Embed Size (px)
Citation preview

Planta (1994)195:210 216 P l ~ _ _ t ~
~ Springer-Verlag 1994
The induction of the CO2-concentrating mechanism is correlated with the formation of the starch sheath around the pyrenoid of Chlamydomonas reinhardtii Ziyadin Ramazanov ~, Mamta Rawat ~, Margaret C. Henk 2, Catherine B. Mason 1, Sharon W. Matthews ~, James V. Moroney 1
Department of Botany, Louisiana State University, Baton Rouge, LA 70803, USA 2 Department of Microbiology, Louisiana State University, Baton Rouge, LA 70803, USA
Received: 12 April 1994 Accepted: 29 April 1994
Abstract. The pyrenoid is a prominent proteinaceous structure found in the stroma of the chloroplast in unicel- lular eukaryotic algae, most multicellular algae, and some hornworts. The pyrenoid contains the enzyme ribu- lose-l,5-bisphosphate carboxylase/oxygenase and is sometimes surrounded by a carbohydrate sheath. We have observed in the unicellular green alga Chlamy- domonas reinhardtii Dangeard that the pyrenoid starch sheath is formed rapidly in response to a decrease in the CO2 concentration in the environment. This formation of the starch sheath occurs coincidentally with the induc- tion of the CO2-concentrating mechanism. Pyrenoid starch-sheath formation is partly inhibited by the pres- ence of acetate in the growth medium under light and low-CO2 conditions. These growth conditions also partly inhibit the induction of the CO2-concentrating mecha- nism. When cells are grown with acetate in the dark, the CO2-concentrating mechanism is not induced and the pyrenoid starch sheath is not formed even though there is a large accumulation of starch in the chloroplast stroma. These observations indicate that pyrenoid starch-sheath formation correlates with induction of the CO2-concen- trating mechanism under low-CO 2 conditions. We sug- gest that this ultrastructural reorganization under low- CO 2 conditions plays a role in the CO2-concentrating mechanism C. reinhardtii as well as in other eukaryotic algae.
Key words: Chlamydomonas Carbon-dioxide-concen- trating mechanism Photosynthesis - Pyrenoid - Starch
Introduction
The pyrenoid of algal and hornwort chloroplasts is a large protein complex sometimes surrounded by a sheath
Abbreviations: Low CO2=air containing ambient 0.03% CO2 (v/ v); high CO2=air supplemented with CO2 so that the final CO2 concentration is 5% (v/v); Rubisco- ribulose-1,5-bisphosphate car- boxylase/oxygenase Correspondence to: James V. Moroney; FAX: I (504) 388-8459; Tel: 1 (504) 388-8561; E-mail: [email protected]
of carbohydrate such as starch, amylose or paramylon (Gibbs 1962; Griffiths 1980; Kuchitsu et al. 1988; Okada 1992). It is located in the chloroplast stroma and is often traversed by thylakoid lamellae (Gibbs 1962; McKay and Gibbs 1991). At the-ul t rastructural level, the pyrenoid is a prominent electron-dense body that under- goes division during mitosis. In the past, a number of physiological roles have been suggested for the pyrenoid, including functions in starch biosynthesis or as a protein- storage body (Griffiths 1980; McKay and Gibbs 1991; Okada 1992). These suggested roles have been challenged by immunolocalization and biochemical studies that have shown that most, if not all, of the ribulose-l,5-bis- phosphate carboxylase/oxygenase (Rubisco) in photoau- totrophically growing cells is localized in the pyrenoid (Vladimirova et al. 1982; Lacoste-Royal and Gibbs 1987; Osafune et al. 1990; Kuchitsu et al. 1991; McKay and Gibbs 1991; Okada 1992), therefore indicating that the pyrenoid is the active location of CO2 fixation and not simply a storage structure.
Light-microscopy studies have shown that ceils adapted to low-CO 2 conditions are more likely to have starch around the pyrenoid than cells adapted to high- CO 2 conditions (Miyachi et al. 1986; Kuchitsu et al. 1988; Kuchitsu et al. 1991). In contrast to most higher plants, unicellular algae grown under l o w - C O 2 conditions have developed a means of concentrating CO 2 at the site of Rubisco (Badger et al. 1980). Cells with the CO2-concen- trating mechanism (CCM) acquire the ability to accumu- late inorganic carbon (CO 2 and HCO3) to levels higher than can be obtained by simple diffusion. Algae with the CCM can grow on very low CO 2 concentrations, lower than that tolerated by plants with C3-type photosynthe- sis. The CCM results in an increase in internal CO2 con- centration, which favors the carboxylation reaction of Rubisco over the competitive oxygenation reaction (Bad- ger et al. 1980; Aizawa and Miyachi 1986). In Chlamy- domonas reinhardtii, as well as Chlorella, Scenedesmus and Dunaliella species, the CCM is inducible; only algal cells grown on air-levels of CO 2 (0.03%) have an active CCM (Badger et al. 1980; Aizawa and Miyachi 1986). If algal cells are grown on elevated CO 2 concentrations (1% CO 2 or higher), they have a relatively low affinity for
z. Ramazanov et al.: Pyrenoid starch-sheath formation in Chlamydomonas 211
CO2, a n d pho tosyn thes i s is s t rong ly inh ib i ted by 02 (Badger et al. 1980; Spa ld ing et al. 1983). However , if the algal cells are p laced in an env i ronmen t where CO2 is l imit ing, wi th in 4 h these cells induce a pe r ip lasmic car- bonic a n h y d r a s e as well as four add i t i ona l p ro te ins of u n k n o w n funct ion (Manue l and M o r o n e y 1988), they ex- h ibi t very high affinities for CO2, and their p h o t o s y n t h e - sis is no t a p p r e c i a b l y inh ib i ted by a t m o s p h e r i c levels of 02 (Badger et al. 1980; A i z a w a and Miyach i 1986).
