Embed Size (px)
DESCRIPTION
There is no damage to describe.Hans Brunner's signature of the top right-hand corner of the front end page along with a letter signed by the author on Department of Conservation, Forests and Lands letterhead. This book is designed to interest the reader to the general field of their study and, in particular to provide a workable system of mammal identification based entirely on an appreciation of their structures. – Whilst many of the species considered in this book are indigenous to Australia, the methods and principles discussed are applicable to any situation requiring the identification of unknown fur samples. – Front fold over blurb Size: 4to 9¾" - 12" tall. 176 pages. Please refer to the accompanying picture(s) Illustrator: Microscopic. Quantity Available: 1. Category: Science & Technology; biology; ISBN: 0909605017. ISBN/EAN: 9780909605018.
Citation preview
I I I
Ii
• I
e
I
Hans Brunner and Brian Coman

--~. ----~"--~-,~: ~.::::-~~-- ~---,-. -, "~~---~-~~-~--,-~- ... "--.~---
(likata Press Proprietary Limited Melbourne
First puhlished 1974
© H. BRUNNER & B. COMAN
National Library of Australia Card Number and ISBN 0 909605 01 7
Library of Congress Catalogue Card Number 74fi79009
All rights strictly reserved. No part of this book may be reproduced in any form without theJ'ermission in writing of the publisher.'
Designed by Arthur Stohs
Text set in Monotype Times Roman by Monotrade Pty Ltd Melbourne, Victoria
Printed by Shanghai Printing Press Ltd.~ Hong Kong
I Ii Contents
I I I
I i
Foreword vii
Preface ix
Acknowledgements ix
SECTION A
SECTION B
1. Introduction General Types of hair 2 The hair profile 3 Aspects of hair growth 4
2. The structure of hairs 5 Cross-sectional appearance 5 The medulla 5 The cortex 8 The cuticle 8 Hair pigmentation 10
1. Methods of studying hair structure 13 The wbole mount 13 Obtaining the Cross-section 13 Obtaining cuticular scale casts 14 Photographing hair structures 16
2. A system for bair identification 17 Producing the system 17 Using the system 18
SECTION C 1. A guide to hai, structure for the indigenous and introduced
mammals of Victoria 19
2. A grouping of Victorian mammals hased initially on the cross-sectional appearance of primary guard hairs 20
3. Photographs and descriptions of hairs for each species involved 22 Bibliography 173 Index to Species 175

SECTION A
1 Introduction
GENERAL
There are many situations where mammal identification is most conveniently based on a consideration of hair structure. The identification of ingested hairs is a particularly useful aid in the determination of diet for wild carnivorous animals, as most meals of mammal origin would include at least a small quantity of hair. Bones and other indigestible items are often avoided. Ingested bones, with the possible exception of those from very small prey species, are generally fragmented and of little diagnostic value. By contrast, hairs suffer much less damage during the digestive processes, and even hairs recovered from faeces retain many of the features useful for identification. Indeed, faecal analysis is now a widely used teChnique in food habits and dietary studies.
The hair from a number of mammals is commercially utilized and a knowledge of hair structure is necessary in certain sections of the textile and allied industries. Hair identification may also play some part in the field of forensic science and the policing of wildlife laws. In mammal survey investigations, hair traces on twigs, fences, etc. may weI! provide the only clue to the presence of a particular mammal in the study area.
Although the history of hair structure studies related to mammal identification may be traced back to the late nineteenth century, the first significant contribution in this field was the work of Hausman (1920,1924,1930) in America. Using written descriptions and somewhat simplified drawings, Hausman presented data on the configuration of cuticular scales and medullae for the different hairs of 166 fur-bearing mammals. However, he made no attempt to provide any key as an identification aid and relied solely on a comparison between known and unknown samples.
In the following decade, Mathiak (1938) produced a key to the identification of hairs of the mammals of southern Michigan. The key was based partly on an appreciation of cross-sectional appearance and medulla arrangement, but also relied heavily on measurements of hair diameter and hair length. The major shor1eoming of Mathiak's key stems from his heavy dependence on these hair measurements as diagnostic aids. In the same year, Williams (1938) produced a key to the identification of hairs of moles and shrews.
At about this time, technological advances in the textile industry prompted detailed hair structure studies by the Wool Industries Research Association at Leeds, England. Wildman (1940) published a booklet on animal fibres of industrial importance and discussed their origin and identification. This work was greatly enlarged and repUblished in 1954. This publication has generally been accepted as the definitive work on hair structure. Understandably, Wildman's investigations centred about a detailed description of fibres of commercial importance, and no attempt was made to produce a key or system of identification for unknown hairs. A further account of this work was giv<ln by Appleyard (1960) in his Guide to the identification of animal fibres.
A descriptive key to the identification of hair of Californian mammals .was published by Mayer (1952). The entire key was based on a consideration of dorsal guard hairs which had been taken from one small area on the pelage. In all, some 392 species and subspecies were considered. Although this key was well devised, it relied heavily on the value of hair length and general profile as a diagnostic aid and ignored the value of cross·sectional shape and scale pattern.
Studies on hair structure of Tasmanian marsupials and monotremes by Lyne and McMahon (1951) suggested
,
THE IDENTIFICATION OF MAMMALIAN HAIR
that a detailed microscopic examination of hairs of these mammals would be of some taxonomic value. In this preliminary investigation they made no attempt to produce a key to the hair of the spe<;ies involved. A detailed study, dealing entirely with the hair of the Chiroptera, was reported by Benedict (1957). Her work involved a description of hair structure for 16 families and also included a key to the dorsal hair of the Chiroptera, In more recent years studies On the diagnostIc value of hairs have been reported by Day (1966) and Adorjan and Kolenosky (1969).
These and many other studies have contributed geeatly to our knowledge of mammalian hair and have de";'onstrated the use of hair structure as a diagnostic aid. Despite this, there stil! remain a number of problems associated with the identification of unknown hairs.
. These inolude the following:
1. the considerable variety of hair types encountered, even on an indIvidual mammal;
2. variations in hair structure along the length of an individual hair;
3. an appreciable degree of inter-species overlap, in particular characteristics, limiting the usefulness of some structure;,s as diagnostic aids, and
4. slow methods of study for some aspects of hair structure which are used as 'diagnostic aids.
" It is the aim of this book to present a particular approach to hair identification, which attempts to minimize these problems. The major problem of inter-species overlap is reduced by the use of a photographic reference system. Although many of the mammals represented in this photographic system (Section C) are endemic to Australia, the general principles and methods outlined could be expected to apply to any situation demanding a rapid system for the routine identification of large numbers of hair samples. The book has been primarily designed with the needs of the food-habits investigator in mind.
TYPES OF HAIR
For the purposes of determining the identity of a particular hair sample it is useful to recognize the following "types" of hair:
Vibrissae These are large, stiff hairs that are primarily sensory in function. They are variously referred to as whiskers, sensory hairs~ tactile hairs and sinus hairs. Generally speaking, the vibrissae are widest in their proximal half and taper to a long tip. Lyne (1959) has noticed that, in the Marsupialia, vibrissae have some
2
systematic significance. However, for the purposes of identifying unknown hair samples, vibrissae are of no value as their basic structure is very similar for all mammals.
Bristle hairs On some mammals, such as certain breeds of the domestic pig, the whole pelage is composed of rather stout and rigid hairs which are of uniform diameter along most of their length. Such hairs usually have a very narrOw medulla, Or no medulla at all (see page 36), are oval to circular in cross-section, and commonly have a flagged tip. Although Noback (1951) and others consider the term "'bristleH to be synonymous with protective hair, primary hair and overhair, it is useful for our purposes to recognize bristles as being different from the coarse hairs mentioned below.
Over/wirs A close inspection of most mammal pelages will show that some sparsely scattered hairs are distinctly longer than those comprising the bulk of the pelage. These are termed overhairs. Usually they are circular in cross-section and have little diagnostic value. However, in those cases where their cross-SL"Ctional shape is not circular, they may be useful in identification. As a rule, overhairs are mOre densely pigmented than the other hair types immediately surrounding them.
