Embed Size (px)
Citation preview

Vol. 35, No. 1 Spring/Summer 2002
THEGREAT LAKES
ENTOMOLOGIST
THE GREAT LAKES ENTOMOLOGISTPublished by the Michigan Entomological Society
Volume 35 No. 1ISSN 0090-0222
TABLE OF CONTENTS
Mating success of gypsy moth (Lepidoptera: Lymantriidae) females in southern Wisconsin Ksenia Tcheslavskaia, Carlyle C. Brewster, and Alexei A. Sharov ....................................1
Eosentomon heatherproctorae n. sp. (Protura: Eosentomidae) from OntarioErnest C. Bernard and Michal Guzowski .......................................................................9
Ecological notes on species of Cleridae (Insecta: Coleoptera) associated with the prairie flora of central North America Jonathan R. Mawdsley ............................................................................................15
Size estimate of a local population of Brychius hungerfordi (Coleoptera: Haliplidae) Michael Grant, Brian Scholtens, Robert Vande Kopple, and Bert Ebbers ...................23
Ant-plant relationships in a re-created tallgrass prairie Chris E. Petersen, Dhvani G. Dave, Charise L. Parker, and Renee E. Petersen ..............27
First record of Rhipiceridae (Coleoptera: Polyphaga: Dascilloidea) from Wisconsin Daniel K. Young and Kerry Katovich ...........................................................................33
Longitarsus melanurus (Coleoptera: Chrysomelidae) reproduces on Onosmodium molle (Boraginacea) Andrew H. Williams ...............................................................................................35
Attraction of apple maggot flies (Diptera: Tephritidae) to synthetic fruit volatile compounds and food attractants in Michigan apple orchards Lukasz L. Stenliski and Ocar E. Liburd ....................................................................37
A preservative-free emergent trap for the isotopic and elemental analysis of emergent insects from a wetland system Richard A. MacKenzie and Jerry L. Kaster ................................................................47
Moths of the Douglas Lake Region (Emmet and Cheboygan counties), Michigan: VI. Miscellaneous small families (Lepidoptera) Edward G. Voss ......................................................................................................53
Tarsal anomaly found in an adult Agabus seriatus (Coleoptera: Dytiscidae)J. L. Snitgen ...............................................................................................................63
Occurrence of two species of old world bees, Anthidium manicatum and A. oblongatum(Apoidea: Megachilidae), in northern Ohio and southern Michigan Shane R. Miller, Robert Gaebel, Randall J. Mitchell, and Mike Arduser .....................65
Trap response of michigan social wasps (Hymenoptera: Vespidae) to the feeding attractants acetic acid, isobutanol, and heptyl butyrate H. C. Reed and P. J. Landolt ....................................................................................71
Ground beetles (Coleoptera: Carabidae) inhabiting stands of reed canary grass Phalaris arundinacea on islands in the lower Chippewa River, Wisconsin
Paula K. Kleintjes, Anita M. Christensen, William J. Barnes and Lori A. Lyons ...............79Development and sex ratio of Melittobia australica and M. digitata (Hymenoptera: Eulophidae)
on Megachile rotundata (Hymenoptera: Megachilidae) and trypoxylon politum (Hymenoptera: Sphecidae) Jorge M. González and Robert W. Matthews ..............................................................85
Food plant and distribution of Meligethes saevus (Coleoptera: Nitidulidae)Andrew H. Williams ...................................................................................................93
COVER PHOTOPolistes dominulus (Hymenoptera: Vespidae) photographed August 2001,
Ann Arbor, MI. Photo by Mark F. O’Brien.
PUBLISHED BY
THE MICHIGAN ENTOMOLOGICAL
SOCIETY
TH
E G
RE
AT
LAK
ES E
NT
OM
OLO
GIST
V
ol. 3
5, N
o. 1
Sp
ring
/Sum
mer 2
002

THE MICHIGAN ENTOMOLOGICAL SOCIETY2002-2003 OFFICERS
President James Dunn President Elect Thomas E. Wallenmaier Immediate Past President Mark O’Brien Treasurer M. C. Nielsen Secretary Robert Kriegel Member-at-Large (2002-05)* John Douglass Member-at-Large (2001-04) Gary Parsons Member-at-Large (2000-03) Gwen Pearson Journal Editor Therese Poland Associate Journal Editor Ronald Priest Newsletter Editor Robert Haack Webmaster Mark O’Brien
*In the past two issues the Member-at-Large (2002-05) has incorrectly been listed as Martin Andree. We apologize for any confusion.
The Michigan Entomological Society traces its origins to the old Detroit Entomological Society and was organized on 4 November 1954 to “. . .promote the science of entomology in all its branches and by all feasible means, and to advance cooperation and good fellowship among persons interested in entomology.” The Society attempts to facilitate the exchange of ideas and information in both amateur and professional circles, and encourages the study of insects by youth. Membership in the Society, which serves the North Central States and adjacent Canada, is open to all persons interested in entomology. There are four pay-ing classes of membership:
Student (to 12th grade)—annual dues $5.00Active—annual dues $15.00 Institutional—annual dues $35.00 Sustaining—annual contribution $25.00 or more Life—$300.00
Dues are paid on a calendar year basis (Jan. 1-Dec. 31).
Memberships accepted before July 1 shall begin on the preceding January 1; memberships accepted at a later date shall begin the following January 1 unless the earlier date is requested and the required dues are paid. All members in good standing receive the Newsletter of the Society and The Great Lakes Entomologist, a quarterly journal. All active and sustaining members may vote in Society affairs.
All dues and contributions to the Society are deductible for Federal income tax purposes.
SUBSCRIPTION INFORMATION
Institutions and organizations, as well as individuals not desiring the benefits of membership, may sub-scribe to The Great Lakes Entomologist at the rate of $30.00 per volume. The journal is published quarterly; subscriptions are accepted only on a volume (4 issues) basis. Single copies of The Great Lakes Entomologist are available at $6.00 each, with a 20 percent discount for 25 or more copies sent to a single address.
MICROFILM EDITION: Positive microfilm copies of the current volume of The Great Lakes Ento-mologist will be available at nominal cost, to members and bona fide subscribers of the paper edition only, at the end of each volume year. Please address all orders and inquiries to University Microfilms, Inc., 300 Zeeb Road, Ann Arbor, Michigan 48106, USA.
Inquiries about back numbers, subscriptions and Society business should be directed to the Secretary, Michigan Entomological Society, Department of Entomology, Michigan State University, East Lansing, Michigan 48824-1115, USA. Manuscripts and related correspondence should be directed to the Editor (see inside back cover).
Copyright © 2003. The Michigan Entomological Society.
INSTRUCTIONS FOR AUTHORSSUBJECTS
Papers dealing with any aspect of entomology will be considered for publication in The Great Lakes Entomolo-gist. Appropriate subjects are those of interest to professional and amateur entomologists in the North Central States and Canada, as well as general papers and revisions directed to a larger audience while retaining an interest to readers in our geographic area. All manuscripts are refereed by two reviewers, except for short notes, which are reviewed at the discretion of the Editor.
REQUIREMENTSManuscripts must be typed, double-spaced, with 1” margins on 8 1/2 x 11” or equivalent size paper, and submitted in triplicate. Please use italics rather than underline. Use subheadings sparingly and set them into paragraphs in boldface. Footnotes (except for author’s addresses, which must be on the title page, and treated as a footnote), legends, and captions of illustrations should be typed on separate sheets of paper. Titles should be concise, identifying the order and family discussed. The author of each species must be given fully at least once in the text, but not in the title or abstract. If a common name is used for a species or group, it should be in accordance with the common names published by the Entomological Society of America. The format for references must follow that used in previous issues of The Great Lakes Entomologist. Literature cited is just that - no unpublished manuscripts or internal memos.
FIGURES & TABLESRemember that the printed page area for The Great Lakes Entomologist is 4.5 x 7 inches. Scale your illustra-tions accordingly. Photographs should be glossy finish (transparencies are not acceptable). Drawings, charts, graphs, and maps must be scaled to permit proper reduction without loss of detail. Please reduce figures to a size no greater than 9 x 12” to permit easier handling. Figures should also be sent on disk and must meet the following criteria: (1) gray-scale images must be submitted as 300 dpi TIFF or EPS files ONLY; (2) line art or graphs must be sent as 600 dpi TIFF or EPS files ONLY. Scanned images should be saved in the na-tive application. Never imbed the images in a word-processing document. Captions for figures should be numbered consecutively and typed in order at the end of the manuscript. Captions should not be attached to illustrations or written on the back of images.Tables should be kept as uncluttered as possible, and should fit normally across a page when typeset by the printers. Tables cannot be submitted as Excel files or graphics, but only as text.Contributors should follow the Council of Biology Editors Style Manual, 5th ed., and examine recent issues of The Great Lakes Entomologist for proper format of manuscripts.
MANUSCRIPTS ON DISKManuscripts must be submitted on computer diskette (Macintosh, MS-DOS, Windows) along with one printed copy, after they have been accepted for publication. The files may be formatted in any popularly used word-processing program, but must be submitted in Rich Text Format (RTF) to avoid any translation problems. Special formatting notes for submitting manuscripts on disk: The organizational format for a manuscript is as seen in the recent issues of the journal: TITLE, Author(s), Abstract, Introduction, Methods & Materials, Results, Discussion, Acknowledgments, Literature Cited, Tables, and a List of Figures. Do not use extra spaces between paragraphs or references in the Lit. Cited. The columns of text in tables should be aligned with TABS, not spaces. Some symbols may not translate properly from one computer system to another. Do not use extbols. So long as these symbols are clearly seen in the manuscript, adjustments can be made in the copy sent to the printers.
PAGE CHARGESPapers published in The Great Lakes Entomologist are subject to a page charge of $35.00 per published page. Members of the Society, who are authors without funds from grants, institutions, or industry, and are unable to pay costs from personal funds, may apply to the Society for financial assistance. Application for subsidy must be made at the time a manuscript is initially submitted for publication. Authors will receive page proof, together with an order blank for separates. Extensive changes to the proof by the author will be billed at a rate of $1.00 per line.
COVER ARTWORKCover art or photographs are desired for upcoming issues. They are published free of charge. We only require that they be suitably prepared as described for images above, and that the subject be identified as accurately as possible.
EDITOR’S ADDRESSAll manuscripts for The Great Lakes Entomologist should be sent to the Editor, Therese Poland, USDA Forest Service, 1407 S. Harrison Rd., Rm. 220, E. Landing, MI 48823
OTHER BUSINESSOther correspondence should be directed to the Secretary, Michigan Entomological Society, c/o Dept. of Entomology, Michigan State University, East Lansing, MI 48824-1115.