The C C M has been found in a large n u m b e r of aqua t - ic o rgan i sms (Aizawa and M i y a c h i 1986). A m o n g the eu- k a r y o t i c algae, the presence of a p y r e n o i d in a species cor re la tes well with the abi l i ty to induce a C C M in tha t a lgal species. A c c o r d i n g to Kuch i t su et al. (1991) the con- tent of Rub i sco in the p y r e n o i d is h igher in l o w - C O 2- g rown cells t han in h igh -COz-g rown cells of C. rein- hardtii. The loca l i za t ion of Rub i sco wi thin the p y r e n o i d and the f o r m a t i o n of the s ta rch shea th specifical ly a r o u n d the p y r e n o i d under l o w - C O 2 cond i t ions have ra ised the ques t ions of (i) the role of the py reno id in the funct ioning of the C C M a n d (ii) the co r re l a t ion be tween induc t ion of C C M and p y r e n o i d s t a rch - shea th forma- t ion.
The p a c k a g i n g of Rubisco in the pho tosyn the t i c cell might be i m p o r t a n t for the o p e r a t i o n of a C C M . F o r example , in c y a n o b a c t e r i a where Rub i sco is p a c k a g e d in c a r b o x y s o m e s ( M c K a y et al. 1993), the n u m b e r of car- b o x y s o m e s was h igher in l ow-COz-g rown cells (Turpin et al. 1984). These p h o t o s y n t h e t i c bac te r i a also possess a C C M , a n d recent ly ca rbon ic a n h y d r a s e has been local- ized with Rub i sco in the c a r b o x y s o m e (Price et al. 1992). Since c y a n o b a c t e r i a act ively t ake up b i c a r b o n a t e f rom the e n v i r o n m e n t (Mil ler et al. 1990), the loca l i za t ion of b o t h ca rbon ic a n h y d r a s e and Rub i sco in the car- b o x y s o m e w o u l d favor the gene ra t ion of CO 2 at the site of Rub i sco (Mil ler et al. 1990; R e i n h o l d et al. 1991; Price et el. 1992). Potent ia l ly , the p y r e n o i d could p l ay a s imi lar role in euka ryo t i c algae.
In this repor t , we have inves t iga ted the effect of exter- nal CO2 concen t r a t i ons on the f o r m a t i o n of the p y r e n o i d s ta rch shea th in wi ld - type C. reinhardtii cells. Transmis - s ion e lec t ron m i c r o s c o p y and phys io log ica l measu re - ments indica te tha t in C. reinhardtii the synthesis of the p y r e n o i d s ta rch shea th cor re la tes closely wi th the ap- pea rance of the C C M .
M a t e r i a l s a n d m e t h o d s
Algal culture conditions. Chlamydomonas reinhardtii 137 wild type was obtained from Dr. R.K. Togasaki, Indiana University, and maintained in this laboratory. In liquid culture, the strains were inoculated at a cell density of 4-104 cells mL ~ and were grown in minimal medium, aerated with 5% CO2 in air and continuously illuminated with 300 lamol, m 2. s 1 of white light for 2 d. The cul- tures were then switched to synchronous growth by using a 12 h light/12 h dark cycle for 3 d and maintained at similar cell densities by daily dilution with fresh medium.
Electron microscopy. For transmission electron microscopy, two methods of fixation and embedding were used. In the first proce- dure, cells were fixed for 2 h at 4~ in 2.5% glutaraldehyde added to Hepes-KOH (pH 7.5) buffer. Samples were rinsed in the same
buffer, then postfixed overnight at 4~ in 1% osmium tetroxide. After a rinse in distilled water, the samples were dehydrated in a graded series of ethanol, with 2% uranyl acetate added to the 80% step, and held overnight at 4~ to a final concentration of 100% ethanol. Samples were pelleted by centrifugation between each change. The samples were slowly infiltrated at 4~ with Spurr's resin (Spurr 1969), with seven changes over a period of several days, then polymerized for 12 h at 60~
In the second procedure, cells were fixed in a mixture of 2% glutaraldehyde and 2% osmium tetroxide in half-strength growth medium for 15 min. The cells were then filtered and further fixed in the same fixative mixture in 0.05 M cacodylate buffer (pH 7.1), for 20 min, rinsed for 5 rain, stained en bloc with 0.5% uranyl acetate for 30 min, dehydrated in ethanol, then infiltrated and embedded in LR White medium resin (London Resin Co., Woking, Surrey, UK). Thin sections were cut using a diamond knife mounted on a Sorvall Porter-Blum (Norwalk, Conn., USA) MT-2 or MT 5000 ultramicro- tome, poststained with uranyl acetate and or lead citrate, and exam- ined and photographed on a JEOL (Tokyo, Japan) JEM 100CX transmission electron microscope.
For estimation of the percentage of cells containing complete pyrenoid starch sheaths, cells from well-separated thin sections were individually inspected. Only cells where a clear pyrenoid was visible were counted. If the starch sheath covered more than 95% of the pyrenoid core that cell was considered to have a complete starch sheath.
Photosynthesis assays. The photosynthetic rate of algal cells was measured with an oxygen electrode (Rank Brothers, Cambridge, England). Algae were centrifuged at 4,400.g for 5 min and the pel- leted algae were resuspended at 25 lag chlorophyll, mL l in 4 ml of 25 mM Hepes-KOH (pH 7.3) and transferred to the electrode cham- ber, where they were allowed to consume the inorganic carbon of the buffer and intracellular pool of inorganic carbon (CO2 + HCO3) until no net Oz exchange was observed. Bicarbonate at the indicat- ed concentrations was added and the rate of 02 evolution mea- sured.
