Embed Size (px)
Citation preview

www.elsevier.com/locate/ijpsycho
International Journal of Psychop
The influence of encoding intention on electrophysiological indices
of recognition memory
Johanna Catharina van Hooff *,1
Department of Psychology, University of Portsmouth, King Henry I Street, Portsmouth, PO1 2DY, United Kingdom
Received 13 October 2003; received in revised form 24 May 2004; accepted 28 September 2004
Available online 10 November 2004
Abstract
The main aim of this study was to further specify the encoding and retrieval conditions that determine the success of an
ERP-based memory assessment procedure, originally derived from lie detection studies. We examined whether event-related
brain potentials (ERPs) recorded during successful and unsuccessful retrieval would vary according to intentional (study) and
incidental (repetition) encoding conditions. Participants (N=20) were asked to indicate recognition of previously studied words
(learned targets, p=0.2) and words that were used as distractors in a preceding recognition task (repeated targets, p=0.2). Words
that were recognised elicited a P3 component, which was largely absent for new words and words that failed to be recognised.
Encoding intention was found to increase the P3 amplitude slightly but had no influence on P3 scalp distribution, suggesting
that the differently encoded targets were similarly processed during retrieval but to a different extent. The amplitude difference
was explained in terms of variance in memory trace strength and decision confidence. With respect to negative findings for
repeated items in our earlier study (Van Hooff, J.C., Golden, S. 2002. Validation of an event-related potential memory
assessment procedure: Intentional learning as opposed to simple repetition. J. Psychophysiol., 16, 12–22.), it was suggested that
the instruction to actively retrieve the repeated words was essential for obtaining reliable indications of the presence or absence
of weak memory traces.
D 2004 Elsevier B.V. All rights reserved.
Keywords: P3; Recognition memory; Encoding intention; Memory trace strength; Retrieval instruction
1. Introduction
This experiment is part of a series of studies
examining the sensitivity of a memory assessment
0167-8760/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.ijpsycho.2004.09.010
* Present address: Department of Psychology, University of
Kent, Keynes College, Canterbury, Kent CT2 7NP, UK. Tel.: +44
1227 823097; fax: +44 1227 827030.
E-mail address: [email protected].
procedure, which, in addition to recognition judge-
ments, involves the recording of event-related brain
potentials (ERPs). The rationale of this procedure is
derived from lie detection studies (e.g., Allen et al.,
1992; Farwell and Donchin, 1991) and is based on the
assumption that a P3 ERP component can be reliably
elicited by items that are infrequently presented and
that possess special significance for the participants. In
previous studies, we have demonstrated that, in
hysiology 56 (2005) 25–36

J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–3626
absence of a behavioural indication of recognition,
items could gain special significance, and hence the
ability to elicit a P3, by virtue of previous learning (van
Hooff et al., 1996a) but not by virtue of mere repetition
(van Hooff and Golden, 2002). These results have
implications for the applicability of the ERP-based
memory assessment procedure since they suggest that
it may not be sensitive enough to detect possible weak
memory traces, for example, as a result of suboptimal
encoding conditions (e.g., during sleep or anaesthesia,
or in dual-task situations) or in patients with amnesic
syndrome (cf., Allen, 2002). Themain aim of this study
was to further specify the encoding and retrieval
conditions that determine the success of the ERP-based
memory assessment procedure. More specifically, it
focused on effects of intentional vs. unintentional
encoding and successful and unsuccessful retrieval.
The P3 component is identified as a positive
deflection in the ERP waveform, reaching a maximum
over the central–parietal areas in a 250–800 ms
poststimulus time window. The P3 is typically elicited
during an oddball paradigm and is believed to reflect
processes that are essential for event categorisation.
According to Kok (1997), these processes are con-
trolled by working memory and attention, which refers
to both automatic capturing of attention and active
focussing of attention. To this extent, researchers
sometimes distinguish different P3 subcomponents
where the dnovelty P3T mainly reflects involuntary
attention shifts to changes in the environment (Spencer
et al., 1999) while the centro-parietal dP3bT (or
dclassical P3T) mainly reflects processes associated
with the evaluation of task relevant stimuli (Kok,
2001). Furthermore, the novelty P3 has a more fronto-
central distribution than P3b and is believed to be
functionally related to the P3a subcomponent, which
indexes the automatic detection of deviant stimuli that
are not task relevant (Squires et al., 1975). In the rest of
this report, the P3 is referred to as the total collection of
these subcomponents, which frequently overlap in time
and which are not always easily distinguishable.
Targets or task relevant items are subject of focused
attention and can thus be expected to elicit large P3s
when correctly classified as targets. In contrast, non-
targets or task irrelevant items may capture the
participant’s attention and, consequently, may elicit a
P3, depending on the items’ specific attributes and their
relationship to the participant. The P3-evoking ability
of personally relevant items is believed to be an
automatic and involuntary process (Kok, 2001) and
has been used in the past to detect guilty knowledge
(Farwell and Donchin, 1991), deception (Rosenfeld et
al., 1991), concealed learning (Allen et al., 1992), and
feigned amnesia (Rosenfeld et al., 1995). In a typical
experiment of this kind, participants are first con-
fronted with a small set of items (i.e., the critical items),
which subsequently are embedded as nontargets in a
two-choice recognition task. The main finding in these
studies was that despite their nontarget status, the
critical items elicited a P3 component, presumably due
to automatic attention processes.