Guard hairs These are the larger or coarser of those hairs forming the main pelage and include a type often described as shield hairs. In shield hairs the distal part' is noticeably wider and flattened, forming a shield. Guard hairs lacking a shield are of uniform diameter along most of their length, tapering only towards the tip.
The largest of the guard hairs, conveniently termed the primary guard hairs,. are of paramount importance in hair identification, for it is these hairs which generally exhibit the most diagnostically useful features. There is usually a variety of sizes for guard hairs in anyone pelage, ranging down until they are indistinguishable from the underhairs (see below).
Underhairs These are shorter and finer than the guard hairs on any given area of the pelage and are commonly wavy. With few exceptions, they retain the same thickness along their whole length (except the tip). Like the guard hairs, they range in size down to extremely fine hair which is sometimes called veUus or lanugo (Danforth, 1939). Underhairs are generally of little diagnostic value.
h should be pointed out that some mammals have a much wider range of hair types than do others. For instance; in the main pelage, rodents have more types of hair than do the more common breeds of sheep. Furthermore, the relative abundance of overhairs,
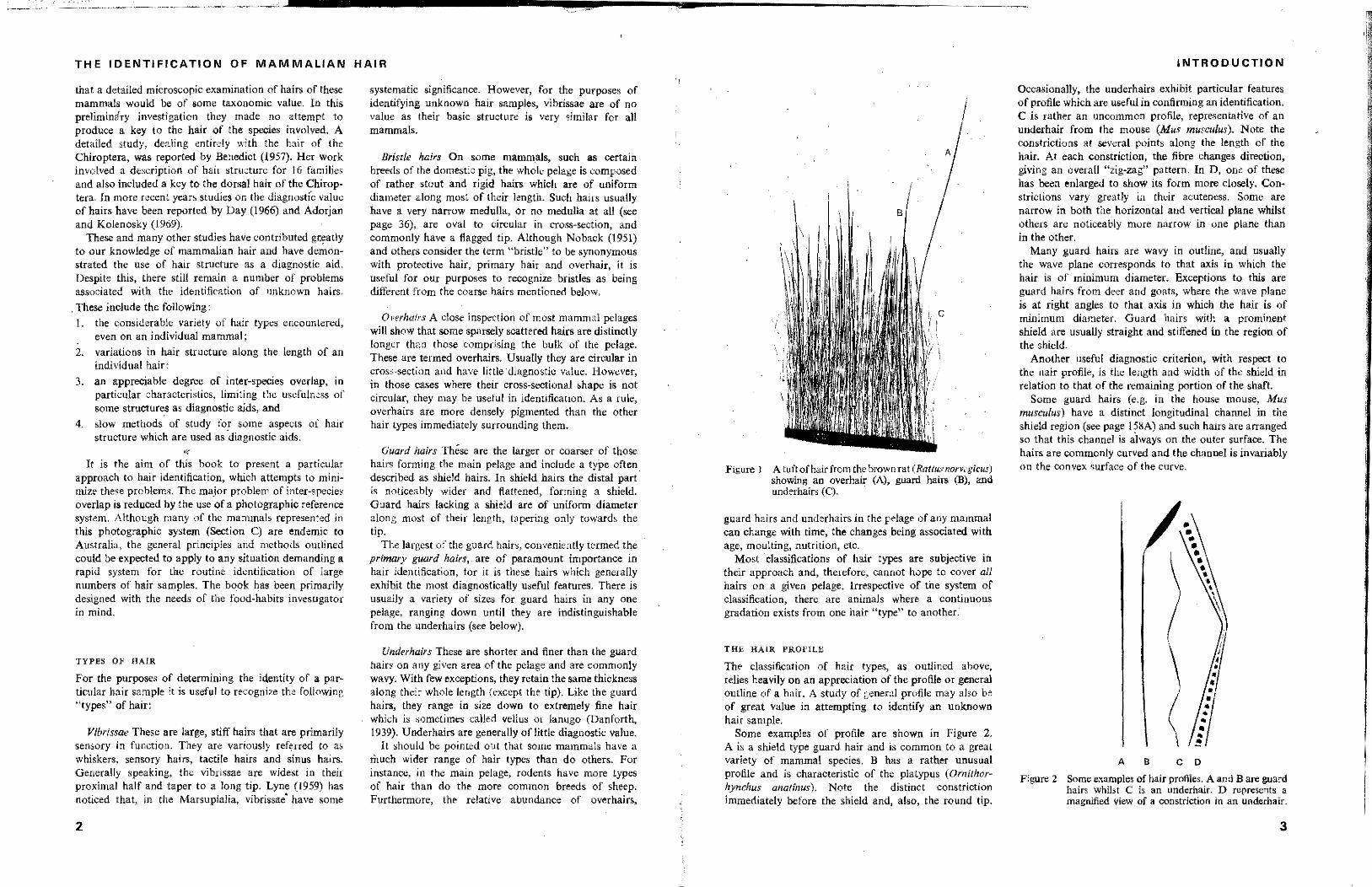
FIgure J A tuftofhairfrom the brown rat (Rallusnorveg<'cus) showing an overhair (A), guard hairs (B), and underhairs (C).
guard hairs and underhairs in the pelage of any mammal can change with time, the changes being associated with age, moulting, nutrition, etc.
Most classifications of hair types are subjective in their approach and, therefore, cannot hope to cover all hairs on a given pelage. Irrespe<;tive of the system of classification. there are animals where a continuous gradation exists from one hair "type" to another.
THE HAlR PROFILE
The classification of hair types, as outlined above, relies heavily on an appreciation of the profile or general outline of a hair. A study of general profile may also be of great value in attempting to identify an unknown hair sample.
Some examples of profile are shown in Figure 2. A is a shield type guard hair and is commOn to a great variety of mammal species. B has a rather unusual profile and is characteristic of the platypus (OrnUhorhynchus anatinus). Note the distinct constriction immediately before the shield and, also, the round tip.
INTRODUCTION
Occasionally, the underhairs exhibit particular features of profile which are useful in confirming an identification. C is rather an uncommon profile, representatiVe of an underhair from the mouse (Mus musculus). Note the constrictions at several points along the length of the hair. At each constriction, the fibre changes direction, giving an overall "zig-zag" pattern. In D, one of these has been enlarged to show its form more closely. Constrictions vary greatly in their acuteness. Some are narrow in both the horizontal and vertical plane whilst others are noticeably more narrow in one plane than in the other.
Many guard hairs are wavy in outline, and usually the wave plane corresponds to that axis in which the hair is of minimum diameter. Exceptions to this are guard hairs from deer and goats, where the wave plane is at right angles to that axis in which the hair is of minimum diameter. Guard hairs with a prominent shield are usually straight and stiffened in the region of the shield.
Another useful diagnostic criterion, with respect to the hair profile, is the length and width of the shield in relation to that of the remaining portion of the shaft
Some guard hairs (e.g. in the house mouse, Mus musculus) have a distinct long; tumnal channel in the shield region (see page 15BA) and such hairs are arranged so that this channel is always on the outer surface. The hairs are commonly curved and the channel is invariably on the convex surface of the curve.
Figure 2
A B
\ ~
C D
Some examples of hair profiles. A and B are guard hairs whilst C is an underhair. D represents a magnIfied view of a constriction in an underhair.
3
~""~~<-<~~-==:::::::--------------~-----------------------
THE IDENTIFICATION OF MAMMALIAN HAIR
Cross~sectlons A-A --_.