Other methods. Chlorophyll concentrations were determined spec- trophotometrically. The immunoblot assay was performed accord- ing to the protocol from Bio-Rad Laboratories (Richmond, Calif.,
250
"r
"F 200
'7 o3 E _ 150 0
mO 100 _= 0
so
0 1 2 3 4 5
0 , , T i m e , h
0 1 2 3 4 5
NaHCO 3 , mM
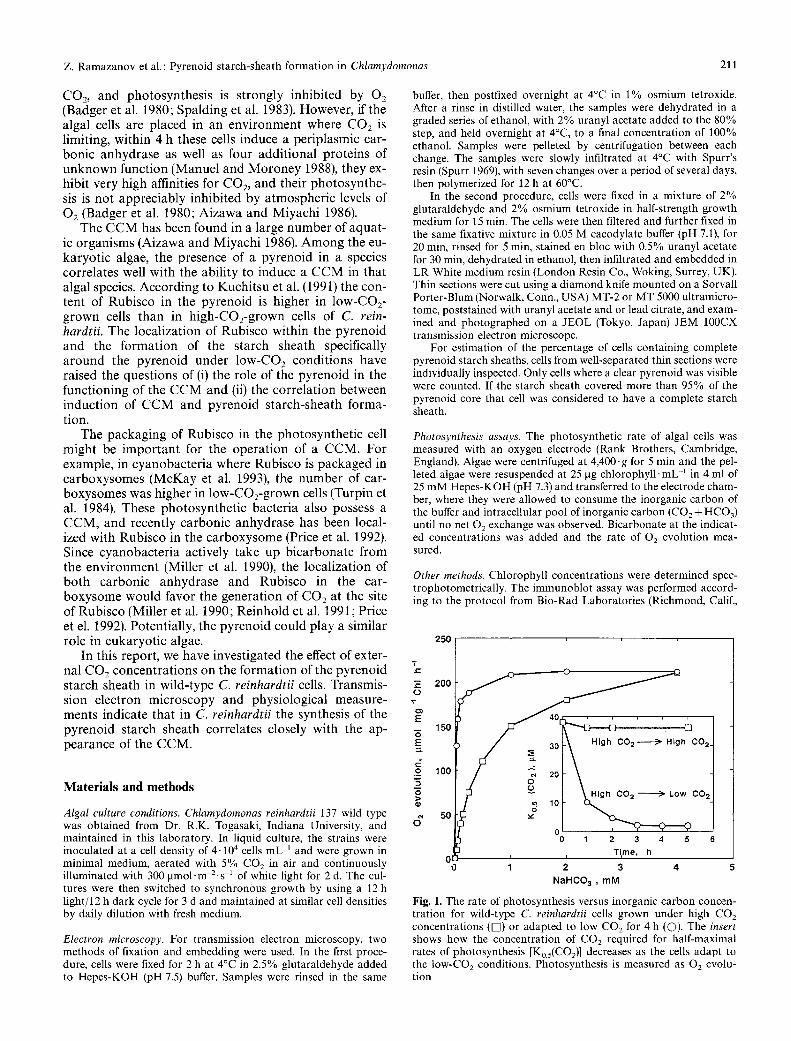
Fig. 1. The rate of photosynthesis versus inorganic carbon concen- tration for wild-type C. reinhardtii cells grown under high CO2 concentrations (IN) or adapted to low COa for 4 h (C)). The insert shows how the concentration of CO2 required for half-maximal rates of photosynthesis [Ko.5(CO2) ] decreases as the cells adapt to the low-CO2 conditions. Photosynthesis is measured as 02 evolu- tion
212 Z. Ramazanov et al.: Pyrenoid starch-sheath formation in Chlamydomonas
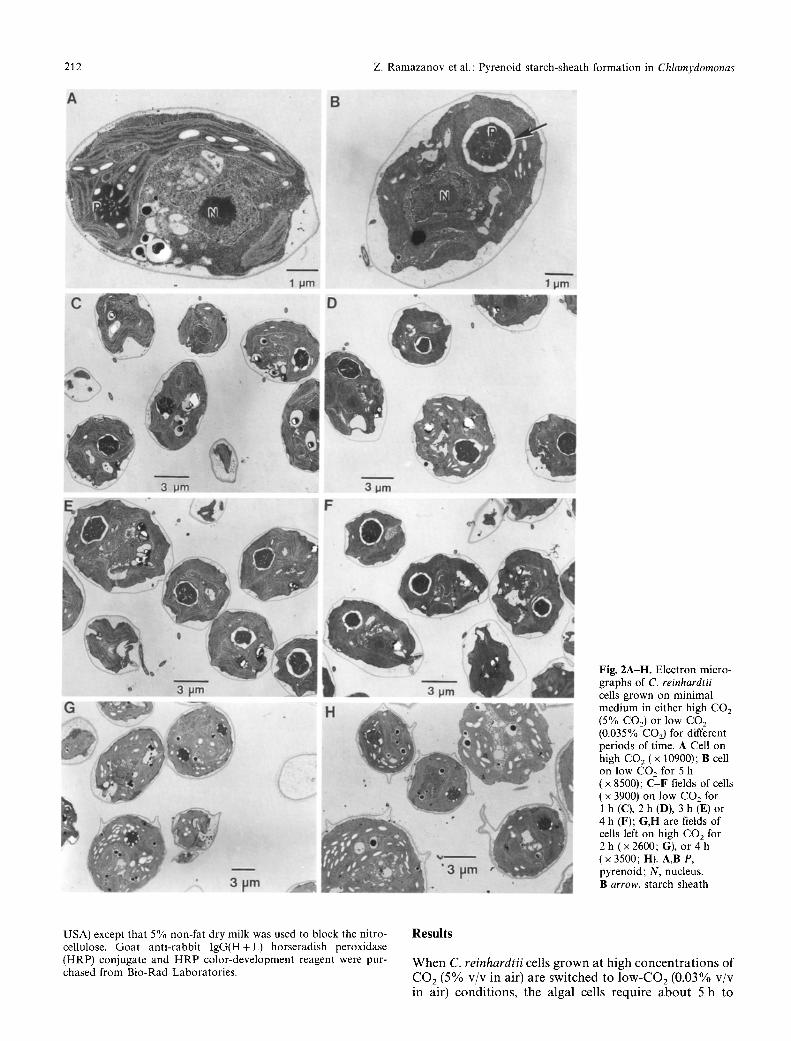
Fig. 2A-H. Electron micro- graphs of C. reinhardtii cells grown on minimal medium in either high CO2 (5% CO2) or low C O 2
(0.035% CO2) for different periods of time. A Cell on high CO 2 ( x 10900); B cell on low CO 2 for 5 h ( x 8500); C-F fields of cells ( x 3900) on low CO2 for 1 h (C), 2 h (D), 3 h (E) or 4 h (F); G,H are fields of cells left on high CO2 for 2 h (x 2600; G), or 4 h ( • 3500; H). A,B P, pyrenoid; N, nucleus. B arrow, starch sheath
USA) except that 5% non-fat dry milk was used to block the nitro- cellulose. Goat anti-rabbit IgG(H+L) horseradish peroxidase (HRP) conjugate and HRP color-development reagent were pur- chased from Bio-Rad Laboratories.
