Embed Size (px)
Citation preview

The Impact of Gamma Radiation on Sediment Microbial Processes
Ashley R. Brown,a Christopher Boothman,a Simon M. Pimblott,b,c Jonathan R. Lloyda
Williamson Research Centre for Molecular Environmental Science and Research Centre for Radwaste and Decommissioning, School of Earth, Atmospheric, andEnvironmental Sciences, University of Manchester, Manchester, United Kingdoma; Dalton Cumbrian Facility, Westlakes Science and Technology Park, Cumbria, UnitedKingdomb; School of Chemistry, University of Manchester, Manchester, United Kingdomc
Microbial communities have the potential to control the biogeochemical fate of some radionuclides in contaminated land sce-narios or in the vicinity of a geological repository for radioactive waste. However, there have been few studies of ionizing radia-tion effects on microbial communities in sediment systems. Here, acetate and lactate amended sediment microcosms irradiatedwith gamma radiation at 0.5 or 30 Gy h�1 for 8 weeks all displayed NO3
� and Fe(III) reduction, although the rate of Fe(III) re-duction was decreased in 30-Gy h�1 treatments. These systems were dominated by fermentation processes. Pyrosequencing indi-cated that the 30-Gy h�1 treatment resulted in a community dominated by two Clostridial species. In systems containing noadded electron donor, irradiation at either dose rate did not restrict NO3
�, Fe(III), or SO42� reduction. Rather, Fe(III) reduction
was stimulated in the 0.5-Gy h�1-treated systems. In irradiated systems, there was a relative increase in the proportion of bacte-ria capable of Fe(III) reduction, with Geothrix fermentans and Geobacter sp. identified in the 0.5-Gy h�1 and 30-Gy h�1 treat-ments, respectively. These results indicate that biogeochemical processes will likely not be restricted by dose rates in such envi-ronments, and electron accepting processes may even be stimulated by radiation.
In many countries, including the United Kingdom, the currentpolicy for the long-term disposal of intermediate-level radioac-
tive waste is to a deep geological disposal facility (GDF). In UKdisposal concepts for higher-strength rocks and lower-strengthsedimentary rocks, much of the intermediate-level radioactivewaste is immobilized with a cementitious grout in stainless steelcontainers that are then surrounded with a cementitious backfillprior to closure of the facility (1). The vicinity of a GDF will not bea sterile environment, and microbial activity in the surroundinggeosphere could have important implications for the evolution ofbiogeochemical processes, including microbial gas generation andutilization, microbially induced corrosion of waste containers andcontents, and the mobility of radionuclides (2). In addition, therewill be elevated concentrations of potential electron donors in andaround the repository, including organics from the degradation ofcellulose in the waste (3), and also molecular hydrogen from theradiolysis of water and the anaerobic corrosion of steel drums (4).Indeed, the availability of alternative electron acceptors will likelynot be limited, since nitrate can be present in nuclear waste mate-rials (5), and Fe(III) will be present due to aerobic corrosion ofwaste components and engineered infrastructure during the op-erational phase of the GDF.
The stimulation of an Fe(III)-reducing community due to anincrease in electron donors and acceptors is of particular interestsince this may promote the reduction and precipitation of redox-active radionuclides via the production of biogenic Fe(II)-bearingphases (2, 6). Indeed, many key Fe(III)-reducing species may alsopossess cytochromes and hydrogenases capable of directly reduc-ing multivalent elements, such as Tc(VII), Np(V), and U(VI),with radionuclides of interest in safety assessments (7, 8). Becausethese processes could lower the mobility of these elements, themicrobial ecology and potential for Fe(III) reduction in geodis-posal environments has been the focus of recent research.
These environments, and the microbially driven processes thatoccur within them, may be subject to significant radiation doses.Firm values for total absorbed doses and dose rates are difficult topredict since they are likely to be highly heterogeneous and depen-
dent on the activity of the waste, the radiation type, decay dynam-ics, and the absorbing materials in the waste. For example, Cana-dian researchers predict the maximum dose rate to be 52 Gy h�1 atthe surface of a waste container (9). Similarly, Allard and Calas(10) suggest that dose rates in silicate clays used for backfill mate-rial may be in the order of 72 Gy h�1 over the first 1,000 years ofthe repository lifetime. For Swedish spent fuel disposal, on theother hand, the maximum estimate of dose rate outside the can-ister is 0.5 Gy h�1 over the first 1,000 years, followed by significantdecay after this (11).
Significant radiation fluxes may also be associated with nearsurface sites contaminated by radionuclides; for example, activi-ties up to 0.37 GBq kg�1 have been measured at contaminatedU.S. Department of Energy sites (12, 13). Again, it is difficult topredict how activities such as this relate to dose rates and totalabsorbed doses; however, as a reference, it has been calculated byparticle track calculation and Monte Carlo simulation that activ-ities of 8.1 MBq kg�1 90Sr and 9.6 MBq kg�1 137Cs in Chernobylsoils equate to dose rates of 51.7 and 14.8 Gy year�1, respectively(14).
Ionizing radiation is potentially lethal to organisms since theenergies involved are sufficient to cause strand breaks in DNA.Despite this, most bacteria encode conventional enzymatic DNArepair mechanisms, rendering much of the damage repairable.However, cytoplasmic water radiolysis generates quantities of re-
Received 19 February 2015 Accepted 29 March 2015
Accepted manuscript posted online 3 April 2015
Citation Brown AR, Boothman C, Pimblott SM, Lloyd JR. 2015. The impact ofgamma radiation on sediment microbial processes. Appl Environ Microbiol81:4014 –4025. doi:10.1128/AEM.00590-15.
Editor: S.-J. Liu
Address correspondence to Jonathan R. Lloyd, [email protected].
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AEM.00590-15
4014 aem.asm.org June 2015 Volume 81 Number 12Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

active oxygen species (e.g., HO�, H2O2, and O2��), which may
react indiscriminately with essential biomolecules, such as nucleicacids, proteins, and lipids, causing damage (15–17). Indeed, radi-ation-induced protein oxidation has been quantifiably related tobacterial viability (17).
When the generation of reactive oxygen species exceeds thescavenging capacity of the cell, oxidative stress is incurred. This,when combined with the inability of a cell’s metabolism to replen-ish damaged molecules as a result of radiation stress, likely resultsin fatality. The dose at which this occurs in a specific species is veryvariable and, as such, there has long been a focus on determiningradiation sensitivity in environmentally important species. Forexample, the extreme radiation resistance of Deinococcus radio-durans and the sensitivity of subsurface bacterial species such asFe(III)-reducing Shewanella sp. have been assessed (15, 18). How-ever, many of these studies were conducted with pure cultures athigh acute doses, and while acute-dose laboratory studies maypredict canister vicinities to be sterile (9, 19), survival may actuallybe possible under dose rates more relevant to nuclear environ-ments. For example, under a chronic dose rate of �2 Gy h�1,microorganisms isolated from a spent nuclear fuel pond were ca-pable of surviving total absorbed doses five times greater thantolerated in acute-dose experiments (�426 Gy h�1) (20, 21). Fur-thermore, microbes from the indigenous endolithic communityof a proposed repository were capable of surviving low gammadoses in a viable-but-nonculturable state (22), such that resusci-tation may be possible when environmental conditions becomemore favorable (23). This highlights the importance of gatheringlow dose rate data, particularly since lower dose rates may allowspecies to respond via upregulating repair mechanisms (24) oreven adapting over geological timescales, relevant to radwaste dis-posal scenarios. Similarly, the survival data from pure culturestudies may not be applicable to relatively nutrient limited sedi-ments, where there is competition from different species of thecommunity, and where radiation is perhaps not the only selectivestress. Indeed, Bruhn et al. (21) showed that the survival of theusually radioresistant D. radiodurans in a mixed culture was some-what limited, probably as a result of competition with Pseudomo-nas spp. However, this study was conducted in a rich tryptic soybroth medium that is far from representative of in situ GDF con-ditions.
Although it is important to examine the radiation tolerance ofmicrobial community members, radiation may also impact uponthe extracellular environment, which may consequently influencethe capacity for microbial processes. For instance, the radiolysis ofwater generates molecular hydrogen, which may be used as anelectron donor for a range of microbial electron accepting pro-cesses (2, 25–27). Furthermore, radiation has been shown to breakdown natural organic matter in soils, resulting in increases in dis-solved organic carbon (DOC) (28, 29). This radiolytic degrada-tion of organic matter may enhance the bioavailability of organiccarbon for microbial metabolism.
The oxidation state of potential electron acceptors may also bealtered by ionizing radiation. For instance, irradiation led toFe(III) reduction in a range of materials, including clays and go-ethite (28, 30–32). On the other hand, irradiation induced oxida-tion of Fe in steel and aqueous Fe(II) solutions led to the genera-tion of the Fe(II)/(III) oxides lepidocrocite, maghemite, andmagnetite (33, 34). Such changes to the oxidation state of Fe may
have important implications for the bioavailability of Fe(III) formicrobial respiration.
Even when no radiation-induced oxidation or reduction is ob-served, Fe(III) in both ferrihydrite and hematite may be mademore available for microbial reduction via alteration to the crys-talline structure (35). Since Fe is likely to be a significant compo-nent of waste packaging and repository infrastructure, suchradiation effects could have important implications to deep sub-surface microbial communities. With regard to other electron ac-ceptors, many studies have shown a decrease in the concentrationof nitrate in irradiated soils (36). On the other hand, sulfate con-centrations increased in a soil by 17% after 30 kGy of gammairradiation, although this was attributed to releases from lysedcells (37).
It is therefore evident that radiation may impact upon bothcellular physiology and the bioavailability of growth substrates,i.e., electron donors, acceptors, and presumably nutrients (36).However, despite the potential consequences to the evolution ofbiogeochemical processes in nuclear environments, there is a lackof information on the combined effect of all of these radiologicalprocesses on microbial metabolism at low dose rates. Here, weaddress the impact of low-dose chronic gamma irradiation upon asediment microbial community and the biogeochemical processescontrolled by this community both during irradiation andthroughout a subsequent recovery stage. In addition, Fe(II) con-centrations were probed to assess the ability of an irradiated com-munity to carry out Fe(III) reduction. To our knowledge, this wasconducted using the lowest dose rate over the longest irradiationperiod of any comparable study to date. Two dose rates were used:30 Gy h�1, representative of dose rates at radwaste canister sur-faces, and 0.5 Gy h�1, simulating dose rates further afield, or afterdecay of radiation levels and microbial repopulation of the repos-itory vicinity. This is in sharp contrast to the acute radiation levelsused in other pure culture studies.
MATERIALS AND METHODSSediment collection. Sediment samples were taken from a location rep-resentative of the Quaternary, unconsolidated alluvial flood-plain depos-its in the vicinity of the UK Sellafield reprocessing site. This site was se-lected because our group has extensive experience studying thebiogeochemistry of sediment from this area. Samples were collected fromthe shallow subsurface at a locality �2 km from the Sellafield site, in theCalder Valley, Cumbria (38–40). Samples were transferred to sterile con-tainers, sealed, and stored in the dark at 4°C prior to use.