Growth Stages
A_
,
A
A
fo.- _A
A
3 •
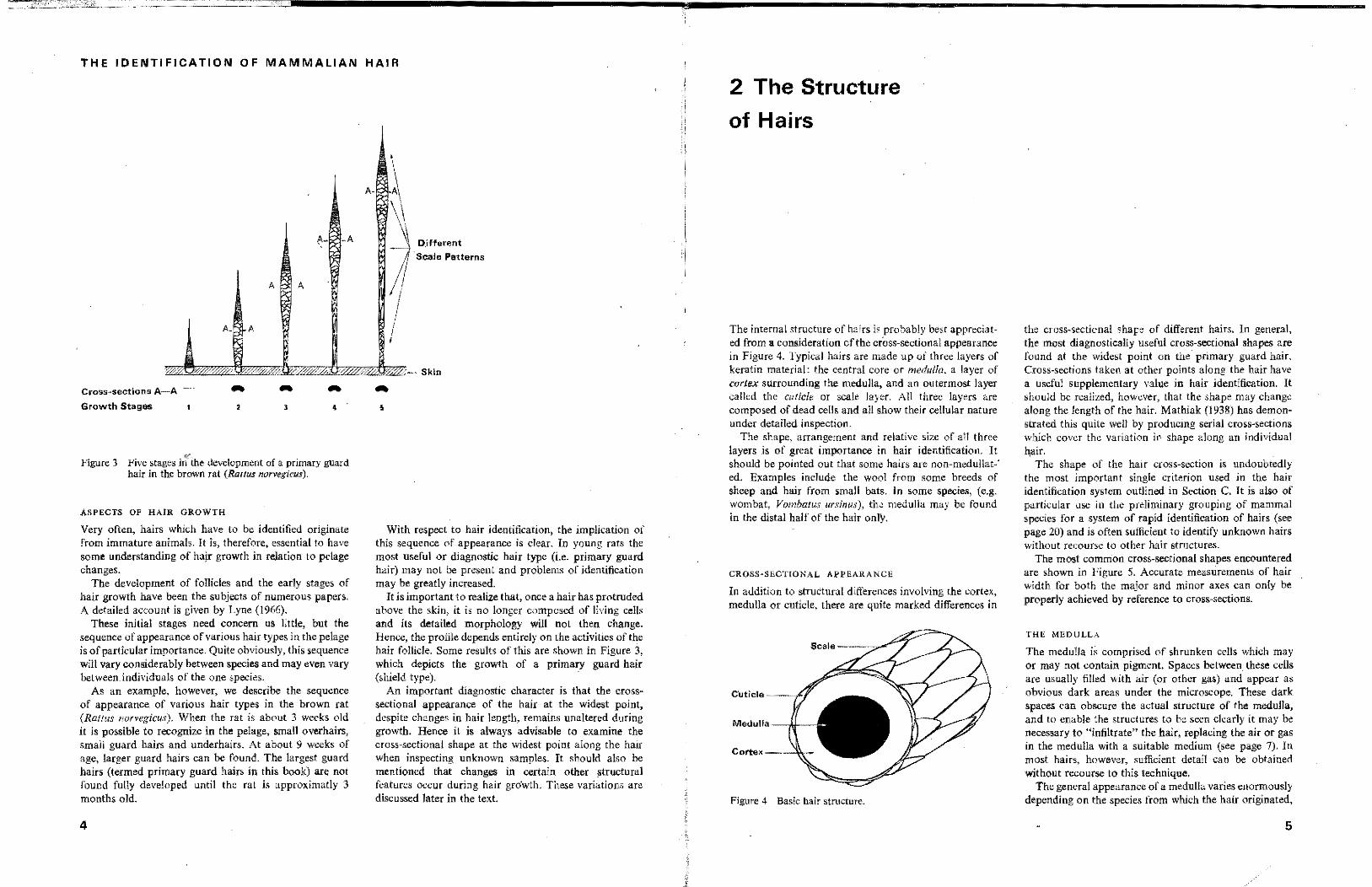
Figure 3 Five stages iI~ the development of a primary guard hair in the brown rat (Rattus narvcgicus),
ASPECTS OF HAIR GROWTH
Very often, hairs which have to be identified originate from immature animals, It is~ therefore? essential to have some understanding of hajr groMh in relation to pelage changes.
The development of follicles and the early stages of hair growth have been the subjects of numerous papers. A detailed account is given by Lyne (1966).
These initial stages need concern us little, but the sequence of appearance of various hair types in the pelage is of particular importance. Quite obviously, this sequence will vary considerably between species and may even vary between< individuals of the one species.
As an example, however, we describe the sequence of appearance of various hair types in the brown rat (Rallus norvegicus). When the rat is about 3 weeks old it is possible to recognize in the pelage, small overhairs, small guard hairs and underhairs. At about 9 weeks of age, larger guard hairs can be found. The largest guard hairs (termed primary guard hairs in this book) are not found fully developed until the rat is approximatly 3 months old.
4
A.
, ,
1 ,
~ i Djfferent
Scah!! Patterns
/1 I I , I I I
Skin
•
With respect to hair identification, the implication of this sequence of appearance is clear. In young rats the most useful or diagnostic hair type (Le. primary guard hair) may not be present and problems of identification may be greatly increased.
It is important to realize that, once a hair has protruded above the skin, it is no longer composed of living cells and its detailed morphology will not then change. Hence, the profile depends entirely on the activities of the hair follicle. Some results of this are shown in Figure 3, which depicts the growth of a primary guard hair (shield type). .
An important diagnostic character is that the crosssectional appearance of the hair at the widest point, despite changes in hair length, remains unaltered during growth. Hence it is always advisable to examine the cross-sectional shape at the widest point along the hair when inspecting unknown samples. It should also be mentioned that changes in certain other structuml features occur during hair gro'wth. These variations are discussed later in the text.
2 The Structure
of Hairs
The internal structure of hairs is probably best appreciated from a consideration of the cross-sectional appearance in Figure 4. Typical hairs are made up of three layers of keratin material: the central core or medulla, a layer of cortex surrounding the medulla, and an outermost layer called the cuticle or scale layer. All three layers are composed of dead cells and all show their cellular nature under detailed inspection.
The shape, arrangement and relative size of all thrce layers is of great importance in hair identification. It should be pointed out that some hairs are non-medullat-' ed. Examples include the wool from some breeds of sheep and hair from small bats. In some species, (e.g. wombat, Vombatus ursin"s), the medulla may be found in the distal half of the hair only.
CROSS~SECTIONAL APPEARANCE
In addition to struCtural differences involving the cortex, medulla or cuticle, there are quite marked differences in
Cuticle ---.<'J
Medulla ---/tI--
Cortex----'i~
Figure 4 Basic hair structure.
the cross-sectional shape of different hairs. In general, the most diagnostically useful cross-sectional shapes are found at the widest point on the primary guard hair. Cross-sections taken at other points along the hair have a useful supplementary value in hair identification. It should be realized, however, that the shape may change along the length of the hair. Mathiak (1938) has demonstrated this quile well by producing serial cross-sections which cover the variation in shape along an individual hair.
The shape of the hair cross-section is undoubtedly the most important single criterion used in the hair identification system outlined in Section C It is also of particular use in the preliminary grouping of mammal species for a system of rapid identification of hairs (see page 20) and is often sufficient to identify unknown hairs without recourse to other hair structures.
