Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY
Printed in U.S.A. Vol. 256, No. 11, Issue of June 10, pp. 5926-5930, 1981
The Isolation, Purification, and Characterization of the Collagen of Cysticercus cellulosae"
(Received for publication, April 15,1980, and in revised form, February 10,1981)
Alfonso Torre-BlancoS and I V O M e Toledo5 From the Departamento de Investigacion Clinica, Instituto Nacional de Pediatria Sistema Nacionalpara el Desavuollo Integral de la Familia and Escuela Nacional de Estudios Profesionales Cuautitlan, Universidad Nacional Aut0nom.a de M&>co, Mexico, Distrito Federal
Insoluble collagen fibrils were obtained from Cysti- cercus cellulosae after homogenization and treatment with NaCl/mercaptoethanol solutions and were solu- bilized after limited pepsin digestion. Solubilized Cys- ticercus collagen shows two different a subunits in sodium dodecyl sulfate-polyacrylamide gel electropho- resis with an apparent molecular weight of 100,000 and is readily degraded by bacterial collagenase. The amino acid composition is characteristic of collagen except that it contains no hydroxyproline. Segment long spac- ing crystallites measuring 280 nm in length were pre- pared. These segments showed a band pattern different from that of vertebrate and other invertebrate colla- gens. The denaturation temperature at neutral pH was 35 "C and correlated with the total pyrrolidine content as observed for other collagens. An intrinsic viscosity value of 15.3 dl/g was obtained for this collagen. Its possible evolutionary relationship with other collagens is discussed.
Comparative studies involving nonvertebrate collagen can contribute considerably to our understanding of the evolution of the collagen molecule. In addition, such studies can reveal adaptations of the same basic molecule to different functions.
Numerous investigations have been performed on the oc- currence of collagen in invertebrate species; characterizations, however, have been thorough in only a few cases, notably the cuticular collagens of annelids and nematodes (1).
The aim of this investigation was to gain insight into the comparative biochemistry of collagen. To this end it was decided to study the interstitial connective tissue of a species representative of a primitive phylum. It was expected that interstitial collagens would differ from cuticular collagens, given their very different functions.
In this paper the isolation, purification, and characterization of solubilized collagen from the platyhelminth cysticercus cellulosae, the larval stage of Taenia solium, are described. C. cellulosae lives in the muscular, subcutaneous, and nervous tissues of the pig and some other mammals; it is also the cause of human cysticercosis in the central nervous system (2).
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "aduertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. * Present address, Departmento de Biologia, Facultad de Ciencias, Universidad Nacional Autonoma de Mexico, Mexico.
8 Present address, Instituto de Investigaciones Biomedicas, Univ- ersidad Nacional Autonoma de Mexico, Mexico.
EXPERIMENTAL PROCEDURES
Materials-Live cysticerci were dissected from the muscles of infected pigs obtained from a local slaughterhouse, within 24 h after death, and stored frozen at -15 "C until use.
Pepsin (twice crystallized) and trypsin (type 111, twice crystallized) were obtained from Sigma. Clostridium collagenase (349 unitshl) was obtained from Worthington and electrophoresis reagents were from Bio-Rad. All other chemicals were of analytical grade and were used without any further purification.
Acid-soluble guinea-pig skin collagen was purified according to Trelstad et al. (31, lyophilyzed, and stored at -15 "C. Acid-soluble goat skin collagen was a gift of Dr. Ruy Perez Tamayo (Departmento de Patologia Experimental, Instituto Nacional de la Nutricion, Mex- ico).
Purification of Connective Tissue Fibrils-The cysticerci were thawed, rinsed three times in cold distilled water, and suspended in cold 0.45 M NaCl containing 0.02 M mercaptoethanol; after homoge- nization in an ice bath for 1 min with an Ultra-Turrax, they were extracted in the same solution at 8 "C for 48 h, with continuous agitation. The insoluble tissue was collected by centrifugation at 34,000 X g for 15 min, resuspended in cold 1.0 M NaCl containing 0.02 M mercaptoethanol, and re-extracted at 8 OC for an additional 48 h. The insoluble fibers were collected by centrifugation as above, rinsed several times in cold distilled water, and lyophylized.