Sediment microcosms. To assess the impact of gamma radiation onthe indigenous microorganisms of the sediment, microcosms were pre-pared in sterile 100-ml serum bottles by the addition of a sterile syntheticgroundwater representative of the region (41) to samples of sediment(10 � 0.1 g of sediment, 100 � 1 ml of groundwater buffered at pH 7 usingNaHCO3 at 0.24 g liter�1). After addition of the buffered groundwater,the pH of the microcosms was �6.4. Sodium lactate and sodium acetatewere added as electron donors, where necessary, to give final added con-centrations of 7 mM for each. Thus, a range of microcosm conditions wereproduced, as shown in Table 1. Triplicates of each of the different micro-cosms were then sealed with butyl rubber stoppers prior to irradiation.
Microcosms containing no added electron donor were also irradiatedprior to the addition of an active Geobacter sulfurreducens culture to in-vestigate the effect of radiation on Fe(III) reduction in the sediments,while evaluating the impact of radiation toxicity on the indigenous mi-croorganisms. After irradiation, microcosms were purged with an N2-CO2 (80:20) gas mixture to render the sediments anoxic to support mi-crobial Fe(III) reduction. Suspensions of G. sulfurreducens (100 �l) were
Microbial Communities and Gamma Radiation
June 2015 Volume 81 Number 12 aem.asm.org 4015Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

added, where necessary, to give a final cell density of �107 cells ml�1.Cultures were initially prepared by growing G. sulfurreducens at 30°C in afully defined anaerobic medium, as described previously (42). Sodiumacetate (20 mM) and fumarate (40 mM) were added as electron donor andacceptor, respectively. After 24 h, late-log/early-stationary-phase cultureswere harvested anaerobically by centrifugation at 4,920 � g for 20 minunder N2-CO2 (80:20) and washed twice with sterile nitrogen-purged 30mM sodium bicarbonate (pH 7.2).
Irradiations. Microcosm irradiations were carried out in the dark atCell 5, AMEC, Harwell, United Kingdom. Co-60 gamma (1.25 MeV) wassupplied to two separate sets of microcosms at dose rates of 0.5 Gy h�1
(�10%) and 30 Gy h�1 (�10%) over a 56-day period. The total absorbeddoses after the 56-day period are given in Table 1. The total absorbed dosein microcosms irradiated at 30 Gy h�1 for 28 days, when aliquots wereremoved for chemical analysis, was 19.2 kGy (�10%). Dose measure-ments were made with instrumentation traceable to national standards.The temperature inside Cell 5 was 18 � 1°C. External control experimentswere maintained at the same temperature. After irradiation, all micro-cosms were returned to the University of Manchester Geomicrobiologylaboratory and incubated in the dark at 19°C.
Geochemical analyses. Experiments were sampled periodically forgeochemical analyses and microbial community analysis using aseptictechniques under anoxic conditions. Microbial Fe(III) reduction wasmonitored in all microcosms by spectrophotometric determination ofFe(II) using the ferrozine assay (43). Biogenic Fe(II) was determined bydigestion of 100 �l of sediment slurry in 5 ml of 0.5 N HCl for 1 h. Thetotal bioavailable Fe was determined by digesting 100 �l of sedimentslurry in 5 ml of 0.25 N HCl and 0.25 N hydroxylamine-HCl, followed bythe ferrozine assay (44).
A sediment slurry (2 ml) from each replicate microcosm was centri-fuged at 3,000 � g for 3 min. The supernatant was used for analysis by ionchromatography, and the sediment was used for microbiological charac-terization. Samples were stored at �20°C prior to analysis.
Ion chromatography. Chloride, nitrate, nitrite, sulfate, phosphate,and organic acids were measured using a Dionex IC5000 system with aDionex capillary AS11-HC-4�m column. Aliquots (0.4 �l) were injectedinto a potassium hydroxide mobile phase with a flow rate of 0.015 mlmin�1 and a gradient of 1 to 36 mM KOH over 40 min.
16S amplicon pyrosequencing and data analysis. Samples for 16SrRNA gene pyrosequencing were taken from the microcosm that was themost representative of the mean of chemical analyses of the three repli-cates. DNA was isolated from microcosm samples (200 �l of slurry) usingthe MoBio PowerSoil DNA isolation kit (MoBio Laboratories, Inc., Carls-
bad, CA) according to the manufacturer’s instructions. PCR of the V1-V2hypervariable region of the bacterial 16S rRNA gene was performed usinguniversal bacterial primers 27F (45) and 338R (46), synthesized byIDTdna (Integrated DNA Technologies, BVBA, Leuven, Belgium).
The fusion forward primer (5=-CCATCTCATCCCTGCGTGTCTCCGACTCAGXXXXXXXXAGAGTTTGATGMTGGCTCAG-3=) contained the454 Life Sciences “Lib-L Primer A,” a 4 base “key” sequence (TCAG), aunique eight-base barcode “MID” sequence for each sample (XXXXXXXX),and bacterial primer 27F. The reverse fusion primer (5=-CCTATCCCCTGTGTGCCTTGGCAGTCTCAGTGCTGCCTCCCGTAGGAGT-3=) con-tained the 454 Life Sciences “Lib-L Primer B,” a four-base “key” sequence(TCAG), and bacterial primer 338R. The PCR amplification was performedin 50-�l volume reactions using 0.5 �l (2.5 U) of FastStart high-fidelity DNApolymerase (Roche Diagnostics GmbH, Mannheim, Germany), 1.8 mMMgCl2, 200 �M concentrations of each deoxynucleoside triphosphate, 0.4�M concentrations of each forward and reverse fusion primer, and 2 �l ofDNA template. The PCR conditions included an initial denaturing step at95°C for 2 min, followed by 35 cycles of 95°C for 30 s, 55°C for 30 s, 72°C for45 s, and a final elongation step at 72°C for 5 min.
PCR products were loaded in an agarose gel and, following gel electro-phoresis, bands of the correct fragment size (�410 bp) were excised,cleaned up using a QIAquick gel extraction kit (Qiagen, GmBH, Hilden,Germany), and eluted in 30 �l of DNase-free H2O. The cleaned-up PCRproducts from the present study (22 samples in total) were quantified byusing an Agilent 2100 bioanalyzer (Agilent Technologies, Inc., SantaClara, CA) and pooled so that the mixture contained equal amounts ofDNA from each sample. The emulsion PCR and pyrosequencing run wereperformed at the University of Manchester sequencing facility using a 454Life Sciences GS Junior system (Roche).
The 454 pyrosequencing reads were analyzed using Qiime 1.6.0 release(47). Low-quality reads (mean quality score of �25) and short sequences(�300 bp) were discarded, and both forward and reverse primers wereremoved from further analysis. Denoising and chimera removal were per-formed during operational taxonomic unit (OTU) picking (at 97% se-quence similarity) using “usearch” (48) in Qiime, and a representativesequence for each OTU was identified. The taxonomic classification of allreads was performed in Qiime using the Ribosomal Database Project(RDP) at a 80% confidence threshold (48), while the closest GenBankmatch for the OTU that contained the highest number of reads (the rep-resentative sequence for each OTU was used) was identified by BLASTNnucleotide search.
RESULTS AND DISCUSSIONBiogeochemistry of irradiated microcosms containing addedelectron donor. To assess the impact of chronic gamma irradia-tion on the biogeochemical processes in the sediment, a series ofmicrocosms were prepared with or without added electron donorand irradiated for 56 days at 0.5 and 30 Gy h�1. In control andirradiated microcosms spiked with lactate and acetate (final addedconcentrations of 7 mM each), electron acceptor usage progressedin the order nitrate � Fe(III) � sulfate during the irradiationperiod (Fig. 1). Treatment with gamma radiation at 30 Gy h�1 didnot appear to affect the reduction of nitrate, which was removedcompletely from porewaters after 28 days in both treated and con-trol microcosms. However, 0.5 N HCl extractable Fe(II) concen-trations in microcosms after treatment with 30 Gy h�1 for 56 dayswere �0.5 mM compared to �2 mM in nonirradiated controls.This limited Fe(III) reduction was likely due to decreased viabilityof Fe(III)-reducing microorganisms arising from a total absorbeddose of 38.6 kGy. After completion of the irradiation, the levels of0.5 N HCl extractable Fe(II) increased gradually over 176 days tolevels comparable to those in nonirradiated microcosms, suggest-ing that significant Fe(III) reduction was still possible, albeit at aslower rate, even after the maximum radiation dose was applied.
TABLE 1 Initial microcosm compositions and treatments
Expt system
Mean doserate(Gy h�1)b
Mean totalabsorbed dose(kGy)b
Electron donoramendmenta
Lactate(mM)
Acetate(mM)
Sediment electrondonor
Nonirradiated 0 7 70.5 0.6 7 730 38.6 7 7
Sediment Nonirradiated 0 0 00.5 0.6 0 030 38.6 0 0
Sediment G.sulfurreducens
Nonirradiated 0 0 00.5 0.6 0 030 38.6 0 0
a Lactate and acetate were added, where required, to give the final concentrationsshown.b � 10% relative standard deviation.
Brown et al.
