Embed Size (px)
Citation preview

Core Oligosaccharides of Plesiomonas shigelloides O54:H2(Strain CNCTC 113/92)STRUCTURAL AND SEROLOGICAL ANALYSIS OF THE LIPOPOLYSACCHARIDE CORE REGION,THE O-ANTIGEN BIOLOGICAL REPEATING UNIT, AND THE LINKAGE BETWEEN THEM*
Received for publication, December 13, 2001, and in revised form, January 11, 2002Published, JBC Papers in Press, February 16, 2002, DOI 10.1074/jbc.M111885200
Tomasz Niedziela‡, Jolanta Lukasiewicz§, Wojciech Jachymek‡§, Monika Dzieciatkowska§,Czeslaw Lugowski§, and Lennart Kenne‡¶
From the ‡Swedish University of Agricultural Sciences, Uppsala SE-75007, Sweden and the §L. Hirszfeld Institute ofImmunology and Experimental Therapy, Wroclaw PL-53114, Poland
The structure of the core oligosaccharide moiety of the lipopolysaccharide (LPS) of Plesiomonasshigelloides O54 (strain CNCTC 113/92) has been investigated by 1H and 13C NMR, fast atombombardment mass spectrometry (MS)/MS, matrix-assisted laser-desorption/ionization time-of-flight MS, monosaccharide and methylation analysis, and immunological methods. It was con-cluded that the main core oligosaccharide of this strain is composed of a decasaccharide with thefollowing structure:
L-�-D-Hepp �-D-Galp1 12 27 4
�-D-Galp-(134)-�-D-GlcpN-(134)-�-D-GalpA-(133)-L-�-D-Hepp-(133)-L-�-D-Hepp-(135)-Kdo6 21 11 1
�-D-Glcp �-D-Glcp
STRUCTURE 1
in which L-�-D-Hepp is L-glycero-�-D-manno-heptopyranose. The nonasaccharide variant of the coreoligosaccharide (�10%), devoid of �-D-Glcp substituting the �-D-GlcpN at C-6, was also identified.The core oligosaccharide substituted at C-4 of the outer core �-D-Glcp residue with the single O-polysaccharide repeating unit was also isolated yielding a hexadecasaccharide structure. The deter-mination of the monosaccharides involved in the linkage between the O-specific polysaccharide partand the core, as well as the presence of 33)-D-�-D-Hepp-(13 instead of 33,4)-D-�-D-Hepp-(13 in therepeating unit, revealed the structure of the biological repeating unit of the O-antigen. The coreoligosaccharides are not substituted by phosphate residues and represent novel core type of bacterialLPS that is characteristic for the Plesiomonas shigelloides serotype O54. Serological screening of 69different O-serotypes of P. shigelloides suggests that epitopes similar to the core oligosaccharide ofserotype O54 (strain CNCTC 113/92) might also be present in the core region of the serotypes O24(strain CNCTC 92/89), O37 (strain CNCTC 39/89) and O96 (strain CNCTC 5133) LPS.
Plesiomonas shigelloides is a Gram-negative, flagellated,rod-shaped bacterium. This ubiquitous and facultatively an-aerobic organism has been isolated from such sources as fresh-
water, surface water, and many wild and domestic animals.The infections correlate strongly with the surface water con-tamination and are particularly common in tropical and sub-tropical habitats (1).
Human infections with P. shigelloides are mostly related todrinking untreated water, eating uncooked shellfish (2, 3), andvisiting countries with low sanitary standards (4, 5). Recentstudies implicated P. shigelloides as an opportunistic pathogenin immunocompromised hosts (6) and especially neonates (6–10). However, it has also been associated with diarrheal illness(11) and other diseases in normal hosts. P. shigelloides hasbeen isolated from an assortment of clinical specimens, includ-ing cerebrospinal fluid, wounds, and respiratory tract. It causesgastrointestinal and localized infections originating from in-fected wounds, which can disseminate to other parts of thebody (12). The cases of meningitis and bacteremia (10) causedby P. shigelloides are of special interest due to theirseriousness.
* This work was supported by grants from the Swedish NaturalScience Research Council, the Swedish Council of Forestry and Agri-cultural Research and Grants 6-P04A-051-14 and 6-P04A-069-19 fromthe State Committee of Scientific Research, Poland. The collaborationbetween Polish and Swedish groups was supported by funds from TheRoyal Swedish Academy of Sciences and The Polish Academy of Sci-ences. The costs of publication of this article were defrayed in part bythe payment of page charges. This article must therefore be herebymarked “advertisement” in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
Part of this work was presented at 20th International CarbohydrateSymposium, August 27 through September 1, 2000, Hamburg, Germany.
¶ To whom correspondence should be addressed: Dept. of Chemistry,Swedish University of Agricultural Sciences, P. O. Box 7015, UppsalaSE-75007, Sweden. Tel.: 46-18-671-573; Fax: 46-18-673-477; E-mail:[email protected].
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 14, Issue of April 5, pp. 11653–11663, 2002© 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org 11653
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
P. shigelloides has been traditionally classified as a member ofthe Vibrionaceae family based on phenotypic characteristics suchas polar flagella, oxidase production, and fermentation properties
(1). However, phylogenetic analysis and assessment of the genusPlesiomonas deducted from small rRNA sequences indicate acloser relationship with members of Enterobacteriaceae (13).
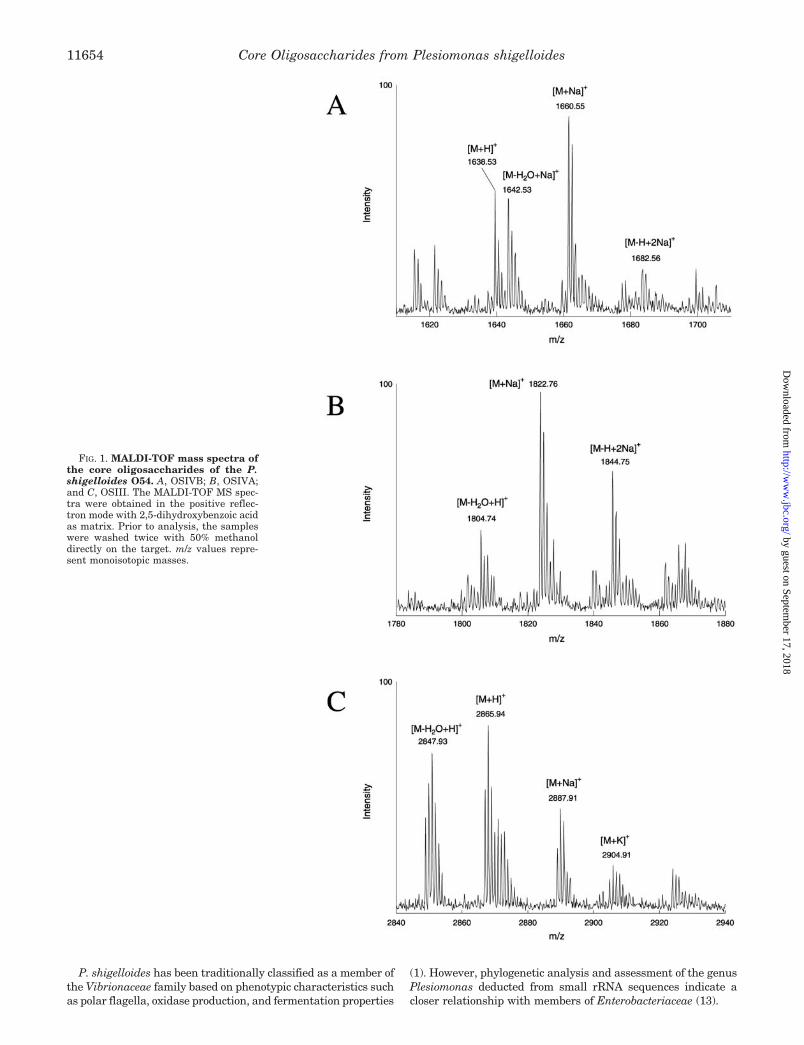
FIG. 1. MALDI-TOF mass spectra ofthe core oligosaccharides of the P.shigelloides O54. A, OSIVB; B, OSIVA;and C, OSIII. The MALDI-TOF MS spec-tra were obtained in the positive reflec-tron mode with 2,5-dihydroxybenzoic acidas matrix. Prior to analysis, the sampleswere washed twice with 50% methanoldirectly on the target. m/z values repre-sent monoisotopic masses.
Core Oligosaccharides from Plesiomonas shigelloides11654
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

The serotyping scheme of P. shigelloides was proposed byAldova, Shimada, and Sakazaki (14–19). Some O-antigenshave shown cross-reactivity with antisera directed against li-popolysaccharides (LPS)1 of Shigella sonnei, Shigella dysente-riae 1, 7 and 8, Shigella boydi 2, 9, and 13 and Shigella flexneri6 (15, 20). Two P. shigelloides strains were found to share thestructure with O-antigens of S. flexneri and S. dysenteriae (20,21). The unique structures of the O-specific polysaccharidesand core oligosaccharides remain unknown, except those ofO-specific polysaccharides from strains 22074, 12254 (21), andCNCTC 113/92 (22). The O-specific polysaccharide of strainCNCTC 113/92 LPS (serotype O54) is composed of a hexasac-charide repeating unit with the following structure:
The core oligosaccharide is important for biological and phys-ical properties of the overall lipopolysaccharide and plays asignificant role in interactions with the host. Thus we nowreport on structural and immunochemical studies of the coreoligosaccharides isolated from P. shigelloides strain CNCTC113/92 LPS.