The above studies have in common that the critical
items were infrequently presented (cf., oddball para-
digm) and were made extra important by means of a
deceive or conceal instruction. Furthermore, the items
were typically well encoded through enactment (Far-
well and Donchin, 1991) or elaborate study (Allen et
al., 1992), or alternatively, were of an autobiograph-
ical nature (Rosenfeld et al., 1995). Following on
from these studies, we demonstrated that neither a
deceive instruction nor an association with a crime-
related or autobiographical event was needed to obtain
comparable results (van Hooff et al., 1996a). Aurally
presented, neutral words from one semantic category
that was previously studied were found to elicit a
reliable P3 component, even when these words did
not require a behavioural recognition response
(studied nontargets). This was replicated in a sub-
sequent study for visually presented words that were
not semantically related, but a similar effect could not
be observed for items that were repeated from an
earlier phase of the experiment (repeated nontargets;
van Hooff and Golden, 2002). This seemed to imply
that the type or depth of encoding crucially affected
the items’ ability to elicit a P3. It was therefore
concluded that the ERP-based memory assessment
procedure might be less suited to demonstrate the
presence of weak memory traces (here modelled by
repetition) in the absence of overt behavioural
recognition responses. Accordingly, this would then
also provide an explanation as to why we did not find
ERP evidence for memories for intraoperatively
presented words, using a highly similar memory
assessment procedure (van Hooff et al., 1996b). There
is at least one reason to doubt this conclusion,
however, because participants were not asked to

J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–36 27
actively try to retrieve the repeated items or to
cognitively distinguish between the different types
of nontargets. Consequently, it could have been the
absence of any retrieval effort that had caused the
nonsignificant findings in the van Hooff and Golden
(2002) study. Furthermore, the experimental setup did
not allow us to separate repeated items that may have
been partially recognised from those that were not
recognised to any extent. The current study was
designed to clarify these issues, using a comparable
study-test recognition paradigm as in our previous
studies but now including a condition in which
participants were asked to provide an affirmative
recognition response for the repeated items. More
specifically, participants first studied a short list of
words and then took part in two consecutive
recognition tests, of which, the second one is of our
primary interest (cf., van Hooff and Golden, 2002).
Target items in this second test were the words from
the study list (learned targets) as well as words that
served as distractors in the first test (repeated targets).
This setup would enable us to compare ERPs,
obtained in one and the same test condition, for
successfully recognised targets that had received
different types/levels of encoding (studied hits vs.
repeated hits). Furthermore, anticipating that a con-
siderable number of repeated targets would not be
recognised, it would also enable us to compare ERPs
for items classified as dnewT that were or were not
presented in an earlier task condition (repeated misses
vs. correctly rejected new items).
Based on earlier ERP studies manipulating the depth
of encoding, it was expected that repeated items that are
recognised as previously presented would elicit a P3,
but this P3 would probably be smaller than the one
elicited by successfully recognised studied items, due
to either a weaker representation inmemory (cf., Bentin
et al., 1992), a lower decision confidence (cf., Finnigan
et al., 2002), or a combination of both. Expectations
with regard to ERPs for repeated items that are not
recognised as previously presented were less straight-
forward. Based on our previous findings (van Hooff
and Golden, 2002), it could be hypothesised that they
would not elicit a differential ERP response; however,
this would be in contrast to findings by Rugg et al.
(1998) and Walla et al. (1999). Rugg et al. namely
reported that perceptually encoded words that were not
recognised as doldT produced more positive-going
ERPs (300–500 ms post stimulus) at the parietal
electrode positions, which, it was claimed, may be bacorrelate of memory in the absence of conscious
recognitionQ (Rugg et al., 1998, p. 596). Although this
finding could not be replicated in a more recent study
(Rugg et al., 2000), Walla et al. (1999) found a similar
but slightly earlier effect for semantically encoded
words that failed to be recognised. In addition, Walla
and colleagues also found a later frontal and right
parietal/occipital effect for the missed items, which
they believed to be associated with ban enhanced effortto retrieve item representations of the prior word
exposuresQ (Walla et al., 1999, p. 132). An important
difference between these studies and our previous
study (van Hooff and Golden, 2002) was that they
required their participants to behaviourally discrim-
inate between repeated and new items. In the present
study, we incorporated this task requirement, and so,
this would enable us to examine whether an active
retrieval effort would indeed be crucial for obtaining
significant ERP differences between repeated and new
items in the absence of a behavioural recognition
response.
2. Method
2.1. Participants
Twenty-two participants took part in the experi-
ment, but due to technical difficulties during record-
ing, only data from 20 participants were used for
analysis. All participants were undergraduate psy-
chology students (9 males, 11 females) and took part
in the experiment to gain course credits. They were
aged between 19 and 26 years (mean=21.1 years,
S.D.=2.21). All participants had English as their first
language, reported to be right handed, and had
normal or corrected-to-normal vision.
2.2. Stimuli and tasks
Stimuli were chosen from the same set of words as
in our previous study (van Hooff and Golden, 2002).
They consisted of 11 lists of six words each, and a set of
nine filler words. All 75 words were four- to seven-
letter nouns with a medium occurrence frequency
(mean 48.4 and range of 27–88 per million) according

J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–3628
to the norms of Johansson and Hofland (1989). To
ensure comparability between the 11-word lists, each
list consisted of three one-syllable words, two two-
syllable words, and one three-syllable word. The mean
word occurrence frequency for each list ranged
between 42 and 57. The words in each list were not
semantically related. Three or four words in each list
were concrete words. The stimuli were presented in
lowercase white letters (12 point ROM2 font) on a
black background (STIM software). They were cen-
trally presented on a PC monitor, which was placed at a
comfortable distance (approximately 70 cm) in front of
the participants. This resulted in horizontal visual
angles between 88 and 128.The experiment consisted of one study period and
two subsequent recognition tests. During the study
period, six words from one list were presented for
1520 ms with stimulus onset asynchronies (SOAs) of
2000 ms. The list was presented as many times as
needed with the same word order. Following this
study period, participants were required to conduct a
two-choice recognition test. This test consisted of
random presentation of the words from five lists,
including the list previously studied (e.g., lists A, B,
C, D, and E, of which list A was studied). The second
test consisted of random presentation of words from
eight lists. One of these eight lists was studied prior to
the first test (e.g., dstudiedT list A), and one was used
as a distractor list in the first test (e.g., drepeatedT listB); the other six lists were not presented previously
(e.g., dnewT lists F, G, H, I, J, and K). The lists were
rotated in a counterbalanced design, so that, for each
individual participant, other lists were studied,
repeated, and new. In this second test, words from
the studied list and the repeated list served as targets.
