Embed Size (px)
Citation preview

lable at ScienceDirect
Soil Biology & Biochemistry 77 (2014) 268e275
Contents lists avai
Soil Biology & Biochemistry
journal homepage: www.elsevier .com/locate/soi lbio
The initial lignin:nitrogen ratio of litter from above and below groundsources strongly and negatively influenced decay rates of slowlydecomposing litter carbon pools
Christine Walela a, *, Heiko Daniel a, Brian Wilson a, b, Peter Lockwood a,Annette Cowie a, c, d, Steven Harden e
a School of Environmental and Rural Sciences, Agronomy and Soil Science, University of New England, NSW 2351, Australiab NSW Office of Environment and Heritage, PO Box 0221, Armidale, NSW 2351, Australiac Rural Climate Solutions, University of New England, NSW 2351, Australiad Department of Primary Industries, NSW, Australiae NSW Department of Primary Industries, Tamworth Agricultural Institute, NSW, Australia
a r t i c l e i n f o
Article history:Received 10 February 2014Received in revised form5 June 2014Accepted 11 June 2014Available online 10 July 2014
Keywords:Carbon sequestrationCarbon poolsBiochemical qualityLitter decompositionLigninNitrogenAustraliaLand use
* Corresponding author.E-mail address: [email protected] (C. W
http://dx.doi.org/10.1016/j.soilbio.2014.06.0130038-0717/© 2014 Published by Elsevier Ltd.
a b s t r a c t
Understanding the interactions between the initial biochemical composition and subsequent decom-position of plant litter will improve our understanding of its influence on microbial substrate use toexplain the flow of organic matter between soil carbon pools. We determined the effects of land use(cultivation/native woodland/native pasture), litter type (above and below ground) and their interactionon the initial biochemical composition (carbon, nitrogen, water soluble carbon, lignin, tannin and cel-lulose) and decomposition of litter. Litter decomposition was studied as the mineralization of C fromlitter by microbial respiration and was measured as CO2eC production during 105 d of laboratory in-cubation with soil. A two-pool model was used to quantify C mineralization kinetics. For all litter types,the active C pool decay rate constants ranged from 0.072 d�1 to 0.805 d�1 which represented relativelyshort half-lives of between 1 and 10 days, implying that this pool contained compounds that wererapidly mineralized by microbes during the initial stages of incubation. Conversely, the decay rateconstants for the slow C pool varied widely between litter types within and among land uses rangingfrom 0.002 d�1 and 0.019 d�1 representing half-lives of between 37 and 446 days. In all litter types, theinitial lignin:N ratio strongly and negatively influenced the decay rate of the slow C pool which impliedthat the interaction between these two litter quality variables had important controls over the decom-position of the litter slow C pool. We interpret our results to suggest that where the flow of C from theactive pool to the slow pool is largely driven by microbial activity in soil, the rate of transfer of C will belargely controlled by the quality of litter under different land-use systems and particularly the initiallignin:N ratio of the litter. Compared with native pastures and cultivation, above and below ground litterfrom native woodland was characterized by higher lignin:N ratio and more slowly decomposing slow Cpools which implies that litter is likely to persist in soils, however based on the sandy nature of the soilsin this study, it is likely to lack protection from microbial degradation in the long term.
© 2014 Published by Elsevier Ltd.
1. Introduction
Plant litter decomposition plays an important role in the globalcarbon (C) cycle through its controls on fluxes of carbon dioxide(CO2) from soils and its influences on the build-up of soil organiccarbon (SOC) (Prescott, 2010). In order to effectively offset
alela).
greenhouse gas emissions through soil C sequestration, the build-up of stable SOC fractions is important (Cotrufo et al., 2013).Recently, advances have been made in our understanding of theprocesses of litter decomposition and factors that control the pro-portion of litter derived C that is incorporated into the stable SOCfraction (Cotrufo et al., 2013). It has been suggested that stableorganic matter compounds are formed in soils as a result of mi-crobial transformations of labile substrates during the early stagesof litter decomposition and that these microbial products interactwith soil minerals to form strong and stable organoemineral

C. Walela et al. / Soil Biology & Biochemistry 77 (2014) 268e275 269
complexes (Bahri et al., 2008; Grandy and Neff, 2008; Rubino et al.,2010). This mechanism contrasts with the conventional processmodel whereby SOM stability is imparted by the inherently recal-citrant SOC components that preferentially accumulate in SOMduring decomposition (Berg and McClaugherty, 2008).
Any factor that can act to constrain microbial activity, might inturn have a significant effect on the flow of organic matter betweenC pools (Sanderman et al., 2010). Biological processes are largelyregulated by climate (moisture and temperature) (Lavelle et al.,1993) and in arid and semi-arid environments, such as thosecommonly found in Australia, it is possible that microbial activityand litter degradation might be constrained by water availability.Under such conditions, microbes might therefore preferentiallydecompose the most energetically favourable organic matter sub-strates entering the soil thereby regulating the flow of organicmatter between C pools (Manzoni et al., 2012). In this case, complexplant structural compounds will incur a higher energy cost todegrade, (Bahri et al., 2008; Manzoni et al., 2012), and mighttherefore accumulate in soils and contribute to the slow cycling SOCfraction. Such a mechanism would generate the same outcome asthe conventional two-phase mineralization kinetics model. Litterchemistry might therefore still play an important role in influ-encing the decomposition kinetics and therefore the flowof organicmatter between SOC pools.
The majority of research on the biochemical composition of litterand its effect on decay has concentrated on above ground inputs asthis litter is deposited on the soil surface thereby strongly contrib-uting to SOC in the upper horizons (Lorenz et al., 2005). However,below ground C input also contributes to SOC with some reportsshowing that roots contribute more C to SOC than above groundinputs (Mafongoya et al., 1998; Wilhelm et al., 2004). Comparing thebiochemical composition of both above and below ground litter withtheir respective decay kinetics may improve our understanding ofthe differential contribution of plant litter materials to SOC(Mafongoya et al.,1998; Johnson et al., 2007). Further, the litter decaykinetics may be used in C cycle models estimating the movement ofplant derived C to SOC pools (Johnson et al., 2007). One of themethods used to study litter decomposition is through laboratoryincubations which assesses the mineralization of C from litter res-idue due to microbial activity and is measured as CO2eC production(Zeng et al., 2010; Cotrufo et al., 2010). This allows the relationshipbetween plant quality and decomposition kinetics to be investigatedunder controlled conditions with variables such as temperature andmoisture content (Johnson et al., 2007).