After each of the extraction steps a sample of the insoluble residue was fixed in 2.5% glutaraldehyde (in 0.1 M sodium cacodylate buffer, pH 7.4), followed by postfiiation in 1% osmium tetroxide. The samples were embedded in Epon, sectioned, positively stained, and examined under a Jeol 100 B electron microscope operated at 60 kV, on Formvar-coated copper grids.
SolubiZization of Cysticercus Collagen-Lyophylized, purified fi- brils from Cysticercus were suspended in cold 0.5 M acetic (5 mg/ml) and pepsin was added (enzymembstrate ratio, 1:50, w/w). After 48 h of digestion with continuous stirring at 8 "C, the suspension was centrifuged at 34,000 X g for 15 min and the clear supernatant was dialyzed against several changes of 0.01 M Na2HP04 (pH 8.6). The white precipitate that formed was collected by centrifugation and redissolved in cold 0.5 M acetic acid. The solution was centrifuged 60 min at 105,000 X g to remove insoluble aggregates.
Estimation of Protein Content-The protein concentration of pu- rified solubilized Cysticercus collagen was measured by the method of Lowry et al. (22) using purified acid-soluble guinea-pig skin collagen as standard.
Viscosity Measurements-All the viscosity measurements were carried out in Cannon-Fenske type viscometers submerged in a water bath. The water bath temperature was always controlled within 0.1 "C of the stated temperature; the solutions were centrifuged 60 min at 105,000 X g prior to viscometry to remove insoluble aggregates.
Denaturation of Cysticercus Collagen-The denaturation temper- ature ( T D ) of Cysticercus collagen was determined by measuring the decrease in viscosity following denaturation (4). The solution of Cysticercus collagen in 0.1 M phosphate buffer, pH 7.2 (protein concentration 0.5 mg/ml), was incubated 30 rnin at each temperature before readings were taken; blank readings with buffer were taken at the same temperatures.
Electrophoresis-Sodium dodecyl sulfate-polyacrylamide gel elec- trophoresis was performed according to Hayashi and Nagai (5 ) ; stain- ing of gels was carried out with Coomassie brilliant blue (5, 6).
5926

Invertebrate Collagen 5927
Intrinsic Viscosity-The intrinsic viscosity of Cysticercus collagen was determined in 0.5 M acetic acid at 23 "C.
SLS crystallites. Acid-soluble guinea-pig skin, goat skin, and Cys- ticercus solubilized collagens, dissolved in 0.05 M acetic acid (0.5 mg/ ml), were dialyzed overnight in the cold room against 0.48 ATP in 0.05 M acetic acid. After diluting the resulting suspension to a faint cloudiness the crystallites were deposited on carbon-coated copper grids. Positive staining with phosphotungstic acid and uranyl acetate was performed according to Bruns and Gross (7). Negative staining was achieved with 2% sodium phosphotungstate, pH 7, for 4 min. The grids were examined under the electron microscope as described. The magnification was determined with a cross-lined replica of a diffrac- tion grating having 54,800 lines/inch (E. F. Fullam, Inc.)
Digestion of Cysticercus Soluble Collagen with Bacterial Colla- genase and Trypsin-Acid-soluble guinea-pig skin and Cysticercus collagens were dissolved in 0.05 M Tris, 0.15 M NaCI, 5 mM CaC12, pH 7.2 (collagenase buffer); 0.25 M glucose was added only to guinea-plg skin collagen to inhibit fibril formation (8) since Cysticercus soluble collagen does not form fibrils in this condition. The rate of collagen degradation was measured by viscometry at 30 "C. Trypsin (1 mg/ ml) or bacterial collagenase (0.5 mg/ml) were dissolved in collagenase buffer and 50 p liters of either enzyme were added to collagen previously equilibrated at 30 "C, the incubation mixture volume was 4 ml with a final collagen concentration of 0.65 mg/ml.