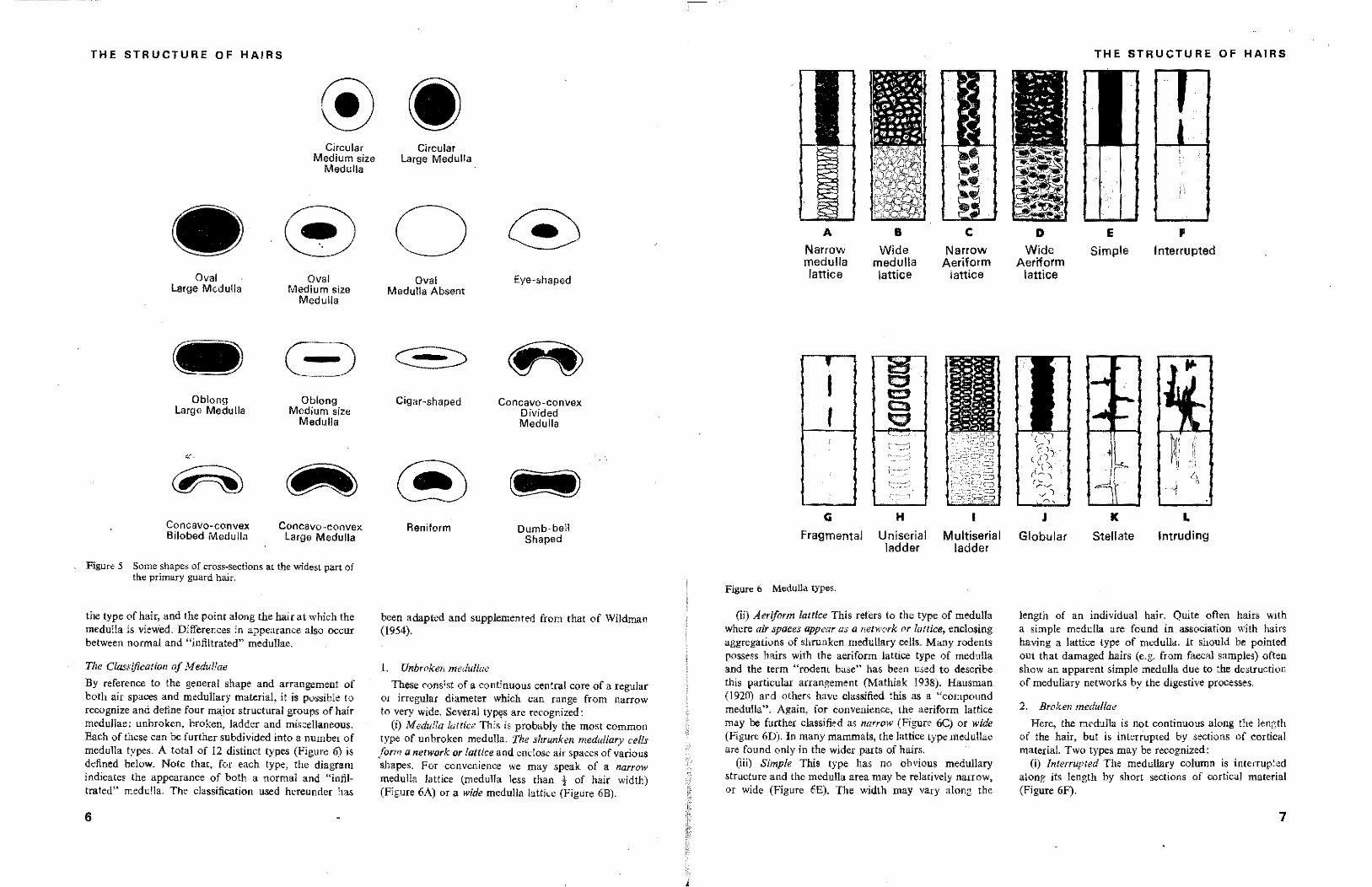
The most common cross~sectional shapes encountered are shown in Figure 5. Accurate measurements of hair width for both the major and minor axes can only be properly achieved by reference to cross-sections.
THE MEDULLA
The medulla is comprised of shrunken ceUs which may or may not contain pigment. Spaces between these ceUs are usually filled with air (or other gas) and appear as obvious dark areas under the microscope. These dark spaces can obscure the actual structure of the medulla, and to ~'Ilable the structures to be seen clearly it may be necessary to "infiltrate~' the hair, replacing the air or gas in the medulla with a suitable medium (see page 7). In most hairs, however, sufficient detail can be obtained without recourse to this technique.
The general appeamnce of a medulla varies enormously depending on the species from which the hair originated,
5
THE STRUCTURE OF HAIRS
Circular Medium size
Medulla
Circular Large Medulla
®o Oval
Large Medulla
Oblong Large Medulla
·r
~ Concavo~convex B ilobed Medu lIa
Oval Medium size
Medulla
Oblong Medium size
Medulla
~ Concavo -convex Large Medu lIa
Figure 5 Some shapes of cross:-sections at the widest part of the primary guard hair.
the type of hair, and the point along the hair at which the medulla is vievied. Differences in appearance also occur between normal and "infiltrated" medullae.
The Classification oj Medullae
By reference to the general shape and arrangement of both air spaces and medullary material, it is possible to recognize and define four major structural groups of hair medullae: unbroken, broken, ladder and miscellaneous. Each of these can be further subdivided into a number of medulla types. A total of 12 distinct types (Figure 6) is defined below. Note that, for each type, the diagram indicates the appearance of both a normal and "inftltrated" medulla. The classification used hereunder has
6
Oval Medulla Absent
Cigar-shaped
® Reniform
Eye-shaped
Concavo-convex Divided Medulla
~ Dumb-bell
Shaped
been adapted and supplemented from that of Wildman (1954).
I. Unbroken medullae
These consist of a continuous central core of a regular or irregnlar diameter which can range from narrow to very wide. Several typ<?S are recognized:
(I) Medulla Iotrice This is probably the most Common type of unbroken medulla. The shrunken medullary cells form a network or lattice and enclose air spaces of various shapes. For convenience we may speak of a narrow medulla lattice (medulla less than 1 of hair width) (Figure 6A) or a wide medulla lattice (Figure 6B).
A
Narrow medulla lattice
, I t
.
G
B
Wide medulla lattice
H
Fragmental Uniserial ladder
Figure 6 Medulla types.
C
Narrow Aeriform
lattice
Multiserial ladder
(ii) Aeriform lattice This relers 10 the type of medulla where air spaces appear as a network or lattice, enclosing aggregations of shrunken medullary cells. Many rodents possess hairs with the aeriform lattice type of medulla and the term "rodent base" has been used to describe this particular arrangement (Mathiak 1938). Hausman (1920) and otbers have classified this as a "compound medulla". Again, for convenience, the aeriform lattice may be further classified as narrow (Fignre 6C) or wide (Figure 60). In many mammals, the lattice type medullae are found only in the wider parts of hairs.
(iii) Simple This type has no obvious medullary structure and the medulla area may be relatively narrow, or wide (Figure 6E). The width may vary along the
THE STRUCTURE OF HAIRS
D Wide
Aeriform lattice
J
Globular
E
Simple
K
Stellate
F
Interrupted
L
Intruding
length of an individual hair. Quite often hairs with a simple medulla are found in association with hairs having a lattice type of medulla. It should be pointed out that damaged hairs (e.g. from faecal samples) often show an apparent simple medulla due to the destruction of medullary networks by the digestive processes.
2. Broken medullae
Here, the medulla is not continuous along the length of the hair, but is interrupted by sections of cortical materiaL Two types may be recognized:
(I) Interrupted The medullary column is interrupted along its length by short sections of cortical material (Fignre 6F).
7

THE IDENTIFICATION OF MAMMALIAN HAIR
(ii) Fragmented In this type the medullary column is interrupted by long sections of cortical tissue, The isolated medulla sections show little differentiation and their structure could be termed simple (Figure 6G).
3, Ladder medullae
This classification includes those medullae having one' or more rows of air spaces. Two distinct types can be defined: .
(i) Uniserialladder The medulla has a single row of air spaces and these can be angular, rounded, flattened or cup-shaped (Figure 6H). This type is .usually found in underhairs and the proximal part of Some oU,er hair types,
(li) Multiserial ladder The medulla has tWo or more distinct rows of mostly uniform air spaces (Figure 61), Such a med,ulla is characteristic of lagomorph hairs,
4, Miscellaneous medullae
These are generally uncommon and represented in the main pelage of only a small number of mammals or restricted to hairs taken from extremities (e.g, tail), The following types may be recognized:
(i) Globular This type has an aggregation of globular air spaces (Figure 6J). It may be found in SOme hairs from the platypus (Ornillwrhynchus analiflus), wombat (Vambatus ursinus) and fur seal (Arctacephalus doriferus).
(ii) Stellate Here, an essentially "simple" medulla has finger-like projectiorts radiating out into the cortex (Figure 6K). This type is often found in the tail hairs of larger mammals, and is usually best appreciated in cross-sections.
(iii) intruding This type has narrow and irregularly placed air spaces projecting into the cortex. The spaces may project in several directions and are not necessarily found in the centre of the hair (Figure 6L),
THE CORTEX
Like the medulla, the cortex of the hair is composed of dead cells, but the individual cells are usually not visible under the light microscope since they are packed into a rigid and almost homogeneous hyaline mass (Hausman 1932), Due to a high degree of cornification, the cortex has a low reftactive index and, in the absence of pigment, is translucent. It is sometimes possible (e,g, by retting wool fibres) to discern outlines of individual cortical cells, and these are generally polygonal in shape. However, few data are available on the way in which cortical cells are arranged in various types of hair,
.Because the cortex lacks discernible structure, under the light microscope, it is of little value for identification purposes, However, in those hairs where the cortex is pigmented, the size and arrangement of pigment granules
8
may be an important diagnostic criterion, Very often too, the ratio of cortex width to medulla width (shown in cross-section) can be of use in the identification process.