EXPERIMENTAL PROCEDURES
Bacteria—Plesiomonas shigelloides strain CNCTC 113/92, classifiedas serovar O54:H2 according to Aldova’s antigenic scheme (14–17, 19)and 68 different P. shigelloides O-serotypes (O1, O2, O4–O6, O9, O11–O13, O15, O17, O19, O21, O22, O24–O28, O33–O46, O48, O50, O51,O56, O58, O59, O62, O64–O68, O70–O72, O74–O77, O81–O86, O91–O98), i.e. a group representative for all currently known serotypes, wereobtained from the Institute of Hygiene and Epidemiology, Prague,Czech Republic. The bacteria were grown and harvested as describedpreviously (22, 23).
Lipopolysaccharide and Core Oligosaccharides—LPS was extractedfrom bacterial cells by the hot phenol/water method (24) and purified asreported earlier (23). The yield of LPS was 2% of the dry bacterial mass.LPS (200 mg) was degraded by treatment with 1.5% acetic acid con-taining 2% SDS at 100 °C for 15 min. The reaction mixture was freeze-dried, the SDS removed by extraction with 96% ethanol, and the residuesuspended in water and centrifuged. The supernatant was fractionatedon Bio-Gel P-10, where O-specific polysaccharide separated fromshorter chains (OSIII, 33 mg) and core oligosaccharides (OSIV, 12 mg).The core oligosaccharides were further fractionated by chromatographyon Bio-Gel P-2 yielding two oligosaccharides: OSIVA (8.9 mg) andOSIVB (0.9 mg). The gel permeation chromatography was performed oncolumns (1.6 � 100 cm) of Bio-Gel P-10 and Bio-Gel P-2, equilibratedwith 0.05 M pyridine/acetic acid buffer, pH 5.6. Eluates were monitoredwith a Knauer differential refractometer and all fractions were checkedby 1H NMR spectroscopy and matrix-assisted laser-desorption/ioniza-tion time-of-flight (MALDI-TOF) mass spectrometry (MS) andfreeze-dried.
For a rapid screening of LPS from different P. shigelloides serotypesproteinase K-digested whole cell lysates were obtained by the methoddescribed earlier (25) with the following modifications. Bacteria weregrown on solid medium, harvested, and suspended in PBS to a turbiditygiving A600 nm � 0.6. A portion (1.5 ml) of the suspension was centri-fuged, and the pellet was resuspended in 200 �l of lysing buffer (0.05 M
Tris-HCl, pH 6.8, containing 4% SDS and 4% glycerol) and heated for 10min at 100 °C. Proteinase K (EC 3.4.21.64, Sigma-Aldrich) (�200 �g) inthe lysing buffer (80 �l) was added, followed by overnight incubation at21 °C. The digested bacterial lysate was boiled for 20 min prior toelectrophoresis.
Analytical Procedures—The LPS was analyzed by SDS-PAGE ac-cording to the method of Laemmli (26) with modifications as describedpreviously (27). The LPS bands were visualized by the silver stainingmethod (28). Sugars were analyzed as their alditol acetates by GC-MS
(23, 29). The absolute configurations of the sugars were determined asdescribed by Gerwig et al. (30, 31) using (�)-2-butanol for the formationof 2-butyl glycosides. The trimethylsilylated butyl glycosides were thenidentified by comparison with authentic samples (produced from re-spective sugar and (�)-2-butanol) on GC-MS. Carboxyl reduction of thenative oligosaccharide was carried out according to the method of Tay-lor et al. (32) as described previously (23). Methylations were performedboth on N-acetylated and carboxyl-reduced oligosaccharides and onlyN-acetylated oligosaccharides according to the method of Hakomori(33). The methyl ester groups of the latter methylated oligosaccharideswere reduced with Superdeuteride (LiB(C2H5)3
2H) as described by Bhatet al. (34). The methylated sugars were analyzed as partially methyl-ated alditol acetates by GC-MS as previously described (23). GC-MSwas carried out with a Hewlett-Packard 5971A system using an HP-1fused-silica capillary column (0.2 mm � 12 m) and a temperatureprogram 1503 270 °C at 8 °C min�1. Amino acid analysis was carriedout as described (35, 36). The core oligosaccharide (1 mg) was hydro-lyzed with 6 M hydrochloric acid at 100 °C for 24 h and concentrated todryness. Subsequently, n-butanol (0.5 ml) and acetyl chloride (50 �l)were added, and the reaction was carried out at 120 °C for 20 min,followed by the evaporation to dryness. Heptafluorobutyric anhydride(100 �l) was added, and the mixture was heated for 5 min at 150 °C.The N-heptafluorobutyryl n-butyl ester derivative of amino acid wasanalyzed by GC-MS on the same system as described above, but atemperature program 100 3 270 °C at 5 °C min�1.
N-Acetylation—Oligosaccharide OSIII (5 mg) was dissolved in satu-rated NaHCO3 (2 ml) at 0 °C and treated with acetic anhydride (3 � 100�l, with 10-min intervals). Reaction mixture was stored for additional30 min at 0 °C, the product purified on a column (1.6 � 100 cm) ofBio-Gel P-2 and the N-acetylated OSIII oligosaccharide examined byNMR spectroscopy and MALDI-TOF MS.
Mass Spectrometry—MALDI MS of the investigated oligosaccha-rides, in positive or negative mode, was run on a Bruker Reflex IIItime-of-flight instrument. Conjugates of core oligosaccharides with BSAwere analyzed using a Kratos Kompact-SEQ instrument. 2,5-Dihy-droxybenzoic acid and sinapinic acid were used as matrices for analysesof oligosaccharides and glycoconjugates, respectively.
NMR Spectroscopy—NMR spectra of the oligosaccharides were ob-tained for 2H2O solutions and H2O solutions containing 10% of 2H2O, at35 °C on Bruker DRX 400 and DRX 600 spectrometers. All spectra wereobtained using acetone (�H 2.225, �C 31.05) as internal reference. Thecore oligosaccharide fractions were repeatedly exchanged with 2H2Owith intermediate lyophilization. The data were acquired and processedusing standard Bruker software. The processed spectra were assignedwith the help of the SPARKY program (37). The signals were assignedby one- and two-dimensional experiments (COSY, clean-TOCSY,NOESY, ROESY, HMBC, HSQC-DEPT, and HSQC with and withoutcarbon decoupling). In the clean-TOCSY experiments the mixing times
1 The abbreviations used are: LPS, lipopolysaccharide; MALDI-TOF,matrix-assisted laser-desorption/ionization time-of-flight; MS, massspectrometry; PBS, phosphate buffered saline; GC, gas chromatogra-phy; COSY, correlated spectroscopy; TOCSY, total correlation spectros-copy; NOESY, nuclear Overhauser effect spectroscopy; ROESY, rotat-ing frame nuclear Overhauser effect spectroscopy; HMBC,heteronuclear multiple bond correlation; HMQC, heteronuclear multi-ple quantum coherence; HSQC, heteronuclear single quantum coher-ence; DEPT, distortionless enhancement by polarization transfer; LD-Hep, L-glycero-D-manno-heptose; Kdo, 3-deoxy-D-manno-oct-2-ulosonicacid; BSA, bovine serum albumin; OS, oligosaccharide; ELISA, enzyme-linked immunosorbent assay.
34)-D-�-D-Hepp-(133)-6d-�-D-Hepp-(134)-�-L-Rhap-(133)-�-D-GlcpNAc-(133 21 �1 OAc
�-L-Rhap411
�-D-Galf
STRUCTURE 2
Core Oligosaccharides from Plesiomonas shigelloides 11655
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

used were 30, 60, and 100 ms. The delay time in the HMBC was 60 ms andthe mixing times in the NOESY and ROESY experiments were 200 ms.
Preparation of Oligosaccharide Conjugates with BSA—The core oli-gosaccharide (OSIVA) was isolated and purified as described above. Theconjugation was carried out as described previously (38). Briefly, coreoligosaccharide OSIVA (2.5 mg) solutions in H2O (100 �l) was mixedwith an equal volume of BSA (1 mg) solution in H2O. Dimethylform-amide was added to a final concentration of 2%, and the mixture wasfreeze-dried. Dry preparation was heated at 110 °C for 30 min, dis-solved in PBS (1 ml), and dialyzed against PBS (3 � 1 liter). Theproducts were analyzed by MALDI-TOF MS, and their antigenic prop-erties were determined in the immunoblotting test, using polyclonalanti-P. shigelloides CNCTC 113/92 antibodies.
Immunization Procedures and Serological Methods—Rabbits wereimmunized with the P. shigelloides core oligosaccharide-BSA conjugate,suspended in a complete Freund adjuvant, and polyclonal antibodiesagainst the conjugates were obtained by the procedures previouslydescribed (39). Enzyme-linked immunosorbent assay (ELISA), usingLPS as solid-phase antigen, was performed by a modification (40) of themethod described by Voller et al. (41). Immunoblotting was done aspreviously described (23). A goat anti-rabbit IgG conjugated with alka-line phosphatase (Bio-Rad) was used as the second antibody and p-nitrophenyl phosphate and 5-bromo-4-chloro-3-indolyl phosphate/nitroblue tetrazolium were applied as detection systems for ELISA andimmunoblotting, respectively.