Amino Acid Analysis-The amino acid composition was kindly determined by Dr. Alejandro Alagon, Rockefeller University, N. Y. Amino acid composition was obtained by the method described by Moore and Stein (21). Duplicate samples were hydrolyzed in 6 N HCI at 110 "C for 20 and 48 h in sealed and evacuated tubes and analyzed in a Beckman Model 120 B amino acid analyzer modified for high sensitivity. Corrections were made for the destruction of threonine and serine and for the slow release of valine.
RESULTS
Purification of Cysticercus Collagen Fibrils-Electron mi- croscopical examination showed that the insoluble residue after the second extraction step consisted, almost exclusively, of fibrils identical with those observed in the intercellular space of Cysticercus tissues (Fig. 1). Connective tissue fibrils of Cysticercus measured 7 to 15 nm in diameter and did not show the typical cross-striation pattern of vertebrate colla- gens. In addition to fibrils, occasional cellular debris was observed in the insoluble residue; a further extraction step in 3 M urea containing 0.2 M mercaptoethanol did not modify this picture.
Solubilization-Lyophylized purified fibrils were almost completely insoluble in 0.5 M acetic acid, 8 M urea containing 0.2 M mercaptoethanol or 6 M guanidine hydrochloride; after 48 h of continuous stirring at 8 "C more than 85% of the initial material remained in insoluble form in the case of acetic acid and more than 90% remained after urea/mercaptoethanol or guanidine. The protein solubilized by acetic acid is a high molecular weight material that remains in the upper part of sodium dodecyl sulfate-polyacrylamide gels and presumably it is extensively cross-linked.
Pepsin, on the other hand, effectively solubilized the puri- fied fibrils, forming a very viscous solution. After dialysis against 0.01 M Na2HP04 (to inactivate pepsin) a white precip- itate appeared; this precipitate readily dissolved in 0.5 M acetic acid, forming a viscous solution.
Denaturation-Fig. 2 shows the change in specific viscosity, as a function of temperature, of a solution of Cysticercus collagen in 0.1 M phosphate buffer, pH 7.2; an abrupt fall in viscosity is observed at 36 "C. The solution always remained clear and transparent at every temperature, indicating that under these conditions solubilized Cysticercus collagen does not aggregate to form fibrils.
The melting curve was obtained by incubating for only 30 min at each temperature; this procedure is responsible for the sharpening at the low temperature end of the transition zone and leads to an increase of about 1 "C in the apparent TI) (4).
FIG. 1. Collagen fibrils of C. cellulosae. A, purified Cysticercus collagen fibrils (95,000); B. and C, sections through the bladder wall of a Cysticercus showing interstitial collagen fibrils (X 95,000 and 140,000, respectively).
4 20 temp * C
36
FIG. 2. Denaturation curve of Cysticercus collagen as meas- ured by viscosity in 0.1 M phosphate buffer, pH 7.2. The incu- bation time at each temperature was 30 min.
Digestion by Trypsin and Bacterial Collagenase-The rate of degradation of soluble Cysticercus collagen by trypsin and bacterial collagenase was determined measuring the loss in viscosity consecutive to the enzymatic scission of the molecule (9). Fig. 3 shows the change in viscosity of Cysticercus soluble collagen and guinea-pig skin collagen in the presence of either enzyme. The decrease in viscosity under the same conditions
5928 Invertebrate Collagen
but in the absence of either enzyme was l%/h for guinea-pig skin collagen and 2%/h for Cysticercus collagen.