THE CUTICLE
The cuticle or outer layer of a hair consists' of a single layer of generally transparent, overlapping scales. In rare cases, the scales may be pigmented (e.g. some Chiroptera-Benedict 1957), Along most of the hair length, scales are flattened against the body of the hair, with their free edges pointing towards the hair tip. At the base of the hair, however, scales may be more "open" in arrangement and a cross-section in this region shows the cuticle to have a rippled or jagged outline, with each ripple corresponding to an individual seale, In this sense, the surface of the hair may be likened to that of a pine cone, where seed receptacles near the base of the cone are partly opened and those towards the tip are tightly closed,
An unusual arrangement of scales can be observed on hairs originating from the foot pad of some lagomorphs. Here, the scales are positioned at a considerable angle to the long axis of the hair and are tightly packed. This has the effect of producing an extremely thick cuticular layer, the advantage of which is obvious in such a situation,
The size and shape of scales, and their pattern of arrangement around the hair, are useful criteria for identification purposes, In relation to scale size, a measurement termed the scale index has been used by some workers (Hausman 1930, Mayer 1952). This can be defined as the ratio of the free proximodistal length of a scale to the diameter of the hair shaft. This measurement is of doubtful value as a diagnostic aid. Along quite short distances of an individual hair, scale sizes may vary considerably, with a resultant change in the scale index. Furthermore, such a measurement conveys no information on the actual scale shape (i,e. a short and wide scale on a thin hair might have the same index as a long and narrow scale on a wide hair), although Khemelevskaya (1965) has suggested that the index might be of some u.e if a measurement of scale width was included.
A manual for hair identification, based almost entirely on photographs of scale shapes and arf'dngements, has been published by Adorjan and Kolenosky (1969), and many other workers depend heavily on aspects of the cuticle in attempting to identify unknown hairs.
The classification of cuticular scales and scale patte,.s
The scheme of classification shown hereunder is based on the following main features: I, Terms which describe the form of the scale margin,
THE STRUCTURE OFHAIRS
FORM OF SCALE MARGINS
A. Smooth B. Crenate C. Rippled . D. Scalloped E. Dentate
DISTANCE BETWEEN SC ALE MARGINS
g;q ~ F, Distant G, Near H. Close
SCALE PATTE RU
I. Simple Coronal
O. Regular Wave
J, Diamond K, Narrow Petal Diamond Petal
p, Irregular Wave
Q, Single Chevron
Figure 7 Various. shapes and arrangements of cuticular scales.
2, Terms which describe the distance between the external margins of scales.
3. Terms descriptive of the general scale patterns.
The scheme as outlined, is generally similar to that of Wildman (1954), although some additions and modifications have been made, All scale patterns described have been based on an appreciation of scale casts rather than rolled impressions (see page 15 for an explanation of these terms),
Before considering detailed aspects of scale classification, it is useful to recognize that there are 'two broad
L Broad Petal
R. Double Chevron
M. Regular Mosaic
r
~ ~'2: ,
N. Flattened Irregular Mosaic
T, Transitional
groups of scales; the coronal group and the imbricate group, A coronal scale completely encircles the hair shaft, whilst an imbricate scale extends only part of the way round the shaft circumference, It follows that several imbricate scales may be required in order to span the circumference of the hair at any point along its length. An example of each type described is shown in Figure 7, (I--coronal, J-T-imbricate),
L Scale margins
The scale margin may be defined as the free distal edge of an individual scale,
9

THE IDENTIFICATION OF MAMMALIAN HAIR
Smooth margins are without indentations and appear as a smooth line (Figure 7A).
Crenate margins have shallow but relatively pointed indentations (Figure 7 B).
Rippled margins have deeper indentations but the profile is generally rounded (Figure 7 C).
Scal/oped margins consist of a series of curves with generally rounded peaks and pointed troughs (Figure 70).
Denlate margins have large tooth-like projections and are found only on coronal scales (Figure 7 E).
2. Distance between margins (Figure 7 F-H).
In describing the arrangement of cuticular scales, it may be desirable to indicate whether the free edge of one scale and that of adjacent scales above and below are close together or well separated. For convenience we may speak' of distant, near and close scale margins. these being purely arbitrary divisions. The distant type are those where the visible width of the scale is not more than three times the visible length (i.e. the distance between one free distal scale margin and the next). In near types, the ratio of width to length might be Irom 3 to about 8, and in c/ose types it is greater than about 8. Scales in the proximal half of a hair are commonly longer (i.e. more distant than those in the distal half).
3. Scale patterns
The majority of harfs used for identification purposeS have cuticular scales of the imbricate type, and these are arranged in a variety of patterns, around the perimeter of the hair. Patterns may change markedly along the length of the hair with a corresponding change in the appearance of individual scales.
For anyone hair, an examination of the cuticle may reveal one or more of the following scale patterns:
I. Petal types Wildman (1954) described the general appearance of these as being similar to the patterns formed by a series of overlapping flower petals. Very often, the individual scales may appear to be diarnondshaped. In these cases, the pattern can be termed diamond petal (Figure 7. J-K). The seales forming this partkular pattern can range from being wide and short to being narrow and long. Another type of petal pattern, termed the broad petal (Figure 7 L), can be recognized, where the individual scales have rounded distal edges and are often of irregular sizes.
2. Mosaic types These patterns have an overall angular appearance. The term mosaic, as applied to the appearance of the pattern, is self explanatory. Sometimes, the width of individual scales is similar to their length (i.e. visible length and width) and we may speak of
10
regular mosaic patterns (Figure 7 M). Hence, irregular mosaic patterns occur when individual scales forming the pattern are of dissimilar sizes and shapes. It is also useful to recognize another common type of mosaic pattern, namely the flattened mosaic. The irregular mosaic pattern shOwn in Figure 7 N is also a flattened mosaic. Here, individual scales appear to be much wider than long (near), but still form an angular mosaic arrangement.
3. Waved types On some hairs, the free distal edges of the scales are presented in such a fashion as to appear like a series of waves. For all imbricate scales, the waves are not continuous but are interrupted by virtue of scale overlap. A true continuous wave can only be observed when rolled impressions (see page 15) are made from hairs with coronal scales. Waves may be either regular (Figure 7 0) or irregular (Figure 7 P) in appearance and, depending on their amplituc\e, may be desscribed as shallow, medium or deep. In cases of irregular waves, the amplitude of successive waves changes noticeably. Other types of waves have been identified and described by Wildman (1954). These include the single and double chevron (Figure 7 Q-R), and the streaked ware (Figure 7 S). In the single chevrqn, either the crests or troughs are pointed. whilst in the dou hie chevron, both crests and troughs are pointed and the Waves tend to have a somewhat larger amplitude. The streaked wave pattern has alternating series of deep and shallower waves and the change in wave form corresponds to a change in the contour of the hair circumference. The streaked wave pattern can often be observed on the outer surface of those guard hairs having a longitudinal channel (see page 1670).
4. Transitional patterns (Figure 7 T). By reference to Figure 3 (page 4), it will be noticed that the scale pattern changes markedly as one progresses from the tip to the root of an individual hair. At those points where one type of scale pattern changes into another, the pattern is often termed transitional. Some actual examples are shown on pages 67 and 69. Note that the transition from one pattern to another can take place over a very short distance along the hair.
HAIR PIGMENTATION
Although not of primary significance as a diagnostic criterion, hair pigment and its distribution can often be of use to confirm an identification made on the basis of other hair characteristics. In isolated cases, the particular
. appearance of hair pigment may be quite characteristic of a certain species and hance, be of great value in identification (Figure 8).
. It,,' f
I " [I
Figure 8 Cross-sectional appearance of hairs of the euro. (Macropus robustus) showing prominent pigment granules.
Pigment distribution
Pigment may be present in the cortex or medulla or both regions of a hair and there are isolated occasions when cuticular scales may be pigmented (Benedict 1957). An important aspect is the way in which the pigment is distributed along the length of an individual hair. Many mammalian hairs present a banded appearance and both Stains (1958) and Mayer (1952) have utilized aspects of colour banding in their keys to the identification of hairs. Again, the location of pigments in hair crosssections may be of importance, and Dreyer (1966) has produced a detailed classification of pigment distribution based on an appreciation of cross-sections.