Because the targets ( p=1/8 for each list, respectively)
appeared less frequently then the nontargets ( p=6/8),
this was essentially an oddball task.
In both tests, words were presented for 306 ms
with SOAs of 2000 ms. Each test consisted of eight
blocks with different word orders. In all test blocks,
each word was presented only once. Because the
words were identical for each test block, all words
(targets and nontargets) were repeated eight times.
Because of possible influences of an orienting
response at the beginning of each test block, they
began with two out of the nine filler words, which
were discarded from further analysis. Each block also
finished with one of the filler words. YES–NO
responses had to be given for each word presented
by pressing either the left or right mouse button with
the index and middle finger, respectively. Left and
right were reversed for half the participants.
2.3. Procedure
Written consent was obtained from all participants
before the start of the experiment. Participants were
informed that the experiment would involve two word
recognition tests to obtain information about memory
processes in the brain. The recording procedure was
explained in detail using a standardised instruction
form and it was emphasised that participants could
withdraw from the experiment at any time if they
wished to do so. The specific aim and the exact nature
of the manipulations were not disclosed until after the
experiment.
Participants were first instructed to memorise a list
of six words which appeared on the monitor in front
of them. It was emphasised that they could view the
words as many times as required and that no record-
ings were taken at this stage of the study. After three
presentations of the study list, participants were asked
to recall the words in the order presented and in the
reversed order. If participants were unable to do this,
the list was presented another three times. If they had
no difficulties recalling the words in both orders, they
were asked to do so again after 1 min to ensure
storage in long-term memory. Once this was com-
pleted successfully, the instructions for the first test
were displayed on the monitor. Participants were
instructed to provide a YES response to the words
they recently studied and a NO response to all other
words. They were informed that words requiring a
YES response would appear less frequently than
words requiring a NO response, and therefore, they
should pay close attention. Participants were
requested to give their responses as accurately and
quickly as possible, using the specified mouse
buttons. To obtain a sufficient number of trials for a
proper signal-to-noise ratio, the test was repeated
eight times, with the same words in each test block but
different word orders. Participants could start each test
block at their own pace by pressing a mouse button.
Participants were furthermore asked to relax as much
as possible and to make no excessive movements.

J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–36 29
After completion of the first test and a short break
(approximately 2 min), new instructions were dis-
played on the monitor. Participants were this time
asked to provide a YES response to the words they
had previously studied (studied targets) and to the
words that had been presented as distractors in the
first test (repeated targets). A NO response was
required for all other words (nontargets). Participants
were informed that this test would take longer to
complete than the first test, and that again, they could
expect more words requiring a NO response than
words requiring a YES response. This test was also
repeated eight times and participants could start each
test block at their own pace. This second test was the
main focus of the current study. Because of the
described manipulation and because participants were
kept uninformed about the requirements of the second
test at the start of the experiment, it was assumed that
the studied targets and the repeated targets had
received different levels of encoding, producing
strong and weak memory traces, respectively.
2.4. EEG recording and ERP analysis
The scalp EEG was recorded from 19 tin electrodes
embedded in a strech-lycra cap (Electrocap Interna-
tional). Recording locations were based on the
International 10–20 system and included Fp1, Fp2,
F3, F4, F7, F8, Fz, T3, T4, C3, C4, Cz, T5, T6, P3,
P4, Pz, O1, and O2. Linked ear lobes were used as
reference, and Fpz served as ground. Vertical eye
movements (EOG) were recorded bipolarly between
electrodes placed above and below the right eye.
Impedances were kept below 5 kV. EEG and EOG
signals were recorded and amplified using a band pass
filter of 0.1 (24 dB) and 30 Hz (48 dB; Neuroscan 4.0
software). The amplified signals were digitised on-
line at a sampling rate of 200 Hz.
EEG data were corrected for eye movements,
using Neuroscan software. Recordings were then
visually inspected, and recording epochs containing
artifacts, such as excessive drift or muscle tone
interference, were removed before further analyses.
EEG epochs were subsequently created starting 100
ms prior to stimulus onset to 1500 ms following
stimulus onset. These epochs were then baseline-
corrected to a prestimulus baseline of �100 to 0 ms
and automatically checked for possible remaining
artifacts (amplitudesb�60 or N60 AV). Accepted
epochs were averaged for each channel time-locked
to the onset of the stimuli. Averaged ERP waveforms
were obtained for each word list and each response
category (dhitsT and dmissesT) separately.
3. Results
Repeated measures ANOVAs were used to analyse
the results, supplemented with Bonferroni pairwise
comparisons where appropriate. Because of easier
notification, the standard noncorrected degrees of
freedom are indicated for the obtained F-values in
the next section; however, these were adjusted using
the Greenhouse–Geisser correction method. Because
the second recognition task was the main focus of our
study, behavioural and ERP data are discussed for this
test only.
3.1. Behavioural data
The mean percentage of correct responses for the
new words (i.e., correct rejections) was 95.5%
(S.D.=4.8), whereas this was 85.6% (S.D.=17.4) for
the studied words and 54.3% (S.D.=22.0) for the
repeated words. As expected, recognition accuracy
(pHit–pFalse Alarm; Snodgrass and Corwin, 1988)
was higher for the studied targets than for the repeated
targets (t=5.61, df=19, pb0.001). Because of the
oddball character of the task, participants could
mistakenly respond NO to studied or repeated items
for two reasons: (1) because they did not recognise
them as being previously presented (dtrueT miss), or
(2) because they were affected by the negative
response bias (dunintentionalT miss). To distinguish
these two types of hypothetical misses, a variable was
created that indicated the number of words that was
missed six times or more (out of a possible eight, thus
75% of the time). If this was the case, it was believed
that this word was a dtrueT miss. It was found that the
number of dtrueT misses was substantially larger for
the repeated words (M=1.85, S.D.=1.84) as compared
to the studied words (M=0.20, S.D.=0.62; t=3.94,
df=19, pb0.01).