2002 THE GREAT LAKES ENTOMOLOGIST 1
1Department of Entomology, Virginia Polytechnic Institute and State University Blacksburg, VA 24061-0319, USA
MATING SUCCESS OF GYPSY MOTH (LEPIDOPTERA: LYMANTRIIDAE) FEMALES IN SOUTHERN WISCONSIN
Ksenia Tcheslavskaia1, Carlyle C. Brewster1, and Alexei A. Sharov1
ABSTRACTMating success of laboratory-reared gypsy moth Lymantria dispar (L.)
females exposed for 24 hr on tree boles and its relationship to male moth counts in pheromone-baited traps was studied in southern Wisconsin. The relation-ship between mating probability of gypsy moth females and male moth counts in traps corresponded to an exponential model that can be used for predicting mating probabilities in sparse isolated populations. Relative attractiveness of females compared with traps was 0.23, which is similar to earlier estimated relative attractiveness of females in Virginia. The mortality of females from predation, however, was found to be significantly lower in Wisconsin than in Virginia, which may contribute to a larger degree of mating success. Increased long-distance dispersal of males could also contribute to the increased mating success of females. The higher rate of spread of gypsy moth populations in Wis-consin compared with other areas may be due to the increased mating success caused by the lower female mortality and higher long-distance dispersal of males.
INTRODUCTION The gypsy moth, Lymantria dispar (Linnaeus), is a pest of hardwood
forests. The insect was introduced accidentally into the United States near Boston, Massachusetts from France (Liebhold et al. 1989) and has spread at a rate of approximately 20 km/yr to the south, west and north from where it was introduced (Liebhold et al. 1992). In 1993 USDA Forest Service initiated the gypsy moth management program. An objective of the program is to slow the spread of the insect by suppressing solitary low-density populations (McFad-den and McManus 1991, Leonard and Sharov 1995), which establish beyond the population front, grow, and contribute to the main population and to the movement of the front (Sharov et al. 1998).
Very few studies have been devoted to understanding the population dynamics mechanisms specific to low-density gypsy moth populations. One such mechanism is mating success of females, which appears to be a critical inverse density-dependent factor (Sharov et al. 1995). Mating success depends mainly on the local abundance of adult males and is correlated highly with the rate of male moth capture in pheromone traps (Sharov et al. 1995). Mating success, therefore, can be predicted from male moth capture in pheromone-baited traps. Sharov et al. (1995) used this approach to study the mating success of gypsy moths in the Appalachian Mountains, Virginia. Another important factor that affects low-density gypsy moth populations is predation, e.g., bird predation on adult males. However, ants were shown to be the most important predators of adult gypsy moth females (Sharov et al. 1995). Ant predators decrease the mating probability of females by decreasing their longevity.
The male moth catches in pheromone-baited traps in Wisconsin range from <1 in the Western and middle part of the state to 30 – 300 in the East (Fig. 1). The spread of gypsy moth populations in southern Wisconsin also occurs much faster than in the Appalachian Mountains, Virginia (Sharov 1998). It has been hypothesized that this difference in the rate of spread between the two regions may be due to the higher mating success of females and subsequent higher rate of growth in low-density populations in Wisconsin (Sharov et al. 1999). In contrast to the Appalachian Mountains, male moth counts in pheromone traps in Wisconsin

2 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
tend to decrease more gradually with increasing distance from the population front (Régniére and Sharov 1998). This suggests that male moth dispersal may be more extensive in Wisconsin than in the Appalachian Mountains. In this paper we describe the study of the relationship between mating success of gypsy moth females and male moth captures in pheromone traps. In order to understand the mechanism of mating success of gypsy moth females in Wisconsin, we compared this mechanism with the mating success observed in the study done previously by Sharov et al. (1995) in the Appalachian Mountains, Virginia.
MATERIALS AND METHODSA study was conducted in Kettle Moraine State Forest and nearby forested
sites in Wisconsin between July 25 and August 6, 2000. Based on male moth catches in pheromone-baited traps (Fig. 1), seven study plots were established at various distances from an advancing gypsy moth front (Fig. 2). Three plots were located in Waukesha County and four plots in Walworth County.
Mating success of gypsy moth females was evaluated using tethered virgin females. Gypsy moth females were raised from pupae obtained from USDA-APHIS Otis Methods Development Center (Otis, MA). Virgin 1-day old females were tethered by a 10-15 cm thread tied to the base of the front wing and attached to a tree by a pushpin (Sharov et al. 1995). Two lines of 10-13 tethered females per line were established in each plot. The lines were separated by a distance of 20 m, and the distance between females in a line was 20-25 m (Fig. 3). A barrier of tanglefoot glue was applied in the radius of 25 cm around females in some of the plots to protect individuals from natural predators (e.g., ants). Females were left on trees for 24 hr, after which they were removed and their fertilization status was determined via dissection. In some plots, where male trap catches were = 0.5 males per trap, females were left on trees for two days. Females that were collected were stored in the vials for 24 hours and then dissected to check for the presence of sperm in the spermatheca. Egg masses also
Fig. 1. Male gypsy moth counts in traps in Wisconsin, 1999

2002 THE GREAT LAKES ENTOMOLOGIST 3
Fig. 2. Map of experimental plots in Wisconsin
Fig. 3. A study plot in Wisconsin showing the layout of tethered gypsy moth females and pheromone-baited traps.

4 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
were analyzed for embryonation in those cases where we were uncertain about whether mating had occurred. Non-fertilized eggs do not develop but remain white, whereas, fertilized eggs become dark and a larva can be observed inside the egg under a dissecting microscope.
Male moth abundance was determined using pheromone-baited traps (Fig. 3). Two pheromone-baited traps were placed in each plot at a distance of 100 m from the lines of females to avoid competition between the two pheromone sources (Fig. 3). Larger distances could not be used because of the fragmented forest landscape. The pheromone traps were checked before placing tethered females on trees and at the time females were collected.
The relationship between mating success of females and male moth capture was described by the exponential model (Sharov et al. 1995):
P(t) = 1 – exp(–s×t×M) (1),where P(t) is the proportion of females that are mated during time t (days), M is the male catch per trap per day, and s is a parameter that can be interpreted as the relative attractiveness of virgin females to males compared with pheromone-baited traps. The parameter s was estimated using nonlinear regression of P(t) versus (t × M) (least squares method). The fit of the model was checked using the coefficient of determination (R2).
Predation rate on gypsy moth females in Wisconsin was estimated by counting the proportion of females removed by predators per day among indi-viduals not protected with a tanglefoot glue barrier. A Chi-square (χ2) test was used to compare the mortality of gypsy moth females in Wisconsin with that in Virginia using the data collected by Sharov in 1995 (Sharov et al. 1995).
RESULTSThe average trap capture rate in the study plots in Wisconsin, ranged from
0.1 to 42.5 male moths per trap per day; the mating probability of gypsy moth females ranged from 0 to 0.95 (Fig. 4). Mating success of females increased with increasing male moth captures in traps. The relative attractiveness of females compared with pheromone traps, s, was 0.23 with a 95% confidence interval of 0.16 to 0.30 (R2 = 0.816).
In Wisconsin, the mortality of gypsy moth females from predation ranged between 0 to 35% per plot per day (Table 1) with an average of 14.2 ± 2.5%.
DISCUSSIONThis first study of gypsy moth mating success in Wisconsin indicates that
the mating probability of females can be predicted from male moth capture in pheromone traps. This may be important for determining the extent to which moth captures are associated with stable reproducing isolated colonies of the gypsy moth in the areas beyond the expanding population front. The threshold number of male moths captured per season that is associated with a stable population was determined using the model (Sharov et al 1995):
1 – exp(–s×t×M) = exp(–r) (2),where s is the parameter that describes the relative attractiveness of virgin females to males compared with pheromone-baited traps, t is female calling time (days), and M is male moth catch per trap per season. The calling time t of females depends on the dynamics of pheromone emission but also may be reduced by predation. Females are usually attractive during the first three days of their lives, and thereafter their attractiveness declines sharply (Collins and Potts 1932, Richerson and Cameron 1974). Considering the predation rate of 14.2% per day as the only mortality cause, the average survival time of females in Wisconsin would be -1/ln(1-0.142) = 6.53 days. This time is longer than the

2002 THE GREAT LAKES ENTOMOLOGIST 5
Table 1. Mortality of gypsy moth females in Wisconsin
Date Plot # Females analyzed % Mortality
29-Jul-00 13 20 4.8 31-Jul-00 1 20 0 31-Jul-00 2 20 0 31-Jul-00 4 23 4.2 01-Aug-00 16 13 35 02-Aug-00 12 13 35 02-Aug-00 16 15 25 03-Aug-00 12 16 20 03-Aug-00 14 17 15 04-Aug-00 1 20 0 04-Aug-00 12 19 5 04-Aug-00 13 19 4.8 04-Aug-00 14 16 20 05-Aug-00 12 20 0 05-Aug-00 14 15 25 05-Aug-00 16 14 30 06-Aug-00 1 20 0 06-Aug-00 2 18 10 06-Aug-00 4 18 10 06-Aug-00 13 19 9.5 06-Aug-00 14 17 15 06-Aug-00 16 15 25
Fig. 4. Mating probability of gypsy moth females and males captured in pheromone-baited traps.

6 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
calling time (3 days). Thus, predation does not appear to limit female calling time in Wisconsin.
The threshold number of males associated with a stable population, M, was also calculated using average population growth rate r = 1.65 (Sharov et al. 1995). This yielded M = 0.23 males per day. Assuming a 3-week average flight period, this is equivalent to 4.8 males/trap/season. Thus, male moth catches below 5 moths per trap are usually not associated with a reproducing popula-tion in this particular location. However, for monitoring purposes, a catch of 5 moths per trap may be an indication of a reproducing population that is located at some distance from the trap (e.g., 1-2 km away).
The results of the study of the mating success of gypsy moth and male moth capture in the pheromone-baited traps in Wisconsin were compared with the results of an analogous study conducted in Appalachian Mountains, Virginia. In the study in Virginia, s was estimated as 0.15 with a 95% confidence interval of 0.09 - 0.23 (Sharov et. al 1995). The confidence intervals of s in Wisconsin and in Virginia overlap (Fig. 4), suggesting that there is no significant difference in the relationships of mating success of gypsy moth females and male moth counts in traps in the two regions. Female mortality from predation in Virginia ranged from 30 to 94% per day with an average predation of 52 ± 5%. Mortality of gypsy moth females from predation, therefore, was significantly lower in Wisconsin than in Virginia (p < 0.001). The threshold number of males associated with a stable population, M, was estimated to be lower in Wisconsin (M = 4.8 males/trap/season) than in Virginia (M = 7 and 15 males/trap/season with and without predation respectively). Therefore, the rate of population spread in Wisconsin is higher than in Virginia.
In the study in Virginia, predation considerably reduced the waiting time of females t = 1.36 days compared to Wisconsin, where t = 6.53 days (Sharov et al. 1995). This suggests that females live longer and have a higher chance of mating in Wisconsin than in Virginia.
In the light of the above discussion we may conclude, that the higher rate of population spread in southern Wisconsin compared with the Appalachian Mountains, Virginia may be due to the increased mating success of females, which is probably caused by the increased long-distance dispersal of males and the longevity of females.
ACKNOWLEDGEMENTSWe thank Kevin Thorpe (USDA, ARS, Beltsville, MD) for providing equip-
ment and help; Gleb Tcheslavski for the help with the fieldwork; Frank Martin (USDA-APHIS Otis Methods Development Center, Otis, MA) for supplying gypsy moth pupae; Department of Entomology at Virginia Tech for providing equipment. This research was funded by USDA Forest Service, Forest Health.
LITERATURE CITEDCollins, C. W. and S. F. Potts. 1932. Attractants for the flying gypsy moths as an aid in
locating new infestations. U.S. Dep. Agric. Tech. Bull. 336.Leonard, D. and A. A. Sharov. 1995. Slow-the-spread project update: Developing a process
for evaluation, pp. 82-85 . In: S. Fosbroke and K. Gottschalk (eds.), Proceedings of the 1995 Gypsy Moth Research Forum. U.S. For. Serv. Gen. Tech. Rep.
Liebhold, A. M., V. Mastro and P. W. Schaefer. 1989. Learning from the legacy of Leopold Trouvelot. Bull. Entomol. Soc. Am. 35: 20-21.
Liebhold, A. M., J. A. Halverson and G. A. Elmes. 1992. Gypsy moth invasion in North America: A quantitative analysis. J. Biogeogr. 19: 513-520.