RESULTS
Isolation and Chemical Analysis of Core Oligosaccharides—The LPS of P. shigelloides CNCTC 113/92 was isolated byconventional methods and analyzed by SDS-PAGE, showingfractions consisting of core oligosaccharide substituted withdifferent numbers of oligosaccharide repeating units as well asunsubstituted core oligosaccharides. The O-specific polysaccha-ride and core oligosaccharides were liberated by mild acidichydrolysis of the LPS and isolated by gel filtration on Bio-GelP-10. In addition to the polysaccharide fraction, which wasanalyzed previously (22), two fractions with lower molecularmass components were obtained, i.e. OSIII (yield, 16.5% ofLPS) and OSIV (yield, 6% of LPS). The fraction OSIV wasfurther separated on Bio-Gel P-2 giving the two main oligosac-charides OSIVA (yield, 4.5% of LPS) and OSIVB (yield, 0.5% ofLPS). Because the initial NMR investigation indicated thepresence of uronic acid, Kdo, and one non-acetylated glucosa-mine residue in the oligosaccharides, all subsequent sugar andmethylation analyses were done on N-acetylated and carboxyl-reduced oligosaccharides to detect these residues. Compositionanalysis of the carboxyl-reduced and N-acetylated oligosaccha-ride OSIVB together with determination of the absolute con-figuration revealed the presence of LD-Hep, D-Glc, D-Gal, andD-GlcN (relative proportions of 2.7:1.8:2.9:0.8) in the carboxyl-reduced OSIVB oligosaccharide. Methylation analysis was per-formed on this carboxyl-reduced and N-acetylated OSIVB butalso on only N-acetylated OSIVB. The methyl esters of thelatter methylated material were reduced with Superdeuteridegenerating two deuterium on C-6 of the former uronic acid.These analyses showed the presence of 2,3,7-trisubstituted LD-Hepp, 3,4-disubstituted LD-Hepp, terminal LD-Hepp, 4-substi-tuted D-GlcpN, terminal D-Glcp, terminal D-Galp, 4-substitutedD-GalpA, and 5-substituted Kdo (relative proportions 0.8:1.1:1.0:0.7:0.9:1.8:0.6:0.7) in the original core oligosaccharideOSIVB.
In oligosaccharide OSIVA the ratio of terminal D-Glcp wastwice as high as in OSIVB and 4,6-disubstituted D-GlcpN wasidentified instead of 4-substituted D-GlcpN. All other compo-nents and ratios were found to be the same as in OSIVB. Thesubstitution positions and the ring forms were supported byNMR data (see below).
The MALDI-TOF mass spectra of the oligosaccharides (Fig.1, A and B) showed main ions at m/z 1660.55 [M�Na]�,1638.53 [M�H]�, and 1642.53 [M-H2O�Na]� for OSIVB and
m/z 1822.76 [M�Na]�, 1844.75 [M-H�2Na]�, and 1804.74[M-H2O�Na]� for OSIVA. This suggests a nonasaccharide inOSIVB and a decasaccharide in OSIVA differing only in onehexose unit (162.21Da difference). The nine sugars, two Gal,one Glc, three Hep, one GalA, one GlcN, and one Kdo, givetogether a monoisotopic mass of 1637.52 and an average massof 1638.41. The mass spectrum of the isolated OSIII (Fig. 1C)component showed main ions at m/z 2865.94 [M�H]�, 2887.91[M�Na]�, and 2847.93 [M-H2O�H]�. The mass difference of1065.15 between OSIII and OSIVA can be explained by onerepeating unit of the O-specific polysaccharide substitutingthe core. The mass of OSIII thus supports a hexadecasaccha-ride structure with one repeating unit linked to the coreoligosaccharide.
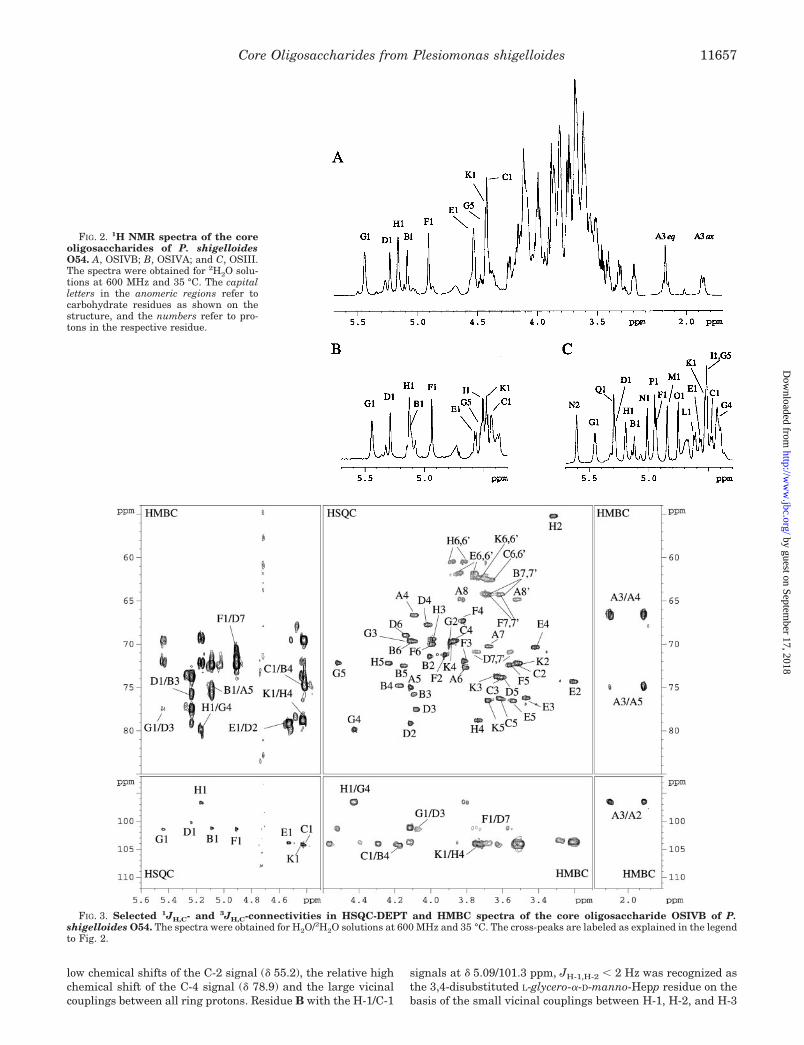
NMR Analysis of the Core Oligosaccharide OSIVB andOSIVA—The 1H (Fig. 2A) and HSQC-DEPT (Fig. 3) NMR spec-tra of the core oligosaccharide OSIVB contained main signalsfor eight anomeric protons and carbons, and in addition a Kdospin system confirming a nonasaccharide (the sugar residuesare indicated by capital letters as shown in the structure below,and these letters refer to the corresponding sugars through theentire text, tables, and figures). The 1H (Fig. 2B) and HSQC-DEPT NMR spectra of the core oligosaccharide OSIVA con-tained main signals for nine anomeric protons and carbons anda Kdo spin system, thus confirming a decasaccharide structure.Because all the 1H NMR spectra were complex and containedoverlapping signals, the major signals and spin systems wereassigned by COSY, TOCSY with different mixing times, andHSQC experiments. By comparing the chemical shifts withpreviously published NMR data for respective monosaccha-rides (42–44) and considering the 3JH, H values for the couplingbetween ring protons, estimated from the cross-peaks in thetwo-dimensional spectra, the sugars could be identified andtheir anomeric configuration determined.
Starting with the signal for the anomeric proton, H-1, theCOSY spectrum identified the H-2 signal and the TOCSY spec-tra with different mixing times the H-3 to H-7 signals. The H-7signals of heptose residues were identified in the TOCSY ex-periments starting with the assigned H-3 and H-4 signals. TheHSQC-TOCSY experiments were used for unambiguous as-signment of overlapping signals. From the assigned 1H signalsand the one-bond C–H connectivities, the carbon signals wereassigned in the gradient-enhanced HSQC-DEPT spectrum(Fig. 3), and the linkage positions were determined from thehigh chemical shifts of the signals from the substituted car-bons. The CH2 carbon signals were readily identified in theHSQC-DEPT experiment from negative cross-peaks. An un-equivocal identification of the H-7, C-7 as a negative cross-peakin the HSQC-DEPT experiment was further confirmed in theHSQC-TOCSY experiment. By these procedures all the spinsystems comprising 1H and 13C resonances were determined(Table I).
Residue G with the H-1/C-1 signals at � 5.44/101.5 ppm andnon-resolved JH-1,H-2 coupling, was assigned as the 4-substi-tuted �-D-GalpA residue based on the characteristic five protonspin system, the high chemical shifts of the H-5 (� 4.52), H-4 (�4.43), H-3 (� 4.12), and C-4 (� 79.9) signals, the large vicinalcouplings between H-2 and H-3 and small vicinal couplingbetween H-3, H-4, and H-5. Residue D with the H-1/C-1 signalsat � 5.23/100.3 ppm, JH-1,H-2 � 2 Hz was recognized as the2,3,7-substituted L-glycero-�-D-manno-Hepp residue from the1H and 13C chemical shifts, small vicinal couplings betweenH-1, H-2, and H-3, and the relatively high chemical shifts of theC-2 (� 79.2), C-3 (� 77.4), and C-7 (� 70.9) signals. Residue Hwith the H-1/C-1 signals at � 5.17/96.8 ppm, JH-1,H-2 3.6 Hz wasassigned as the 4-substituted �-D-GlcpN residue based on the
Core Oligosaccharides from Plesiomonas shigelloides11656
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
low chemical shifts of the C-2 signal (� 55.2), the relative highchemical shift of the C-4 signal (� 78.9) and the large vicinalcouplings between all ring protons. Residue B with the H-1/C-1
signals at � 5.09/101.3 ppm, JH-1,H-2 � 2 Hz was recognized asthe 3,4-disubstituted L-glycero-�-D-manno-Hepp residue on thebasis of the small vicinal couplings between H-1, H-2, and H-3
FIG. 2. 1H NMR spectra of the coreoligosaccharides of P. shigelloidesO54. A, OSIVB; B, OSIVA; and C, OSIII.The spectra were obtained for 2H2O solu-tions at 600 MHz and 35 °C. The capitalletters in the anomeric regions refer tocarbohydrate residues as shown on thestructure, and the numbers refer to pro-tons in the respective residue.