Mean reaction times (RTs) were calculated for each
stimulus and response category. For the new words,
only one list was selected to account for the larger

J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–3630
number of stimuli in this category. This list corre-
sponded to the one also selected for the ERP averages
(see further) and was counterbalanced across partic-
ipants. Mean RTs for correct responses were quickest
for the new words (M=544 ms, S.D.=89.7), slightly
slower for the studied targets (M=625ms, S.D.=105.6),
and slowest for the repeated targets [M=721 ms,
S.D.=107.2; F(2,38)=74.11, pb0.001]. All Bonferroni
corrected pairwise comparisons were significant at
pb0.001. For the studied targets (N=10), mean RTs for
correct responses (mean RT hits=618 ms) were
significantly slower than for incorrect responses (mean
RT misses=434 ms; t=4.82, df=9, pb0.01).2 For the
repeated targets (N=20), a similar effect was found
(mean RT hits=721 ms, mean RT misses=580 ms,
t=4.91, df=19, pb0.001). Because of these RT differ-
ences, it seems likely that, for both types of targets, at
least some of the misses were due to premature
responding (previously classified as unintentional
misses).
3.2. ERP data
Individual ERPs were averaged separately for each
stimulus and response category. For the new words,
only one list was selected for averaging (counter
balanced across participants) to approach a compara-
ble number of trials as for the studied and repeated
word lists. In agreement with previous studies (e.g.,
Rugg et al., 1998), a minimum number of 16 artifact-
free trials were considered sufficient to obtain a
reliable ERP. Because five participants did not
recognise many of the repeated targets, this criterion
resulted in that ERP comparisons for correct
responses were based on 15 participants only. Mean
number of trials contributing to the ERPs were 38.1
(studied hits), 28.3 (repeated hits), and 40.7 (correct
rejections), respectively. Similarly, 10 participants
performed very well and did not miss many of the
repeated targets; thus ERP comparisons for misses
were based on the other 10 participants who produced
a sufficient number of missed repeated targets (these
10 included the five left out in the first comparison).
2 Mean RTs were considered reliable when calculated on the
basis of at least five trials. Half of the participants did not meet this
criterion for the learned misses, and hence, this analysis was based
on data from 10 participants only.
Mean number of trials was 24.5 (repeated misses) and
27.2 (correct rejections).
3.2.1. Correct responses (studied and repeated hits)
Grand average ERP waveforms (N=15) are shown
in Fig. 1 for all electrode positions. As can be
observed in this figure, the three stimulus categories
elicited comparable early, visual evoked responses
(80–250 ms). In addition, a large positive wave,
starting around 400 ms poststimulus and reaching a
maximum at 520 ms, was present for all stimuli but
most clearly so for the recognised studied words and
the recognised repeated words. These positive waves
were present at all electrode positions but were largest
over the central and parietal areas. Because of these
characteristics and because they were elicited by
infrequent target stimuli, these waves were considered
to correspond to the P3 component. Visual inspection
of Fig. 1 reveals that the upward slope of the P3 seems
to occur somewhat earlier for the studied words as
compared to the repeated words. In contrast, the
downward slow seems to be prolonged for the
repeated words, particularly at the frontal electrode
positions. P3 amplitude appears largest for the studied
targets, slightly smaller for the repeated targets, and
considerably smaller for the nontargets.
The P3 peak was defined as the most positive peak
in the 450–650 ms post stimulus period. For statistical
analyses, P3 peak amplitude and latency were
obtained from three coronal chains of electrodes F3–
Fz–F4 (frontal), C3–Cz–C4 (central), P3–Pz–P4
(parietal). The three coronal chains were chosen to
enable analysis of scalp distribution differences in the
anterior–posterior and the lateral–medial direction. All
anticipated effects were most visible at these electrode
locations, and no additional effects could be observed
at the other electrodes (see Fig. 1). Repeated measures
ANOVAs were performed with stimulus category
(studied, repeated, new), anterior–posterior electrode
position (frontal, central, and parietal), and laterality
(left, central, and right), as within subjects factor.