In this work, we studied the decomposition of above and belowground litter from three contrasting land uses namely; nativewoodland, native pastures and cultivation. Our aims were to a)determine the initial biochemical composition (litter chemistry) ofabove and below ground litter from the three land uses; b) deter-mine the C mineralization dynamics of decomposing plant litterover time and c) examine the interactions between litter chemistryand C mineralization kinetics.
2. Materials and methods
2.1. Site characteristics
This study was conducted near the township of Uralla in theNorthern Tablelands of New South Wales, Australia. Uralla lies at30.64�S, 151.49�E at an altitude of 1012 m and receives a meanannual rainfall of 807mm and has ameanmaximum andminimumtemperature of 26.4 �C and 12.5 �C respectively (1901e2012;Bureau of Meteorology, 2012). The soils of the area are largelyderived from Permian granite and are classified as yellow Chro-mosols (Isbell, 2002) equivalent to Alfisols (Soil survey staff, 1999).
2.2. Field sampling of above and below ground litter
Site clusters containing the three land uses namely, nativewoodland, native pastures and cultivation were randomly selectedat three separate locations (after Wilson et al. 2010, 2011). Aboveand below ground plant litter samples were collected from threerandomly selected plots within each land use, giving a total numberof 54 (3 sites � 3 land uses � 2 litter types � 3 reps) samples. Thenative pastures were dominated by native grasses such as Micro-laena stipoides. Above ground biomass from this land use consistedof standing dead biomass and was sampled from quadratsmeasuring 50 � 50 cm randomly placed in the paddocks (afterSanaullah et al., 2010). Above ground biomass within each quadratwas cut above the soil surface and collected into plastic bags.Thereafter, roots were collected by excavating soil from withinquadrats from the upper 15 cm soil layer. Native woodland con-sisted of a mixture of Eucalyptus species including Yellow box(Eucalyptus melliodora) and Blakely's Red Gum (Eucalyptus blakelyi).Above ground litter from this land use consisted of senescent leaveswhich were sampled from the ground within a randomly selected25 � 25 m plot. Sampling for below ground root litter was donewithin the same plot by randomly selecting three trees, establish-ing a distance of 3 m from each tree base and excavating roots from1.0 m soil depth.
The paddocks under cultivation had been predominantly crop-ped with forage oats (Avena sativa) for over 20 years. At the time oflitter sampling these paddocks were fallow and therefore forageoats (A. sativa) were grown in glasshouse pots until physiologicalmaturity from which standing dead biomass and roots were har-vested. The soil used for sowing forage oats (A. sativa) was sampledfrom the field within the plough layer (0e15 cm). At physiologicalmaturity, above ground litter was sampled by first discarding thestems bearing grains and harvesting only standing dead biomass.Roots were separated from the soil. In the laboratory, all aboveground litter from all land uses was sorted to remove any greenfoliage and retain only brown litter. Roots were separated from soilby sieving through <4 mm and picked out with forceps. All plantabove and below ground litter were oven-dried at 60 �C to constantweight (~48 h). Subsamples were taken from each above groundlitter type and passed through <4 mm.
2.3. Litter chemistry laboratory analysis
The initial biochemical composition of all plant litter wasdetermined on subsamples. Total C and N were determined by drycombustion using a CN LECO-1000 autoanalyser (LECO Corporation,St. Joseph, Mich). Water soluble carbon (WSC) was extractedfollowing methods described by Don and Kalbitz (2005). Briefly,0.2 g, of <4 mm above and below ground litter was weighed into130 ml specimen jars. Litter samples were soaked in 100 ml of ultrapure water for 24 h at 25 �C. The mixture was then filtered using a<45 mm nylon filter into 70 ml specimen jars. The solution sampleswere analysed for WSC using a Shimadzu TOC-5000 A analyzer.Acid detergent fibre (ADF) and lignin were determined by thesulphuric acid procedure using Foss FibreCap (tm) (AOAC, method973.18). In this study, lignin refers to the portion determined as acidunhydrolysable residue (AUR). Tannin was determined by extrac-tion with an aqueous methanol solution and measured by color-imetry in the presence of FolineDenis reagent (AOAC, method952.03). Cellulose was calculated from Acid detergent fibre-lignin.
2.4. Carbon mineralization
The kinetics of C mineralization were studied through a labo-ratory incubation which was performed under controlled

C. Walela et al. / Soil Biology & Biochemistry 77 (2014) 268e275270
conditions of 25 �C for 105 d. A sandy soil (yellow Chromosol) takenfrom our study site from a paddock under forage oats cultivationhistory (>20 years) was used as a medium for the incubation ofplant materials. Briefly, approximately 5 kg soils were sampledfrom the top layer (0e5 cm) and gently sieved through a 4 mmmesh on the same day and stored at 4 �C. An earlier assessment ofthe soil taken from this top layer showed that the soil contained5.6% clay, 10.4% silt 84% sand, 1.2% C, soil pH (water) of 5.3 and aninitial mineral nitrogen of 2.84 mg N kg�1 soil. Prior to the mainincubation experiment, soils (only) were pre-incubated in 2 L glassjars for one month to avoid capturing the initial flush of CO2eCwhich is associated with initial disturbance of soil.
In order to allow decomposition under non-limiting conditionsof N availability (Recous et al., 1995; Trinsoutrot et al., 2000) themineral N content of the soil in each jar was adjusted to80 mg N kg�1 soil. This was done by adding 77.16 mg N kg�1 soil asKNO3. Soils were then moistened to 60% of their water holdingcapacity (WHC) and plant litter (1.0 g) was incorporated homoge-neously and mixed thoroughly with the moist soil. The soil withoutaddition of any plant material was used as control. Triplicate blanks(empty jars) were incubated in the same way, but without soil-plant mixture. The CO2 produced by the plant litter amended soiland control treatments was trapped in 20 ml 1 M NaOH in 70-mlspecimen jars which were placed at the bottom of the jars. A smallvial containing 10 ml of deionised water was also placed at thebottom of the jar to maintain humid conditions and prevent soildesiccation. To ensure that the jars remained air-tight, they werereinforced with cling wrap before closing them tight.