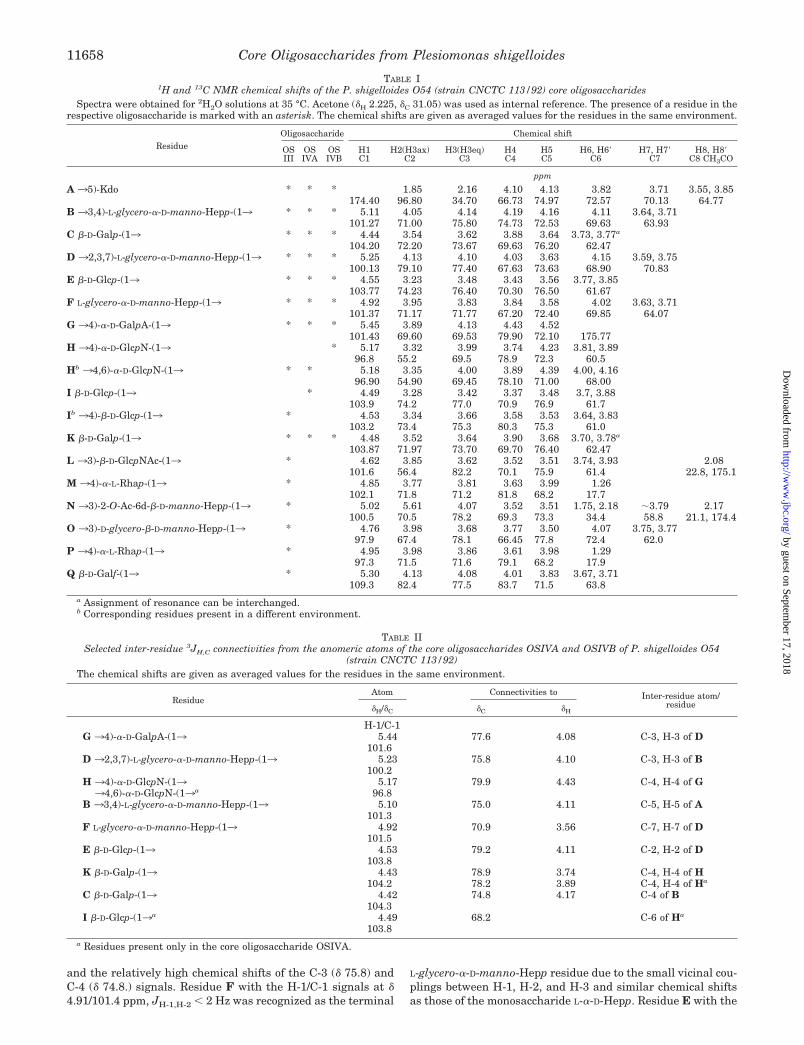
FIG. 3. Selected 1JH,C- and 3JH,C-connectivities in HSQC-DEPT and HMBC spectra of the core oligosaccharide OSIVB of P.shigelloides O54. The spectra were obtained for H2O/2H2O solutions at 600 MHz and 35 °C. The cross-peaks are labeled as explained in the legendto Fig. 2.
Core Oligosaccharides from Plesiomonas shigelloides 11657
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
and the relatively high chemical shifts of the C-3 (� 75.8) andC-4 (� 74.8.) signals. Residue F with the H-1/C-1 signals at �4.91/101.4 ppm, JH-1,H-2 � 2 Hz was recognized as the terminal
L-glycero-�-D-manno-Hepp residue due to the small vicinal cou-plings between H-1, H-2, and H-3 and similar chemical shiftsas those of the monosaccharide L-�-D-Hepp. Residue E with the
TABLE I1H and 13C NMR chemical shifts of the P. shigelloides O54 (strain CNCTC 113/92) core oligosaccharides
Spectra were obtained for 2H2O solutions at 35 °C. Acetone (�H 2.225, �C 31.05) was used as internal reference. The presence of a residue in therespective oligosaccharide is marked with an asterisk. The chemical shifts are given as averaged values for the residues in the same environment.
ResidueOligosaccharide Chemical shift
OSIII
OSIVA
OSIVB
H1C1
H2(H3ax)C2
H3(H3eq)C3
H4C4
H5C5
H6, H6�C6
H7, H7�C7
H8, H8�C8 CH3CO
ppm
A 35)-Kdo * * * 1.85 2.16 4.10 4.13 3.82 3.71 3.55, 3.85174.40 96.80 34.70 66.73 74.97 72.57 70.13 64.77
B 33,4)-L-glycero-�-D-manno-Hepp-(13 * * * 5.11 4.05 4.14 4.19 4.16 4.11 3.64, 3.71101.27 71.00 75.80 74.73 72.53 69.63 63.93
C �-D-Galp-(13 * * * 4.44 3.54 3.62 3.88 3.64 3.73, 3.77a
104.20 72.20 73.67 69.63 76.20 62.47D 32,3,7)-L-glycero-�-D-manno-Hepp-(13 * * * 5.25 4.13 4.10 4.03 3.63 4.15 3.59, 3.75
100.13 79.10 77.40 67.63 73.63 68.90 70.83E �-D-Glcp-(13 * * * 4.55 3.23 3.48 3.43 3.56 3.77, 3.85
103.77 74.23 76.40 70.30 76.50 61.67F L-glycero-�-D-manno-Hepp-(13 * * * 4.92 3.95 3.83 3.84 3.58 4.02 3.63, 3.71
101.37 71.17 71.77 67.20 72.40 69.85 64.07G 34)-�-D-GalpA-(13 * * * 5.45 3.89 4.13 4.43 4.52
101.43 69.60 69.53 79.90 72.10 175.77H 34)-�-D-GlcpN-(13 * 5.17 3.32 3.99 3.74 4.23 3.81, 3.89
96.8 55.2 69.5 78.9 72.3 60.5Hb 34,6)-�-D-GlcpN-(13 * * 5.18 3.35 4.00 3.89 4.39 4.00, 4.16
96.90 54.90 69.45 78.10 71.00 68.00I �-D-Glcp-(13 * 4.49 3.28 3.42 3.37 3.48 3.7, 3.88
103.9 74.2 77.0 70.9 76.9 61.7Ib 34)-�-D-Glcp-(13 * 4.53 3.34 3.66 3.58 3.53 3.64, 3.83
103.2 73.4 75.3 80.3 75.3 61.0K �-D-Galp-(13 * * * 4.48 3.52 3.64 3.90 3.68 3.70, 3.78a
103.87 71.97 73.70 69.70 76.40 62.47L 33)-�-D-GlcpNAc-(13 * 4.62 3.85 3.62 3.52 3.51 3.74, 3.93 2.08
101.6 56.4 82.2 70.1 75.9 61.4 22.8, 175.1M 34)-�-L-Rhap-(13 * 4.85 3.77 3.81 3.63 3.99 1.26
102.1 71.8 71.2 81.8 68.2 17.7N 33)-2-O-Ac-6d-�-D-manno-Hepp-(13 * 5.02 5.61 4.07 3.52 3.51 1.75, 2.18 �3.79 2.17
100.5 70.5 78.2 69.3 73.3 34.4 58.8 21.1, 174.4O 33)-D-glycero-�-D-manno-Hepp-(13 * 4.76 3.98 3.68 3.77 3.50 4.07 3.75, 3.77
97.9 67.4 78.1 66.45 77.8 72.4 62.0P 34)-�-L-Rhap-(13 * 4.95 3.98 3.86 3.61 3.98 1.29
97.3 71.5 71.6 79.1 68.2 17.9Q �-D-Galf-(13 * 5.30 4.13 4.08 4.01 3.83 3.67, 3.71
109.3 82.4 77.5 83.7 71.5 63.8a Assignment of resonance can be interchanged.b Corresponding residues present in a different environment.
TABLE IISelected inter-residue 3JH,C connectivities from the anomeric atoms of the core oligosaccharides OSIVA and OSIVB of P. shigelloides O54
(strain CNCTC 113/92)The chemical shifts are given as averaged values for the residues in the same environment.
ResidueAtom Connectivities to Inter-residue atom/
residue�H/�C �C �H
H-1/C-1G 34)-�-D-GalpA-(13 5.44 77.6 4.08 C-3, H-3 of D
101.6D 32,3,7)-L-glycero-�-D-manno-Hepp-(13 5.23 75.8 4.10 C-3, H-3 of B
100.2H 34)-�-D-GlcpN-(13 5.17 79.9 4.43 C-4, H-4 of G34,6)-�-D-GlcpN-(13a 96.8
B 33,4)-L-glycero-�-D-manno-Hepp-(13 5.10 75.0 4.11 C-5, H-5 of A101.3
F L-glycero-�-D-manno-Hepp-(13 4.92 70.9 3.56 C-7, H-7 of D101.5
E �-D-Glcp-(13 4.53 79.2 4.11 C-2, H-2 of D103.8
K �-D-Galp-(13 4.43 78.9 3.74 C-4, H-4 of H104.2 78.2 3.89 C-4, H-4 of Ha
C �-D-Galp-(13 4.42 74.8 4.17 C-4 of B104.3
I �-D-Glcp-(13a 4.49 68.2 C-6 of Ha
103.8a Residues present only in the core oligosaccharide OSIVA.