2002 THE GREAT LAKES ENTOMOLOGIST 7
McFadden, M. W. and M. E. McManus. 1991. An insect out of control? The potential for spread and establishment of the gypsy moth in new forest areas in the United States, pp. 172-186. In: Y.N. Baranchikov, W.J. Mattson, F.P. Hain, and T.L. Payne (eds.), Forest insect guilds: patterns of interaction with host trees. U.S. For. Serv. Gen. Tech. Rep. NE - 153.
Régniére, J. and A. Sharov. 1998. Phenology of Lymantria dispar (Lepidoptera: Lymant-riiade), male flight and the effect of moth dispersal in heterogeneous landscapes. Int. J. Biometeorol. 41: 161-168.
Richerson, J. V. and E. A. Cameron. 1974. Differences in pheromone release and sexual behavior between laboratory-reared and wild gypsy moth adults [Porthetria dispar]. Environ. Entomol. 3: 475-481.
Sharov, A. A., A. M. Liebhold and F. W. Ravlin. 1995. Prediction of gypsy moth (Lepidop-tera: Lymantriidae) mating success from pheromone trap counts. Environ. Entomol. 24: 1239-1244.
Sharov, A. A. 1998. Decision-Support system for STS (www.ento.vt.edu/~sharov/stsdec/)Sharov, A. A., B. C. Pijanowski, A. M. Liebhold and S. H. Gage. 1999. What affects the
rate of gypsy moth (Lepidoptera: Lymantriidae) spread: Winter temperature or forest susceptibility. Agric. Entomol. 1: 37-45.

8 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1

2002 THE GREAT LAKES ENTOMOLOGIST 9
1Department of Entomology and Plant Pathology, Tennessee Agricultural Experiment Station, University of Tennessee, 205 Plant Sciences, 2431 Center Drive, Knoxville, TN 37996-4560.2Department of Biology, Queen’s University, Kingston, Ontario K7L 3N6, Canada.
EOSENTOMON HEATHERPROCTORAE N. SP. (PROTURA: EOSENTOMIDAE) FROM ONTARIO
Ernest C. Bernard1 and Michal Guzowski2
ABSTRACTA new species of eosentomid proturan, Eosentomon heatherproctorae, is
described from Ontario. This new species is a member of the transitorium species-group. It differs from related North American species of this group in the following combination of characters: labral setae absent, foretarsal sensilla d and t3 long and stout, no anterior setae on the sternite of the eighth abdominal seg-ment, and six setae on the sternites of the ninth and tenth abdominal segments.
____________________Among 37 Protura collected from 27 sites at the Queen’s University
Biological Station near Kingston, Ontario, were 10 specimens representing an undescribed species of Eosentomon Berlese. These specimens proved to be members of the transitorium-group sensu Tuxen (1964), characterized by a sharply bent caput processus in the female genital apparatus. This species group is primarily European and North African (Szeptycki 1986), with only a few species known from North America: E. bernardi Nosek, E. richardi Ber-nard, and E. vermontense Nosek (Bernard 1990, Nosek and Kevan 1984). The purpose of this paper is to describe the new species and differentiate it from similar members of the genus.
MATERIALS AND METHODSProtura were extracted from soil cores and preserved in 80% ethanol.
Specimens were cleared and expanded in Marc Andre I fluid at 60 °C for 15 min-utes, then mounted in Marc Andre II medium on glass slides (Christiansen and Bellinger 1998). Mounts were cured in a 60 °C oven for four days, then ringed with Glyptal. Observations were made with phase and differential contrast microscopy. Drawings were made with the aid of a drawing tube.
Eosentomon heatherproctorae new speciesColor and dimensions: Adult body pale amber, mean length (range) 710
µm (609-809); head length without labrum 98 µm (92-103); foretarsal length with-out claw (n = 13) 63 µm (60-64). LR = 11.5; PR = 6.2; TR (n = 12) = 5.7 (5.0-6.3).
Morphology: Pseudoculus broadly oval, with three diffuse, longitudinal lines on the posterior half. Clypeal apodeme with convex, slightly clavate lateral arms weakly connected to each other anteriorly (Fig. 1). Labrum with truncate apices angling inward to a V-shaped notch; labral setae absent (Fig. 2). Central pair of rostral setae inflated (Fig. 2), the inflation more visible laterally than dorsally. Digits of galea rounded, the outer digit curved slightly outward and longer than the other two digits; both lobes of lacinia curved, outer lobe not visibly dentate, inner lobe not hooked (Fig. 3). Mandible with two apical and one subapical teeth (Fig. 4).
Empodium of foretarsus about equal to length of claw. Empodia of middle and hind legs less than one-eighth the lengths of their ungues.
Central lobe of precosta on abdominal segments IV and V narrow, slightly

10 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
narrowed at the midpoint but not incised (Fig. 9). Female squama genitalis (Fig. 11) with short, broad styli; proximo-lateral sclerotizations well-developed; caput processus prominent, sharply angled toward inner stylus margin; corpus processus narrow, weakly striate proximally, extending to inner stylus margin but not extending outward past the caput processus; median sclerotizations absent; filum processus much shorter than stylus; stylus apices finely rounded.
Chaetotaxy: Cephalic setae aa and pa present, anterior sensillum as present. Setae sp and p about equal in length (Fig. 1).
On mesonotum (Fig. 5), seta p1 slightly longer than p1’; p2 slightly longer than p2’; seta p3’ setiform. Metanotal setae (Fig. 6) similar to those of mesono-tum ; bases of p5 and p5’ touching.
On foretarsus (Figs. 12,13), sensillum b’1 absent, c’ spatulate, thick; sen-sillum a short and slender; b weakly spatulate and almost reaching the base of β6; c slightly expanded, reaching to the base of γ3; d very long, reaching nearly to α7; f1 narrowly spatulate, f2 slender, almost equal in length to f1; e and g clearly spatulate. Sensillum t1 equidistant to α3 and α3’, t2 slender; t3 long and thick, nearly reaching base of claw. Sensillum a’ weakly spatulate, b’2 slender and reaching well past α6. On metatarsus (Fig. 10), seta D2 spiniform, less stout than D5.
Abdominal setal composition given in Table 1. Tergum I setal formula 3:1:2, p1’ longer than p1; p3’ and p3’’ minute but swollen (Fig. 7). On terga II-VI, p1’ and p2’ longer than p1 and p2; p4’ more than half the length of p4 and setiform. On tergum VII, p1’ setiform, shorter than p1; p4 and p4’ of equal length and robustness. On tergum VIII, pc and p1’ setiform, p1” aristate, p2’ linear (Fig. 8).
Diagnosis: Eosentomon heatherproctorae n. sp. is a member of the tran-sitorium group as defined by Tuxen (1964). Females in this group have the caput processus strongly reflexed toward the inner edge of the stylus, making a birdhead-like shape. The new species differs from other North American members of this group (E. bernardi Nosek in Nosek & Kevan, E. richardi Ber-nard, and E. vermontense Nosek in Nosek & Kevan) in the lack of labral setae (present in E. bernardi and E. richardi, unknown in E. vermontense), absence of anterior setae on sternum VIII (2 anterior setae in the other species), and six setae on sterna IX-X (four in E. bernardi and E. vermontense, six in E. richardi). Foretarsal sensillum d is much longer in E. heatherproctorae n. sp. than in the other three North American species.
The presence of the accessory head setae aa and pa places E. heatherproc-torae n. sp. in the subset named the “delicatum” group by Szeptycki (1985), where it will key to E. wanda Szeptycki. The new species differs from E. wanda in the following ways: labral setae absent, foretarsal sensillum d long and stout (short in E. wanda), sensillum t1 equidistant to α3' and α3 (in E. wanda, t1 very close to α3'), ten anterior setae on tergum VII (four in E. wanda).
Types: Holotype female collected from soil cores 13 July 1996, Chaffey’s Lock, Queen’s University Biological Station (44°33’45”N, 76°19’21”W), near Kingston, Ontario. Nine paratypes (2 females, 5 males, 2 maturus juniors) col-lected at same locality from 14 June to 13 July 1996, M. Guzowski, collector. Holotype and 4 paratypes deposited in the Canadian National Collection; five paratypes deposited in the apterygote section of the University of Tennessee Entomology Collection.
It is our pleasure to name this species after Dr. Heather Proctor, who directed the research that led to the collection of this new species.

2002 THE GREAT LAKES ENTOMOLOGIST 11
Figures 1-11. Eosentomon heatherproctorae n. sp. 1) Dorsum of head, left side. 2) Labrum and rostral setae, dorsal view. 3) Galea and lacinia of maxilla. 4) Apex of mandible. 5) Mesonotum, posterior margin of left side. 6) Metanotum, posterior mar-gin of left side. 7) Abdominal tergite I, right side. 8) Abdominal tergite VIII, right side. 9) Precosta of abdominal tergum V. 10) Metatarsus. 11) Squama genitalis. Scale bars = 30 µm.

12 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Figures 12, 13. Eosentomon heatherproctorae n. sp. 12) Foretarsus, dorsal view. 13) Foretarsus, ventral view. Scale bar = 15 µm.
Table 1. Abdominal chaetotaxy of Eosentomon heatherproctorae n. sp.
I II III IV-VI VII VIII IX-X XI XII Dorsal Anterior setae 4 10 10 8 10 6 8 8 12 Posterior setae 12 14 14 16 16 9
Ventral Anterior setae 4 6 6 6 6 0 6 8 9 Posterior setae 4 4 10 10 10 7

2002 THE GREAT LAKES ENTOMOLOGIST 13
ACKNOWLEDGEMENTWe thank Dr. Heather Proctor for making this research opportunity possible.
LITERATURE CITEDBernard, E. C. 1990. New species, clarifications, and changes in status within Eosen-
tomon Berlese (Hexapoda: Protura: Eosentomidae) from the United States. Proc. Biol. Soc. Wash. 103:861-890.
Christiansen, K. A. and P. F. Bellinger. 1998. The Collembola of North America North of the Rio Grande. 2nd ed. Grinnell College, Grinnell, IA.
Nosek, J. and D. K. M. Kevan. 1984. New species of Eosentomon Berlese, 1909 (Protura: Eosentomidae) from Quebec and Vermont. Can. Ent. 116:19-26.
Szeptycki, A. 1995. Polish Protura. II. Eosentomon delicatum Gisin, 1945, and related species. Polskie Pismo Ent. 55:139-186.
Szeptycki, A. 1986. Polish Protura. IV. Eosentomon “transitorium” group. Polskie Pismo Ent. 56:481-530.
Tuxen, S. L. 1964. The Protura. Hermann, Paris.