A main effect of stimulus category [F(2,28)=39.6,
pb0.001] confirmed that the P3 peak was largest for the
studied hits (mean=8.05 AV), somewhat smaller for the
repeated hits (mean=7.38 AV), and smallest for the
correctly rejected new words (mean=4.44 AV). Bon-ferroni corrected pairwise comparisons showed that
both types of recognised targets (studied and repeated)
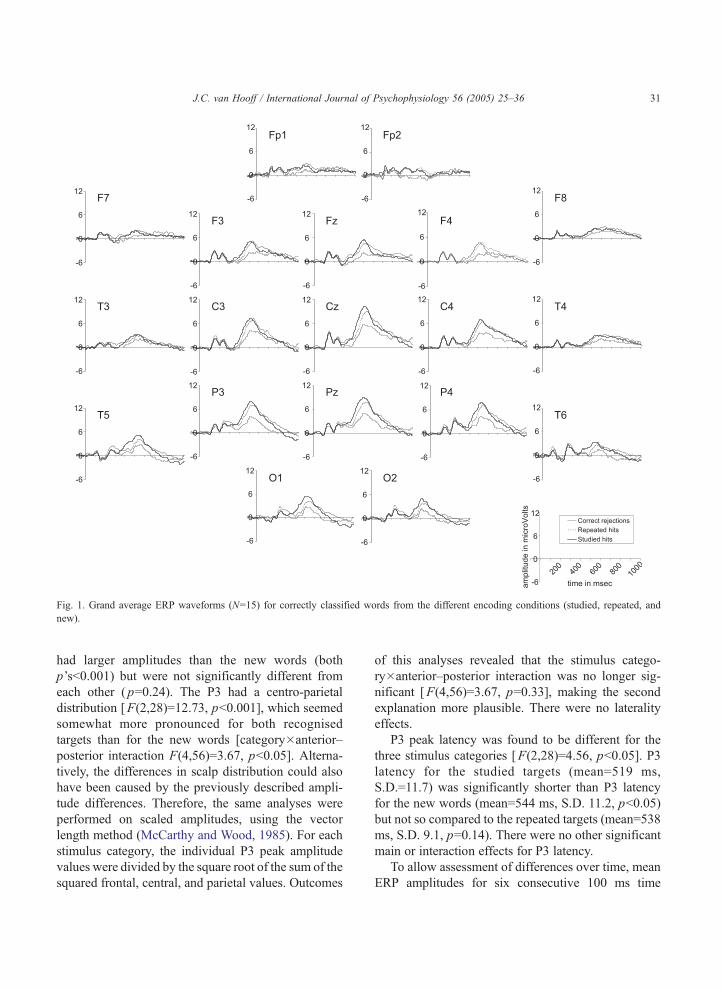
Fig. 1. Grand average ERP waveforms (N=15) for correctly classified words from the different encoding conditions (studied, repeated, and
new).
J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–36 31
had larger amplitudes than the new words (both
p’sb0.001) but were not significantly different from
each other ( p=0.24). The P3 had a centro-parietal
distribution [F(2,28)=12.73, pb0.001], which seemed
somewhat more pronounced for both recognised
targets than for the new words [category�anterior–
posterior interaction F(4,56)=3.67, pb0.05]. Alterna-
tively, the differences in scalp distribution could also
have been caused by the previously described ampli-
tude differences. Therefore, the same analyses were
performed on scaled amplitudes, using the vector
length method (McCarthy and Wood, 1985). For each
stimulus category, the individual P3 peak amplitude
values were divided by the square root of the sum of the
squared frontal, central, and parietal values. Outcomes
of this analyses revealed that the stimulus catego-
ry�anterior–posterior interaction was no longer sig-
nificant [F(4,56)=3.67, p=0.33], making the second
explanation more plausible. There were no laterality
effects.
P3 peak latency was found to be different for the
three stimulus categories [F(2,28)=4.56, pb0.05]. P3
latency for the studied targets (mean=519 ms,
S.D.=11.7) was significantly shorter than P3 latency
for the new words (mean=544 ms, S.D. 11.2, pb0.05)
but not so compared to the repeated targets (mean=538
ms, S.D. 9.1, p=0.14). There were no other significant
main or interaction effects for P3 latency.
To allow assessment of differences over time, mean
ERP amplitudes for six consecutive 100 ms time
J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–3632
windows (250–350, 350–450, 450–550, 550–650,
650–750, and 750–850 ms) were calculated for the
same three coronal chains of electrodes. The same
ANOVAs, as described above, were performed, for
which a summary of the main effects is shown in
Table 1.
The main effects of stimulus category and sub-
sequent pairwise comparisons (Bonferroni corrected)
confirmed our earlier observation that the P3 was
larger for the correctly identified targets (both studied
and repeated) than for the new words. In addition, the
P3 reached higher amplitudes for the studied targets
than the repeated targets ( pb0.05) in the 450–550 ms
time window. In the 650–750 ms time window
(downward slope of the P3), the repeated targets,
but not the studied targets, had significantly larger
amplitudes than the new words ( pb0.001). The main
effect of anterior–posterior position, followed by
pairwise comparisons, showed that the P3 had
primarily a centro-parietal distribution.
A stimulus category�anterior–posterior electrode
position interaction was present for the last four time
windows (all p’sb0.05), suggesting possible differ-
ences in anterior–posterior scalp distribution for
studied targets, repeated targets, and new nontargets.
A reanalysis on scaled amplitudes (procedure
described above) revealed that there were still signifi-
cant stimulus category�anterior–posterior interactions
for the 650–750 ms [F(4,56)=5.32, pb0.05] and 750–
850 ms [F(4,56)=4.30, pb0.05] time windows (i.e.,
downward slope of the P3). Inspection of mean
amplitudes revealed that this interaction referred to a
different anterior–posterior scalp distribution for the
targets as compared to the nontargets. More specifi-
Table 1
Repeated measures ANOVA main effects of category (learned targets, re
central, and parietal), and laterality (left, middle, and right) for mean ERP
ERP time window Category Anterior–
F(2,28) p Description F(2,28)
250–350 0.69 ns 19.14
350–450 8.92 ** ST/RTNN 5.10
450–550 46.15 *** STNRTNN 12.54
550–650 13.26 *** ST/RTNN 13.95
650–750 7.47 ** RTNN 9.82
750–850 1.64 ns 5.28
ns—nonsignificant; *—pb0.05; **—pb0.01; ***—pb0.001. Descript
ST—studied targets; RT—repeated targets; N—new words; F—frontal; C
cally, during these time windows, ERPs elicited by the
targets (both studied and repeated) were characterised
by a centro-parietal maximum, whereas ERPs elicited
by the nontargets were characterised by a central
maximum and a parietal minimum.