Core Oligosaccharides from Plesiomonas shigelloides11658
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
H-1/C-1 signals at � 4.53/103.9 ppm, JH-1,H-2 7.8 Hz was recog-nized as terminal �-D-Glcp from the similarity of the 1H and13C chemical shifts with those of �-D-Glcp and the large vicinalcouplings between all protons in the sugar ring. Residue K withthe H-1/C-1 signals at � 4.43/104.1 ppm, JH-1,H-2 7.8 Hz as wellas residue C with the H-1/C-1 signals at � 4.42/104.3 ppm,JH-1,H-2 7.8 Hz were assigned as terminal �-D-Galp residuesdue to the large coupling between H-1, H-2, and H-3 and thesmall vicinal coupling between H-3, H-4, and H-5, and chemicalshifts similar to those of �-D-Galp. Residue A was identified asa 5-substituted Kdo on the basis of characteristic deoxy protonsignals, found at � 1.86 ppm (H-3ax) and � 2.16 ppm (H-3eq),and a high chemical shift of the C-5 signal (� 75.0 ppm). InOSIVA an additional terminal �-D-Glcp (residue I), � 4.49/103.9ppm, JH-1,H-2 7.8 Hz, was found and residue H with the H-1/C-1signals at � 5.16/97.0 ppm, JH-1,H-2 3.6 Hz was found to beadditionally substituted at C-6 due to characteristic downfieldshift of the C-6 signal (� 68.2 ppm). The 1JC-1,H-1 values ob-tained from an HMQC experiment, confirmed the �-pyranosylconfiguration for residues B (173 Hz), D (175 Hz), F (173 Hz),
G (173 Hz), and H (174 Hz) and �-pyranosyl configuration forresidues C, E, and K (163 Hz for all these residues). The resultsare in agreement with data from the sugar and methylationanalyses.
In some of the batches of core oligosaccharides a glycine wasidentified, by the presence of an additional carbonyl signal at �169.2 ppm, and a negative CH2 signal (H� � 3.96 ppm, C� � 41.6ppm) in the HSQC-DEPT spectrum. The presence of glycine insome of the preparations of the core oligosaccharides was con-firmed by amino acid analysis and mass spectrometry. How-ever, only MS data (data not shown) suggested that the glycinewas linked to the isolated core oligosaccharides.
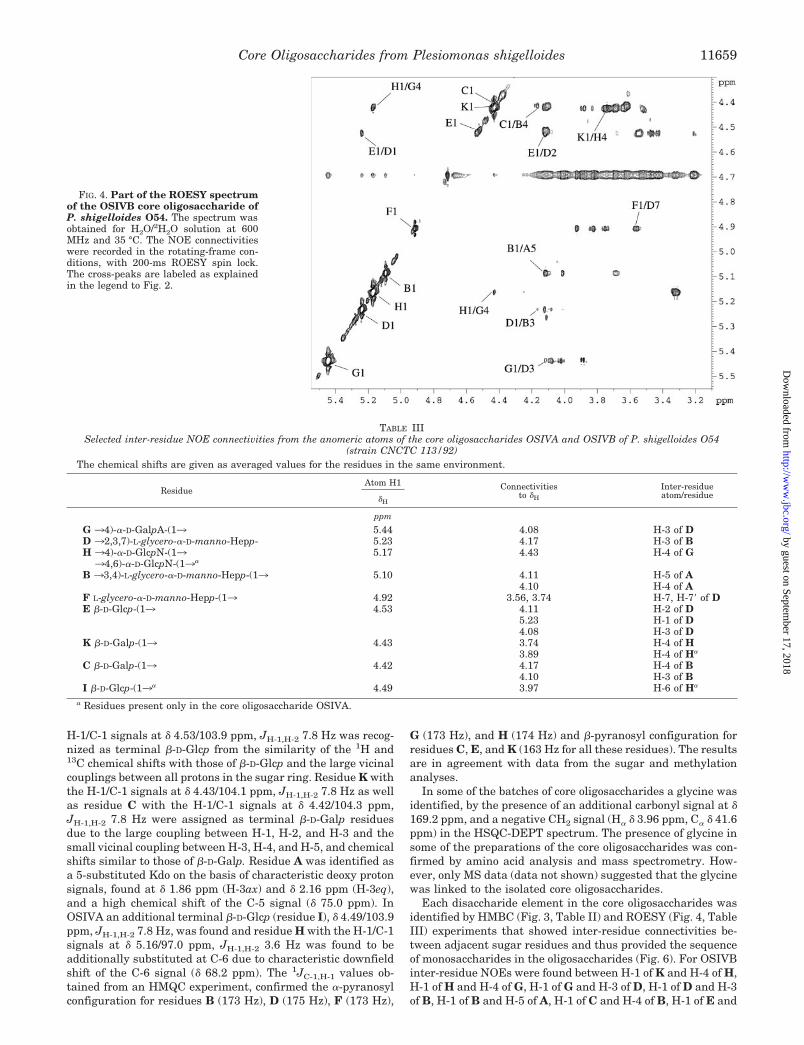
Each disaccharide element in the core oligosaccharides wasidentified by HMBC (Fig. 3, Table II) and ROESY (Fig. 4, TableIII) experiments that showed inter-residue connectivities be-tween adjacent sugar residues and thus provided the sequenceof monosaccharides in the oligosaccharides (Fig. 6). For OSIVBinter-residue NOEs were found between H-1 of K and H-4 of H,H-1 of H and H-4 of G, H-1 of G and H-3 of D, H-1 of D and H-3of B, H-1 of B and H-5 of A, H-1 of C and H-4 of B, H-1 of E and
FIG. 4. Part of the ROESY spectrumof the OSIVB core oligosaccharide ofP. shigelloides O54. The spectrum wasobtained for H2O/2H2O solution at 600MHz and 35 °C. The NOE connectivitieswere recorded in the rotating-frame con-ditions, with 200-ms ROESY spin lock.The cross-peaks are labeled as explainedin the legend to Fig. 2.
TABLE IIISelected inter-residue NOE connectivities from the anomeric atoms of the core oligosaccharides OSIVA and OSIVB of P. shigelloides O54
(strain CNCTC 113/92)The chemical shifts are given as averaged values for the residues in the same environment.
ResidueAtom H1 Connectivities
to �H
Inter-residueatom/residue�H
ppm
G 34)-�-D-GalpA-(13 5.44 4.08 H-3 of DD 32,3,7)-L-glycero-�-D-manno-Hepp- 5.23 4.17 H-3 of BH 34)-�-D-GlcpN-(13 5.17 4.43 H-4 of G34,6)-�-D-GlcpN-(13a
B 33,4)-L-glycero-�-D-manno-Hepp-(13 5.10 4.11 H-5 of A4.10 H-4 of A
F L-glycero-�-D-manno-Hepp-(13 4.92 3.56, 3.74 H-7, H-7� of DE �-D-Glcp-(13 4.53 4.11 H-2 of D
5.23 H-1 of D4.08 H-3 of D
K �-D-Galp-(13 4.43 3.74 H-4 of H3.89 H-4 of Ha
C �-D-Galp-(13 4.42 4.17 H-4 of B4.10 H-3 of B
I �-D-Glcp-(13a 4.49 3.97 H-6 of Ha
a Residues present only in the core oligosaccharide OSIVA.
Core Oligosaccharides from Plesiomonas shigelloides 11659
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
H-2 of D, and H-1 of F and H-7,7� of D. In OSIVA connectivitiesbetween H-1 of the additional glucose (residue I) and H-6 of Hwere established in NOESY experiment.
The HMBC spectra showed cross-peaks between the ano-meric proton and the carbon at the linkage position and be-tween the anomeric carbon and the proton at the linkage posi-tion (Table II), which confirmed the structure of the core nona-and decasaccharide in the LPS of P. shigelloides strain CNCTC113/92 (Fig. 6).