The 2 L glass jars were placed in the dark in a constant tem-perature (25 �C) room for 105 days. In order to determine the CO2evolved from different treatments during the incubation period,the alkali traps were changed periodically on days 2, 5, 8, 14, 21, 28,35, 49, 63, 77 and 105. Prior to replacing fresh alkali traps and ateach sampling time, jars were left open for 1 h and allowed toequilibrate with the incubation room air in order to maintain aer-obic conditions. The moisture of the soil-plant litter mixture andthe soil only treatment was checked by periodic weighing of jarsduring each alkali-changing event and replenished by addingdeionised water and maintaining the moisture at field capacity 60%of their WHC. The alkali traps were then titrated with 0.5 M HCLusing an automatic double-point titration (pH 8.3 and 3.7) (TIM 840Titration Manager). The C mineralization of the added plant littermaterials was calculated as the difference in the amount ofCO2eCevolved from the soil-litter and the basal CO2eCevolved fromunamended soil (control) (Wang et al., 2004; Johnson et al., 2007).This calculation assumes any potential priming effects from theaddition of plant materials are negligible (Kuzyakov et al., 2000;Wang et al., 2004).
2.5. Fitting kinetic models to determine C mineralization kinetics
To quantify the kinetics of C mineralization in above and belowground litter, curves were fitted with a double exponential modelusing R software. Prior to fitting the model, the amount of % Cremaining in the soil from the plant litter relative to the initial Ccontents was first calculated following Equation (1) (Johnson et al.,2007; Sanaullah et al., 2010).
%Cremaining ¼ ½ðCinitial � CO2 � CevolvedÞ=Cinitial�100 (1)
Hence %C remaining was calculated as the fraction of the initialC (Cinitial) added in plant litter after subtracting the Cevolved as CO2(CO2eCevolved).
The double exponential model was fitted on the %C remainingdata using Equation (2) (Johnson et al., 2007).
Ct ¼ Ca expð � katÞ þ ð100� CaÞexpð � kstÞ (2)
where Ct is the percentage of C remaining at time t, Ca is the initialpercentage of C in the active pool, (100�Ca) is the initial percentageof C in the slow C pool, Ka (%C d�1) is the active pool decompositionrate constant and Ks (%C d�1) is the slow pool decomposition rateconstant. Half-lives were calculated as: HL ¼ 0.693/k where HL istime in days and k corresponds to either Ka or Ks. In order to validatethe best curve fit, the residual standard error (RSE) of the curvefitting was calculated.
2.6. Statistical analysis
The effects of land use (i.e. cultivation/native woodland/nativepasture), litter type (i.e. above and below ground) and their inter-action on the initial biochemical composition (i.e. carbon, nitrogen,water soluble carbon, lignin, tannin and cellulose) and percentageof C remaining after 105 days were analysed by ANOVA (Genstat,version 13.3). With three samples per land use at a site the blockterms were site and land use within site, except in those initialbiochemical composition variables with only one composite sam-ple per land use at a site where the only block term was site. Datawere square root transformed, where required, to normalizethe residuals. Land use by litter type means are shown in Tables 1and 2with differences determined by LSD at P ¼ 0.05 indicated byletters, with means within a column having a letter in common notbeing significantly different. Decomposition kinetics were esti-mated by fitting Equation (2) to Cmineralization data using the nonlinear mixed effects (nlme) function in the R software program.Pearson correlation coefficients were used to investigate the rela-tionship between initial biochemical composition of plant litter andC mineralization parameters including, Ca, Ka and Ks.
3. Results
3.1. Biochemical composition of above and below ground plant litter
Highly significant (P < 0.001) land use and litter type in-teractions were found for the initial biochemical composition oflitter which included; total carbon, total nitrogen, C/N ratio, cellu-lose, water soluble carbon (Table 1). Similarly, significant land useand litter type interactions were found for tannin (P ¼ 0.034),tannin/N ratio (P ¼ 0.005), lignin/N ratio (P ¼ 0.007), (lignin andtannin)/N ratio (P ¼ 0.006) and Cellulose/N ratio (P ¼ 0.002)(Table 1). These results indicate that the differences in biochemicalcomposition between and within land uses were dependent onlitter type. Significant land use main effects were found for ligninand lignin þ tannin (P < 0.001). The litter type main effect wassignificant for lignin (P ¼ 0.023) and nearly significant forlignin þ tannin (P ¼ 0.054), with no significant land use and littertype interaction observed (Table 1). The results for lignin andlignin þ tannin indicate that the three land uses had consistentdifferences in these properties across litter types.
The biochemical composition of the litter residues obtainedfrom the three land uses is shown in Table 2. Total C concentrationranged from 29.5% in oat roots to 48.8% in native woodland leaves.The magnitude of total C concentration in above ground litter wasin the order of native woodland > native pastures ¼ cultivation(P < 0.05) while that of below ground was in the order of nativewoodland > native pastures > cultivation (P < 0.05). Above groundlitter residues from the three land uses, consistently had largerconcentrations of total C than their below ground components.Total N concentration ranged from 0.46% in oat straw to 1.32% innative woodland leaves. Strong differences were similarly observed
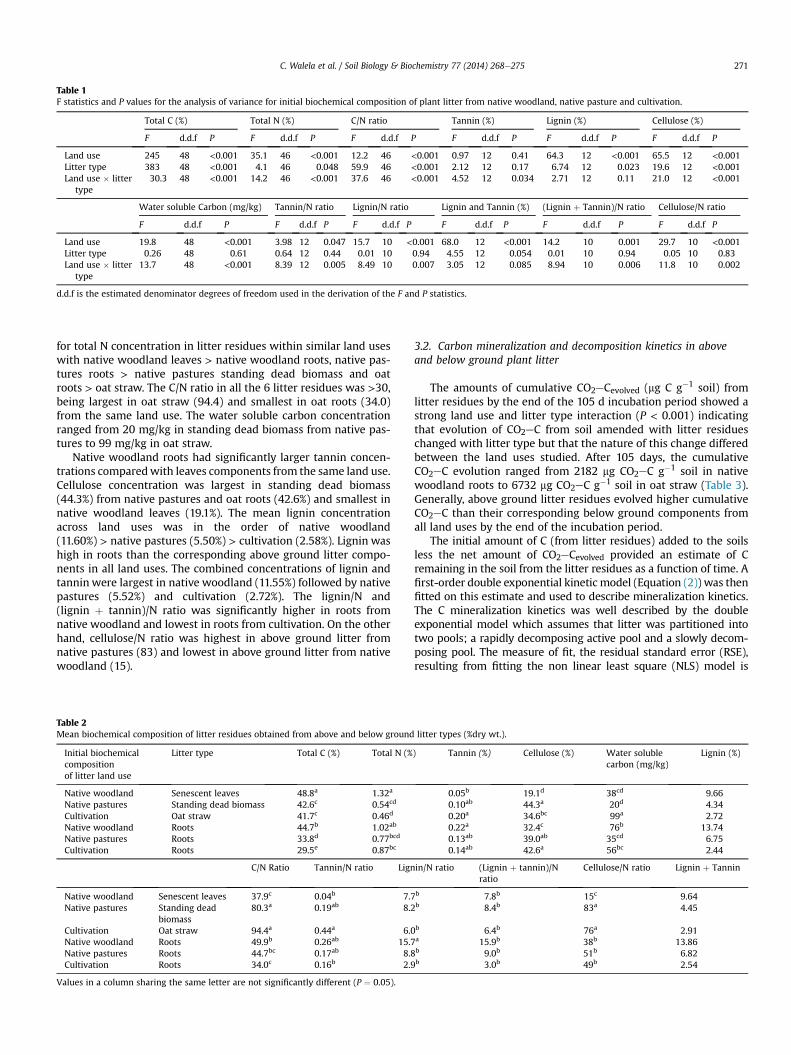
Table 1F statistics and P values for the analysis of variance for initial biochemical composition of plant litter from native woodland, native pasture and cultivation.
