Embed Size (px)
Citation preview

THE JOURNAL OF BIOIAGICAL CHEMISTRY Vol. 254. No. 23, Issue of December 10, pp. 12209-12218, 1979 Printed in U.S. A.
Carbohydrate Structure of Yeast Invertase DEMONSTRATION OF A FORM WITH ONLY CORE OLIGOSACCHARIDES AND A FORM WITH COMPLETED POLYSACCHARIDE CHAINS*
(Received for publication, June 12, 1979)
Ludwig Lehle,$ Robert E. Cohen, and Clinton E. Balloug
From the Department of Biochemistry, University of California, Berkeley, California 94720
Invertase, extracted from broken cells of Saccharo- myces cerevisiae X-2180 mnn2 mannan mutant, was separated into a fraction insoluble in 75% ammonium sulfate (P75 invertase, 36% carbohydrate) and a soluble fraction (S75 invertase, 53% carbohydrate). The latter reacted with antibodies specific for the al + 6-linked mannose of the mannoprotein outer chain, whereas the P75 invertase failed to react with this antiserum al- though it did react with serum against terminal al + 3-linked mannose units that are characteristic of the mannoprotein core. A bacterial endo-al + 6-mannan- ase removed the outer chains from the S75 invertase and converted it to a form that was similar in electro- phoretic and immunochemical properties to the P75 invertase, whereas the endomannanase had little effect on the latter invertase. The results suggest that the P75 invertase is a form of the enzyme to which only the core oligosaccharide units had been added, and the S75 invertase represents an enzyme fraction to which the polysaccharide outer chains were also attached. A strong anomeric PMR signal for unsubstituted al -+ 6- linked mannose in the S75 invertase, and a much re- duced signal in the P75 invertase and endomannanase- digested S75 invertase, support these conclusions. Endo-N-acetyl-fi-glucosaminidase digestion of the S75 and P75 invertases, as well as of a purified wild type yeast invertase, produced an apparently identical se- ries of 3 to 4 carbohydrate-containing proteins that were separable by polyacrylamide gel electrophoresis in sodium dodecyl sulfate but that migrated as a singIe band on isoelectric focusing. The bands ranged from about 63,000 to 69,000 daltons and differed by the size of one or more carbohydrate core units each of 15 mannoses and 1 N-acetylglucosamine. The results sug- gest that the external invertase molecules contain some core units without attached outer chains, and that the cells contain a precursor form of the enzyme to which only the core units have been added. In support of this conclusion, PMR spectra and chromatographic pat- terns show that the core fragments from the P75, S75, and wild type invertases are essentially identical.
Because of their high mannose content, yeast extracellular
* This work was supported by National Science Foundation Grant PCM77-27334 and United States Public Health Service Grant AI- 12522 (to C. E. B.), and by a research fellowship from the Deutsche Forschungsgemeinschaft (to L. L.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accord- ance with 18 USC. Section 1734 solely to indicate this fact.
$ Present address, Fachbereich Biologie und Vorklinische Medizin, Universittitsstrasse, 8400 Regensburg (G.F.R.).
5 To whom inquiries should be addressed.
glycoproteins are called mannoproteins (1). The most exten- sively studied of these mannoproteins is the external invertase of Saccharomyces cereuisiae, an enzyme that is located out- side the plasma membrane in loose association with the cell wall (2). S. cereuisiae external invertase appears to be a dimer of 270,000 daltons with approximately 50% D-IIXinnOSe (3) that is attached to the protein as 18 to 20 asparagine-linked poly- saccharide units, or 9 to 10 units/protein subunit (4,5). Endo- N-acetyl-P-glucosaminidase H is reported to remove the car- bohydrate chains of invertase, by cleaving the di-N-acetylchi- tobiose unit linked to asparagine, to release a protein subunit of about 60,000 daltons (5) and the polysaccharide chains that can be separated into two size classes of about 26 or 52 mannose units that are terminated at the reducing end by a single N-acetylglucosamine (4). Although the intact external invertase migrates as a diffuse band on polyacrylamide gels in sodium dodecyl sulfate, partial deglycosylation by the endo- glucosaminidase produces 3 or 4 components that move as sharper bands and appear to differ by 3,000 to 4,000 daltons (5). These do not appear to be products of proteolysis, but they could represent artifactual products that differ from each other by integral numbers of carbohydrate chains.
Previous studies (6) have demonstrated that S. cerevisiae mannoprotein polysaccharide chains are differentiated into a core structure that is analogous, if not identical, to the man- nose-rich chains of mammalian glycoproteins (7), and an outer chain that is built up on the core by extension of the al 4 6- linked mannose backbone. Our ability to distinguish between these two parts of the molecule depends on an altered man- noprotein made by the mnn2 mutant of S. cereuisiae (8), which possesses an intact branched core but has a linear unbranched outer chain that can be removed by an endo-al + 6-mannanase (9), thus allowing isolation of the core. The mnn2 mannoprotein can be recognized by its reaction with antiserum against the al + 6-linked mannose chain (a), whereas exposure of the core by removal of the outer chain enhances reaction with antiserum against terminal al + 3- linked mannose (6).
We have investigated the invertase in the S. cereuisiae mnn2 mutant and have demonstrated a form that possesses essentially only the carbohydrate core units, and another form that possesses some completed polysaccharide chains and some apparently unmodified core units. The results support a biosynthetic pathway in which the carbohydrate core is added to the protein, probably as a unit (10, ll), followed by a stepwise elaboration of the outer chain, probably by addition of single mannose units (12). Some core units are so located or are of such a nature that they are not susceptible to modification and, thus, lack the outer chain. We demonstrate that baker’s yeast invertase has a similar structure, suggesting that our observations have general validity and are not a consequence of the mnn2 mannan mutation.
12209
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from

12210 Carbohydrate Structure of Yeast Invertase
EXPERIMENTAL PROCEDURES
Materials-Bio-Gel P-4 (-400 mesh), Bio-Gel P-60 (100 to 200 mesh), Bio-Gel P-100 (100 to 200 mesh), Bio-Gel P-200 (100 to 200 mesh), ampholytes for isoelectric focusing, and material for polyacryl- amide electrophoresis were from Bio-Rad; DEAE-Sephadex A-50, CM-Sephadex C-50, DEAE-Sephacel, and agarose were from Phar- macia; phenylmethylsulfonyl fluoride was from Sigma; and ultrapure urea came from Schwarz/Mann. Bacillus circulans ATCC 29101 and Streptomyces plicatus ATCC 27800 were from the American Type Culture Collection, and various yeast strains were provided by Lun Ballou from the laboratory collection. Bulk cell wall mnn2’ mannan was precipitated by Fehling’s solution, subjected to p-elimination and chromatographed on Bio-Gel P-2 to remove oligosaccharides linked to serine and threonine (13). Escherichia coli RNA polymerase was a gift from George Kassavetis of this department, and wild type invertase (Grade VII) was from Sigma. Glucostat Special reagent was obtained from Worthington. Acetonitrile, UV grade, was from Burdick and Jackson Laboratories, and deuterated solvents were from Bio- Had and Aldrich Chemical Co. All other chemicals were of reagent grade and were obtained from commercial sources.