4016 aem.asm.org June 2015 Volume 81 Number 12Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
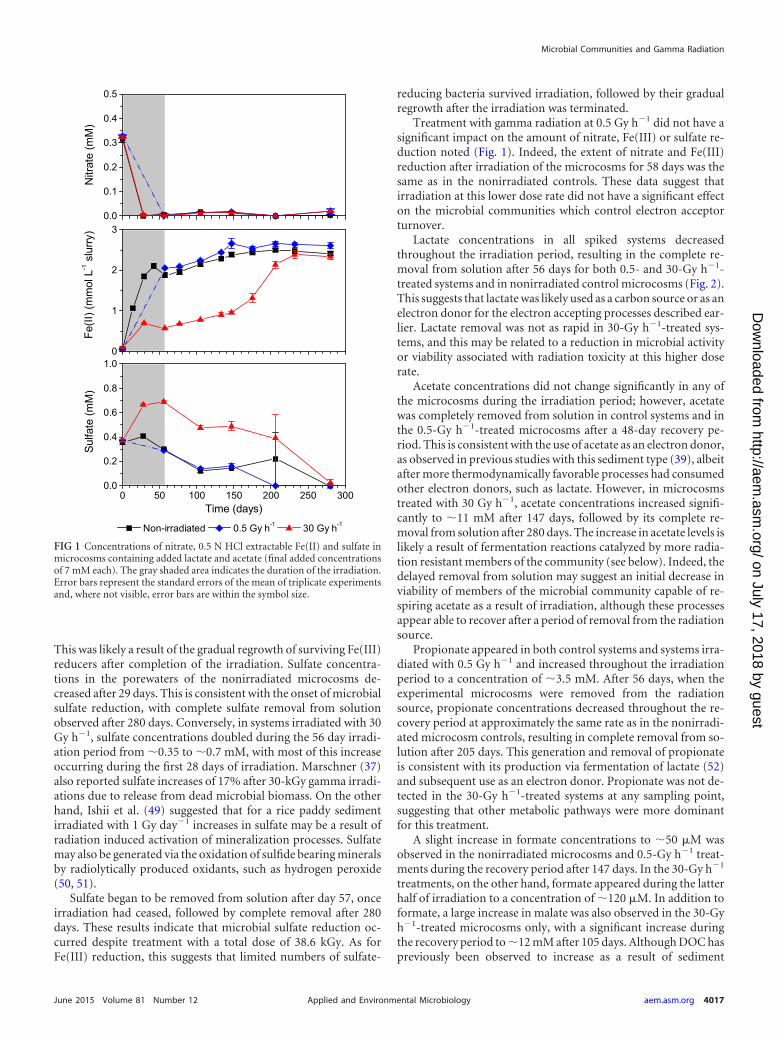
This was likely a result of the gradual regrowth of surviving Fe(III)reducers after completion of the irradiation. Sulfate concentra-tions in the porewaters of the nonirradiated microcosms de-creased after 29 days. This is consistent with the onset of microbialsulfate reduction, with complete sulfate removal from solutionobserved after 280 days. Conversely, in systems irradiated with 30Gy h�1, sulfate concentrations doubled during the 56 day irradi-ation period from �0.35 to �0.7 mM, with most of this increaseoccurring during the first 28 days of irradiation. Marschner (37)also reported sulfate increases of 17% after 30-kGy gamma irradi-ations due to release from dead microbial biomass. On the otherhand, Ishii et al. (49) suggested that for a rice paddy sedimentirradiated with 1 Gy day�1 increases in sulfate may be a result ofradiation induced activation of mineralization processes. Sulfatemay also be generated via the oxidation of sulfide bearing mineralsby radiolytically produced oxidants, such as hydrogen peroxide(50, 51).
Sulfate began to be removed from solution after day 57, onceirradiation had ceased, followed by complete removal after 280days. These results indicate that microbial sulfate reduction oc-curred despite treatment with a total dose of 38.6 kGy. As forFe(III) reduction, this suggests that limited numbers of sulfate-
reducing bacteria survived irradiation, followed by their gradualregrowth after the irradiation was terminated.
Treatment with gamma radiation at 0.5 Gy h�1 did not have asignificant impact on the amount of nitrate, Fe(III) or sulfate re-duction noted (Fig. 1). Indeed, the extent of nitrate and Fe(III)reduction after irradiation of the microcosms for 58 days was thesame as in the nonirradiated controls. These data suggest thatirradiation at this lower dose rate did not have a significant effecton the microbial communities which control electron acceptorturnover.
Lactate concentrations in all spiked systems decreasedthroughout the irradiation period, resulting in the complete re-moval from solution after 56 days for both 0.5- and 30-Gy h�1-treated systems and in nonirradiated control microcosms (Fig. 2).This suggests that lactate was likely used as a carbon source or as anelectron donor for the electron accepting processes described ear-lier. Lactate removal was not as rapid in 30-Gy h�1-treated sys-tems, and this may be related to a reduction in microbial activityor viability associated with radiation toxicity at this higher doserate.
Acetate concentrations did not change significantly in any ofthe microcosms during the irradiation period; however, acetatewas completely removed from solution in control systems and inthe 0.5-Gy h�1-treated microcosms after a 48-day recovery pe-riod. This is consistent with the use of acetate as an electron donor,as observed in previous studies with this sediment type (39), albeitafter more thermodynamically favorable processes had consumedother electron donors, such as lactate. However, in microcosmstreated with 30 Gy h�1, acetate concentrations increased signifi-cantly to �11 mM after 147 days, followed by its complete re-moval from solution after 280 days. The increase in acetate levels islikely a result of fermentation reactions catalyzed by more radia-tion resistant members of the community (see below). Indeed, thedelayed removal from solution may suggest an initial decrease inviability of members of the microbial community capable of re-spiring acetate as a result of irradiation, although these processesappear able to recover after a period of removal from the radiationsource.
Propionate appeared in both control systems and systems irra-diated with 0.5 Gy h�1 and increased throughout the irradiationperiod to a concentration of �3.5 mM. After 56 days, when theexperimental microcosms were removed from the radiationsource, propionate concentrations decreased throughout the re-covery period at approximately the same rate as in the nonirradi-ated microcosm controls, resulting in complete removal from so-lution after 205 days. This generation and removal of propionateis consistent with its production via fermentation of lactate (52)and subsequent use as an electron donor. Propionate was not de-tected in the 30-Gy h�1-treated systems at any sampling point,suggesting that other metabolic pathways were more dominantfor this treatment.
A slight increase in formate concentrations to �50 �M wasobserved in the nonirradiated microcosms and 0.5-Gy h�1 treat-ments during the recovery period after 147 days. In the 30-Gy h�1
treatments, on the other hand, formate appeared during the latterhalf of irradiation to a concentration of �120 �M. In addition toformate, a large increase in malate was also observed in the 30-Gyh�1-treated microcosms only, with a significant increase duringthe recovery period to �12 mM after 105 days. Although DOC haspreviously been observed to increase as a result of sediment
FIG 1 Concentrations of nitrate, 0.5 N HCl extractable Fe(II) and sulfate inmicrocosms containing added lactate and acetate (final added concentrationsof 7 mM each). The gray shaded area indicates the duration of the irradiation.Error bars represent the standard errors of the mean of triplicate experimentsand, where not visible, error bars are within the symbol size.
Microbial Communities and Gamma Radiation
June 2015 Volume 81 Number 12 aem.asm.org 4017Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
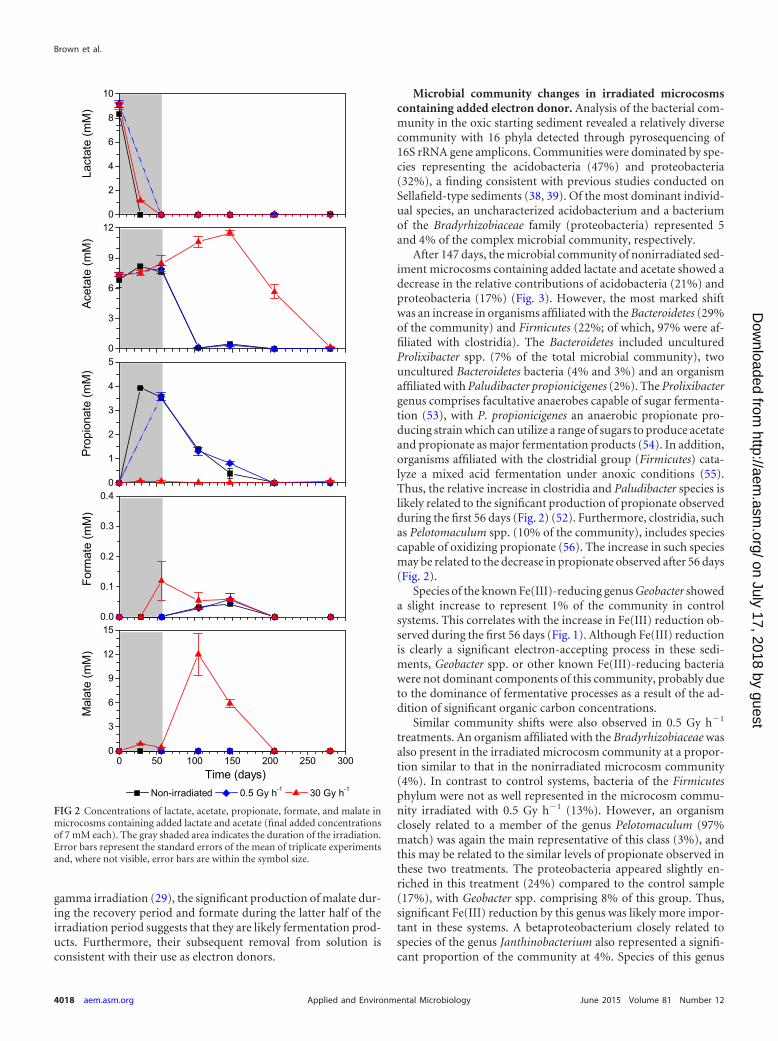
gamma irradiation (29), the significant production of malate dur-ing the recovery period and formate during the latter half of theirradiation period suggests that they are likely fermentation prod-ucts. Furthermore, their subsequent removal from solution isconsistent with their use as electron donors.
Microbial community changes in irradiated microcosmscontaining added electron donor. Analysis of the bacterial com-munity in the oxic starting sediment revealed a relatively diversecommunity with 16 phyla detected through pyrosequencing of16S rRNA gene amplicons. Communities were dominated by spe-cies representing the acidobacteria (47%) and proteobacteria(32%), a finding consistent with previous studies conducted onSellafield-type sediments (38, 39). Of the most dominant individ-ual species, an uncharacterized acidobacterium and a bacteriumof the Bradyrhizobiaceae family (proteobacteria) represented 5and 4% of the complex microbial community, respectively.
After 147 days, the microbial community of nonirradiated sed-iment microcosms containing added lactate and acetate showed adecrease in the relative contributions of acidobacteria (21%) andproteobacteria (17%) (Fig. 3). However, the most marked shiftwas an increase in organisms affiliated with the Bacteroidetes (29%of the community) and Firmicutes (22%; of which, 97% were af-filiated with clostridia). The Bacteroidetes included unculturedProlixibacter spp. (7% of the total microbial community), twouncultured Bacteroidetes bacteria (4% and 3%) and an organismaffiliated with Paludibacter propionicigenes (2%). The Prolixibactergenus comprises facultative anaerobes capable of sugar fermenta-tion (53), with P. propionicigenes an anaerobic propionate pro-ducing strain which can utilize a range of sugars to produce acetateand propionate as major fermentation products (54). In addition,organisms affiliated with the clostridial group (Firmicutes) cata-lyze a mixed acid fermentation under anoxic conditions (55).Thus, the relative increase in clostridia and Paludibacter species islikely related to the significant production of propionate observedduring the first 56 days (Fig. 2) (52). Furthermore, clostridia, suchas Pelotomaculum spp. (10% of the community), includes speciescapable of oxidizing propionate (56). The increase in such speciesmay be related to the decrease in propionate observed after 56 days(Fig. 2).
Species of the known Fe(III)-reducing genus Geobacter showeda slight increase to represent 1% of the community in controlsystems. This correlates with the increase in Fe(III) reduction ob-served during the first 56 days (Fig. 1). Although Fe(III) reductionis clearly a significant electron-accepting process in these sedi-ments, Geobacter spp. or other known Fe(III)-reducing bacteriawere not dominant components of this community, probably dueto the dominance of fermentative processes as a result of the ad-dition of significant organic carbon concentrations.