Total C (%) Total N (%) C/N ratio Tannin (%) Lignin (%) Cellulose (%)
F d.d.f P F d.d.f P F d.d.f P F d.d.f P F d.d.f P F d.d.f P
Land use 245 48 <0.001 35.1 46 <0.001 12.2 46 <0.001 0.97 12 0.41 64.3 12 <0.001 65.5 12 <0.001Litter type 383 48 <0.001 4.1 46 0.048 59.9 46 <0.001 2.12 12 0.17 6.74 12 0.023 19.6 12 <0.001Land use � litter
type30.3 48 <0.001 14.2 46 <0.001 37.6 46 <0.001 4.52 12 0.034 2.71 12 0.11 21.0 12 <0.001
Water soluble Carbon (mg/kg) Tannin/N ratio Lignin/N ratio Lignin and Tannin (%) (Lignin þ Tannin)/N ratio Cellulose/N ratio
F d.d.f P F d.d.f P F d.d.f P F d.d.f P F d.d.f P F d.d.f P
Land use 19.8 48 <0.001 3.98 12 0.047 15.7 10 <0.001 68.0 12 <0.001 14.2 10 0.001 29.7 10 <0.001Litter type 0.26 48 0.61 0.64 12 0.44 0.01 10 0.94 4.55 12 0.054 0.01 10 0.94 0.05 10 0.83Land use � litter
type13.7 48 <0.001 8.39 12 0.005 8.49 10 0.007 3.05 12 0.085 8.94 10 0.006 11.8 10 0.002
d.d.f is the estimated denominator degrees of freedom used in the derivation of the F and P statistics.
C. Walela et al. / Soil Biology & Biochemistry 77 (2014) 268e275 271
for total N concentration in litter residues within similar land useswith native woodland leaves > native woodland roots, native pas-tures roots > native pastures standing dead biomass and oatroots > oat straw. The C/N ratio in all the 6 litter residues was >30,being largest in oat straw (94.4) and smallest in oat roots (34.0)from the same land use. The water soluble carbon concentrationranged from 20 mg/kg in standing dead biomass from native pas-tures to 99 mg/kg in oat straw.
Native woodland roots had significantly larger tannin concen-trations comparedwith leaves components from the same land use.Cellulose concentration was largest in standing dead biomass(44.3%) from native pastures and oat roots (42.6%) and smallest innative woodland leaves (19.1%). The mean lignin concentrationacross land uses was in the order of native woodland(11.60%) > native pastures (5.50%) > cultivation (2.58%). Lignin washigh in roots than the corresponding above ground litter compo-nents in all land uses. The combined concentrations of lignin andtannin were largest in native woodland (11.55%) followed by nativepastures (5.52%) and cultivation (2.72%). The lignin/N and(lignin þ tannin)/N ratio was significantly higher in roots fromnative woodland and lowest in roots from cultivation. On the otherhand, cellulose/N ratio was highest in above ground litter fromnative pastures (83) and lowest in above ground litter from nativewoodland (15).
Table 2Mean biochemical composition of litter residues obtained from above and below ground
Initial biochemicalcompositionof litter land use
Litter type Total C (%) Total N (%
Native woodland Senescent leaves 48.8a 1.32a
Native pastures Standing dead biomass 42.6c 0.54cd
Cultivation Oat straw 41.7c 0.46d
Native woodland Roots 44.7b 1.02ab
Native pastures Roots 33.8d 0.77bcd
Cultivation Roots 29.5e 0.87bc
C/N Ratio Tannin/N ratio Lign
Native woodland Senescent leaves 37.9c 0.04b 7.7Native pastures Standing dead
biomass80.3a 0.19ab 8.2
Cultivation Oat straw 94.4a 0.44a 6.0Native woodland Roots 49.9b 0.26ab 15.7Native pastures Roots 44.7bc 0.17ab 8.8Cultivation Roots 34.0c 0.16b 2.9
Values in a column sharing the same letter are not significantly different (P ¼ 0.05).
3.2. Carbon mineralization and decomposition kinetics in aboveand below ground plant litter
The amounts of cumulative CO2eCevolved (mg C g�1 soil) fromlitter residues by the end of the 105 d incubation period showed astrong land use and litter type interaction (P < 0.001) indicatingthat evolution of CO2eC from soil amended with litter residueschanged with litter type but that the nature of this change differedbetween the land uses studied. After 105 days, the cumulativeCO2eC evolution ranged from 2182 mg CO2eC g�1 soil in nativewoodland roots to 6732 mg CO2eC g�1 soil in oat straw (Table 3).Generally, above ground litter residues evolved higher cumulativeCO2eC than their corresponding below ground components fromall land uses by the end of the incubation period.
