Embed Size (px)
Citation preview

The histaminergic neurotransmitter system
– its development and role in neuronal plasticity in
zebrafish, Danio rerio
Jenny Maria Sundvik
Neuroscience Center and
Institute of Biomedicine, Anatomy
Faculty of Medicine
University of Helsinki
Finland
and
Helsinki Biomedical Graduate Program
ACADEMIC DISSERTATION
To be publicly discussed with the permission of the Faculty of Medicine, University
of Helsinki, in Lecture Hall 2, Haartman Institute, Haartmaninkatu 3,
on August 24th
2012, at 12 noon.
Helsinki 2012

Supervised by
Professor Pertti Panula
Neuroscience Center and Institute of Biomedicine, Anatomy
Faculty of Medicine
University of Helsinki
Finland
Thesis committee
Professor Esa Korpi
Institute of Biomedicine, Pharmacology
Faculty of Medicine
University of Helsinki
Finland
Dr. Tarja Stenberg
Institute of Biomedicine, Physiology
Faculty of Medicine
University of Helsinki
Finland
Reviewed by
Assistant Professor Sanna Lehtonen
Haartman Institute, Pathology
Faculty of Medicine
University of Helsinki
Finland
Assistant Professor Niclas Kolm
Department of Ecology and
Evolution, Animal Ecology
Evolutionary Biology Center
Uppsala University
Sweden
Opponent
Dr. Catherina Becker
Center for Neuroregeneration
The University of Edinburgh
United Kingdom
© Maria Sundvik 2012
ISBN 978-952-10-8136-1 (paperback)
ISBN 978-952-10-8137-8 (PDF)
http://ethesis.helsinki.fi
Unigrafia, Helsinki University Print
Helsinki 2012

To my family


Table of contents
List of original publications........................................................................................................... 7
Author contribution ......................................................................................................................... 7
Abstract .................................................................................................................................................. 8
Abbreviations ................................................................................................................................... 10
1 Introduction .................................................................................................................................. 12
2 Review of the literature ........................................................................................................... 14
2.1 Neuronal networks and plasticity ............................................................................... 14
2.1.1 Aminergic neurotransmitter systems............................................................... 15
2.1.2 Development of the histaminergic neurotransmitter system ............... 16
2.1.3 Synthesis, storage, release, and catabolism of the aminergic
neurotransmitters .................................................................................................... 17
2.1.4 Histaminergic receptor-signaling system ....................................................... 19
2.2 Neurotransmitter systems under normal conditions and disease ............... 21
2.2.1 Alertness and wakefulness .................................................................................... 21
2.2.2 Histamine and neurodegenerative diseases .................................................. 22
2.3 Zebrafish as a vertebrate model organism ............................................................. 25
2.3.1 Comparative neuroanatomy of fish and mammals ..................................... 26
2.3.2 Zebrafish behavior and motor functions ......................................................... 28
2.3.3 Behavioral pharmacology ...................................................................................... 30
2.3.4 Modification of gene expression with morpholino
oligonucleotides ........................................................................................................ 31
2.3.5 Modification of gene expression by inducing mutations into the
genome .......................................................................................................................... 31
2.3.6 Spatially and temporally targeted gene expression ................................... 32
2.3.7 Appropriateness of modeling human disease in a simple
vertebrate .................................................................................................................... 33
3 Aims of the study ........................................................................................................................ 34
4 Methods ........................................................................................................................................... 35
4.1 Animals ................................................................................................................................... 35
4.2 Manipulation of gene expression by morpholino oligonucleotides (III) ... 35
4.3 Pharmacological treatments ......................................................................................... 35
4.3.1 Treatment with histamine receptor ligands or α-
fluoromethylhistidine (III) ................................................................................... 36
4.3.2 Deprenyl, p-chlorophenylalanine, and fluvoxamine treatments (V) .. 36
4.3.3 γ-Secretase inhibitor DAPT treatments (IV).................................................. 36
4.4 Behavioral tests ................................................................................................................... 37
4.4.1 Locomotor activity of zebrafish (I, III–V) ........................................................ 37
4.4.2 Place preference (IV) ............................................................................................... 37
4.4.3 Place preference in a water column (V) .......................................................... 37
4.4.4 Locomotor activity of larval zebrafish over a 24-h period (III) ............ 38
4.4.5 Dark-induced flash responses in larval zebrafish (III, IV) ....................... 38
4.4.6 Optomotor response (III) ....................................................................................... 38
4.5 Lipophilic dye DiI tracing (III) ...................................................................................... 38

4.6 Real-time polymerase chain reaction ........................................................................ 39
4.6.1 Cloning of zebrafish hrh1, hrh2, and hrh3 receptors in adult
zebrafish (I) ................................................................................................................. 39
4.6.2 Expression of hrh1, hrh2, and hrh3 mRNA (I) .............................................. 39
4.6.3 Cloning full-length hdc cDNA and synthesis of capped
hdc mRNA (III) ........................................................................................................... 39
4.6.4 Quantitative real-time polymerase chain reaction (III, IV) ..................... 40
4.6.5 Touchdown RT-PCR for identification of mutation in psen1-/- (IV) .. 40
4.7 Sequence comparisons and analysis of histamine receptors (I) ................... 41
4.8 [125I]iodoaminopotentidine binding to hrh2 (I) ................................................... 41
4.9 Whole-mount in situ hybridization (III, IV) ............................................................ 41
4.10 Immunohistochemistry (II–V) ................................................................................... 42
4.10.1 IHC with antibodies from different species on whole-mount larval
brains (II–V) ................................................................................................................ 42
4.10.2 IHC on whole-mount adult brains (II, IV) ..................................................... 42
4.10.3 IHC with two antibodies from the same species on cryosections
(II, III) ............................................................................................................................ 43
4.10.4 Enzyme histochemistry and activity assay (V) .......................................... 44
4.11 High-pressure liquid chromatography .................................................................. 44
4.11.1 Catecholamine and serotonin HPLC (V) ....................................................... 44
4.11.2 Histamine HPLC (III) ............................................................................................. 44
4.12 Microscopy ......................................................................................................................... 44
4.13 Image analysis ................................................................................................................... 45
4.14 Statistical methods .......................................................................................................... 46
5 Results ............................................................................................................................................. 47
5.1 Identification and characterization of the histamine receptors in
zebrafish and the behavioral effects of their ligands (I, III) ............................ 47
5.2 Cotransmitters of the histaminergic neurons (II, III) ......................................... 48
5.3 Histaminergic system in alertness/wakefulness (III) ........................................ 49
5.4 γ-Secretase regulates the development of histamine neurons (IV) ............. 50
5.5 Neuronal plasticity and the histamine system (III, IV) ...................................... 52
5.6 Monoamine oxidase affects locomotor activity via serotonin (V) ................ 54
6 Discussion ...................................................................................................................................... 56
6.1 The methods – drawbacks and strengths ................................................................ 56
6.2 Neuronal circuits underlying behavior (III, V) ...................................................... 57
6.3 The histaminergic system in zebrafish compared with that in
mammals (I–III) .................................................................................................................. 59
6.4 The potential role of histamine in neurodegenerative disease (III, IV) ..... 63
7 Conclusions .................................................................................................................................... 66
8 Future prospects ......................................................................................................................... 68
Acknowledgements ....................................................................................................................... 70
References ......................................................................................................................................... 72
APPENDIX Original publications

7
List of original publications
I Peitsaro N, Sundvik M, Anichtchik OV, Kaslin J, Panula P, 2007,
“Identification of zebrafish histamine H1, H2 and H3 receptors and effects of
histaminergic ligands on behavior”, Biochemical Pharmacology, vol. 73, no.
8, pp. 1205-1214.
II Sundvik M, Panula P, 2012, “The organization of the histaminergic system in
adult zebrafish (Danio rerio) brain: neuron number, location and co-
transmitters”, Journal of Comparative Neurology, accepted for publication.
III Sundvik M, Kudo H, Toivonen P, Rozov S, Chen YC, Panula P, 2011, “The
histaminergic system regulates wakefulness and orexin/hypocretin neuron
development via histamine H1 receptor in zebrafish”, The FASEB Journal,
vol. 25, vol. 12, pp. 4338-4347.
IV Sundvik M, Chen YC, Panula P, “Presenilin1 regulates histamine neuron
development and behavior in zebrafish, Danio rerio”, submitted.
V Sallinen V, Sundvik M, Reenilä I, Peitsaro N, Khrustalyov D, Anichtchik O,
Toleikyte G, Kaslin J, Panula P, 2009, “Hyperserotonergic phenotype after
monoamine oxidase inhibition in larval zebrafish”, Journal of
Neurochemistry, vol. 109, no. 2, pp. 403-415. This publication is included in
Dr. Ville Sallinen’s doctoral thesis.
Author contribution
I Sundvik set up a method for assessing behavior in larval zebrafish, designed
the behavioral experiment, conducted it, and analyzed the results.
II Sundvik designed the study partially, conducted all experiments, analyzed
the data, and wrote the manuscript.
III Sundvik set up behavioral methods for zebrafish larvae, conducted the
behavioral, molecular, and immunohistological studies, analyzed the results,
and wrote the manuscript.
IV Sundvik established the association between the histaminergic system and
psen1, designed the study, carried out the experiments, and wrote the
manuscript.
V Sundvik set up a method for assessing behavior in larval zebrafish, designed
the behavioral experiments, conducted them, analyzed the results, and took
part in writing the manuscript.

8
Abstract
The histaminergic neurotransmitter system is a modulatory system which
regulates many functions in the vertebrate brain. In the mammalian brain,
histaminergic neurons fire during waking and are involved in maintaining sleep-
wake homeostasis. Furthermore, histamine has been implicated in
neurodegenerative disorders as resulting from either up- or downregulation of the
neurotransmitter system and has been detected in patients suffering from
Alzheimer’s disease, Parkinson’s disease, and narcolepsy. Histamine is produced
from the amino acid ι-histidine by histidine decarboxylase (HDC). The aims of this
study were to characterize the histaminergic system in zebrafish, a nonmammalian
vertebrate, and to identify novel mechanisms that are mediated by the
histaminergic system.
We found that histamine receptor ligands altered the behavior in larval zebrafish
in a dose-dependent manner. Cloning and sequence analysis of the histaminergic
receptors indicated that the zebrafish histamine receptors were similar to the
corresponding mammalian receptors. We also studied the neurotransmitter
phenotype of histaminergic neurons by immunohistochemistry and confocal
microscopy and found that these neurons resemble those of rats.
Immunohistochemical comparison of the histaminergic system in male and female
adult brain revealed no significant differences in location, distribution, or neuron
number, and thereby no differences between the sexes were observed. We
studied the functioning of the histaminergic system in hdc knockdown animals
(morpholino oligonucleotide approach), and found that the neurotransmitter was
important for mediating rapid behavioral responses to environmental cues, and
that these behavioral responses were driven via the histamine receptor, hrh1. We
studied the interaction of histamine with a wakefulness associated neuropeptide
system, hypocretin/orexin (hcrt), which is implicated in narcolepsy. We reported a
novel mechanism for histamine that regulates development of hcrt neurons.
Transient knockdown of hdc with morpholino oligonucleotides and
pharmacological inactivation of hrh1 with pyrilamine reduced the number of hcrt
mRNA-positive neurons in zebrafish larvae. This effect was rescued by
overexpression of hdc mRNA. Since histaminergic neurons play important roles in
the central nervous system, we were interested in identifying a factor that
regulates the development of the histaminergic neurons themselves. We found
that in the Alzheimer’s disease-associated gene, presenilin1 (psen1), mutant
zebrafish, the histaminergic system was altered. During the developmental and

9
larval stages, the histamine neuron number was significantly reduced in psen1
mutants, whereas at the young adult age there was a significant increase in
histamine neuron number in psen1 mutants in comparison to wild-type fish. These
results showed us that psen1 was essential for the development and normal
functioning of the histaminergic neurotransmitter system. Histamine is
metabolized by histamine N-methyltransferase (HNMT), followed by monoamine
oxidase (MAO). We studied the effect that pharmacological inhibition of MAO has
on the locomotor behavior of fish. We found that pharmacological inhibition of the
MAO enzyme mainly affects the serotonin system in zebrafish, with characteristic
behavioral consequences that could be rescued.
Taken together, the results compiled in this thesis provide evidence that the
histaminergic neurotransmitter system in zebrafish is similar to that in mammals.
These results show that the zebrafish is an excellent tool in studies that provide
novel insights into neurotransmitter system functions in the vertebrate central
nervous system on both the mechanistic and behavioral levels. We found that the
histaminergic system is important for rapid behavioral responses, affects the
development of the hcrt neurotransmitter system, and is itself regulated in a
temporal manner by psen1.

10
Abbreviations 3D Three-dimentional
5-HIAA 5-Hydroxyindoleacetic acid
AADC Aromatic ι-amino acid decarboxylase
AAS Ascending arousal system
AC Adenylate cyclase
AD Alzheimer’s disease
Akt/GSK3 Serine/threonine protein kinase/ Glycogen synthase kinase 3
ALDH Aldehyde dehydrogrnase
AMPK Adenosine monophosphate-activated protein kinase
ANOVA Analysis of variance
aph-1 Anterior pharynx-defective 1
APP Amyloid precursor protein
Ca2+
Calcium
cAMP Cyclic adenosine monophosphate
cDNA Complementary DNA
CEL-I Plant endonuclease
CNS Central nervous system
COMT Catechol-O-methyl transferase
CREB cAMP response element-binding
DAG Diacylglycerol
DAO Diamine oxidase
DAPT N-[(3,5-Difluorophenyl)acetyl]-L-alanyl-2-phenyl]glycine-1,1-dimethylethylester
Dc Central area of dorsal telencephalon
Dd Dorsal area of dorsal telencephalon
DβH Dopamine β-hydroxylase
DiI Lipophilic tracer dye
Dl Lateral area of dorsal telencephalon
Dm Medial area of dorsal telencephalon
DMSO Dimethyl sulfoxide
ι-DOPA ι-Dihydroxyphenylalanine
DOPAC 3,4-Dihydroxyphenylacetic acid
Dp Posterior area of dorsal telencephalon
dpf Days postfertilization
E3 Embryo medium
EDAC 1-Ethyl-3,3(dimethyl-aminopropyl) carbodiimide
ENU N-ethyl-N-nitrosurea
α-FMH α-Fluoromethylhistidine
GABA γ-aminobutyric acid
gad1 Glutamate decarboxylase 1, or gad67
gad2 Glutamate decarboxylase 2, or gad65
Gal4 Transcriptional activator, galactose utilization
HA Histamine
Hc Caudal zone of periventricular hypothalamus
hcrt Hypocretin/orexin
hcrt1 Hypocretin receptor subtype 1
hcrt2 Hypocretin receptor subtype 2
HDC Histidine decarboxylase
HNMT Histamine N-methyltransferase
hpf Hours postfertilization
HPLC High-pressure liquid chromatography
hrh1 Histamine receptor subtype 1
hrh2 Histamine receptor subtype 2
hrh3 Histamine receptor subtype 3
hrh4 Histamine receptor subtype 4
IHC Immunohistochemistry
IP3 Inositol triphosphate

11
MAO Monoamine oxidase
MAPK Mitogen-activated protein kinase
mENK Methionine-enkephalin
MID2cm Mauthner neuron homologue, middle dorsal 2 contralateral medial longitudinal fasciculus
interneuron in rhombomere 5
MID3cm Mauthner neuron homologue, middle dorsal 3 contralateral medial longitudinal fasciculus
interneuron in rhombomere 6
MO Morpholino oligonucleotide
mRNA Messenger RNA
NF-κB Nuclear factor κ-light-chain-enhancer of activated B cells
nMLF Medial longitudinal fasciculus
o/n Overnight
PCPA p-Chlorophenylalanine
PD Parkinson’s disease
pen-2 Presenilin enhancer 2
PFA Paraformaldehyde
PKA Protein kinase A
PKC Protein kinase C
PLC Phospholipase C
PNMT Phenylethanolamine Ν-methyl transferase
psen1 Presenilin1
psen2 Presenilin2
qPCR Quantitative real-time polymerase chain reaction
RM ANOVA Repeated measures ANOVA
RT Room temperature
RT-PCR Real-time polymerase chain reaction
SEM Standard error of mean
slc17a6a Solute carrier family 17 (sodium-dependent inorganic phosphate cotransporter), member 6a,
or vesicular glutamate transporter 2b
slc17a6b solute carrier family 17 (sodium-dependent inorganic phosphate cotransporter), member 6b,
or vesicular glutamate transporter 2a
slc17a7 solute carrier family 17 (sodium-dependent inorganic phosphate cotransporter), member 7,
or vesicular glutamate transporter 1
sOVA Succinylated ovalbumin
std Standard
TALEN Transcription activator-like effector nuclease
TeTx Tetanus toxin
TH Tyrosine hydroxylase
TILLING Targeting induced localized lesions in genomes
TL Tubingen longfin
TMN Tuberomammillary nucleus
TPH Tryptophan hydroxylase
TRH Thyrotropin-releasing hormone
TSA Tyramide signal amplification
UAS Upstream activating sequence
VMAT2 Vesicular monoamine transporter 2
WISH Whole-mount in situ hybridization
WT Wild-type
ZFN Zinc finger nuclease
ZT Zeitberger time

Introduction
12
1 Introduction
The basis of modern neuroscience was laid about a century ago when the Spanish
neuroanatomist Santiago Ramon y Cajal suggested that neurons are the
anatomical and functional units that together form the central nervous system
(CNS). His work focused on the structure and organization of the CNS and
contributed to the acceptance of the neuron doctrine. The neuron doctrine states
that neurons are the fundamental structural and functional units of the CNS, are
distinct units that are comprised of dendrites, somata, and axons, and that
information flows along the neuron from dendrites to axon. Specific behaviors are
produced by integration of neuronal networks and sensory inputs. Failure to
produce appropriate responses to certain environmental stimuli can be
detrimental for the individual. Hence, it is of the outmost importance that the
individual responds in a correct way. Coordination of all the neurons in the CNS is a
complex task and therefore elucidating how these networks of molecular and
neuronal mechanisms interact is a field of constant study and holds many
unrevealed aspects.
The histaminergic system is linked to many neuronal pathways and, by interaction
with receptors in different brain areas, is involved in several complex behaviors,
from hormonal control to cognition. Although the histaminergic system has been
studied for decades, the development of the neurons and the exact functions and
molecular mechanisms of the interactions with other neurotransmitter systems
are still poorly understood. Diseases in which the histaminergic system has been
implicated are Alzheimer’s disease (AD), Parkinson’s disease (PD), schizophrenia,
narcolepsy, and epilepsy. Whether the histaminergic system is involved in the
pathogenesis of these diseases is still unknown. An analysis of linkage in a two-
generation pedigree showed that a mutation in histidine decarboxylase (hdc), the
rate-limiting enzyme in histamine synthesis, is associated with Tourette’s
syndrome (Ercan-Sencicek et al. 2010). This is the first evidence that histamine is
indeed associated with the etiology of a specific disease. These results open an
avenue for speculations and one might argue that histamine plays a crucial role in
modulating and fine-tuning of motor functions. If this is the case, this could
partially explain why the histaminergic transmission and system is altered in
diseases that affect motor functioning, such as in PD. In PD, cognitive impairments
are also evident. Taken together, it is still unclear why the histaminergic system is
altered in diseases with cognitive decline, such as AD and PD, in the sleep disorder

Introduction
13
narcolepsy, or in alertness- and neuronal excitability-associated diseases such as
schizophrenia and epilepsy, respectively. Although the role of histamine in
neuronal plasticity, e.g. during neurodegeneration, has been studied, there are still
many unanswered questions to be solved.
In this thesis, I addressed the factors that affect the development of the
histaminergic system and how this system contributes to mediating of specific
behaviors. I set out to study the neuronal histaminergic system in a vertebrate
species, the zebrafish Danio rerio. The zebrafish has, during the recent decades,
emerged as a highly suitable vertebrate model for biomedical and neuroscientific
research, due to its main features: ex utero development, amenability to genetic
manipulation, and cost-effectiveness. Neuronal and molecular networks
underlying behaviors have been studied successfully in the invertebrates
Caenorhabditis elegans and Drosophila melanogaster, and vertebrate model
organisms, but some unanswered questions still remain.

Review of the literature
14
2 Review of the literature
One of the many challenges in the field of medicine is the identification of specific
connections of neuronal networks that underlie and mediate specific behaviors.
Since the human brain is formed of billions of neurons, it is challenging to pinpoint
the changes in neuronal pathways and the exact mechanisms mediating different
behaviors. Addressing these questions in a vertebrate model organism with lower
neuronal complexity, i.e. the zebrafish, has therefore emerged as an attractive
alternative approach.
2.1 Neuronal networks and plasticity
Neuronal networks, in the biological sense, are those that specific pools of neurons
form with each other to execute certain physiological functions. The ability of the
individual to adapt to a changed environment, as well as the capacity to learn and
remember, are dependent on how well the nervous system is able to form new
and undo old connections, defined as neuronal plasticity. The concept of neuronal
plasticity was initially described in 1890 by James, and the concept was later
refined by several scientists, including Cajal and Hebb (as cited in Berlucchi &
Buchtel 2009). The connections between neurons are formed by specialized
junctions, i.e. synapses. Strengthening or weakening of the synapses and neuronal
circuits underlie learning and memory (Kandel, Schwartz & Jessell 1991).
Alterations in the morphology of the synapses have also been linked to
neurological disorders (Penzes et al. 2011).
The neuronal circuits that underlie behavior in mammals have been extensively
characterized, but the roles of individual neurons in bringing bout precise
behaviors remain to be elucidated (Kandel, Schwartz & Jessell 1991). Behaviors can
vary from simple reflexes to those that require the cooperation of several brain
areas such as the brainstem, telencephalon, cerebellum, basal ganglia, and/or
thalamus (Figure 1). The cortico-striato-thalamic system and cerebellum therefore
control and modulate the functions of the upper motor neurons in mammals. The
release of acetylcholine from the α-motor neurons at myoneuronal junctions
induces muscle contraction and the produced behavior can be recorded. A new
cycle is initiated when somatosensory receptors are activated and the newly
generated information is processed through the spinal cord via modulatory
systems to the motor cortex.

Review of the literature
15
Figure 1. Neuronal circuits that underlie behavior (modified from Kandel, Schwartz &
Jessell 1991). The neuronal pathways that underlie behavior are organized in a hierarchical
manner. The cerebral cortex is the master regulator, integrating motor output control and
somatosensory information. Behaviors can be modulated by the thalamus, basal ganglia,
cerebellum, brain stem, and spinal cord. Dotted lines indicate the connections known in
mammals but still unknown in zebrafish.
2.1.1 Aminergic neurotransmitter systems
Histamine belongs to the group of biogenic amines, together with dopamine,
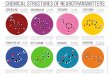
serotonin, adrenaline, and noradrenaline (Figure 2). These neurotransmitter
systems constitute together with γ-aminobutyric acid (GABA) and glutamate, the
classical neurotransmitter systems and networks in the brain. All these
neurotransmitter systems interact to produce complex behaviors such as cognition
and movement, and regulate wake-sleep states (Saper, Scammell & Lu 2005, Haas,
Sergeeva & Selbach 2008, Eriksson et al. 2010). The amines are modulatory
transmitters, since they in general alter the excitability of neurons that are either

Review of the literature
16
excitatory or inhibitory, and mainly exert their actions through volume
transmission (Agnati et al. 1995, Cragg et al. 2001).
Figure 2. Chemical structure of the aminergic neurotransmitters histamine, serotonin, and
dopamine.
Histamine, a biogenic amine, was discovered by Dale and Laidlaw in 1910 and is
one of the phylogenetically oldest neurotransmitters. The first indication that
histamine functions as a neurotransmitter was by the French professor Schwartz in
1970 when he identified the properties and distribution of the histamine
producing enzyme HDC in the rat brain (Schwartz, Lampart & Rose 1970).
Histaminergic neurons were later identified in the tuberomammillary nucleus
(TMN) in the posterior part of the hypothalamus in rat brain independently by two
research groups in 1984 (Panula, Yang & Costa 1984, Watanabe et al. 1984). In the
periphery of the body, histamine is the main mediator of allergic responses acting
through histamine receptor subtype 1 (hrh1). Histamine is involved in an array of
different physiological functions and states, ranging from cognition to hormonal
homeostasis (Haas & Panula 2003, Haas, Sergeeva & Selbach 2008). It was only
recently that a strong association of the neurotransmitter system with a specific
human neuronal disease, Tourette’s syndrome, was established (Ercan-Sencicek et
al. 2010) and shown to be hrh1-mediated (Fernandez et al. 2011). In zebrafish, the
role of histamine has lately been studied in dominance and aggression (Filby et al.
2010, Norton et al. 2011, Pavlidis et al. 2011).
2.1.2 Development of the histaminergic neurotransmitter system
The development of the histaminergic system has been studied in several species,
including zebrafish. In the developing rat brain, histamine is transiently expressed
in the serotonergic raphe nucleus (Vanhala, Yamatodani & Panula 1994). In
zebrafish, the first histaminergic neurons appear in the ventral hypothalamus at 3
days postfertilization (dpf) and efferent projections from these neurons innervate
the dorsal telencephalon by 4 dpf (Eriksson et al. 1998). The histaminergic system
consists of a relatively small, specific, and localized neuron population in the
posterior part of the hypothalamus in zebrafish, and no transient system has been

Review of the literature
17
detected (Eriksson et al. 1998) as in mammals. Considering that the histaminergic
system consists only of a few neurons in the larval zebrafish, the fiber projections
that arise from these neurons are quite elaborate. A detailed study of the intact
fiber network could reveal the precise three-dimensional (3D) innervation
patterns. Although histamine is involved in and regulates many physiological
functions, the factors that regulate the development of this system are not known.
2.1.3 Synthesis, storage, release, and catabolism of the aminergic
neurotransmitters
Histamine is derived from ι-histidine by HDC (EC 4.1.1.22) (Garbarg et al. 1974) and
is packed into vesicles by vesicular monoamine transporter 2 (VMAT2) (Figure 3)
(Merickel & Edwards 1995). After release of histamine into the extracellular space,
histamine is bound to its receptors or inactivated by methylation into tele-
methylhistamine, a process that is mediated via histamine N-methyltransferase
(HNMT, EC 2.1.1.8) (Brown, Tomchick & Axelrod 1959). Tele-methylhistamine is
further oxidized by monoamine oxidase (MAO, EC 1.4.3.4) to methylimidazole
acetaldehyde (Hough & Domino 1979). Two forms of MAO are known, of which
MAO-A metabolizes serotonin (Nicotra et al. 2004) and MAO-B metabolizes
histamine (Schwartz et al. 1991). As in other species, MAO in zebrafish is also
found in the histaminergic neurons (Sallinen et al. 2009), suggesting that MAO may
be responsible for the inactivation of histamine in this species. In nonneuronal
tissue in mammals, histamine is oxidized by diamine oxidase (DAO, EC 1.4.3.3)
(Schayer 1953). In contrast to mammals, DAO expression has been observed in
zebrafish neuronal tissue, specifically in the ventral habenula, which corresponds
to the mammalian lateral habenula (Amo et al. 2010). Histamine synthesis can be
inhibited by α-fluoromethylhistidine (α-FMH), which is an irreversible inhibitor of
HDC (Garbarg et al. 1980). So far, no reuptake system for histamine has been
identified.