Resul ts
W h e n C. reinhardtii cells g r o w n at h igh c o n c e n t r a t i o n s of CO2 (5% v/v in air) are swi tched to low-CO2 (0 .03% v/v in air) cond i t ions , the algal cells requi re a b o u t 5 h to
Z. Ramazanov et al.: Pyrenoid starch-sheath formation in Chlamydomonas 213
25
2O
o ~ is
~g lO
5
0 0
/ 8~
7 lqrr~, h
l I I I I I
12 24 36 48 60 72
Time, h
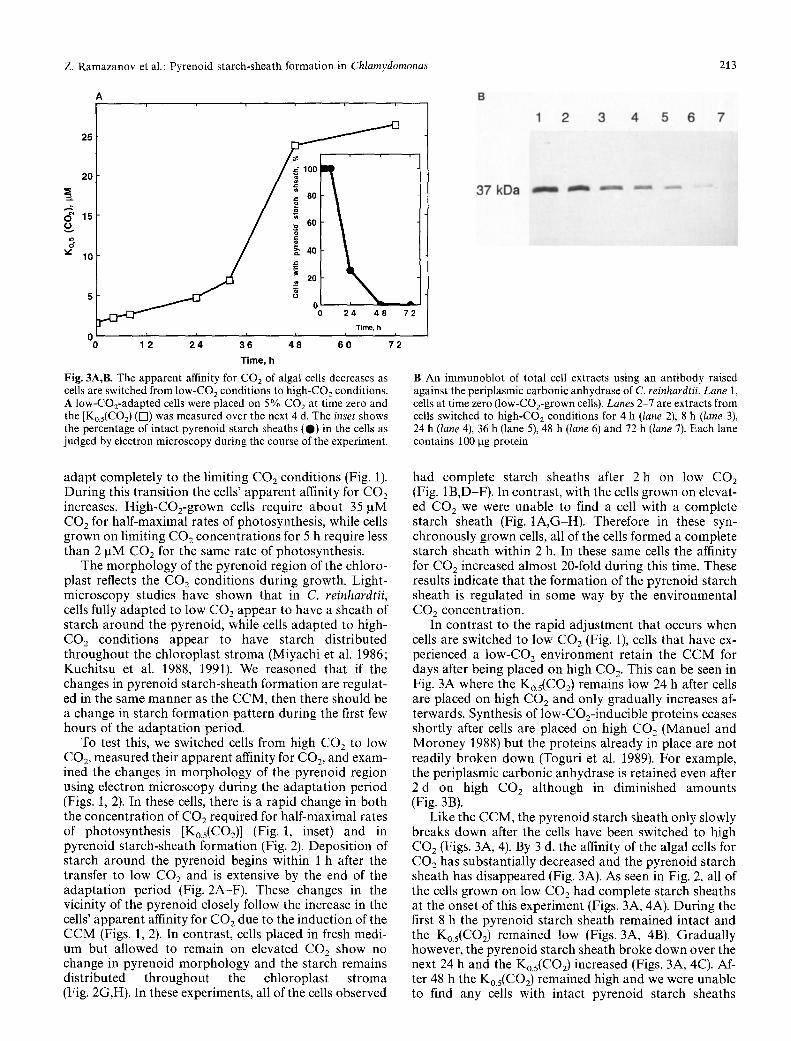
Fig. 3A,B. The apparent affinity for CO2 of algal cells decreases as cells are switched from low-CO 2 conditions to high-CO2 conditions. A low-COz-adapted cells were placed on 5% COz at time zero and the [Ko.5(CO2) ([]) was measured over the next 4 d. The inset shows the percentage of intact pyrenoid starch sheaths (@) in the cells as judged by electron microscopy during the course of the experiment.
adapt completely to the limiting C O 2 conditions (Fig. 1). During this transition the cells' apparent affinity for CO2 increases. High-CO2-grown cells require about 35 gM CO2 for half-maximal rates of photosynthesis, while cells grown on limiting CO2 concentrations for 5 h require less than 2 laM CO2 for the same rate of photosynthesis.
The morphology of the pyrenoid region of the chloro- plast reflects the CO2 conditions during growth. Light- microscopy studies have shown that in C. reinhardtii, cells fully adapted to low CO2 appear to have a sheath of starch around the pyrenoid, while cells adapted to high- CO2 conditions appear to have starch distributed throughout the chloroplast stroma (Miyachi et al. 1986; Kuchitsu et al. 1988, 1991). We reasoned that if the changes in pyrenoid starch-sheath formation are regulat- ed in the same manner as the CCM, then there should be a change in starch formation pattern during the first few hours of the adaptation period.
To test this, we switched cells from high CO 2 to low CO2, measured their apparent affinity for CO2, and exam- ined the changes in morphology of the pyrenoid region using electron microscopy during the adaptation period (Figs. 1, 2). In these cells, there is a rapid change in both the concentration of CO2 required for half-maximal rates of photosynthesis [Ko.5(CO2) ] (Fig. 1, inset) and in pyrenoid starch-sheath formation (Fig. 2). Deposition of starch around the pyrenoid begins within 1 h after the transfer to low CO 2 and is extensive by the end of the adaptation period (Fig. 2A-F). These changes in the vicinity of the pyrenoid closely follow the increase in the cells' apparent affinity for CO 2 due to the induction of the CCM (Figs. 1, 2). In contrast, cells placed in fresh medi- um but allowed to remain on elevated CO2 show no change in pyrenoid morphology and the starch remains distributed throughout the chloroplast stroma (Fig. 2G,H). In these experiments, all of the cells observed
B An immunoblot of total cell extracts using an antibody raised against the periplasmic carbonic anhydrase of C. reinhardtii. Lane l, cells at time zero (low-CO2-grown cells). Lanes 2-7 are extracts from cells switched to high-CO2 conditions for 4 h (lane 2), 8 h (lane 3), 24 h (lane 4), 36 h (lane 5), 48 h (lane 6) and 72 h (lane 7). Each lane contains 100 gg protein
had complete starch sheaths after 2h on low CO2 (Fig. 1 B,D-F). In contrast, with the cells grown on elevat- ed CO 2 we were unable to find a cell with a complete starch sheath (Fig. 1A,G-H). Therefore in these syn- chronously grown cells, all of the cells formed a complete starch sheath within 2 h. In these same cells the affinity for CO 2 increased almost 20-fold during this time. These results indicate that the formation of the pyrenoid starch sheath is regulated in some way by the environmental CO2 concentration.