The initial amount of C (from litter residues) added to the soilsless the net amount of CO2eCevolved provided an estimate of Cremaining in the soil from the litter residues as a function of time. Afirst-order double exponential kinetic model (Equation (2)) was thenfitted on this estimate and used to describe mineralization kinetics.The C mineralization kinetics was well described by the doubleexponential model which assumes that litter was partitioned intotwo pools; a rapidly decomposing active pool and a slowly decom-posing pool. The measure of fit, the residual standard error (RSE),resulting from fitting the non linear least square (NLS) model is
litter types (%dry wt.).
) Tannin (%) Cellulose (%) Water solublecarbon (mg/kg)
Lignin (%)
0.05b 19.1d 38cd 9.660.10ab 44.3a 20d 4.340.20a 34.6bc 99a 2.720.22a 32.4c 76b 13.740.13ab 39.0ab 35cd 6.750.14ab 42.6a 56bc 2.44
in/N ratio (Lignin þ tannin)/Nratio
Cellulose/N ratio Lignin þ Tannin
b 7.8b 15c 9.64b 8.4b 83a 4.45
b 6.4b 76a 2.91a 15.9b 38b 13.86b 9.0b 51b 6.82b 3.0b 49b 2.54
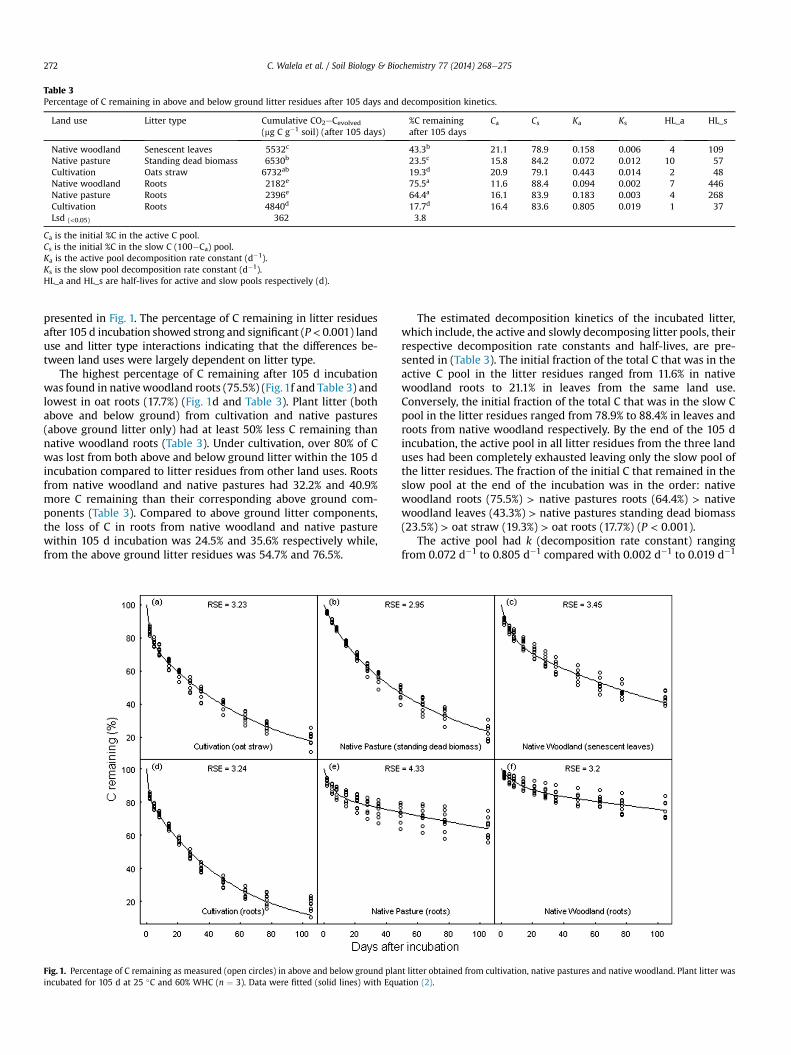
Table 3Percentage of C remaining in above and below ground litter residues after 105 days and decomposition kinetics.
Land use Litter type Cumulative CO2eCevolved(mg C g�1 soil) (after 105 days)
%C remainingafter 105 days
Ca Cs Ka Ks HL_a HL_s
Native woodland Senescent leaves 5532c 43.3b 21.1 78.9 0.158 0.006 4 109Native pasture Standing dead biomass 6530b 23.5c 15.8 84.2 0.072 0.012 10 57Cultivation Oats straw 6732ab 19.3d 20.9 79.1 0.443 0.014 2 48Native woodland Roots 2182e 75.5a 11.6 88.4 0.094 0.002 7 446Native pasture Roots 2396e 64.4a 16.1 83.9 0.183 0.003 4 268Cultivation Roots 4840d 17.7d 16.4 83.6 0.805 0.019 1 37Lsd (<0.05) 362 3.8
Ca is the initial %C in the active C pool.Cs is the initial %C in the slow C (100�Ca) pool.Ka is the active pool decomposition rate constant (d�1).Ks is the slow pool decomposition rate constant (d�1).HL_a and HL_s are half-lives for active and slow pools respectively (d).
C. Walela et al. / Soil Biology & Biochemistry 77 (2014) 268e275272
presented in Fig. 1. The percentage of C remaining in litter residuesafter 105 d incubation showed strong and significant (P< 0.001) landuse and litter type interactions indicating that the differences be-tween land uses were largely dependent on litter type.
The highest percentage of C remaining after 105 d incubationwas found in nativewoodland roots (75.5%) (Fig.1f and Table 3) andlowest in oat roots (17.7%) (Fig. 1d and Table 3). Plant litter (bothabove and below ground) from cultivation and native pastures(above ground litter only) had at least 50% less C remaining thannative woodland roots (Table 3). Under cultivation, over 80% of Cwas lost from both above and below ground litter within the 105 dincubation compared to litter residues from other land uses. Rootsfrom native woodland and native pastures had 32.2% and 40.9%more C remaining than their corresponding above ground com-ponents (Table 3). Compared to above ground litter components,the loss of C in roots from native woodland and native pasturewithin 105 d incubation was 24.5% and 35.6% respectively while,from the above ground litter residues was 54.7% and 76.5%.