Isolation and Purification of Znvertase-Saccharomyces cerevi- sine X2180-IA (wild type) and the mutant mnn2-la (LBI-16A) were grown at 30°C in a medium containing 1% yeast extract, 2% Bacto- peptone, and 2% glucose. For optimal production of invertase, 0.5% sucrose was added immediately after the glucose was depleted. Cells were usually harvested about 2 h later. Alternatively, after growth on glucose, the cells were washed with 0.9% NaCl and resuspended in a medium containing 0.25% yeast extract, 0.125% Bacto-peptone, and 0.5% sucrose. Growth was monitored by absorbance at 578 nm in a Bausch and Lomb Spectronic 100 spectrophotometer, after diluting samples to an absorbance not exceeding 0.2 optical density unit.
Invertase was isolated from the mnn2 strain by a modification of the method of Neumann and Lampen (14). The organism was grown in a 200-liter fermenter that was inoculated with 5 liters of logarith- mically growing cells, and approximately 3.5 kg of cell paste was obtained after the shift to sucrose as described above. All of the purification steps were performed at 5°C unless specified otherwise.
Wild type yeast invertase, purchased from Sigma (Grade VII) was dissolved in water and dialyzed against 10 mM sodium phosphate, pH 6.5. The dialyzed fraction was fractionated on a column (2.5 X 30 cm) of DEAE-Sephacel equilibrated with 10 mM sodium phosphate, pH 6.7. The column was washed with 10 mM buffer and developed with a 12O@ml NaCl gradient from 0 to 0.2 M in 10 mM sodium phosphate buffer, pH 6.7. Fractions with the highest specific activity were pooled and concentrated by ultrafiltration with an Amicon XM-50 filter.
Analytical Methods-Invertase preparations (20 pl), appropriately diluted to allow determination of the initial rates, were added to 200 ~1 of 0.05 M sodium acetate, pH 4.5. The reactions were started by addition of 200 ~1 of prewarmed 7% sucrose, then incubated at 3O”C, and after 10 to 15 min the tubes were heated for 2.5 min in a boiling water bath and the glucose contents were determined with the Glu- costat Special reagent. One unit of P-fructofuranosidase activity is defined as 1 pmol of sucrose hydrolyzed/min at 30°C. Invertase activity of column fractions was determined with the dinitrosalicylate reagent, reduced to one-tenth of the original volumes (15).
Total carbohydrate was determined by the phenol-sulfuric acid method (16), and reducing sugar was measured by the method of Nelson-Somogyi (17), with mannose as a standard for both proce- dures. Protein was assayed according to Lowry et al. (18), with bovine serum albumin as a standard. Carboxymethylation of denatured in- vertase was done as described by Kuhn et al. (19). After this treatment the protein became insoluble; to facilitate enzymic carbohydrate removal (see below), the precipitated protein was dissolved in 0.01 N
NaOH and then adjusted with acid to a pH of 6.5. Removal of Carbohydrate from Znvertase-Two specific glycosi-
dases were used for the structural analysis of the carbohydrate component of invertase. Endo-N-acetyl-/3-glucosaminidase H was pu- rified from the cultural filtrate of Streptomyces plicatus according to Tarentino and Maley (20), whereas endo-al + 6-mannanase was isolated according to Nakajima et al. (9). Both enzyme preparations were free from proteinase activity when tested with ovalbumin as substrate. Carbohydrate release from native and carboxymethylated
’ The abbreviations used are: P75 and S75 invertases, the fractions precipitated and soluble in 75% ammonium sulfate; mnn2, designation for the S. cerevisiae mannan mutant with a defective cul + 2-man- nosyltransferase I activity.
invertase by the endoglucosaminidase was done as described by Trimble and Maley (5). Enzymic removal of the al -+ 6-mannan outer chain was performed according to Nakajima and Ballou (6), in 20 mM sodium citrate, pH 6. Samples were withdrawn from the incubations and frozen for subsequent analysis by gel electrophoresis. The reference core oligosaccharide was prepared from cell wall mnn2 mannoprotein by the action of purified endomannanase and endoglu- cosaminidase H, and it was isolated as previously described (6).
Immunological Methods-Antimannan sera were obtained by im- munizing rabbits intravenously with ethanol-killed yeast cells accord- ing to a published protocol (8). Antisera having immunochemical reactivity toward crl -+ 3-linked nonreducing mannose (anti-X2180 serum), and with specificity for the oil + 6-mannan chain (anti-mnnl, mnn2 serum), were used. Ouchterlony immunodiffusion tests on aga- rose gel (21) were performed at room temperature for 18 h.
Gel Electrophoresis and Isoelectric Focusing-Acrylamide gel electrophoresis (7.5% acrylamide) in sodium dodecyl sulfate was done in the discontinuous system of Laemmli (22), with a running buffer of Tris-HCl, pH 7.2. Samples were reduced with 1% mercaptoethanol in 1% sodium dodecyl sulfate at 100°C for 3 min. Gels were stained for protein with Coomassie brilliant blue R-250. To detect carbohydrate, duplicate gels were treated with the periodic acid-fuchsin stain (23). RNA polymerase PP’-subunits (165,000 and 155,000), o-subunit (90,000), a-subunit (40,000), bovine serum albumin (68,000), ovalbu- min (45,000), catalase (60,000), and aldolase subunit (40,000) were used to calibrate the gels for molecular weight estimations.
Isoelectric focusing was done according to O’Farrell (24) on 4% polyacrylamide gels using a pH range from 3 to 10. The total ampho- line concentration was 2%, comprising the pH ranges 3 to 10, 3 to 6, and 5 to 7 in the ratio of 1:2:2. The gels were electrofocused for 15 h at 300 V and then for 1.5 h at 500 V.
High Pressure Liquid Chromatography-The core oligosaccha- rides were separated on a Waters Associates model ALC/GPC 201 instrument with a pBondapack/Carbohydrate column (0.4 X 30 cm). The solvent was a 55:45 (v/v) mixture of acetonitrile-water. The effluent was monitored with a Waters Associates differential refrac- tometer.
Proton Magnetic Resonance Spectroscopy-PMR spectra were obtained at 180 MHz on an instrument equipped with a Bruker superconducting magnet and Nicolet 1180 computer operated in the Fourier transform mode. Spectra were obtained at a probe tempera- ture of 40.0 + 0.2’C using a 9O’C pulse width, 1500 Hz spectral width, and 2.7 s acquisition time. Data were accumulated into 8192 addresses, and a digital broadening of 0.05 or 0.10 Hz was used to enhance sensitivity. Chemical shifts are expressed relative to sodium 3-(tri- methylsilyl)-propanesulfonate referenced indirectly to internal ace- tone (6 = 2.217 ppm at 4O.O”C). Samples were prepared by passing the aqueous solutions through l-ml columns of Chelex 100 (Bio-Rad) to remove metal contaminants, exchanging them three times with D20 by lyophilization, and dissolving the product in 100.0% DzO (Aldrich) to a final concentration varying from 0.1 to 2.0 mM.
RESULTS
Derepression of External Invertase Synthesis
The formation of external invertase activity in S. cerevisiae X2180 is under catabolite repression when cells are grown in the presence of glucose (2). With the mnn2 mutant strain used in this study, invertase production increased more than 20- fold above the basal level if glucose-grown cells were shifted to a sucrose-containing medium shortly before glucose was exhausted (data not shown). Essentially the same result was obtained by adding sucrose, to a concentration of 0.5%, directly to the original medium after the glucose concentration was reduced to less than 0.1% (Fig. 1). This enabled us to produce invertase from cells grown in a large fermenter and thus obtain sufficient material for structural’analysis on the carbohydrate component of the mutant mannoproteins. A maximal rate of invertase formation was usually observed 1 to 2 h after sucrose addition.