The rate and extent of degradation of both collagens with bacterial collagenase is very similar. Even when Cysticercus collagen appears to be susceptible to the attack by trypsin, there is a remarkable difference between the rate of degrada- tion by trypsin and by bacterial collagenase; even without considering that the trypsin concentration is higher than that of bacterial collagenase, the time for the loss of 50% of the
20 40 60 mm
FIG. 3. Susceptibility of Cysticercus collagen and guinea-pig skin collagen to bacterial collagenase and trypsin as measured by viscosity at 30 "C and neutral pH. Cysticercus collagen was incubated in the presence of trypsin (0) and bacterial collagenase (0); guinea-pig skin collagen was incubated in the presence of trypsin (M) and bacterial collagenase (0).
gr N-
A B-. FIG. 4. SDS-polyacrylamide gel electrophoresis of Cysticer-
cus solubilized collagen ( A ) and acid-soluble guinea-pig skin collagen (B).
0
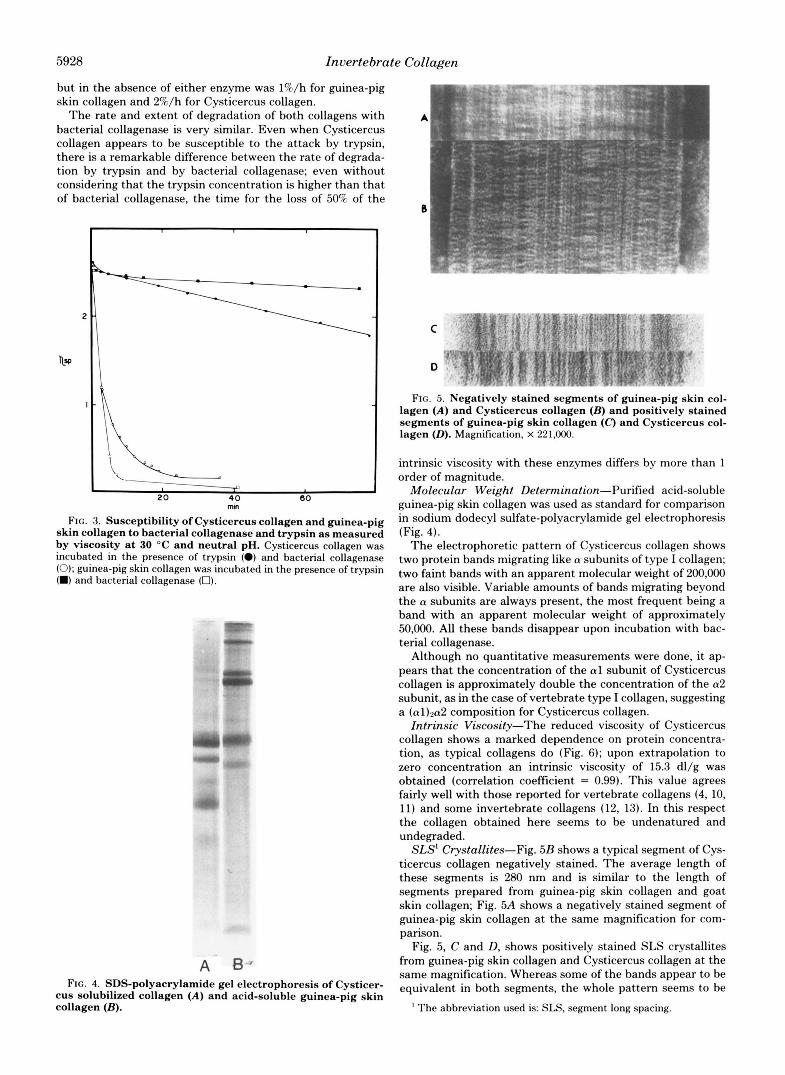
FIG. 5. Negatively stained segments of guinea-pig skin col- lagen (A) and Cysticercus collagen (B) and positively stained segments of guinea-pig skin collagen ( C ) and Cysticercus col- lagen (0). Magnification, X 221.000.
intrinsic viscosity with these enzymes differs by more than 1 order of magnitude.