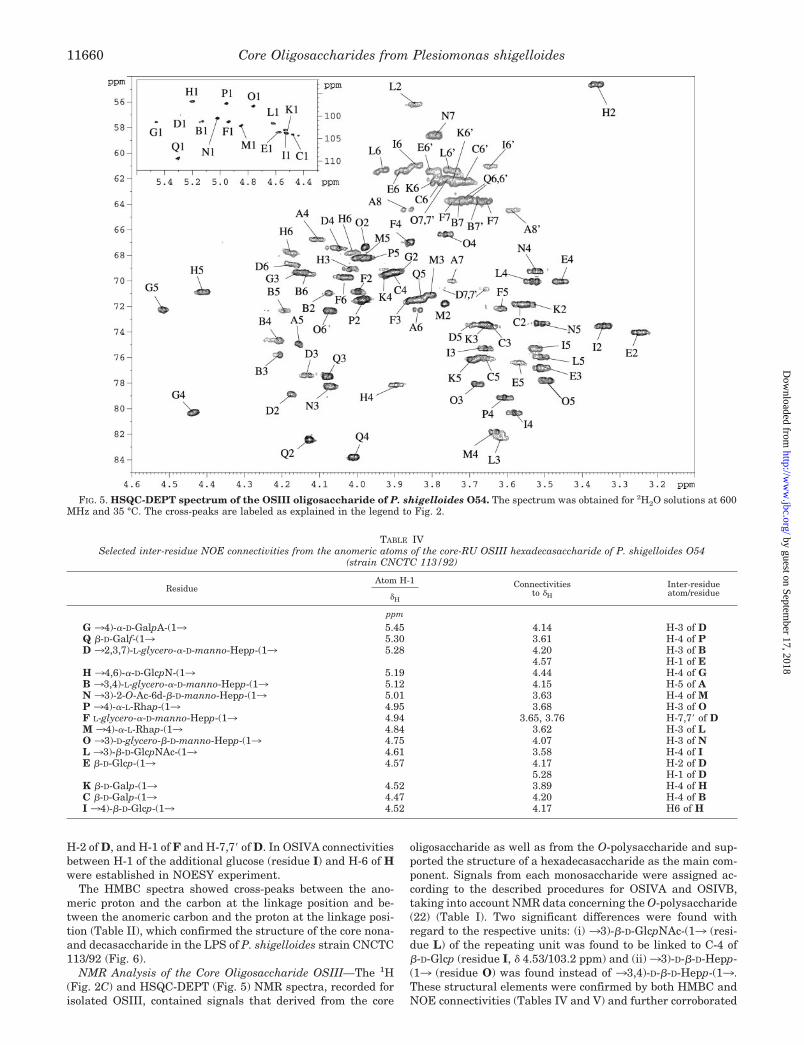
NMR Analysis of the Core Oligosaccharide OSIII—The 1H(Fig. 2C) and HSQC-DEPT (Fig. 5) NMR spectra, recorded forisolated OSIII, contained signals that derived from the core
oligosaccharide as well as from the O-polysaccharide and sup-ported the structure of a hexadecasaccharide as the main com-ponent. Signals from each monosaccharide were assigned ac-cording to the described procedures for OSIVA and OSIVB,taking into account NMR data concerning the O-polysaccharide(22) (Table I). Two significant differences were found withregard to the respective units: (i) 33)-�-D-GlcpNAc-(13 (resi-due L) of the repeating unit was found to be linked to C-4 of�-D-Glcp (residue I, � 4.53/103.2 ppm) and (ii)33)-D-�-D-Hepp-(13 (residue O) was found instead of 33,4)-D-�-D-Hepp-(13.These structural elements were confirmed by both HMBC andNOE connectivities (Tables IV and V) and further corroborated
FIG. 5. HSQC-DEPT spectrum of the OSIII oligosaccharide of P. shigelloides O54. The spectrum was obtained for 2H2O solutions at 600MHz and 35 °C. The cross-peaks are labeled as explained in the legend to Fig. 2.
TABLE IVSelected inter-residue NOE connectivities from the anomeric atoms of the core-RU OSIII hexadecasaccharide of P. shigelloides O54
(strain CNCTC 113/92)
ResidueAtom H-1 Connectivities
to �H
Inter-residueatom/residue�H
ppm
G 34)-�-D-GalpA-(13 5.45 4.14 H-3 of DQ �-D-Galf-(13 5.30 3.61 H-4 of PD 32,3,7)-L-glycero-�-D-manno-Hepp-(13 5.28 4.20 H-3 of B
4.57 H-1 of EH 34,6)-�-D-GlcpN-(13 5.19 4.44 H-4 of GB 33,4)-L-glycero-�-D-manno-Hepp-(13 5.12 4.15 H-5 of AN 33)-2-O-Ac-6d-�-D-manno-Hepp-(13 5.01 3.63 H-4 of MP 34)-�-L-Rhap-(13 4.95 3.68 H-3 of OF L-glycero-�-D-manno-Hepp-(13 4.94 3.65, 3.76 H-7,7� of DM 34)-�-L-Rhap-(13 4.84 3.62 H-3 of LO 33)-D-glycero-�-D-manno-Hepp-(13 4.75 4.07 H-3 of NL 33)-�-D-GlcpNAc-(13 4.61 3.58 H-4 of IE �-D-Glcp-(13 4.57 4.17 H-2 of D
5.28 H-1 of DK �-D-Galp-(13 4.52 3.89 H-4 of HC �-D-Galp-(13 4.47 4.20 H-4 of BI 34)-�-D-Glcp-(13 4.52 4.17 H6 of H
Core Oligosaccharides from Plesiomonas shigelloides11660
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
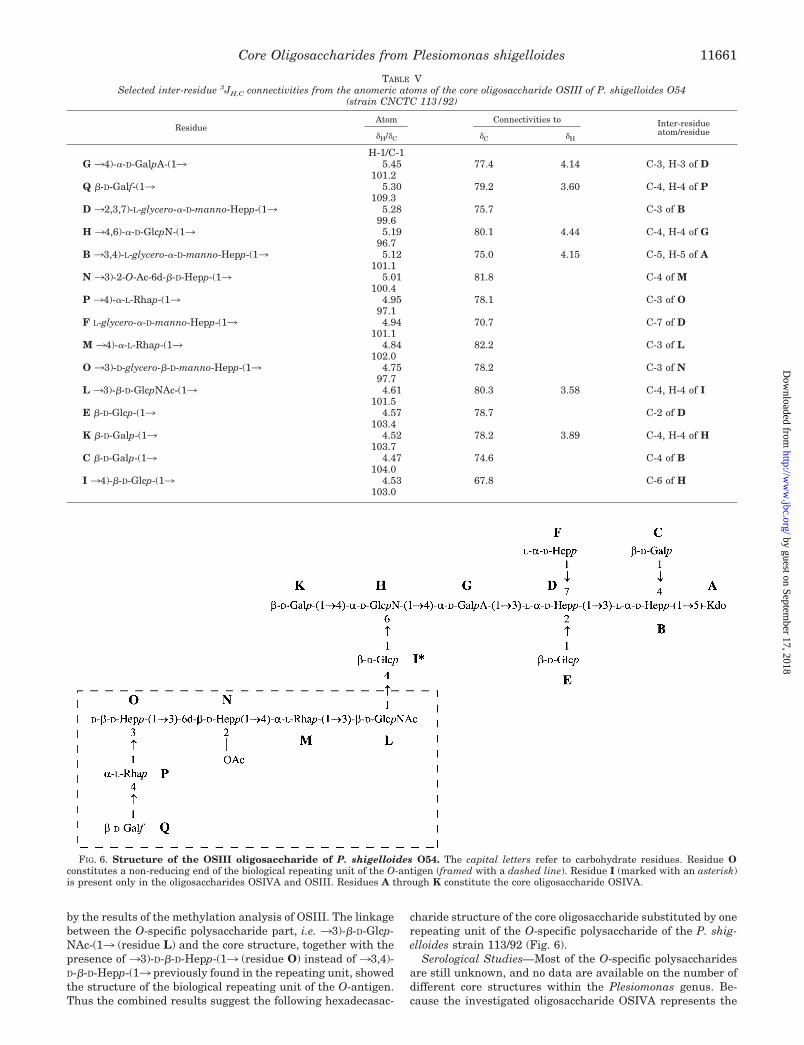
by the results of the methylation analysis of OSIII. The linkagebetween the O-specific polysaccharide part, i.e. 33)-�-D-Glcp-NAc-(13 (residue L) and the core structure, together with thepresence of 33)-D-�-D-Hepp-(13 (residue O) instead of 33,4)-D-�-D-Hepp-(13 previously found in the repeating unit, showedthe structure of the biological repeating unit of the O-antigen.Thus the combined results suggest the following hexadecasac-
charide structure of the core oligosaccharide substituted by onerepeating unit of the O-specific polysaccharide of the P. shig-elloides strain 113/92 (Fig. 6).
Serological Studies—Most of the O-specific polysaccharidesare still unknown, and no data are available on the number ofdifferent core structures within the Plesiomonas genus. Be-cause the investigated oligosaccharide OSIVA represents the
FIG. 6. Structure of the OSIII oligosaccharide of P. shigelloides O54. The capital letters refer to carbohydrate residues. Residue Oconstitutes a non-reducing end of the biological repeating unit of the O-antigen (framed with a dashed line). Residue I (marked with an asterisk)is present only in the oligosaccharides OSIVA and OSIII. Residues A through K constitute the core oligosaccharide OSIVA.
TABLE VSelected inter-residue 3JH,C connectivities from the anomeric atoms of the core oligosaccharide OSIII of P. shigelloides O54
(strain CNCTC 113/92)
ResidueAtom Connectivities to Inter-residue
atom/residue�H/�C �C �H
H-1/C-1G 34)-�-D-GalpA-(13 5.45 77.4 4.14 C-3, H-3 of D
101.2Q �-D-Galf-(13 5.30 79.2 3.60 C-4, H-4 of P
109.3D 32,3,7)-L-glycero-�-D-manno-Hepp-(13 5.28 75.7 C-3 of B
99.6H 34,6)-�-D-GlcpN-(13 5.19 80.1 4.44 C-4, H-4 of G
96.7B 33,4)-L-glycero-�-D-manno-Hepp-(13 5.12 75.0 4.15 C-5, H-5 of A
101.1N 33)-2-O-Ac-6d-�-D-Hepp-(13 5.01 81.8 C-4 of M
100.4P 34)-�-L-Rhap-(13 4.95 78.1 C-3 of O
97.1F L-glycero-�-D-manno-Hepp-(13 4.94 70.7 C-7 of D
101.1M 34)-�-L-Rhap-(13 4.84 82.2 C-3 of L
102.0O 33)-D-glycero-�-D-manno-Hepp-(13 4.75 78.2 C-3 of N
97.7L 33)-�-D-GlcpNAc-(13 4.61 80.3 3.58 C-4, H-4 of I
101.5E �-D-Glcp-(13 4.57 78.7 C-2 of D
103.4K �-D-Galp-(13 4.52 78.2 3.89 C-4, H-4 of H
103.7C �-D-Galp-(13 4.47 74.6 C-4 of B
104.0I 34)-�-D-Glcp-(13 4.53 67.8 C-6 of H
103.0
Core Oligosaccharides from Plesiomonas shigelloides 11661
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from
first complete structure of a Plesiomonas shigelloides core,anti-OSIVA-BSA polyclonal antibodies were used to scan byimmunoblotting all the currently available P. shigelloidesstrains comprising 69 different O-serotypes for the presence ofepitopes similar to those found in the P. shigelloides O54(strain CNCTC 113/92).