In contrast to the rapid adjustment that occurs when cells are switched to low CO2 (Fig. 1), cells that have ex- perienced a l o w - C O 2 environment retain the CCM for days after being placed on high CO 2. This can be seen in Fig. 3A where the Ko.5(CO2) remains low 24 h after cells are placed on high CO2 and only gradually increases af- terwards. Synthesis of low-CO2-inducible proteins ceases shortly after cells are placed on high CO 2 (Manuel and Moroney 1988) but the proteins already in place are not readily broken down (Toguri et al. 1989). For example, the periplasmic carbonic anhydrase is retained even after 2 d on high CO2 although in diminished amounts (Fig. 3B).
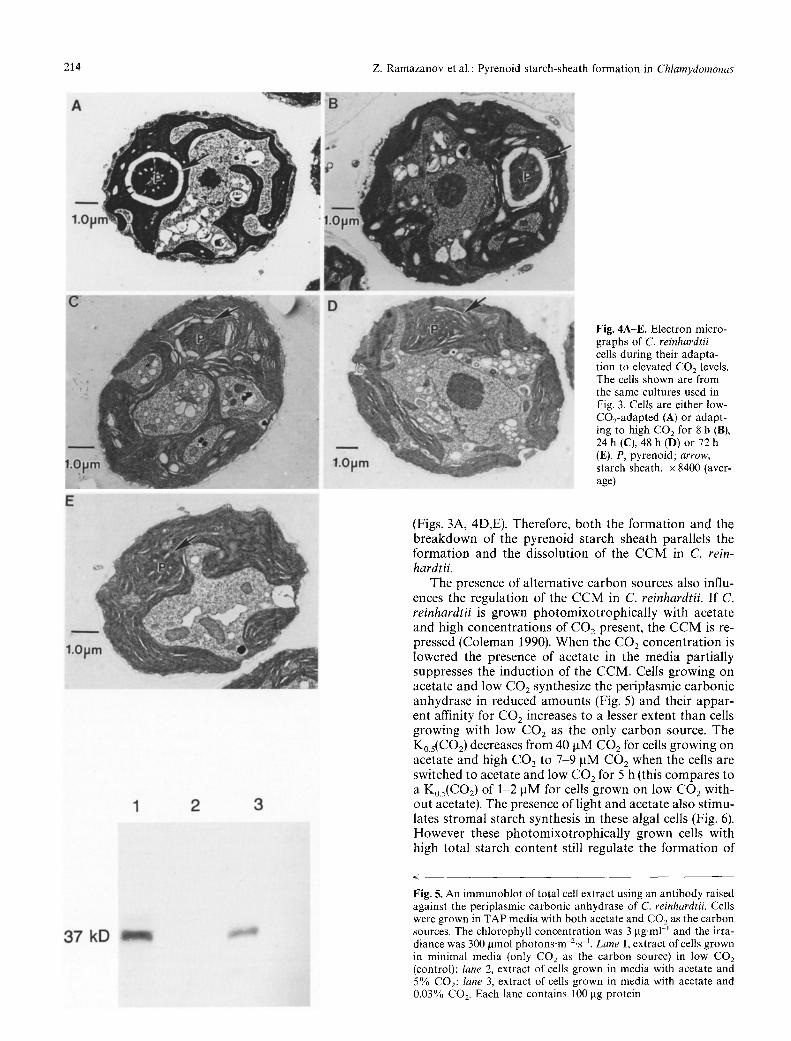
Like the CCM, the pyrenoid starch sheath only slowly breaks down after the cells have been switched to high CO2 (Figs. 3A, 4). By 3 d, the affinity of the algal cells for CO2 has substantially decreased and the pyrenoid starch sheath has disappeared (Fig. 3A). As seen in Fig. 2, all of the cells grown on low CO2 had complete starch sheaths at the onset of this experiment (Figs. 3A, 4A). During the first 8 h the pyrenoid starch sheath remained intact and the Ko.5(CO2) remained low (Figs. 3A, 4B). Gradually however, the pyrenoid starch sheath broke down over the next 24 h and the K0.5(C02) increased (Figs. 3A, 4C). Af- ter 48 h the K0.5(CO2) remained high and we were unable to find any cells with intact pyrenoid starch sheaths
214 Z. Ramazanov et al.: Pyrenoid starch-sheath formation in Chlamydomonas
Fig. 4A-E. Electron micro- graphs of C. reinhardtii cells during their adapta- tion to elevated CO2 levels. The cells shown are from the same cultures used in Fig. 3. Cells are either low- CO2-adapted (A) or adapt- ing to high CO 2 for 8 h (B), 24 h (C), 48 h (D) or 72 h (E). P, pyrenoid; arrow, starch sheath. • 8400 (aver- age)
(Figs. 3A, 4D,E). Therefore, both the format ion and the b reakdown of the pyrenoid starch sheath parallels the format ion and the dissolution of the C C M in C. rein- hardtii.