Purification of Invertases Soluble and Insoluble in 75% Ammonium Sulfate
During the mnn2 invertase purification, we noticed that the
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from

Carbohydrate Structure of Yeast Invertase 12211
251 I I I I I Sucrose
- 2.5
120
Time (hours)
FIG. 1. Kinetics of external invertase formation in the mnn2 mutant of S. cereukiae. Cells were grown in the yeast extract/ peptone/glucose medium, and sucrose was added to a concentration of 0.5% at the indicated time. Samples were removed at intervals and analyzed for cell mass (As7,, closed circles), glucose in the medium (mg/ml, open circles), and invertase activity (units/ml of cells with an absorbance of 10 at 578 nm, open triangles).
enzyme activity separated into two fractions. The key step involved a fractionation with 75% ammonium sulfate, appar- ently due to differences in carbohydrate content of the en- zymes. Most of the activity remained soluble (here called S75 invertase), whereas about 10% was precipitated (called P75 invertase). The P75 fraction initially also contained a small amount of internal carbohydrate-free invertase that was re- moved during DEAE-Sephadex chromatography. The frac- tionation followed the flow diagram in Fig. 2, and details are given in the following paragraphs.
Isolation of S75 Invertase-S. cerevisiae mnn2 cells (3.3 kg) were suspended in an equal volume of 0.01 M sodium phosphate buffer, pH 6.5, containing 1 mu phenylmethylsul- fonyl fluoride. The cells were broken by five passages through a prechilled Manton-Gaulin press at 12,000 p.s.i. The homog- enate was centrifuged for 90 min at 13,700 X g, giving a turbid supernatant liquid of 5,100 ml, which was kept overnight at 4°C. To remove ribosomes and nucleic acid, 10 ml of 10% streptomycin sulfate solution/IO g of protein was added slowly and with constant stirring. After an additional 5 h, the solution was centrifuged for 60 min at 13,700 x g.
The supernatant solution was stirred and heated rapidly to 5O”C, maintained at this temperature for 30 min, and then cooled in an ice bath and centrifuged for 60 min at 13,700 x g. To the heat-treated supernatant solution, solid ammonium sulfate was added slowly with continuous stirring to 75% of saturation at 4°C. After 5 h, the precipitate was separated by centrifugation for 60 min at 13,700 X g and was saved for isolation of the P75 invertase (see below). The supernatant solution containing the soluble invertase was dialyzed against 10 mM phosphate buffer, pH 6.5, using an Amicon Diaflo- Hollow Fiber system with the HP-10 cartridge.
The dialyzed enzyme solution was adsorbed on a DEAE- Sephadex column (4 x 35 cm) that had been equilibrated with 10 mM sodium phosphate, pH 6.5. After the column was washed with the equilibration buffer until all unbound protein was removed, a 2-liter linear gradient of 0 to 0.4 M NaCl in 10 mM sodium phosphate buffer was used for elution. Fractions with the highest activity were pooled and dialyzed against water.
After dialysis, the enzyme solution was adjusted to 10 mM
sodium citrate, pH 3.7, and applied to an SP-Sephadex C-50 column (4 x 35 cm) that had been equilibrated with 10 mM
sodium citrate, pH 3.7. The column was washed with one column-volume of the equilibrating buffer before the enzyme was eluted with a 2-liter linear gradient of 0 to 0.3 M NaCl in 10 mM citrate buffer. The active fractions were combined and concentrated in an Amicon ultrafiltration cell with an XM-50 filter.
The enzyme fraction from above was loaded onto a Bio-Gel P-200 column (2 x 90 cm) equilibrated with 10 mM sodium phosphate, pH 6.9, containing 0.1 M NaCI. The column was eluted with this buffer and the active fractions were pooled, dialyzed against water, and lyophilized. Table I summarizes a typical purification.
Isolation of P75 Invertase-The invertase fraction insolu- ble in 75% ammonium sulfate (see above) was used for the purification of invertase containing only core carbohydrate units. The pellet was dissolved in 10 mM sodium phosphate buffer, pH 6.5, and to the solution ammonium sulfate was added to 40% of saturation. The supernatant liquid was di- alyzed against water, adjusted to pH 6.5 with sodium phos- phate buffer, and the invertase was purified to homogeneity by chromatography on DEAE-Sephadex, SP-Sephadex, and finally by gel filtration on Bio-Gel P-200 (2 x 90 cm) under conditions used for the S75 invertase. The yield of P75 in- vertase was 10% of the S75 form.
Properties of the Invertase Preparations-Fig. 3 shows the elution profile of the two invertases on SP-Sephadex, and at this stage they were nearly homogeneous. Whereas the spe- cific activity of the S75 fraction was constant throughout the peak fractions, the carbohydrate component varied and de-
Crude cell extract
1 i; ~~;~~~2~“;~~Y~~~ Pelteted
Supernatant fraction fraction
i 40% Ammonium sulfate HoIlk+fiber supernatant fraction concentrate
i OEAE-Sephadex
:r;~~~
DEAE-Sephadex
1 SP-Sephadex
4 1 Bio-Gel P-ZOO
1 Carbohydrate- i P75 Invertase free invertase S75 Invertase
FIG. 2. Flow diagram showing the steps in the fractionation of the S. cerevisiae X2180 mnn2 invertase. Details are given in the text.
TABLE I
Purification of S75 invertase from the S. cerevisiae mnn2 mutant
Purification step Total protein
Total activ-
ity Specific RWXW- activity -Y
Crude extract Streptomycin sulfate frac-
tion Heat treatment 75% Ammonium sulfate su-
pernatant hollow fiber concentrate
DEAE-Sephadex A-50 SP-Sephadex C-50 Bio-Gel P-200
261,375 217,150
78,000 13.490
54 55 1,020 19 18 43 2,410 15 16 41 2,560 14
rnits x units/mg/ 10-3 min
294 1.13 283 1.3
220 2.8 141 10.5
5%
100 96
74 48
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from

Carbohydrate Structure of Yeast Invertase
Fraction
FIG. 3. Chromatography of invertase preparations on SP- Sephadex C-50. Invertase fractions, S75 in A and P75 in B, were loaded on the column and eluted with a 0 to 0.3 M NaCl gradient. Invertase activity (A520, closed circles), carbohydrate (A,w, open circles), protein (Az~, open triangles), and conductivity (mmho, closed triangles) are shown.
creased on the right side of the peak. The P75 invertase was more homogeneous and had a lower carbohydrate content (Table II). Presumably, the reduced carbohydrate content was responsible for the stronger binding to the column; the S75 fraction was eluted at a conductivity of 1.8 mmho, whereas the P75 fraction required a higher salt concentration.