Similar community shifts were also observed in 0.5 Gy h�1
treatments. An organism affiliated with the Bradyrhizobiaceae wasalso present in the irradiated microcosm community at a propor-tion similar to that in the nonirradiated microcosm community(4%). In contrast to control systems, bacteria of the Firmicutesphylum were not as well represented in the microcosm commu-nity irradiated with 0.5 Gy h�1 (13%). However, an organismclosely related to a member of the genus Pelotomaculum (97%match) was again the main representative of this class (3%), andthis may be related to the similar levels of propionate observed inthese two treatments. The proteobacteria appeared slightly en-riched in this treatment (24%) compared to the control sample(17%), with Geobacter spp. comprising 8% of this group. Thus,significant Fe(III) reduction by this genus was likely more impor-tant in these systems. A betaproteobacterium closely related tospecies of the genus Janthinobacterium also represented a signifi-cant proportion of the community at 4%. Species of this genus
FIG 2 Concentrations of lactate, acetate, propionate, formate, and malate inmicrocosms containing added lactate and acetate (final added concentrationsof 7 mM each). The gray shaded area indicates the duration of the irradiation.Error bars represent the standard errors of the mean of triplicate experimentsand, where not visible, error bars are within the symbol size.
Brown et al.
4018 aem.asm.org June 2015 Volume 81 Number 12Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
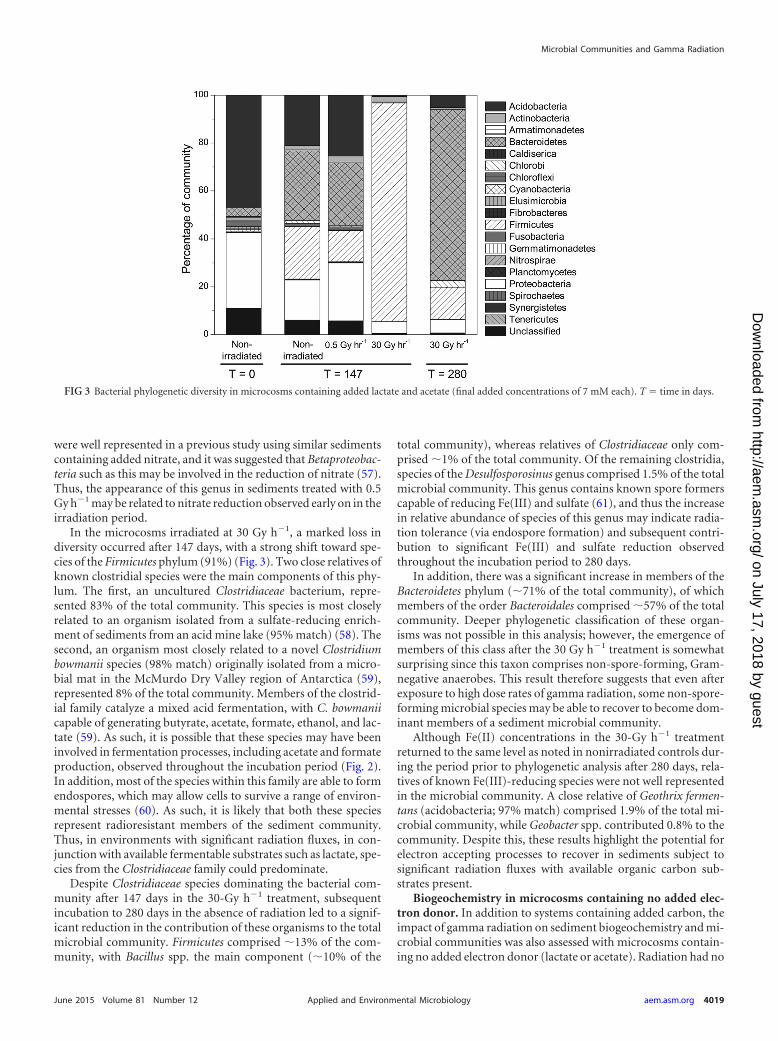
were well represented in a previous study using similar sedimentscontaining added nitrate, and it was suggested that Betaproteobac-teria such as this may be involved in the reduction of nitrate (57).Thus, the appearance of this genus in sediments treated with 0.5Gy h�1 may be related to nitrate reduction observed early on in theirradiation period.
In the microcosms irradiated at 30 Gy h�1, a marked loss indiversity occurred after 147 days, with a strong shift toward spe-cies of the Firmicutes phylum (91%) (Fig. 3). Two close relatives ofknown clostridial species were the main components of this phy-lum. The first, an uncultured Clostridiaceae bacterium, repre-sented 83% of the total community. This species is most closelyrelated to an organism isolated from a sulfate-reducing enrich-ment of sediments from an acid mine lake (95% match) (58). Thesecond, an organism most closely related to a novel Clostridiumbowmanii species (98% match) originally isolated from a micro-bial mat in the McMurdo Dry Valley region of Antarctica (59),represented 8% of the total community. Members of the clostrid-ial family catalyze a mixed acid fermentation, with C. bowmaniicapable of generating butyrate, acetate, formate, ethanol, and lac-tate (59). As such, it is possible that these species may have beeninvolved in fermentation processes, including acetate and formateproduction, observed throughout the incubation period (Fig. 2).In addition, most of the species within this family are able to formendospores, which may allow cells to survive a range of environ-mental stresses (60). As such, it is likely that both these speciesrepresent radioresistant members of the sediment community.Thus, in environments with significant radiation fluxes, in con-junction with available fermentable substrates such as lactate, spe-cies from the Clostridiaceae family could predominate.
Despite Clostridiaceae species dominating the bacterial com-munity after 147 days in the 30-Gy h�1 treatment, subsequentincubation to 280 days in the absence of radiation led to a signif-icant reduction in the contribution of these organisms to the totalmicrobial community. Firmicutes comprised �13% of the com-munity, with Bacillus spp. the main component (�10% of the
total community), whereas relatives of Clostridiaceae only com-prised �1% of the total community. Of the remaining clostridia,species of the Desulfosporosinus genus comprised 1.5% of the totalmicrobial community. This genus contains known spore formerscapable of reducing Fe(III) and sulfate (61), and thus the increasein relative abundance of species of this genus may indicate radia-tion tolerance (via endospore formation) and subsequent contri-bution to significant Fe(III) and sulfate reduction observedthroughout the incubation period to 280 days.
In addition, there was a significant increase in members of theBacteroidetes phylum (�71% of the total community), of whichmembers of the order Bacteroidales comprised �57% of the totalcommunity. Deeper phylogenetic classification of these organ-isms was not possible in this analysis; however, the emergence ofmembers of this class after the 30 Gy h�1 treatment is somewhatsurprising since this taxon comprises non-spore-forming, Gram-negative anaerobes. This result therefore suggests that even afterexposure to high dose rates of gamma radiation, some non-spore-forming microbial species may be able to recover to become dom-inant members of a sediment microbial community.
Although Fe(II) concentrations in the 30-Gy h�1 treatmentreturned to the same level as noted in nonirradiated controls dur-ing the period prior to phylogenetic analysis after 280 days, rela-tives of known Fe(III)-reducing species were not well representedin the microbial community. A close relative of Geothrix fermen-tans (acidobacteria; 97% match) comprised 1.9% of the total mi-crobial community, while Geobacter spp. contributed 0.8% to thecommunity. Despite this, these results highlight the potential forelectron accepting processes to recover in sediments subject tosignificant radiation fluxes with available organic carbon sub-strates present.
Biogeochemistry in microcosms containing no added elec-tron donor. In addition to systems containing added carbon, theimpact of gamma radiation on sediment biogeochemistry and mi-crobial communities was also assessed with microcosms contain-ing no added electron donor (lactate or acetate). Radiation had no
FIG 3 Bacterial phylogenetic diversity in microcosms containing added lactate and acetate (final added concentrations of 7 mM each). T time in days.
Microbial Communities and Gamma Radiation
June 2015 Volume 81 Number 12 aem.asm.org 4019Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
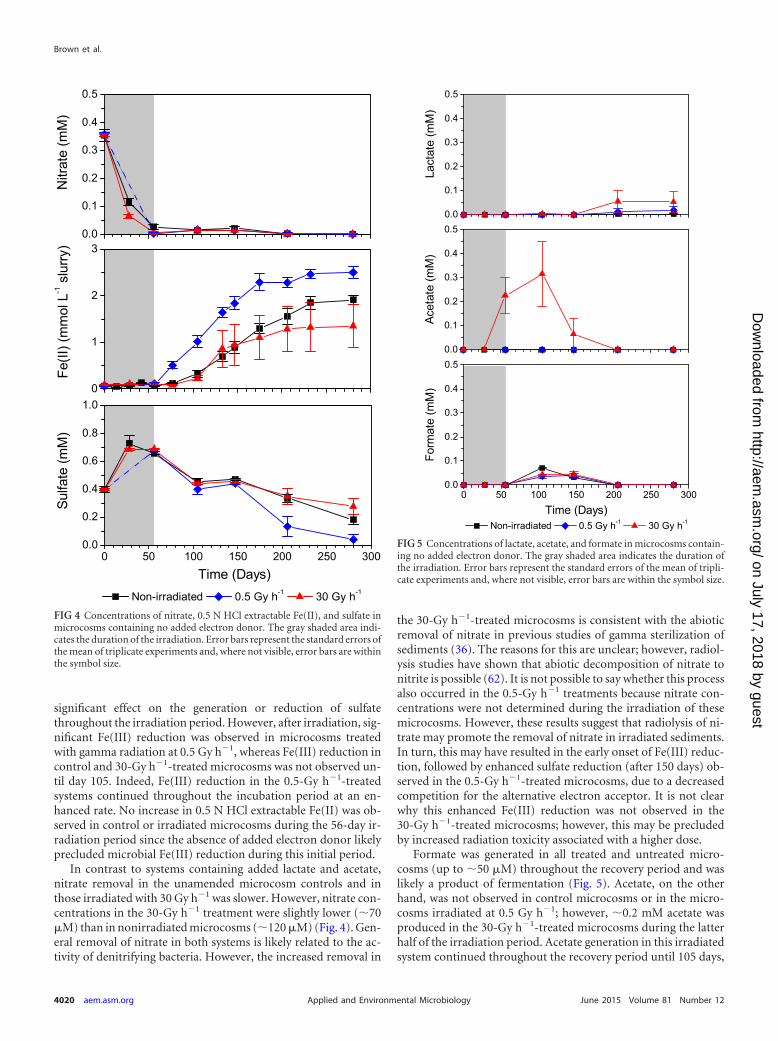
significant effect on the generation or reduction of sulfatethroughout the irradiation period. However, after irradiation, sig-nificant Fe(III) reduction was observed in microcosms treatedwith gamma radiation at 0.5 Gy h�1, whereas Fe(III) reduction incontrol and 30-Gy h�1-treated microcosms was not observed un-til day 105. Indeed, Fe(III) reduction in the 0.5-Gy h�1-treatedsystems continued throughout the incubation period at an en-hanced rate. No increase in 0.5 N HCl extractable Fe(II) was ob-served in control or irradiated microcosms during the 56-day ir-radiation period since the absence of added electron donor likelyprecluded microbial Fe(III) reduction during this initial period.