Pigment colour
Two pigment types are known to occur in mammalian hair: eumelanin and pheomelanin. Eumelanin results in brown or black hair, whilst pheomelanin gives a yellow or reddish colour. Despite this, hairs may display
THE STRUCTURE OF HAIRS
a variety of colour hues, with black, brown and yellow being the most common or basic colours (Fitzpatrick ot al. 1958). Pigment type
It is possible to describe the presence of pigment in a hair as either diffuse, granular or aggregate; these terms being self-explanatory. Granular or aggregate pigment may be arranged in discrete and well defined areas Or in a streaked pattern. Individual granules may be large or small. Figure 8 gives an example of large granules of pigment in the cortex.
The use of pigmentation as a diagnostic aid in hair structure is subject to many limitations. For anyone mammal, there may be a noticeable variation in colour between hairs from different parts of the body. Again, the distribution of pigment may change considerably along the length of the hair. Colour mutations and fading are some other factors which limit the use of pigment in identification of unknown hairs.
The pelage of certain mammals will fluoresce under ultra-violet light, and Latham (1953) has used this char~cteristic to separate pelts of the least weasel (Mustela rixosa) fwm those o.f two other species in the genus. However, this phenomenon appears to be quite restricted and only oflimited application in identification of mammalian hair. '
11

-~-~~~~ ... ----------,----------------
SECTION B
1 Methods
of Studying
Hair Structure
THE WHOLE MO{;NT
When hairs are mounted on a microscope slide, it is generally possible to observe not only the general profile of the hair but also the appearance of the medulla (if present) and the distribution of pigment.
If the hair sample is taken from a study skin and is to be used as a reference, several precautions are necessary. First, a full range of bail's (i.e. a tuft of hairs) should be selected from each of several parts of the body and removed from the pelt in such a manner as to ensure that the maximum accessible length of each hair has been obtained. This can be achieved by using a very sharp razor blade and cutting flush to the skin surface.
The hairs must be cleaned, and 70 % alcohol or an alcohol/ether mixture (equal proportions) is useful for this purpose. Dried hairs can then be mounted .in a suitable medium on a clean glass slide. Several mounting media are available and the merits of each have been outlined by Wildman (1954). For reference slides, a permanent mountant such as DePeX (Gurr) is the most useful. When mounting the hair tuft, it is necessary to ensure that individual hairs are well separated so that a "hair forest" situation is avoided. Depending on the length of individual hairs, it may be necessary to use very long coverslips or to cut the hairs into two or more sections and mount each section in order. This technique of cutting hairs is also used to observe infiltrated medullae, as the mounting medium will. infiltrate the medullary spaces for a short distance from the cut end.
F or routine identification work (Le. preparing mounts of unknown hairs) it is generally more practicable to usc temporary mounting procedures. This allows for the re-use of glass slides and, also, is considerably faster than using permanent mounting. If possible, a tuft containing the full range of hairs should be selected, cleaned and
mounted. Paraffin oil is a most convenient medium for temporary mounts. An important advantage of the temporary mount is that hairs can subsequently be removed from the slide and used for othar purposes (e.g. cross-sectioning). This is of particular value when the unknown sample contains only a small number of individual hairs.
OBTAINING THE CROSS-SECTION
A number of methods are available for obtaining the hair cross~section and these vary from simple hand sectioning techniques through to the use of speci~lIy designed microtomes.
Where thin sections are required, it is usual to emp'loy fibre microtomes. Several such instruments bave been devised and they are widely used in the field of commercial fibre microscopy. A brief description of these instruments has been given by Wildman (1954) and Stoves (1957).
For the purposes of bair identification, very thin sections are not required and simple methods of sectioning can be used. Hand sectioning has been successfully employed by Mathiak (1938) and many others, but the most efficient of these simpler techniques would appear to be a modification of the plate method described by Ford and Simmens (1959). All cross-sections photographed or described in this book were obtained by the use of this technique,
The plate method
The apparatus consists of a stainless steel microscope slide of approximate dimensions 76 X 25 X O· 5 mm with two to six holes, each of 0·8 mm diameter, drilled at equal intervals along the centre as shown in Figure 9. These holes are carefully contoured on the top and
13

'·',:::Ei,'lI _______________________ -, __ -------------------
THE IDENTIFICATION OF MAMMALIAN HAIR
Figure 9 The plate method of cross-section;ng (see text).
bottom, to eliminate rough edges. The finished plate must he kept highly p~lished and perfectly flat.
For sectioning, a loop of thin nylon thread (approx. diam. 0·3 mm) is passed up through one of the holes (Fig. 9A) and threads of cellulose acetate yarn (330 decitex) are inserted into the loop (Fig. 9B). These threads are pulled a short distance down through the hole with the aid of the nylon thread. The hair tuft for identification, which should include a range of guard and underhairs, is then inserted into the centre of the yarn bundle protruding from the top of the plate (Fig. 9C). Hairs and packing material can then be pulled any desired distance through the plate by means of the nylon yarn.
The cross-sections are obtained by using a degreased safety razor blade to cut the protruding hair bundle flush to the plate on both sides. The blade is held at about 35 degrees to the plate (Fig, 9C), The cross-sections are viewed in situ after placing paraffin oil and a coverslip directly over the sectioning hole, Sections should be viewed from that side of the plate upon which the second cut is made (i.e, the underside),
In some samples, where large hairs have only a thin or narrow cortex layer, the shape of the hair in the cross-section may become distorted as a result of undue packing pressure in the sectioning hole, Such distortion can be overcome by using fewer hairs and dissolving
14
the packing material with a few drops of cellulose acetate solution (2~A % in acetone) immediately prior to cutting tbe section, This relieves undue pressure and the dissolved yarn resets in a few second" thus preventing any part of the bair bundle from falling out of the plate.
The plate method has a number of distinct advantages over other sectioning methods, It is rapid, and the experienced operator can produce cross-sections ready for microscopic examination within two minutes, Very short hairs (such as are often found on the head, reet and ears) and damaged hairs, can be cross-sectioned with relative ease, Because cross-sectional appearance often varies along the length of an individual hair, longer hairs can be preent into sections before they are inserted in the plate, For a permanent record, plates can be sealed and labelled. A particular advantage of the plate method is that the plate can he re-used indefinitely simply by pushing out the contents of the sectioning hole with a mounted needle.
OBTAINING CUTICULAR SCALE CASTS
As a rule, the surface structure or scale pattern of a hair cannot be properly appreciated from the whole mount preparation, Exceptions occur in hairs which have little or no medulla and are either non-pigmented or only slightly pigmented.
METHODS OF STUDYING HAIR STRUCTURE
Procedures for studying scale patterns usually involve the use of special media to obtain a cast or impression of the actual hair surface, These are often produced directly on to glass slides and viewed under the microscope, Wildman (1954) differentiates between the terms "cast" and "irnpressionH and restricts the use of the latter tenn to those cases where the entire circumference of the hair is represented, Hence a scale cast refers to the situation where only part of the hair circumference is represented,
Impressions are obtained by carefully rolling the hair over the surface of a suitable medium on a slide so that the scale pattern around the full circumference of the hair can he appreciated. Whilst this may be desirable, the technique is rather time-consuming and of limited application in routine hair identification work. Very short hairs, damaged hairs and hairs which are extremely flattened in cross-section can be accurately rolled only with the greatest difficulty,
[t is more efficient, in hair identification work, to employ the "scale cast" technique, Here, the hair is ~
placed on or pressed into a suitable medium which will then reproduce the surface structure for that part of the hair in contact with the medium. Various media are available, including gelatin, celluloid, vinyl acetate polymers and various specialized products such as ethofoi! and permount (Wildman 1954).