The presence of alternative ca rbon sources also influ- ences the regulation of the C C M in C. reinhardtii. If C. reinhardtii is grown photomixot rophica l ly with acetate and high concentra t ions of CO2 present, the C C M is re- pressed (Coleman 1990). When the CO2 concentra t ion is lowered the presence of acetate in the media partially suppresses the induct ion of the CCM. Cells growing on acetate and low CO2 synthesize the periplasmic carbonic anhydrase in reduced amounts (Fig. 5) and their appar- ent affinity for CO 2 increases to a lesser extent than cells growing with low CO 2 as the only ca rbon source. The K0.5(C02) decreases f rom 40 txM CO2 for cells growing on acetate and high CO 2 to 7-9 gM CO2 when the cells are switched to acetate and low CO2 for 5 h (this compares to a K0.5(CO2) of 1-2 laM for cells grown on low CO 2 with- out acetate). The presence of light and acetate also stimu- lates s tromal starch synthesis in these algal cells (Fig. 6). However these photomixot rophica l ly grown cells with high total starch content still regulate the format ion of
Fig. 5. An immunoblot of total cell extract using an antibody raised against the periplasmic carbonic anhydrase of C. reinhardtii. Cells were grown in TAP media with both acetate and CO2 as the carbon sources. The chlorophyll concentration was 3 gg.ml 1 and the irra- diance was 300 lamol photons.m 2.s ~. Lane 1, extract of cells grown in minimal media (only CO2 as the carbon source) in low CO2 (control); lane 2, extract of cells grown in media with acetate and 5% CO2; lane 3, extract of cells grown in media with acetate and 0.03% CO2. Each lane contains 100 gg protein
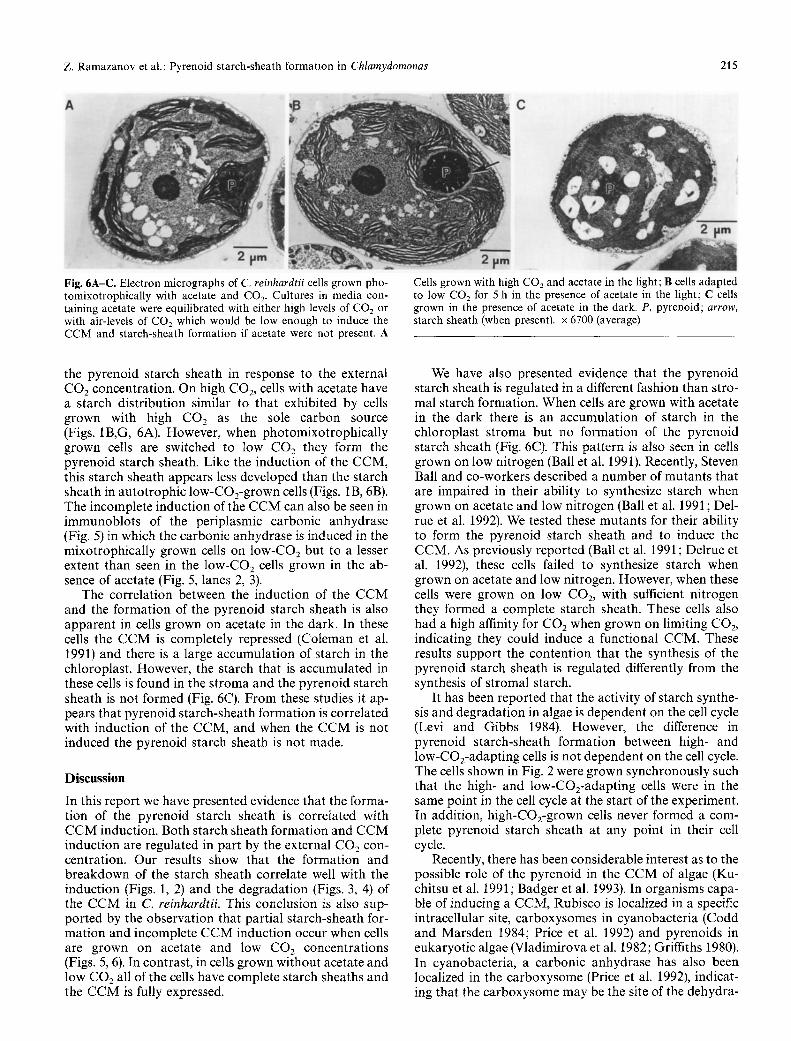
Z. Ramazanov et al.: Pyrenoid starch-sheath formation in Chlamydomonas 215
Fig. 6A-C. Electron micrographs of C. reinhardtii ceils grown pho- tomixotrophically with acetate and CO2. Cultures in media con- taining acetate were equilibrated with either high levels of CO2 or with air-levels of CO2 which would be low enough to induce the CCM and starch-sheath formation if acetate were not present. A
Cells grown with high CO2 and acetate in the light; B cells adapted to low CO2 for 5 h in the presence of acetate in the light; C cells grown in the presence of acetate in the dark. P, pyrenoid; arrow, starch sheath (when present), x 6700 (average)
the pyrenoid starch sheath in response to the external CO2 concentration. On high CO2, cells with acetate have a starch distribution similar to that exhibited by ceils grown with high CO 2 as the sole carbon source (Figs. 1B,G, 6A). However, when photomixotrophically grown cells are switched to low CO 2 they form the pyrenoid starch sheath. Like the induction of the CCM, this starch sheath appears less developed than the starch sheath in autotrophic low-COz-grown cells (Figs. 1B, 6B). The incomplete induction of the CCM can also be seen in immunoblots of the periplasmic carbonic anhydrase (Fig. 5) in which the carbonic anhydrase is induced in the mixotrophically grown cells on low-CO2 but to a lesser extent than seen in the low-CO2 cells grown in the ab- sence of acetate (Fig. 5, lanes 2, 3).
The correlation between the induction of the CCM and the formation of the pyrenoid starch sheath is also apparent in cells grown on acetate in the dark. In these cells the CCM is completely repressed (Coleman et al. 1991) and there is a large accumulation of starch in the chloroplast. However, the starch that is accumulated in these cells is found in the stroma and the pyrenoid starch sheath is not formed (Fig. 6C). From these studies it ap- pears that pyrenoid starch-sheath formation is correlated with induction of the CCM, and when the CCM is not induced the pyrenoid starch sheath is not made.