3.2.2. Incorrect responses (repeated misses)
Grand average ERPs (N=10) for correctly rejected
new words and missed repeated words are displayed in
Fig. 2 for electrode positions F3, F4, P3, and P4. Please
note that dtrueT misses and dincidentalT misses were
both included in these averages because of insufficient
trials in each separate category. Based on behavioural
results, it could be calculated that proportionally, about
43% of the trails were dtrueT misses. Visual inspection
of this figure, reveals that ERPs elicited by the repeated
targets that were missed seemed to be more positive
going in the 550–750 ms period than those elicited by
new words, especially over the frontal areas. In the
period thereafter (750–900 ms), the repeated misses
were still more positive going but for the parietal
electrode positions only. Because of its relatively late
occurrence and its scalp distribution, it is unlikely that
this wave corresponds to the classical P3 component.
Similar repeated measures ANOVAs were carried
out on the same time windows, as described earlier.
Significant main effects of stimulus category were
found for the 550–650 ms period [F(1,9)=8.46,
pb0.05], the 650–750 ms time period [F(1,9)=16.87,
pb0.01], and the 750–850 time period [F(1,9)=11.92,
pb0.01]. This confirmed the observation that the
nonrecognised repeated words elicited a more positive
going ERP waveform than the new words in the later
stages of the recording epoch. There were no signifi-
peated targets, and new items), anterior–posterior position (frontal,
amplitude measures for the different time windows
posterior Laterality
p Description F(2,28) p Description
*** FbCbP 1.74 ns
* 3.32 0.083 RiNLe
** FbC/P 3.72 *
*** FbC/P 0.93 ns
** FbC 1.51 ns
* F/PbC 2.81 0.092
ions only include significant Bonferroni pairwise comparisons.
—central; P—parietal; Ri—right; Le—left.
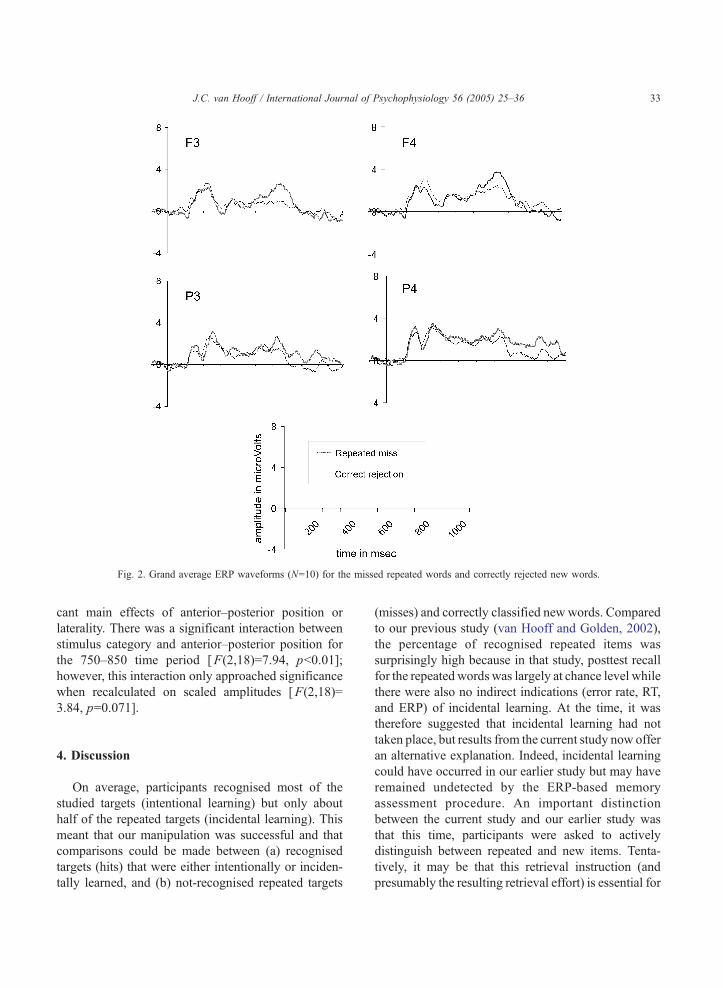
Fig. 2. Grand average ERP waveforms (N=10) for the missed repeated words and correctly rejected new words.
J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–36 33
cant main effects of anterior–posterior position or
laterality. There was a significant interaction between
stimulus category and anterior–posterior position for
the 750–850 time period [F(2,18)=7.94, pb0.01];
however, this interaction only approached significance
when recalculated on scaled amplitudes [F(2,18)=
3.84, p=0.071].
4. Discussion
On average, participants recognised most of the
studied targets (intentional learning) but only about
half of the repeated targets (incidental learning). This
meant that our manipulation was successful and that
comparisons could be made between (a) recognised
targets (hits) that were either intentionally or inciden-
tally learned, and (b) not-recognised repeated targets
(misses) and correctly classified newwords. Compared
to our previous study (van Hooff and Golden, 2002),
the percentage of recognised repeated items was
surprisingly high because in that study, posttest recall
for the repeated words was largely at chance level while
there were also no indirect indications (error rate, RT,
and ERP) of incidental learning. At the time, it was
therefore suggested that incidental learning had not
taken place, but results from the current study now offer
an alternative explanation. Indeed, incidental learning
could have occurred in our earlier study but may have
remained undetected by the ERP-based memory
assessment procedure. An important distinction
between the current study and our earlier study was
that this time, participants were asked to actively
distinguish between repeated and new items. Tenta-
tively, it may be that this retrieval instruction (and
presumably the resulting retrieval effort) is essential for

J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–3634
the success of the ERP-based memory assessment
procedure. Moreover, Rugg et al. (2000) described
retrieval effort as bthe mobilization of processing
resources in service of attempts to retrieve memoryQ(p. 673), and it might be these extra resources that are
essential not only for a behavioural recognition
response but also for the generation of a differential
ERP response. In future applications, the ERP-based
memory assessment procedure should therefore
include an explicit recognition instruction in order to
judge more reliably the presence or absence of weak
memory traces as a result of repetition or incidental
learning. This would also be more in line with studies
investigating spared memory functions in patients with
amnesic syndrome (cf., Lalouschek et al., 1997), in
patients undergoing general anaesthesia (van Hooff et
al., 1995, 1996b), or in patients with dissociative
identity disorder (Allen and Movius, 2000).