Review of the literature
18
Figure 3. Synthesis, storage, release, and catabolism of histamine. ι-Histidine is taken up
into neurons, where it is converted to histamine by HDC. Histamine is packed into vesicles
by VMAT2 in the presynaptic terminals, and after release histamine binds to hrh1, hrh2,
and hrh3 receptors expressed in the brain. In neuronal tissue, histamine is cleared from the
extracellular space by HNMT and MAO. Histamine is also degraded by DAO (pathway
indicated in gray).
Serotonin is derived from ι-tryptophan by two modifications mediated by
tryptophan hydroxylase (TPH, EC 1.14.16.4) (Lovenberg, Jequier & Sjoerdsma
1967) and aromatic ι-amino acid decarboxylase (AADC, EC 4.1.1.28) (Lovenberg,
Weissbach & Udenfriend 1962). Two forms of TPH exist, of which TPH2 is
expressed in the brain and is responsible for the production of neuronal serotonin
(Walther et al. 2003). Serotonin is packed into vesicles by VMAT2, transported to

Review of the literature
19
the nerve terminals, secreted, and bound to serotonin receptors. Serotonin can be
taken up into the cells by serotonin reuptake transporters (Torres, Gainetdinov &
Caron 2003). Inactivation of serotonin is mediated by MAO (Sjoerdsma et al. 1955)
by formation of 5-hydroxyindoleacetic acid (5-HIAA). The synthesis and catabolism
of serotonin can be manipulated by different chemicals, such as p-
chlorophenylalanine (PCPA), a selective irreversible inhibitor of TPH (Koe &
Weissman 1966, Jequier, Lovenberg & Sjoerdsma 1967), ι-deprenyl, which is a
MAO inhibitor (Fowler, Oreland & Callingham 1981), and fluvoxamine, a selective
serotonin reuptake inhibitor (Claassen et al. 1977).
Catecholamines (dopamine, noradrenaline, and adrenaline) are all derived from
the same amino acid, ι-tyrosine, by the following four enzymatic reactions:
tyrosine hydroxylase (TH, EC 1.14.16.2) forming ι-dihydroxyphenylalanine (ι-DOPA)
(Nagatsu, Levitt & Udenfriend 1964), AADC forming dopamine (Lovenberg,
Weissbach & Udenfriend 1962), dopamine β-hydroxylase (DβH, EC 1.14.17.1)
forming noradrenaline (Levin, Levenberg & Kaufman 1960) and
phenylethanolamine Ν-methyl transferase (PNMT, EC 2.1.1.28) forming adrenaline
(Axelrod 1962). The main enzymes in the degradation of amines are MAO and
catechol-O-methyl transferase (COMT, EC 2.1.1.6).
An interesting feature of rodent histaminergic neurons is that they express the
dopamine-producing enzyme, ι-DOPA (Yanovsky et al. 2011). Furthermore
histaminergic neurons take up ι-DOPA, the precursor of dopamine, and are
dopamine-immunoreactive (Yanovsky et al. 2011), suggesting that these neurons
can also produce dopamine.
2.1.4 Histaminergic receptor-signaling system
Aminergic neurotransmission is mediated through G-protein-coupled receptors.
Histamine acts through the four G-protein-coupled histaminergic receptor
subtypes 1, 2, 3, and 4 (hrh1, hrh2, hrh3, and hrh4) (Haas & Panula 2003, Haas,
Sergeeva & Selbach 2008). Of the known receptors, hrh1, hrh2, and hrh3 are
expressed in the vertebrate brain. hrh4 has also been detected in the mammalian
brain, but the highest level of hrh4 messenger RNA (mRNA) is found in the spinal
cord (Strakhova et al. 2009). hrh1 is mainly implicated in regulation of wakefulness
in rodents (Monti, Pellejero & Jantos 1986, Yanai et al. 1998) and hrh2 is mainly
involved in cognitive functions in rodents (Dai et al. 2007). hrh3 is expressed in
mammalian TMN histaminergic neurons and activation of hrh3 reduces the firing
rate of the neurons and thereby reduces the release of histamine (Arrang, Garbarg

Review of the literature
20
& Schwartz 1983, Arrang, Morisset & Gbahou 2007). Furthermore, hrh3 can act as
a presynaptic heteroreceptor (Schwartz et al. 1990) and regulates the release of
several other neurotransmitters (Blandina et al. 1996, Yamamoto et al. 1997,
Schlicker, Werthwein & Zentner 1999, Jang et al. 2001, Pillot et al. 2002b).
Histamine-mediated neuronal excitation is achieved mainly via activation of hrh1
and hrh2 receptors in mammals, whereas hrh3 activation inhibits histamine
release in rats (Arrang, Garbarg & Schwartz 1983). Of the histaminergic receptors,
the crystalline structure of the human HRH1 together with a first-generation hrh1
antagonist, doxepin, has been determined (Shimamura et al. 2011), providing
detailed information on the molecular properties necessary for specific binding of
ligands to hrh1. The histaminergic receptors signal through different intracellular
signal transduction pathways (Haas, Sergeeva & Selbach 2008). In Table 1 the
features of the different histamine receptors are compared. The properties of the
zebrafish histamine receptor subtypes are not well characterized and require
further study.
Table 1. Properties and role of histamine receptors in mammals (modified from Haas,
Sergeeva & Selbach 2008). For abbreviations in the table, please see list on page 8–9.
Feature hrh1 hrh2 hrh3
G protein Gq/11 Gαs Gi/o
Signal transduction PLC, IP3, DAG, Ca2+
,
PKC, AMPK, NF-κB
AC, cAMP, PKA, CREB AC, cAMP, MAPK,
Akt/GSK3
Cellular function Postsynaptic
excitability
and plasticity
Postsynaptic excitability
and plasticity
Presynaptic transmitter
release
and plasticity
Systemic function Behavioral
state and
reinforcement
(arousal),
working memory,
feeding rhythms,
energy metabolism,
endocrine control
Learning
and memory
Blood-brain barrier
control, behavioral
state and
reinforcement
Pathophysiology Disorders of sleep,
eating, memory,
mood, addiction,
pain and neuro-
inflammation
Schizophrenia, pain and
neuro-inflammation
Disorders of sleep,
mood, memory, eating,
addiction, pain and
neuro-inflammation

Review of the literature
21
2.2 Neurotransmitter systems under normal conditions and disease
2.2.1 Alertness and wakefulness
Alertness and wakefulness are specific states and important for the survival of the
individual. Almost a century ago, the Viennese neurologist von Economo noted
that lesions in the junction between the telencephalon and mesencephalon were
associated with profound sleepiness in his patients (von Economo 1930). Later, it
was found that this area harbors the aminergic and peptidergic systems that
constitute the ascending arousal system (AAS) and activate the cerebral cortex and
thalamus (Saper, Scammell & Lu 2005). Histamine in the TMN is part of the AAS
and histaminergic neurons fire during the waking period in mammals (Mochizuki et
al. 1992, Takahashi, Lin & Sakai 2006). The rest of the components of the AAS
include the dopaminergic ventral tegmental area, serotonergic raphe nucleus,
noradrenergic locus coeruleus (Saper, Scammell & Lu 2005), and the
hypocretin/orexin (hcrt) of the lateral hypothalamus (de Lecea et al. 1998). Hcrt
signals through two G-protein-coupled receptor subtypes 1 and 2 (hcrt1 and hcrt2)
in rodents and fish (Trivedi et al. 1998, Panula 2009). Of these two receptors, hcrt2
is expressed at high densities in mammalian histaminergic neurons (Yamanaka et
al. 2002). In rodents histamine and hcrt may be responsible for different aspects of
arousal/wakefulness: histamine mainly responsible for cognitive and motivated
behaviors and hcrt for motor activity (Anaclet et al. 2009). Reduction in the hcrt
level causes narcolepsy in humans (Peyron et al. 2000). The symptoms of
narcolepsy have only been reproduced in hcrt-/- mice, not in hdc-/- mice (Anaclet
et al. 2009). Serotonin, dopamine, galanin, GABA, thyrotropin-releasing hormone
(TRH) and methionine-enkephalin (mENK) are also known to affect wakefulness by
either promotion or inhibition of the AAS (Cirelli 2009). The sleep-promoting
center in the brain is the ventrolateral preoptic nucleus, which consists of
GABAergic and galaninergic neurons (Gaus et al. 2002). These neurons innervate
the nuclei of the AAS (Sherin et al. 1998) and are active during sleep (Szymusiak et
al. 1998). Together with the AAS clusters, these sleep-promoting neurons regulate
alertness and wakefulness in mammals.
Although the above-mentioned neurotransmitter nuclei are located in close
proximity, not many of the neurotransmitters are colocalized. To better
understand the role histaminergic neurons play in different behaviors or
physiological states, the cotransmitters of histaminergic neurons have been
identified in different species to varying degrees. Histamine is not colocalized with
serotonin, dopamine, or hcrt in adult zebrafish and the systems have different

Review of the literature
22
innervation areas (Kaslin & Panula 2001, Kaslin et al. 2004). Serotonergic fibers and
dopaminergic fibers mainly innervate the ventral telencephalon, whereas
histaminergic fibers innervate the dorsal telencephalon (Kaslin & Panula 2001,
Kaslin et al. 2004). The coexistence of histamine with galanin, GABA, TRH, and
mENK has been studied in rodents (Airaksinen et al. 1992, Kukko-Lukjanov &
Panula 2003), but not in zebrafish. In rats, histamine coexist with galanin, TRH, and
GABA in histaminergic neurons (Airaksinen et al. 1992). In guinea pig, histamine is
coexpressed with GABA and mENK in histaminergic neurons, whereas in mice
histamine is colocalized with galanin and GABA in histaminergic neurons
(Airaksinen et al. 1992). In human TMN, the coexistence of histamine with galanin
and GABA was studied, and it was found that histamine is colocalized with GABA,
but not with galanin (Trottier et al. 2002).
2.2.2 Histamine and neurodegenerative diseases
Aminergic neurotransmitters have been indicated in several neurological diseases,
including narcolepsy, AD, PD (Eriksson et al. 2010), and most recently in Tourette’s
syndrome (Ercan-Sencicek et al. 2010). Narcolepsy is a neurodegenerative disease
characterized by fragmented sleep and unpredictable sleeping bouts throughout
the day, and to date there is no cure for this disease (De la Herran-Arita, Guerra-
Crespo & Drucker-Colin 2011). The neurological background is a deficient hcrt
system in humans (Peyron et al. 2000). The etiology is unknown, although research
indicates an autoimmune origin (Scammell 2003, Hallmayer et al. 2009, De la
Herran-Arita, Guerra-Crespo & Drucker-Colin 2011). It is thought that the
mechanism by which hcrt induces wakefulness is by exciting histaminergic neurons
via hcrt2 receptors (Bayer et al. 2001). Cataplexy, i.e. loss of muscle tone, is often
associated with narcolepsy and is usually triggered by emotional excitation (Anic-
Labat et al. 1999). Narcoleptic patients have traditionally been treated with
stimulants, such as amphetamine (Zaharna, Dimitriu & Guilleminault 2010). Due to
the addictive potential of amphetamine, a new class of drugs has been designed.
Modafinil and armodafinil are the best available drugs to treat narcolepsy,
although the mechanism of action is not known (Zaharna, Dimitriu & Guilleminault
2010). The European Medicines Agency has approved selective serotonin reuptake
inhibitors and tricyclic antidepressants for treatment of cataplexy, while the
American Food and Drug Administration has not (Zaharna, Dimitriu &
Guilleminault 2010).

Review of the literature
23
Narcolepsy has been modeled in zebrafish by disruption of the hcrt system, either
by hcrt2 receptor null mutation, hcrt2-/-, (Yokogawa et al. 2007) or hcrt
overexpression (Prober et al. 2006). Both models produce a similar insomnia
phenotype. In humans affected with sleep disorders, histamine and hcrt
concentrations are both reduced (Nishino et al. 2009) and patients with
neurodegenerative disorders display insomnia (Dauvilliers 2007). A comparative
representation of both neurotransmitter systems and their projections in human
and zebrafish brain is available in Figure 4.
Figure 4. Comparative presentation of hcrt and histamine in the central nervous system of
humans (left) and zebrafish (right). The histaminergic and hcrt neurotransmitter systems
are conserved between the two species. The fibers from each neurotransmitter system
innervate the neurons of the other in both humans and zebrafish. The projection pattern of
the neurons is similar in these species, since the fibers innervate the majority of the brain
regions. Please note that the brains are not depicted in correct scale in regard to each
other.
AD is the most common neurodegenerative disease worldwide, with cognitive
decline and dementia as the main manifestations. The neurological hallmarks
include reduced brain size, neurofibrillary tangles, and amyloid plaques (Bishop, Lu
& Yankner 2010). The majority of AD cases are sporadic (Blennow, de Leon &
Zetterberg 2006) and affects persons older than 65 years. The early-onset familial
form of AD has an onset around 30 years (Blennow, de Leon & Zetterberg 2006).
The familial forms of AD are due to mutations in the following genes: amyloid
precursor protein (app), presenilin1 (psen1) and presenilin2 (psen2). Both PSEN1
and PSEN2 are part of the γ-secretase complex (Kimberly et al. 2003), which is
expressed within the membranes of the neurons, including the cell membrane,
endoplasmic reticulum, Golgi apparatus, and mitochondria (Kovacs et al. 1996, Xu
et al. 2002, Hansson et al. 2004) (Figure 5). The cognitive decline associated with

Review of the literature
24
AD is mainly due to reduced acetylcholine levels in the brain, which results from
reduced numbers of neurons and neuronal dysfunction (Whitehouse et al. 1982).
Figure 5. The γ-secretase complex is composed of four proteins, PSEN1 or PSEN2,
presenilin enhancer 2 (PEN-2), nicastrin and anterior pharynx-defective 1 (APH-1), and
resides in the membranous organelles of the cell and the cell membrane. These proteins
are not depicted in the correct relation to each other in the figure.
The γ-secretase complex is thought to have many functions, including cleavage of
APP (Borchelt et al. 1996), regulation of the calcium (Ca2+) homeostasis (Guo et al.
1996, Stutzmann et al. 2004), cleavage of the Notch receptor (Levitan & Greenwald
1995), and involvement in Wnt/β-catenin signaling (Serban et al. 2005). Notch
signaling is important in embryonic neural development and in the adult brain
(Ables et al. 2011). In zebrafish, inhibition of γ-secretase with DAPT (N-[(3,5-
Difluorophenyl)acetyl]-L-alanyl-2-phenyl]glycine-1,1-dimethylethylester) affects
Notch signaling (Geling et al. 2002). PSEN1 functions also independently of the γ-
secretase complex. The regulation of Wnt/β-catenin signaling is an example of this
(De Strooper & Annaert 2010). In this case, PSEN1 is important for degradation of
β-catenin (Kang et al. 2002) and lack of PSEN1 is associated with an increase in β-
catenin signaling (Xia et al. 2001). Wnt/β-catenin signaling, as is Notch signaling, is
an important signaling cascade during embryonic development, but also plays a
role in the adult CNS (Inestrosa & Arenas 2010) and in the progression of AD
(Boonen, van Tijn & Zivkovic 2009).
Interestingly, amyloid formation is regulated by hcrt and the sleep-wake cycle,
indicating that there may be a correlation between the AAS and pathogenesis of
AD (Kang et al. 2009). Kang et al. (2009) showed that lack of hcrt would protect
against AD, but a study on the coexistence of narcolepsy with AD found that also
narcoleptics are affected by AD (Scammell et al. 2011). In postmortem brains of AD
patients, the histamine content is reduced (Panula et al. 1998, Shan et al. 2012).
The underlying mechanism for this alteration in histamine content is not known.

Review of the literature
25
One possible mechanism may be associated with hypoxia, since human HDC
expression is regulated by hypoxia-inducible factor 1 (Jeong et al. 2009) and
hypoxia in general is associated with the progression of neurodegenerative
disorders, such as AD (Zlokovic 2011). So far, it is not known if neuronal HDC can
be induced by hypoxia. Furthermore, histaminergic neurons exert neuroprotective
effects in organotypic slice preparations during excitotoxic insults (Kukko-Lukjanov
et al. 2006). Other possible mechanisms include alteration in neurogenesis and/or
proliferation during development (Wen et al. 2002, Chevallier et al. 2005,
Rodriguez, Jones & Verkhratsky 2009) or activity-dependent neurotransmitter
specification (Spitzer 2012), that later on in adulthood may affect the balance of
neurotransmitter systems in normal and diseased brains. Histamine receptor
expression has been examined in the prefrontal cortex of AD patients. In human
brain samples, hrh1 binding was decreased (Higuchi et al. 2000), hrh2 mRNA was
unaltered (Perry et al. 1998, Shan et al. 2012), and hrh3 mRNA was increased (Shan
et al. 2012). The increase in hrh3 mRNA was only observed in female AD patients
(Shan et al. 2012). Taken together, understanding of the mechanisms that underlie
altered histaminergic neurotransmission will provide further insight into AD
progress. There is no cure for AD currently, and treatments can only ease the
symptoms of the disease. Acetylcholinesterase inhibitors and N-methyl-D-aspartic
acid antagonists (Wollen 2010) are widely used treatments. Stimulatory therapies,
which include physical exercise, socialization, cognitive training, and music, are
emerging as new therapeutics (Wollen 2010). Since the life expectancy of our
population is increasing, new therapies, but especially better understanding of the
etiology of the disease, are needed to combat the progression of AD. In zebrafish,
the role of psen1 has been studied by transiently inhibiting gene expression.
Transient knockdown of psen1 in zebrafish (Nornes et al. 2003, 2008, 2009)
phenocopies the features reported for the psen1 knockout mice (Shen et al. 1997),
i.e. disrupted CNS formation and Notch signaling.
2.3 Zebrafish as a vertebrate model organism
The zebrafish belongs to the bony fishes (Teleosts) of the ray-finned fishes
(Actinopterygii), which comprise the majority of all living fish. The actinopterygian
fishes and mammals shared an ancestor about 500 million years ago (Metscher &
Ahlberg 1999, Grillner & Jessell 2009). The zebrafish is a vertebrate model
organism that was initially used in developmental biology studies during the 1970s.
During the last decade, the popularity of this vertebrate model has grown in the
field of neuroscience (Panula et al. 2006, 2010, Sager, Bai & Burton 2010). The

Review of the literature
26
main advantages, such as its small size (approximately 4 cm in length) and
transparent ex utero development in combination with feasibilities of genetic
manipulation, have rendered it a good model for studying neuronal circuits
underlying behavior. Importantly, the neuroanatomy of zebrafish correlates well
with that of other vertebrates studied, which is one of the reasons it can be
utilized as a model for studying the mechanisms underlying human disease. In
addition, tracing and lesion studies have also been undertaken to elucidate the
neuroanatomy and the role of different brain areas in fish and how these can be
correlated with other vertebrates. The similarity in organization of various
neurotransmitter systems among several vertebrate species has further motivated
the use of zebrafish as a model in neuroscience. Due to the small size of the
zebrafish, 3D imaging of whole neurotransmitter systems is possible. These
methods, combined with targeted stimulation or ablation of single neurons, render
the zebrafish a powerful tool for studying the role of specific neurons (Burgess &
Granato 2007b). For example, Ca2+ imaging can be used to correlate the activity of
a neuron with specific stimuli (O'Malley, Kao & Fetcho 1996, Orger et al. 2008).
Other possible tools that are currently available include optogenetics, which allows
noninvasive activation of neurons within millisecond ranges (Szobota et al. 2007,
Douglass et al. 2008), and high-throughput pharmacological experiments
combined with a behavioral assay (Rihel et al. 2010). The availability of a large
number of transgenic and mutant fish lines combined with the above-mentioned
appealing approaches for studying behavior makes this an excellent model
organism for use in neuroscience.
2.3.1 Comparative neuroanatomy of fish and mammals
The telencephalon develops in actinopterygian fishes via eversion, whereas in
other vertebrates it develops via evagination. The difference in development was
postulated to be the reason for the nonlaminar telencephalon in actinopterygian
fishes (Ito & Yamamoto 2009). In actinopterygian fishes, the telencephalon can be
grossly divided into the dorsal telencephalon, i.e. the pallium, and the ventral
telencephalon, i.e. the subpallium (Nieuwenhuys 1963, 2010, Nieuwenhuys, ten
Donkelar & Nicholson 1998) (Figure 6). Based on tracing and gene expression
studies the basal ganglia, including the striatum, was suggested to be located in
the ventral telencephalon (Rink & Wullimann 2001, Mueller, Wullimann & Guo
2008, Ganz et al. 2011). Toxicity studies, neuronal connectivity, and tracing studies
have suggested that the teleost correlate to the mammalian substantia nigra,
which is part of the basal ganglia, is located in the diencephalon of larval zebrafish

Review of the literature
27
(Kaslin & Panula 2001, Rink & Wullimann 2004, Sallinen et al. 2009). The dorsal
part of the dorsal telencephalon (Dd, Figure 6) in goldfish may correspond to the
mammalian cerebral cortex (Vargas, Lopez & Portavella 2009). Recent results
indicate that the Dd region does not exist in zebrafish and that the central part of
the dorsal telencephalon (Dc) corresponds to the mammalian cortex (Mueller et al.
2011). Based on lesion studies of the medial pallium, the area corresponding to the
mammalian amygdala may be located in the medial region of the dorsal
telencephalon (Dm, Figure 6) in goldfish (Portavella et al. 2002, Portavella & Vargas
2005). Similar lesion studies of the lateral pallium suggest that the phylogenetically
conserved area that corresponds to the mammalian hippocampus in goldfish is in
the lateral region of the dorsal telencephalon (Dl, Figure 6) (Portavella, Torres &
Salas 2004, Portavella & Vargas 2005). The dorsal telencephalon is highly
innervated by histaminergic fibers (Kaslin & Panula 2001). In contrast,
dopaminergic, serotonergic, and hcrt fibers innervate the ventral telencephalon
(Kaslin & Panula 2001, Kaslin et al. 2004).
Figure 6. Comparative analysis of the telencephalic areas in fish (left) and mammals (right).
Dd = dorsal area of dorsal telencephalon, Dm = medial area of dorsal telencephalon, Dl =
lateral area of dorsal telencephalon, Dc = central area of dorsal telencephalon, Dp =
posterior area of dorsal telencephalon. In zebrafish, the area that is suggested to
correspond to the mammalian cortex is Dc, whereas studies in goldfish proposed Dd.
The thalamic complex in mammals is a major structure of the diencephalon and
consists of three regions: prethalamus, zona limitans intrathalamica, and thalamus.
The development of the thalamus is evolutionarily conserved, as suggested by a
comparative analysis of mouse, chicken, and zebrafish. The study demonstrated
that homologous genes are responsible for development of the thalamic nuclei in
these three different species (Scholpp & Lumsden 2010).