The S75 invertase possessed a specific activity of 2,500 units/mg, and the P75 form had a similar activity (2,400 units/ mg). This agrees with the finding of Smith and Ballou (15) that carbohydrate-depleted invertase obtained by exoman- nanase digestion retained full activity. However, the P75 invertase was more difficult to solubilize after lyophilization than was the S75 invertase. Both invertase fractions migrated as homogeneous but, for glycoproteins, typically diffuse bands on acrylamide gels in sodium dodecyl sulfate (Fig. 4). The S75 invertase had a carbohydrate content of 53% and an average mass around 120,000 daltons, whereas the P75 invertase with 36% carbohydrate migrated in the 90,000-dalton region. Sub- sequent experiments demonstrated that the P75 fraction rep- resented an invertase that carried only the carbohydrate cores, whereas the S75 invertase was a larger form in which some of the core units carried attached outer chains of polysaccharide.
Removal of the Carbohydrate Component of the Invertase Preparations
The mnn2 mutant mannoproteins possess an unbranched al + &linked outer chain that can be removed by an endo- al -+ 6-mannanase, and a branched core structure that can be removed by an endo-N-acetyl+-glucosaminidase (6). As seen in Fig. 4, lane 5, after treatment with endomannanase the S75 band migrated on the gel at a new position similar to that of the P75 invertase. The P75 invertase was apparently un- changed by the same treatment, although some sharpening of the P75 band did occur in one long-term incubation with the endomannanase. A subsequent digestion of either invertase with endoglucosaminidase released additional carbohydratti, and both the S75 and P75 invertases then migrated as 3 or more sharp protein- and carbohydrate-staining bands. The patterns were similar and the bands appeared to differ in mass by about 3,000 daltons, or approximately the size of a core unit of 15 mannoses and one N-acetylglucosamine (25). The fastest band migrated at about 63,000 daltons, whereas car- bohydrate-free invertase is reported to have &Z, = 60,000 (5).
It is unlikely that these multiple bands are products of
proteolysis, because isoelectric focusing patterns showed no difference before and after endoglucosaminidase digestion (data not shown). It is probable that the bands represent forms of invertase possessing variable numbers of residual core units that are not readily accessible to the enzyme. Fig. 4 shows that an identical pattern was obtained after endo- glucosaminidase treatment of purified wild type invertase, although the endomannanase had no effect on this enzyme because the outer chain of the mannan component is branched. From the number of protein bands, it appears that some of the invertase molecules have at least 4 unsubstituted core units/monomer in the native state. Calculations given below suggest the number may be even higher.
The endoglucosaminidase digestion products of the S75 invertase were separated into three fractions by gel filtration on Bio-Gel P-60 (Fig. 5), and Fractions A and C were purified by rechromatography on Bio-Gel P-100 and P-4, respectively. Polyacrylamide gel electrophoresis of Fraction A in sodium dodecyl sulfate gave a pattern (not shown) that was identical to that in Fig. 4 for the endoglucosaminidase limit digest. The carbohydrate in Fraction C was eluted from Bio-Gel P-4 in the same volume as core fragments prepared from mnn2 bulk cell wall mannoprotein, indicating a size of 13 to 18 sugars (25), and analysis by high pressure liquid chromatography and PMR (see below) confirmed that Fraction C was a mixture of core units.
The material in Fraction B was degraded by endomanna- nase digestion to a mixture of core fragments and mannose plus al -+ 6-mannooligosaccharides (Fig. 6), all expected products of the action of this enzyme. The core fragments were similar to those from the mnn2 mannoprotein. Thus,
TABLE II Properties of invertase preparations
IlNWtW2 Protein Carbohydrate Subunit size range
% 5% x lo+
mnn2-S75 47 53 go-140 mnn2-P75 64 36 75-100 Wild type 45 55 95-155
I23 456789 FIG. 4. Polyacrylamide gel electrophoresis in sodium dode-
cyl sulfate of invertase preparations. Lanes 1 to 3 are wild type invertases, Lanes 4 to 6 are mnn2S75 invertase, and Lanes 7 to 9 are mnn2-P75 invertase. The first sample of each fraction is the native enzyme, the second is after endomannanase digestion, and the third is following endoglucosaminidase digestion. For the enzyme diges- tions, 0.5 mg of invertase was incubated 24 h at 45°C under a drop of toluene in 5 mM sodium citrate, pH 6, containing 10 pM CaClz and 0.25 unit of endomannanase in a total volume of 0.1 ml, after which a 5-g sample was taken for electrophoresis. To the remainder, 20 ~1 of 50 mM sodium citrate, pH 5.5, and 0.02 unit of endoglucosaminidase were added, the mixture was digested at 38°C for 48 h, and again subjected to ilectrophoresis. Digestion with an additional 0.02 unit of endoglucosaminidase did not change the pattern.
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from

Carbohydrate Structure of Yeast Invertase 12213
a discrepancy, however, in the estimates of the chain lengths for the smaller fragments, to which we have assigned the average size of 15 sugar units. The value of 26 determined by Tarentino et al. for the small oligosaccharide (4) could result from cross-contamination of the two fractions because their
-3
-2
-I_
I.0
:.o c .- a ', cc
0
0.
0.
0. 2 e -0 F 8 5 0.
0
.3-
.2-
I.1 -
OO 5
Fraction
FIG. 5. Separation of endoglucosaminidase digestion prod- ucts of S75 invertase. The invertase (12 mg), in 0.5 ml of 50 mM sodium citrate, pH 5.5, was digested at 38°C with 0.25 unit of endo- glucosaminidase. After 48 h, another 0.1 unit of endoglucosaminidase was added and the digestion was continued for 48 h, after which the products were chromatographed on a Bio-Gel P-60 column (1 X 90 cm) by elution with water. Carbohydrate (A,,) is shown in open circles and protein (A& is shown in closed circles. Fraction A (27% of the mannose) is residual invertase with some carbohydrate still attached, Fraction B (61% of the mannose) is core carbohydrate units with attached outer chains of varying length, and Fraction C (12% of the mannose) is carbohydrate core units without outer chains.
Fraction B represents the large polysaccharide chains of the invertase molecule.
Fractions A, B, and C from the endoglucosaminidase diges- tion of S75 invertase accounted for 27%, 61%, and 12% of the total carbohydrate. Endomannanase digestion of Fraction B indicated that 18% of the carbohydrate was in core units, the remainder being in al + 6-linked outer chains. Thus, of the chains released by the endoglucosaminidase, the ratio of core- sized units to cores with attached outer chains is 12/(61 X 0.18), or 1.1 to 1. We assume that the carbohydrate remaining on the invertase consists exclusively of core units averaging about 15 mannoses (25) because this material failed to react with al + g-specific antiserum (see below) and migrated on gels as a series of sharp bands differing in size by about 3000 daltons (Fig. 4). Moreover, after carboxymethylation of Frac- tion A, additional carbohydrate was released by endogluco- saminidase digestion, all of which was eluted from a Bio-Gel P-60 column in the position of core oligosaccharides (data not shown). Consequently, for every 2.1 chains released by the enzyme, approximately 2.5 chains (1.1 X 27 + 12) resisted cleavage. From this, we calculate that only about 2 of the 9 carbohydrate chains/invertase monomer had attached outer chains.