Both types of recognised targets elicited a reliable
P3 component with a centro-parietal maximum and a
peak latency of about 520–540 ms post stimulus. This
is in agreement with our previous studies (van Hooff et
al., 1996a; van Hooff and Golden, 2002) and other
studies using comparable oddball recognition para-
digms (e.g., Allen et al., 1992; Farwell and Donchin,
1991). Although P3 peak amplitude did not differ
between the studied and repeated targets, mean
amplitudes in the 450–550 ms time window were
somewhat larger for the studied targets as compared to
the repeated targets. As mentioned in the Introduction,
this might have been the result of a stronger memory
trace strength (cf., Bentin and Moscovitch, 1990;
Bentin et al., 1992) or a higher decision confidence
(Finnigan et al., 2002). An interpretation in terms of
memory trace strength would fit in best with the
model of the P3 amplitude presented by Kok (1997,
2001). In Kok’s model, the P3 is considered to reflect
a target identification mechanism that can be con-
ceptualised as ba set of neural elements or
drecognition unitsT that form a neural networkQ, theprimary function of which is bto compare stimulus
attributes with an internal representation of the targetQ(Kok, 2001, p. 571). Accordingly, a P3 will only be
elicited when some kind of connection has been
established between the perceptual and memory
systems. Thus, when a target is presented and a
matching process has been triggered, the recognition
units will be activated and more units will be
activated as a result of stronger memory traces.
Hence, in our experiment, the studied items may have
elicited somewhat larger P3s than the repeated items
because they had formed stronger memory traces.
This amplitude difference and the absence of scalp
distribution differences furthermore suggests that
intentional study and repetition have generated
memory traces that give rise to retrieval mechanisms
that differ quantitatively rather than qualitatively. This
is an important finding that can be contrasted with
(nonoddball) studies using perceptual- and semantic-
encoding conditions to create weak and strong
memory traces, respectively (e.g., Rugg et al., 1998,
2000). Results from these studies namely suggested
that while recognition of perceptually (or shallowly)
encoded words relied primarily on familiarity pro-
cesses (as reflected by an early decreased negativity
over the frontal areas), recognition of semantically (or
deeply) encoded words relied on familiarity and
recollection processes (as reflected by an additional
late increased positivity over the left central–parietal
areas).
An alternative explanation for the P3 amplitude
difference could be that participants were more
confident in categorising the studied targets as
compared to the repeated targets. This interpretation
is supported by the behavioural results showing a
higher recognition accuracy, a lower number of dtrueTmisses, and a faster mean RT for the studied targets as
compared to the repeated targets. It is also in
agreement with Finnigan et al.’s (2002) suggestion
that the amplitude of a late positive component (LPC)
is modulated by decisional factors. However, unlike
Finnigan et al., we did not observe a preceding or
partially overlapping effect (which they claimed to be
a N400 and not a P3 effect) that they associated with
memory trace strength. It may therefore be speculated
that variance in memory trace strength and decision
confidence both have contributed to the observed P3
amplitude difference between learned and repeated
targets (if indeed they can be presumed to be
manifestations of two different processes). Task
characteristics that might have promoted a combina-
tion or overlap of effects were the oddball format and
the requirement for a quick rather than a delayed
response. In a future study, the inclusion of confidence
ratings should be able to shed more light on this issue,
since even Finnigan et al. seemed less certain to

J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–36 35
associate the LPC amplitude effect to decision
confidence than to decision accuracy.
Sometimes, latency jitter has been suggested to
artificially create amplitude differences between mem-
ory conditions (e.g., Spencer et al., 2000). Latency jitter
refers to bthe variability in the latency of the ERP
component across the individual trials in an
experimentQ and is of particular concern when averagedERP deflections seem to be more dpeakedT in one
condition as compared to the other (Spencer et al.,
2000, p. 495). In our experiment, this was not the case
since the width of the P3 for the studied targets seems
highly comparable with that of the repeated targets
(Fig. 1).
The repeated items that were not recognised
elicited a late positive deflection, which characteristics
did not correspond to those of the classic P3
component nor to any of its subcomponents. It had
a frontal rather than centro-parietal maximum, and its
maximum occurred relatively late; around 650 ms (as
compared to 520–540 ms for recognised items) and
after the response had been given (mean RT 580 ms).
It therefore appeared too late to index an implicit
memory or a partial recognition process, as described
by Rugg et al. (1998) and Walla et al. (1999). Instead,
it is more likely that the increased frontal positivity is
a reflection of some kind of postretrieval or decision-
related process. The presence of such process may
have been facilitated by the strong negative response
bias, which consequently also led to a large number of
unintentional misses. These misses presumably origi-
nated from participants pressing the YES button
prematurely while at the same time (or shortly
thereafter) realising that it was the incorrect response.
Nevertheless, even if about half of the trials that
contributed to the ERPs for repeated misses was
elicited by such premature responses (and the other
half by dtrueT misses), it was striking that, for these
items, no earlier positivity was observed that
resembled a P3-related deflection and that could have
been indicative of at least a partial activation of the
relevant memory trace. It can therefore be speculated
that the increased frontal positivity for the repeated
misses indexes some kind of postretrieval evaluation
process but which must have had access to memory
traces that were apparently not capable of attracting
sufficient attention resources to generate the correct
overt recognition response and to elicit a P3 compo-
nent. The frontal maximum of this component
supports this interpretation, since the prefrontal cortex
has been associated with postretrieval monitoring and
verification processes (Rugg et al., 2000).
In conclusion, behavioural and ERP results dem-
onstrated that the ERP-based memory assessment
procedure is sensitive to both study intention and
retrieval performance. Intentional encoding (study) as
compared to incidental encoding (repetition) led to
less errors, quicker responses, and larger P3 ampli-
tudes. Essentially, these were all quantitative differ-
ences suggesting that activation of the created
memory traces depended on similar neural systems.