14 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1

2002 THE GREAT LAKES ENTOMOLOGIST 15
ECOLOGICAL NOTES ON SPECIES OF CLERIDAE (INSECTA: COLEOPTERA) ASSOCIATED WITH THE PRAIRIE FLORA OF CENTRAL NORTH AMERICA
Jonathan R. Mawdsley1
ABSTRACTThe association of eighteen species of Cleridae (Coleoptera) with tallgrass
and shortgrass prairie ecosystems in central North America is reported for the first time. New flower visitation, habitat association, distribution, and/or rearing records are reported for Enoclerus analis (LeConte), E. coccineus coc-cineus (Schenkling), E. cordifer (LeConte), E. rosmarus (Say), E. zonatus (Klug), Placopterus thoracicus pallipes (Wolcott), Trichodes bibalteatus LeConte, T. bicinctus Green, T. nutalli (Kirby), Phyllobaenus dubius (Wolcott), P. knausii (Wickham), P. pubescens (LeConte), P. subfasciatus (LeConte), Isohydnocera albocincta (Horn), I. brunnea (Horn), I. curtipennis (Newman), I. tricondylae (LeConte) and Wolcottia pedalis (LeConte). Diagnostic characters are presented to separate adults of P. dubius, a species endemic to the northern shortgrass prairie region, from the common and widespread P. pubescens.
INTRODUCTIONMembers of the beetle family Cleridae (Insecta: Coleoptera) are often con-
sidered forest insects, since many species in this family are predators of bark and wood-boring beetles (Coleoptera: Scolytidae, Buprestidae, Cerambycidae) that feed on trees and woody plants (Balduf 1935). The existence of a group of clerid species that are closely associated with the native prairie or grassland flora in central North America has not been noted prior to the present paper. Clerids are not mentioned in early ecological studies of prairie insects, such as those of Shelford (1913) or Adams (1915), and more recent studies by prairie ecologists and clerid taxonomists have also overlooked the association of clerid species with prairie habitats. The following notes on eighteen species of Cleridae that are undoubtedly associated with prairies and the prairie flora were compiled after field studies in Colorado and Illinois, and the examination of several important North American collections of clerid beetles.
MATERIALS AND METHODSSpecies of Cleridae included in the present report have either been col-
lected at known prairie remnants, or collected in association with plant species typical of tallgrass or shortgrass prairie ecosystems (such as those listed by Sampson 1921, Owensby 1980, Vance et al. 1984, and Runkel 1989). Museum collections of clerid beetles examined for this study include: Cornell University Insect Collection (CUIC), National Museum of Natural History, Smithsonian Institution (NMNH), Snow Entomological Museum, University of Kansas (SEMK); University Museum, University of Colorado at Boulder (UCBC). For the new ecological records, I list the collection in which voucher specimens are deposited, as well as the locality where the voucher specimens were collected.
Information on the geographic distributions of species of Cleridae discussed in this paper can be found in the catalogue of Corporaal (1950) and the checklist of Barr (1975). The Corporaal catalogue also includes citations for anatomical descriptions and illustrations.
1Division of Entomology, Department of Systematic Biology, National Museum of Natu-ral History, Smithsonian Institution, Washington, DC 20560-0187 USA

16 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Botanical nomenclature follows the online databases of the Missouri Bo-tanical Garden (available on the World Wide Web at http://mobot.mobot.org/W3T/Search/vast.html).
The list of species below probably does not include all species of Cleridae found in North American grasslands. Many additional clerid species breed in trees attacked by bark or wood-boring beetles in prairie groves or open savannas, and adults of these species may at times be found on the flowers and foliage of prairie plants. I discuss several additional clerid species that are especially likely to be found at prairie remnants at the end of this paper. There is, at present, no comprehensive guide to the species of Cleridae from North America, and therefore no way to insure accurate identification of all species that might be encountered. The key to genera of Nearctic Cleridae by Barr (1962), the revision of North American Trichodes by Foster (1976), and the review and illustrations of species of Cleridae from Ohio by Knull (1951) will help prairie ecologists and coleopterists identify species of Cleridae found at prairie sites.
ECOLOGY OF PRAIRIE CLERIDS: AN OVERVIEWAdults of the clerid species listed below are found on flowers and foliage in
North American grasslands during spring, summer, and early fall. These adult clerids are predators of small beetles, wasps, flies, true bugs, aphids, and other insects found on vegetation; adults of Trichodes species may also feed on pollen. My field studies and information associated with museum specimens indicate that two of the most common predatory clerid species (Enoclerus rosmarus and Phyllobaenus pubescens) are generalist predators as adults. Other grassland clerid species may be more specialized in their predatory habits. Adults of most of the prairie clerid species can be found on a wide range of plant species, including plants with which their larvae are not known to be associated. Some flower-visiting adult clerids (particularly species in the genera Trichodes and Enoclerus) are densely pubescent and may serve as pollinators of native plant species, much like their relatives in the family Melyridae (Grant and Grant 1965).
For most of the clerid species included here, little is known of the lar-val biology and life history. Published accounts describe portions of the life histories of Enoclerus rosmarus, E. zonatus, Trichodes bibalteatus, T. nutalli, Phyllobaenus pubescens, and Isohydnocera curtipennis (Böving and Champlain 1920, Sabrosky 1934, Foster 1971, 1976). Larvae of these clerids exhibit a wide range of prey preferences: T. nutalli larvae feed on grasshopper egg masses; T. bibalteatus larvae parasitize nests of megachilid bees; while larvae of E. ros-marus, E. zonatus, I. curtipennis, and P. pubescens prey on immature stages of stem-boring and/or gall-forming insects. Most prairie clerids have annual life cycles, with mating and oviposition in spring and early summer, larval devel-opment during summer and fall, pupation either in fall or in early spring, and adult emergence in spring and early summer. Family Cleridae Subfamily Clerinae Enoclerus analis (LeConte)
Notes: This is one of the most poorly known species of Enoclerus in North America, represented in most museum collections by few specimens. Wickham and Wolcott (1912) note that adults have been collected on flowers in grassland areas. Foster and Barr (1972) report the collection of an adult on Melilotus albus Lamarck. The larvae are unknown.
New rearing record: Adult reared from Sporobolus sp. (NM: Mesilla Park; NMNH).

2002 THE GREAT LAKES ENTOMOLOGIST 17
Enoclerus coccineus coccineus (Schenkling)Notes: Adults are usually collected on flowers, where they are quite active
and readily take flight or tumble if disturbed (Wickham and Wolcott 1912). In New Mexico, the larvae are important predators of Crossidius pulchellus LeConte (Coleoptera: Cerambycidae) and other borers attacking roots of Gutierrezia sarothrae (Pursh) Britton and Rusby (see http://taipan.nmsu.edu/snakeweed/bugs.html). A disjunct population of this species from the Colorado Plateau was described by Barr (1976) as E. coccineus desertus Barr.
New flower visitation records: Argemone squarrosa Greene (NM: 20 miles NW Melrose; NMNH), Cleome sp. (WY: New Castle; NMNH), Helianthus annuus L. (TX: 3 miles W Estelline; SEMK), Monarda citriodora Cervantes ex Lagasca y Segura (TX: Devils River; NMNH), Opuntia engelmanni Engelmann (TX: Devils River; NMNH), Opuntia sp. (TX: 5 miles S Kingsville; SEMK), Ratibida columnaris (Sims) D. Don (TX: Devils River; NMNH), Sophia obtuse Greene (NM: 8.5 miles E Batil; NMNH). Enoclerus cordifer (LeConte)
Notes: This is another poorly known species of Enoclerus associated with dry grasslands in central and southwestern North America. LeConte (1849) was the first to publish biological information for this species, stating that adults were collected on the foliage of Artemisia tridentata Nuttall. Wickham and Wolcott (1912) noted that adults are found on flowers. S. P. Cover of the Museum of Comparative Zoology (pers. comm.) reports that adults can also found under objects on the ground in desert grasslands in the mountains of southeastern Arizona. The record below of an adult of this species feeding on eggs of a grass-hopper in the genus Melanoplus (Orthoptera: Acrididae) is the first record of any species of Enoclerus feeding on grasshopper eggs. Species of the clerid genera Trichodes Herbst (Corporaal 1950; Foster 1976) and Aulicus Spinola (Barr and Foster 1979) are also known to feed on grasshopper egg masses.
New predation record: Adult collected while feeding on eggs of Melanoplus sp. (KS: Wichita; NMNH). Enoclerus rosmarus (Say)
Notes: This is one of the most widespread and frequently collected spe-cies of Cleridae in eastern North America. Little is known about its biology, although adults are usually collected on flowers or foliage (Wickham and Wolcott 1912; Knull 1951). Wolcott (1910) reported that adults were collected on foliage in damp meadows (possibly wet prairies) in Indiana. Böving and Champlain (1920) described the mature larva and reared adults from Rhus sp., while Dillon and Dillon (1961) noted that adults were especially abundant on the flowers of Conyza canadensis (L.) Cronquist. Adults from Illinois prairies that I kept in captivity fed readily on small floricolous beetles of the families Curculionidae, Dermestidae, and Mordellidae, as well as aphids, small flies, and small species of Hemiptera. I also observed adults at prairie sites in northern Illinois preying on small braconid wasps.
New flower visitation records: Asclepias syriaca L. (IL: West Chicago Prairie; CUIC), Melilotus officinalis (L.) Pallas (KS: Lawrence; SEMK), Solidago sp. (IL: West Chicago Prairie; NMNH), Tradescantia virginiana L. (IL: West Chicago Prairie; NMNH).
New rearing record: Adult reared from abandoned gall in Solidago sp. (OH: Congress; NMNH). Enoclerus zonatus (Klug)
Notes: Adults are usually found on inflorescences, stems, and leaves of species of the genera Agave, Yucca, or Dasylirion; their larvae feed on weevils and other insects boring in the stems and seed pods of these plants (Foster 1971; Foster and Barr 1972). Adults have also been collected at lights at night.

18 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
New flower visitation records: Asclepias sp. (KS: Medicine Lodge; NMNH), Yucca glauca Nuttall (CO: Boulder; UCBC). Placopterus thoracicus pallipes Wolcott
Notes: The taxonomy of the genus Placopterus is currently under review and this taxon, which is largely allopatric with the nominate form, may repre-sent a distinct species (W. F. Barr, pers. comm.). Although nothing has been published on the biology of P. thoracicus pallipes, the nominate form from eastern North America is a generalist predator associated with a wide range of habitat types and prey species (Böving and Champlain 1920; Foster and Barr 1972).
Distribution: I examined specimens from IA: Lake Okoboji; KS: Atchison, Manhattan, Onaga; MI: Detroit; MO: St. Louis; ND: Bottineau County, Section 3, Township 160; SD: Big Stone City (all NMNH). Trichodes bibalteatus LeConte
Notes: This is the largest species of Trichodes in North America. Adults of species in this Holarctic and African genus occur on flowers, sometimes abun-dantly. As with many Trichodes, larvae of T. bibalteatus are predators in nests of leaf-cutter bees (Hymenoptera: Megachilidae; Foster 1976). The revision of North American Trichodes by Foster (1976) provides additional information on the ecology and distribution of T. bibalteatus and a key to Nearctic species of this genus.
Published flower visitation records: Acacia greggii Gray, Acacia sp., Coreopsis tinctoria Nuttall, Gossypium sp., Helianthus sp., Opuntia sp., Pithe-colobium sp., Prosopis glandulosa Torrey, Ratibida columnaris (Sims) D. Don, Ratibida sp., Rudbeckia sp., Verbesina encelioides (Cavanilles) Gray (Foster 1976).
New flower visitation records: Bidens sp. (TX: 11 miles N Uvalde; SEMK), Condalia obtusifolia (Hooker) Weber (TX: Hidalgo Co., Starr Co.; NMNH), Coreopsis cardeminaefolia (De Candolle) Nuttall (TX: San Antonio; NMNH), Hedeoma sp. (TX: Cotulla; NMNH), Hymenocarpus sp. (TX: near Uvalde; NMNH), Monarda citriodora Cervantes ex Lagasca y Segura (TX: Clarendon; NMNH), Rudbeckia amplexicaulis Vahl (TX: Denton; NMNH), Rudbeckia sp. (TX: Dallas; NMNH). Trichodes bicinctus Green
Published flower visitation records: Ratibida columnaris (Sims) D. Don (Foster 1976).
New flower visitation record: Engelmannia pinnatifida Nuttall (OK: 7 miles SW Lone Wolf; SEMK). Trichodes nutalli (Kirby)
Notes: This species has a widespread but somewhat patchy distribution; adults may occasionally be abundant when they are encountered. Further studies of the biology of T. nutalli are needed; the only known larval host for this species is Chloealtis conspersa (Harris) (Orthoptera: Acrididae) (Foster 1976). Besides prairies, adults of T. nutalli have also been collected on flowers in pine barrens, boreal wetlands, and montane alpine habitats in northeastern North America.
Published flower visitation records: Achillea millefolium L., Apocynum sp., Asclepias sp., Aster sp., Chrysanthemum leucanthemum L., Helianthus sp., Heracleum maximum Bartram, Melilotus albus Lamarck, Melilotus sp., Poten-tilla recta L., Rosa sp., Rubus sp., Rudbeckia hirta L., Spiraea sp. (Foster 1976).
New flower visitation records: Geranium parryi (Engelmann ex A. Gray) A. Heller (CO: Boulder; NMNH), Heracleum lanatum Michaux (CO: Boulder;