In contrast to systems containing added lactate and acetate,nitrate removal in the unamended microcosm controls and inthose irradiated with 30 Gy h�1 was slower. However, nitrate con-centrations in the 30-Gy h�1 treatment were slightly lower (�70�M) than in nonirradiated microcosms (�120 �M) (Fig. 4). Gen-eral removal of nitrate in both systems is likely related to the ac-tivity of denitrifying bacteria. However, the increased removal in
the 30-Gy h�1-treated microcosms is consistent with the abioticremoval of nitrate in previous studies of gamma sterilization ofsediments (36). The reasons for this are unclear; however, radiol-ysis studies have shown that abiotic decomposition of nitrate tonitrite is possible (62). It is not possible to say whether this processalso occurred in the 0.5-Gy h�1 treatments because nitrate con-centrations were not determined during the irradiation of thesemicrocosms. However, these results suggest that radiolysis of ni-trate may promote the removal of nitrate in irradiated sediments.In turn, this may have resulted in the early onset of Fe(III) reduc-tion, followed by enhanced sulfate reduction (after 150 days) ob-served in the 0.5-Gy h�1-treated microcosms, due to a decreasedcompetition for the alternative electron acceptor. It is not clearwhy this enhanced Fe(III) reduction was not observed in the30-Gy h�1-treated microcosms; however, this may be precludedby increased radiation toxicity associated with a higher dose.
Formate was generated in all treated and untreated micro-cosms (up to �50 �M) throughout the recovery period and waslikely a product of fermentation (Fig. 5). Acetate, on the otherhand, was not observed in control microcosms or in the micro-cosms irradiated at 0.5 Gy h�1; however, �0.2 mM acetate wasproduced in the 30-Gy h�1-treated microcosms during the latterhalf of the irradiation period. Acetate generation in this irradiatedsystem continued throughout the recovery period until 105 days,
FIG 4 Concentrations of nitrate, 0.5 N HCl extractable Fe(II), and sulfate inmicrocosms containing no added electron donor. The gray shaded area indi-cates the duration of the irradiation. Error bars represent the standard errors ofthe mean of triplicate experiments and, where not visible, error bars are withinthe symbol size.
FIG 5 Concentrations of lactate, acetate, and formate in microcosms contain-ing no added electron donor. The gray shaded area indicates the duration ofthe irradiation. Error bars represent the standard errors of the mean of tripli-cate experiments and, where not visible, error bars are within the symbol size.
Brown et al.
4020 aem.asm.org June 2015 Volume 81 Number 12Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
but by the end of the incubation period acetate had largely beenremoved from the solution. The production of acetate (during thelatter part of irradiation only) and its subsequent removal suggestsits production by microbial fermentation, followed by its oxida-tion as an electron donor. It is possible that these processes oc-curred in the nonirradiated systems and in the 0.5-Gy h�1-treatedsystems; however, acetate may have been metabolized as quicklyas it was formed. Thus, the detection of acetate in the 30-Gy h�1-treated systems may be a result of radiation toxicity in acetate-oxidizing species.
It is unclear from these data whether the enhanced reduction ofFe(III) in the 0.5-Gy h�1-treated microcosms was related to anincrease in the availability of organic electron donors, increases inthe bioavailability of Fe(III), or a decrease in electron acceptorcompetition arising from enhanced nitrate removal.
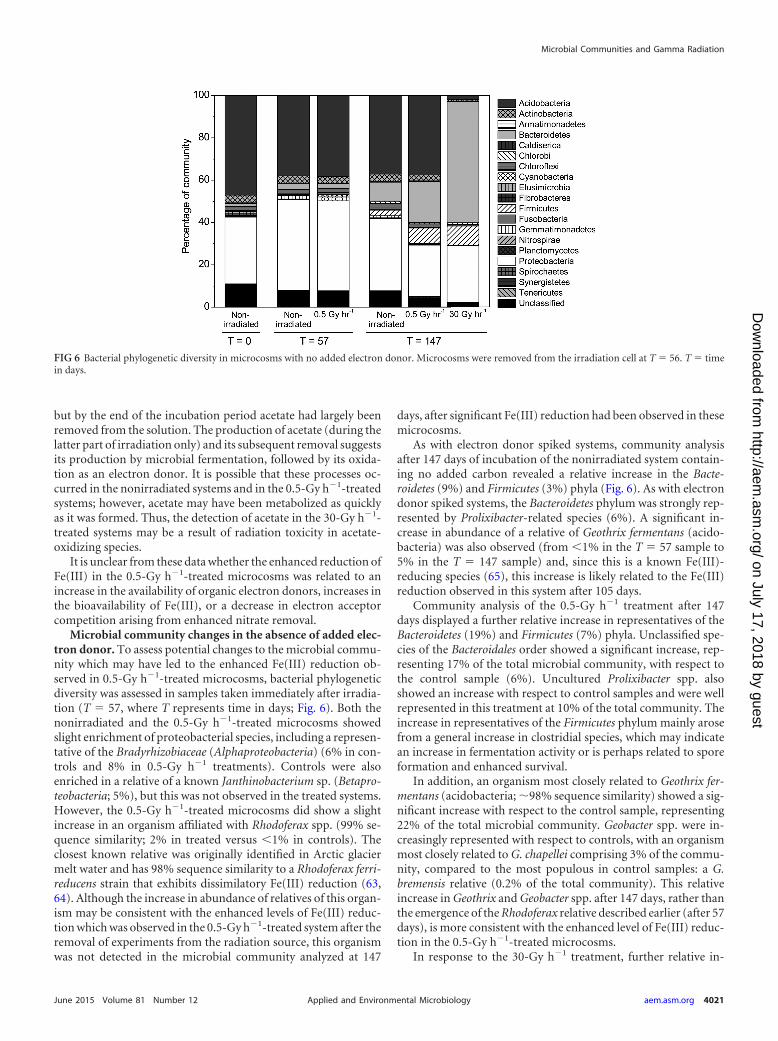
Microbial community changes in the absence of added elec-tron donor. To assess potential changes to the microbial commu-nity which may have led to the enhanced Fe(III) reduction ob-served in 0.5-Gy h�1-treated microcosms, bacterial phylogeneticdiversity was assessed in samples taken immediately after irradia-tion (T 57, where T represents time in days; Fig. 6). Both thenonirradiated and the 0.5-Gy h�1-treated microcosms showedslight enrichment of proteobacterial species, including a represen-tative of the Bradyrhizobiaceae (Alphaproteobacteria) (6% in con-trols and 8% in 0.5-Gy h�1 treatments). Controls were alsoenriched in a relative of a known Janthinobacterium sp. (Betapro-teobacteria; 5%), but this was not observed in the treated systems.However, the 0.5-Gy h�1-treated microcosms did show a slightincrease in an organism affiliated with Rhodoferax spp. (99% se-quence similarity; 2% in treated versus �1% in controls). Theclosest known relative was originally identified in Arctic glaciermelt water and has 98% sequence similarity to a Rhodoferax ferri-reducens strain that exhibits dissimilatory Fe(III) reduction (63,64). Although the increase in abundance of relatives of this organ-ism may be consistent with the enhanced levels of Fe(III) reduc-tion which was observed in the 0.5-Gy h�1-treated system after theremoval of experiments from the radiation source, this organismwas not detected in the microbial community analyzed at 147
days, after significant Fe(III) reduction had been observed in thesemicrocosms.
As with electron donor spiked systems, community analysisafter 147 days of incubation of the nonirradiated system contain-ing no added carbon revealed a relative increase in the Bacte-roidetes (9%) and Firmicutes (3%) phyla (Fig. 6). As with electrondonor spiked systems, the Bacteroidetes phylum was strongly rep-resented by Prolixibacter-related species (6%). A significant in-crease in abundance of a relative of Geothrix fermentans (acido-bacteria) was also observed (from �1% in the T 57 sample to5% in the T 147 sample) and, since this is a known Fe(III)-reducing species (65), this increase is likely related to the Fe(III)reduction observed in this system after 105 days.
Community analysis of the 0.5-Gy h�1 treatment after 147days displayed a further relative increase in representatives of theBacteroidetes (19%) and Firmicutes (7%) phyla. Unclassified spe-cies of the Bacteroidales order showed a significant increase, rep-resenting 17% of the total microbial community, with respect tothe control sample (6%). Uncultured Prolixibacter spp. alsoshowed an increase with respect to control samples and were wellrepresented in this treatment at 10% of the total community. Theincrease in representatives of the Firmicutes phylum mainly arosefrom a general increase in clostridial species, which may indicatean increase in fermentation activity or is perhaps related to sporeformation and enhanced survival.
In addition, an organism most closely related to Geothrix fer-mentans (acidobacteria; �98% sequence similarity) showed a sig-nificant increase with respect to the control sample, representing22% of the total microbial community. Geobacter spp. were in-creasingly represented with respect to controls, with an organismmost closely related to G. chapellei comprising 3% of the commu-nity, compared to the most populous in control samples: a G.bremensis relative (0.2% of the total community). This relativeincrease in Geothrix and Geobacter spp. after 147 days, rather thanthe emergence of the Rhodoferax relative described earlier (after 57days), is more consistent with the enhanced level of Fe(III) reduc-tion in the 0.5-Gy h�1-treated microcosms.
In response to the 30-Gy h�1 treatment, further relative in-
FIG 6 Bacterial phylogenetic diversity in microcosms with no added electron donor. Microcosms were removed from the irradiation cell at T 56. T timein days.
Microbial Communities and Gamma Radiation
June 2015 Volume 81 Number 12 aem.asm.org 4021Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
creases were observed in the Bacteroidetes and Firmicutes phyla(Fig. 6). Unclassified species from the order Bacteroidales repre-sented 37% of the total microbial community. This comprisedtwo dominant species: the first (12% of the community) was mostclosely related to an uncultured bacterium isolated from mosspillars at an Antarctic lake (66) and the second was an unculturedProlixibacter sp. (11%). Paludibacter spp., also of the order Bacte-roidales, represented 5% of the total community and species of thefamily Chitinophagaceae (Bacteroidetes phylum) comprised 7% ofthe total community. Two uncultured sphingobacteria (Bacte-roidetes phylum) represented 14% of the community. Interest-ingly, these observations are similar to those from microcosmsamended with acetate and lactate and irradiated at 30 Gy h�1, inwhich members of the Bacteroidales represented �57% of the totalmicrobial community after 280 days. These results suggest thatmembers of the Bacteroidetes phylum may exhibit high levels ofradiation resistance and potentially represent a group of respira-tory generalists capable of dominating a community after signifi-cant radiation stress.