Repeated items that did not receive a recognition
response could be differentiated from new items based
on their ERPs. This ERP difference is believed to be
associated with a postretrieval evaluation process, but
this needs further investigation.
Acknowledgement
I would like to thank Graham Wade for his help
with data collection.
References
Allen, J.J.B., 2002. The role of psychophysiology in clinical
assessment: ERPs in the evaluation of memory. Psychophysio-
logy 39, 261–280.
Allen, J.J.B., Movius, H.L., 2000. The objective assessment of
amnesia in dissociative identity disorder using event-related
potentials. Int. J. Psychophysiol. 38, 21–41.
Allen, J.J., Iacono, W.G., Danielson, K.D., 1992. The identification
of concealed memories using event related potential and implicit
behavioural measures: a methodology for prediction in the face
of individual differences. Psychophysiology 29, 504–522.
Bentin, S., Moscovitch, M., 1990. Psychophysiological indices of
implicit memory performance. Bull. Psychon. Soc. 28, 346–352.
Bentin, S., Moscovitch, M., Heth, I., 1992. Memory with and
without awareness: performance and electrophysiological evi-
dence of savings. J. Exper. Psychol., Learn., Mem., Cogn. 18,
359–366.
Farwell, L.A., Donchin, E., 1991. The truth will out: interrogative
polygraphy (Lie-detection) with event-related brain potentials.
Psychophysiology 28, 531–547.
Finnigan, S., Humphreys, M.S., Dennis, S., Geffen, G., 2002. ERP
dold/newT effects: memory strength and decisional factor(s).
Neuropsychologia 40, 2288–2304.
Johansson, S., Hofland, K., 1989. Frequency Analysis of English
Vocabulary and Grammar. Claredon Press, Oxford.

J.C. van Hooff / International Journal of Psychophysiology 56 (2005) 25–3636
Kok, A., 1997. Event-related potential (ERP) reflections of mental
resources: a review and synthesis. Biol. Psychol. 45, 19–56.
Kok, A., 2001. On the utility of P3 amplitude as a measure of
processing capacity. Psychophysiology 38, 557–577.
Lalouschek, W., Goldenberg, G., Merterer, A., Beisteiner, R.,
Lindinger, G., Lang, W., 1997. Brain/behaviour dissociation
on old/new distinction in a patient with amnesic syndrome.
Electroencephalogr. Clin. Neurophysiol. 104, 222–227.
McCarthy, G., Wood, C.C., 1985. Scalp distributions of event-related
potentials: an ambiguity associated with analysis of variance
models. Electroencephalogr. Clin. Neurophysiol. 62, 203–208.
Rosenfeld, J.P., Angell, A., Johnson, M., Qian, J., 1991. An ERP-
based, control-question lie detector analog: algorithms for
discriminating effects within individuals’ average waveforms.
Psychophysiology 28, 319–335.
Rosenfeld, J.P., Ellwanger, J., Sweet, J., 1995. Detecting simulated
amnesia with event-related brain potentials. Int. J. Psychophysiol.
19, 1–11.
Rugg, M.D., Mark, R.E., Walla, P., Schloerscheidt, A.M., Birch,
C.S., Allan, K., 1998. Dissociation of the neural correlates of
implicit and implicit memory. Nature 392, 595–598.
Rugg, M.D., Allan, K., Birch, C.S., 2000. Electrophysiological
evidence for the modulation of retrieval orientation by depth of
study processing. J. Cogn. Neurosci. 12, 664–678.
Snodgrass, J.G., Corwin, J., 1988. Pragmatics of measuring
recognition memory: applications to dementia and amnesia.
J. Exp. Psychol. Gen. 117, 34–50.
Spencer, K.M., Dien, J., Donchin, E., 1999. A componential
analysis of the ERP elicited by novel events using a dense
electrode array. Psychophysiology 36, 409–414.
Spencer, K.M., Vila Abad, E., Donchin, E., 2000. On the search for
the neurophysiological manifestation of recollective experience.
Psychophysiology 37, 494–506.
Squires, N.C., Squires, K.C., Hillyard, S.A., 1975. Two varieties of
long-latency positive waves evoked by unpredictable auditory
stimuli in man. Electroencephalogr. Clin. Neurophysiol. 38,
387–401.
van Hooff, J.C., Golden, S., 2002. Validation of an event-related
potential memory assessment procedure: intentional learning as
opposed to simple repetition. J. Psychophysiol. 16, 12–22.
van Hooff, J.C., De Beer, N.A.M., Brunia, C.H.M., Cluitmans,
P.J.M., Korsten, H.H.M., Tavilla, G., Grouls, R., 1995.
Information during cardiac surgery: an event-related potential
study. Electroencephalogr. Clin. Neurophysiol. 96, 433–452.
van Hooff, J.C., Brunia, C.H.M., Allen, J.J.B., 1996a. Event-related
potentials as indirect measures of recognition memory. Int. J.
Psychophysiol. 21, 15–31.
van Hooff, J.C., de Beer, N.A.M., Brunia, C.H.M., Cluitmans,
P.J.M., Korsten, H.H.M., 1996b. Detection of information
processing during general anaesthesia by means of event related
potentials. In: Bonke, B., Bovill, J.G., Moerman, N. (Eds.).
Memory and Awareness in Anaesthesia, vol. III. Van Gorcum,
Assen, NL, pp. 207–218.
Walla, P., Endl, W., Lindinger, G., Deecke, L., Lang, W., 1999.
Implicit memory within a word recognition task: an event-
related potential study in human subjects. Neurosci. Lett. 269,
129–132.





