2002 THE GREAT LAKES ENTOMOLOGIST 19
NMNH), Solidago sp. (WI: Waupaca; NMNH). Subfamily Hydnocerinae Phyllobaenus dubius (Wolcott)
Diagnosis: Similar to P. pubescens (diagnosed and illustrated by Knull 1951) in coloration and general appearance, but body size usually larger (length 4.6-5.0 mm, compared to 3.3-4.7 mm in P. pubescens), frons and pronotum much more coarsely and densely punctate, and elytra proportionately longer and more slender.
Known distribution: Iowa and Nebraska (Barr 1975); also Montana and Colorado (reported here for the first time).
Notes: Based on the few collection records published to date, this species appears to be endemic to the northern shortgrass prairie region of central North America. Further investigations of this species’ distribution and ecology are needed.
Distribution: CO: Boulder Co., Eldorado Open Space, 1967 m, 1.X.1996, malaise trap, creekside (1, UCBC); Jefferson Co., Doudy Draw Open Space, 27.VI.1994, malaise trap (1, UCBC). MT: Roosevelt Co., 15 miles NE Bainville, 12.VI.1996, sweeping vegetation in marsh (1, CUIC); no locality specified (1, NMNH). Phyllobaenus knausii (Wickham)
Notes: The bright reddish-orange, black, and white coloration of this spe-cies is unusual in the genus Phyllobaenus, but is similar to the coloration of P. tricolor (Schaeffer). The elytra of P. tricolor have a median white transverse band of hairs that is lacking in P. knausii.
New flower visitation record: Sapindus drummondii Hooker and Arnott (KS: Meade Co.; SEMK). Phyllobaenus pubescens (LeConte)
Notes: Based on the large number of specimens examined by the author, this species appears to be one of the most common and abundant clerid species associated with the prairie flora of North America. Adults and larvae have been previously recorded as predators of cotton boll weevils (Coleoptera: Cur-culionidae: Anthonomus spp.), gall-forming insects, and cocoons of species in the wasp genus Microbracon Ashmead (Hymenoptera: Braconidae) (Corporaal 1950; Knull 1951). More recent studies indicate that P. pubescens may be an important predator of insect pests in commerical cotton and sunflower planta-tions. Rogers et al. (1986) reported that larvae of this species were predators of the larvae of Stibadium spumosum Grote (Lepidoptera: Noctuidae), a pest of commercial sunflower plantations in the south-central USA, while Farrar (1985) reported that adults and larvae of P. pubescens were predators of Heliothis zea (Boddie) (Lepidoptera: Noctuidae), a pest of cotton.
New flower visitation records: Amorpha fruticosa Torrey (TX: Dallas; NMNH), Asclepias latifolia (Torrey) Rafinesque (TX: Dallas; NMNH), Aster tanacetifolius Kunth (TX: Raymondsville, 5 miles SW Tivoli; SEMK), “cactus” (TX: 19 miles N Tivoli; SEMK), Callirhoe involucrata (Torrey and A. Gray) Tor-rey (TX: Victoria; NMNH), Cassia sp. (TX: Victoria; NMNH), Cicuta maculata L. (KS: Lone Star Lake; SEMK), Croton punctatus Jacquin (TX: Cameron Co.; NMNH), Gaillardia sp. (TX: Boca Chica, McDade; SEMK), Helianthus annuus L. (wild, not cultivated) (IL: Monsanto; MO: Matthews, New Madrid, Porta-geville; TX: 6 miles N Raymondsville, 29 miles S Sarita; NMNH), Helianthus petiolaris Nuttall (KS: Lewis; SEMK), Helianthus sp. (TX: Victoria; NMNH), Malvastrum apicatum (L.) Gray (TX: Cameron Co.; NMNH), Melilotus offici-nalis (L.) Pallas (KS: Arkansas City; SEMK), Monarda citriodora Cervantes ex Lagasca y Segura (TX: Raymondsville, Wolfe City; SEMK), Monarda pectinata

20 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Nuttall (TX: Matagorda; SEMK), Monarda punctata L. (TX: 17 miles N Vernon; SEMK), Opuntia sp. (TX: Victoria; NMNH), Parthenium hysterophorus L. (TX: Hidalgo Co.; NMNH), Polytaenia nuttalli De Candolle (OK: 2 miles S Jennings; SEMK), Quamasia sp. (TX: Denton; NMNH), Rudbeckia amplexicaulis Vahl (TX: Victoria; NMNH), Rudbeckia grandiflora (Sweet) Gmelin ex De Candolle (TX: Dallas; NMNH), Rudbeckia hirta L. (KS: 9 miles N Caldwell; SEMK), Rudbeckia sp. (OK: Ardmore; SEMK), Sideranthus rubiginosus (Torrey and A. Gray) Britton ex Rydberg (OK: Ardmore; NMNH), Vernonia interior Smith (KS: Manhattan; NMNH).
New rearing records: Adult reared from trap-nest of Megachile brevis Say (KS: Johnson Co.; NMNH), adult reared from stem of Iva ciliata Willdenow (TX: Victoria; NMNH), adult reared from flower head of Rudbeckia amplexicaulis Vahl (TX: Dallas; NMNH). Phyllobaenus subfasciatus (LeConte)
Notes: This species belongs to a taxonomically difficult group in the ge-nus Phyllobaenus DeJean that is greatly in need of revision. Another valid, described member of this group associated with grassland communities is P. maritimus (Wolcott) from the Atlantic coast of northeastern North America. All species in this group are metallic coppery-black or bluish-black in color and have rugosely punctate elytra with two transverse bands of pale pubescence, one at mid-elytron, the other at apical third or fourth.
New flower visitation records: Amorpha canescens Nuttall (KS: 3 miles S Sawyer; SEMK), Helianthus petiolaris Nuttall (KS: 5 miles N Bennington; SEMK), Sapindus drummondii Hooker and Arnott (KS: 8 miles W Hardtner; SEMK). Isohydnocera albocincta (Horn)
Notes: Nothing has been published on this species’ biology.New habitat association: “Prairie vegetation” (TX: 2 miles E Reisel;
NMNH). Isohydnocera brunnea (Horn)
Notes: Nothing has been published on this species’ biology.New flower visitation record: Echinacea angustifolia De Candolle (KS:
Hutchinson; SEMK). Isohydnocera curtipennis (Newman)
Notes: Sabrosky (1934) provides a detailed account of the biology of this species. Larvae prey on gall-making insects associated with plants of the genus Solidago; the adults are found on flowers and stems of the host plants and other vegetation. Three voucher specimens from Sabrosky’s study are in NMNH. Isohydnocera tricondylae (LeConte)
Notes: According to W. F. Barr (pers. comm.), this species is associated with prairies in central North America. Nothing has been published about its biology.
Distribution: IA: Ames; KS: Manhattan, Onaga, Topeka (all NMNH). Wolcottia pedalis (LeConte)
Notes: Nothing has been published about the biology of this species. S. W. Lingafelter (pers. comm.) indicates that it is common at tallgrass prairie sites in Kansas, where adults occur on flowers and foliage of many different plant species.
New habitat association: Tallgrass prairie (KS: Konza Tallgrass Prairie; NMNH).
Additional species of Cleridae associated with prairies and grasslands.As noted above, other species of Cleridae will probably be recorded from

2002 THE GREAT LAKES ENTOMOLOGIST 21
Nearctic grasslands in the future. In particular, five species of the subfamily Hydnocerinae probably also occur in savanna or prairie ecosystems. Phyllobae-nus pallipennis (Say), P. unifasciatus (Say), P. verticalis (Say), and P. humeralis (Say) breed in trunks and limbs of oaks (Quercus spp.) and hickories (Carya spp.), but adults may occur on flowers and foliage of nearby herbaceous plants. Knull (1951) provided illustrations and diagnoses of these species, all of which can be readily separated from the other representatives of this genus discussed above. One other Nearctic clerid that is probably found in prairie habitats is Isohydnocera tabida (LeConte). Little is known about the biology of this species, which was also figured and diagnosed by Knull (1951).
W. F. Barr (pers. comm.) notes the presence of a complex of clerid species associated with desert grasslands in the southwestern United States. Although these species fall outside the geographic scope of this paper, I mention them here as evidence that the association of clerids with grassland habitats is not restricted to central North America. Clerid species associated with desert grass-lands in the southwestern U. S. include Aulicus dentipes Schaeffer, A. nigriven-tris Schaefer, Phyllobaenus longus (LeConte), P. rudis (Gorham), P. wickhami (Wolcott), Isohydnocera chiricahuana Knull, and Wolcottia sobrina (Fall).
ACKNOWLEDGMENTSSupport for the author was provided by a Smithsonian Institution Postdoc-
toral Research Fellowship and Research Associateship, sponsored by T. L. Erwin. Funding for fieldwork associated with this project was provided in part by a grant from the Theodore Roosevelt Memorial Fund at the American Museum of Natural History. C. M. Carter, A. S. Mawdsley, J. L. Mawdsley, R. D. Mawdsley, J. Shoger, and M. Shoger are to be thanked for providing assistance with field studies of clerids at prairie sites. S. P. Cover of Harvard University provided information on the biology of Enoclerus cordifer, while S. W. Lingafelter provided information on the biology of Wolcottia pedalis. I would like to thank E. R. Hoebeke and J. K. Liebherr (CUIC), N. J. Vandenberg (NMNH), R. W. Brooks (SEMK), and V. Scott (UCBC) for permitting me to examine specimens in their care. Finally, W. F. Barr provided significant information on grassland clerids as well as helpful comments on an earlier draft of the manuscript.
LITERATURE CITEDAdams, C. C. 1915. An ecological study of prairie and forest invertebrates. Publisher not
indicated, Urbana, Illinois. 280 pp.Balduf, W. V. 1935. The bionomics of entomophagous Coleoptera. John S. Swift, St.
Louis. 220 pp.Barr, W. F. 1962. A key to the genera and a classification of the North American Cleridae
(Coleoptera). Coleopterists’ Bulletin 16:121-127.Barr, W. F. 1975. Cleridae. North American Beetle Fauna Project 73:1-18.Barr, W. F. 1976. Descriptions and taxonomic notes on Enoclerus and some allied genera
(Coleoptera: Cleridae). Melanderia 24:18-35.Barr, W. F. and Foster, D. E. 1979. Revision of the genus Aulicus (Coleoptera: Cleri-
dae). Melanderia 33:1-31.Böving, A. G. and A. B. Champlain. 1920. Larvae of North American beetles of the family
Cleridae. Proceedings of the United States National Museum 57:575-649 + pls. 42-53.Corporaal, J. B. 1950. Coleopterorum Catalogus, Pars 23 (Editio Secunda), Cleridae. W.
Junk, The Hague. 373 pp.Dillon, L. S. and Dillon, E. S. 1961. A manual of common beetles of eastern North America.
Row, Peterson, Evanston, Illinois. viii + 824 pp.