Fig. 1. Percentage of C remaining as measured (open circles) in above and below ground planincubated for 105 d at 25 �C and 60% WHC (n ¼ 3). Data were fitted (solid lines) with Equ
The estimated decomposition kinetics of the incubated litter,which include, the active and slowly decomposing litter pools, theirrespective decomposition rate constants and half-lives, are pre-sented in (Table 3). The initial fraction of the total C that was in theactive C pool in the litter residues ranged from 11.6% in nativewoodland roots to 21.1% in leaves from the same land use.Conversely, the initial fraction of the total C that was in the slow Cpool in the litter residues ranged from 78.9% to 88.4% in leaves androots from native woodland respectively. By the end of the 105 dincubation, the active pool in all litter residues from the three landuses had been completely exhausted leaving only the slow pool ofthe litter residues. The fraction of the initial C that remained in theslow pool at the end of the incubation was in the order: nativewoodland roots (75.5%) > native pastures roots (64.4%) > nativewoodland leaves (43.3%) > native pastures standing dead biomass(23.5%) > oat straw (19.3%) > oat roots (17.7%) (P < 0.001).
The active pool had k (decomposition rate constant) rangingfrom 0.072 d�1 to 0.805 d�1 compared with 0.002 d�1 to 0.019 d�1
t litter obtained from cultivation, native pastures and native woodland. Plant litter wasation (2).

C. Walela et al. / Soil Biology & Biochemistry 77 (2014) 268e275 273
for the slow pool. The above decomposition rates of the active poolrepresented relatively short half-lives for all litter types from allland uses and ranged from 1 to 10 days. Generally, the active pool ofoat roots and straw had the fastest decomposition rates with half-lives of 1 and 2 days respectively (Table 3). Half-lives for the slowpool varied widely between litter types within and among landuses and ranged from 37 to 446 days (Table 3). Half-lives for theslow pool were highest in native woodland roots (446 days) fol-lowed by native pastures roots (268 days). These half-lives werehigher than those of their corresponding above ground compo-nents which were 109 and 57 days respectively. Shorter half-livesfor the slow pool were found for oat roots (37 days) than oatstraw (48 days).
3.3. Relationship between biochemical composition of above andbelow ground litter and carbon mineralization kinetics
The results on the relationship between biochemical composi-tion of litter and C mineralization kinetic parameters are presentedin Table 4. Across all litter types from all land uses, Ca was notsignificantly correlated to any of the plant litter biochemicalcompositional constituents that were measured. Highly significantnegative correlations were observed between Ka and lignin(r ¼ �0.58, P < 0.01), lignin/N ratio (r ¼ �0.51, P < 0.01),lignin þ tannin (r ¼ �0.57, P < 0.01), (lignin þ tannin)/N ratio(r ¼ �0.58, P < 0.01) and total carbon (r ¼ �0.59, P < 0.01). The Kswas similarly significantly and negatively correlated to total carbon(r ¼ �0.41, P < 0.01), total nitrogen (r ¼ �0.33, P < 0.05), lignin(r ¼ �0.59, P < 0.01), lignin/N ratio (r ¼ �0.65, P < 0.01),lignin þ tannin (r ¼ �0.59, P < 0.01) and (lignin þ tannin)/N ratio(r ¼ �0.64, P < 0.01) (Table 4).
4. Discussion
4.1. Biochemical composition of above and below ground plant litter
The differences in initial biochemical composition of plant lit-ters indicate significant quality differences between litter typeswithin and among the land uses studied. Plant litter (leaves and
Table 4Correlation coefficients (r) between biochemical composition of plant litter anddecomposition kinetics.
Mineralization curve parameters
Ca Ka Ks
Biochemical composition of litter residues (54 combinations of 2 littertypes � 3 land uses � 3 sites � 3 reps)
Total carbon 0.08 �0.59** �0.41**Total nitrogen �0.01 �0.16 �0.33*C/N ratio 0.10 �0.09 0.18Water soluble carbon �0.05 0.22 0.13Biochemical composition of litter residues (18 combinations of 2 litter
types � 3 land uses � 3 sites)Tannin 0.07 0.25 �0.10Tannin/N ratio 0.29 0.29 0.05Lignin �0.26 �0.58* �0.59**lignin/N ratio �0.04 �0.51* �0.65**Lignin and Tannin �0.26 �0.57* �0.59**(Lignin and Tannin)/N ratio �0.03 �0.50* �0.64**Cellulose 0.07 0.41 �0.05Cellulose/N ratio 0.39 0.23 �0.01ADF �0.07 0.11 �0.44
To be significant jrj has to be greater than.*0.2686 (5%), **0.3482 (1%) for 54 combinations; and.*0.4683 (5%), **0.5897 (1%) for 18 combinations.
roots) from native woodland was found to contain significantlylarger concentrations of lignin and tannin compounds comparedwith litter types from other land uses which were also significantlydifferent from each other, a result which is consistent with otherstudies (Rowell et al., 2001; Wang et al., 2004). Generally, forestlitter has been reported as having higher lignin and tannin contentcompared with crop residues or pastures/grass an observationmainly attributed to the presence of woody tissues in forest litter asopposed to large amounts of polysaccharides in crop and grass litter(Lorenz et al., 2005). Both lignin and tannin compounds are knownto contribute to the recalcitrance of plant litter by physically andchemically protecting the more easily degradable structural poly-saccharides (cellulose) from microbial degradation (Mafongoyaet al., 1998; Lorenz et al., 2005). The straw from oats had signifi-cantly larger amounts of WSC, compounds that are of high qualityand are primarily responsible for promotion of microbial growthand activity (Smith, 1994).
Our results indicate that litter from cultivation and native pas-tures may be considered to be relatively high in quality, with theabove ground litter being of higher quality than the below groundcomponents in the respective land uses. Although they had highC:N ratios, plant litters from the two land uses were largely char-acterized by large amounts of easily decomposable carbon fractionssuch as WSC, cellulose compounds, low lignin:N and(lignin þ tannin):N ratio. Conversely, plant litter from nativewoodland may be considered to be relatively low in quality as itcontained large amounts of biochemically recalcitrant compoundssuch as lignin and tannins, with below ground litter containinglarger amounts of these compounds compared with above groundlitter. These litter quality differences between plant litter typeswithin and among land uses would further be expected to impacton their subsequent C mineralization.