Digestion of wild type invertase with the endoglucosamini- dase and chromatography on Bio-Gel P-60 gave a pattern similar to that of the S75 invertase (Fig. 7). Again, three fractions were obtained, corresponding to protein with resid- ual carbohydrate attached (A), large oligosaccharides (B), and core-sized units (C). Tarentino et al. (4) observed two size classes of released carbohydrate similar to Fractions B and C. Because the large wild type carbohydrate chains cannot be degraded by the endomannanase to yield cores, we were unable to determine the stoichiometry of these two chain types. From glucosamine analyses, Tarentino et al. (4) con- cluded these were produced in equimolar amounts, which agrees with our findings for the S75 mnn2 invertase. There is
80 120
Fraction
FIG. 6. Separation of endomannanase digestion products of Fraction 5B from Fig. 5. Fraction 5B was digested for 72 h under toluene in 0.5 ml of 50 mM sodium citrate, pH 6.0, containing 50 pM CaCl* and 2 units of endomannanase. The products were separated on a Bio-Gel P-4 column (1.3 x 115 cm) by elution with water. Mannose and (Al+ 6-mannooligosaccharides eluted in Fractions 100 to 140, whereas the core units from which the outer chains were removed eluted in Fractions 80 to 100. The latter represented 18% of the carbohydrate in Fraction 5B.
0) zi
0.10 0.2
G 2 2%
.E
z s 2 0
0.05 0.1
0 0
Fraction
FIG. 7. Separation of endoglucosaminidase digestion prod- ucts of wild type invertase. The wild type invertase (10 mg) was digested and the products were fractionated under the conditions described in the legend to Fig. 5.
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from
12214 Carbohydrate Structure of Yeast Invertase
separation procedure did not have a high resolving power for molecules of this size.
Immunochemical Properties of Invertases
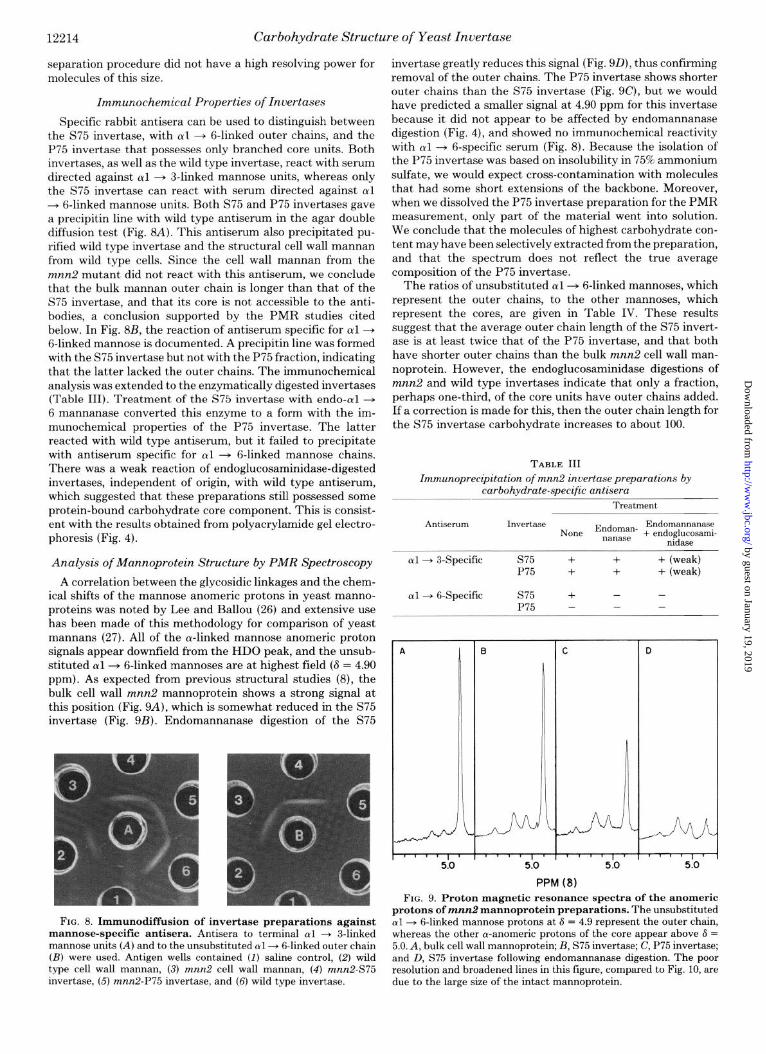
Specific rabbit antisera can be used to distinguish between the S75 invertase, with cul + g-linked outer chains, and the P75 invertase that possesses only branched core units. Both invertases, as well as the wild type invertase, react with serum directed against al + 3-linked mannose units, whereas only the S75 invertase can react with serum directed against al --+ 6-linked mannose units. Both S75 and P75 invertases gave a precipitin line with wild type antiserum in the agar double diffusion test (Fig. 8A). This antiserum also precipitated pu- rified wild type invertase and the structural cell wall mannan from wild type cells. Since the cell wall mannan from the mnn2 mutant did not react with this antiserum, we conclude that the bulk mannan outer chain is longer than that of the S75 invertase, and that its core is not accessible to the anti- bodies, a conclusion supported by the PMR studies cited below. In Fig. 8B, the reaction of antiserum specific for al + 6-linked mannose is documelited. A precipitin line was formed with the S75 invertase but not with the P75 fraction, indicating that the latter lacked the outer chains. The immunochemical analysis was extended to the enzymatically digested invertases (Table III). Treatment of the S75 invertase with endo-al + 6 mannanase converted this enzyme to a form with the im- munochemical properties of the P75 invertase. The latter reacted with wild type antiserum, but it failed to precipitate with antiserum specific for al + 6-linked mannose chains. There was a weak reaction of endoglucosaminidase-digested invertases, independent of origin, with wild type antiserum, which suggested that these preparations still possessed some protein-bound carbohydrate core component. This is consist- ent with the results obtained from polyacrylamide gel electro- phoresis (Fig. 4).
Analysis of Mannoprotein Structure by PMR Spectroscopy
A correlation between the glycosidic linkages and the chem- ical shifts of the mannose anomeric protons in yeast manno- proteins was noted by Lee and Ballou (26) and extensive use has been made of this methodology for comparison of yeast mannans (27). All of the a-linked mannose anomeric proton signals appear downfield from the HDO peak, and the unsub- stituted al+ 6-linked mannoses are at highest field (S = 4.90 ppm). As expected from previous structural studies (8), the bulk cell wall mnn2 mannoprotein shows a strong signal at this position (Fig. 9A), which is somewhat reduced in the S75 invertase (Fig. 9B). Endomannanase digestion of the S75
FIG. 8. Immunodiffusion of invertase preparations against mannose-specific antisera. Antisera to terminal al + 3-linked mannose units (A) and to the unsubstituted cul + &linked outer chain (B) were used. Antigen wells contained (I) saline control, (2) wild type cell wall mannan, (3) mnn2 cell wall mannan, (4) mnn2-S75 invertase, (5) mnn2-P75 invertase, and (6) wild type invertase.
invertase greatly reduces this signal (Fig. 9D), thus confirming removal of the outer chains. The P75 invertase shows shorter outer chains than the S75 invertase (Fig. 9C), but we would have predicted a smaller signal at 4.90 ppm for this invertase because it did not appear to be affected by endomannanase digestion (Fig. 4), and showed no immunochemical reactivity with ~yl -+ 6-specific serum (Fig. 8). Because the isolation of the P75 invertase was based on insolubility in 75% ammonium sulfate, we would expect cross-contamination with molecules that had some short extensions of the backbone. Moreover, when we dissolved the P75 invertase preparation for the PMR measurement, only part of the material went into solution. We conclude that the molecules of highest carbohydrate con- tent may have been selectively extracted from the preparation, and that the spectrum does not reflect the true average composition of the P75 invertase.