22 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Farrar, R. R. 1985. A new record of the clerid, Phyllobaenus pubescens (LeConte) (Cole-optera), as a predator of Heliothis zea (Boddie) (Lepidoptera: Noctuidae) on cotton. Coleopterists’ Bulletin 39(1):33-34.
Foster, D. E. 1971. Observations on the biologies of Enoclerus abdominalis (Chevrolat) and Enoclerus opifex (Gorham) (Coleoptera: Cleridae). Coleopterists’ Bulletin 25:127-129.
Foster, D. E. and Barr, W. F. 1972. Notes on the distribution and bionomics of some North American Cleridae. Journal of the Kansas Entomological Society 45(1):122-125.
Foster, D. E. 1976. Revision of North American Trichodes (Herbst) (Coleoptera: Cleridae). Special Publications of the Museum, Texas Tech University 11:1-86.
Grant, V. and Grant, K. A. 1965. Flower pollination in the phlox family. Columbia Uni-versity Press, New York. xi + 180 pp.
Knull, J. N. 1951. The checkered beetles of Ohio (Coleoptera: Cleridae). Ohio Biological Survey Bulletin 8(2):269-350.
LeConte, J. L. 1849. Synopsis of the coleopterous insects of the group Cleridae which in-habit the United States. Annals of the Lyceum of Natural History of New York 5:9-35.
Owensby, C. E. 1980. Kansas prairie wildflowers. Iowa State University Press, Ames, Iowa. vi + 124 pp.
Rogers, C. E., Perkins, W. D. and Seiler, G. T. 1986. Bionomics of Stibadium spu-mosum (Lepidoptera: Noctuidae), a pest of sunflower in the southern plains. Environmental Entomology 15(6):1275-1278.
Runkel, S. T. 1989. Wildflowers of the tallgrass prairie: the upper midwest. Iowa State University Press, Ames, Iowa. xii + 279 pp.
Sabrosky, C. W. 1934. Notes on the larvae and larval habit of Isohydnocera curtipennis (Newmn.) (Coleoptera: Cleridae). Journal of the Kansas Entomological Society 7(2):65-68.
Sampson, H. C. 1921. An ecological survey of the prairie vegetation of Illinois. Publisher not indicated, Urbana, Illinois, 55 pp.
Shelford, V. E. 1913. Animal communities in temperate America, as illustrated in the Chicago region: A study in animal ecology. University of Chicago Press, Chicago. xiii + 368 pp.
Vance, F. R., Jowsey, J. R. and J. S. McLean, J. S. 1984. Wildflowers of the northern Great Plains. University of Minnesota Press, Minneapolis. 336 pp.
Wickham, H. F. and Wolcott, A. B. 1912. Notes on Cleridae from North and Central America. Bulletin from the Laboratories of Natural History, State University of Iowa 6(3):49-67.
Wolcott, A. B. 1910. Family XLII, Cleridae, The Checkered Beetles. Indiana Department of Geology and Natural Resources Bulletin 1:846-862.

2002 THE GREAT LAKES ENTOMOLOGIST 23
SIZE ESTIMATE OF A LOCAL POPULATION OF BRYCHIUS HUNGERFORDI (COLEOPTERA: HALIPLIDAE)
Michael Grant 1, Brian Scholtens 2, Robert Vande Kopple 1, and Bert Ebbers 3
ABSTRACTMuch of the natural history of the endangered crawling water beetle,
Brychius hungerfordi, is unknown. Population estimates have only been sur-mised. We have measured, using a mark and recapture technique, a localized population of the beetle in a pool on the East Branch of the Maple River, Em-met County, Michigan. The average population during July 2001 was found to be 1052 individuals with a range of 43 (highest estimate – lowest estimate). A three-year study of the relative abundance of the beetle in the same pool sug-gests that the population has remained relatively constant.
____________________The Federally endangered crawling water beetle, Brychius hungerfordi
(Spangler), is the only Brychius species in eastern North America (Strand et al. 1994). It is known from only four sites in Northern Michigan and one in Ontario (Grant et al. 2000 and references therein). Efforts to locate the beetle in the Upper Peninsula of Michigan, Minnesota and Wisconsin have been unsuccessful (Hilsenhoff and Brigham 1978, Brigham 1982, White 1986).
A literature search produced only one reference to a population estimate for the beetle. A visual population estimate at the type locale of 200-500 individuals was reported by D. S. White (pers. comm.). He noted that other areas likely con-tained fewer numbers.
We report on the use of a mark and recapture technique for estimating a Brychius hungerfordi population in a pool located approximately 2.5 km up-stream from the type locale.
MATERIALS AND METHODSThe study site is a pool approximately 219 m 2 in area located on the East
Branch of the Maple River in Emmet County, Michigan. Beetles were collected using a long-handled D-net (30-cm diameter) from around the perimeter of the pool to a depth of approximately 1.5 m. A uniform sweeping motion with the lip of the net held slightly above the substrate produced strong turbulence that dislodged beetles from the bottom. Quickly returning the net over the same area captured them. The net contents were emptied into a white enamel pan and sorted.
The mark and recapture experiment was conducted from 13-19 July 2001. The beetles were dried by placing them on a disk of filter paper in the bottom of a small petri dish. Once the beetle appeared dry, the petri dish was placed onto the stage of a dissecting microscope and a small dot of Testors paint applied to the elytra using a 26-gauge needle fitted onto a syringe. The paint was allowed to dry for several minutes before the marked beetle was returned to the storage bucket. Captured beetles were re-distributed into the pool at the sampling locations from which they were removed, in approximately the same proportion as they were found. The marked beetles did not exhibit any unusual behavior during their time 1 The University of Michigan Biological Station, Pellston 49769.2 Department of Biology, College of Charleston, Charleston, South Carolina 29424-0001.3 Great Lakes Ecosystems, 4989 E. Burt Lake Rd., Indian River, Michigan 49749.

24 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
in captivity. Beetles captured on the first sampling day (13 July) were marked with white paint and those on the second sampling day (16 July) with orange. Beetles captured on the final day (19 July) were not marked. Preliminary experi-ments using Peltodytes in an aquarium indicated that a small drop of Testors paint placed on a dry beetle would adhere to the elytra for about seven days. We used both the Lincoln index and the Bailey Triple-Catch methods of estimating population size. Details of the techniques are described in Southwood (1978).
Seasonal relative abundance estimates were determined monthly through-out the spring, summer and fall over a three-year period, from 1999 through 2001. During each sampling interval, the pool was systematically searched for exactly 60 minutes. Relative abundance is expressed as the number of beetles captured per hour.
Assuming relative abundance is proportional to population size, a constant of proportionality can be found by dividing the population estimate, determined using the mark and recapture technique, by the relative abundance data col-lected during the same time period. Since the sampling effort and collecting area was standardized for all monthly abundance measurements, multiplying abundance estimates by the calibration factor results in a population estimate for the given time period.
RESULTS AND DISCUSSIONResults for the mark and recapture experiment are shown in Table 1.
The Lincoln Index was calculated once using data from days one and two, and a second time using data from days two and three; Bailey’s Triple Catch Method was used for samples collected on day three. All three techniques gave good agreement with an average of 1052 individuals and range of 43 (highest estimate – lowest estimate).
Assumptions in the model are that the population is closed, i.e., no immi-gration or emigration occurs and that there were no births or mortalities during the sampling period. Occasional searches of the river immediately downstream from the pool failed to produce any marked beetles and upstream culverts blocked passage. Given the relatively short time frame of the experiment, it is unlikely that births or deaths significantly affected results.
The relative abundance measurements are shown in Figure 1. Converting the maximum relative abundance measurement for each year into a population estimate shows no clear trend in the beetle population in the pool. During the three years surveyed, peak populations occurred during different seasons but remained fairly constant at around 1100 individuals.
This work represents the first time a Brychius hungerfordi population has been estimated using generally accepted measurement techniques. Although only one pool on the East Branch of the Maple was surveyed, it appears that
Table 1. Sampling Data From Mark-Recapture Experiment, 13-19 July 2001
Day No. Marked (Color) No. Recaptured % Recaptured And Released Containing Marks (Color) 1 81 (White) 0 — 2 101 (Orange) 8 (White) 9.9 3 0 8 (White) and 9 (Orange) 16.8
First Estimate (Lincoln Index) = 1023Second Estimate (Lincoln Index) = 1066Third Estimate (Bailey’s Triple Catch Method) = 1066Average 1052 Range 43 (highest estimate- lowest estimate)

2002 THE GREAT LAKES ENTOMOLOGIST 25
Figure 1. Seasonal Relative Abundance Of Brychius hungerfordi Study Pool on East Branch of Maple River, Emmet County, Michigan.

26 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
the beetle population in this river may be larger than originally estimated and the population relatively stable.
ACKNOWLEDGEMENTSThe authors would like to thank the Department of the Interior, U.S.
Fish and Wildlife Service and the Michigan Department of Natural Resources, Wildlife Division, for providing sampling permits PRT 839782 and PN 1304 respectively. We also appreciate the comments from reviewers whose sugges-tions improved this paper.
LITERATURE CITEDBrigham, W.U. 1982. Aquatic Coleoptera, p 10.1-10.136. In: Brigham, R., W. U. Brigham
and A. Grilka, (eds.). Aquatic insects and oligochaets of North and South Carolina. Midwest aquatic entomologist. Mahomet, IL.
Grant, M.E., R. Vande Kopple and B. Ebbers. 2000. New distribution record for the en-dangered crawling water beetle Brychius hungerfordi (Coleoptera: Haliplidae) and notes on seasonal abundance and food preferences. Great Lakes Entomol. 33: 165-168.
Hilsenhoff, W.L. and W.U. Brigham. 1978. Crawling water beetles of Wisconsin (Coleop-tera: Haliplidae). Great Lakes Entomol. 11(1): 11-12.
Southwood, T. R. E. 1978. Ecological methods with particular reference to the study of insect populations, p. 92-105. Chapman and Hall, London.
Strand, R.M. and P.J. Spangler. 1994. The natural history, distribution, and larval description of Brychius hungerfordi (Coleoptera: Haliplidae). Proc. Entomol. Soc. Wash. 96(2): 208-213.
White, D.S. 1986. The Status of Brychius hungerfordi and Stenelmis douglasensis in Michigan. A report to The Nature Conservancy, Michigan Field Office, Small Grants Program; 8 pp.

2002 THE GREAT LAKES ENTOMOLOGIST 27
ANT-PLANT RELATIONSHIPS IN A RE-CREATED TALLGRASS PRAIRIE
Chris E. Petersen1, Dhvani G. Dave1, Charise L. Parker1, and Renee E. Petersen1
ABSTRACTRelationships between the distributions of nesting ants and flora were
investigated in a re-created tallgrass prairie located in northeastern Illinois. One-third of 167 randomly selected 1m2 quadrats in the prairie contained ant nests. Basal coverage of ground by tall grass shoots was greater in quadrats having ant nests than in those lacking ant nests. In addition, clumps of big bluestem (Andropogon gerardii) that were inhabited by the ant, Acanthomyops clavigera, had higher median rank densities than adjacent clumps lacking nests. The clumps also showed a negative correlation between big bluestem shoots/cm2 and area occupied by the clump at ground level. These significant findings relating to the distribution of nesting ants and tall grasses indicate ants have ecologic importance in re-created prairie that warrants greater scrutiny among restoration scientists.
____________________Ants (Formicidae) have profound effects on most terrestrial systems as soil
movers (Baxter and Hole 1967), modifiers of the physical and chemical proper-ties of soil in which they nest (Baxter and Hole 1967, Culver and Beattie 1983, Lesica and Kannowski 1998, Newman and Wolff 1990), consumers (Soule and Knapp 1996), protectors of certain plants and herbivorous insects (Hammond 1995, Huxley 1980), and seed dispersal agents (Handel et al. 1981). Through these effects, ants influence the composition of and succession within plant communities (Beattie and Culver 1977, Rissing 1986). Despite their recognized importance in natural systems, the ecological importance of ants in restored systems has received little attention.
Restoration offers to conserve biodiversity by the recovery of degraded land through the reintroduction of its former components i.e., species and ecologi-cal processes (Dobson et al. 1997). Too often, the term “restoration” is used to describe re-creation projects, in which the original substrate has been destroyed and recovery of the natural system must begin anew. In DuPage County, Illinois, the location of the following study, re-creation projects have offered to preserve biodiversity of tallgrass prairie that once covered much of the county. Today, only 3.5 hectares of the original 74,000 hectares of tallgrass prairie remains in the county (IDENR 1994), similar to the demise that has occurred throughout the historic range of the ecosystem (Samson and Knopf 1994). With the amount of remnant prairie being so small, species preservation has undoubtedly been compromised giving significance to re-creation efforts. However, these efforts have been biased to recovery of tallgrass prairie vegetation. This is unfortunate as the establishment and successional change of tallgrass prairie vegetation is influenced by other organisms such as ants (Howe 1994, IDENR 1994, Ham-mond 1995, Newman and Wolff 1990).
Over 100 species of ants have been found in tallgrass prairie (Trager 1998). Ants are so abundant in the prairie that they can surpass grasshoppers in total insect biomass. Their presence in restored and re-created sites is apparent in the form of visual observation of individuals and mounds even within a few
1Natural Sciences Division, College of DuPage, 425 Fawell Blvd., Glen Ellyn, IL 60187