Review of the literature
28
The hypothalamus is located below the thalamus and is connected to the pituitary
(Nieuwenhuys, ten Donkelar & Nicholson 1998). By release of neuropeptides and
neurotransmitters, the hypothalamus maintains the homeostasis of the organism
and is involved in endocrine and metabolic control of body functions and
behaviors. For example, arousal is tightly regulated by the nuclei in the
hypothalamus (Scammell 2003). The mammalian and zebrafish hypothalamus are
not anatomically very alike, but the equivalents of the neuronal nuclei that are
found in the mammalian hypothalamus are also found in zebrafish (Machluf,
Gutnick & Levkowitz 2011). The ventrocaudal hypothalamus is the site of the
histaminergic neurons, and some of the serotonergic, dopaminergic, and hcrt
neuron populations are also located in this region in zebrafish (Kaslin & Panula
2001, Kaslin et al. 2004). To evaluate the role of the cerebellum in modulation of
motor behaviors, the cerebellar anatomy has been described and mutations that
affect the development of the cerebellum have been produced (Bae et al. 2009).
2.3.2 Zebrafish behavior and motor functions
At as early as 17 hours postfertilization (hpf), zebrafish larvae show spontaneous
movement (Brustein et al. 2003). The activity of the fish increases during the
course of a few days and at 4–5 dpf, a mature beat-and-glide swimming pattern
can already be observed. At this age, the larvae begin to hunt for food, since the
nutrients of the yolk sac have been almost completely consumed. Fundamental
behaviors, such as prey capture (Budick & O'Malley 2000) and sleep (Prober et al.
2006, Zhdanova 2006), can be assessed in zebrafish larvae.
The neuronal circuits in the spinal cord and brainstem that underlie behavior have
been studied in zebrafish (discussed below). In contrast, the involvement of the
telencephalon in execution of behaviors has not been assessed (Figure 1). To date,
there are no identified neuronal circuits that connect the telencephalon with the
brainstem (Figure 1). The optic tectum of zebrafish corresponds to the superior
colliculus of mammals and in both cases the main task is to integrate visual
information with motor output (Isa 2002, Sato et al. 2007, Del Bene et al. 2010).
The brainstem nuclei regulate the lower motor neurons and mediate modulatory
inputs from the telencephalon and cerebellum. The spinal cord harbors the motor
and sensory connections and it is this area that coordinates several reflexes.
To link specific neurons with neuronal networks and to identify the circuits
underlying behavior, specific behaviors can be dissected into smaller units. This
allows the identification of which movement patterns zebrafish larvae combine to

Review of the literature
29
produce specific behaviors. Dependent on the behavior studied, movement
patterns can be identified by different methods. Identification of the movement
patterns zebrafish combine to produce behavior, such as prey capture or startle
response, requires high-frequency imaging (500–1000 frames per second). Others,
such as sleep, place preference, and general activity, can be assessed with low-
frequency imaging (5 frames per second or less). Different stimuli elicit distinct
responses in zebrafish larvae. Acoustic/vibrational stimuli induce a rapid escape
response (Burgess & Granato 2007b), whereas dark flashes induces a slower
response (Burgess & Granato 2007a). For navigational purposes, adjustment
moves such as turns and scoots (Burgess, Schoch & Granato 2010) and slow-
swimming with the pectoral fins can be detected (Budick & O'Malley 2000).
The reticulospinal neurons in the brainstem of larval zebrafish project to the spinal
cord to regulate execution of many behaviors. The Mauthner neuron (Figure 7) is
the best known and is thought to be the sole driver of the escape response, which
is also called the startle response. The startle response of freely moving larval
zebrafish can be divided into short-latency and long-latency startle responses
(Burgess & Granato 2007b). Both types of startle responses last for several
milliseconds and are completed in less than 1 second after stimulation. The short-
latency startle response is mediated by the Mauthner neuron, whereas the long-
latency startle response is not (Burgess & Granato 2007b). Other neurons of the
brainstem, such as MID2cm (Mauthner neuron homologue, middle dorsal 2
contralateral medial longitudinal fasciculus interneuron in rhombomere 5) and
MID3cm (Mauthner neuron homologue, middle dorsal 3 contralateral medial
longitudinal fasciculus interneuron in rhombomere 6) cooperate with the
Mauthner neuron in giving the escape/startle response a direction (Figure 7)
(O'Malley, Kao & Fetcho 1996). Stimulation of the head causes both the Mauthner
neuron and MID2cm or MID3cm neuron to respond, whereas when the tail is
stimulated, only the Mauthner neuron is activated (O'Malley, Kao & Fetcho 1996).
Among the medial longitudinal fasciculus (nMLF) neurons active during forward
swimming have been identified (Orger et al. 2008). In the same study, neurons that
execute right or left movements were also identified. A schematic drawing of the
anatomical position of the neurons described is presented in Figure 7 and their
connections via commisural interneurons to the lower motor neurons in the spinal
cord are also illustrated.

Review of the literature
30
Figure 7. Lateral view of a zebrafish brain. The main brain areas are indicated and the
nMLF, Mauthner neuron, MID2cm, MID3cm, and the relationship to the histaminergic
neurons and their projections are illustrated. Furthermore, the interaction of the lower
motor neurons with the interneurons is visualized.
In both larval and adult zebrafish, the response to changes in the environment can
be assessed. This can either be studied by moving white and black stripes
unidirectionally under the freely moving fish, which causes an optomotor
response, or by fixing the animals and monitoring their eye movements, which in
turn causes an optokinetic response. These two assays have successfully been used
to screen for visual system defects in mutants (Brockerhoff et al. 1995, Muto et al.
2005). Changes in illumination mediate a Mauthner neuron-independent
behavioral response that has been classified as an О-bend, which may be strictly
navigational (Burgess & Granato 2007a, Burgess, Schoch & Granato 2010). In this
study, we assessed a similar response, i.e. an increase in locomotor activity when
zebrafish larvae are exposed to sudden changes in illumination. We call this
response a dark-induced flash response.
2.3.3 Behavioral pharmacology
Pharmacological experiments are easily performed in zebrafish larvae, since they
can be immersed in drugs. High-throughput screening of thousands of molecules
combined with a behavioral readout has been successfully developed to study
sleep-wake regulation in zebrafish larvae (Rihel et al. 2010). Depending on the
properties of the drug, an appropriate solvent must be used to ease the uptake of
the drug into the larvae. Furthermore, the blood-brain barrier in zebrafish begins
to develop at 3 dpf (Xie et al. 2010), but is not fully functional until 9 dpf (Jeong et
al. 2008). The timeline for the development and maturation of the blood-brain
barrier therefore offers a window when drugs are easily taken up into the brain.

Review of the literature
31
2.3.4 Modification of gene expression with morpholino oligonucleotides
To transiently inhibit gene expression, morpholino oligonucleotides (MOs) have
been developed. The MOs can be designed to inhibit gene expression via two
different approaches: 1) by inhibiting the binding of the ribosomal subunits to the
mRNA initiation site by targeting the MO upstream of the initiation site or 2) by
interfering with the splicing of the premature mRNA by targeting the MO to the
exon-intron or vice versa boundary. The MOs are injected into zebrafish embryos
at the 1–4-cell stage. Since the effect of the MO is only transient and usually ceases
within the first week of life, the effect of transient downregulation can only be
assessed in larval zebrafish. Care must be taken that the expression of the gene of
interest is downregulated throughout the observation time. This can be done by
verifying loss of the protein by immunohistochemistry (IHC) or Western blotting. If
a splice site MO is used, then the expression of mature mRNA can be assessed by
real-time polymerase chain reaction (RT-PCR). MOs quite often also induce off-
target effects, such as edema and increased cell death (Robu et al. 2007, Eisen &
Smith 2008). Different approaches to control the specificity of the MO have
therefore been proposed. For example, several MOs could be used to induce the
same phenotype and activation of the p53 pathway could be assessed by
quantitative real-time polymerase chain reaction (qPCR) or by coinjecting MO the
that targets the p53 pathway, and rescue of the phenotype is recommended by
injection of target gene mRNA. There are some problems with these approaches,
since some target genes may indeed be involved in the upregulation of cell death
pathways (Sallinen et al. 2010).
2.3.5 Modification of gene expression by inducing mutations into the genome
Chemical mutagenesis was applied to zebrafish almost two decades ago (Driever et
al. 1996) and this approach required forward genetics to identify the mutation-
carrying gene. To speed up the process, the TILLING (Targeting induced localized
lesions in genomes) tool was developed. TILLING is a reverse genetic approach by
which knockout animals can be obtained. TILLING was initially set up for
Arabidopsis thaliana (McCallum et al. 2000, Colbert et al. 2001) and later modified
to allow target-selected mutagenesis in zebrafish (Wienholds et al. 2002, 2003).
Random gain-of-function or loss-of-function mutations are induced into genomes
by treating male zebrafish with a highly potent mutagen, N-ethyl-N-nitrosourea
(ENU, chemical formula C3H7N3O2), which targets the spermatogonial stem cells
(Driever et al. 1996). After a recovery period the ENU-treated males are mated

Review of the literature
32
with healthy wild-type (WT) female zebrafish to produce an F1 offspring
generation. Genomic DNA is prepared from fin clips of this live library or from
sperm kept in a frozen library, and the individuals are screened for mutations in a
specific gene. The plant endonuclease CEL-I can be used for detection of the
mutations, since it detects all of the currently known sequence variants, including
deletions, insertions, and missense alterations (Oleykowski et al. 1998). Since ENU
is a highly potent mutagen that induces random mutations throughout the
genome, it is advisable to outcross identified carriers for several generations to
reduce the number of background mutations.
Designed targeted mutagenesis is possible via the zinc finger nuclease (ZFN) and
TALEN (Transcription activator-like effector nuclease) methods (Caroll, Zhang
2011). These two methods have higher specificity for the genomic target sequence
and thereby produce fewer off-target effects (Doyon et al. 2008, Meng et al. 2008)
than random mutagenesis induced with ENU. ZFN and TALEN function in a similar
manner. They are heterodimers that consist of a catalytic unit (FokI endonuclease)
that introduces double-stranded breaks at specific genomic sites, and a targeting
unit that is a DNA-binding unit consisting of several DNA-binding modules. The
double-stranded breaks are then repaired by the DNA repair machinery, which
introduces small insertions or deletions that can cause frameshift knockout
mutations (Bibikova et al. 2003). The difference between ZFN and TALEN is that
each ZFN DNA-binding module recognizes three nucleotides, whereas the TALEN
DNA-binding module recognizes only one, resulting in higher specificity (Caroll &
Zhang 2011). The ZFN method has been available for a decade (Smith et al. 2000),
but due to the high uncertainty with the ZFN design, it has not been used
extensively in zebrafish research. Due to the favorable aspects and increased
knowledge of the methods, the ZFN and TALEN methods will most likely also
become more commonly used in zebrafish research.
2.3.6 Spatially and temporally targeted gene expression
Methods for targeted gene expression in desired tissues have also been described.
The medaka (Oryzias latipes) Tol2 transposon system in combination with the
Gal4/UAS (upstream activating sequence) system has been used to express genes
downstream of the UAS in specific neurons (Halpern et al. 2008, Asakawa &
Kawakami 2009, Suster et al. 2009). In this manner, the neural circuits underlying
the touch response in zebrafish were unraveled. Inhibition of neurotransmitter
release from Gal4-positive hindbrain interneurons and Rohon-Beard sensory

Review of the literature
33
neurons by expression of tetanus toxin (TeTx) under the UAS resulted in lack of a
touch response (Asakawa et al. 2008, Asakawa & Kawakami 2009). This method for
spatially targeted gene expression can also be further modified to express genes in
a temporal manner (Halpern et al. 2008). This method requires, as does TILLING,
large facilities for housing and screening the different Gal4 fish lines that later on
are crossed with UAS fish.
2.3.7 Appropriateness of modeling human disease in a simple vertebrate
The molecular pathways underlying many disorders in humans are still unknown
and new inexpensive models are needed to contribute in the quest to reveal these
mechanisms. Invertebrates such as Caenorhabditis elegans and Drosophila
melanogaster have already provided insight into some of the mechanisms
underlying human disease processes. The zebrafish was initially used as a model
organism in developmental studies but has during the past decade become more
widely used to model various human diseases, ranging from disorders of the
nervous system, e.g. attention deficit hyperactivity disorder (Lange et al. 2012), to
leukemia (Ridges et al. 2012). The zebrafish is, due to its small size, less expensive
to keep in large quantities than are rodents. In addition, zebrafish may by the
public be considered as a more ethical choice than mammals as disease models.
The major drawback of zebrafish in modeling human disease is that it has
undergone the genome duplication event, and therefore some of the genes in the
zebrafish are duplicated, whereas in other cases one gene in zebrafish carries the
properties of both human homologues. An example of this is the enzyme
responsible for converting ι-tyrosine to ι-DOPA, TH, which in zebrafish is duplicated
(Chen, Priyadarshini & Panula 2009). So far, the function of zebrafish TH2 is
unknown and zebrafish TH1 converts ι-tyrosine to ι-DOPA, just as in humans.
Another example is MAO in zebrafish, which carries features of both human MAO-
A and MAO-B (Anichtchik et al. 2006). Naturally, the novel insights obtained in
zebrafish must be studied in mammalian model organisms and humans before
conclusions regarding the findings are applicable to humans and human disease.
Taken together, the above-mentioned results indicate that the zebrafish is an
appropriate tool for modeling human disease to answer mechanistic questions.

Aims of the study
34
3 Aims of the study
The main goal of this thesis project was to study the functions of neuronal
histamine; the interactions of the system with other neurotransmitter systems, its
role in neuronal plasticity, and the neuronal circuits underlying behavior. The
zebrafish was chosen as a model organism and various methods for quantifying
larval zebrafish behavior were established.
The specific aims of the thesis were the following:
I Identify and characterize the histamine receptors in zebrafish and
the behavioral effects of their ligands,
II Identify the cotransmitters of the histaminergic neurons,
III Study the role of histamine in alertness and wakefulness,
IV Elucidate the factors that regulate the development of histamine
neurons by characterizing the system in psen1 knockout fish, and
V Assess the role of MAO in behaviors driven by amines.

Methods
35
4 Methods
Please find more detailed information regarding the materials and methods in the
original publications.
4.1 Animals
Several zebrafish strains were used in this thesis. For the studies in I–III and V, a
WT strain that originated from Abo Akademi University and which was used in the
following studies (Kaslin & Panula 2001, Peitsaro et al. 2003, Kaslin et al. 2004,
Sallinen et al. 2009, 2010) was used. For the results in unpublished form (IV), a
homozygote TILLING mutant line carrying a premature stop codon in the psen1
gene (hu2547) was used. This line was a kind gift from Dr. Zivkovic (Hubrecht
Institute of Developmental Biology and Stem Cell Research, Royal Netherlands
Academy of Arts and Sciences, Amsterdam, and University Medical Center,
Utrecht, The Netherlands). The mutation was induced in the Tubingen longfin (TL)
strain and we therefore used the TL strain as the control WT line in studies with
the mutant. Fish handling and housing (+26–28 °C, 14:10 light-dark cycle) were
done as previously described (Westerfield 2007). The permits for the experiments
were obtained from the Animal Experiment Board of the Regional State
Administrative Agency for Southern Finland, in agreement with the ethical
guidelines of the European Convention.
4.2 Manipulation of gene expression by morpholino oligonucleotides (III)
We designed an MO that targeted the initiation site of the hdc mRNA (atg), 5’-
CATCCCGTCACTCAGAGAAGATTAG-3’. A standard (std) control MO was used as the
control (both MOs from Gene Tools). At 5–6 hpf, the success rate of the injection
was observed in a fluorescence microscope, since the MOs were tagged with
fluorescein. Any uninjected embryos within the MO-injected groups were excluded
from the experiment. The injected embryos were grown until 5 dpf or older, when
the experiments were conducted. Uninjected WT animals were used as additional
controls.
4.3 Pharmacological treatments
For the pharmacological studies, zebrafish larvae were dechorionated with forceps
at 24 hpf and raised in 6-well plates with 3 ml of embryo medium (E3: 5 mM NaCl,

Methods
36
0.17 mM KCl, 0.33 mM CaCl2, 0.33 mM MgSO4, all reagents from Sigma-Aldrich) to
which the drug was added. The group size varied between n=10–35. If the drug
treatment was started at 5 dpf or later, the fish were raised in petri dishes until the
appropriate age. The E3 medium with the drug was usually changed once daily.
4.3.1 Treatment with histamine receptor ligands or α-fluoromethylhistidine (III)
Zebrafish larvae were treated with 1, 10, or 100 μM pyrilamine (Sigma-Aldrich),
cimetidine (Sigma-Aldrich), thioperamide or immepip. The thioperamide and
immepip were kind gifts from Professors Rob Leurs and Henk Timmerman (Vrije
Universiteit, Amsterdam, The Netherlands). The control groups were kept in E3
medium. The treatments were initiated at 2–3 hpf, 1 dpf, or 2 h before the
behavior was assessed. The locomotor activity was observed at 5 dpf. The α-FMH
was a kind gift from Dr. Kollonitch (Merck) and the treatments were 1 mM for 1–5
or 1–7 dpf before behavior (please see 4.4) or hcrt whole-mount in situ
hybridization (WISH, please see 4.9).
4.3.2 Deprenyl, p-chlorophenylalanine, and fluvoxamine treatments (V)
Fish exposed to solutions containing deprenyl (Sigma-Aldrich) at 1, 10, or 100 μM
were further divided into three groups. The fish were exposed to the drug for
different times, either 0–7 dpf, 0–5 dpf or only 2 h preceding testing. For deprenyl
and PCPA (Sigma-Aldrich) treatment, the fish were divided into six groups and
exposed to 150 μM or 1.5 mM PCPA for 1–5 dpf and/or 100 μM deprenyl 0–5 dpf.
The fluvoxamine (Sigma-Aldrich) treatments were also done in combination with
deprenyl (100 μM 0–5 dpf) or alone by exposing zebrafish to 100 μM fluvoxamine
2 h before the animals were killed at 5 dpf. The control groups received no drugs.
4.3.3 γ-Secretase inhibitor DAPT treatments (IV)
The DAPT in solution (Tocris Bioscience) was diluted in dimethyl sulfoxide (DMSO)
to reach a final concentration of 2.5 mM. At 1 dpf, 30 fish per treatment group
were dechorionated, moved to 6-well plates, and kept in 3 ml of E3 medium
containing either 1% DMSO or DAPT. After the treatment, the fish were killed and
fixed with 4% 1-ethyl-3,3(dimethyl-aminopropyl) carbodiimide (EDAC; CMS
Chemicals) diluted in 0.1 M phosphate buffer, pH 7.0.

Methods
37
4.4 Behavioral tests
To assess the effect of histamine on behavior, several behavioral assays were
developed during the course of this thesis. The behavioral experiments were done
during the light period of the day, unless otherwise mentioned. The zeitberger
time (ZT) onset was at 9.00 in the morning (ZT0) and lights went off at 23.00 in the
evening (ZT14). The behavioral data were either collected by i) observing the fish
and manually scoring the specific behaviors or ii) a video camera and further
analyzed with EthoVision 3.1 software (Noldus Information Technology). Advanced
calibration of the arenas was applied in each case when the EthoVision software
was used. All behavioral experiments were done at room temperature (RT) and
repeated three times with different fish.
4.4.1 Locomotor activity of zebrafish (I, III–V)
The larvae were placed in individual hemisphere-shaped chambers (diameter 4 cm,
volume 12 ml) or in the wells of a 48-well plate (volume 1 ml) and analyzed for 10
min at a sample rate of 5–25 frames per second. The total distance moved (cm),
turn angle (degrees), and movement (percentage of time fish spent moving) were
calculated from the coordinates acquired, using the EthoVision software. System
noise was removed, using an input filter of 0.2 cm for minimum distance moved.
The tracks were excluded if the sample size (i.e. the number of frames in which the
animal was detected) did not exceed 90% of the maximum sample size or if
reflections were detected. The locomotor activity (total distance moved, cm) of
adult zebrafish was analyzed in the same manner as for the larvae, with the only
difference being that the arenas used were larger (diameter 22 cm, volume 2 l).
4.4.2 Place preference (IV)
Place preference was assessed in the same arenas as the locomotor activity. The
arenas were divided into several zones (in EthoVision) and the time the fish spent
in each zone during the tracking period was determined.
4.4.3 Place preference in a water column (V)
A vertical place preference test was done in a 2-ml cylinder filled with E3 medium.
Two groups were used: a control group without drug treatment and a ι-deprenyl
group with treatment of 100 μM ι-deprenyl for 0–5 days. Ten 5-day-old larvae
were placed in a cylinder and allowed to accommodate for 5 min. The cylinder was

Methods
38
divided into upper and lower parts. The number of fish in the lower part was
counted every 5 min for 25 min.
4.4.4 Locomotor activity of larval zebrafish over a 24-h period (III)
The activity and frequency of movement were observed during a 24-h period. The
zebrafish larvae (5 dpf) were placed individually in the wells of white 96-well plates
5 hours before tracking, which was started at ZT14. A video camera connected to a
computer with EthoVision software was used for tracking the fish illuminated by
infrared lights. Two parameters were analyzed: frequency of movement and total
distance moved per hour.
4.4.5 Dark-induced flash responses in larval zebrafish (III, IV)
In this behavioral test, we studied whether light per se affects the activity of fish.
We set up an assay to approach this, which was similar to assays in previous
studies in which related behaviors were assessed (Burgess & Granato 2007a,
Burgess & Granato 2007b, Emran, Rihel & Dowling 2008). Light-induced decrease
in locomotor activity and dark-induced flash response of morphant fish were
assessed by observing 5 dpf fish during 2 min periods of lights on and off for 10
min. The fish were transferred to 48-well plates on the day of observation,
illuminated by infrared lights, and their movement was recorded with the same
system as described above.
4.4.6 Optomotor response (III)
An optomotor response assay was developed for the initial zebrafish mutation
screen to identify visual system defects in mutants. We used a simplified version of
the optomotor assay by manually moving a paper with black and white stripes (1-
cm-wide stripes) for 30 seconds under a plate with 6–7 dpf zebrafish larvae and
observing whether or not the larvae adjusted their direction of swimming in the
direction of movement of the stripes.
4.5 Lipophilic dye DiI tracing (III)
Micropipettes were filled with 20% carbocyanine dye DiI (Molecular
Probes/Invitrogen Corporation) solution (diluted in 1:1 ethanol:DMSO). After
anesthetization of the fish, the needle was placed perpendicularly to the body axis
and 2 nl of the tracer was injected, by PV830 Pneumatic PicoPump (World
Precision Instruments Inc.) at a pressure of 30 psi for 5 ms, through the skin to the

Methods
39
superficial part of the right dorsal telencephalon of 9 dpf larvae. The injection was
done under a stereomicroscope and targeting was verified immediately with a
Leica MZFBL III fluorescence microscope (Leica Microsystems GmbH). After the
injection, the larvae were kept alive 1 h, killed and fixed with ice-cold 4% EDAC.
4.6 Real-time polymerase chain reaction
4.6.1 Cloning of zebrafish hrh1, hrh2, and hrh3 receptors in adult zebrafish (I)
Sanger Institute zebrafish blast pages (http://www.ensembl.org/Danio_rerio) were
used to identify sequences that are similar to the human hrh1, hrh2, and hrh3
receptors. The coding sequences predicted for the receptors were amplified, using
PCR. RNA isolated from adult zebrafish brains was used as a template for the
reactions. The RNA was extracted with RNAwiz™ (Ambion Inc.) and
complementary DNA (cDNA) was reverse-transcribed, by the First-Strand Synthesis
Kit (Amersham Biosciences) or by the SuperScript™ First-Strand Synthesis System
(Invitrogen Corporation) according to the manufacturers’ instructions. The
following primers were used: hrh1 (DQ647806) forward 5′-
TGTCTCTCCTCACCGTCATC-3′ and reverse 5′-CAATCCCTTCATTTACCCGCTCT-3′; hrh2
(DQ647808) forward 5′-ATGCAATTTATATTCAGCGAT-3′ and reverse 5′-
ACTACCTTTTACTCCCATTAGC-3′; hrh3 (DQ647807) forward 5′-
ATGGAGAGAGAAAACGCG-3′ and reverse 5′-AATACTTCTGGTCAATGTTC-3′. The
amplified PCR products were sequenced from both directions (PRISM 310; Applied
Biosystems Inc.).
4.6.2 Expression of hrh1, hrh2, and hrh3 mRNA (I)
RNA was extracted from fish at different developmental stages (3, 6, 11, 24, 48,
and 72 hpf, 5, 7, and 10 dpf) from freshly isolated brains, eyes, gills, intestine,
heart, liver, muscle, and spleen from 10 adult fish and reverse-transcribed as
described above. cDNA originating from 0.25 μg total RNA was used for the PCR
reactions, which were carried out with the same primer pairs as above for hrh1,
hrh2, and hrh3.
4.6.3 Cloning full-length hdc cDNA and synthesis of capped hdc mRNA (III)
The 5' half of zebrafish hdc cDNA was amplified with primers, forward 5’-
ATGCAGCCGCAGGAGTACAT-3’ and reverse 5’-GGCTGCCGGGTCACGACT-3’, with
the partial hdc clone (Yokogawa et al. 2007) as PCR template. The 3' half was
amplified with forward 5’-GTCCTGAGCTGCGCTATTTC-3’ and reverse 5’-