Molecular Weight Determination-Purified acid-soluble guinea-pig skin collagen was used as standard for comparison in sodium dodecyl sulfate-polyacrylamide gel electrophoresis (Fig. 4).
The electrophoretic pattern of Cysticercus collagen shows two protein bands migrating like a subunits of type I collagen; two faint bands with an apparent molecular weight of 200,000 are also visible. Variable amounts of bands migrating beyond the a subunits are always present, the most frequent being a band with an apparent molecular weight of approximately 50,000. All these bands disappear upon incubation with bac- terial collagenase.
Although no quantitative measurements were done, it ap- pears that the concentration of the a1 subunit of Cysticercus collagen is approximately double the concentration of the a2 subunit, as in the case of vertebrate type I collagen, suggesting a (al)aa2 composition for Cysticercus collagen.
Intrinsic Viscosity-The reduced viscosity of Cysticercus collagen shows a marked dependence on protein concentra- tion, as typical collagens do (Fig. 6); upon extrapolation to zero concentration an intrinsic viscosity of 15.3 dl/g was obtained (correlation coefficient = 0.99). This value agrees fairly well with those reported for vertebrate collagens (4, 10, 11) and some invertebrate collagens (12, 13). In this respect the collagen obtained here seems to be undenatured and undegraded.
SLS' Crystallites-Fig. 5B shows a typical segment of Cys- ticercus collagen negatively stained. The average length of these segments is 280 nm and is similar to the length of segments prepared from guinea-pig skin collagen and goat skin collagen; Fig. 5A shows a negatively stained segment of guinea-pig skin collagen at the same magnification for com- parison.
Fig. 5, C and D, shows positively stained SLS crystallites from guinea-pig skin collagen and Cysticercus collagen at the same magnification. Whereas some of the bands appear to be equivalent in both segments, the whole pattern seems to be
' The abbreviation used is: SLS. segment long spacing.

Invertebrate Collagen 5929
amino acid residues, the proportion of imino acids is charac- teristically high and the aromatic amino acids are few. The most remarkable difference with most collagens is the absence of hydroxyproline; this imino acid was not detected in the amino acid analyzer and only traces of it with a highly sensi- tive colorimetric procedure (20).
a 0.2 03 0.4 OllOOml
FIG. 6. Reduced viscosity of Cysticercus collagen as a func- tion of protein concentration. Measurements were done in 0.5 M acetic acid at 23 "C.
TABLE I Amino acid composition of Cysticercus soluble collagen
Amino acid 20-h hydroly- 48-h hydroly- Average SIS= sis"
residues/1000 total amino acid residues
Hydroxyproline 0 0 0 Lysine 14.64 15.36 15 Histidine 4.85 4.73 5 Hydroxylysineb 18.80 19.20 19 Arginine 54.96 55.21 55 Aspartic acid 54.85 54.65 55 Threonine 28.35 21.03 28' Serine 27.04 22.38 27' Glutamic acid 97.50 97.68 98 Proline 216.00 218.60 217 Glycine 313.80 310.80 312 Alanine 55.21 55.81 56 Half-cystine 0 0 0 Valine 27.95 28.32 28' Methionine 23.50 22.60 23 Isoleucine 21.28 21.88 22 Leucine 25.66 25.91 26 Tyrosine 6.28 6.34 6 Phenvlalanine 8.22 8.33 8
Values are the average of two separate anaysis. The color factor for lysine was used.
'' Corrected values.
essentially different. With the same methods, SLS crystallites from two different mammalian species (guinea-pig and goat) give a similar band pattern which is comparable to that reported by Bruns and Gross for calf skin collagen (7) and by Nordwig and Hayduk for vertebrate and some invertebrate collagens (14).
In addition to complete SLS crystallites, preparations of Cysticercus collagen frequently show other crystallites meas- uring only half that length. These segments form long ribbons and usually aggregate and appear as dimers with a centrosym- metrical band pattern (not shown).