Discussion
In this report we have presented evidence that the forma- tion of the pyrenoid starch sheath is correlated with CCM induction. Both starch sheath formation and CCM induction are regulated in part by the external CO 2 con- centration. Our results show that the formation and breakdown of the starch sheath correlate well with the induction (Figs. 1, 2) and the degradation (Figs. 3, 4) of the C C M in C. reinhardtii. This conclusion is also sup- ported by the observation that partial starch-sheath for- mation and incomplete CCM induction occur when cells are grown on acetate and low CO 2 concentrations (Figs. 5, 6). In contrast, in cells grown without acetate and low CO 2 all of the cells have complete starch sheaths and the CCM is fully expressed.
We have also presented evidence that the pyrenoid starch sheath is regulated in a different fashion than stro- mal starch formation. When cells are grown with acetate in the dark there is an accumulation of starch in the chloroplast stroma but no formation of the pyrenoid starch sheath (Fig. 6C). This pattern is also seen in cells grown on low nitrogen (Ball et al. 1991). Recently, Steven Ball and co-workers described a number of mutants that are impaired in their ability to synthesize starch when grown on acetate and low nitrogen (Ball et al. 1991; Del- rue et al. 1992). We tested these mutants for their ability to form the pyrenoid starch sheath and to induce the CCM. As previously reported (Ball et al. 1991; Delrue et al. 1992), these cells failed to synthesize starch when grown on acetate and low nitrogen. However, when these cells were grown on low CO2, with sufficient nitrogen they formed a complete starch sheath. These cells also had a high affinity for CO2 when grown on limiting CO2, indicating they could induce a functional CCM. These results support the contention that the synthesis of the pyrenoid starch sheath is regulated differently from the synthesis of stromal starch.
It has been reported that the activity of starch synthe- sis and degradation in algae is dependent on the cell cycle (Levi and Gibbs 1984). However, the difference in pyrenoid starch-sheath formation between high- and low-COz-adapting cells is not dependent on the cell cycle. The cells shown in Fig. 2 were grown synchronously such that the high- and low-CO2-adapting cells were in the same point in the cell cycle at the start of the experiment. In addition, high-CO2-grown ceils never formed a com- plete pyrenoid starch sheath at any point in their cell cycle.
Recently, there has been considerable interest as to the possible role of the pyrenoid in the CCM of algae (Ku- chitsu et al. 1991 ; Badger et al. 1993). In organisms capa- ble of inducing a CCM, Rubisco is localized in a specific intracellular site, carboxysomes in cyanobacteria (Codd and Marsden 1984; Price et al. 1992) and pyrenoids in eukaryotic algae (Vladimirova et al. 1982; Griffiths 1980). In cyanobacteria, a carbonic anhydrase has also been localized in the carboxysome (Price et al. 1992), indicat- ing that the carboxysome may be the site of the dehydra-

216 Z. Ramazanov et al.: Pyrenoid starch-sheath formation in Chlamydomonas
t ion of accumula ted b ica rbona te (Reinhold et al. 1991). Transformed cyanobac te r ia that express carbonic anhy- drase in their cy toplasm c a n n o t grow on low CO 2, pre- sumably because the accumula ted b ica rbona te is dehy- drated at an inappropr ia te in t racel lu lar location, short- circui t ing the C C M (Price and Badger 1989). It has also been proposed that the pro te in shell that su r rounds the ca rboxysome might be a barr ier to diffusion of CO, gen- erated within the carboxysome by the dehydra t ion of H C O 3 (Reinhold et al. 1991).
We propose that the pyrenoid may serve as the site where accumula ted b ica rbona te is preferentially dehy- drated to CO 2 in C. reinhardtii as well as other eukaryot ic algae. This react ion would generate an elevated CO 2 con- cen t ra t ion in the pyrenoid, the locat ion of Rubisco. In this model, the starch sheath su r round ing the Rubisco might aid in ca rboxyla t ion by slowing the diffusion of CO2 out of the pyrenoid. The format ion of the starch sheath in response to low CO 2 is consistent with this role for the pyreno id starch, but such a rote remains to be tested. The observa t ion that chloroplasts isolated from low-CO2-grown cells retain the abil i ty to accumula te in- organic ca rbon suppor ts the idea that the chloroplast plays an i m p o r t a n t role in the C C M (Moroney et al. 1987; Moroney and Mason 1991). The increased CO2 in the pyrenoid would favor the carboxyla t ion react ion of Rubisco over the compet ing oxygenat ion react ion and therefore increase photosynthes is and product ivi ty in those cells possessing a CO2-concentra t ing mechanism. It is no tab le that the pyrenoid is widely found in aquat ic photosynthe t ic organisms and may represent an adapta- t ion by these plants to the l imit ing CO2 condi t ions found in m a n y aquat ic env i ronments .
The authors thank Dr. David J. Longstreth and Dr. Patricia M. Moroney for critically reading the manuscript. This work was sup- ported by National Science Foundation grants IBN-8957037 and IBN-9304662.