The OSIVA oligosaccharide, obtained by mild acidic hydrol-ysis and separation by size exclusion chromatography, waslinked covalently to BSA (38) and the reaction product waschecked by MALDI-TOF MS. The MALDI-TOF mass spectrumof the OSIVA conjugated to BSA showed main ion at m/z68,300. The reference spectrum of BSA gave the main ion atm/z 66,510. The mass difference between BSA and the BSAconjugate suggests that mainly one oligosaccharide moleculewas conjugated to one BSA molecule.
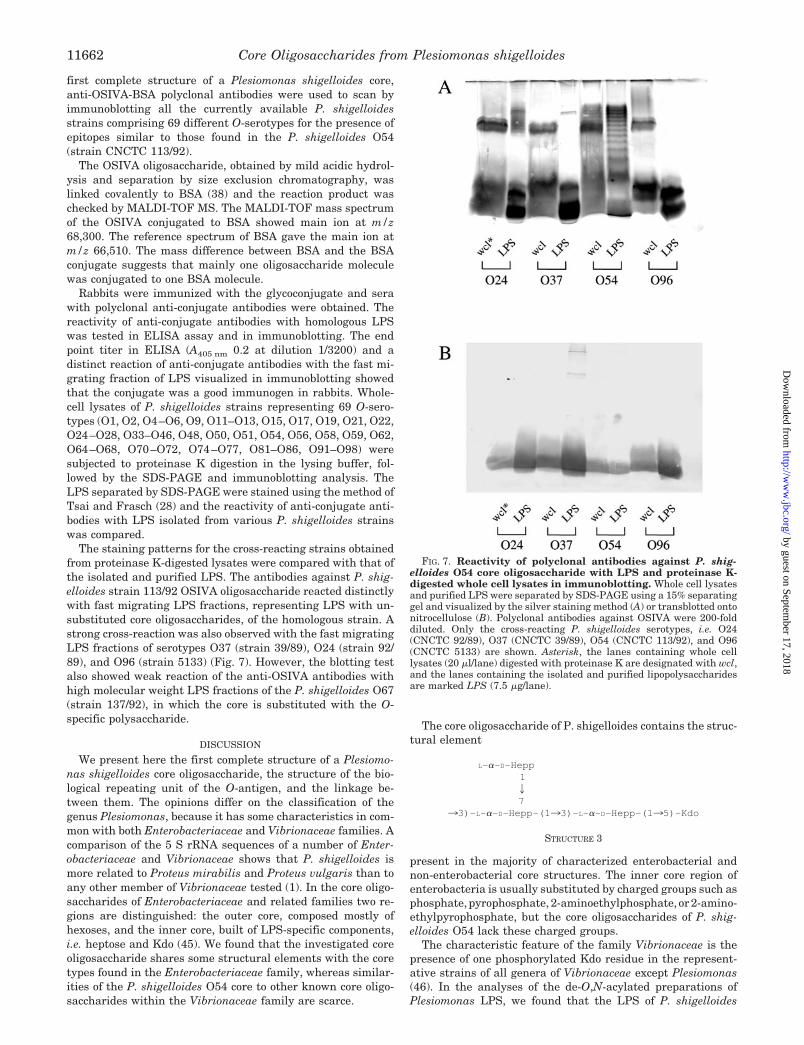
Rabbits were immunized with the glycoconjugate and serawith polyclonal anti-conjugate antibodies were obtained. Thereactivity of anti-conjugate antibodies with homologous LPSwas tested in ELISA assay and in immunoblotting. The endpoint titer in ELISA (A405 nm 0.2 at dilution 1/3200) and adistinct reaction of anti-conjugate antibodies with the fast mi-grating fraction of LPS visualized in immunoblotting showedthat the conjugate was a good immunogen in rabbits. Whole-cell lysates of P. shigelloides strains representing 69 O-sero-types (O1, O2, O4–O6, O9, O11–O13, O15, O17, O19, O21, O22,O24–O28, O33–O46, O48, O50, O51, O54, O56, O58, O59, O62,O64–O68, O70–O72, O74–O77, O81–O86, O91–O98) weresubjected to proteinase K digestion in the lysing buffer, fol-lowed by the SDS-PAGE and immunoblotting analysis. TheLPS separated by SDS-PAGE were stained using the method ofTsai and Frasch (28) and the reactivity of anti-conjugate anti-bodies with LPS isolated from various P. shigelloides strainswas compared.
The staining patterns for the cross-reacting strains obtainedfrom proteinase K-digested lysates were compared with that ofthe isolated and purified LPS. The antibodies against P. shig-elloides strain 113/92 OSIVA oligosaccharide reacted distinctlywith fast migrating LPS fractions, representing LPS with un-substituted core oligosaccharides, of the homologous strain. Astrong cross-reaction was also observed with the fast migratingLPS fractions of serotypes O37 (strain 39/89), O24 (strain 92/89), and O96 (strain 5133) (Fig. 7). However, the blotting testalso showed weak reaction of the anti-OSIVA antibodies withhigh molecular weight LPS fractions of the P. shigelloides O67(strain 137/92), in which the core is substituted with the O-specific polysaccharide.
DISCUSSION
We present here the first complete structure of a Plesiomo-nas shigelloides core oligosaccharide, the structure of the bio-logical repeating unit of the O-antigen, and the linkage be-tween them. The opinions differ on the classification of thegenus Plesiomonas, because it has some characteristics in com-mon with both Enterobacteriaceae and Vibrionaceae families. Acomparison of the 5 S rRNA sequences of a number of Enter-obacteriaceae and Vibrionaceae shows that P. shigelloides ismore related to Proteus mirabilis and Proteus vulgaris than toany other member of Vibrionaceae tested (1). In the core oligo-saccharides of Enterobacteriaceae and related families two re-gions are distinguished: the outer core, composed mostly ofhexoses, and the inner core, built of LPS-specific components,i.e. heptose and Kdo (45). We found that the investigated coreoligosaccharide shares some structural elements with the coretypes found in the Enterobacteriaceae family, whereas similar-ities of the P. shigelloides O54 core to other known core oligo-saccharides within the Vibrionaceae family are scarce.
The core oligosaccharide of P. shigelloides contains the struc-tural element
L-�-D-Hepp127
33)-L-�-D-Hepp-(133)-L-�-D-Hepp-(135)-Kdo
STRUCTURE 3
present in the majority of characterized enterobacterial andnon-enterobacterial core structures. The inner core region ofenterobacteria is usually substituted by charged groups such asphosphate, pyrophosphate, 2-aminoethylphosphate, or 2-amino-ethylpyrophosphate, but the core oligosaccharides of P. shig-elloides O54 lack these charged groups.
The characteristic feature of the family Vibrionaceae is thepresence of one phosphorylated Kdo residue in the represent-ative strains of all genera of Vibrionaceae except Plesiomonas(46). In the analyses of the de-O,N-acylated preparations ofPlesiomonas LPS, we found that the LPS of P. shigelloides
FIG. 7. Reactivity of polyclonal antibodies against P. shig-elloides O54 core oligosaccharide with LPS and proteinase K-digested whole cell lysates in immunoblotting. Whole cell lysatesand purified LPS were separated by SDS-PAGE using a 15% separatinggel and visualized by the silver staining method (A) or transblotted ontonitrocellulose (B). Polyclonal antibodies against OSIVA were 200-folddiluted. Only the cross-reacting P. shigelloides serotypes, i.e. O24(CNCTC 92/89), O37 (CNCTC 39/89), O54 (CNCTC 113/92), and O96(CNCTC 5133) are shown. Asterisk, the lanes containing whole celllysates (20 �l/lane) digested with proteinase K are designated with wcl,and the lanes containing the isolated and purified lipopolysaccharidesare marked LPS (7.5 �g/lane).
Core Oligosaccharides from Plesiomonas shigelloides11662
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

possesses two Kdo residues2 in the inner core. We also foundthat none of the sugar residues within the core oligosaccharideof the P. shigelloides LPS was substituted by phosphate groups.
The isolation of the OSIII oligosaccharide, i.e. the completecore oligosaccharide substituted with one repeating unit, notonly showed the structure of the biological repeating unit of theO-antigen but also allowed for the identification of the linkagebetween the O-specific polysaccharide and the core region. The33)-�-D-GlcpNAc-(13 (residue L) found at the reducing end ofthe O-specific polysaccharide repeating unit is linked to O-4 (C-4at � 80.3 ppm) of �-D-Glcp-(13 (residue I) of the core oligosac-charide OSIVA. The anomeric configuration of that linkage isretained as in the O-polysaccharide (�-configuration).