Amino Acid Composition-The amino acid composition of Cysticercus soluble collagen is given in Table I. In spite of some differences with vertebrate and some invertebrate col- lagens, Cysticercus collagen shows the characteristics of col- lagen molecules. Glycine accounts for nearly a third of all
DISCUSSION
The results of this investigation of some biochemical and biophysical properties of Cysticercus collagen have shown that it can be properly classified in the collagen group of molecules. The amino acid composition, high intrinsic viscos- ity, susceptibility to bacterial collagenase, and the ability to aggregate as SLS crystallites unambiguously label this protein as a member of the collagen family.
There is very good agreement between the subunit molec- ular weight, the size of the molecule as revealed by the electron microscope, and the value of the intrinsic viscosity. These data suggest a structure for Cysticercus collagen similar to that of vertebrate collagens: a molecule 280 nm long with a molecular weight of 300,000 formed by three subunits with individual molecular weights of 100,000 each.
The finding of SLS crystallites measuring about half the size of normal segments correlates with the presence of bands with an apparent molecular weight of 50,000 in electrophore- sis; they probably represent a degradation product of the solubilization process.
Considering its electrophoretic pattern, Cysticercus colla- gen appears to have two different types of (Y subunits, but further investigation by chromatography on cellulose ion ex- changers is needed to confm this suggestion. Although there is only scattered information on the subunit composition of interstitial invertebrate collagens, it would appear that the more primitive phyla show only one type of a subunit, as in the case of the sea anemones Actinza (14, 15) and Metridzum (16), whereas acantocephala (13), arthropoda, and echino- dermata (1) show two different types of a subunits; the phylum mollusca would seem to be at an intermediate position having members in both groups (1). Being derived from a member of a very primitive phylum we would expect the Cysticercus collagen to show only one type of (Y subunit; in spite of this its composition seems to be similar to that of collagen from more evolved phyla.
The denaturation temperature of Cysticercus collagen agrees fairly well, as expected, with the environment of this parasite. Recent evidence obtained with peptide models and with nonhydroxylated collagen indicates that hydroxyproline contributes more than proline to thermal stability (19); never- theless, the behavior of the present collagen agrees with the previous finding showing an almost linear relationship be- tween the TO values and the total pyrrolidine content (17).
Considering the thermal stability showed by Crysticercus collagen it is surprising to find the absence of hydroxyproline in its amino acid composition. The other collagen with a known amino acid composition from a platyhelminth contains almost the same amount of pyrrolidine residues as does Cys- ticercus collagen, but the former shows an usual degree of hydroxylation: about 46% (14); in spite of this, the thermal stability of both collagens is very similar. With respect to other amino acids the composition of both collagens compares fairly well, including hydroxylysine content. To our knowledge only the collagenous silk of a sawfly (18) and the present collagen seem to be devoid of hydroxyproline.
The absence of hydroxyproline coupled to the presence of hydroxylysine suggests that prolyl hydroxylase activity could be absent in C. cellulosae, whereas lysyl hydroxylase activity is normal. Work is under way in our laboratory to explore that

5930 Invertebrate Collagen
possibility and to test if Cysticercus collagen is a substrate for prolyl hydroxylase.
The difficulty in solubilizing Cysticercus collagen fibrils in the absence of pepsin and the examination of the material dissolved by 0.5 M acetic acid suggest that Cysticercus collagen may be extensively cross-linked although future research is needed to determine the nature of these cross-links, our results suggest that they are not disulfide bonds. At present we cannot rule out the possibility that the solubilization process with pepsin affects Cysticercus collagen, making it susceptible to attack by trypsin.