References
Aizawa, K., Miyachi, S. (1986) Carbonic anhydrase and CO2 concentrat- ing mechanisms in microalgae and cyanobacteria. FEMS Micro. Rev. 39, 215-233
Badger, M.R., Kaplan, A., Berry, J.A. (1980) Internal inorganic carbon pool of Chlamydomonas reinhardtii: Evidence for a carbon dioxide concentrating mechanism. Plant Physiol. 66, 407-413
Badger, M.R., Pfanz, H., Bfidel, B., Heber, U., Lange, O.L. (1993) Evi- dence for the functioning of photosynthetic CO2-concentrating mechanism in lichens containing green algal and cyanobacterial photobionts. Planta 191, 57-70
Ball, S., Marianne, T., Dirick, L., Fresnoy, M., Delrue, B., Decq, A. (1991) A Chlamydomonas reinhardtii tow-starch mutant is defective for 3- phosphoglycerate activation and orthophosphate inhibition of ADP-glucose pyrophosphorylase. Planta 185, 17-26
Codd, G.A., Marsden, W.J.N. (1984) The carboxysomes (polyhedral bod- ies) of autotrophic prokaryotes. Biol, Rev. 59, 389-422
Coleman, J.R. (1990) The molecular biology of carbonic anhydrase ex- pression in Chlamydomonas reinhardtii. In: Current research in pho- tosynthesis, vol. IV, pp. 455-462, M. Baltscheffsky, ed, Kluwer Aca- demic Publishers, Dordrecht
Coleman, J.R., Luinenburg, I., Majeau, N., Provart, N. (1991) Sequence analysis and regulation of expression of a gene coding for carbonic anhydrase in Chlamydomonas reinhardtii. Can. J. Bot. 69, 1097-1102
Delrue, B., Fontaine, T., Routier, F., Decq, A., Wieruszeski, J.-M., van den Koornhuyse, N. Maddelein, M.-L., Fournet, B., Ball S. (1992) Waxy Chlamydomonas reinhardtii: Monocellular algal mutants de- fective in amytose biosynthesis and granule-bound starch synthase
activity accumulate a structurally modified amylopectin. J. BacterioL 174, 3612 3620
Gibbs, S.P. (1962) The ultrastructure of the pyrenoids of green algae. L Ultrastruct. Res. 74 262-272
Griffiths, D.J. (1980) The pyrenoid and its role in algal metabolism. Sci. Progr. 66, 537 553
Kuchitsu, K., Tsuzuki, M., Miyachi, S. (1988) Changes of starch localiza- tion within the chloroplast induced by changes in carbon dioxide concentration during growth of Chlamydomonas reinhardtii: inde- pendent regulation of pyrenoid starch and stroma starch. Plant Cell Physiol. 29, 1269-1278
Kuchitsu, K., Tsuzuki, M., Miyachi, S. (1991) Polypeptide composition and enzyme activities of the pyrenoid and its regulation by CO 2 concentration in unicellular green alga. Can. J. Bot. 69, 1062-1069
Lacoste-Royal, G., Gibbs, S.P. (1987) lmmnnocytochemical localization of ribulose 1,5-bisphosphate carboxylase in the pyrenoid and thy- lakoid region of the chloroplast of Chlamydomonas reinhardtii. Plant Physiol. 83, 602-606
Levi, C., Gibbs, M. (1984) Starch degradation in synchronously grown Chlamydomonas reinhardtii and characterization of the amylase. Plant Physiol. 74, 459-463
Manuel, L.J., Moroney, J.V. (1988) Inorganic carbon accumulation in Chlamydomonas reinhardtii: new proteins are made during adaption to tow CO 2. Plant Physiol. 88, 491--496
McKay, R.M.L., Gibbs, S.P. (1991) Composition and function of pyrenoids: cytochemical and immunocytochemical approaches. Can. J. Bot. 69, 1040-1052
McKay, R.M.L., Gibbs, S.P., Espie, G.S. (1993) Effect of dissolved inor- ganic carbon on the expression of carboxysomes localization of Ru- bisco and the mode of inorganic carbon transport in cells of the cyanobacterium Synechococcus UTEX 625. Arch. Microbiol. 159, 21 29
Miller, A.G., Espie, G.S, Canvin, D.T. (1990) Physiological aspects of CO2 and HCO3 transport by cyanobacteria; a review. Can. J. Bet. 68, 1291- 1302
Miyachi, S., Tsuzuki, M., Maruyama, I., Gantar, M., Miyachi, S., Mat- sushima, H. (1986) Effects of CO 2 concentration during growth on the intracellular structure of Chlorella and Scenedesmus (Chlorophy- ta). J. Phycol. 22, 313 319
Moroney, J.V., Kitayama, M., Togasaki, R.K., Tolbert, N.E. (1987) Evi- dence for inorganic carbon transport by intact chloroplasts of Chlamydomonas reinhardtii. Plant Physiol. 83, 460-463
Moroney, J.V., Mason, C.B. (1991) The role of the chloroplast in inorgan- ic carbon acquisition by Chtamydomonas reinhardtii. Can. J. Bot. 69, 1017 1024
Okada, M. (1992) Recent studies on the composition and the activity of algal pyrenoids. Progr. PhycoL Res. 8, 117-138
Osafune, "12, Yokota, A., Sumida, S., Hase, E. (1990) Immunogold local- ization of ribulose-l,5-bisphosphate carboxylase with reference to pyrenoid morphology in chloroplasts of synchronized Euglena gra- ellis cells. Plant Physiol. 92, 802-808
Price, G.D., Badger, M.R. (1989) Expression of human carbonic anhy- drase in the cyanobacterium Synechococcus PCC 7942 creates a high CO,-requiring phenotype. Plant Physiol. 91, 505 513
Price, G.D., Coleman, J.R, Badger, M.R. (1992) Association of carbonic anhydrase activity with carboxysomes isolated from the cyanobac- terium Synechococcus PCC7942. Plant Physiol. 100, 784-793
Reinhold, L., Kosloff, R., Kaplan, A. (1991) A model for inorganic car- bon fluxes and photosynthesis in cyanobacterial carboxysomes. Can. J, Bot. 69, 984-988
Spalding, M.H., Spreitzer, R J,, Ogren, W.L. (1983) Carbonic anhydrase deficient mutant of Chlamydomonas reinhardtii requires elevated car- bon dioxide concentration for photoautotrophic growth. Plant Phys- iol. 73, 268-272
Spurr, A.R. (1969) A low-viscosity resin embedding medium for electron microscopy. J. Ultrastruct. Res. 26, 31-34
Toguri, T., Muto, S., Mihara, S., Miyachi, S. (1989) Synthesis and degra- dation of carbonic anhydrase in a synchronized culture of Chlamy- domonas reinhartii. Plant Cell Physiol. 30, 533 539
Turpin, D.H., Miller, A.G., Canvin, D.T. (1984) Carboxysome content of Synechococcus leopoliensis (Cyanophyta) in response to inorganic carbon. J. Phycol. 20, 249-253
Vladimirova, M.G., Markelova, A.G., Semenenko, V.E. (1982) Identifica- tion of ribulose bisphosphate carboxylase location in the pyrenoids of unicellular algae by the cytoimmunofluorescent method. Fiziol. Rast. (Moscow) 29, 941-950





