The herein reported core oligosaccharide structure is thefirst one described for the LPS of the genus Plesiomonas. How-ever, we have found that, despite the fact that Plesiomonas isclassified within the Vibrionaceae family, the core region of theP. shigelloides O54 (strain CNCTC 113/92) LPS possesses somecharacteristic structural features of the enterobacterial coreoligosaccharides, and it differs substantially from those de-scribed for other genera among Vibrionaceae (47–49).
The results obtained from serological screening of 69 P. shig-elloides O-serotypes suggest that structural elements of thecore oligosaccharide of serotype O54, recognized by the poly-clonal antibodies directed against the core oligosaccharide con-jugate, are only shared by the strains of the serotypes O24(strain CNCTC 92/89), O37 (strain CNCTC 39/89), and O96(strain CNCTC 5133). The presence of many non-reactive coreoligosaccharides suggests the lack of a uniform core structureamong the screened P. shigelloides O-serotypes. Such observa-tions were reported previously for bacteria belonging to othergenera, e.g. Escherichia, Proteus, Citrobacter, and Salmonella,possessing diverse core types within a species (45, 50).
REFERENCES
1. Farmer, J. J., III, Arduino, M. J., and Hickman-Brenner, F. W. (1992) in TheProkaryotes (Balows, A., Truper, H. G., Dworkin, M., Wim, H., andSchleifer, K. H., eds) Vol. 3, pp. 3012–3043, Springer-Verlag, New York
2. Van Houten, R., Farberman, D., Norton, J., Ellison, J., Kiehlbauch, J., Morris,T., and Smith, P. (1998) Morb. Mortal. Wkly. Rep. 47, 394–396
3. Levy, D. A., Bens, M. S., Craun, G. F., Calderon, R. L., and Herwaldt, B. L.(1998) Morb. Mortal. Wkly. Rep. 47, 1–34
4. Yamada, S., Matsushita, S., Dejsirilert, S., and Kudoh, Y. (1997) Epidemiol.Infect. 119, 121–126
5. Rautelin, H., Sivonen, A., Kuikka, A., Renkonen, O. V., Valtonen, V., andKosunen, T. U. (1995) Scand. J. Infect. Dis. 27, 495–498
6. Lee, A. C. W., Yuen, K. Y., Ha, S. Y., Chiu, D. C. K., and Lau, Y. L. (1996)Pediatr. Hematol. Oncol. 13, 265–269
7. Riley, P. A., Parasakthi, N., and Abdullah, W. A. (1996) Clin. Infect. Dis. 23,206–207
8. Clark, R. B., Westby, G. R., Spector, H., Soricelli, R. R., and Young, C. L. (1991)J. Infect. 23, 89–92
9. Delforge, M. L., Devriendt, J., Glupczynski, Y., Hansen, W., and Douat, N.(1995) Clin. Infect. Dis. 21, 692–693
10. Fujita, K., Shirai, M., Ishioka, T., and Kakuya, F. (1994) Acta Paediatr. Jpn.36, 450–452
11. Bravo, L., Monte, R., Ramirez, M., Garcia, B., Urbaskova, P., and Aldova, E.(1998) Cent. Eur. J. Public Health 6, 67–70
12. Korner, R. J., Macgowan, A. P., and Warner, B. (1992) Zentralbl. Bakteriol.277, 334–339
13. Ruimy, R., Breittmayer, V., Elbaze, P., Lafay, B., Boussemart, O., Gauthier,M., and Christen, R. (1994) Int. J. Syst. Bacteriol. 44, 416–426
14. Aldova, E. (1987) Zentralbl. Bakteriol. 265, 253–26215. Aldova, E. (1992) Syst. Appl. Microbiol. 15, 70–7516. Aldova, E., Benackova, E., Shimada, T., and Danesova, D. (1992) Syst. Appl.
Microbiol. 15, 247–24917. Shimada, T., Arakawa, E., Itoh, K., Kosako, Y., Inoue, K., Zhengshi, Y., and
Aldova, E. (1994) Curr. Microbiol. 28, 351–35418. Shimada, T., and Sakazaki, R. (1978) Jpn. J. Med. Sci. Biol. 31, 135–14219. Aldova, E., and Shimada, T. (2000) Folia Microbiol. 45, 301–30420. Albert, M. J., Ansaruzzaman, M., Qadri, F., Hossain, A., Kibriya, A., Haider,
K., Nahar, S., Faruque, S. M., and Alam, A. N. (1993) J. Med. Microbiol. 39,211–217
21. Linnerborg, M., Widmalm, G., Weintraub, A., and Albert, M. J. (1995) Eur.J. Biochem. 231, 839–844
22. Czaja, J., Jachymek, W., Niedziela, T., Lugowski, C., Aldova, E., and Kenne, L.(2000) Eur. J. Biochem. 267, 1672–1679
23. Petersson, C., Niedziela, T., Jachymek, W., Kenne, L., Zarzecki, P., andLugowski, C. (1997) Eur. J. Biochem. 244, 580–586
24. Westphal, O., and Jann, K. (1965) Methods Carbohydr. Chem. 5, 83–8925. Hitchcock, P. J., and Brown, T. M. (1983) J. Bacteriol. 154, 269–27726. Laemmli, U. K. (1970) Nature 227, 680–68527. Niedziela, T., Petersson, C., Helander, A., Jachymek, W., Kenne, L., and
Lugowski, C. (1996) Eur. J. Biochem. 237, 635–64128. Tsai, C. M., and Frasch, C. E. (1982) Anal. Biochem. 119, 115–11929. Sawardeker, J. S., Sloneker, J. H., and Jeanes, A. R. (1965) Anal. Chem. 37,
1602–160430. Gerwig, G. J., Kamerling, J. P., and Vliegenthart, J. F. G. (1978) Carbohydr.
Res. 62, 349–35731. Gerwig, G. J., Kamerling, J. P., and Vliegenthart, J. F. (1979) Carbohydr. Res.
77, 10–1732. Taylor, R. L., Shively, J. E., and Conrad, H. E. (1976) Methods. Carbohydr.
Chem. 7, 149–15133. Hakomori, S. (1964) J. Biochem. (Tokyo) 55, 205–20834. Bhat, U. R., Krishnaiah, B. S., and Carlson, R. W. (1991) Carbohydr. Res. 220,
219–22735. MacKenzie, S. L., and Tenaschuk, D. (1974) J. Chromatogr. 97, 19–2436. MacKenzie, S. L., and Hogge, L. R. (1977) J. Chromatogr. 132, 485–49337. Goddard, T. D., and Kneller, D. G. (2001) SPARKY, 3rd Ed., University of
California, San Francisco38. Boratynski, J., and Roy, R. (1998) Glycoconj. J. 15, 131–13839. Lugowski, C., and Romanowska, E. (1978) Eur. J. Biochem. 91, 89–9740. Jennings, H. J., and Lugowski, C. (1981) J. Immunol. 127, 1011–101841. Voller, A., Draper, C., Bidwell, D. E., and Bartlett, A. (1975) Lancet 1, 426–42842. Gorin, P. A., and Mazurek, M. (1975) Can. J. Chem. 53, 1212–122743. Jansson, P. E., Kenne, L., and Widmalm, G. (1989) Carbohydr. Res. 188,
169–19144. Susskind, M., Brade, L., Brade, H., and Holst, O. (1998) J. Biol. Chem. 273,
7006–701745. Rietschel, E. T., Kirikae, T., Schade, F. U., Mamat, U., Schmidt, G., Loppnow,
H., Ulmer, A. J., Zahringer, U., Seydel, U., Dipadova, F., Schreier, M., andBrade, H. (1994) FASEB J. 8, 217–225
46. Kondo, S., Haishima, Y., and Hisatsune, K. (1992) Carbohydr. Res. 231, 55–6447. Shaw, D. H., Hart, M. J., and Luderitz, O. (1992) Carbohydr. Res. 231, 83–9148. Knirel, Y. A., and Kochetkov, N. K. (1993) Biochemistry. (Mosc.) 58, 84–9949. Kondo, S., Zahringer, U., Seydel, U., Sinnwell, V., Hisatsune, K., and
Rietschel, E. T. (1991) Eur. J. Biochem. 200, 689–69850. Olsthoorn, M. M. A., Petersen, B. O., Schlecht, S., Haverkamp, J., Bock, K.,
Thomas-Oates, J. E., and Holst, O. (1998) J. Biol. Chem. 273, 3817–38292 L. Kenne, manuscript in preparation.
Core Oligosaccharides from Plesiomonas shigelloides 11663
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Czeslaw Lugowski and Lennart KenneTomasz Niedziela, Jolanta Lukasiewicz, Wojciech Jachymek, Monika Dzieciatkowska,
REPEATING UNIT, AND THE LINKAGE BETWEEN THEM LIPOPOLYSACCHARIDE CORE REGION, THE O-ANTIGEN BIOLOGICAL
STRUCTURAL AND SEROLOGICAL ANALYSIS OF THE O54:H2 (Strain CNCTC 113/92):Plesiomonas shigelloidesCore Oligosaccharides of
doi: 10.1074/jbc.M111885200 originally published online January 16, 20022002, 277:11653-11663.J. Biol. Chem.
10.1074/jbc.M111885200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/277/14/11653.full.html#ref-list-1
This article cites 49 references, 4 of which can be accessed free at
by guest on September 17, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