Of the key Fe(III)-reducing species in the microcosms irradi-ated at 30 Gy h�1, 18% of the total community was affiliated withknown Geobacter species. However, unlike in the microcosms ir-radiated at 0.5 Gy h�1, Geothrix species were not well represented,comprising �0.1% of the total microbial community. In addition,a close relative of the Herbaspirillum frisingense (Betaproteobacte-ria) comprised 5% of the total community (98% sequence simi-
larity). This species is capable of nitrate reduction and nitrogenfixation and can oxidize a broad range of sugars and alcohols (67).As in the microcosms irradiated at 0.5 Gy h�1, the increase inFirmicutes mainly arose from a general increase in clostridial spe-cies.
These results indicate that despite sediments receiving a totalabsorbed dose of nearly 40 kGy, Fe(III) reduction was still possiblein sediments without added electron donor. Furthermore, irradi-ation of these sediments resulted in significant increases in abun-dance of Fe(III)-reducing species compared to nonirradiated sys-tems. This suggests that, although Fe(III) reduction was notenhanced in the 30-Gy h�1-treated systems, these sediments maybe poised for Fe(III) reduction.
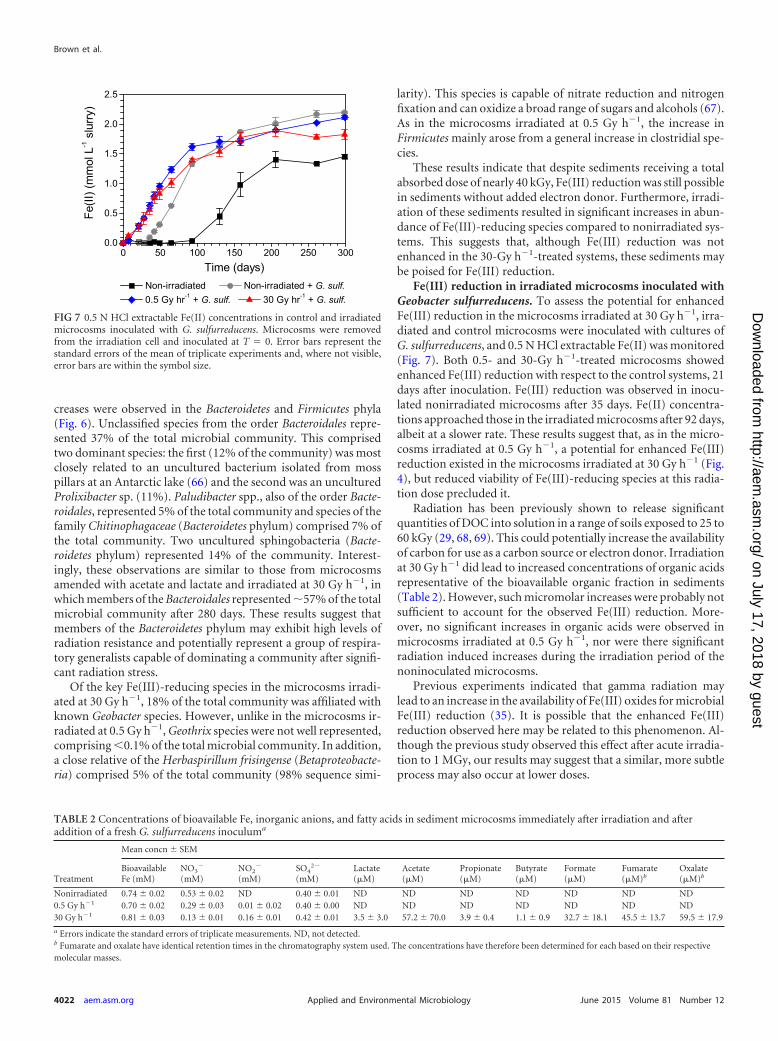
Fe(III) reduction in irradiated microcosms inoculated withGeobacter sulfurreducens. To assess the potential for enhancedFe(III) reduction in the microcosms irradiated at 30 Gy h�1, irra-diated and control microcosms were inoculated with cultures ofG. sulfurreducens, and 0.5 N HCl extractable Fe(II) was monitored(Fig. 7). Both 0.5- and 30-Gy h�1-treated microcosms showedenhanced Fe(III) reduction with respect to the control systems, 21days after inoculation. Fe(III) reduction was observed in inocu-lated nonirradiated microcosms after 35 days. Fe(II) concentra-tions approached those in the irradiated microcosms after 92 days,albeit at a slower rate. These results suggest that, as in the micro-cosms irradiated at 0.5 Gy h�1, a potential for enhanced Fe(III)reduction existed in the microcosms irradiated at 30 Gy h�1 (Fig.4), but reduced viability of Fe(III)-reducing species at this radia-tion dose precluded it.
Radiation has been previously shown to release significantquantities of DOC into solution in a range of soils exposed to 25 to60 kGy (29, 68, 69). This could potentially increase the availabilityof carbon for use as a carbon source or electron donor. Irradiationat 30 Gy h�1 did lead to increased concentrations of organic acidsrepresentative of the bioavailable organic fraction in sediments(Table 2). However, such micromolar increases were probably notsufficient to account for the observed Fe(III) reduction. More-over, no significant increases in organic acids were observed inmicrocosms irradiated at 0.5 Gy h�1, nor were there significantradiation induced increases during the irradiation period of thenoninoculated microcosms.
Previous experiments indicated that gamma radiation maylead to an increase in the availability of Fe(III) oxides for microbialFe(III) reduction (35). It is possible that the enhanced Fe(III)reduction observed here may be related to this phenomenon. Al-though the previous study observed this effect after acute irradia-tion to 1 MGy, our results may suggest that a similar, more subtleprocess may also occur at lower doses.
FIG 7 0.5 N HCl extractable Fe(II) concentrations in control and irradiatedmicrocosms inoculated with G. sulfurreducens. Microcosms were removedfrom the irradiation cell and inoculated at T 0. Error bars represent thestandard errors of the mean of triplicate experiments and, where not visible,error bars are within the symbol size.
TABLE 2 Concentrations of bioavailable Fe, inorganic anions, and fatty acids in sediment microcosms immediately after irradiation and afteraddition of a fresh G. sulfurreducens inoculuma
Treatment
Mean concn � SEM
BioavailableFe (mM)
NO3�
(mM)NO2
�
(mM)SO4
2�
(mM)Lactate(�M)
Acetate(�M)
Propionate(�M)
Butyrate(�M)
Formate(�M)
Fumarate(�M)b
Oxalate(�M)b
Nonirradiated 0.74 � 0.02 0.53 � 0.02 ND 0.40 � 0.01 ND ND ND ND ND ND ND0.5 Gy h�1 0.70 � 0.02 0.29 � 0.03 0.01 � 0.02 0.40 � 0.00 ND ND ND ND ND ND ND30 Gy h�1 0.81 � 0.03 0.13 � 0.01 0.16 � 0.01 0.42 � 0.01 3.5 � 3.0 57.2 � 70.0 3.9 � 0.4 1.1 � 0.9 32.7 � 18.1 45.5 � 13.7 59.5 � 17.9
a Errors indicate the standard errors of triplicate measurements. ND, not detected.b Fumarate and oxalate have identical retention times in the chromatography system used. The concentrations have therefore been determined for each based on their respectivemolecular masses.
Brown et al.
4022 aem.asm.org June 2015 Volume 81 Number 12Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

On the other hand, the enhanced Fe(III) reduction may also berelated to the removal of nitrate by radiolysis, as in the irradiationof sediments containing no added G. sulfurreducens cells or elec-tron donors. Nitrate concentrations in irradiated microcosms(0.13 mM in 30-Gy h�1 treatments and �0.3 mM in 0.5-Gy h�1
treatments) were significantly lower immediately after irradiationthan in nonirradiated systems (�0.5 mM). Again, these results areconsistent with radiation enhanced removal of nitrate and theearly onset of Fe(III) reduction, as discussed previously.
Implications for the geodisposal of radioactive waste. In thepresent study, we have highlighted microbial activities under doserates representative of gamma radiation emitted from radioactivewaste canister surfaces in the near field of a geological disposalfacility. We then assessed microbial activities under a simulatedrecovery period that would exist after significant radioactive decayhad occurred.
Previous studies suggested that microbial activity will be sup-pressed in these environments. For instance, studies of survival ofmicroorganisms from clay buffer material have suggested thattypically only 10% of the population survives after doses of �1.6kGy (9) and that the dose rate may not have a significant impact onthe viability of microbial populations (19). On the other hand,indigenous members of an endolithic microbial community froma proposed high-level radioactive waste repository may have beenable to survive in a nonculturable state after irradiation (9.34 kGyat 1.63 Gy min�1), to be rejuvenated when conditions becomefavorable (22, 23).
In contrast, the results presented here indicate that a sedimentcommunity can survive long-term gamma irradiation and thatcomponents of these communities can remain active and catalyzebiogeochemical processes, including Fe(III) reduction. We haveshown this to be the case for doses of up to �38 kGy using a lower,environmentally relevant dose rate of 30 Gy h�1. Indeed, the doserate had a strong influence on the community structure in systemswith or without added carbon. This demonstrates the importanceof acquiring low dose rate data, particularly since lower dose ratesmay allow species to respond via upregulating repair mechanisms(24) or adapting over the geological timescales involved.
Radiation led to significant changes in the microbial commu-nities, with fermentative bacteria, such as clostridia, dominant insystems with added carbon. Such changes may be important inenvironments where there is an excess of carbon substrates, suchas in cellulosic wastes (3). Despite this loss of diversity, theseresults suggest that Fe(III) reduction can still be an importantelectron accepting process in such sediments. Furthermore, in en-vironments with lower electron donor concentrations, an Fe(III)-reducing community may be selected by radiation. This may oc-cur both directly, by making Fe(III) more bioavailable throughradiation-induced changes to the mineralogy, or indirectly, byradiation-induced removal of other electron acceptors, such asnitrate, which may lead to the early onset of microbial Fe(III)reduction. Regardless, a relative increase in Fe(III)-reducing spe-cies was also observed in irradiated systems that did not displayenhanced Fe(III) reduction. These results have positive implica-tions for the geodisposal of radioactive waste, whereby the stimu-lation of an Fe(III)-reducing community by radiation mayenhance the reduction and subsequent precipitation of radionu-clides by direct enzymatic or indirect [e.g., biogenic Fe(II)-medi-ated] mechanisms. Furthermore, the oxidation of molecular hy-drogen by the radiolysis of water coupled to the enhanced
reduction of alternative electron acceptors by low-dose gammaradiation could provide the basis of a novel ecosystem in the deepbiosphere. Future studies will focus on the radiolysis of recalci-trant organic matter and the potential for enhanced carbon min-eralization by subsurface microbial communities. Further workwould be required to assess how these altered communities mayaffect the mobility of key radionuclides.
ACKNOWLEDGMENTS
This study was funded by a BBSRC studentship awarded to A.R.B. andCASE funding from Radioactive Waste Management, Ltd. Irradiationswere carried out by AMEC, Harwell, Oxfordshire, United Kingdom, andwe are grateful for the assistance of Victoria Smith and Alan Hollinrake.The work of Clare Thorpe in sediment collection and Alastair Bewsher inIC analysis is also greatly appreciated.