The ratios of unsubstituted al--+ 6-linked mannoses, which represent the outer chains, to the other mannoses, which represent the cores, are given in Table IV. These results suggest that the average outer chain length of the S75 invert- ase is at least twice that of the P75 invertase, and that both have shorter outer chains than the bulk mnn2 cell wall man- noprotein. However, the endoglucosaminidase digestions of mnn2 and wild type invertases indicate that only a fraction, perhaps one-third, of the core units have outer chains added. If a correction is made for this, then the outer chain length for the S75 invertase carbohydrate increases to about 100.
TABLE III Zmmunoprecipitation of mnn2 invertasepreparations by
carbohydrate-specific antisera
Treatment
Antiserum IWWtaSC3 Endomannanase None Endoman* + endoglucosami- nanase nidaae
al--+ 3-Specific s75 + + + (weak) P75 + + + (weak)
al + 6-Specific s75 + - - P75 - - -
PPM (8) FIG. 9. Proton magnetic resonance spectra of the anomeric
protons of mnn2 mannoprotein preparations. The unsubstituted al -+ &linked mannose protons at 6 = 4.9 represent the outer chain, whereas the other a-anomeric protons of the core appear above 6 = 5.0. A, bulk cell wall mannoprotein; B, S75 invertase; C, P75 invertase; and D, S75 invertase following endomannanase digestion. The poor resolution and broadened lines in this figure, compared to Fig. 10, are due to the large size of the intact mannoprotein.
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from

Carbohydrate Structure of Yeast Invertase 12215
The PMR spectrum (Fig. 10) of the unfractionated core oligosaccharide from the S75 invertase was almost identical with that from the cell wall mannoprotein, which supports the conclusion that they are constructed in essentially the same way. The various anomeric proton signals in Fig. 10 have been assigned by comparison with reference compounds and are listed in Table V. The spectra confirm that the core oligosac- charide is highly branched, with at most one unsubstituted oil + B-linked mannose per N-acetylglucosamine unit. The other signals are consistent with the cul -+ 2 and cwl + 3 linkages, and the branched cul 4 6-linked mannoses, that are known to be present in the cell wall mannoprotein core (25). Unambiguous identification of Peak H as the anomeric proton signal for a 3,6-di-O-substituted a-mannopyranosyl residue characterized the compounds as core-derived because man- nose in this linkage is found only in the core (6). The presence of a single /3-mannopyranosyl unit (Peak I) is also character- istic of the core structure. The integrations in Table V give an average chain length of 14, which agrees with the value of 15
TABLE IV
Analysis of mannoprotein preparations by PMR Ratio of
“II- Mall.
Mannoprotein branched nclses per Outer Carbohy- al + 6. POlY-
linked saccha- chain drate con-
mannose ride lengthh tent’
to core chain’ mannosese
Cell wall mnn2 mannan S75 invertase P75 invertase’ Endomannanase-di-
gested S75 invertase
R
7.6:1 137 122 d 2.2:1 50 35 ii (53) 1.1:1 33 18 47 (36) 0.3:1 20 5 -d
” From integration of the anomeric proton signals. * Assuming a core unit of 15 mannoses (25). ’ Calculated assuming 9 polysaccharide chains/invertase monomer
of 60,000 daltons (5). The values in parentheses were determined by direct analysis.
’ Not determined. ‘Approximately half of the P75 invertase was insoluble after
lyophilization. The PMR spectrum was done on the subfraction that was soluble and, we expect, had a higher than average carbohydrate content. This would explain the long outer chain length, obtained from the PMR data, and the high calculated carbohydrate content compared with the measured value.
II
II 0 ) I’ n ‘I r I v I ” 5.L) 5.2 5.0 L1.8 PPM
FIG. 10. Proton magnetic resonance spectra of the anomeric protons of the mnn2 mannoprotein core oligosaccharides. Top tracing, oligosaccharide from the bulk cell wall mannoprotein; and bottom tracing, oligosaccharide from the S75 invertase. The signals A to J were assigned as shown in Table V.
TABLE V
PMR assignments for core oligosaccharide anomeric protons Chemical
Peak shift Integra- (6 in tions” Proposed linkage assignments”
PPd
A 5.34 0.91 (1) +*aMan+”
B 5.29 1.03 (1) +‘eMan-t’ C 5.24 0.66 (1) +4aGlcNAc D 5.13’ 3.98 (4) aMan+“, +“cYMan-t“, +“cYMan+“,
t’
aMan+” t’
E 5.09” 0.86 (1) +“aMan+
t” aMan
F 5.04 3.42 (3) aMan-+‘, +“oiMan+” G 4.91’ 1.25 (1) +‘uMan+” aMan+” H 4.87 1.25 (1) +“&an+“’
t3
I 4.77 .29 (1) +?PMan+’ t?
J 4.71 -< +“fiGlcNAc
a Areas relative to the 3 N-acetyl protons (6 = 2.048) of N-acetyl- glucosamine, with nearest integral values in parentheses.
‘Determined by a comparison of standard oligosaccharides (R. Cohen and C. E. Ballou, manuscript in preparation).
‘This signal corresponds to mannose units of more than one linkage type, so the structural assignments are ambiguous.
’ The signal corresponds to the terminal mannose in 1 + 3-linkage. e Not determined.
Time FIG. 11. Fractionation of invertase core preparations by
high pressure liquid chromatography. A is the core fraction prepared from mnn2 S75 invertase by digestion with purified endo- mannanase and endoglucosaminidase, and B is a core reference ob- tained from mnn2 cell wall mannoprotein in a similar manner. The other patterns are for invertases digested only with the endoglucos- amidase. C is from the S75 invertase and is Fraction C from Fig. 5, D is from P75 invertase and corresponds to a preparation similar to Fraction C in Fig. 5, and E is from wild type baker’s yeast invertase and is Fraction C from Fig. 7. The 4 major peaks in B differ from each other by 1 mannose unit and range from 13 on the left to 16 on the right, each containing a single N-acetylglucosamine residue as well.
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from

12216 Carbohydrate Structure of Yeast Inuertase
to 16 obtained from chromatographic analysis of the same preparations.
Chromatographic Comparison of Invertase Core Preparations
The core preparation from cell wall mnn2 mannoprotein, obtained by sequential digestion with the endomannanase and an endoglucosaminidase, is heterogeneous and can be resolved by gel filtration into about 7 homologs (25). We have com- pared the high pressure liquid chromatography fractionation patterns of the cores from the various invertases (Fig. 11). It is apparent that the core units released by the endoglucosa- minidase H are heterogeneous and that the distribution of fragments in the different preparations is almost identical. A slight difference is noted in the core pattern of the reference mannoprotein that was digested with both the endomanna- nase and endoglucosaminidase. The result is consistent with a reduction in average size of the fragments by one or two hexoses that could have been due to the action of the endo- mannanase on terminal (~1 --+ &linked mannose units. The S75 invertase gave a similar pattern when so treated. The size distribution of the core oligosaccharides averages 15 mannoses and one N-acetylglucosamine, considerably smaller than the smallest fragments reported by Tarentino et al. (4), but iden- tical with those reported by Nakajima and Ballou (25) for the cell wall mannoprotein.