Methods
40
GTTATTTAGTAGGCAGCGGTGTG-3’ primers by RT-PCR from total RNA extracted
from adult zebrafish hypothalamus. The full-length hdc cDNA was acquired by
splice overlap extension (SOE) PCR, with forward 5’-ATGCAGCCGCAGGAGTACAT-3’
and reverse 5’-GTTATTTAGTAGGCAGCGGTGTG-3’ primers. All the above-
mentioned PCR reactions were done with Phusion high-fidelity DNA polymerase
(Finnzymes), which makes blunt end PCR products. The 3’ end of the full-length
fragment acquired was modified to increase the stability of the fragment with an
adenosine-tailing procedure that adds 3´ adenosine overhangs. This modification
was accomplished with GoTaq master mix (Promega), followed by
thymine/adenine-cloning. The modified full-length hdc cDNA was cloned into a
pGEM-Teasy vector and sequenced. It was inserted into a pMC vector at the EcoRI
and SpeI restriction enzyme sites (Fink et al. 2006). Capped RNA (mRNA) was
synthesized with T7 mMESSAGE mMACHINE (Ambion) after linearization of the
cloned plasmid with a NarI and purified with Qiagen RNeasy Mini Kit (Qiagen),
after DNase I digestion for removing template DNA. The purified capped hdc RNA
(mRNA) was injected into the yolk of embryos at the 1–4 cell stage.
4.6.4 Quantitative real-time polymerase chain reaction (III, IV)
RNA was isolated from 30 pooled 5 dpf larvae using a Qiagen RNeasy® Mini Kit,
following the protocol supplied by the manufacturer (Qiagen). RNA quality and
amount were analyzed spectrophotometrically and cDNA was prepared by
SuperScript III® (Invitrogen), primers used for qPCR were the following: hcrt A
(NM001077392) forward 5’-TCTACGAGATGCTGTGCCGAG-3’ and reverse 5’-
CGTTTGCCAAGAGTGAGAATC-3’ (Kaslin et al. 2004), galanin (NM_015973) forward
5’-GACCAACTGATACTCAGGATGCA-3’ and reverse 5´-
ATCCCGAGTGTTTCTGTCAGAA-3’, and β-actin (AF057040) forward 5’-
CGAGCAGGAGATGGGAACC-3’ and reverse 5’-CAACGGAAACGCTCATTGC-3’ (Keegan
et al. 2002).
4.6.5 Touchdown RT-PCR for identification of mutation in psen1-/- (IV)
Adult WT and psen1-/- (hu2547) zebrafish were anesthetized in clove oil and
genomic DNA was isolated from finclips with buffer (50 mM KCl, 2.5 mM MgCl, 10
mM Tris-HCl, 0.45% NP40, 0.45% Tween 20, 0.01% gelatine, and 1.6 µg/ml
poteinase K). For identification of the mutation, standard touchdown RT-PCR was
performed with the following primer pairs: forward 5’-GAAACCCATTTGGGAAGTG-
3’ and reverse 5’-AAATACAAGCCTACACACAACC-3’, and forward 5’-
AGGAAACAGCTATGACCAT GAACGCAGAAGAATGAACG-3’ and reverse 5’-

Methods
41
TGTAAAACGACGGCCAGT CAAGTTTGACATTTGCATGG-3’. The mutation was located
to exon 5, where it induced a premature stop codon.
4.7 Sequence comparisons and analysis of histamine receptors (I)
Multisequence alignments were performed, using ClustalW on Web pages
provided by the European Molecular Biology Laboratory-European Bioinformatics
Institute, Hixton, Cambridgeshire, UK, and the sequence analyses were carried out
with the GENSCAN Web server at Massachusetts Institute of Technology,
Cambridge, MA, USA. Sequence comparisons were carried out with ALIGN
provided by European Molecular Biology Laboratory-European Bioinformatics
Institute.
4.8 [125I]iodoaminopotentidine binding to hrh2 (I)
[125I]iodoaminopotentidine was synthesized as described (Leurs et al. 1994) and
was a kind gift from Professors Leurs and Timmerman (Vrije Universiteit).
4.9 Whole-mount in situ hybridization (III, IV)
Zebrafish MAO (AY185211) (Anichtchik et al. 2006), hcrt (NM_001077392) (Kaslin
et al. 2004), hrh1 and hrh3 (DQ647806 and DQ647807) (Peitsaro et al. 2007) were
cloned into pGEM-Teasy vector as described earlier. The partial coding region of
hdc (NM001102593) (Yokogawa et al. 2007) was cloned into pGEM-Teasy vector,
using forward 5’-GCAGCCGCAGGAGTACATGC-3’ and reverse 5’-
GGCTGCCGGGTCACGACT-3’ primers. gad1 (NM194419.1), gad2 (NM001017708.2),
slc17a6a (NM001009982.1), slc17a6b (NM001128821.1), and slc17a7
(NM001098755.1) (the last five abbreviated in the list on page 8–9) were cloned
and were kind gifts from Dr. Higashijima (Higashijima, Mandel & Fetcho 2004)
(State University of New York, Albany, NY, USA). The Notch1a (NP_571516.1) clone
was a kind gift from Dr. Brand (Center for Regenerative Therapies Dresden,
Biotechnology Center and University of Technology Dresden, Germany). Antisense
and sense digoxigenin-labeled RNA probes were synthesized using the digoxigenin
RNA labeling kit (Roche Diagnostics), following the instructions of the
manufacturer. In situ hybridization was performed as previously described (Thisse
& Thisse 2008) with some modifications as described (Chen, Priyadarshini & Panula
2009, Kudo et al. 2009).

Methods
42
4.10 Immunohistochemistry (II–V)
For optimal staining for histamine IHC, it was necessary to use the bifunctional
fixative EDAC, due to the properties of the anti-histamine antibody (Auvinen &
Panula 1988, Panula et al. 1989, 1990). For histamine IHC, the samples were fixed
in 4% EDAC and 0.1% paraformaldehyde (PFA; Sigma-Aldrich) in 0.1 M phosphate
buffer, pH 7.0, overnight (o/n) at +4 °C. For other antibody staining, PFA fixation (2
or 4%) in 0.1 M phosphate buffer, pH 7.4, o/n at +4 °C was used. Brains from larval
zebrafish were dissected after fixation and processed for IHC immediately. Adult
brains that were intended for sectioning were incubated in 20% sucrose in 0.1 M
phosphate buffer, pH 7.4, o/n at +4 °C, then embedded and sectioned at 20 µm
with a Leica cryostat. The sections were stored at -20 °C until further processing
for IHC. No sucrose treatment was performed on the brains that were processed as
whole-mounts. The preabsorption controls for some of the primary antisera were
reported earlier (Panula et al. 1990, Kaslin & Panula 2001, Kaslin et al. 2004).
4.10.1 IHC with antibodies from different species on whole-mount larval brains
(II–V)
The following primary antibodies were used and incubated o/n at +4 °C: rabbit
anti-histamine 19C (Panula et al. 1990) 1:10 000, mouse anti-TH 1:1000
(LOT22941, DiaSorin Inc.), rabbit anti-serotonin 1:1000 (S5545, Sigma-Aldrich),
rabbit anti-active caspase 3 1:500 (clone C92-605, 559565, BD Biosciences), rabbit
anti-hcrt A 1:1000 (AB3704, Millipore/Chemicon), rabbit anti-β-catenin (clone
2206, LOT056K4763, Sigma-Aldrich), and rabbit anti-galanin antibody 1:1000–
1:5000 (AB1985 and AB5909, Millipore/Chemicon). The secondary antibodies were
highly cross-purified Alexa-fluorophore-conjugated goat antirabbit or antimouse
antibodies (with 488, 561, or 647 fluorophores; Molecular Probes) incubated o/n
at +4 °C.
4.10.2 IHC on whole-mount adult brains (II, IV)
IHC on whole-mount adult brains was performed as for larval brains; the only
difference was that the incubation times were prolonged (primary antibody
incubations 2–4 days). The following primary antibodies were used: rabbit anti-
histamine 19C (Panula et al. 1990) 1:10 000, mouse anti-TH 1:1000 (Diasorin Inc.),
and rat anti-serotonin 1:250 (MAB352, Millipore/Chemicon). The secondary
antibodies were highly cross-purified Alexa-fluorophore-conjugated goat

Methods
43
antirabbit, antimouse and/or antirat antibodies (with 488 or 561 fluorophores;
Molecular Probes).
4.10.3 IHC with two antibodies from the same species on cryosections (II, III)
A tyramide signal amplification kit (TSA kit; PerkinElmer Life and Analytical
Sciences) was used to enable double-staining of samples with primary antibodies
from the same species. The preabsorption controls are listed in Table 2. IHC with
TSA was performed according to the manufacturer's instructions. After the TSA-
IHC, the samples were stained to obtain the desired double-staining and mounted
in 1:1 glycerol in phosphate-buffered saline.
Table 2. Preabsorption controls for the antibodies used in the cotransmitter study (more
detailed information can be found in II).
Preabsorption controls
Antigen Preabsorption
conjugate/peptide
Manufacturer,
catalog number
Dilution used
(of conjugate/peptide)
GABA GABA-sOVA Karhunen et al., 1993 20 µg/ml
Galanin Human galanin peptide BACHEM, H-8230 10 µM
Galanin Porcine galanin peptide BACHEM, H-1365 50µM
Histamine Histamine-sOVA Panula et al., 1984 2 µg/ml
mENK mENK peptide Sigma-Aldrich, E-5757/ M-
6638
100 µM
TRH TRH peptide Pennisula Laboratories, H-
4915
10 µM
The following primary antisera and antibodies were used: rabbit anti-histamine
19C 1:50 000 (Panula et al. 1990), rabbit anti-GABA 1H 1:1000 (Karhunen et al.
1993), rabbit anti-TRH #4319 1:1000 (Airaksinen et al. 1992), rabbit anti-mENK #36
1:1000 (Airaksinen et al. 1992), rabbit anti-hcrt A 1:1000 (AB3704,
Millipore/Chemicon), and rabbit anti-galanin antibodies 1:1000–1:5000 (AB1985
and AB5909, Millipore/Chemicon). The secondary antibody was a biotinylated goat
antirabbit antibody 1:750 (Vectastain ABC kit, Vector Laboratories Inc.). Avidin-
conjugated Texas Red (Vector Laboratories) 1:500 and streptavidin-conjugated
fluorescein 1:50 (Molecular Probes) and highly cross-purified Alexa-fluorophore-
conjugated goat antirabbit antibodies (with 488 or 561 fluorophores, Molecular
Probes) were used to visualize the primary staining.

Methods
44
4.10.4 Enzyme histochemistry and activity assay (V)
An MAO oxidase enzyme histochemistry and activity assay was carried out, as
described (Anichtchik et al. 2006).
4.11 High-pressure liquid chromatography
4.11.1 Catecholamine and serotonin HPLC (V)
Catecholamines and their metabolites were measured from snap-frozen samples
that had been stored at -80 °C. A total of 15–25 larvae were pooled and
homogenized in 150 μl of cold solution containing 0.4 M perchloric acid, 0.1%
Na2S2O5, and 0.1% ethylenediaminetetraacetic acid. After centrifugation, an
aliquot of the supernatant was injected into the HPLC system. The HPLC system
that was used for dopamine, noradrenaline, adrenaline, 3,4-dihydroxyphenylacetic
acid, 5-HIAA and 3-methoxytyramine measurements comprised a Waters pump
model 515 (Waters Corporation), Waters Autosampler 717plus, and an ESA
Colochem 5100A electrochemical detector (ESA). The HPLC system used for
serotonin measurements included a Waters pump model 510, Waters
Autosampler 717plus and Concorde electrochemical detector (Waters
Corporation).
4.11.2 Histamine HPLC (III)
Histamine was assayed with HPLC, as described earlier (Eriksson et al. 1998).
Briefly, samples were collected by pooling 10 or 30 larvae from which the eyes and
tails were removed. These samples were frozen on dry ice and stored at -80 °C
until further processing for HPLC. The samples were then homogenized by
sonication in 2% perchloric acid, centrifuged for 30 min at 15 000 g at +4 °C, and
filtered through a 0.45 µm polyvinylidene diflouride filter (Pall Life Sciences) before
loading into the HPLC system. The protein concentration was measured by the
BCA-kit (Thermo Fisher Scientific Inc.) to normalize the results from HPLC
measurement.
4.12 Microscopy
Samples of MAO WISH were examined, using an Olympus microscope (Olympus
Corporation) connected through a charge-coupled device camera to the AnalySIS
software (Soft Imaging Software GbmH). WISH samples of hdc, hrh1, hrh3, hcrt,
gad1, gad2, slc17a6a, slc17a6b, and slc17a7 were analyzed with an inverted Leica

Methods
45
DM IRB microscope. The images were acquired with a Leica DFC490 camera and
Leica Application Suite, MultiFocus option (Leica Microsystems).
The IHC samples were all analyzed with a Leica TCS SP2 AOBS confocal microscope.
The excitation wavelength of the argon laser for samples visualized with
streptavidin-fluorescein and the Alexa 488 fluorophore was 488 nm and the
emission was collected at 500–550 nm. The excitation wavelength of the diode
laser used for samples stained with the Alexa 568 fluorophore was 561 nm and the
emission was collected at 600–700 nm (narrower emission spectra when triple-
staining was used to prevent overlap). A helium/neon laser was used to excite the
Alexa 647 fluorophore at 633 nm and the emission was collected at 640–750 nm.
In all cases, sequential scanning between the frames and frame averaging were
applied to minimize cross-talk between the channels. For the 3D analysis, the
optimal step size calculated by the Leica software was applied.
4.13 Image analysis
The brains were scanned with optimized step size and averaging to allow 3D
rendering of the data in Imaris software (Bitplane AG). In general, maximum
projections of the stacks acquired were produced with Leica software for
comparative analysis. The images were further processed using ImageJ or Fiji. All
processing was applied to whole images. The 3D-rendered images were exported
as snapshots and as video files. Quantification of the histamine neuron number
was done by counting the neurons from all the stacks of images that had been
acquired and scanned with optimal settings.
For the colocalization study (II), a single sagittal section was studied from each
adult zebrafish brain. The section with the most histamine-immunoreactive
neurons was chosen. All the histamine-immunoreactive neurons were counted, as
well as the number of cells containing both histamine and the other neuronal
markers of interest. We were interested in which neurotransmitters are
colocalized with histamine in histaminergic neurons, and not the surrounding
fibers. Thus, we counted the neurons and utilized no software for assessing
colocalization, because it would have detected each pixel and not distinguish
between the fibers and the somata of the neurons. Single focal planes of the
overlay images were analyzed for colocalization with Leica software or open-
source image analysis software Fiji (http://pacific.mpi-
cbg.de/wiki/index.php/Main_Page). The maximum projections of the stacks
acquired were produced with Leica software for comparative analysis.

Methods
46
The anatomical structures were identified and/or numbered, using the atlas of the
adult zebrafish brain (Wullimann, Rupp & Reichert 1996), and the atlas based on
the location of the aminergic neurons in adult zebrafish (Kaslin & Panula 2001). All
pictures were compiled into panels in CorelDraw (Corel Corporation).
4.14 Statistical methods
For statistical analysis between two groups, the Student’s t-test or Mann-Whitney
U-test were used (GraphPad Prism Software or Microsoft Office Excel 2007). For
statistical analysis between several groups, one-way analysis of variance (ANOVA)
was applied with Tukey’s Multiple Comparison post hoc test in GraphPad. To
assess the interaction between time and treatment two-way repeated measures
(RM) ANOVA was applied to the material in GraphPad Prism. The graphs were
plotted as mean values and the variation was indicated with standard error of the
mean (SEM), unless otherwise described.

Results
47
5 Results
Please consult the original publications in the appendix for more detailed
information.
5.1 Identification and characterization of the histamine receptors in
zebrafish and the behavioral effects of their ligands (I, III)
The innervation pattern of the histaminergic neurotransmitter system was studied
previously in zebrafish (Eriksson et al. 1998, Kaslin & Panula 2001). In contrast, the
distribution of the histamine receptors has so far been unknown in zebrafish and
the specific site of action for histaminergic innervation is unknown in any species.
Thus, a detailed map of the distribution of the receptors could reveal the site of
action of the histaminergic neurons. The mRNA of the three histamine receptors
hrh1, hrh2, and hrh3, were already expressed from 3 hpf onwards in the
developing zebrafish embryo, as detected with RT-PCR. Histamine receptor mRNA
in situ hybridization revealed expression patterns in different parts of the zebrafish
brain (Figure 8). hrh1 was mainly expressed in the dorsal telencephalon, with low
levels of expression in the ventral telencephalon and diencephalon of larval fish
brain. hrh2 binding was detected with [125I]iodoaminopotentidine in adult fish
brain sections in the optic tectum, brainstem, and caudal hypothalamus, which
also harbors the histaminergic hdc-positiv, neurons. The receptor with the most
restricted neuronal expression pattern was hrh3, being located only in the dorsal
telencephalon and ventrocaudal hypothalamus of larval zebrafish. Furthermore,
we identified the histamine receptor-dense area as glutamatergic, because
markers for the glutamatergic neurons were mainly found in the dorsal
telencephalon and the GABAergic markers in the ventral telencephalon.
Figure 8. Histamine receptor expression in the zebrafish brain. Lateral view of the brain.
The hrh3 mRNA had the most restricted expression pattern, whereas hrh1 mRNA was
expressed both in the telencephalon and diencephalon. The data is a merged presentation
of the findings in I and III.

Results
48
To assess the functionality of the receptors, the zebrafish larvae were treated with
histamine receptor ligands. Treatment with different concentrations of the hrh1
antagonist pyrilamine, hrh2 antagonist cimetidine, and hrh3 inverse agonist
thioperamide and hrh3 agonist immepip resulted in altered behavior, which
suggested that the receptors were functional. Each of the histamine receptor
ligands tested significantly reduced the locomotor activity of zebrafish larvae
during a short 10 min tracking period, whereas the turn angle of the fish was not
altered. Pyrilamine at the highest dose, 100 µM, caused the strongest effect
compared with that of control animals (p ≤ 0.001). Surprisingly cimetidine
decreased the movement at all doses in comparison to that of control animals (1
µM, p ≤ 0.05, 10 µM, p ≤ 0.05 and 100 µM, p ≤ 0.001). Both thioperamide and
immepip strongly affected on movement with the highest dose (100 µM, p ≤ 0.001
and p ≤ 0.05, respectively), compared with the control animals.
5.2 Cotransmitters of the histaminergic neurons (II, III)
A quantitative comparison of the histaminergic system in the brains of adult male
and female zebrafish was done to rule out sex-related differences in the animals.
No significant difference between the sexes was found (p ≤ 0.57) and therefore
further studies were done on groups of fish in which both sexes were represented.
To assess the similarity of zebrafish histaminergic neurons with other species, the
cotransmitters of these cells were studied. We found that the pattern of the
various neurotransmitters in zebrafish was similar to that documented in rats
(Airaksinen et al. 1992). Galanin was the most commonly observed neuropeptide
that coexists with histamine, since about 30% of the histaminergic neurons
observed were positive for both markers. GABA and histamine were colocalized in
14% of the neurons and TRH coexisted with histamine in about 7% of the neurons.
mENK and hcrt were never observed to coexist with histamine.
In the whole-mount adult brain, the contacts between the histaminergic system
and the serotonergic and dopaminergic systems were assessed. Histamine neurons
surround the caudal zone of the periventricular hypothalamus (Hc), which harbors
both serotonergic and dopaminergic neurons (Kaslin & Panula 2001). Histamine-
immunoreactive fibers are scarce in the region that harbors the serotonergic and
dopaminergic neurons. We observed that the histaminergic fibers innervated only
the ventrolateral part of the Hc.

Results
49
5.3 Histaminergic system in alertness/wakefulness (III)
Histamine neurons are active during the waking period and silenced during sleep
(Lin et al. 2011). Translation inhibition of hdc, which is essential for the conversion
of the amino acid ι-histidine to histamine, with MOs resulted in significantly lower
histamine levels in zebrafish larvae at 5 dpf when morphants were compared with
control animals (75% reduction in hdc MO group, p ≤ 0.05). Furthermore, the
histamine immunoreactivity was almost completely abolished in hdc MO-injected
animals, compared with both WT controls and std MO-injected fish. At 24 hpf and
6 dpf, the hdc MO-injected fish showed no abnormal gross phenotypes. At 5 dpf,
cell death was not increased, nor was any defects observed in other related
neurotransmitter systems, such as the dopaminergic system. Inhibition of hdc
impaired locomotor activity during the light period (Figure 9). Two-way RM ANOVA
revealed a significant interaction between the WT and hdc MO groups (p ≤ 0.001,
Figure 9), whereas the std MO did not induce a significant effect. These results
indicate that the effect was specific to the hdc MO.
Figure 9. Transient inhibition of hdc induced a decline in locomotor activity during the light
phase of a 24-h period in 5–6 dpf old larvae. Findings compiled from III.
Since the activity of the larvae was altered during the light period, we
hypothesized that the decreased activity would be due to attenuated alertness
rather than a motor dysfunction. We assessed the optomotor response of the hdc
MO-injected fish and found that it was not significantly altered compared with that
of WT siblings. To assess whether the decreased activity attenuated alertness, we
subjected the zebrafish larvae to periods of lights on and off. The dark-induced
flash response (consisting of an O-bend (Burgess & Granato 2007a)) is elicited in
WT fish as a response to the onset of the dark period and lasts for less than a
second. Inhibition of hdc interfered with the response of larval fish to
environmental cues and the dark-induced flash response was therefore impaired in
hdc MO-injected fish when compared with noninjected WT control fish (Figure 10).

Results
50
This difference was confirmed by a significant time and treatment interaction (p ≤
0.05, Figure 10) when WT and hdc MO fish were compared. In line with these
observations, the same impairment of the response to environmental cues could
be detected when WT larvae were treated with the hrh1 antagonist pyrilamine (p ≤
0.001, Figure 10). The dark-induced flash response was recovered in hdc MO-
injected larvae that were coinjected with hdc mRNA (p>0.05).
Figure 10. Histamine drives the dark-induced flash response in larval zebrafish in a hrh1-
dependent manner. The data are compiled from III.
The strongest mRNA expression of hrh1 was detected in the dorsal telencephalon
of larval zebrafish and this area is highly innervated by histaminergic afferent
fibers. We hypothesized that the dorsal telencephalon would be the main area
that controls alertness-associated behavior. To further study the neuronal circuits
that underlie this behavior, we traced the connections of the dorsal telencephalon
by injecting a lipophilic dye, DiI, into this area. We found that the fibers from the
dorsal telencephalon innervated the histaminergic ventrocaudal hypothalamus.
The distribution of TH-immunoreactive neurons was used as an anatomical map to
locate the targets of the DiI-labeled fibers. No fibers were positive for both DiI and
histamine. This fact supports the idea that the target area of the dorsal
telencephalon would indeed be the ventrocaudal hypothalamus, since it was not
the histaminergic fibers per se that were labeled with DiI.
5.4 γ-Secretase regulates the development of histamine neurons (IV)
The histaminergic neurons play an important role in alertness and many
neurological disorders (Haas, Sergeeva & Selbach 2008). However, little is known
about which factors mediate the development of this system. We found that,
when γ-secretase functioning is disturbed due to a loss of function mutation in
psen1, the development of the histaminergic neurotransmitter system was altered
(Figure 11). Expression analysis of hdc mRNA at 7 dpf revealed significantly fewer

Results
51
hdc-positive neurons in psen1-/- than in WT controls (p ≤ 0.001). Similarly, at 7 and
8 dpf the histamine-immunoreactive neuron number in the psen1-/- fish was lower
than in WT fish (p ≤ 0.001 and p ≤ 0.01, respectively).
Figure 11. Loss of function mutation of psen1, which is part of the γ-secretase complex,
impaired the development of the histaminergic neurotransmitter system at the larval
stage. DAPT treatment induced the same phenotype. The γ-secretase complex consists of
PSEN1, nicastrin, APH-1, and PEN-2. The psen1 mutant zebrafish carries a premature stop
codon in the third extracellular loop. The part of psen1 that is missing in the mutant is
visualized in the image as a dotted line. Unpublished results, IV.
The same impairment of development of the histaminergic neurons was found
when larval zebrafish were treated with an inhibitor of γ-secretase, DAPT (Figure
11). DAPT treatment for 1–5 dpf negatively affected development of the
histaminergic neurons and when compared with WT and DMSO treated siblings
there were significantly fewer histamine-positive neurons in the DAPT-treated
animals (p ≤ 0.05). A shorter treatment time with DAPT from 6–8 dpf had the same
effect on development of the histaminergic system (p ≤ 0.05).
To verify that psen1-/- specifically affected the development of the histaminergic
system, we also assessed the development of the galaninergic (mRNA and protein
level) and dopaminergic (by TH-IHC) systems. There was no difference in the
development of these systems when WT and psen1-/- animals were compared at 7
dpf. Furthermore, we could not detect an increase in cell death in the psen1-/-
compared with the WT siblings.

Results
52
5.5 Neuronal plasticity and the histamine system (III, IV)
We found that in psen1-/- zebrafish the histaminergic neurotransmitter system
was temporally and spatially altered in the brain compared with WT fish (Figure
12). At the larval stages, fewer histamine-positive neurons in the psen1-/- fish
were observed than in the WT (p ≤ 0.001). At increasing age, the difference
between the WT and the mutants was reduced. In 2-month-old animals, the
histamine neuron number in the psen1-/- fish was not significantly different from
that in the WT (p > 0.05). This state was not stable and changed with time, since
the histamine neuron number was significantly (about 30%) increased in psen1-/-
fish compared with the WT controls (p ≤ 0.001) when the animals were 1 year old
(Figure 12).
Figure 12. The development of the histaminergic neurotransmitter system is temporally
altered in psen1-/- zebrafish. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001. Unpublished results, IV.
At the age when the psen1-/- fish had significantly lower numbers of histaminergic
neurons, the behavior of the animals was also changed. At 6 dpf the psen1-/-
larvae failed to exhibit the dark-induced flash response, a histamine-mediated
behavior. Furthermore, at 7 dpf the psen1-/- larvae swam significantly less than
the WT fish (p ≤ 0.001). The locomotor activity of adult psen1-/- fish was unaltered
compared with the WT controls. However, the mutants exhibited an increased
thigmotaxis (p ≤ 0.05), since they avoided the inner zone of the arena. The
behavioral phenotypes were associated with the changes in the histaminergic
system.
After hdc knockdown, we found that the hcrt system did not develop as in WT
siblings. Histamine regulated the development of the hcrt neurons, since hdc MO-
injections reduced hcrt mRNA, as measured both by in situ hybridization (Figure
13, p ≤ 0.05, WT and hdc MO) and qPCR (p ≤ 0.05). The hcrt neuron number was

Results
53
rescued when the embryos were coinjected with hdc MO and hdc mRNA. Injection
of 800 pg of hdc mRNA alone induced an increase in the hcrt neuron number
compared with WT controls (p ≤ 0.05).
Figure 13. Rescue of hcrt neurons in zebrafish after injection of hdc mRNA. *p ≤ 0.05, **p ≤
0.01. The data are compiled from III.
Treatment of WT larvae with the inhibitor of HDC, α-FMH, induced the same
reduction in hcrt neuron number (Figure 14 b, p ≤ 0.05) that was observed after
inhibition of hdc with MO. In situ hybridization of hcrt and hrh1 in separate
samples revealed that these two transcripts are found in the same region (Figure
14 a). This finding led us to question whether hcrt neurons could be regulated in a
hrh1-dependent manner. Indeed, the regulation of the developing hcrt
neurotransmitter system was hrh1-dependent. This was evident, because the hrh1
antagonist pyrilamine reduced the hcrt neuron number (Figure 14 c, p ≤ 0.001).