REFERENCES1. Nuclear Decommissioning Authority. 2010. Geological disposal: near-
field evolution status report NDA/RWMD/033. Nuclear Decommission-ing Authority, Harwell, Oxfordshire, United Kingdom.
2. Lloyd JR. 2003. Microbial reduction of metals and radionuclides. FEMS Mi-crobiol Rev 27:411–425. http://dx.doi.org/10.1016/S0168-6445(03)00044-5.
3. Glaus MA, Van Loon LR. 2008. Degradation of cellulose under alkalineconditions: new insights from a 12 year degradation study. Environ SciTechnol 42:2906 –2911. http://dx.doi.org/10.1021/es7025517.
4. Libert M, Bildstein O, Esnault L, Jullien M, Sellier R. 2011. Molecularhydrogen: an abundant energy source for bacterial activity in nuclearwaste repositories. Phys Chem Earth A B C 36:1616 –1623. http://dx.doi.org/10.1016/j.pce.2011.10.010.
5. Truche L, Berger G, Albrecht A, Domergue L. 2013. Abiotic nitratereduction induced by carbon steel and hydrogen: implications for envi-ronmental processes in waste repositories. Appl Geochem 28:155–163.http://dx.doi.org/10.1016/j.apgeochem.2012.10.010.
6. Lloyd JR, Renshaw JC. 2005. Microbial transformations of radionuclides:fundamental mechanisms and biogeochemical implications. Met IonsBiol Syst 44:205–240.
7. Lloyd JR, Sole VA, Van Praagh CVG, Lovley DR. 2000. Direct andFe(II)-mediated reduction of technetium by Fe(III)-reducing bacteria.Appl Environ Microbiol 66:3743–3749. http://dx.doi.org/10.1128/AEM.66.9.3743-3749.2000.
8. Lloyd JR, Yong P, Macaskie LE. 2000. Biological reduction and removalof Np(V) by two microorganisms. Environ Sci Technol 34:1297–1301.http://dx.doi.org/10.1021/es990394y.
9. Stroes-Gascoyne S, Lucht LM, Borsa J, Delaney TL, Haveman SA,Hamon CJ. 1994. Radiation resistance of the natural microbial popula-tion in buffer materials. MRS Online Proc Libr 353. http://dx.doi.org/10.1557/PROC-353-345.
10. Allard T, Calas G. 2009. Radiation effects on clay mineral properties.Appl Clay Sci 43:143–149. http://dx.doi.org/10.1016/j.clay.2008.07.032.
11. SKB. 2006. Buffer and backfill process report for the safety assessmentSR-Can. SKB Technical Report TR-06 –18. Svensk KärnbränslehanteringAB, Stockholm, Sweden.
12. Riley RG, Zachara JM, Wobber FJ. 1992. Chemical contamination onDOE lands and selection of contaminated mixtures for subsurface scienceresearch. U.S. Department of Energy, Washington, DC.
13. Fredrickson JK, Zachara JM, Balkwill DL, Kennedy D, Li SMW, Ko-standarithes HM, Daly MJ, Romine MF, Brockman FJ. 2004. Geomi-crobiology of high-level nuclear waste-contaminated vadose sediments atthe Hanford Site, Washington State. Appl Environ Microbiol 70:4230 –4241. http://dx.doi.org/10.1128/AEM.70.7.4230-4241.2004.
14. Niedrée B, Vereecken H, Burauel P. 2013. Radiation-induced impacts onthe degradation of 2,4-D and the microbial population in soil micro-cosms. J Environ Radioact 115:168 –174. http://dx.doi.org/10.1016/j.jenvrad.2012.08.008.
15. Ghosal D, Omelchenko MV, Gaidamakova EK, Matrosova VY,Vasilenko A, Venkateswaran A, Zhai M, Kostandarithes HM, Brim H,Makarova KS, Wackett LP, Fredrickson JK, Daly MJ. 2005. How radi-ation kills cells: survival of Deinococcus radiodurans and Shewanella one-idensis under oxidative stress. FEMS Microbiol Rev 29:361–375. http://dx.doi.org/10.1016/j.femsre.2004.12.007.
Microbial Communities and Gamma Radiation
June 2015 Volume 81 Number 12 aem.asm.org 4023Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

16. Du J, Gebicki JM. 2004. Proteins are major initial cell targets of hydroxylfree radicals. Int J Biochem Cell Biol 36:2334 –2343. http://dx.doi.org/10.1016/j.biocel.2004.05.012.
17. Daly MJ, Gaidamakova EK, Matrosova VY, Vasilenko A, Zhai M,Leapman RD, Lai B, Ravel B, Li SMW, Kemner KM, Fredrickson JK.2007. Protein oxidation implicated as the primary determinant of bacte-rial radioresistance. PLoS Biol 5:769 –779. http://dx.doi.org/10.1371/journal.pbio.0050092.
18. Daly MJ, Gaidamakova EK, Matrosova VY, Vasilenko A, Zhai M, Ven-kateswaran A, Hess M, Omelchenko MV, Kostandarithes HM, MakarovaKS, Wackett LP, Fredrickson JK, Ghosal D. 2004. Accumulation of Mn(II)in Deinococcus radiodurans facilitates gamma-radiation resistance. Science306:1025–1028. http://dx.doi.org/10.1126/science.1103185.
19. Lucht LM, Stroes-Gascoyne S. 1996. Characterization of the radiationand heat resistance of the natural microbial population in buffer materialsand selected pure cultures. Atomic Energy of Canada Limited, technicalrecord. TR-744/COG-96 –171. Atomic Energy of Canada, Ltd., ChalkRiver, Ontario, Canada.
20. Bruhn DF, Breckenridge CR, Tsang MN, Watkins CS, Windes WE,Roberto FF, Wright PJ, Pinhero PJ, Brey RR. 1999. Irradiation ofmicrobes from spent nuclear fuel storage pool environments. Global 99,Jackson Hole, WY.
21. Bruhn DF, Frank SM, Roberto FF, Pinhero PJ, Johnson SG. 2009.Microbial biofilm growth on irradiated, spent nuclear fuel cladding. JNuclear Mater 384:140 –145. http://dx.doi.org/10.1016/j.jnucmat.2008.11.008.
22. Pitonzo BJ, Amy PS, Rudin M. 1999. Effect of gamma radiation on nativeendolithic microorganisms from a radioactive waste deposit site. RadiatRes 152:64 –70. http://dx.doi.org/10.2307/3580050.
23. Pitonzo BJ, Amy PS, Rudin M. 1999. Resuscitation of microorganismsafter gamma irradiation. Radiat Res 152:71–75. http://dx.doi.org/10.2307/3580051.
24. Qiu XY, Daly MJ, Vasilenko A, Omelchenko MV, Gaidamakova EK,Wu LY, Zhou JZ, Sundin GW, Tiedje JM. 2006. Transcriptome analysisapplied to survival of Shewanella oneidensis MR-1 exposed to ionizingradiation. J Bacteriol 188:1199 –1204. http://dx.doi.org/10.1128/JB.188.3.1199-1204.2006.
25. Pedersen K. 1997. Microbial life in deep granitic rock. FEMS MicrobiolRev 20:399 – 414. http://dx.doi.org/10.1111/j.1574-6976.1997.tb00325.x.
26. Galès G, Libert M-F, Sellier R, Cournac L, Chapon V, Heulin T. 2004.Molecular hydrogen from water radiolysis as an energy source for bacte-rial growth in a basin containing irradiating waste. FEMS Microbiol Lett240:155–162. http://dx.doi.org/10.1016/j.femsle.2004.09.025.
27. Lin l-H, Slater GF, Sherwood Lollar B, Lacrampe-Couloume G, OnstottTC. 2005. The yield and isotopic composition of radiolytic H2, a potentialenergy source for the deep subsurface biosphere. Geochim CosmochimActa 69:893–903. http://dx.doi.org/10.1016/j.gca.2004.07.032.
28. Bank TL, Kukkadapu RK, Madden AS, Ginder-Vogel MA, Baldwin ME,Jardine PM. 2008. Effects of gamma-sterilization on the physico-chemicalproperties of natural sediments. Chem Geol 251:1–7. http://dx.doi.org/10.1016/j.chemgeo.2008.01.003.
29. Schaller J, Weiske A, Dudel EG. 2011. Effects of gamma-sterilization onDOC, uranium and arsenic remobilization from organic and microbialrich stream sediments. Sci Total Environ 409:3211–3214. http://dx.doi.org/10.1016/j.scitotenv.2011.05.014.
30. Ladrière J. 1998. Irradiation effects detected by Mössbauer spectroscopyin iron complexes. Hyperfine Interact 113:411– 418. http://dx.doi.org/10.1023/A:1012652522689.
31. Gournis D, Mantaka-Marketou AE, Karakassides MA, Petridis D. 2000.Effect of �-irradiation on clays and organoclays: a Mössbauer and XRDstudy. Phys Chem Miner 27:514 –521. http://dx.doi.org/10.1007/s002690000089.
32. Plötze M, Kahr G, Stengele RH. 2003. Alteration of clay minerals: gam-ma-irradiation effects on physicochemical properties. Appl Clay Sci 23:195–202. http://dx.doi.org/10.1016/S0169-1317(03)00103-0.
33. Daub K, Zhang X, Noel JJ, Wren JC. 2011. Gamma-radiation-inducedcorrosion of carbon steel in neutral and mildly basic water at 150 degrees.C Corros Sci 53:11–16. http://dx.doi.org/10.1016/j.corsci.2010.09.048.
34. Yakabuskie PA, Joseph JM, Keech P, Botton GA, Guzonas D, Wren JC.2011. Iron oxyhydroxide colloid formation by gamma-radiolysis. PhysChem Chem Phys 13:7198 –7206. http://dx.doi.org/10.1039/c1cp20084d.
35. Brown AR, Wincott PL, LaVerne JA, Small JS, Vaughan DJ, PimblottSM, Lloyd JR. 2014. The impact of � radiation on the bioavailability of
Fe(III) minerals for microbial respiration. Environ Sci Technol 48:10672–10680. http://dx.doi.org/10.1021/es503249r.
36. McNamara NP, Black HIJ, Beresford NA, Parekh NR. 2003. Effects ofacute gamma irradiation on chemical, physical and biological propertiesof soils. Appl Soil Ecol 24:117–132. http://dx.doi.org/10.1016/S0929-1393(03)00073-8.
37. Marschner B. 1993. Microbial contribution to sulphate mobilization afterliming an acid forest soil. J Soil Sci 44:459 – 466. http://dx.doi.org/10.1111/j.1365-2389.1993.tb00468.x.
38. Law GT, Geissler A, Boothman C, Burke IT, Livens FR, Lloyd JR,Morris K. 2010. Role of nitrate in conditioning aquifer sediments fortechnetium bioreduction. Environ Sci Technol 44:150 –155. http://dx.doi.org/10.1021/es9010866.