For general work, the use of polyvinYl acetate (pva) or vinyl acetate copolymers is recommended. These have several advantages, They are inexpensive, easily obtained and readily soluble in water, Moreover, they have excellent casting properties and will faithfully reproduce surface structures, A large range of such polymers is available commercially and the particular choice is hest left to the individual,
A simple casting technique
A long glass coverslip (50X20xO,2 mm) is coated
-.= ... ==============
Figure 10 Obtaining the scale cast (see text),
with a thin layer of diluted pva, using a fine flat brush. Casts are not produced directly onto standard microscope slides because the thickness of the glass will not allow viewing of inverted casts at high magnifications, The pva has good levelling qualities and will produce a film of fairly even thickness. A 50 % solution of pva in distilled water is generally adequate, although more concentrated solutions may be necessary for very coarse hairs.
The selected hairs should be cleaned in an alcohol-ether mixture and dried between good quality absorbent paper. The individual hairs are conveniently handled with fine forceps and a mounted needle. Several hairs should he placed on the coverslip, some with tips overhanging and others with the hair root or lower shaft region overhanging (Figure 10), All hairs should he gently pressed into the medium so that adequate surface contact is ensured,
After some 20 minutes at room temperature, hairs can be removed from the coverslip. This is effected by grasping the overhanging section of each hair with forceps and carefully lifting it out of the medium. For viewing the casts, the coverslip is inverted and placed on a normal microscope slide. Such an arrangement will show the scale pattern as it would normally appear on the hair surface (i.e, convex view),
Waved or twisted hairs must he treated with special
15

~-----~-~-----""----------------------'!lIP-------------------
THE IDENTIFICATION OF MAMMALIAN HAIR
care if useful casts are to be produced. It is generally advisable to fasten both ends of such hairs with small pieces of gummed paper so that they are correctly positioned for embedding and are laid fiat along their entire length.
In the case of very fine hairs (e.g. of some Chiroptera) the strong adhesive property of pva prevents removal of the hair from the coverslip. It is then advisable to use a gelatin solution in place of pva. A coverslip is coated with a thin film of gelatin solution (3 % in water)_ Hairs are laid out in the manner described above and stripped from the glass when the medium has dried sufficiently.
When studying scale casts it is important to view the scale pattern at various points along the length of the hair, and the points of transition between one type of scale pattern and another. It must also be appreciated that some hairs have recognisable "sides" (e.g. a COncave and a convex surface) and that such hairs have a tendency to consistently present only one "side" to the embedding medium. Such a problem can be overcome by deliberate
. manipulation of hairs whilst they are being laid in the embedding niedi urn.
Scale casts cannot be successfully stored for long periods and it is therefore essential that photographic records are kept for comparative work.
PliQTOGRAPHlNG HAIR STRUCTURES
It is not intended ifere to deal with the subject of photomicrography in any detail but, rather, to mention some of the general principles involved in producing photographs for use in a hair identification system. The reader is advised to consult more specialized texts for information on photographic equipment, film types, lighting etc. Some useful technical information on photomicrography of hair is given by Appleyard (1954).
The photographs of hair structure in Section C of this book were produced by the use of a 5" X 4" bellows camera using cut films in a double-sided holder_ All photographs of hair cross-sections, whole mounts and scale casts repreSi'nt a total magnification of 468 (±3).
Large format cameras allow for contact printing from negatives and enlargements should seldom be necessary. Nevertheless, modern film in 35 mm cameras should produce negatives of a high standard, allowing for considerable enlargement without loss of detail.
It is important that all photographs of hair structure be produced at the one magnification. This allows for direct comparison between photographs and is par, ticularly useful when measuring the maximum diameter of primary guard hairs from different species.
Although colour transparencies of hair structures (particularly cross-sections) may be of use in the identification process, the assessment of colour is difficult and
16
intra-species variation in hair colour may be considerabJe. For these reasons, colour photographs have not been included in the photographic system outlined.
The photographs of hair profile shown in Section C were taken after projecting the image of hairs on to a white paper screen and carefully drawing the outline of the hair. A standard 35 mm projector was used and the hairs were held between two glass plates. The enlarged drawings of hairs were subsequently placed over an appropriate scale and photographed_
2 A System
for Hair
Identification
When hair identification is employed as a routine technique, the system adopted must have the capacity to screen a range of hair structures for all mammals likely to be present in the particular situation or area under study.
The identification can be effected by comparing the unknown sample with the full range of known hair samples. This direct comparison technique is, however, tedious and slow and quite unsuitable for routine analysis work.
One might consider the use of descriptive dichotomous keys. Unfortunately, these have several shortcomings, when applied to mammalian hairs~ Like the direct comparison method, they are slow and often difficult to interpret Their major drawback, however, stems from the fact that they are unable to fully represent the intra· specific variation in hair structures that is characteristic of most mammals.
One solution to these problem, is to photograph the most diagnostic characteristics of hair structure for all mammals concerned and use these photographs as a reference system. Useful aspects of microscopic structure for single hairs and groups of hairs can be represented and appreciated directly from the photographs. This eliminates the tedious "procedure of direct comparison with specimens of known and unknown hairs but, at the same time, allows for greater representation of structure variations than does a written key.
It must be remembered that no system of hair identification can be fully effective. Sometimes, hair structures for closely related mammals appear identical in all respects and no differentiation can be made. Again, there are occasions when an unknown sample is devoid of those hairs which exhibit the most diagnostic features of structure (usually primary guard hairs). In these cases, precise identification may not be possible.
PRODUCING THE SYSTEM
It is necessary, at the very begInning, to point out that the construction of a photographic reference system and its subsequent use~ requires some experience on the part of the operator. It is only experience which allows one to recognize the most diagnostic features of hair structure for each species considered or to identify a particular type of hair in an unknown sample (i.e. primary guard hair, overhair etc.)~ The time taken to achieve this expertise, will depend largely on the number of species which must be represented in the system and the complexity of hair structures encountered. .
The first requirement for the construction of the reference system is a set of accurately identified study skins, representative of all animals likely to be encountered in the identification work. The methods of preparing study skins have been outlined by Dimpel (1971) and many others, and are outside the scope of thip book.
Hair samples should normally be taken from the rump area of each skin but, if hair structure differs markedly from place to place on the species examined, it will be necessary to include hair samples from other parts of the body. Each sample must indude all hair types present in the main pelage (i.e. overhairs, guard hairs and underhairs).
For most mammals, the following features of structure should be carefully examined and photographed:
(a) the general profile of primary and smaller guard hairs and an underhaif.
(b) the cross-sectional appearance of primary and smaller guard hairs at their widest point (the cross' sectional appearance of underhairs can also be represented in this one photograph).
(c) the medulla arrangement for primary and smaller
17

THE IDENTIFICATION OF MAMMALIAN HAIR
guard hairs at several points along their length.
(d) the scale pattern at several points along the length of a primary guard hair.
Where large numbers of photographs are required to cover the four points mentioned above, it is usually possible to ecOn lize by retaining only those particular aspects of medulla arrangement, scale pattern, etc. which appear to have the greatest diagnostic value.
The photographs of scale pattern, medulla arrangement and cross-sectional shape for hairs from all the mammals considered should be taken at the one standard magnification. Various magnifications can be chosen fur all photographs of hair profile, but in each case a meaSurement scale should be included. The use of .,standard magnifications will enable direct comparison between photographs. and give some idea of relative sizes for various hairs and their components.
In any reference system containing a relatively large number of species (say in exceSS of 30), it should be
. possible to split up the reference system into several major groups of mammals. This grouping can be carried out on the basis of gross differences in the appearance of one or more hair structures. In the case of the mammals which the authors studied, the division was best effected on the basis of differences in cross-sectional shape at the widest point along the length of the primary guard hairs. In other situations it may be more convenient to divide on the basis of general '{profile of guard hsirs, medulla arrangement or other hair characteristics.
Within each resulting group, it may be possible to carry out further subdivision. For instance, in our experience, some groups can be subdivided on the basis of medulla appearance, presence of constrictions, presence of divided medulla and other characteristics.
Such a division into groups and sub-groups substantiaJJy reduces the number of photographs which must be scrutinized before an identification can be made. At the same time, it is necessary to point out the danger of excessive subgrouping. In effect this produces a situation similar to that which results from attempting to use a full descriptive key and introduces the difficulties inherent in that system.