Results
54
Figure 14. Development of the hcrt neurons in zebrafish. a) Histamine receptor mRNA
expression in comparison to hcrt neurons. b) Treatment of larvae with α-FMH caused
impairment in the developing hcrt system. c) Histamine regulated hcrt neuron
development in a hrh1-dependent manner, since the hrh1 antagonist pyrilamine interfered
with the development of the hcrt system, *p ≤ 0.05, ***p ≤ 0.001. The data are compiled
from III.
5.6 Monoamine oxidase affects locomotor activity via serotonin (V)
In zebrafish, MAO is found in dopaminergic, serotonergic, and histaminergic
neurons (Sallinen et al. 2009). The role of MAO was assessed in amine-driven
behaviors, such as locomotor activity. Chronic (0–7 dpf) treatment of zebrafish
larvae with the MAO inhibitor ι-deprenyl reduced the locomotor activity of larval
zebrafish in a dose-dependent manner at 7 dpf. When the larvae were allowed to
recover for 2 days, the behavior also returned to the level of the controls. Acute (2
h) treatment just prior to assessment of behavior induced a significant decline in
movement. We also assessed the vertical place preference in a water column of ι-
deprenyl-treated larvae and found that these swam closer to the water surface
than the controls (p ≤ 0.0001). Interestingly, we found that inhibition of MAO
increased the serotonin content 100-fold, but did not affect the metabolism of
other amines (dopamine and noradrenaline) that were measured with HPLC. The
increase in serotonin was associated with a decline in locomotor activity and an
increase in ectopic serotonin-positive neurons (Figure 15).

Results
55
Figure 15. MAO drives serotonin-mediated behaviors in larval zebrafish. ι-Deprenyl inhibits
MAO and thereby the conversion of serotonin to its metabolite, 5-HIAA. This causes an
increase in serotonin concentration and decrease in locomotor activity. PCPA is an inhibitor
of TPH and by exposure of larvae to both chemicals (ι-deprenyl and TPH), PCPA inhibits the
elevation of serotonin. The data are compiled from V.
To verify whether serotonin was responsible for the phenotypes observed, we
treated larvae with drugs that interfere with serotonin synthesis (TPH inhibitor
PCPA) and uptake (selective serotonin reuptake inhibitor fluvoxamine). The
behavioral deficit was rescued by treatment of the ι-deprenyl-exposed larvae with
PCPA. HPLC analysis revealed that PCPA treatment rescued the effect that ι-
deprenyl had on serotonin increase and therefore allowed serotonin to remain at
control levels. The ectopic serotonin-positive neurons were abolished when the ι-
deprenyl-treated animals were treated with the selective serotonin reuptake
inhibitor fluvoxamine.

Discussion
56
6 Discussion
6.1 The methods – drawbacks and strengths
To date, there are several available zebrafish strains; however the interstrain
differences have not been studied extensively. In zebrafish, the differences in the
genetic backgrounds of the various WT strains do not affect spontaneous
locomotor activity (Bretaud et al. 2011). The zebrafish strain that was used for four
of the publications in this thesis has previously been used in several studies
(Eriksson et al. 1998, Kaslin & Panula 2001, Kaslin et al. 2004, Anichtchik et al.
2006, Sallinen et al. 2009, 2010, Perala et al. 2010) and the behaviors observed for
the adult and larval fish are reproducible (above-mentioned publications
compared with results in I–IV).
Genetic manipulation of gene expression can either be transient through MOs or
then stable by induction of mutations into genomes, followed by forward or
reverse genetic screening of the animals for mutations. At a glance, the MO
approach seems to be the easiest. The main strength with MOs is that they
constitute a quick and inexpensive approach to study the function of specific
genes. The main problem is that many of the MOs have short-lasting effects that
are no longer effective when the fish are 1-week-old and start to exhibit
quantifiable behaviors. Furthermore, many MOs induce unspecific off-target
effects and therefore it has been stressed that several MOs should be used to
reproduce the phenotypes of interest (Robu et al. 2007). We did this, but found
that the splice site blocking MOs did not recapitulate the phenotype observed with
initiation site blocking MOs, although the phenotype was phenocopied by other
means of intervention of histaminergic signaling (III). Therefore many different
approaches to recapitulate the same phenotype provide a good approach. The
behavioral analysis was done mainly in 1-week-old larval zebrafish. At this age, the
fish already hunt for food, since they have consumed the nutrients from the yolk
sac, and show many quantifiable behaviors. The drugs can be taken up at this late
stage (I, III), because the blood-brain barrier of the zebrafish is not fully functional
at 7 dpf and the initiation site blocking MO we used was still effective at this age
(III). The behavioral effects observed were therefore due to the MO and/or
pharmacological interventions used. In the behavioral experiments, the number of
individuals used per group in each experiment was quite high and was needed to
reduce the standard deviation and variation seen within the groups. This is one of

Discussion
57
the major drawbacks with the behavioral assay approach, but can quite easily be
circumvented since fish lay many eggs per clutch.
IHC with two primary antibodies from the same species was used in the
colocalization study (II). The drawback of this approach is that we had to use
antibodies from the same species, which could have resulted in some false-positive
detection. This approach worked, because we used the appropriate controls
required. We also ensured that there was no overlap between the light paths,
neither excitation nor emission, of the confocal microscope. While scanning the
images, care was taken to ensure that the best possible resolution was used and
also that the averaging of each channel was performed to reduce background
interference and thereby excess detection. In a few cases, deconvolution was
applied to the images, but we observed that this did not affect the final outcome
of the image, suggesting that the IHC staining procedure and confocal microscope
settings were optimal.
6.2 Neuronal circuits underlying behavior (III, V)
In this thesis, we observed that histamine and serotonin affected behavior. We
found that hdc knockdown abolished the dark-induced flash response and that
pharmacological inhibition of hrh1 resulted in the same behavioral phenotype.
These results indicated that histamine via hrh1 is an important mediator of rapid
behavioral responses that are elicited by sudden changes in the environment. In
agreement with our results, studies using hdc knockout mice have shown that in
situations when increased alertness is required, hdc knockouts fail to respond
(Ohtsu et al. 2001, Parmentier et al. 2002, Dere et al. 2004). Knockout of hrh1 in
mice results in reduced locomotor activity (Inoue et al. 1996). Contrasting results
have been reported in rats, since hrh1 antagonism with pyrilamine increases the
startle response and attenuates prepulse inhibition (Roegge et al. 2007). This
discrepancy between the genetic and pharmacological models may be due to
differences in uptake of or sensitivity properties of the hrh1 ligand. The dorsal
telencephalon showed the highest hrh1 and hrh3 mRNA expression in zebrafish.
The expression patterns of inhibitory GABAergic and excitatory glutamatergic
neuron markers revealed that the dorsal telencephalon was mainly glutamatergic.
IHC analysis showed that the dorsal telencephalon in zebrafish larvae also harbors
GABAergic neurons (Perala et al. 2010). Taken together, these data suggest that
hrh1 and hrh3 are mainly expressed in glutamatergic postsynaptic
nonhistaminergic neurons in the dorsal telencephalon in zebrafish. Previous

Discussion
58
studies with mammals showed that histamine receptors are expressed in a laminar
manner in the cortex of rodents (Pillot et al. 2002a, Haas, Sergeeva & Selbach
2008) and humans (Jin & Panula 2005, Jin, Anichtchik & Panula 2009). The dorsal
telencephalon of the zebrafish brain is highly innervated by histaminergic fibers
(Kaslin & Panula 2001). Our tract-tracing study of the projections from the dorsal
telencephalon with the lipophilic dye DiI revealed that the fibers terminated in the
histaminergic ventrocaudal hypothalamus and other areas in zebrafish. None of
the fibers were doubly positive for both DiI and histamine. Antero- and retrograde
tracing identified the limbic telencephalon as the main input area in rat
histaminergic tuberomammillary neurons (Ericson, Blomqvist & Kohler 1991). Only
a few fibers were reported to enter the core of the TMN (Ericson, Blomqvist &
Kohler 1991) in the same manner as we observed in zebrafish. Based on these
data, we propose that in zebrafish a hypothalamic-telencephalic loop exists that
mediates the alertness-associated behavior we observed as a dark-induced flash
response.
In larval fish, lack of histamine affected alertness-associated behavior, whereas
inhibition of MAO with ι-deprenyl increased mainly serotonin and reduced
locomotion. In mammals, the main substrate for MAO is dopamine, which is one of
the amines that mediate locomotion. PD is a result of degeneration of the
dopaminergic neurons in the substantia nigra. The main symptom of PD is the
motor deficit that emerges when a large proportion of the niagral dopaminergic
neurons have died. Modeling PD with neurotoxins in zebrafish causes a locomotor
deficit that can be partially rescued by ι-deprenyl treatment (Sallinen et al. 2009).
Deprenyl-mediated inhibition of MAO in rats elevated dopamine levels (Molinengo
& Ghi 1997), whereas in zebrafish we found that the main substrate for MAO was
serotonin. In humans, altered serotonin levels are associated with depression,
anxiety, and the serotonin syndrome. The serotonin syndrome is caused by
increased levels of serotonin in the brain, leading to consciousness impairment,
including coma, agitation, rigidity, and tachycardia (Radomski et al. 2000, Ables &
Nagubilli 2010). Administration of the serotonin receptor agonist into the raphe
nucleus in rats reduced spontaneous locomotor activity (Hillegaart 1990) and
injections into the periventricular nucleus of the hypothalamus increased inactivity
(Osei-Owusu et al. 2005). The effect is thought to be mediated via reduced activity
of the serotonergic projections in areas controlling motor function in the
telencephalon (Hillegaart 1990). In our experiments, inhibition of MAO with ι-
deprenyl increased the serotonin level in the brains of zebrafish larvae and was
associated with increased heart rate, reduced locomotion, and preference for

Discussion
59
remaining close to the water surface. The reduction in movement could be dose-
dependently reversed when the larvae were treated with a TPH inhibitor.
Furthermore, the increase in serotonin level measured by HPLC was associated
with expression of ectopic serotonin-immunoreactive neurons in the ventral
hypothalamus. Treatment with a selective serotonin reuptake inhibitor,
fluvoxamine, restored the neuron phenotype by abolished ectopic serotonin
immunoreactivity. Taken together, uptake of serotonin in ectopic neurons of the
ventral hypothalamus induced a behavioral alteration that could be restored by
normalized serotonin levels.
Based on tract tracing and IHC data, we found afferent and efferent projections
that provided connections between the dorsal telencephalon and the ventrocaudal
hypothalamus. This telencephalo-hypothalamic loop may be crucial for execution
of motivated behaviors. Recent studies in zebrafish have identified specific brain
areas that are involved in mediating certain behaviors. For example, filtering of
visual information in the optic tectum by GABAergic interneurons is important in
prey capture behavior (Del Bene et al. 2010). Interestingly, hrh1 (Yanai et al. 1998)
and hdc knockout (Dere et al. 2004) in mice cause increases in serotonin turnover.
These rodent data interlink the role of reduced histamine via hrh1 signaling with
an increase in the serotonin metabolite 5-HIAA. The increase in 5-HIAA
concentration was only observed in the brainstem of hrh1 knockout mice, whereas
the turnover rate of 5-HIAA/serotonin was increased in the telencephalon,
hippocampus, mesencephalon, and brainstem (Yanai et al. 1998). In hdc knockout
mice, serotonin turnover was increased in the frontal cortex, a brain area linked
with attention and alertness (Dere et al. 2004). Collectively, the results from our
study provide novel data regarding two neuronal circuits by which specific
aminergic neurotransmitter systems affect neuronal plasticity and specific
behaviors in the zebrafish.
6.3 The histaminergic system in zebrafish compared with that in
mammals (I–III)
Histaminergic neurons are important in mediating various behavioral responses.
Therefore, it was important to study which neurotransmitters colocalize with
histamine and how similar the neurons of zebrafish are in comparison with
mammalian histaminergic neurons (data summarized in Table 3). First, we
quantified the histaminergic neuron number in both female and male adult
zebrafish brain and observed no differences between the sexes. This allowed us to
use both males and females for the cotransmitter analysis without taking into

Discussion
60
account the gender difference. We found that the cotransmitters of histamine in
histaminergic neurons are galanin, GABA, and TRH. A previous study of
histaminergic neurons revealed that there are discrepancies between different
rodent species (Airaksinen et al. 1992). In rats, the cotransmitters of histaminergic
neurons are galanin, GABA, and TRH (Airaksinen et al. 1992). In mice the
neurotransmitters that coexist with histamine are galanin and GABA, whereas in
guinea pig histamine colocalizes with GABA and mENK (Airaksinen et al. 1992).
Only one study has addressed whether galanin and GABA are cotransmitters of
tuberomammillary histaminergic neurons in humans and it showed that GABA
coexists with histamine (Trottier et al. 2002). In this study on postmortem samples
from both males and females, no galanin was found in histaminergic neurons
(Trottier et al. 2002). Fibers of each of the neurotransmitter systems studied
innervated the histaminergic neurons and may affect the activity of the histamine
neurons. Previous studies in rodents have shown that galanin has an inhibitory
effect on histamine release (Arrang et al. 1991) and that sleep deprivation
increases galanin levels (Toppila et al. 1995). Furthermore, TRH excites both
histaminergic (Parmentier et al. 2009) and hcrt neurons (Gonzalez et al. 2009).
Interestingly, the stimulating effect of TRH is abolished in hdc knockout animals
(Parmentier et al. 2009), which suggests that the wake-promoting effect of TRH is
histamine-mediated. The role of mENK in the histaminergic system has not been
studied. Galanin and GABA may form the main components of the sleep-
promoting system (McGinty & Szymusiak 2003). Neurons of these two systems are
located in the ventrolateral preoptic nucleus. These innervate and exert an
inhibitory effect on the activity of histaminergic neurons (Sherin et al. 1998). It has
long been known that via this projection the GABAergic neurons regulate the
sleep-wake cycle (McGinty & Szymusiak 2003). We also assessed the anatomical
relationship between histamine, serotonin, and dopamine in whole-mount adult
zebrafish brains. In this case, we found that the serotonin and dopamine neurons
are found in the same area, which is devoid of histaminergic neurons.
Furthermore, we could detect no neurons that were positive for both histamine
and TH or histamine and serotonin. This is in accordance with what was reported
from studies on sections of the adult zebrafish brain (Kaslin & Panula 2001) and rat
(Vanhala, Yamatodani & Panula 1994).

Discussion
61
Table 3. Comparison of the cotransmitters in histaminergic neurons in several species.
References to original publications are found in the text for human, rat, guinea pig, and
mouse. Zebrafish data presented as reported in II.
Cotransmitters of histaminergic neurons
Human Rat Guinea pig Mouse Zebrafish
hcrt ? - ? ? -
Galanin + + - + +
GABA - + + + +
mENK ? - + - -
TRH ? + - - +
TH ? - ? ? -
5-HT ? - ? ? -
The histamine receptors were expressed in the zebrafish embryo from 3 hpf
onwards and the histamine receptor subtypes were found throughout the brain.
As in mammals (Haas, Sergeeva & Selbach 2008), zebrafish hrh1 and hrh3 were
mainly expressed in the dorsal telencephalon. Faint expression of hrh1 was
detected in the lateral and ventrocaudal hypothalamus and hrh3 was expressed in
histaminergic ventrocaudal hypothalamus as well. The histamine receptors in
zebrafish larvae seem to be functional, since ligands targeting the receptors had
behavioral consequences in zebrafish. The behavioral consequences observed in
zebrafish were in accordance with those reported for mice (summary in Table 4).
In hrh1 knockout mice, spontaneous locomotor and exploratory behaviors are
reduced during the active period (Inoue et al. 1996) and in stressful situations
(Yanai et al. 1998). The motor coordination of hrh1 knockout animals was reported
to be normal (Inoue et al. 1996, Yanai et al. 1998), suggesting that hrh1 may
mediate the locomotor activities associated with emotion. In line with these
observations, we observed a reduction in spontaneous locomotor activity after
treatment of zebrafish larvae with pyrilamine, suggesting that the functionality of
hrh1 was conserved between the species. hrh2 knockout mice develop normally
and are fertile (Kobayashi et al. 2000). The CNS phenotype of hrh2 knockout mice
results in reduced spontaneous movement, increased restlessness in familiar
environments, since the knockouts exhibit increased rearing behavior, and lower
susceptibility for electrically induced convulsions compared with that in WT mice
(Fukushima et al. 2003). Similarly, in our study on zebrafish larvae, the hrh2
antagonist cimetidine reduced spontaneous locomotor activity and movement at

Discussion
62
low doses. Mice devoid of hrh3 also develop normally, are fertile, and display
overall reduced locomotor activity and normal circadian rhythm (Toyota et al.
2002).
Table 4. Comparison of the rodent and zebrafish histaminergic systems. References to
original publications regarding the mouse are found in the text. Zebrafish data presented
as reported in I and III.
Inhibition either via genetic (mouse) or pharmacological (zebrafish) intervention
Mouse Zebrafish
hdc Fails to respond in situations that require
increased alertness
Fails to respond in situations that
require increased alertness
hrh1 Reduced spontaneous locomotion Reduced spontaneous movement,
impaired response to environmental
cues
hrh2 Reduced spontaneous movement, increased
restlessness in familiar environments
Reduced spontaneous movement
hrh3 Reduced spontaneous movement Reduced spontaneous movement
The activity of hrh3 affects the release of amines, neuropeptides, and other
substances such as GABA and acetylcholine (Haas, Sergeeva & Selbach 2008),
which is why the levels of amines and their metabolites were measured in hrh3
knockout animals (Toyota et al. 2002). Only the histamine level was reduced in the
cortex of hrh3 knockout mice (Toyota et al. 2002). Thioperamide, an unselective
antagonist of hrh3, has been used in pharmacological studies in rats to increase
wakefulness (Monti et al. 1991) and to increase turnover of histamine in the cortex
of rats (Yates et al. 1999). Thioperamide treatment of zebrafish larvae decreased
spontaneous movement, a result in contrast to that reported for rats. Interestingly,
immepip which is a hrh3 agonist, reduced spontaneous locomotor activity. These
results indicate that hrh3 antagonism/agonism in zebrafish must be further
studied, e.g. with receptor binding and electrophysiology, to validate the exact
properties and specificity of the hrh3 receptor. Taken together, the functionality of
the histamine receptors in the zebrafish was generally conserved, since
pharmacological inhibition of two of the receptors produced similar behaviors, as
was previously described in mice.
So far, two studies have assessed the role of histaminergic ligands in larval
zebrafish behavior (I and III) and one study assessed the involvement of
histaminergic ligands in sleep-wake regulation in zebrafish larvae (Rihel et al.
2010). These findings are in line with each other and suggest that the zebrafish can

Discussion
63
be used as a model to study the role of histaminergic neurons under different
conditions.
6.4 The potential role of histamine in neurodegenerative disease (III, IV)
In this thesis study, deletion of psen1 led to altered development of the
histaminergic system that persisted until adulthood. These findings linked
abnormalities in histaminergic system development with AD associated gene
psen1. The main symptom of AD is dementia and cognitive decline, which are due
to progressive degeneration of neuronal tissue (Bishop, Lu & Yanker 2010).
Histamine receptors are involved in memory formation and hrh3 antagonists have
been implicated in improving cognitive functions (Haas & Panula 2003, Haas,
Sergeeva & Selbach 2008). We found that in psen1-/- larvae the histaminergic
neuron number was lower than in WT siblings. This reduced histaminergic neuron
number was not associated with increased cell death or alterations in other
neurotransmitter systems, such as dopamine or galanin. Notch signaling was
earlier reported to be involved in development of the dopaminergic system in
zebrafish (Mahler, Filippi & Driever 2010). We did not, however, observe any
changes in the dopaminergic neurotransmitter system between the psen1-/- fish
and WT fish. CNS defects and abnormal phenotype are characteristic of psen1
knockout mice, which die at birth (Shen et al. 1997). We observed no major
defects in the gross phenotype of psen1-/- larval fish compared with WT fish. The
role of psen1 has been studied by MO in zebrafish and these studies have shown
that transient inhibition of psen1 and psen2 affects Notch signaling to varying
degrees (Nornes et al. 2009). Notch is a crucial regulator of neural development in
many species (Pierfelice, Alberi & Gaiano 2011) and we found here that the
development of histaminergic neurons was regulated via γ-secretase activity,
which affects Notch signaling. The reduced number of histaminergic neurons in
psen1-/- was associated with a decrease in locomotor activity and lack of robust
response to environmental cues. We furthermore observed that the histamine
neuron number was higher in psen1-/- at 1 year of age than in WT fish. This
neuronal phenotype was also associated with increased thigmotaxis, suggesting
increased alertness or anxiety. Reduced anxiety is observed in adult zebrafish
when the histamine level in the brain is reduced with α-FMH (Peitsaro et al. 2003).
The mechanism underlying the increase in histaminergic neurons may be increased
proliferation. In line with this hypothesis, mutations in psen1 increase proliferation
in mouse models of AD (Veeraraghavalu et al. 2010) and histaminergic signaling
itself potentiates proliferation and neuronal fate in vitro (Molina-Hernandez &

Discussion
64
Velasco 2008). Taken together, we found that the histaminergic system was plastic
throughout life and that this plasticity was mediated by psen1 and Notch1a. Notch
signaling was recently shown to control regeneration of motor neurons in adult
zebrafish (Dias et al. 2012). Alternatively, the plasticity observed in the
histaminergic system in this AD model could be a consequence of
neurotransmitter respecification (Spitzer 2012).
Furthermore, we found that histamine had a bidirectional effect on the
development of hcrt neurons, which is the main neurotransmitter system affected
in narcolepsy. In our studies, hdc knockdown and hrh1 antagonism resulted in
decreased hcrt neuron number in the treated animals compared with WT siblings
at 5 dpf. The deficit was rescued with coinjection of hdc MO and mRNA, whereas
hdc mRNA alone led to an increase in the number of hcrt neurons. In August 2010,
a survey was initiated due to the increased incidents of narcolepsy in Finnish and
Swedish children and the link between Pandemrix, a H1N1 influenza vaccine, and
the most severe form of narcolepsy, i.e. narcolepsy associated with cataplexy, was
established in persons under 20 years of age (Dauvilliers et al. 2010, Kallweit et al.
2011, Kornum, Faraco & Mignot 2011). Narcolepsy is thought to be an
autoimmune disease (Scammell 2003, De la Herran-Arita, Guerra-Crespo &
Drucker-Colin 2011). In the cerebrospinal fluid of narcoleptic patients, the
histamine level is reduced (Nishino et al. 2009) and in hrh1 knockout animals, the
hcrt levels are also reduced (Lin et al. 2002). Previously, it was shown that hcrt
excites histaminergic neurons (Eriksson et al. 2001), whereas the opposite has not.
Histamine and hcrt are responsible for different aspects of alertness, histamine for
motivated alertness and hcrt for locomotion-mediated alertness (Anaclet et al.
2009). Histamine is either neuroprotective (Kukko-Lukjanov et al. 2006) or
neurotoxic (Vizuete et al. 2000). The neurotoxic effect of histamine has been
documented in the dopaminergic neurons in the substantia nigra (Vizuete et al.
2000). Alternatively, histamine could exert a neurotrophic effect, since we found
that lack of histamine interferes with the development of the hcrt system. These
results may have implications for the use of antihistamines during pregnancy.
Since antihistamines are widely used in the treatment of allergy and pass through
the placenta easily from the mother to the fetus, the effect of antihistamine
medication during pregnancy on the developing child should be studied. An
intriguing finding linked AD pathology to hcrt a few years ago, when changes in the
β-amyloid concentration in brain interstitial fluid was correlated with wakefulness
in mice and infusions with hcrt increased the amount (Kang et al. 2009). Here, we
showed that histamine affects the hcrt system that underlies narcolepsy.

Discussion
65
Furthermore, we showed a spatiotemporal alteration in the histaminergic system,
which was due to impaired psen1 function, a gene associated with AD. Therefore,
combining knowledge of the interaction between the histaminergic and hcrt
systems and the effect that hcrt has on the formation of β-amyloids in a circadian
manner, could provide further information on the mechanisms underlying the
progression of neurodegenerative diseases.
Taken together, we identified for the first time a factor that affects the developing
histaminergic neurotransmitter system. These results suggest that histaminegic
neuron development is regulated by γ-secretase, and that histamine itself drives
the development of the hcrt system. These results offer a new avenue for study of
the etiology of AD and narcolepsy, respectively.