DISCUSSION
The bulk cell wall mannoprotein of S. cerevisiae possesses large polymannose chains linked through di-N-acetylchito- biose units to asparagine in the protein, for which an average, generalized structure has been established (1). The mnn2 mutant of S. cerevisiae makes an altered mannan that lacks the branched outer chain (8). Such a mannan can be degraded by the combined action of an endo-al + 6-mannanase and an endo-N-acetyl#-glucosaminidase (Fig. 12) to release the core fragment (6). The core preparation from bulk cell wall mannan is heterogeneous and is composed of oligosaccharides that contain 12 to 17 mannose units (25).
The glycosylated invertase isolated from the mnn2 mutant was separated into two fractions, one soluble in 75% ammo- nium sulfate (S75) and one precipitated by the salt (P75). The S75 invertase is similar to the bulk cell wall mnn2 mannopro- tein, whereas the P75 invertase appears to possess the core carbohydrate units with very little or none of the outer chain added. The endo-al -+ 6-mannanase converted the S75 in-
[M * M * H + M * Ml"*!4 * M-t M-t "* bGNA&NAc+kn SacchaFomyces
i : :, i cerevisiae mm2 mannoprotein
+ +
FIG. 12. Structural relationships between the polysaccha- ride chains of S. cerevisiae X2180 wild type mannoprotein, the mnn2 mutant, and the core oligosaccharide obtained from the latter by enzymic degradation. M, mannose.
vertase to material that was similar to the P75 fraction, but this enzyme had no observable effect on the latter or on baker’s yeast invertase.
Although the endo-N-acetyl+glucosaminidase H acted on both the S75 and P75 invertases, it did not release all of the carbohydrate. Instead, it produced a mixture of invertase molecules that appeared to differ by about 3,000 daltons, and ranged from 63,000 to 69,000 daltons. Since the carbohydrate- free invertase has an M, of about 60,000 (5), these products appear to contain variable numbers of carbohydrate chains with about 15 mannoses each, that is, approximately the size of the largest core unit (25). It is unlikely that these acrylamide gel bands result from proteolysis, because they traveled as a single band on isoelectric focusing. A similar spectrum of 3 or 4 bands was observed by Trimble and Maley (5) and, as they reported, we found that complete removal of the carbohydrate required further digestion of the carboxymethylated invertase.
The difference between the S75 and P75 invertases was confirmed by immunochemical analysis. The S75 invertase gave a precipitin line with anti-mnn2 mannan serum, in which the specificity is known to be directed against ~yl + 6-linked mannose units (8), whereas the P75 invertase failed to react with this serum. The P75 invertase reacted well with antise- rum to wild type mannan, however, which is specific for terminal al -+ 3-linked mannose (28). This linkage is known to be present in the core of the bulk cell wall mnn2 mannan (6). The S75 invertase also reacted with this antiserum, al- though the mnn2 mannoprotein did not, which indicates that the core fragments of this invertase are exposed to the anti- body. The failure of the bulk cell wall mnn2 mannoprotein to react with this serum probably means that its longer outer chains inhibit antibody approach to the core units or that there are few unsubstituted core units in the molecule.
The PMR spectra of the two invertase preparations were consistent with these conclusions. The spectrum of mnn2 mannoprotein shows a prominent anomeric proton signal (6 = 4.90) that is characteristic of an unsubstituted al -+ 6- linked hexose (29). Both S75 and P75 invertases show the same signal, although at a lower intensity, and the ratio of this signal to those for the other mannose anomeric protons was higher in the S75 invertase spectrum than in the P75 spectrum, as expected if it possessed a longer unsubstituted outer chain. Endo-cyl + 6-mannanase digestion of the S75 invertase almost eliminated the anomeric proton signal at 4.90 ppm, which shows that this enzyme is able to remove essen- tially all of the outer chains in the mnn2 invertase. The PMR spectrum of the invertase core fragments and their size distri- bution upon high pressure liquid chromatography confirmed their structural similarity to the core fragments isolated from the bulk cell wall mannoprotein (25).
External invertase from S. cerevisiae X2180, which is sim- ilar to baker’s yeast, is irnmunochemically identical to the bulk cell wall mannoprotein (15), and presumably it has a similar mannan structure. Tarentino et al. (4) showed that most of the carbohydrate could be released from baker’s yeast external invertase by the action of the endo-N-acetyl+glu- cosaminidase H, and the released carbohydrate could be sep- arated into two components with about 26 and 52 mannose units/N-acetylglucosamine. Trimble and Maley (5) concluded that the invertase molecule had, per subunit, about 9 such carbohydrate chains. We have obtained similar results, except we find that the small oligosaccharide fraction is very heter- ogeneous and that the molecules released contain less man- nose and approximate the size of the core fragments obtained from the mnn2 mannan. It is also notable that the endoglu- cosaminidase digest of wild type invertase shows, on acryl- amide gels in sodium dodecyl sulfate, a spectrum of 3 or 4
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from

Carbohydrate Structure of Yeast Invertase 12217
bands that appear to be identical to those obtained from the mnn2 invertases. This suggests that baker’s yeast external invertase also contains some carbohydrate chains consisting only of the core, whereas others have been enlarged by addi- tion of the outer chain.
The observations we report here have important implica- tions concerning the structure and biosynthesis of mannopro- teins and, perhaps, other glycoproteins. A general mechanism appears to be involved in the addition of the mannose-rich core unit to glycoproteins, in which the structure is assembled on a dolichyl pyrophosphate carrier (Dol-Pz-GlcNAczMan,) and then transferred as a unit to protein (11, 30). In mam- malian systems, this unit may contain 12 mannoses and 1 to 3 glucoses (31,32,33), and all of the glucose and several of the mannoses may be removed in the processing of the carbohy- drate unit to synthesize the more complex oligosaccharide chains that contain N-acetylglucosamine, galactose, and N- acetylneuraminic acid (34, 35, 36, 37). Yeasts have not been shown to contain such complex oligosaccharide chains, but they do modify the mannose-rich core by addition of the outer chain in a process that apparently involves the stepwise trans- fer of single mannose units from GDP-mannose (12). Our results suggest that the P75 invertase fraction may represent molecules to which the core fragments have been added, but on which the outer chains have not yet been attached or completed, whereas the S75 invertase fraction represents com- pleted molecules that have been exported from the cell.
A further implication of our results is that an outer chain is not attached uniformly to all core units. Several explanations of this observation are available. One possibility is that, after addition of the core, the protein chains fold in such a way that some of the core units are no longer accessible to the enzymes that build the outer chain. Another possibility is that the invertase molecules are processed at a fixed rate, and that the availability of mannosyl donor is limited such that all chains do not get extended. The fact that a yeast strain such as the mnn2 mutant appears to grow normally indicates that large variations in the extent of glycosylation of the mannoprotein outer chains can be tolerated by the organism.
The synthesis of invertase in S. cerevisiae is repressed when the yeast is grown on glucose, and many studies of this enzyme have employed the genetically derepressed FH4C strain (3). To facilitate use of the various mannan mutants in our collec- tion for studies of invertase synthesis, we have grown the cells on glucose until the hexose is depleted, then added sucrose and incubated the cultures until invertase formation reaches a maximum about 1 h later. This procedure worked satisfac- torily when carried out in a 200-liter fermenter, thus allowing a large-scale isolation of the enzyme. The invertase was pre- pared from broken cells, so the origin of the P75 fraction is uncertain. It is known that the majority of invertase activity in S. cereuisiae is external to the plasma membrane, so the S75 fraction is probably derived from the periplasm (2). Be- cause the mannosyltransferases are membrane-bound (12,38, 39), it is possible that the outer chains are added during vesicle or membrane association of the incomplete invertase mole- cules. If so, then we expect that the P75 invertase may be derived from a vesicular or membrane fraction of the cell. Recently, a membrane-associated form of invertase has been shown to possess the kinetic characteristics of a precursor of external invertase (40). It has a native molecular weight of 19C,OOO and could correspond to the P75 invertase described here.