28 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
years of initial recovery (Petersen et al. 1998, Trager 1990). The objective of this study was to explore relationships between the distributions of nesting ants and tallgrass prairie flora. Our study was intended to be preliminary in scope and provide a baseline for more discriminating studies in the future.
MATERIALS AND METHODSThe study site was the Russell Kirt Tallgrass Prairie located on the campus
of College of DuPage, Illinois. Re-creation efforts of the 7.5-hectare preserve began in 1984 with seed broadcasting and seedling transplant on top of clay and construction debris deposited from campus construction. The flora consists of some 150 species that are characteristic of the mesic prairie and wetlands that once covered the area (Swink and Wilhelm 1994). Grasses most frequently encountered at this site are big bluestem (Andropogon gerardii Vitman), In-dian grass (Sorghastrum nutans (L.) Nash), and prairie dropseed (Sporobolus heterolepis Gray). Kirt (1989) provides a comprehensive listing of plants found within the preserve that has been burned each spring.
From June 14 until July 8, 2001, the area covered by plant shoots at ground level and the presence of ant nests were recorded from each of 137 1-m2 square quadrats taken along randomly selected transects. Three to five evenly spaced quadrats were sampled along each of the 28 transects that ranged from 10m to 30m in length. A quadrat was only visited once. Mann-Whitney tests were used to compare surface coverage of grasses, forbs, and all plants (grasses and forbs) within quadrats having ant nests to those without. Significant differences (α< 0.05) in median rank contribution were expected where interactions in some form occur between nesting ants and plants growing upon and immediately around the nests.
Relationships between plant distribution and nesting ants were also ex-plored with two mound-building species, Acanthomyops clavigera (Roger) and Formica nitidiventris Emery. The nests of these species were integrated with and inseparable from clumps of big bluestem in which the nests were located. Clumps were distinguished as aggregates of tall grasses separated by at least 20 cm of bare substrate from other clumps. The clumps of big bluestem inhabited by these ants were distinguished by the added height (up to several centime-ters) and loose texture of the soil as compared to clumps lacking ant nests. The number of shoots of big bluestem projecting from a clump, and also the area of the clump, were measured in each ant-inhabited clump and the closest clump of big bluestem without ant nests. It was assumed one shoot emerging from the ground was one individual, although perhaps a genetic clone of others in the clump. Wilcoxon paired-sample tests were used to compare median ranks of big bluestem counts/cm2 between clumps having nests and those without. Again, significant statistical results were expected where ant-big bluestem in-teractions occur. Spearman rank correlation was used to test for relationships between clump area and the count of big bluestem shoots/cm2. Big bluestem is known to self-shade (Jurik and Kliebenstein 2000), leading to the expectation that clump area would be negatively correlated to density. Our interest was how the presence of ant nests affects the correlation.
RESULTSNests of six ant species were found in a third of the 137 quadrats (Table 1).
Nests of Lasius alienus (Foerster) were the most common, being either located in close proximity of one another (within centimeters) or possessing multiple entrances spread over several meters. L. alienus was not found to nest within clumps of vegetation, but only on barren ground. In contrast, nests of the next most common species, A. clavigera and F. nitidiventris, were found within clumps of big bluestem, Indian grass, and prairie dropseed. Coverage of ground
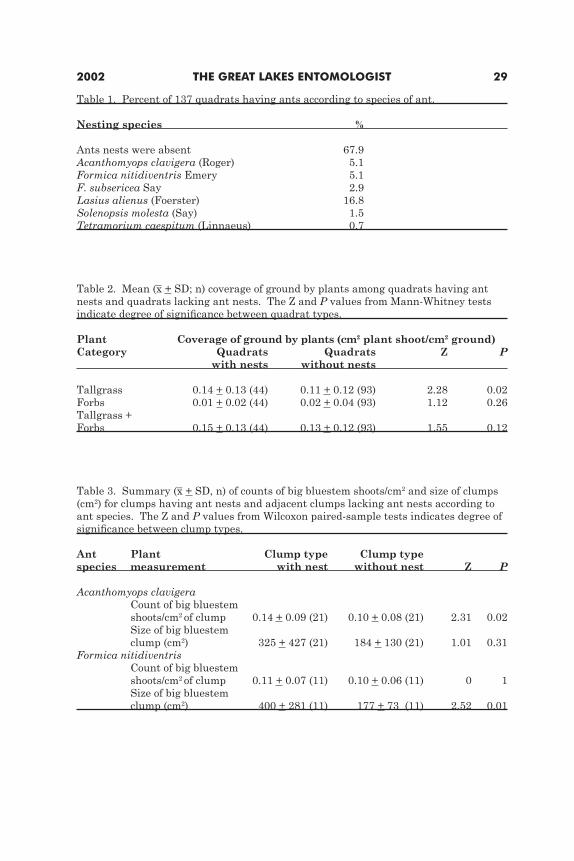
2002 THE GREAT LAKES ENTOMOLOGIST 29
Table 1. Percent of 137 quadrats having ants according to species of ant.
Nesting species %
Ants nests were absent 67.9Acanthomyops clavigera (Roger) 5.1Formica nitidiventris Emery 5.1F. subsericea Say 2.9Lasius alienus (Foerster) 16.8Solenopsis molesta (Say) 1.5Tetramorium caespitum (Linnaeus) 0.7
Table 2. Mean (_x + SD; n) coverage of ground by plants among quadrats having ant nests and quadrats lacking ant nests. The Z and P values from Mann-Whitney tests indicate degree of significance between quadrat types.
Plant Coverage of ground by plants (cm2 plant shoot/cm2 ground)Category Quadrats Quadrats Z P with nests without nests
Tallgrass 0.14 + 0.13 (44) 0.11 + 0.12 (93) 2.28 0.02Forbs 0.01 + 0.02 (44) 0.02 + 0.04 (93) 1.12 0.26Tallgrass +Forbs 0.15 + 0.13 (44) 0.13 + 0.12 (93) 1.55 0.12
Table 3. Summary (_x + SD, n) of counts of big bluestem shoots/cm2 and size of clumps (cm2) for clumps having ant nests and adjacent clumps lacking ant nests according to ant species. The Z and P values from Wilcoxon paired-sample tests indicates degree of significance between clump types.
Ant Plant Clump type Clump typespecies measurement with nest without nest Z P
Acanthomyops clavigera Count of big bluestem shoots/cm2 of clump 0.14 + 0.09 (21) 0.10 + 0.08 (21) 2.31 0.02 Size of big bluestem clump (cm2) 325 + 427 (21) 184 + 130 (21) 1.01 0.31Formica nitidiventris Count of big bluestem shoots/cm2 of clump 0.11 + 0.07 (11) 0.10 + 0.06 (11) 0 1 Size of big bluestem clump (cm2) 400 + 281 (11) 177 + 73 (11) 2.52 0.01

30 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
by these grasses was highest in plants having ant nests (Table 2). The median rank coverage of forbs which included lead plant (Amorpha canescens Pursh), white and red prairie clover (Dalea candida Michx. ex Willd. and D. purpurea Vent.), rattlesnake master (Eryngium yuccifolium Michx.), and beebalm (Mo-narda fistulosa L.), were not significantly different between quadrats having nests and those without.
Big bluestem clumps having nests of A. clavigera had significantly higher median rank counts of shoots/cm2 than clumps lacking nests (Table 3). No dif-ference in shoot counts/cm2 was observed between clumps having nests of F. nitidiventris and those without. The area covered by big bluestem clumps tended to be larger when ant nests were present, and was significantly larger where F. nitidiventris nested. A significant negative relationship between counts of big bluestem shoots/cm2 and clump area was found where A. clavigera nested (Table 4). In the same type of analysis, but with F. nitidiventris, significance was just missed.
DISCUSSIONAll of the ant species observed in this study are widely distributed in
North America and are known from a variety of habitats to include oak and pine forests, pasture, and natural tallgrass prairie (Gregg 1944, Talbot 1934, Trager 1998). The feeding habits of A. clavigera, F. nitidiventris, F. subsericea Say, and L. alienus include scavenging for dead macroinvertebrates and tend-ing aphids (Burrill and Smith 1919, Greg 1944, Ross et al. 1971, Wheeler and Wheeler 1986). A. clavigera in particular is inclined to remain subterranean, tending and, at times, consuming root feeding aphids (Trager 1998). Solenopsis molesta (Say) and Tetramorium caespitum (Linnaeus) are scavengers and seed predators (Ross et al. 1971, Trager 1998). S. molesta is also known to live within mounds of larger ant species where it feeds on discarded matter and immature stages of host ants.
Greater coverage of ground by tall grasses where ant nests occur and the higher density of big bluestem shoots in clumps having nests of A. clavigera are evidence of a positive relationship between grasses and ants in the 16-year old re-created prairie. However, it remains unknown whether and how the tall grass or the ant community promotes the other. Nesting ants may loosen soil, promoting aeration, drainage, and thus tall grass growth. Seed predation and aphid tending by ant colonies may also function to restrict tall grass competi-tors, allowing greater tall grass growth in clumps. Alternatively, tall grass may provide food to nesting ants in the form of tissues and exudates via aphids. Growth may be limited in larger clumps of big bluestem due to self-shading and greater attraction of root-feeding aphids, accounting for the negative correlation between clump area and shoot density.
Table 4. Spearman Rank Order Correlation analysis of big bluestem clump area (cm2) and counts of big bluestem shoots/cm2 according to the presence of ant nests.
Ant species R T P N
Acanthomyops clavigera Clumps with nest -0.59 3.17 <0.01 21 Clumps without nests -0.29 1.31 0.21 21Formica nitidiventris Clumps with nests -0.56 2.05 0.07 11 Clumps without nests -0.22 0.66 0.52 11

