, Elsevier Scientific Publishing
.", ,"-
(1949) Science 110,396-398
Natl. Acad. Sci. U. S. A. 71, 278-282
Invest. 76,2144-2150
. " Co., Amsteidam
22. Gravzel. A. I.. Fuhr. J. E.. and London. I. M. (1967) Biochem. Biophys. Res. Cbmmun. 28,705-710
. .
23. Dailey, H. A., and Fleming, J. E. (1983) J. Biol. Chem. 258 , 11453-11459 24. Ali, F. M. K., May, A,, Jones, B. M., and Jacobs, A. (1983) Br. J. Haematol.
25. Lajtha, L. G., and Suit, H. D. (1955) Br. J. Haematol. 1 , 55-61 26. May, A,, de Souza, P., and Jacobs, A. (1982) in The Bzochemistry and
Physiology of Iron (Saltman, P., and Hegenauer, J., eds) pp. 249-253,
27. Ponka, P., and Schulman, H. M. (1986) in Porphyrins and Porphyrias Elsevier/North Holland, New York
(Nordmann, Y., ed) Vol. 134, pp. 56-67, Colloque INSERM, John Libbey, London
28. Ellory, J. C., Johnes, S. E. M., and Young, J. D. (1981) J. Physiol. (Lond.) 320,403-422
29. Rappoport, S. M. (1986) in The Reticulocyte, pp. 40-64, CRC Press Inc., Rocn Rnton. FI.
63,227-235
~- 30. Ross, J., and Sautner, S . (1976) Cell 8 , 513-520 31. Bonanou-Tzedaki, S. A,, Sohi, M. N., and Amstein, H. R. V. (1984) Eur.
J. Biochem. 144,589-596





