Conclusions
66
7 Conclusions
In this thesis the role of the histaminergic neurotransmitter system in neuronal
plasticity was examined (Figure 16).
• We found that the histaminergic system resembles that of other
vertebrates, since the similarity between zebrafish and rodent histamine
receptors is reasonably high. Moreover, histaminergic ligands have
behavioral effects on larval zebrafish that mimic previously documented
behaviors that have been observed in mammals.
• The phenotype of the histaminergic neurons in zebrafish resembles that of
mammals, and the zebrafish histaminergic neurons have the same
cotransmitters as the histaminergic neurons in rats.
• Behavioral experiments revealed that histamine mediated the response of
animals to environmental cues by altered alertness. We proposed that this
rapid response, the dark-induced flash response, was partially controlled
by the dorsal telencephalon. The dorsal telencephalon of zebrafish was
innervated by a dense network of histaminergic fiber projections and
expressed hrh1 and hrh3. This brain area sent afferent projections that
terminated at the histaminergic neurons. Together these formed a
telencephalic-hypothalamic loop. The hrh1 antagonist pyrilamine
abolished the dark-induced flash response.
• We found that histamine deficiency interfered negatively with the
development of the hcrt system in a hrh1-dependent manner. Another
novel finding was the identification of γ-secretase and possibly Notch
signaling as factors that regulated the development of histaminergic
neurons.
• We found that MAO, also expressed in histaminergic neurons, mainly
affects the metabolism of serotonin. We identified increased serotonin
levels and ectopic serotonin immunoreactivity in the ventral hypothalamus
as mediators of decreased locomotor activity in larval zebrafish.

Conclusions
67
In conclusion, the main findings in this thesis are the identification of new
regulatory mechanisms for histamine. Firstly, we found that histamine affects the
development of the hcrt system. Secondly, we found that the developing
histaminergic system itself is regulated in a γ-secretase-dependent manner. These
results are raising new important aspects of study in the etiology of
neurodegenerative disorders, such as narcolepsy and AD.
Figure 16. Summary of the main results presented in this thesis. Histaminergic neurons
containing histamine (HA) and several cotransmitters project from the ventrocaudal
hypothalamus to the dorsal telencephalon. In this area, hrh1 and hrh3 are highly
expressed. The neurons that project from the target area of histaminergic neurons are
mainly glutamatergic and innervate the ventrocaudal hypothalamus. In this region,
serotonergic neurons are also found and ectopic expression of serotonin as a result of
inhibition of MAO decreases locomotor activity. Inhibition of γ-secretase downregulates
the histaminergic neuron number in larval zebrafish, whereas inhibition of histamine
synthesis downregulates the number of hcrt-positive neurons in the lateral hypothalamus
in a hrh1-mediated manner and impairs the behavioral responses to environmental cues.

Future prospects
68
8 Future prospects
Tract tracing revealed a hypothalamic–telencephalic loop in zebrafish similar to
that detected in rodents (Ericson, Blomqvist & Kohler 1991). Recently, a study
reported that the Dc in zebrafish could be the mammalian cortex equivalent
(Mueller et al. 2011). Further studies in comparative neurology and hodology that
can be correlated with behavioral outputs are needed for a better understanding
of neuronal functions in the zebrafish brain. Although the MO approach was
successful in this study, knockout of hdc by TILLING, Gal4/UAS, ZFN, or TALEN
would provide favorable additional models for assessing the role of hdc and the
histaminergic system in development and disease. Furthermore, the role of MAO
in histaminergic neurons could be studied with Gal4/UAS:TeTx transgenic
zebrafish.
We showed that the dark-flash response was mediated by hrh1 and therefore hrh1
knockouts could in future experiments be studied to verify the results of the
pharmacological experiments. Interestingly, increased serotonin turnover was
detected in the brainstem of hdc (Dere et al. 2004) and hrh1 knockout mice (Yanai
et al. 1998). When extrapolated to zebrafish, this could provide a link between the
two neuronal circuits that we identified. Therefore, the serotonin turnover could
be assessed in animals treated with the hrh1 antagonist pyrilamine. Since hrh1
antagonism affected the hcrt neurons, this system should be studied in both hdc
and hrh1 knockouts. The fact that histamine affects the development of the hcrt
neurons allows studies of the role of histamine in development and etiology of
narcolepsy. As pointed out earlier, a mutation in hdc was associated with
Tourette’s syndrome (Ercan-Sencicek et al. 2010) and we found that lack of hdc
abolished the dark-induced flash response in zebrafish larvae. Taken together,
these results, the responsiveness to changes in the environment in zebrafish and
fine-tuning of the motor responses, appear to be mediated at least partially via the
histaminergic neurotransmitter system. Furthermore, these results indicate that
histamine could be needed for fine-tuning of motor functions, since Tourette’s
syndrome patients suffer from involuntary uncontrolled small muscle contractions,
i.e. tics. Therefore, it would be important to study the role of the histaminergic
neurotransmitter system in prepulse inhibition and the startle response in
zebrafish larvae. Since the startle response is driven by Mauthner neurons, the
innervation of histaminergic neuron projections of the Mauthner neurons needs to
be determined before the behavioral studies are conducted.

Future prospects
69
Since psen1 knockout affects the developing histaminergic system, the effect of
this mutation could also disturb the hcrt system. A consensus of increased
incidence of narcolepsy in AD patients is not available, since the two studies
conducted reported contrasting results (Fronczek et al. 2011, Scammell et al.
2011). The effect that psen1 knockout has on cell proliferation in rodents has not
been reported for zebrafish. Therefore, the effect of histaminergic ligands on
behavior and proliferation in psen1-/- and WT zebrafish should be determined.
Finally, we reported that inhibition of γ-secretase with DAPT reduced the
histamine neuron number. In zebrafish, DAPT mainly affects the ability of γ-
secretase to cleave Notch (Geling et al. 2002), and studying the histaminergic
neurotransmitter system in Notch mutants would give a definite answer on
whether or not the impaired development of histaminergic neurons is Notch-
mediated.
Taken together, the results acquired from the pharmacological and transient
knockdown approaches should be verified in stable knockout zebrafish models.
The novel mechanistic findings should further be validated in other model
organisms and humans, to eventually be able to draw conclusions applicable to
humans, based on these specific findings.

Thank you
70
Acknowledgements
This work has been carried out at the Neuroscience Center and Institute of
Biomedicine, Anatomy at the Faculty of Medicine, University of Helsinki. I started
my PhD project in late 2007 and had a break for five months during 2008 to obtain
a teacher’s degree at the Faculty of Pedagogy, Abo Akademi University. My study
was supported by the Academy of Finland, the Finnish Technology Development
Fund, the Sigrid Juselius Foundation, the Helsinki Biomedical Graduate Program,
the University of Helsinki Chacellor’s Funds, the University of Helsinki Funds, the
Magnus Ehrnrooth Foundation, the Swedish Cultural Foundation in Finland, the
European Histamine Research Society, the International Federation of Societies for
Histochemistry and Cytochemistry, and the International Federation of Cell
Biology.
I had the great opportunity and pleasure to be supervised by Professor Pertti
Panula. I am very grateful for your never ending support and encouragement, not
only regarding pursuing a PhD, but also for encouraging me to go through with my
studies to obtain a biology and geography teacher’s degree. Your genuine interest
and passion for science always brought my faith back into my long lasting project.
I would like to thank my thesis committee members Professor Esa Korpi and
Docent Tarja Stenberg for their supportive and constructive comments on the
progress of my thesis. I would also like to thank reviewers Assistant Professors
Sanna Lehtonen and Niclas Kolm for their excellent comments on my thesis, and
opponent Dr. Catherina Becker for accepting the invitation to travel all the way to
Helsinki to discuss my research.
A sincere thank you to all of my coauthors, Dr. Oleg Anichtchik, Dr. Yu-Chia Chen,
Dr. Jan Kaslin, Dr. Denis Khrustalyov, Dr. Hisaaki Kudo, Dr. Nina Peitsaro, Professor
Pertti Panula, Dr. Ilkka Reenilä, Stanislav Rozov, Dr. Ville Sallinen, Pauliina Toivonen
and Gabija Tolekyte. It has been nice to work together with you on our quest to
unravel “the mysteries” within neuroscience!
I would like to thank all the current and past members of the pp-group both at Abo
Akademi University (Abbi, Adrian, Bodil, Dominique, Jan, Jari K, Jin, Kaj, Kimmo,
Levente, Maria Ö, Minna-Maija, Minni, Nina, Oleg, Pirjo, Tina, Tiina-Kaisa,
Veronica) and University of Helsinki (Anna, Anu, Dennis, Gabija, Henri K, Henri P,
Hiroshi, Hisaaki, Ilmari, Jari R, Jenni, Jenny, Juha, Luisa, Maxim, Madhusmita, Maria

Thank you
71
V, Michalis, Michele, Olga, Olli, Oskar, Paula, Pauliina, Piotr, Reeta, Saara, Stanislav,
Susanna, Svetlana, Tiia, Thomas, Tuomas, Veera, Ville, Yarosalv, Yu-Chia, Yumiko)
that I had the pleasure to interact with, for introducing me to the wonderful world
of neuroscience. Thank you for the time spent inside and outside the lab. I really
enjoyed working with and getting to know all of you! I would especially like to
mention Dr. Saara Nuutinen and Dr. Jari Rossi who kindly commented on the first
versions of my thesis. Mikko Liljeström and Dr. Mika Hukkanen at the Biomedicum
Imaging Unit are thanked for their never ending patience regarding questions
about confocal microscopes and image processing. I furthermore would like to
thank all the scientists who visited the pp-group in Helsinki, especially Dr. Mario de
Bono, Dr. Harry Burgess, Professor Robert Cornell, Professor Helmut Haas, Dr.
Marnie Halpern, Dr. Petronella Kettunen, Dr. Donald O’Malley, Professor Hiroshi
Ohtsu, Professor Michalis Pavlidis, Dr. Gonzalo de Polavieja, Professor Manuel
Portavella, and Dr. Maximiliano Suster for the discussions we had regarding
science and zebrafish. Friends in and outside science – you all contributed to this
thesis. I am very grateful for all the fun together, and for having you in my life!
My deepest gratitude goes to my loved ones, to my family including all possible
extensions. Your joyousness, and endless support and faith in me keep on amazing
me. You will always be in my heart.
Maria Sundvik
Helsingfors, 2012

72
References
Ables, A.Z. & Nagubilli, R. 2010, "Prevention, recognition, and management of serotonin syndrome", American Family Physician, vol. 81, no. 9, pp. 1139-1142.
Ables, J.L., Breunig, J.J., Eisch, A.J. & Rakic, P. 2011, "Not(ch) just development: Notch signalling in the adult brain", Nature Reviews Neuroscience, vol. 12, no. 5, pp. 269-283.
Agnati, L.F., Zoli, M., Stromberg, I. & Fuxe, K. 1995, "Intercellular communication in the brain: wiring versus volume transmission", Neuroscience, vol. 69, no. 3, pp. 711-726.
Airaksinen, M.S., Alanen, S., Szabat, E., Visser, T.J. & Panula, P. 1992, "Multiple neurotransmitters in the tuberomammillary nucleus: comparison of rat, mouse, and guinea pig", The Journal of comparative neurology, vol. 323, no. 1, pp. 103-116.
Amo, R., Aizawa, H., Takahoko, M., Kobayashi, M., Takahashi, R., Aoki, T. & Okamoto, H. 2010, "Identification of the zebrafish ventral habenula as a homolog of the mammalian lateral habenula", The Journal of Neuroscience, vol. 30, no. 4, pp. 1566-1574.
Anaclet, C., Parmentier, R., Ouk, K., Guidon, G., Buda, C., Sastre, J.P., Akaoka, H., Sergeeva, O.A., Yanagisawa, M., Ohtsu, H., Franco, P., Haas, H.L. & Lin, J.S. 2009, "Orexin/hypocretin and histamine: distinct roles in the control of wakefulness demonstrated using knock-out mouse models", The Journal of Neuroscience, vol. 29, no. 46, pp. 14423-14438.
Anichtchik, O., Sallinen, V., Peitsaro, N. & Panula, P. 2006, "Distinct structure and activity of monoamine oxidase in the brain of zebrafish (Danio rerio)", The Journal of comparative neurology, vol. 498, no. 5, pp. 593-610.
Anic-Labat, S., Guilleminault, C., Kraemer, H.C., Meehan, J., Arrigoni, J. & Mignot, E. 1999, "Validation of a cataplexy questionnaire in 983 sleep-disorders patients", Sleep, vol. 22, no. 1, pp. 77-87.
Arrang, J.M., Garbarg, M. & Schwartz, J.C. 1983, "Auto-inhibition of brain histamine release mediated by a novel class (H3) of histamine receptor", Nature, vol. 302, no. 5911, pp. 832-837.
Arrang, J.M., Gulat-Marnay, C., Defontaine, N. & Schwartz, J.C. 1991, "Regulation of histamine release in rat hypothalamus and hippocampus by presynaptic galanin receptors", Peptides, vol. 12, no. 5, pp. 1113-1117.
Arrang, J.M., Morisset, S. & Gbahou, F. 2007, "Constitutive activity of the histamine H3 receptor", Trends in pharmacological sciences, vol. 28, no. 7, pp. 350-357.
Asakawa, K., Suster, M.L., Mizusawa, K., Nagayoshi, S., Kotani, T., Urasaki, A., Kishimoto, Y., Hibi, M. & Kawakami, K. 2008, "Genetic dissection of neural circuits by Tol2 transposon-mediated Gal4 gene and enhancer trapping in zebrafish", Proceedings of the National Academy of Sciences of the United States of America, vol. 105, no. 4, pp. 1255-1260.
Asakawa, K. & Kawakami, K. 2009, "The Tol2-mediated Gal4-UAS method for gene and enhancer trapping in zebrafish", Methods (San Diego, Calif.), vol. 49, no. 3, pp. 275-281.
Auvinen, S. & Panula, P. 1988, "Development of histamine-immunoreactive neurons in the rat brain", The Journal of comparative neurology, vol. 276, no. 2, pp. 289-303.
Axelrod, J. 1962, "Purification and properties of phenylethanolamine-N-methyl transferase", The Journal of biological chemistry, vol. 237, pp. 1657-1660.
Bae, Y.K., Kani, S., Shimizu, T., Tanabe, K., Nojima, H., Kimura, Y., Higashijima, S. & Hibi, M. 2009, "Anatomy of zebrafish cerebellum and screen for mutations affecting its development", Developmental biology, vol. 330, no. 2, pp. 406-426.
Bayer, L., Eggermann, E., Serafin, M., Saint-Mleux, B., Machard, D., Jones, B. & Muhlethaler, M. 2001, "Orexins (hypocretins) directly excite tuberomammillary neurons", The European journal of neuroscience, vol. 14, no. 9, pp. 1571-1575.
Berlucchi, G. & Buchtel, H.A. 2009, "Neuronal plasticity: historical roots and evolution of meaning", Experimental brain research, vol. 192, no. 3, pp. 307-319.
Bibikova, M., Beumer, K., Trautman, J.K. & Carroll, D. 2003, "Enhancing gene targeting with designed zinc finger nucleases", Science (New York, N.Y.), vol. 300, no. 5620, pp. 764.
Bishop, N.A., Lu, T. & Yankner, B.A. 2010, "Neural mechanisms of ageing and cognitive decline", Nature, vol. 464, no. 7288, pp. 529-535.

73
Blandina, P., Giorgetti, M., Bartolini, L., Cecchi, M., Timmerman, H., Leurs, R., Pepeu, G. & Giovannini, M.G. 1996, "Inhibition of cortical acetylcholine release and cognitive performance by histamine H3 receptor activation in rats", British journal of pharmacology, vol. 119, no. 8, pp. 1656-1664.
Blennow, K., de Leon, M.J. & Zetterberg, H. 2006, "Alzheimer's disease", Lancet, vol. 368, no. 9533, pp. 387-403.
Boonen, R.A., van Tijn, P. & Zivkovic, D. 2009, "Wnt signaling in Alzheimer's disease: up or down, that is the question", Ageing research reviews, vol. 8, no. 2, pp. 71-82.
Borchelt, D.R., Thinakaran, G., Eckman, C.B., Lee, M.K., Davenport, F., Ratovitsky, T., Prada, C.M., Kim, G., Seekins, S., Yager, D., Slunt, H.H., Wang, R., Seeger, M., Levey, A.I., Gandy, S.E., Copeland, N.G., Jenkins, N.A., Price, D.L., Younkin, S.G. & Sisodia, S.S. 1996, "Familial Alzheimer's disease-linked presenilin 1 variants elevate Abeta1-42/1-40 ratio in vitro and in vivo ", Neuron, vol. 17, no. 5, pp. 1005-1013.
Bretaud, S., Macraild, S., Ingham, P.W. & Bandmann, O. 2011, "The Influence of the Zebrafish Genetic Background on Parkinson's Disease-Related Aspects", Zebrafish, vol. 3, pp. 103-108.
Brockerhoff, S.E., Hurley, J.B., Janssen-Bienhold, U., Neuhauss, S.C., Driever, W. & Dowling, J.E. 1995, "A behavioral screen for isolating zebrafish mutants with visual system defects", Proceedings of the National Academy of Sciences of the United States of America, vol. 92, no. 23, pp. 10545-10549.
Brown, D.D., Tomchick, R. & Axelrod, J. 1959, "The distribution and properties of a histamine-methylating enzyme", The Journal of biological chemistry, vol. 234, pp. 2948-2950.
Brustein, E., Saint-Amant, L., Buss, R.R., Chong, M., McDearmid, J.R. & Drapeau, P. 2003, "Steps during the development of the zebrafish locomotor network", Journal of physiology, vol. 97, no. 1, pp. 77-86.
Budick, S.A. & O'Malley, D.M. 2000, "Locomotor repertoire of the larval zebrafish: swimming, turning and prey capture", The Journal of experimental biology, vol. 203, no. Pt 17, pp. 2565-2579.
Burgess, H.A. & Granato, M. 2007a, "Modulation of locomotor activity in larval zebrafish during light adaptation", The Journal of experimental biology, vol. 210, no. Pt 14, pp. 2526-2539.
Burgess, H.A. & Granato, M. 2007b, "Sensorimotor gating in larval zebrafish", The Journal of Neuroscience, vol. 27, no. 18, pp. 4984-4994.
Burgess, H.A., Schoch, H. & Granato, M. 2010, "Distinct retinal pathways drive spatial orientation behaviors in zebrafish navigation", Current biology, vol. 20, no. 4, pp. 381-386.
Caroll, D. & Zhang, B. 2011, "Primer and interviews: advances in targeted gene modification. Interview by Julie C. Kiefer", Developmental dynamics : an official publication of the American Association of Anatomists, vol. 240, no. 12, pp. 2688-2696.
Chen, Y.C., Priyadarshini, M. & Panula, P. 2009, "Complementary developmental expression of the two tyrosine hydroxylase transcripts in zebrafish", Histochemistry and cell biology, vol. 132, no. 4, pp. 375-381.
Chevallier, N.L., Soriano, S., Kang, D.E., Masliah, E., Hu, G. & Koo, E.H. 2005, "Perturbed neurogenesis in the adult hippocampus associated with presenilin-1 A246E mutation", The American journal of pathology, vol. 167, no. 1, pp. 151-159.
Cirelli, C. 2009, "The genetic and molecular regulation of sleep: from fruit flies to humans", Nature Reviews Neuroscience, vol. 10, no. 8, pp. 549-560.
Claassen, V., Davies, J.E., Hertting, G. & Placheta, P. 1977, "Fluvoxamine, a specific 5-hydroxytryptamine uptake inhibitor", British journal of pharmacology, vol. 60, no. 4, pp. 505-516.
Colbert, T., Till, B.J., Tompa, R., Reynolds, S., Steine, M.N., Yeung, A.T., McCallum, C.M., Comai, L. & Henikoff, S. 2001, "High-throughput screening for induced point mutations", Plant Physiology, vol. 126, no. 2, pp. 480-484.
Cragg, S.J., Nicholson, C., Kume-Kick, J., Tao, L. & Rice, M.E. 2001, "Dopamine-mediated volume transmission in midbrain is regulated by distinct extracellular geometry and uptake", Journal of neurophysiology, vol. 85, no. 4, pp. 1761-1771.
Dai, H., Kaneko, K., Kato, H., Fujii, S., Jing, Y., Xu, A., Sakurai, E., Kato, M., Okamura, N., Kuramasu, A. & Yanai, K. 2007, "Selective cognitive dysfunction in mice lacking histamine H1 and H2 receptors", Neuroscience research, vol. 57, no. 2, pp. 306-313.

74
Dale, H.H. & Laidlaw, P.P. 1910, "The physiological action of beta-iminazolylethylamine", The Journal of Physiology Online, vol. 41, no. 5, pp. 318-344.
Dauvilliers, Y. 2007, "Insomnia in patients with neurodegenerative conditions", Sleep medicine, vol. 8 Suppl 4, pp. S27-34.
Dauvilliers, Y., Montplaisir, J., Cochen, V., Desautels, A., Einen, M., Lin, L., Kawashima, M., Bayard, S., Monaca, C., Tiberge, M., Filipini, D., Tripathy, A., Nguyen, B.H., Kotagal, S. & Mignot, E. 2010, "Post-H1N1 narcolepsy-cataplexy", Sleep, vol. 33, no. 11, pp. 1428-1430.
De la Herran-Arita, A.K., Guerra-Crespo, M. & Drucker-Colin, R. 2011, "Narcolepsy and orexins: an example of progress in sleep research", Frontiers in neurology, vol. 2, pp. 26.
de Lecea, L., Kilduff, T.S., Peyron, C., Gao, X., Foye, P.E., Danielson, P.E., Fukuhara, C., Battenberg, E.L., Gautvik, V.T., Bartlett, F.S.,2nd, Frankel, W.N., van den Pol, A.N., Bloom, F.E., Gautvik, K.M. & Sutcliffe, J.G. 1998, "The hypocretins: hypothalamus-specific peptides with neuroexcitatory activity", Proceedings of the National Academy of Sciences of the United States of America, vol. 95, no. 1, pp. 322-327.
De Strooper, B. & Annaert, W. 2010, "Novel research horizons for presenilins and gamma-secretases in cell biology and disease", Annual Review of Cell and Developmental Biology, vol. 26, pp. 235-260.
Del Bene, F., Wyart, C., Robles, E., Tran, A., Looger, L., Scott, E.K., Isacoff, E.Y. & Baier, H. 2010, "Filtering of visual information in the tectum by an identified neural circuit", Science, vol. 330, no. 6004, pp. 669-673.
Dere, E., De Souza-Silva, M.A., Spieler, R.E., Lin, J.S., Ohtsu, H., Haas, H.L. & Huston, J.P. 2004, "Changes in motoric, exploratory and emotional behaviours and neuronal acetylcholine content and 5-HT turnover in histidine decarboxylase-KO mice", The European journal of neuroscience, vol. 20, no. 4, pp. 1051-1058.
Dias, T.B., Yang, Y.J., Ogai, K., Becker, T. & Becker, C.G. 2012, "Notch signaling controls generation of motor neurons in the lesioned spinal cord of adult zebrafish", The Journal of Neuroscience, vol. 32, no. 9, pp. 3245-3252.
Douglass, A.D., Kraves, S., Deisseroth, K., Schier, A.F. & Engert, F. 2008, "Escape behavior elicited by single, channelrhodopsin-2-evoked spikes in zebrafish somatosensory neurons", Current biology, vol. 18, no. 15, pp. 1133-1137.
Doyon, Y., McCammon, J.M., Miller, J.C., Faraji, F., Ngo, C., Katibah, G.E., Amora, R., Hocking, T.D., Zhang, L., Rebar, E.J., Gregory, P.D., Urnov, F.D. & Amacher, S.L. 2008, "Heritable targeted gene disruption in zebrafish using designed zinc-finger nucleases", Nature biotechnology, vol. 26, no. 6, pp. 702-708.
Driever, W., Solnica-Krezel, L., Schier, A.F., Neuhauss, S.C., Malicki, J., Stemple, D.L., Stainier, D.Y., Zwartkruis, F., Abdelilah, S., Rangini, Z., Belak, J. & Boggs, C. 1996, "A genetic screen for mutations affecting embryogenesis in zebrafish", Development, vol. 123, pp. 37-46.
Eisen, J.S. & Smith, J.C. 2008, "Controlling morpholino experiments: don't stop making antisense", Development (Cambridge, England), vol. 135, no. 10, pp. 1735-1743.
Emran, F., Rihel, J. & Dowling, J.E. 2008, "A behavioral assay to measure responsiveness of zebrafish to changes in light intensities", Journal of visualized experiments, vol. 20, pii. 923, doi. 10.3791/923.
Ercan-Sencicek, A.G., Stillman, A.A., Ghosh, A.K., Bilguvar, K., O'Roak, B.J., Mason, C.E., Abbott, T., Gupta, A., King, R.A., Pauls, D.L., Tischfield, J.A., Heiman, G.A., Singer, H.S., Gilbert, D.L., Hoekstra, P.J., Morgan, T.M., Loring, E., Yasuno, K., Fernandez, T., Sanders, S., Louvi, A., Cho, J.H., Mane, S., Colangelo, C.M., Biederer, T., Lifton, R.P., Gunel, M. & State, M.W. 2010, "L-histidine decarboxylase and Tourette's syndrome", The New England journal of medicine, vol. 362, no. 20, pp. 1901-1908.
Ericson, H., Blomqvist, A. & Kohler, C. 1991, "Origin of neuronal inputs to the region of the tuberomammillary nucleus of the rat brain", The Journal of comparative neurology, vol. 311, no. 1, pp. 45-64.
Eriksson, K.S., Peitsaro, N., Karlstedt, K., Kaslin, J. & Panula, P. 1998, "Development of the histaminergic neurons and expression of histidine decarboxylase mRNA in the zebrafish brain in the absence of all peripheral histaminergic systems", The European journal of neuroscience, vol. 10, no. 12, pp. 3799-3812.