SLS crystallites have been reported in at least three inver- tebrate species: two coelenterates, Actiniu (14) and Metridiun (16), and the platyhelminth Fusciolu (14). In all cases the band pattern is indistinguishable from that of vertebrate collagens, indicating a similar distribution of polar amino acids along the molecule. In contrast with these observations, SLS crystallites prepared from Cysticercus collagen show a differ- ent pattern. Although the finding was initidly surprising, it shouId be noted that the three collagens previously mentioned aggregate to form fibrils exhibiting a definite band period under the electron microscope, whereas Cysticercus collagen forms very thin fibrils with no discernible band period at all. It seems reasonable to imagine that a changed distribution of charges in the molecule leads to a different form of aggregation at the fibril level.
In spite of some deviations from the vertebrate norm, already discussed, the collagen here described is closer to vertebrate collagens than to invertebrate cuticular collagens. Other interstitial invertebrate collagens already mentioned (1, 13-16) also seem to conform to the vertebrate rather than to the invertebrate cuticular pattern. These observations suggest that interstitial collagen is a remarkably stable molecule throughout evolution and that cuticular collagens are proba- bly the result of adaptations to a very different function.
Acknowledgments-we wish to thank Dr. A. Alagon and Dr. L. Possani for the amino acid analysis and Mr. Hector Oropeza for the
processing of samples for electron microscopy. We are indebted to Dr. R. Perez-Tamayo, Dr. I. Montfort, and Dr. A Pardo for their many helpful suggestions, encouragement, and criticism. We grate- fully acknowledge Dr. M. Rojkind for critically reading this manu- script.
REFERENCES 1. Adams, E. (1978) Science 202,591-598 2. Slais, J. (1970) Morphology and Pathogenicity of the Bladder
3. Trelstad, R. L., Kang, A. H., Igarashi, S., and Gross, J. (1970)
4. von Hippel, P. H. (1967) in Treatise on Collagen (Ramachandran,
5. Hayashi, T., and Nagai, Y. (1979) J. Biochem. 86,453-459 6. Blakesley, R. W., and Boezi, J. A. (1971) Anal. Biochem. 82,580-
7. Bruns, R. R., and Gross, J . (1973) Biochemistry 12,808-815 8. Terato, K., Nagai, Y., Kaeanishi, K., and Yamamoto, S. (1976)
Biochim. Bwphys. Acta 445,753-762 9. Perez-Tamayo, R. (1973) in Molecular Pathology of Connective
Tissues (Perez-Tamayo, R., and Rojkind, M., eds) Marcel Dek- ker, New York
10. Bensusan, H. B., Mumaw, V. R., and Scanu, A. W. (1962) Bio- chemistry 1,215-223
11. Veis, A., and Drake, M. P. (1963) J. Biol. Chem. 238,2003-2011 12. Fujimoto, D. (1968) Biochim. Biophys. Acta 168,537-543 13. Cain, G. D. (1970) Arch. Biochem. Biophys. 141, 264-270 14. Nordwig, A., and Hayduk, U. (1969) J. Mol. Biol. 44, 161-172 15. Nordwig, A., Nowack, H., and Hieber-Rogall, E. (1973) J. Mol.
16. Katzman, R. L., and Kang, A. H. (1972) J. Biol. Chem. 247,5486-
17. Josse, J., and Harrington, W. F. (1964) J. Mol. Biol. 9,269-287 18. Spiro, R. G., Lucas, F., and R u d d , K. M. (1971) Nature 231,54-
55 19. Prockop, D. J., Berg, R. A., Kivirikko, K. I. and Uitto, J . (1976) in
Biochemistry of Collagen (Ramachandran, G. N., and Reddi, A. H. eds) Plenum, New York
Worms, W. Junk N. V. Publishers, The Hague
Biochemistry 9,4993-4998
G. N., ed) Vol. 1, Academic Press, New York
582
Evol. 2, 175-180
5489
20. Woessner, J. F. (1961) Arch. Biochem. Biophys. 93,440-447 21. Moore, S., and Stein, W. H. (193) Methods Enzymol. 6,819-831 22. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.
(1951) J. Biol. Chem. 193,265-275




