4.2. Carbon mineralization and decomposition kinetics in aboveand below ground plant litter
The addition of litter to the soil had strong and significant effectson C mineralization during the 105 d laboratory incubation. Thedifferences in the amount of residue C remaining in soils amendedwith different litter types implied that the litter materials used inthis study decomposed differently (Kuzyakov et al., 2000; Johnsonet al., 2007; Trinsoutrot et al., 2000). The C mineralization kineticsof the litter used in this study was well described by a doubleexponential model which partitioned litter mineralization into twoorganic C pools; the active and slow pools, which differed both insize and decomposition rates. However, recent advances in theunderstanding of mineralization kinetics suggest that duringdecomposition of these pools considerable interaction takes placebetween them and that microbial activity plays a major roleregulating the flow of C between these pools (Grandy and Neff,2008).
The differences observed between the size of the active andslow C pools reflect differences in the amounts of C inputs enteringinto the respective pools. Over 80% of the C was in the slow pool,while <20% of the C was in the active pool in the above and belowground litter in all land uses. Johnson et al. (2007) found pool sizesof similar magnitude, 25e35% active and 65e75% slowpool from anincubation study carried out for 498 d under controlled conditionssimilar to ours.
The active C pool had decomposition rates ranging from0.072 d�1 to 0.805 d�1 which represented relatively short half-livesof between 1 and 10 days across all litter types. This result impliesthat irrespective of litter type the active C pool is largely composedof easily degradable compounds which are available for rapid mi-crobial decomposition during the initial stages of decomposition.

C. Walela et al. / Soil Biology & Biochemistry 77 (2014) 268e275274
Wang et al. (2004) reported decomposition rate constants rangingfrom 0.042 d�1 to 0.24 d�1 for the active C pool for similar littertypes and land uses in Australia.
Although over 80% of the initial C remaining was reported asbeing in the slow C pool irrespective of litter type, there were sig-nificant differences in the decomposition rate of this pool amongthe land-uses and litter types studied, suggesting that litter qualityhas a significant impact on SOC cycling in the long term. Comparedwith native pastures and cultivation, the litter obtained fromabove- and below-ground components under native woodland wasassociated with larger half-lives of 109 and 446 days respectivelyindicating that litter was decomposing more slowly. These slow-pool decomposition rates are comparable with those reported byWang et al. (2004).
The decomposition rate of the slow C pool was correlatedstrongly and negatively with the initial lignin:N ratio of plant littersuggesting that the interaction between these two litter qualityvariables had important controls over decomposition of this pool. Anumber of mechanisms have been used to explain how lignin and Ninteract to influence decomposition of litter. For example, Talbotet al. (2012) suggested that lignin may form cross-linkages to themore labile N thereby chemically protecting N from hydrolysisduring litter decay (Talbot et al., 2012). Alternatively, lignin mayphysically protect the more labile cell wall components (e.g.structural polysaccharides and N) from microbial attack duringlitter decomposition when lignin is deposited in cell walls withinthe hemicelluloseeprotein matrices (Boerjan et al., 2003).
Our current understanding of the mechanism of organic matterdecomposition and transformation in soils involves a progressivetransfer of C from active to slow pools (Cotrufo et al., 2010). Weinterpret our results to suggest that where this flow of C from theactive pool to the slow pool is largely driven bymicrobial activity insoil, the rate of transfer of C will be largely controlled by the qualityof litter under different land-use systems and particularly the initiallignin:N ratio of the litter. The mechanisms of SOM stabilisation inthe long term will undoubtedly also be moderated by other factorsincluding the physical characteristics of the soil where organicmatter interact with soil minerals to form stable SOM (Rabbi et al.,2013, 2014). The sandy nature of the soils (5.6% clay, 10.4% silt, 84%sand) used in this study is likely to lack this protection mechanism.In Australian environments, another key constraint to SOCdecomposition is moisture availability. In the unpredictableAustralian climate, where soil moisture is often in deficit, microbialactivity and SOC decomposition processes will most probably beconstrained by this moisture limitation and SOC cycling mecha-nisms are likely to be episodic in line with climate patterns and, inparticular, precipitation.
5. Conclusions
Our laboratory incubation study showed that, the differences indecomposability between litter types was determined by differ-ences in the decomposition rate of the slow C pool. The decom-position rate of the slow C pool in litter was strongly and negativelycorrelated with the initial lignin:N ratio of plant litter suggestingthat the interaction between these two litter quality variables hadimportant controls over litter decomposition. Compared with otherlitter types, above and below ground litter from native woodlandhad higher initial lignin:N ratio and were associated with moreslowly decomposing slow C pools with longer half-lives of 109 and446 days respectively. Compared with native pastures and culti-vation, the more slowly decomposing litter from native woodlandis more likely to persist in soils, however based on the sandy natureof the soils, it is likely to lack protection frommicrobial degradationin the long term.
Acknowledgements
We gratefully acknowledge the financial support by the FordFoundation International Fellowship Program and University of NewEngland (UNE) for providing academic and research funds to theauthor. We are grateful to the land owners who granted us access totheir land to conduct our research and to technical staff at UNE, inparticular Leanne Lisle and Gary Cluley, for their assistance withlaboratory chemical analyses. We are also grateful to Bruce McCor-kell from the department of Primary Industries, Tamworth Agricul-tural Institute, NSW Australia, for partly assisting in statistics.
References
Berg, B., McClaugherty, C., 2008. Plant Litter: Decomposition, Humus Formation,Carbon Sequestration. Springer Verlag, Heidelberg, pp. 107e135.
Bahri, H., Rasse, D.P., Rumpel, C., Dignac, M.F., Bardoux, G., Mariotti, A., 2008. Lignindegradation during a laboratory incubation followed by (13) C isotope analysis.Soil Biol. Biochem. 40, 1916e1922.
Boerjan, W., Ralph, J., Baucher, M., 2003. Lignin biosynthensis. Annu. Rev. Plant Biol.54, 519e546.
Bureau of Meteorology, 2012. Averages for Uralla Climate Averages for AustraliaSites. Australia Government.
Cotrufo, M.F., Ngao, J., Marzaioli, F., Piermatteo, D., 2010. Inter-comparison ofmethods for quantifying above-ground leaf litter decomposition rates. PlantSoil 334, 365e376.
Cotrufo, M.F., Wallenstein, M.D., Boot, M.C., Denef, K., Paul, E., 2013. The microbialefficiency-matrix stabilization (MEMS) framework integrates plant litterdecomposition with soil organic matter stabilization: do labile plant inputsform stable soil organic matter? Glob. Change Biol. 19, 988e995.