75
Eriksson, K.S., Sergeeva, O., Brown, R.E. & Haas, H.L. 2001, "Orexin/hypocretin excites the histaminergic neurons of the tuberomammillary nucleus", The Journal of Neuroscience, vol. 21, no. 23, pp. 9273-9279.
Eriksson, K.S., Sergeeva, O.A., Haas, H.L. & Selbach, O. 2010, "Orexins/hypocretins and aminergic systems", Acta physiologica, vol. 198, no. 3, pp. 263-275.
Fernandez, T.V., Sanders, S.J., Yurkiewicz, I.R., Ercan-Sencicek, A.G., Kim, Y.S., Fishman, D.O., Raubeson, M.J., Song, Y., Yasuno, K., Ho, W.S., Bilguvar, K., Glessner, J., Chu, S.H., Leckman, J.F., King, R.A., Gilbert, D.L., Heiman, G.A., Tischfield, J.A., Hoekstra, P.J., Devlin, B., Hakonarson, H., Mane, S.M., Gunel, M. & State, M.W. 2011, "Rare Copy Number Variants in Tourette Syndrome Disrupt Genes in Histaminergic Pathways and Overlap with Autism", Biological psychiatry, vol. 71, no. 5, pp. 392-402.
Filby, A.L., Paull, G.C., Hickmore, T.F. & Tyler, C.R. 2010, "Unravelling the neurophysiological basis of aggression in a fish model", BMC genomics, vol. 11, pp. 498-515.
Fink, M., Flekna, G., Ludwig, A., Heimbucher, T. & Czerny, T. 2006, "Improved translation efficiency of injected mRNA during early embryonic development", Developmental dynamics, vol. 235, no. 12, pp. 3370-3378.
Fowler, C.J., Oreland, L. & Callingham, B.A. 1981, "The acetylenic monoamine oxidase inhibitors clorgyline, deprenyl, pargyline and J-508: their properties and applications", The Journal of pharmacy and pharmacology, vol. 33, no. 6, pp. 341-347.
Fronczek, R., van Geest, S., Frolich, M., Overeem, S., Roelandse, F.W., Lammers, G.J. & Swaab, D.F. 2011, "Hypocretin (orexin) loss in Alzheimer's disease", Neurobiology of aging, In press.
Fukushima, Y., Shindo, T., Anai, M., Saitoh, T., Wang, Y., Fujishiro, M., Ohashi, Y., Ogihara, T., Inukai, K., Ono, H., Sakoda, H., Kurihara, Y., Honda, M., Shojima, N., Fukushima, H., Haraikawa-Onishi, Y., Katagiri, H., Shimizu, Y., Ichinose, M., Ishikawa, T., Omata, M., Nagai, R., Kurihara, H. & Asano, T. 2003, "Structural and functional characterization of gastric mucosa and central nervous system in histamine H2 receptor-null mice", European journal of pharmacology, vol. 468, no. 1, pp. 47-58.
Ganz, J., Kaslin, J., Freudenreich, D., Machate, A., Geffarth, M. & Brand, M. 2011, "Subdivisions of the adult zebrafish subpallium by molecular marker analysis", The Journal of comparative neurology, vol. 520, no. 3, pp. 633-655.
Garbarg, M., Barbin, G., Bischoff, S., Pollard, H. & Schwartz, J.C. 1974, "Evidence for a specific decarboxylase involved in histamine synthesis in an ascending pathway in rat brain", Agents and Actions, vol. 4, no. 3, pp. 181.
Garbarg, M., Barbin, G., Rodergas, E. & Schwartz, J.C. 1980, "Inhibition of histamine synthesis in brain by alpha-fluoromethylhistidine, a new irreversible inhibitor: in vitro and in vivo studies", Journal of neurochemistry, vol. 35, no. 5, pp. 1045-1052.
Gaus, S.E., Strecker, R.E., Tate, B.A., Parker, R.A. & Saper, C.B. 2002, "Ventrolateral preoptic nucleus contains sleep-active, galaninergic neurons in multiple mammalian species", Neuroscience, vol. 115, no. 1, pp. 285-294.
Geling, A., Steiner, H., Willem, M., Bally-Cuif, L. & Haass, C. 2002, "A gamma-secretase inhibitor blocks Notch signaling in vivo and causes a severe neurogenic phenotype in zebrafish", EMBO reports, vol. 3, no. 7, pp. 688-694.
Gonzalez, J.A., Horjales-Araujo, E., Fugger, L., Broberger, C. & Burdakov, D. 2009, "Stimulation of orexin/hypocretin neurones by thyrotropin-releasing hormone", The Journal of physiology, vol. 587, no. Pt 6, pp. 1179-1186.
Grillner, S. & Jessell, T.M. 2009, "Measured motion: searching for simplicity in spinal locomotor networks", Current opinion in neurobiology, vol. 19, no. 6, pp. 572-586.
Guo, Q., Furukawa, K., Sopher, B.L., Pham, D.G., Xie, J., Robinson, N., Martin, G.M. & Mattson, M.P. 1996, "Alzheimer's PS-1 mutation perturbs calcium homeostasis and sensitizes PC12 cells to death induced by amyloid beta-peptide ", Neuroreport, vol. 8, no. 1, pp. 379-383.
Haas, H. & Panula, P. 2003, "The role of histamine and the tuberomamillary nucleus in the nervous system", Nature Reviews Neuroscience, vol. 4, no. 2, pp. 121-130.
Haas, H.L., Sergeeva, O.A. & Selbach, O. 2008, "Histamine in the nervous system", Physiological Reviews, vol. 88, no. 3, pp. 1183-1241.
Hallmayer, J., Faraco, J., Lin, L., Hesselson, S., Winkelmann, J., Kawashima, M., Mayer, G., Plazzi, G., Nevsimalova, S., Bourgin, P., Hong, S.C., Honda, Y., Honda, M., Hogl, B., Longstreth,

76
W.T.,Jr, Montplaisir, J., Kemlink, D., Einen, M., Chen, J., Musone, S.L., Akana, M., Miyagawa, T., Duan, J., Desautels, A., Erhardt, C., Hesla, P.E., Poli, F., Frauscher, B., Jeong, J.H., Lee, S.P., Ton, T.G., Kvale, M., Kolesar, L., Dobrovolna, M., Nepom, G.T., Salomon, D., Wichmann, H.E., Rouleau, G.A., Gieger, C., Levinson, D.F., Gejman, P.V., Meitinger, T., Young, T., Peppard, P., Tokunaga, K., Kwok, P.Y., Risch, N. & Mignot, E. 2009, "Narcolepsy is strongly associated with the T-cell receptor alpha locus", Nature genetics, vol. 41, no. 6, pp. 708-711.
Halpern, M.E., Rhee, J., Goll, M.G., Akitake, C.M., Parsons, M. & Leach, S.D. 2008, "Gal4/UAS transgenic tools and their application to zebrafish", Zebrafish, vol. 5, no. 2, pp. 97-110.
Hansson, C.A., Frykman, S., Farmery, M.R., Tjernberg, L.O., Nilsberth, C., Pursglove, S.E., Ito, A., Winblad, B., Cowburn, R.F., Thyberg, J. & Ankarcrona, M. 2004, "Nicastrin, presenilin, APH-1, and PEN-2 form active gamma-secretase complexes in mitochondria ", The Journal of biological chemistry, vol. 279, no. 49, pp. 51654-51660.
Higashijima, S., Mandel, G. & Fetcho, J.R. 2004, "Distribution of prospective glutamatergic, glycinergic, and GABAergic neurons in embryonic and larval zebrafish ", The Journal of comparative neurology, vol. 480, no. 1, pp. 1-18.
Higuchi, M., Yanai, K., Okamura, N., Meguro, K., Arai, H., Itoh, M., Iwata, R., Ido, T., Watanabe, T. & Sasaki, H. 2000, "Histamine H(1) receptors in patients with Alzheimer's disease assessed by positron emission tomography", Neuroscience, vol. 99, no. 4, pp. 721-729.
Hillegaart, V. 1990, "Effects of local application of 5-HT and 8-OH-DPAT into the dorsal and median raphe nuclei on motor activity in the rat", Physiology & Behavior, vol. 48, no. 1, pp. 143-148.
Hough, L.B. & Domino, E.F. 1979, "Tele-methylhistamine oxidation by type B monoamine oxidase", The Journal of pharmacology and experimental therapeutics, vol. 208, no. 3, pp. 422-428.
Inestrosa, N.C. & Arenas, E. 2010, "Emerging roles of Wnts in the adult nervous system", Nature Reviews Neuroscience, vol. 11, no. 2, pp. 77-86.
Inoue, I., Yanai, K., Kitamura, D., Taniuchi, I., Kobayashi, T., Niimura, K., Watanabe, T. & Watanabe, T. 1996, "Impaired locomotor activity and exploratory behavior in mice lacking histamine H1 receptors", Proceedings of the National Academy of Sciences of the United States of America, vol. 93, no. 23, pp. 13316-13320.
Isa, T. 2002, "Intrinsic processing in the mammalian superior colliculus", Current opinion in neurobiology, vol. 12, no. 6, pp. 668-677.
Ito, H. & Yamamoto, N. 2009, "Non-laminar cerebral cortex in teleost fishes?", Biology letters, vol. 5, no. 1, pp. 117-121.
Jang, I.S., Rhee, J.S., Watanabe, T., Akaike, N. & Akaike, N. 2001, "Histaminergic modulation of GABAergic transmission in rat ventromedial hypothalamic neurones", The Journal of physiology, vol. 534, no. 3, pp. 791-803.
Jeong, J.Y., Kwon, H.B., Ahn, J.C., Kang, D., Kwon, S.H., Park, J.A. & Kim, K.W. 2008, "Functional and developmental analysis of the blood-brain barrier in zebrafish", Brain research bulletin, vol. 75, no. 5, pp. 619-628.
Jeong, H.J., Moon, P.D., Kim, S.J., Seo, J.U., Kang, T.H., Kim, J.J., Kang, I.C., Um, J.Y., Kim, H.M. & Hong, S.H. 2009, "Activation of hypoxia-inducible factor-1 regulates human histidine decarboxylase expression", Cellular and molecular life sciences, vol. 66, no. 7, pp. 1309-1319.
Jequier, E., Lovenberg, W. & Sjoerdsma, A. 1967, "Tryptophan hydroxylase inhibition: the mechanism by which p-chlorophenylalanine depletes rat brain serotonin", Molecular pharmacology, vol. 3, no. 3, pp. 274-278.
Jin, C.Y., Anichtchik, O. & Panula, P. 2009, "Altered histamine H3 receptor radioligand binding in post-mortem brain samples from subjects with psychiatric diseases", British journal of pharmacology, vol. 157, no. 1, pp. 118-129.
Jin, C.Y. & Panula, P. 2005, "The laminar histamine receptor system in human prefrontal cortex suggests multiple levels of histaminergic regulation", Neuroscience, vol. 132, no. 1, pp. 137-149.
Kallweit, U., Hidalgo, H., Engel, A., Baumann, C.R., Bassetti, C.L. & Dahmen, N. 2011, "Post H1N1 vaccination narcolepsy-cataplexy with decreased CSF beta-amyloid", Sleep medicine, vol. 13, no. 3, pp. 323.
Kandel, E.R., Schwartz, J.H. & Jessell, T.M. (eds) 1991, Principles of Neural Science, Appelton & Lange, East Norwalk, Connecticut, pp. 531-547, 997-1007.

77
Kang, D.E., Soriano, S., Xia, X., Eberhart, C.G., De Strooper, B., Zheng, H. & Koo, E.H. 2002, "Presenilin couples the paired phosphorylation of beta-catenin independent of axin: implications for beta-catenin activation in tumorigenesis", Cell, vol. 110, no. 6, pp. 751-762.
Kang, J.E., Lim, M.M., Bateman, R.J., Lee, J.J., Smyth, L.P., Cirrito, J.R., Fujiki, N., Nishino, S. & Holtzman, D.M. 2009, "Amyloid-beta dynamics are regulated by orexin and the sleep-wake cycle", Science, vol. 326, no. 5955, pp. 1005-1007.
Karhunen, T., Airaksinen, M.S., Tuomisto, L. & Panula, P. 1993, "Neurotransmitters in the nervous system of Macoma balthica (Bivalvia)", The Journal of comparative neurology, vol. 334, no. 3, pp. 477-488.
Kaslin, J. & Panula, P. 2001, "Comparative anatomy of the histaminergic and other aminergic systems in zebrafish (Danio rerio)", The Journal of comparative neurology, vol. 440, no. 4, pp. 342-377.
Kaslin, J., Nystedt, J.M., Ostergard, M., Peitsaro, N. & Panula, P. 2004, "The orexin/hypocretin system in zebrafish is connected to the aminergic and cholinergic systems", The Journal of Neuroscience, vol. 24, no. 11, pp. 2678-2689.
Keegan, B.R., Feldman, J.L., Lee, D.H., Koos, D.S., Ho, R.K., Stainier, D.Y. & Yelon, D. 2002, "The elongation factors Pandora/Spt6 and Foggy/Spt5 promote transcription in the zebrafish embryo", Development, vol. 129, no. 7, pp. 1623-1632.
Kimberly, W.T., LaVoie, M.J., Ostaszewski, B.L., Ye, W., Wolfe, M.S. & Selkoe, D.J. 2003, "Gamma-secretase is a membrane protein complex comprised of presenilin, nicastrin, Aph-1, and Pen-2 ", Proceedings of the National Academy of Sciences of the United States of America, vol. 100, no. 11, pp. 6382-6387.
Kobayashi, T., Tonai, S., Ishihara, Y., Koga, R., Okabe, S. & Watanabe, T. 2000, "Abnormal functional and morphological regulation of the gastric mucosa in histamine H2 receptor-deficient mice", The Journal of clinical investigation, vol. 105, no. 12, pp. 1741-1749.
Koe, B.K. & Weissman, A. 1966, "p-Chlorophenylalanine: a specific depletor of brain serotonin", The Journal of pharmacology and experimental therapeutics, vol. 154, no. 3, pp. 499-516.
Kornum, B.R., Faraco, J. & Mignot, E. 2011, "Narcolepsy with hypocretin/orexin deficiency, infections and autoimmunity of the brain", Current opinion in neurobiology, vol. 21, no. 6, pp. 897-903.
Kovacs, D.M., Fausett, H.J., Page, K.J., Kim, T.W., Moir, R.D., Merriam, D.E., Hollister, R.D., Hallmark, O.G., Mancini, R., Felsenstein, K.M., Hyman, B.T., Tanzi, R.E. & Wasco, W. 1996, "Alzheimer-associated presenilins 1 and 2: neuronal expression in brain and localization to intracellular membranes in mammalian cells ", Nature medicine, vol. 2, no. 2, pp. 224-229.
Kudo, H., Liu, J., Jansen, E.J., Ozawa, A., Panula, P., Martens, G.J. & Lindberg, I. 2009, "Identification of proSAAS homologs in lower vertebrates: conservation of hydrophobic helices and convertase-inhibiting sequences", Endocrinology, vol. 150, no. 3, pp. 1393-1399.
Kukko-Lukjanov, T.K. & Panula, P. 2003, "Subcellular distribution of histamine, GABA and galanin in tuberomamillary neurons in vitro", Journal of chemical neuroanatomy, vol. 25, no. 4, pp. 279-292.
Kukko-Lukjanov, T.K., Soini, S., Taira, T., Michelsen, K.A., Panula, P. & Holopainen, I.E. 2006, "Histaminergic neurons protect the developing hippocampus from kainic acid-induced neuronal damage in an organotypic coculture system", The Journal of Neuroscience, vol. 26, no. 4, pp. 1088-1097.
Lange, M., Norton, W., Coolen, M., Chaminade, M., Merker, S., Proft, F., Schmitt, A., Vernier, P., Lesch, K.P. & Bally-Cuif, L. 2012, "The ADHD-susceptibility gene lphn3.1 modulates dopaminergic neuron formation and locomotor activity during zebrafish development", Molecular psychiatry, In press.
Leurs, R., Smit, M.J., Menge, W.M. & Timmerman, H. 1994, "Pharmacological characterization of the human histamine H2 receptor stably expressed in Chinese hamster ovary cells", British journal of pharmacology, vol. 112, no. 3, pp. 847-854.
Levin, E.Y., Levenberg, B. & Kaufman, S. 1960, "The enzymatic conversion of 3,4-dihydroxyphenylethylamine to norepinephrine", The Journal of biological chemistry, vol. 235, pp. 2080-2086.
Levitan, D. & Greenwald, I. 1995, "Facilitation of lin-12-mediated signalling by sel-12, a Caenorhabditis elegans S182 Alzheimer's disease gene ", Nature, vol. 377, no. 6547, pp. 351-354.

78
Lin, L., Wisor, J., Shiba, T., Taheri, S., Yanai, K., Wurts, S., Lin, X., Vitaterna, M., Takahashi, J., Lovenberg, T.W., Koehl, M., Uhl, G., Nishino, S. & Mignot, E. 2002, "Measurement of hypocretin/orexin content in the mouse brain using an enzyme immunoassay: the effect of circadian time, age and genetic background", Peptides, vol. 23, no. 12, pp. 2203-2211.
Lin, J.S., Anaclet, C., Sergeeva, O.A. & Haas, H.L. 2011, "The waking brain: an update", Cellular and molecular life sciences, vol. 68, no. 15, pp. 2499-2512.
Lovenberg, W., Weissbach, H. & Udenfriend, S. 1962, "Aromatic L-amino acid decarboxylase", The Journal of biological chemistry, vol. 237, pp. 89-93.
Lovenberg, W., Jequier, E. & Sjoerdsma, A. 1967, "Tryptophan hydroxylation: measurement in pineal gland, brainstem, and carcinoid tumor", Science, vol. 155, no. 759, pp. 217-219.
Machluf, Y., Gutnick, A. & Levkowitz, G. 2011, "Development of the zebrafish hypothalamus", Annals of the New York Academy of Sciences, vol. 1220, pp. 93-105.
Mahler, J., Filippi, A. & Driever, W. 2010, "DeltaA/DeltaD regulate multiple and temporally distinct phases of notch signaling during dopaminergic neurogenesis in zebrafish", The Journal of Neuroscience, vol. 30, no. 49, pp. 16621-16635.
McCallum, C.M., Comai, L., Greene, E.A. & Henikoff, S. 2000, "Targeted screening for induced mutations", Nature biotechnology, vol. 18, no. 4, pp. 455-457.
McGinty, D. & Szymusiak, R. 2003, "Hypothalamic regulation of sleep and arousal", Frontiers in bioscience, vol. 8, pp. 1074-1083.
Meng, X., Noyes, M.B., Zhu, L.J., Lawson, N.D. & Wolfe, S.A. 2008, "Targeted gene inactivation in zebrafish using engineered zinc-finger nucleases", Nature biotechnology, vol. 26, no. 6, pp. 695-701.
Merickel, A. & Edwards, R.H. 1995, "Transport of histamine by vesicular monoamine transporter-2", Neuropharmacology, vol. 34, no. 11, pp. 1543-1547.
Metscher, B.D. & Ahlberg, P.E. 1999, "Zebrafish in context: uses of a laboratory model in comparative studies", Developmental biology, vol. 210, no. 1, pp. 1-14.
Mochizuki, T., Yamatodani, A., Okakura, K., Horii, A., Inagaki, N. & Wada, H. 1992, "Circadian rhythm of histamine release from the hypothalamus of freely moving rats", Physiology & Behavior, vol. 51, no. 2, pp. 391-394.
Molina-Hernandez, A. & Velasco, I. 2008, "Histamine induces neural stem cell proliferation and neuronal differentiation by activation of distinct histamine receptors", Journal of neurochemistry, vol. 106, no. 2, pp. 706-717.
Molinengo, L. & Ghi, P. 1997, "Behavioral and neurochemical effects induced by subchronic l-deprenyl administration", Pharmacology, biochemistry, and behavior, vol. 58, no. 3, pp. 649-655.
Monti, J.M., Pellejero, T. & Jantos, H. 1986, "Effects of H1- and H2-histamine receptor agonists and antagonists on sleep and wakefulness in the rat", Journal of neural transmission, vol. 66, no. 1, pp. 1-11.
Monti, J.M., Jantos, H., Boussard, M., Altier, H., Orellana, C. & Olivera, S. 1991, "Effects of selective activation or blockade of the histamine H3 receptor on sleep and wakefulness", European journal of pharmacology, vol. 205, no. 3, pp. 283-287.
Mueller, T., Wullimann, M.F. & Guo, S. 2008, "Early teleostean basal ganglia development visualized by zebrafish Dlx2a, Lhx6, Lhx7, Tbr2 (eomesa), and GAD67 gene expression", The Journal of comparative neurology, vol. 507, no. 2, pp. 1245-1257.
Mueller, T., Dong, Z., Berberoglu, M.A. & Guo, S. 2011, "The dorsal pallium in zebrafish, Danio rerio (Cyprinidae, Teleostei)", Brain research, vol. 1381, pp. 95-105.
Muto, A., Orger, M.B., Wehman, A.M., Smear, M.C., Kay, J.N., Page-McCaw, P.S., Gahtan, E., Xiao, T., Nevin, L.M., Gosse, N.J., Staub, W., Finger-Baier, K. & Baier, H. 2005, "Forward genetic analysis of visual behavior in zebrafish ", PLoS genetics, vol. 1, no. 5, pp. e66.
Nagatsu, T., Levitt, M. & Udenfriend, S. 1964, "Tyrosine Hydroxylase the Initial Step in Norepinephrine Biosynthesis", The Journal of biological chemistry, vol. 239, pp. 2910-2917.
Nicotra, A., Pierucci, F., Parvez, H. & Senatori, O. 2004, "Monoamine oxidase expression during development and aging", Neurotoxicology, vol. 25, no. 1-2, pp. 155-165.
Nieuwenhuys, R. 1963, "The Comparative Anatomy of the Actinopterygian Forebrain", Journal fur Hirnforschung, vol. 13, pp. 171-192.

79
Nieuwenhuys, R., ten Donkelar, H.J. & Nicholson, C. 1998, The Central Nervous System of Vertebrates, Springer, Berlin.
Nieuwenhuys, R. 2010, "The development and general morphology of the telencephalon of actinopterygian fishes: synopsis, documentation and commentary", Brain structure & function, vol. 215, no. 3-4, pp. 141-157.
Nishino, S., Sakurai, E., Nevsimalova, S., Yoshida, Y., Watanabe, T., Yanai, K. & Mignot, E. 2009, "Decreased CSF histamine in narcolepsy with and without low CSF hypocretin-1 in comparison to healthy controls", Sleep, vol. 32, no. 2, pp. 175-180.
Nornes, S., Groth, C., Camp, E., Ey, P. & Lardelli, M. 2003, "Developmental control of Presenilin1 expression, endoproteolysis, and interaction in zebrafish embryos", Experimental cell research, vol. 289, no. 1, pp. 124-132.
Nornes, S., Newman, M., Verdile, G., Wells, S., Stoick-Cooper, C.L., Tucker, B., Frederich-Sleptsova, I., Martins, R. & Lardelli, M. 2008, "Interference with splicing of Presenilin transcripts has potent dominant negative effects on Presenilin activity", Human molecular genetics, vol. 17, no. 3, pp. 402-412.
Nornes, S., Newman, M., Wells, S., Verdile, G., Martins, R.N. & Lardelli, M. 2009, "Independent and cooperative action of Psen2 with Psen1 in zebrafish embryos", Experimental cell research, vol. 315, no. 16, pp. 2791-2801.
Norton, W.H., Stumpenhorst, K., Faus-Kessler, T., Folchert, A., Rohner, N., Harris, M.P., Callebert, J. & Bally-Cuif, L. 2011, "Modulation of Fgfr1a signaling in zebrafish reveals a genetic basis for the aggression-boldness syndrome", The Journal of Neuroscience, vol. 31, no. 39, pp. 13796-13807.
Ohtsu, H., Tanaka, S., Terui, T., Hori, Y., Makabe-Kobayashi, Y., Pejler, G., Tchougounova, E., Hellman, L., Gertsenstein, M., Hirasawa, N., Sakurai, E., Buzas, E., Kovacs, P., Csaba, G., Kittel, A., Okada, M., Hara, M., Mar, L., Numayama-Tsuruta, K., Ishigaki-Suzuki, S., Ohuchi, K., Ichikawa, A., Falus, A., Watanabe, T. & Nagy, A. 2001, "Mice lacking histidine decarboxylase exhibit abnormal mast cells", FEBS letters, vol. 502, no. 1-2, pp. 53-56.
Oleykowski, C.A., Bronson Mullins, C.R., Godwin, A.K. & Yeung, A.T. 1998, "Mutation detection using a novel plant endonuclease", Nucleic acids research, vol. 26, no. 20, pp. 4597-4602.
O'Malley, D.M., Kao, Y.H. & Fetcho, J.R. 1996, "Imaging the functional organization of zebrafish hindbrain segments during escape behaviors ", Neuron, vol. 17, no. 6, pp. 1145-1155.
Orger, M.B., Kampff, A.R., Severi, K.E., Bollmann, J.H. & Engert, F. 2008, "Control of visually guided behavior by distinct populations of spinal projection neurons", Nature neuroscience, vol. 11, no. 3, pp. 327-333.
Osei-Owusu, P., James, A., Crane, J. & Scrogin, K.E. 2005, "5-Hydroxytryptamine 1A receptors in the paraventricular nucleus of the hypothalamus mediate oxytocin and adrenocorticotropin hormone release and some behavioral components of the serotonin syndrome", The Journal of pharmacology and experimental therapeutics, vol. 313, no. 3, pp. 1324-1330.
Panula, P., Yang, H.Y. & Costa, E. 1984, "Histamine-containing neurons in the rat hypothalamus", Proceedings of the National Academy of Sciences of the United States of America, vol. 81, no. 8, pp. 2572-2576.
Panula, P., Pirvola, U., Auvinen, S. & Airaksinen, M.S. 1989, "Histamine-immunoreactive nerve fibers in the rat brain", Neuroscience, vol. 28, no. 3, pp. 585-610.
Panula, P., Airaksinen, M.S., Pirvola, U. & Kotilainen, E. 1990, "A histamine-containing neuronal system in human brain", Neuroscience, vol. 34, no. 1, pp. 127-132.
Panula, P., Rinne, J., Kuokkanen, K., Eriksson, K.S., Sallmen, T., Kalimo, H. & Relja, M. 1998, "Neuronal histamine deficit in Alzheimer's disease", Neuroscience, vol. 82, no. 4, pp. 993-997.
Panula, P., Sallinen, V., Sundvik, M., Kolehmainen, J., Torkko, V., Tiittula, A., Moshnyakov, M. & Podlasz, P. 2006, "Modulatory neurotransmitter systems and behavior: towards zebrafish models of neurodegenerative diseases", Zebrafish, vol. 3, no. 2, pp. 235-247.
Panula, P. 2009, "Hypocretin/orexin in fish physiology with emphasis on zebrafish", Acta physiologica,vol. 198, no. 3, pp. 381-386.
Panula, P., Chen, Y.C., Priyadarshini, M., Kudo, S., Semenova, S., Sundvik, M. & Sallinen, V. 2010, "The comparative neuroanatomy and neurochemistry of zebrafish CNS systems of relevance to human neuropsychiatric diseases", Neurobiology of disease, vol. 40, no. 1, pp. 46-57.