39. Thorpe CL, Law GT, Boothman C, Lloyd JR, Burke IT, Morris K. 2012.The synergistic effects of high nitrate concentrations on sediment bioreduc-tion. Geomicrobiol J 29:484–493. http://dx.doi.org/10.1080/01490451.2011.581332.
40. Thorpe CL, Lloyd JR, Law GTW, Burke IT, Shaw S, Bryan ND, MorrisK. 2012. Strontium sorption and precipitation behaviour during biore-duction in nitrate impacted sediments. Chem Geol 306-307:114 –122.http://dx.doi.org/10.1016/j.chemgeo.2012.03.001.
41. Wilkins MJ, Livens FR, Vaughan DJ, Beadle I, Lloyd JR. 2007. Theinfluence of microbial redox cycling on radionuclide mobility in the sub-surface at a low-level radioactive waste storage site. Geobiology 5:293–301.http://dx.doi.org/10.1111/j.1472-4669.2007.00101.x.
42. Lloyd JR, Leang C, Hodges Myerson AL, Coppi MV, Cuifo S, Methe B,Sandler SJ, Lovley DR. 2003. Biochemical and genetic characterization ofPpcA, a periplasmic c-type cytochrome in Geobacter sulfurreducens.Biochem J 369:153–161. http://dx.doi.org/10.1042/BJ20020597.
43. Stookey LL. 1970. Ferrozine: a new spectrophotometric reagent for iron.Anal Chem 42:779 –781. http://dx.doi.org/10.1021/ac60289a016.
44. Lovley DR, Phillips EJP. 1986. Availability of ferric iron for microbialreduction in bottom sediments of the freshwater tidal Potomac river. ApplEnviron Microbiol 51:751–757.
45. Lane DJ. 1991. 16S/23S rRNA sequencing, p 115–176. In Stackebrandt E,Goodfellow M (ed), Nucleic acid techniques in bacterial systematics.Wiley, Chichester, United Kingdom.
46. Hamady M, Walker JJ, Harris JK, Gold NJ, Knight R. 2008. Error-correcting barcoded primers for pyrosequencing hundreds of samples inmultiplex. Nat Methods 5:235–237. http://dx.doi.org/10.1038/nmeth.1184.
47. Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD,Costello EK, Fierer N, Pena AG, Goodrich JK, Gordon JI, Huttley GA,Kelley ST, Knights D, Koenig JE, Ley RE, Lozupone CA, McDonald D,Muegge BD, Pirrung M, Reeder J, Sevinsky JR, Turnbaugh PJ, WaltersWA, Widmann J, Yatsunenko T, Zaneveld J, Knight R. 2010. QIIMEallows analysis of high-throughput community sequencing data. NatMethods 7:335–336. http://dx.doi.org/10.1038/nmeth.f.303.
48. Edgar RC. 2010. Search and clustering orders of magnitude faster thanBLAST. Bioinformatics 26:2460 –2461. http://dx.doi.org/10.1093/bioinformatics/btq461.
49. Ishii N, Fuma S, Tagami K, Honma-Takeda S, Shikano S. 2011. Re-sponses of the bacterial community to chronic gamma radiation in a ricepaddy ecosystem. Int J Radiat Biol 87:663– 672. http://dx.doi.org/10.3109/09553002.2010.549534.
50. Lin L-HH, Wang P-LL, Rumble D, Lippmann-Pipke J, Boice E, PrattLM, Sherwood Lollar B, Brodie EL, Hazen TC, Andersen GL, DeSantisTZ, Moser DP, Kershaw D, Onstott TC. 2006. Long-term sustainabilityof a high-energy, low-diversity crustal biome. Science 314:479 – 482. http://dx.doi.org/10.1126/science.1127376.
51. Lefticariu L, Pratt LA, LaVerne JA, Schimmelmann A. 2010. Anoxicpyrite oxidation by water radiolysis products: a potential source of biosus-taining energy. Earth Planet Sci Lett 292:57– 67. http://dx.doi.org/10.1016/j.epsl.2010.01.020.
52. Seeliger S, Janssen PH, Schink B. 2002. Energetics and kinetics of lactatefermentation to acetate and propionate via methylmalonyl-CoA or acry-lyl-CoA. FEMS Microbiol Lett 211:65–70. http://dx.doi.org/10.1111/j.1574-6968.2002.tb11204.x.
53. Holmes DE, Nevin KP, Woodard TL, Peacock AD, Lovley DR. 2007.Prolixibacter bellariivorans gen. nov., sp. nov., a sugar-fermenting, psy-chrotolerant anaerobe of the phylum Bacteroidetes, isolated from a ma-rine-sediment fuel cell. Int J Syst Evol Microbiol 57:701–707. http://dx.doi.org/10.1099/ijs.0.64296-0.
Brown et al.
4024 aem.asm.org June 2015 Volume 81 Number 12Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

54. Imachi H, Sekiguchi Y, Kamagata Y, Loy A, Qiu YL, Hugenholtz P,Kimura N, Wagner M, Ohashi A, Harada H. 2006. Non-sulfate-reducing, syntrophic bacteria affiliated with Desulfotomaculum cluster Iare widely distributed in methanogenic environments. Appl Environ Mi-crobiol 72:2080 –2091. http://dx.doi.org/10.1128/AEM.72.3.2080-2091.2006.
55. Moat AG, Foster JW, Spector MP. 2002. Microbial physiology, 4th ed.Wiley-Liss, New York, NY.
56. Imachi H, Sakai S, Ohashi A, Harada H, Hanada S, Kamagata Y, SekiguchiY. 2007. Pelotomaculum propionicicum sp. nov., an anaerobic, mesophilic,obligately syntrophic propionate-oxidizing bacterium. Int J Syst Evol Mi-crobiol 57:1487–1492. http://dx.doi.org/10.1099/ijs.0.64925-0.
57. Geissler A, Law GTW, Boothman C, Morris K, Burke IT, Livens FR,Lloyd JR. 2011. Microbial communities associated with the oxidation ofiron and technetium in bioreduced sediments. Geomicrobiol J 28:507–518. http://dx.doi.org/10.1080/01490451.2010.515287.
58. Meier J, Piva A, Fortin D. 2012. Enrichment of sulfate-reducing bacteriaand resulting mineral formation in media mimicking pore water metal ionconcentrations and pH conditions of acidic pit lakes. FEMS MicrobiolEcol 79:69 – 84. http://dx.doi.org/10.1111/j.1574-6941.2011.01199.x.
59. Spring S, Merkhoffer B, Weiss N, Kroppenstedt RM, Hippe H, Stacke-brandt E. 2003. Characterization of novel psychrophilic clostridia from anAntarctic microbial mat: description of Clostridium frigoris sp. nov., Clos-tridium lacusfryxellense sp. nov., Clostridium bowmanii sp. nov. and Clos-tridium psychrophilum sp. nov. and reclassification of Clostridium lara-miense as Clostridium estertheticum subsp. laramiense subsp. nov. Int J SystEvol Microbiol 53:1019 –1029. http://dx.doi.org/10.1099/ijs.0.02554-0.
60. Wiegel J, Tanner R, Rainey FA. 2006. An introduction to the familyClostridiaceae, p 654 – 678. In Dworkin MM, Falkow S, Rosenberg E,Schleifer K-H, Stackebrandt E (ed), The prokaryotes. Springer, NewYork, NY.
61. Pester M, Brambilla E, Alazard D, Rattei T, Weinmaier T, Han J, LucasS, Lapidus A, Cheng J-F, Goodwin L, Pitluck S, Peters L, OvchinnikovaG, Teshima H, Detter JC, Han CS, Tapia R, Land ML, Hauser L,Kyrpides NC, Ivanova NN, Pagani I, Huntmann M, Wei C-L, Daven-port KW, Daligault H, Chain PSG, Chen A, Mavromatis K, MarkowitzV, Szeto E, Mikhailova N, Pati A, Wagner M, Woyke T, Ollivier B,
Klenk H-P, Spring S, Loy A. 2012. Complete genome sequences ofDesulfosporosinus orientis DSM765T, Desulfosporosinus youngiaeDSM17734T, Desulfosporosinus meridiei DSM13257T, and Desulfosporosi-nus acidiphilus DSM22704T. J Bacteriol 194:6300 – 6301. http://dx.doi.org/10.1128/JB.01392-12.
62. Daniels M, Wigg EE. 1969. Radiation chemistry of the aqueous nitratesystem: II. Scavenging and pH effects in the cobalt-60 gamma radiolysis ofconcentrated sodium nitrate solutions. J Phys Chem 73:3703–3709.
63. Finneran KT, Johnsen CV, Lovley DR. 2003. Rhodoferax ferrireducens sp.nov., a psychrotolerant, facultatively anaerobic bacterium that oxidizesacetate with the reduction of Fe(III). Int J Syst Evol Microbiol 53:669 –673. http://dx.doi.org/10.1099/ijs.0.02298-0.
64. Vardhan Reddy PV, Shiva Nageswara Rao SS, Pratibha MS, Sailaja B,Kavya B, Manorama RR, Singh SM, Radha Srinivas TN, Shivaji S. 2009.Bacterial diversity and bioprospecting for cold-active enzymes from cul-turable bacteria associated with sediment from a melt water stream ofMidtre Loenbreen glacier, an Arctic glacier. Res Microbiol 160:538 –546.http://dx.doi.org/10.1016/j.resmic.2009.08.008.
65. Coates JD, Ellis DJ, Gaw CV, Lovley DR. 1999. Geothrix fermentans gen.nov., sp. nov., a novel Fe(III)-reducing bacterium from a hydrocarbon-contaminated aquifer. Int J Syst Evol Microbiol 49:1615–1622.
66. Nakai R, Abe T, Baba T, Imura S, Kagoshima H, Kanda H, Kanekiyo A,Kohara Y, Koi A, Nakamura K, Narita T, Niki H, Yanagihara K,Naganuma T. 2012. Microflorae of aquatic moss pillars in a freshwaterlake, East Antarctica, based on fatty acid and 16S rRNA gene analyses.Polar Biol 35:425– 433. http://dx.doi.org/10.1007/s00300-011-1090-2.
67. Kirchhof G, Eckert B, Stoffels M, Baldani JI, Reis VM, Hartmann A.2001. Herbaspirillum frisingense sp. nov., a new nitrogen-fixing bacterialspecies that occurs in C4-fibre plants. Int J Syst Evol Microbiol 51:157–168.
68. Lynch JM. 1982. Limits to microbial growth in soil. J Gen Microbiol128:405– 410.
69. Marschner B, Bredow A. 2002. Temperature effects on release and eco-logically relevant properties of dissolved organic carbon in sterilised andbiologically active soil samples. Soil Biol Biochem 34:459 – 466. http://dx.doi.org/10.1016/S0038-0717(01)00203-6.
Microbial Communities and Gamma Radiation
June 2015 Volume 81 Number 12 aem.asm.org 4025Applied and Environmental Microbiology
on July 17, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from





