In preliminary experiments not documented here, we have used immunoprecipitation with carbohydrate-specific antisera to isolate the S75 and P75 mannoproteins from the soluble and membrane fractions of extracts of whole cells that had
been incubated with [3H]mannose under conditions that al- lowed its incorporation into mannoproteins. Radioactivity precipitated by the al + 6-specific antiserum co-migrated with the S75 invertase on electrophoresis and was found only in the soluble fraction. The radioactivity precipitated by the al -+ 3-specific antiserum co-migrated with the P75 invertase and it was recovered from both the membrane and soluble fractions. From this we conclude that the mannoprotein with only core carbohydrate units is at least partly membrane- bound, whereas the mannoprotein with attached outer chains is all in a soluble form and may originate from the periplasm.
The separation of the P75 and S75 invertase fractions is due to their differing solubilities that depend on the differences in carbohydrate content. These differences may reflect the ex- tent of outer chain glycosylation, although we have not elim- inated the possibility that the invertases also differ in the number of core units. Perhaps both invertase fractions rep- resent a spectrum of molecules with microheterogeneity in their core and outer chain structures.
REFERENCES
1. Ballou, C. E. (1976) Adu. Microb. Physiol. 14, 93-158 2. Lampen, J. 0. (1968) Antonie uan Leeuwenhoeh 34, l-18 3. Gascon, S., Neumann, N. P., and Lampen, J. 0. (1968) J. Biol.
Chem. 243,1573-1577 4. Tarentino, A. L., Plummer, T. H., Jr., and MaIey, F. (1974) J.
Biol. Chem 249,818-824 5. Trimble, R. B., and Maley, F. (1977) J. BioZ. Chem. 252, 4409-
6.
7.
8.
9.
10. 11.
12.
13.
14.
15. 16.
17. 18.
19.
20.
21.
22. 23.
24. 25.
26. 27.
28. 29. 30.
31.
32.
4412 Nakajima, T., and Ballou, C. E. (1974) J. BioZ. Chem. 249, 7685-
7694 Kornfeld, R., and Kornfeld, S. (1976) Anna. Reu. Biochem. 45,
217-237 Raschke, W. C., Kern, K. A., Antalis, C., and BalIou, C. E. (1973)
J. Biol. Chem. 248,4660-4666 Nakajima, T., Maitra, S. K., and Ballou, C. E. (1976) J. BioZ.
Chem. 251,174-181 Lehle, L., and Tanner, W. (1978) Eur. J. Biochem. 83, 563-570 Lehle, L., and Tanner, W. (1978) Biochim. Biophys. Acta 539,
218-229 Nakajima, T., and Ballou, C. E. (1975) Proc. Natl. Acad. Sci. U.
S. A. 72,3912-3916 Nakajima, T., and Ballou, C. E. (1974) J. BioZ. Chem. 249, 7679-
7684 Neuman, N. P., and Lampen, J. 0. (1967) Biochemistry 6, 468-
475 Smith, W. L., and Ballou, C. E. (1974) Biochemistry 13, 355-361 Dubois, M., GiIIes, K. A., Hamilton, J. K., Rebers, P. A., and
Smith, F. (1966) Anal. Chem. 28, 350-356 Somogyi, M. (1952) J. Biol. Chem. 195,19-23 Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.
(1951) J. Biol. Chem. 193, 265-275 Kuhn, R. W., Walsh, K. A., and Neurath, H. (1974) Biochemistry
13,3871-3877 Tarentino, A. L., and Maley, F. (1974) J. Biol. Chem. 249, 811-
817 Kabat, E. A., and Mayer, M. (1961) Experimental Zmmunochem-
istry, 2nd Ed, C. C Thomas, Springfield, Ill. Laemmli, U. K. (1970) Nature 227,680-685 Fairbanks, G., Steck, T. L., and Wallach, D. F. H. (1971) Bio-
chemistry 10,2606-2617 O’Farrell, P. H. (1975) J. BioZ. Chem. 250,4007-4021 Nakajima, T., and BaIIou, C. E. (1975) Biochem. Biophys. Res.
Commun. 66,870-879 Lee, Y. C., and Ballou, C. E. (1965) Biochemistry 4,257-264 Spencer, J. F. T., and Gorin, P. A. J. (1968) J. Bacterial. 96, 180-
183 Ballou, C. E. (1970) J. BioZ. Chem. 245, 1197-1203 Van der Veen, J. M. (1963) J. Org. Chem. 28, 564-566 Waechter, C. J., and Lennarz, W. J. (1976) Annu. Rev. Biochem.
45,95-112 Behrens, N. H., Parodi, A. J., and Leloir, L. F. (1971) Proc. NatZ.
Acad. Sci. U. S. A. 68,2857-2860 Spiro, R. G., Spiro, M. J., and Bhoyroo, V. D. (1976) J. Biol.
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from

12218 Carbohydrate Structure of Yeast Invertase
Chem. 251,6409-6419 37. Chen, W. W., and Lennarz, W. J. (1978) J. Biol. Chem. 253,5780- 33. Li, E., Tabas, I., and Kornfeld, S. (1978) Fed. Proc. 37, 1299 5785 34. Robbins, P. W., Hubbard, S. C., Turco, S. J., and Wirth, D. F. 38. Lehle, L., and Tanner, W. (1974) Biochim. Biophys. Acta 350,
(1977) Cell 12,893~900 225-235 35. Tabas, I., Schlesinger, S., and Kornfeld, S. (1978) J. Biol. Chem. 39. Lehle, L., Bauer, F., and Tanner, W. (1977) Arch. Microbial. 14,
253, 716-722 77-81 36. Hunt, L. A., Etchison, J. R., and Summers, D. F. (1978) Proc. 40. Babczinski, P., and Tanner, W. (1978) Biochim. Biophys. Acta
Natl. Acad. Sci. U. S. A. 75, 754-758 538,426-434
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from

L Lehle, R E Cohen and C E Balloucore oligosaccharides and a form with completed polysaccharide chains.
Carbohydrate structure of yeast invertase. Demonstration of a form with only
1979, 254:12209-12218.J. Biol. Chem.
http://www.jbc.org/content/254/23/12209Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/254/23/12209.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on January 19, 2019http://w
ww
.jbc.org/D
ownloaded from



![WEVP 254 Bis - notices.sad-distribution.fr1].pdf · wevp 254 bis vpmn 254 bis + vppl a 254 bis ed. 06-08 v3 guide d’installation et d'utilisation handleiding voor installatie en](https://img.pdfslide.us/doc/110x75/5bda280709d3f2db058c1a59/wevp-254-bis-1pdf-wevp-254-bis-vpmn-254-bis-vppl-a-254-bis-ed-06-08-v3.jpg)

