80
Parmentier, R., Ohtsu, H., Djebbara-Hannas, Z., Valatx, J.L., Watanabe, T. & Lin, J.S. 2002, "Anatomical, physiological, and pharmacological characteristics of histidine decarboxylase knock-out mice: evidence for the role of brain histamine in behavioral and sleep-wake control", The Journal of Neuroscience, vol. 22, no. 17, pp. 7695-7711.
Parmentier, R., Kolbaev, S., Klyuch, B.P., Vandael, D., Lin, J.S., Selbach, O., Haas, H.L. & Sergeeva, O.A. 2009, "Excitation of histaminergic tuberomamillary neurons by thyrotropin-releasing hormone", The Journal of Neuroscience, vol. 29, no. 14, pp. 4471-4483.
Pavlidis, M., Sundvik, M., Chen, Y.C. & Panula, P. 2011, "Adaptive changes in zebrafish brain in dominant-subordinate behavioral context", Behavioural brain research, vol. 225, no. 2, pp. 529-537.
Peitsaro, N., Kaslin, J., Anichtchik, O.V. & Panula, P. 2003, "Modulation of the histaminergic system and behaviour by alpha-fluoromethylhistidine in zebrafish", Journal of neurochemistry, vol. 86, no. 2, pp. 432-441.
Peitsaro, N., Sundvik, M., Anichtchik, O.V., Kaslin, J. & Panula, P. 2007, "Identification of zebrafish histamine H1, H2 and H3 receptors and effects of histaminergic ligands on behavior", Biochemical pharmacology, vol. 73, no. 8, pp. 1205-1214.
Penzes, P., Cahill, M.E., Jones, K.A., VanLeeuwen, J.E. & Woolfrey, K.M. 2011, "Dendritic spine pathology in neuropsychiatric disorders", Nature neuroscience, vol. 14, no. 3, pp. 285-293.
Perala, N., Peitsaro, N., Sundvik, M., Koivula, H., Sainio, K., Sariola, H., Panula, P. & Immonen, T. 2010, "Conservation, expression, and knockdown of zebrafish plxnb2a and plxnb2b", Developmental dynamics, vol. 239, no. 10, pp. 2722-2734.
Perry, E., Court, J., Goodchild, R., Griffiths, M., Jaros, E., Johnson, M., Lloyd, S., Piggott, M., Spurden, D., Ballard, C., McKeith, I. & Perry, R. 1998, "Clinical neurochemistry: developments in dementia research based on brain bank material", Journal of neural transmission, vol. 105, no. 8-9, pp. 915-933.
Peyron, C., Faraco, J., Rogers, W., Ripley, B., Overeem, S., Charnay, Y., Nevsimalova, S., Aldrich, M., Reynolds, D., Albin, R., Li, R., Hungs, M., Pedrazzoli, M., Padigaru, M., Kucherlapati, M., Fan, J., Maki, R., Lammers, G.J., Bouras, C., Kucherlapati, R., Nishino, S. & Mignot, E. 2000, "A mutation in a case of early onset narcolepsy and a generalized absence of hypocretin peptides in human narcoleptic brains", Nature medicine, vol. 6, no. 9, pp. 991-997.
Pierfelice, T., Alberi, L. & Gaiano, N. 2011, "Notch in the vertebrate nervous system: an old dog with new tricks", Neuron, vol. 69, no. 5, pp. 840-855.
Pillot, C., Heron, A., Cochois, V., Tardivel-Lacombe, J., Ligneau, X., Schwartz, J.C. & Arrang, J.M. 2002a, "A detailed mapping of the histamine H(3) receptor and its gene transcripts in rat brain", Neuroscience, vol. 114, no. 1, pp. 173-193.
Pillot, C., Ortiz, J., Heron, A., Ridray, S., Schwartz, J.C. & Arrang, J.M. 2002b, "Ciproxifan, a histamine H3-receptor antagonist/inverse agonist, potentiates neurochemical and behavioral effects of haloperidol in the rat", The Journal of Neuroscience, vol. 22, no. 16, pp. 7272-7280.
Portavella, M., Vargas, J.P., Torres, B. & Salas, C. 2002, "The effects of telencephalic pallial lesions on spatial, temporal, and emotional learning in goldfish", Brain research bulletin, vol. 57, no. 3-4, pp. 397-399.
Portavella, M., Torres, B. & Salas, C. 2004, "Avoidance response in goldfish: emotional and temporal involvement of medial and lateral telencephalic pallium", The Journal of Neuroscience, vol. 24, no. 9, pp. 2335-2342.
Portavella, M. & Vargas, J.P. 2005, "Emotional and spatial learning in goldfish is dependent on different telencephalic pallial systems", The European journal of neuroscience, vol. 21, no. 10, pp. 2800-2806.
Prober, D.A., Rihel, J., Onah, A.A., Sung, R.J. & Schier, A.F. 2006, "Hypocretin/orexin overexpression induces an insomnia-like phenotype in zebrafish", The Journal of Neuroscience, vol. 26, no. 51, pp. 13400-13410.
Radomski, J.W., Dursun, S.M., Reveley, M.A. & Kutcher, S.P. 2000, "An exploratory approach to the serotonin syndrome: an update of clinical phenomenology and revised diagnostic criteria", Medical hypotheses, vol. 55, no. 3, pp. 218-224.
Ridges, S., Heaton, W.L., Joshi, D., Choi, H., Eiring, A., Batchelor, L., Choudhry, P., Manos, E.J., Sofla, H., Sanati, A., Welborn, S., Agarwal, A., Spangrude, G.J., Miles, R.R., Cox, J.E., Frazer, J.K., Deininger, M., Balan, K., Sigman, M., Muschen, M., Perova, T., Johnson, R., Montpellier,

81
B., Guidos, C.J., Jones, D.A. & Trede, N.S. 2012, "Zebrafish screen identifies novel compound with selective toxicity against leukemia", Blood, In press.
Rihel, J., Prober, D.A., Arvanites, A., Lam, K., Zimmerman, S., Jang, S., Haggarty, S.J., Kokel, D., Rubin, L.L., Peterson, R.T. & Schier, A.F. 2010, "Zebrafish behavioral profiling links drugs to biological targets and rest/wake regulation", Science, vol. 327, no. 5963, pp. 348-351.
Rink, E. & Wullimann, M.F. 2001, "The teleostean (zebrafish) dopaminergic system ascending to the subpallium (striatum) is located in the basal diencephalon (posterior tuberculum)", Brain research, vol. 889, no. 1-2, pp. 316-330.
Rink, E. & Wullimann, M.F. 2004, "Connections of the ventral telencephalon (subpallium) in the zebrafish (Danio rerio)", Brain research, vol. 1011, no. 2, pp. 206-220.
Robu, M.E., Larson, J.D., Nasevicius, A., Beiraghi, S., Brenner, C., Farber, S.A. & Ekker, S.C. 2007, "P53 Activation by Knockdown Technologies", PLoS genetics, vol. 3, no. 5, pp. e78.
Rodriguez, J.J., Jones, V.C. & Verkhratsky, A. 2009, "Impaired cell proliferation in the subventricular zone in an Alzheimer's disease model", Neuroreport, vol. 20, no. 10, pp. 907-912.
Roegge, C.S., Perraut, C., Hao, X. & Levin, E.D. 2007, "Histamine H1 receptor involvement in prepulse inhibition and memory function: relevance for the antipsychotic actions of clozapine", Pharmacology, biochemistry, and behavior, vol. 86, no. 4, pp. 686-692.
Sager, J.J., Bai, Q. & Burton, E.A. 2010, "Transgenic zebrafish models of neurodegenerative diseases", Brain structure & function, vol. 214, no. 2-3, pp. 285-302.
Sallinen, V., Torkko, V., Sundvik, M., Reenila, I., Khrustalyov, D., Kaslin, J. & Panula, P. 2009, "MPTP and MPP+ target specific aminergic cell populations in larval zebrafish", Journal of neurochemistry, vol. 108, no. 3, pp. 719-731.
Sallinen, V., Kolehmainen, J., Priyadarshini, M., Toleikyte, G., Chen, Y.C. & Panula, P. 2010, "Dopaminergic cell damage and vulnerability to MPTP in Pink1 knockdown zebrafish", Neurobiology of disease, vol. 40, no. 1, pp. 93-101.
Saper, C.B., Scammell, T.E. & Lu, J. 2005, "Hypothalamic regulation of sleep and circadian rhythms", Nature, vol. 437, no. 7063, pp. 1257-1263.
Sato, T., Hamaoka, T., Aizawa, H., Hosoya, T. & Okamoto, H. 2007, "Genetic single-cell mosaic analysis implicates ephrinB2 reverse signaling in projections from the posterior tectum to the hindbrain in zebrafish", The Journal of Neuroscience, vol. 27, no. 20, pp. 5271-5279.
Scammell, T.E. 2003, "The neurobiology, diagnosis, and treatment of narcolepsy", Annals of Neurology, vol. 53, no. 2, pp. 154-166.
Scammell, T.E., Matheson, J.K., Honda, M., Thannickal, T.C. & Siegel, J.M. 2011, "Coexistence of narcolepsy and Alzheimer's disease", Neurobiology of aging, vol. 33, no. 7, pp.1318-1319.
Schayer, R.W. 1953, "Studies on histaminemetabolizing enzymes in intact animals", The Journal of biological chemistry, vol. 203, no. 2, pp. 787-793.
Schlicker, E., Werthwein, S. & Zentner, J. 1999, "Histamine H3 receptor-mediated inhibition of noradrenaline release in the human brain", Fundamental & clinical pharmacology, vol. 13, no. 1, pp. 120-122.
Scholpp, S. & Lumsden, A. 2010, "Building a bridal chamber: development of the thalamus", Trends in neurosciences, vol. 33, no. 8, pp. 373-380.
Schwartz, J.C., Lampart, C. & Rose, C. 1970, "Properties and regional distribution of histidine decarboxylase in rat brain", Journal of neurochemistry, vol. 17, no. 11, pp. 1527-1534.
Schwartz, J.C., Arrang, J.M., Garbarg, M., Gulat-Marnay, C. & Pollard, H. 1990, "Modulation of histamine synthesis and release in brain via presynaptic autoreceptors and heteroreceptors", Annals of the New York Academy of Sciences, vol. 604, pp. 40-54.
Schwartz, J.C., Arrang, J.M., Garbarg, M., Pollard, H. & Ruat, M. 1991, "Histaminergic transmission in the mammalian brain", Physiological Reviews, vol. 71, no. 1, pp. 1-51.
Serban, G., Kouchi, Z., Baki, L., Georgakopoulos, A., Litterst, C.M., Shioi, J. & Robakis, N.K. 2005, "Cadherins mediate both the association between PS1 and beta-catenin and the effects of PS1 on beta-catenin stability ", The Journal of biological chemistry, vol. 280, no. 43, pp. 36007-36012.
Shan, L., Bossers, K., Unmehopa, U., Bao, A.M. & Swaab, D.F. 2012, "Alterations in the histaminergic system in Alzheimer's disease: a postmortem study", Neurobiology of aging, In press.
Shen, J., Bronson, R.T., Chen, D.F., Xia, W., Selkoe, D.J. & Tonegawa, S. 1997, "Skeletal and CNS defects in Presenilin-1-deficient mice", Cell, vol. 89, no. 4, pp. 629-639.

82
Sherin, J.E., Elmquist, J.K., Torrealba, F. & Saper, C.B. 1998, "Innervation of histaminergic tuberomammillary neurons by GABAergic and galaninergic neurons in the ventrolateral preoptic nucleus of the rat", The Journal of Neuroscience, vol. 18, no. 12, pp. 4705-4721.
Shimamura, T., Shiroishi, M., Weyand, S., Tsujimoto, H., Winter, G., Katritch, V., Abagyan, R., Cherezov, V., Liu, W., Han, G.W., Kobayashi, T., Stevens, R.C. & Iwata, S. 2011, "Structure of the human histamine H1 receptor complex with doxepin", Nature, vol. 475, no. 7354, pp. 65-70.
Sjoerdsma, A., Smith, T.E., Stevenson, T.D. & Udenfriend, S. 1955, "Metabolism of 5-hydroxytryptamine (serotonin) by monoamine oxidase", Proceedings of the Society for Experimental Biology and Medicine, vol. 89, no. 1, pp. 36-38.
Smith, J., Bibikova, M., Whitby, F.G., Reddy, A.R., Chandrasegaran, S. & Carroll, D. 2000, "Requirements for double-strand cleavage by chimeric restriction enzymes with zinc finger DNA-recognition domains", Nucleic acids research, vol. 28, no. 17, pp. 3361-3369.
Spitzer, N.C. 2012, "Activity-dependent neurotransmitter respecification", Nature Reviews Neuroscience, vol. 13, no. 2, pp. 94-106.
Strakhova, M.I., Nikkel, A.L., Manelli, A.M., Hsieh, G.C., Esbenshade, T.A., Brioni, J.D. & Bitner, R.S. 2009, "Localization of histamine H4 receptors in the central nervous system of human and rat", Brain research, vol. 1250, pp. 41-48.
Stutzmann, G.E., Caccamo, A., LaFerla, F.M. & Parker, I. 2004, "Dysregulated IP3 signaling in cortical neurons of knock-in mice expressing an Alzheimer's-linked mutation in presenilin1 results in exaggerated Ca2+ signals and altered membrane excitability ", The Journal of Neuroscience, vol. 24, no. 2, pp. 508-513.
Suster, M.L., Kikuta, H., Urasaki, A., Asakawa, K. & Kawakami, K. 2009, "Transgenesis in zebrafish with the tol2 transposon system", Methods in molecular biology, vol. 561, pp. 41-63.
Szobota, S., Gorostiza, P., Del Bene, F., Wyart, C., Fortin, D.L., Kolstad, K.D., Tulyathan, O., Volgraf, M., Numano, R., Aaron, H.L., Scott, E.K., Kramer, R.H., Flannery, J., Baier, H., Trauner, D. & Isacoff, E.Y. 2007, "Remote control of neuronal activity with a light-gated glutamate receptor", Neuron, vol. 54, no. 4, pp. 535-545.
Szymusiak, R., Alam, N., Steininger, T.L. & McGinty, D. 1998, "Sleep-waking discharge patterns of ventrolateral preoptic/anterior hypothalamic neurons in rats", Brain research, vol. 803, no. 1-2, pp. 178-188.
Takahashi, K., Lin, J.S. & Sakai, K. 2006, "Neuronal activity of histaminergic tuberomammillary neurons during wake-sleep states in the mouse", The Journal of Neuroscience, vol. 26, no. 40, pp. 10292-10298.
Thisse, C. & Thisse, B. 2008, "High-resolution in situ hybridization to whole-mount zebrafish embryos", Nature protocols, vol. 3, no. 1, pp. 59-69.
Toppila, J., Stenberg, D., Alanko, L., Asikainen, M., Urban, J.H., Turek, F.W. & Porkka-Heiskanen, T. 1995, "REM sleep deprivation induces galanin gene expression in the rat brain ", Neuroscience letters, vol. 183, no. 3, pp. 171-174.
Torres, G.E., Gainetdinov, R.R. & Caron, M.G. 2003, "Plasma membrane monoamine transporters: structure, regulation and function", Nature Reviews Neuroscience, vol. 4, no. 1, pp. 13-25.
Toyota, H., Dugovic, C., Koehl, M., Laposky, A.D., Weber, C., Ngo, K., Wu, Y., Lee, D.H., Yanai, K., Sakurai, E., Watanabe, T., Liu, C., Chen, J., Barbier, A.J., Turek, F.W., Fung-Leung, W.P. & Lovenberg, T.W. 2002, "Behavioral characterization of mice lacking histamine H(3) receptors", Molecular pharmacology, vol. 62, no. 2, pp. 389-397.
Trivedi, P., Yu, H., MacNeil, D.J., Van der Ploeg, L.H. & Guan, X.M. 1998, "Distribution of orexin receptor mRNA in the rat brain", FEBS letters, vol. 438, no. 1-2, pp. 71-75.
Trottier, S., Chotard, C., Traiffort, E., Unmehopa, U., Fisser, B., Swaab, D.F. & Schwartz, J.C. 2002, "Co-localization of histamine with GABA but not with galanin in the human tuberomamillary nucleus", Brain research, vol. 939, no. 1-2, pp. 52-64.
Vanhala, A., Yamatodani, A. & Panula, P. 1994, "Distribution of histamine-, 5-hydroxytryptamine-, and tyrosine hydroxylase-immunoreactive neurons and nerve fibers in developing rat brain", The Journal of comparative neurology, vol. 347, no. 1, pp. 101-114.
Vargas, J.P., Lopez, J.C. & Portavella, M. 2009, "What are the functions of fish brain pallium?", Brain research bulletin, vol. 79, no. 6, pp. 436-440.
Veeraraghavalu, K., Choi, S.H., Zhang, X. & Sisodia, S.S. 2010, "Presenilin 1 mutants impair the self-renewal and differentiation of adult murine subventricular zone-neuronal progenitors via cell-

83
autonomous mechanisms involving notch signaling ", The Journal of Neuroscience, vol. 30, no. 20, pp. 6903-6915.
Vizuete, M.L., Merino, M., Venero, J.L., Santiago, M., Cano, J. & Machado, A. 2000, "Histamine infusion induces a selective dopaminergic neuronal death along with an inflammatory reaction in rat substantia nigra", Journal of neurochemistry, vol. 75, no. 2, pp. 540-552.
von Economo, C. 1930, "Sleep as a problem of localization", The Journal of Nervous and Mental Disease, vol. 71, no. 3, pp. 249-259.
Walther, D.J., Peter, J.U., Bashammakh, S., Hortnagl, H., Voits, M., Fink, H. & Bader, M. 2003, "Synthesis of serotonin by a second tryptophan hydroxylase isoform", Science, vol. 299, no. 5603, pp. 76.
Watanabe, T., Taguchi, Y., Shiosaka, S., Tanaka, J., Kubota, H., Terano, Y., Tohyama, M. & Wada, H. 1984, "Distribution of the histaminergic neuron system in the central nervous system of rats; a fluorescent immunohistochemical analysis with histidine decarboxylase as a marker", Brain research, vol. 295, no. 1, pp. 13-25.
Wen, P.H., Shao, X., Shao, Z., Hof, P.R., Wisniewski, T., Kelley, K., Friedrich, V.L.,Jr, Ho, L., Pasinetti, G.M., Shioi, J., Robakis, N.K. & Elder, G.A. 2002, "Overexpression of wild type but not an FAD mutant presenilin-1 promotes neurogenesis in the hippocampus of adult mice", Neurobiology of disease, vol. 10, no. 1, pp. 8-19.
Westerfield, M. 2007, The Zebrafish Book, 5th Edition; A guide for the laboratory use of zebrafish (Danio rerio), University of Oregon Press, Eugene, http://zfin.org/zf_info/zfbook/zfbk.html.
Whitehouse, P.J., Price, D.L., Struble, R.G., Clark, A.W., Coyle, J.T. & Delon, M.R. 1982, "Alzheimer's disease and senile dementia: loss of neurons in the basal forebrain", Science, vol. 215, no. 4537, pp. 1237-1239.
Wienholds, E., Schulte-Merker, S., Walderich, B. & Plasterk, R.H. 2002, "Target-selected inactivation of the zebrafish rag1 gene", Science, vol. 297, no. 5578, pp. 99-102.
Wienholds, E., van Eeden, F., Kosters, M., Mudde, J., Plasterk, R.H. & Cuppen, E. 2003, "Efficient target-selected mutagenesis in zebrafish", Genome research, vol. 13, no. 12, pp. 2700-2707.
Wollen, K.A. 2010, "Alzheimer's disease: the pros and cons of pharmaceutical, nutritional, botanical, and stimulatory therapies, with a discussion of treatment strategies from the perspective of patients and practitioners", Alternative Medicine Review, vol. 15, no. 3, pp. 223-244.
Wullimann, M.F., Rupp, B. & Reichert, H. (eds) 1996, Neuroanatomy of the Zebrafish Brain. A Topological Atlas., Birkhäuser Verlag, Basel, Switzerland, pp. 1-144.
Xia, X., Qian, S., Soriano, S., Wu, Y., Fletcher, A.M., Wang, X.J., Koo, E.H., Wu, X. & Zheng, H. 2001, "Loss of presenilin 1 is associated with enhanced beta-catenin signaling and skin tumorigenesis", Proceedings of the National Academy of Sciences of the United States of America, vol. 98, no. 19, pp. 10863-10868.
Xie, J., Farage, E., Sugimoto, M. & Anand-Apte, B. 2010, "A novel transgenic zebrafish model for blood-brain and blood-retinal barrier development", BMC developmental biology, vol. 10, pp. 76.
Xu, X., Shi, Y.C., Gao, W., Mao, G., Zhao, G., Agrawal, S., Chisolm, G.M., Sui, D. & Cui, M.Z. 2002, "The novel presenilin-1-associated protein is a proapoptotic mitochondrial protein ", The Journal of biological chemistry, vol. 277, no. 50, pp. 48913-48922.
Yamamoto, Y., Mochizuki, T., Okakura-Mochizuki, K., Uno, A. & Yamatodani, A. 1997, "Thioperamide, a histamine H3 receptor antagonist, increases GABA release from the rat hypothalamus", Methods and findings in experimental and clinical pharmacology, vol. 19, no. 5, pp. 289-298.
Yamanaka, A., Tsujino, N., Funahashi, H., Honda, K., Guan, J.L., Wang, Q.P., Tominaga, M., Goto, K., Shioda, S. & Sakurai, T. 2002, "Orexins activate histaminergic neurons via the orexin 2 receptor", Biochemical and biophysical research communications, vol. 290, no. 4, pp. 1237-1245.
Yanai, K., Son, L.Z., Endou, M., Sakurai, E., Nakagawasai, O., Tadano, T., Kisara, K., Inoue, I., Watanabe, T. & Watanabe, T. 1998, "Behavioural characterization and amounts of brain monoamines and their metabolites in mice lacking histamine H1 receptors", Neuroscience, vol. 87, no. 2, pp. 479-487.

84
Yanovsky, Y., Li, S., Klyuch, B.P., Yao, Q., Blandina, P., Passani, M.B., Lin, J.S., Haas, H.L. & Sergeeva, O.A. 2011, "L-Dopa activates histaminergic neurons", The Journal of physiology, vol. 589, no. Pt 6, pp. 1349-1366.
Yates, S.L., Tedford, C.E., Gregory, R., Pawlowski, G.P., Handley, M.K., Boyd, D.L. & Hough, L.B. 1999, "Effects of selected histamine H3 receptor antagonists on tele-methylhistamine levels in rat cerebral cortex", Biochemical pharmacology, vol. 57, no. 9, pp. 1059-1066.
Yokogawa, T., Marin, W., Faraco, J., Pezeron, G., Appelbaum, L., Zhang, J., Rosa, F., Mourrain, P. & Mignot, E. 2007, "Characterization of sleep in zebrafish and insomnia in hypocretin receptor mutants", PLoS biology, vol. 5, no. 10, pp. e277.
Zaharna, M., Dimitriu, A. & Guilleminault, C. 2010, "Expert opinion on pharmacotherapy of narcolepsy", Expert opinion on pharmacotherapy, vol. 11, no. 10, pp. 1633-1645.
Zhdanova, I.V. 2006, "Sleep in zebrafish", Zebrafish, vol. 3, no. 2, pp. 215-226. Zlokovic, B.V. 2011, "Neurovascular pathways to neurodegeneration in Alzheimer's disease and other
disorders", Nature Reviews Neuroscience, vol. 12, no. 12, pp. 723-738.

85
APPENDIX Original publications