Don, A., Kalbitz, K., 2005. Amounts and degradability of dissolved organic carbonfrom foliar litter at different decomposition stages. Soil Biol. Biochem. 37,2171e2179.
Grandy, A.S., Neff, J.C., 2008. Molecular C dynamics downstream: the biochemicaldecomposition sequence and its impact on soil organic matter structure andfunction. Sci. Total Environ. 404, 297e307.
Isbell, R.F., 2002. The Australian Soil Classification. CSIRO Publishing, Melbourne,Victoria, Australia.
Johnson, J.F., Barbour, N.W., Weyers, S.L., 2007. Chemical composition of cropbiomass impacts its decomposition. Soil Sci. Soc. Am. J. 71, 155e162.
Kuzyakov, Y., Friedel, J.K., Stahr, K., 2000. Review of mechanisms and quantificationof priming effects. Soil Biol. Biochem. 32, 1485e1498.
Lavelle, P., Blanchart, E., Martin, A., Martin, S., Spain, A., Toutain, F., Barois, I.,Schaefer, R., 1993. A hierarchical model for decomposition in terrestrial eco-systems e application to soils of the humid tropics. Biotropica 25, 130e150.
Lorenz, K., Lal, R., Donald, L.S., 2005. In: Norman, A.G. (Ed.), The Depth Distributionof Soil Organic Carbon in Relation to Land Use and Management and the Po-tential of Carbon Sequestration in Subsoil Horizons. Elsevier Academic Press.,California, pp. 35e66.
Mafongoya, P.L., Giller, K.E., Palm, C.A., 1998. Decomposition and nitrogen releasepatterns of tree prunings and litter. Agrofor. Syst. 38, 77e97.
Manzoni, S., Trofymow, J.A., Jackson, R.B., Porporato, A., 2012. Stoichiometric con-trols on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecol.Monogr. 80, 89e106.
Prescott, C.E., 2010. Litter decomposition: what controls it and how can we alter itto sequester more carbon in forest soils? Biogeochemistry 101, 133e149.
Rabbi, S.M.F., Hua, Q., Daniel, H., Lockwood, P., Wilson, B.R., Young, I.M., 2013. Meanresidence time of soil organic matter in aggregates under contrasting land usesbased on radiocarbon measurements. Radiocarbon 55, 127e139.
Rabbi, S.M.F., Wilson, B.R., Lockwood, P.V., Daniel, H., Young, I.M., 2014. Soil organiccarbon mineralization rates in aggregates under contrasting land uses. Geo-derma 216, 10e18.
Recous, S., Robin, D., Darwin, D., Mary, B., 1995. Soil inorganic N availability: effecton maize residue decomposition. Soil Biol. Biochem. 27, 1529e1538.
Rowell, D.M., Prescott, C.E., Preston, C.M., 2001. Decomposition and nitrogenmineralization from biosolids and other organic materials: relationship withinitial chemistry. J. Environ. Qual. 30, 1401e1410.
Rubino, M., Dungait, J.A.J., Evershed, R.P., 2010. Carbon input belowground is themajor C flux contributing to leaf litter mass loss: evidences from a (13) Clabelled leaf litter experiment. Soil Biol. Biochem. 42, 1009e1016.
Sanaullah, M., Chabbi, A., Lemaire, G., Charrier, X., Rumpel, C., 2010. How does plantleaf senescence of grassland species influence decomposition kinetics and littercompounds dynamics? Nutr. Cycle Agroecosyst. 88, 159e171.
Sanderman, J., Farquharson, R., Baldock, J., 2010. Soil Carbon Sequestration Poten-tial: a Review for Australian Agriculture. Report to the Australian GovernmentDepartment of Climate Change and Energy Efficiency. CSIRO, Canberra,Australia.
Smith, J.L., 1994. Cycling of nitrogen through microbial activity. In: Hatfield, J.L.,Stewart, B.A. (Eds.), Soil Biology: Effects on Soil Quality. Advances in Soil Sci-ence. CRC Press, Boca Raton., FL, USA, pp. 91e120.

C. Walela et al. / Soil Biology & Biochemistry 77 (2014) 268e275 275
Soil survey staff, 1999. Soil Taxonomy: a Basic System of Soil Classification forMaking And Interpreting Soil Surveys, second ed.. In: USDA AgriculturalHandbook, No. 436 Government Printer, Washington DC.
Talbot, J.M., Yelle, D.J., Nowick, J., Treseder, K.K., 2012. Litter decay rates are deter-mined by lignin chemistry. Biogeochemistry 108, 279e295.
Trinsoutrot, I., Recous, S., Bentz, B., Lin�eres, M., Ch�eneby, D., Nicolardot, B., 2000.Biochemical quality of crop residues and carbon and nitrogen mineralizationkinetics under nonlimiting nitrogen conditions. Soil Sci. Soc. Am. J. 64,918e926.
Wang, W.J., Baldock, J.A., Dalal, R.C., Moody, P.W., 2004. Decomposition dynamics ofplant materials in relation to nitrogen availability and biochemistry determinedby NMR and wet-chemical analysis. Soil Biol. Biochem. 36, 2045e2058.
Wilhelm, W.W., Johnson, J.M.F., Hatfield, J.L., Voorhees, W.E., Linden, D.R., 2004.Crop and soil productivity response to corn residue removal: a literature re-view. Agron. J. 96, 1e17.
Wilson, B.R., Barnes, P., Koen, T., Ghosh, S., King, D., 2010. Measurement and esti-mation of land-use effects on soil carbon and related properties on a basaltlandscape of northern NSW, Australia. Aust. J. Soil Res. 48, 421e433.
Wilson, B.R., Koen, T., Barnes, P., Ghosh, S., King, D., 2011. Soil carbon and related soilproperties along a soil type and land-use intensity gradient, New South Wales,Australia. Soil Use Manag. 27, 437e447.
Zeng, D., Mao, R., Chang, S., Li, L., Yang, D., 2010. Carbon mineralization of tree leaflitter and crop residues from poplar-based agroforestry systems in NortheastChina: a laboratory study. Appl. Soil Ecol. 44, 133e137.





