USING THE SYSTEM
For efficient identification it is best to have a tuft of the unknown hairs. This should ensure that all desirable hair types are present and, most importantly, will facilitate the location of primary guard hairs. The unknown sample may often contain hairs from more
18
than one species but, fortunately, hair tufts from each mammal usually remain intact and differences in the microscopic appearance of tufts will allow their separation.
When only small numbers of hairs are available or when the sample has been extensively damaged, this procedure of sorting out discrete groups or bundles of hair will not be possible.
Whatever the situation, a range Of mixture of hairs should be selected for closer examination. Whenever possible, the selected hairs should include several of the primary or larger guard hairs. For the sample so selected it will then be necessary to produce whole mounts so that aspects of medulla appearance, pigment distribution and general profile can be observed. The most important step, however, is the production of cross-sections. All hairs in the selected sample (a hair tuft or mixture of hairs) can be sectioned in the one operation. Where guard hairs can be recognized in the sample, the cross-section should be taken in such a way so as to ensure that these hairs are sectioned at their widest point. When poor samples are encountered, it may be necessary to repeat the sectioning process until maximum representation of all possible cross-sectional shapes present in the sample can be achieved.
From an appreciation of cross-sectional appearance, and to a lesser extent general profile, medulla, or other hair characteristics, it should be possible to select quickly the appropriate group or subgroup of mammals to which the unknown hairs belong. Finally, however, it will be necessary to compare aspects of the unknown sample against a series of photomicrographs.
Scale patterns can very often be ignored in the identification process and are generally only used to confirm identifications made on the basis of other criteria. However, there will be some occasions when the scale pattern is of primary diagnostic significance.
Notwithstanding the value of photographs, it is always advisable to have, close at hand, a full set of reference skins. Direct comparisons may be necessary when the unknown hairs are suspected of having originated from extremities on the body (e.g. tail, foot, etc.). These hairs will often be markedly different from those on the main pelage and may not be represented in the photographs.
The identification system, as outlined above, has been used successfully by the authors in the examination of several thousand hair samples from the stomach contents of foxes (Vulpes vulpes L. J, dingoes (Canis jamiliaris dingo) and feral domestic cats. (Coman 1972, 1973: Coman and Brunner 1972). A large number of hairs from faecal samples have also been successfully identified using this system. (Brunner and Coman, unpublished data).
SECTION C
1 A Guide to Hair Structure
for the Indigenous" and
Introducedt Mammals of Victoria
It is important to realize that the photographs in this section represent only some of the multitude of hair structures observed in the hair of anyone species. Generally speaking, only the most diagnostic features have been covered.
It should also be noted that with very few exceptions, hairs from extremitjes) such as the taH. ear or mane, have not been included in the system. Such hairs may be encountered from time to time, especially in food-habits investigations. Usually hairs from the extremities are circular to oval in cross-section with a small medulla and are often of a bristly nature.
For each species considered, a short commentary is given and this attempts to point out particularly useful features of structure. The maximum diameter of guard hairs is also given and a scale, useful for measuring directly from photographs is located below.
Some arrangement of species into groups or sub-groups (based on hair structures) has been possible. In each group or sub-group, the species are arranged within families or genera.
The following key has been used on 80me oflhe crosssection photographs: G Primary Guard.hair· o Overhair U Underhair Micron o 50 100 150 200 250
111111111111111111111111111 Scale for photomicrographs in this section. Each division equals 10 microns.
* Classification according to Ryde (1970, t Classification according to Walker (1964)
,
19

2 A Grouping
of Victorian Mammals
BASED INITIALLY ON THE CROSS-SECTIONAL
APPEARANCE OF PRIMARY GUARD HAIRS
'.
Family Genus and Species
. GROUP Hairs predominantly circular in cross section.
Dasyuridae Dasyurus maculatus page 22 Dasyurus viverrz'n~s 24 Sminthopsis crassicaudata 26
Phalangeridae Acrobates pygmaeus 28 Cercartetus concinnus 30 Cercartetus nallus 32 Burramys parvus 34
i;{'~
GROUP 2 Hairs predominantly oval in cross-section.
Sub-group (a) Hairs with medulla much reduced or absent.
Suidae Phascolomidae Hominidae Bovidae Vespertilionidae
• Sus serofa 36 Vambatus ursinus 38 Homo sapiens 40
*Ovis aries 42 Nyctophi!us geoffroyi 44 Miniopterus schreibersii 46 J::-ptesicus pumilis 48 Chalinolobus gouldii 50 Myotis adversus 52
Sub-group (b) Hairs with very large medulla.
Cervidae • Dama dama 54 * Axis porcinus 56
Sub-group (0) Hairs up to 451' (max. diam.) and with distinct constriction before shield.
Dasyuridae Antechinus j/avipes 58 Antechinus stuartii 60 Antecninus swainsonii 62 Anteehinus minimus 64 Sminthopsis murina 66 Sminthopsis leueapus 68
20
Family Genus and Speceis
Sub-group (d) Hairs larger than 40/-, (max. diam.) and without constrictions.
Macropodidae
Canidae
Equidae Felidae Bovidae
Maeropus glgomeus 70 Maeropus fuliginosus me/anal's 72 Maeropus robustus 74 Macropus rufogrL,eus 76 Megalela rufa 78 Wallahla bieolor 80 Thylogale billardierii 82 Petrogale penicillata 84· Potorous tridactylus 86 Canis familiaris 88
• Vulpes vulpes 90 'Equus wbollus 92 • Felis eatus 94 • Bas taurus 96
GROUP 3 Hairs with eye or lemon-shaped crosssections.
Phalangeridae
Petauridae
Phascolarctidae
Trichosurus Quipecula 98 Trichosurus caninus 100 Pseudocheirus peregrinus 102 Pe/aurus breviceps 104 Pe/aurus nor/oleensis 106 Petaurus aus/ralis to8 Schomobates volans 110 Gymnobe/ideus leadbeateri 112 Phascolaretos emereus 114
GROUP 4 Hairs with predominantly oblong crosssections.
Sub-group (a) Medulla absent.
Taehyg10ssidae Tachyglossus acu/eatus 116
Family Genus and Species
Sub-group (b). Medulla present .
Cervidae
Mustelidae Dasyuridae Muridae
'Ceraus un/color 118 'Cercus e/aphus 120 • Mustela pu/orius 122 Phascogale tapoatafa 124 Hydromys ehrysogaster 126 Notomys miteheliii 128 Pseudomys novaehollandiae 130 Pseudomys a/boeinereus 132 Pseudomys fumeus 134
GROUP 5 Hairs predominantly flattened or cigar-shaped in cross~section,
Otariidae Arctocephalus doriferus 136 Ornithorhynchidae Ornithorhynchus anat/nus 138
GROUP 6 Hairs predominantly renifonn or cone"VO-convex in cross~section.
Sub-group (a) Hairs with divided medulla.
Peramelidae Isoodon obesulus 140 Perameles nasuta 142
. Perameles gunnii 144
Family
Sub-group (b)
Muridae
Caviidae
Sub-group (c)
Genus and Species.
Hairs with bilobed or large medulla.
• Rattus rattus 146 • Rattus norvegicus 148
Rattus fuscipes 150 Rallus lutreolus 152 Pseudomys shortridgei 154 Mastacornys fuscus 156
'Mus musculus 158 'Cavia parcellus 160
Medulla much reduced or absent.
Pteropodidae Pteropus scapulatus 162 Pter!'pus palioeephalus 164
GROUP 7 Hairs predominantly dumb-bell-shaped in cross-section.
Bovidae Leporidae
Capra hircus 166 Oryc/olagus cuniculus 168 Lepus europaeus 170
21

TIGER CAT
Dasyurus maculatus
A, B Cross-sections of various hairs, Maximum diameter of primary guard hairs up to 11Oj!t-,
C, D Whole mounts of primary guard hairs in shield region. E Whole mount of primary guard hair in proximal half. P-H Scale pattern at various points. F, shield region; G,
mid-shaft region; H, near base, ", MOST DIAGNOSTIC FEATURES: Aeriform lattice medulla and
medium pigment distribution.
A
22
&<>1,
?~~ ___ 2 __ 3~_4~~5 __ .6 __ 7~_e~~9--"?MM
c o E
F G H
23





