2002 THE GREAT LAKES ENTOMOLOGIST 31
Clearly, more study is needed to scrutinize these hypotheses, including information about the presence and abundance of root-feeding aphids. Find-ings of significant relationships between the distribution of nesting ants and tall grasses indicate ants have ecologic importance in re-created prairie that may affect the process and outcome of floral establishment. Long-term studies should help identify patterns in ant-plant interactions, including the perennial nature of interactions within particular tall grass clumps.
ACKNOWLEDGEMENTSThe research was supported by a Tellabs Grant. Appreciation is extended
to B. Petersen, L. Randa, and J. Sutherland for their valuable comments in preparation of this manuscript.
LITERATURE CITEDBaxter, F. P. and F. D. Hole. 1967. Ant (Formica cinerea) pedoturbation in a prairie soil.
Soil Sci. Soc. Amer. Proc. 31:425-428.Beattie, A. J. and D. C. Culver. 1977. Effects of the mound nests of the ant, Formica
obscuripes, on the surrounding vegetation. Am. Midl. Nat. 97:390-399.Burrill, A. C. and M. R. Smith. 1919. A key to the species of Wisconsin ants, with notes
on their habits. Ohio J. Sci. 19:279-292.Culver, D. C. and A. J. Beattie. 1983. Effects of ant mounds on soil chemistry and vegeta-
tion patterns in a Colorado montane meadow. Ecology 64:485-492.Dobson, A. P., A. D. Bradshaw, and A. J. M. Baker. 1997. Hopes for the future: restora-
tion ecology and conservation biology. Science 277:515-522.Gregg, R. E. 1944. The ants of the Chicago region. Ann. Entomol. Soc. Amer. 37:447-480.Hammond, P. C. 1995. Conservation of biodiversity in native prairie communities in the
United States. J. Kansas Entomol. Soc. 68:1-6.Handel, S. N., S. B. Fisch, and G. E. Schatz. 1981. Ants disperse a majority of herbs in
a mesic forest community in New York State. Bull. Torrey Bot. Club 108:430-437. Howe, H. F. 1994. Managing species diversity in tallgrass prairie: Assumptions and
implications. Cons. Biol. 8:691-704.Huxley, C. R. 1980. Symbiosis between ants and epiphytes. Biol. Rev. (Camb.) 55:321-
340. Illinois Department of Energy and Natural Resources (IDENR). 1994. The Changing Illinois Environment: Critical Trends Technical Report of the Critical Trends Assessment Project Volume 3: Ecological Resources. IDENR, Springfield, IL.
Jurik, T. W. and H. Kliebenstein. 2000. Canopy architecture, light extinction and self-shading of a prairie grass, Andropogon gerardii. Am. Midl. Nat. 144:51-65.
Kirt, R. R. 1989. Prairie Plants of Northern Illinois: Identification and Ecology. Stipes Publishing Company, Champaign, IL.
Lesica, P. and P. B. Kannowski. 1998. Ants create hummocks and alter structure and vegetation of a Montana fen. Amer. Midl. Nat. 139:58-68.
Newman, L. M. and R. J. Wolff. 1990. Ants of a northern Illinois savanna and degraded avanna woodland, pp. 71-73. In: D. D. Smith and C. A Jacobs (ed.), Proceedings of the Twelfth North America Prairie Conference. University of Northern Iowa, Cedar Rapids, IO.
Petersen, C. E., K. Zwolfer, and J. Fradkin. 1998. Ant fauna of reconstructed tallgrass Prairie in northeastern Illinois. Trans. Ill. State Acad. Sci. 91:85-90.
Rissing, S. W. 1986. Indirect effects of granivory by harvester ants: plants species com-position and reproductive increase near ant nests. Oecologia 68:231-234.

32 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Ross, H. H. , G. L. Rotramel, and W. E. LaBerge. 1971. A synopsis of common and economic Illinois ants, with keys to the genera (Hymenoptera: Formicidae). Illinois Nat. Hist. Surv. Bull. 71:1-22.
Samson, F. and F. Knopf. 1994. Prairie conservation in North America. BioScience 44: 418-421.
Soule, P. T. and P. A. Knapp. 1996. The influence of vegetation removal by Western Har-vester ants (Pogonomyrmex osyheei) in a relict area of sagebrush-steppe in Central Oregon. Am. Midl. Nat. 136:336-345.
Swink, R. and G. Wilhelm. 1994. Plants of the Chicago Region, 4th ed. The Morton Arboretum, Lisle, IL.
Talbot, M. 1934. Distribution of ant species in the Chicago region with reference to ecological factors and physiological toleration. Ecology 15:416-439.
Trager, J. C. 1990. Restored prairies colonized by native prairie ants (Missouri, Illinois). Restoration and Management Notes 8:104-105.
Trager, J. C. 1998. An introduction to ants (Formicidae) of the tallgrass prairie. Missouri Prairie Journal 18:4-8.
Wheeler, G. and J. Wheeler. 1986. The ants of Nevada. Nat. Hist. Mus. Los Angeles, CA.

2002 THE GREAT LAKES ENTOMOLOGIST 33
1 Department of Entomology, University of Wisconsin-Madison, 445 Russell Laborato-ries, Madison, WI 53706 2 Department of Biological Sciences, University of Wisconsin-Whitewater, 800 West Main Street, Whitewater, WI 53190
FIRST RECORD OF RHIPICERIDAE (COLEOPTERA: POLYPHAGA: DASCILLOIDEA) FROM WISCONSIN
Daniel K. Young1 and Kerry Katovich2
ABSTRACTThe Cedar beetle family, Rhipiceridae, is recorded from Wisconsin for the
first time. A single adult female of Sandalus niger was collected on a walk-way along a forested path in south-central Wisconsin on 7 October 2000. This discovery represents not only the first record of Rhipiceridae from Wisconsin, but also the first record of the entire superfamily Dascilloidea, a relatively basal lineage of Elateriformia.
____________________The small and poorly known dascilloid beetle family Rhipiceridae includes
six genera and 57 species, widely distributed in all major zoogeographic regions (Lawrence et al. 1999). Sandalus Knoch (25 species) occurs in North and South America, Africa, Southeast Asia, China, India, and Japan; Rhipicera Latreille (15 species) occurs in Australia, New Caledonia and Brazil;
Chameorhipis Latreille (6 species) occurs in Africa; Ptiocerus Castelaneu (4 species) is restricted to South Africa; Arrhaphipterus Kraatz (6 species) oc-curs in southeastern Europe, North Africa and central Asia; the monotypic Polymerius marmoratus Philippi is endemic to Chile (Katovich, unpubl. notes). Nearctic rhipicerid species, all in Sandalus, include S. californicus LeConte (California, Idaho, and Nevada), S. cribricollis Van Dyke (California), S. niger Knoch (southern Ontario south and west into Texas), S. petrophyus Knoch (north-central United States), and S. porosus LeConte (Arizona, Colorado, New Mexico, and Texas). At least some Neotropical and Nearctic species are known to be attracted to lights.
On 7 October 2000, a single adult female of S. niger was captured on a sidewalk in Stewart Park, in the city of Mount Horeb. The collection site is largely a semi-open, mesic forested habitat located in central Dane County, in south-central Wisconsin. The single specimen upon which this new state record is based, is housed in the Insect Research Collection of the Department of Entomology, University of Wisconsin-Madison. This discovery represents not only the first record of Rhipiceridae from Wisconsin, but also the first record of the entire superfamily Dascilloidea, a relatively basal lineage of Elateriformia. Dascillidae, the only other family of Dascilloidea in the United States is known from a meager fauna in the western states.
Sandalus niger is a hypermetamorphic species, with a triungulinoid first instar and subsequent ectoparasitoid instars developing on cicada nymphs (Craighead 1921, Hicks 1942, and Young 1956). During the summer of 2000, adult cicadas were quite abundant locally, perhaps correlating with the discovery of the adult S. niger.
Elzinga (1977) confirmed the cicada-host relationship, recording the life history of S. niger in Kansas. Adult emergence from soil occurred in late fall, with males frequently flying during warm afternoons, primarily around trees where a female was present. Large numbers of males were often seen attempt-ing to mate with a single female; mating took place on the surface of the tree

34 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
trunk. Rings (1942) recorded 16,846 eggs from a single female S. niger. Eggs were easily washed off plant materials during rain storms (as often appears to be the case with cicada eggs, as well); egg eclosion occurred in the following spring, producing a triungulin. Adults have not been observed to feed.
ACKNOWLEDGMENTSThe authors are grateful to University of Wisconsin undergraduate stu-
dent, Michael Putnam, collector of the specimen.
LITERATURE CITEDCraighead, F. C. 1921. Larva of the North American beetle Sandalus niger Knoch. Proc.
Entomol. Soc. Washington 23: 44-48.Elzinga, R. J. 1977. Observations on Sandalus niger Knoch (Coleoptera: Sandalidae) with
a description of the triungulin larva. J. Kansas Entomol. Soc. 50: 324-328.Hicks, S. D. 1942. Observations on the adult of Sandalus niger Knoch in southern Ontario
(Coleoptera: Rhipiceridae). Can. Entomol. 74: 78-79.Lawrence, J. F., A. M. Hastings, M. J. Dallwitz, T. A. Paine, and E. J. Zurcher. 1999.
Beetles of the world, a key and information system for families and subfamilies. Ver-sion 1.0 for Windows(CSIRO Entomology, Canberra, Australia. CD ROM).
Rings, R. W. 1942. The external anatomy of Sandalus niger Knoch (Coleoptera: Rhipic-eridae). Ann. Entomol. Soc. Am. 35: 411-425.
Young, F. N. 1956. Unusual abundance of Sandalus in southern Indiana. Coleopt. Bull. 9: 74.

2002 THE GREAT LAKES ENTOMOLOGIST 35
LONGITARSUS MELANURUS (COLEOPTERA: CHRYSOMELIDAE) REPRODUCES ON ONOSMODIUM MOLLE (BORAGINACEAE)
Andrew H. Williams1
Longitarsus melanurus (Melsheimer) was first associated with Onosmo-dium Michx. by Popenoe (1877) in Kansas where the sole species is O. molle Michx. (Great Plains Flora Assn. 1986). Recent field observations over several years showed that adults of L. melanurus feed on leaves of O. molle in Wiscon-sin, where these beetles were collected at 43 of the 59 sites where O. molle was found growing naturally (Williams 1996).
On several occasions, L. melanurus adults were found feeding on leaves of Cynoglossum officinale L. (Boraginaceae) at sites where O. molle also occurred (Wil-liams 1996). These beetles were neither observed at these sites on foliage of several native borages: Lithospermum incisum Lehm. (31 sites), L. canescens (Michx.) Lehm. (30 sites), L. carolinense (Walter) MacMillan (1 site) and Hackelia virginiana (L.) I. M. Johnston (15 sites) nor were they seen on the exotics Lappula squarrosa (Retz.) Dumort (2 sites) and Echium vulgare L. (1 site) (Williams 1996). Furth (1994) reported L. melanurus using E. vulgare in Massachusetts and New York.
A simple experiment was done to test whether or not L. melanurus would reproduce on O. molle. Tiny, wild, bare rooted plants were potted and tightly caged. These were maintained free of other plants and without evidence of L. melanurus being present for more than a year. On 11 June 1996, 37 adults collected on O. molle in the wild that same day were introduced. Adults were removed between 27 and 29 June. The caged plants did not again harbor adults until 7 August. Between 7 August and 24 September 1996, 102 adults were removed from these plants. This shows that L. melanurus can reproduce on O. molle, the sole native species on which adults have been reported to feed (Popenoe 1877, Furth 1994, Williams 1996).
Specimens of L. melanurus were donated to the Insect Research Collec-tion of the Entomology Department, University of Wisconsin - Madison. Plant nomenclature herein follows Gleason and Cronquist (1991).
ACKNOWLEDGMENTSThis work was part of my MS thesis research which could not have been
completed without the assistance of E. Williams, R. Christoffel, D. Young, R. Kowal and T. Vale. I’m also grateful to D. Furth and E. Riley for their help determining specimens of L. melanurus early in this process.
LITERATURE CITEDFurth, D. G. 1994. A new case of parthenogenesis in beetles: Longitarsus melanurus
(Melsheimer) (Coleoptera: Chrysomelidae). Jour. N. Y. Entomol. Soc. 102:310-317.Gleason, H. A. and A. Cronquist. 1991. Manual of vascular plants of northeastern United
States and adjacent Canada. 2nd Ed. N. Y. Bot. Garden. Bronx, NY. 910 pp.Great Plains Flora Association. 1986. Flora of the Great Plains. Univ. Press of Kansas.
Lawrence, KS. 1402 pp.Popenoe, E. A. 1877. List of Kansas Coleoptera. Trans. Kan. Acad. Sci. 5:21-40.Williams, A. H. 1996. Conservation of the plant Onosmodium molle A. Michaux (Boragi-
naceae) and the beetle Longitarsus subrufus LeConte (Chrysomelidae) in Wisconsin. MS thesis. Univ. of Wisconsin. Madison, WI. 424 pp. Available from: University Microfilms, Ann Arbor, MI; Order #1382083.
1 Department of Entomology, University of Wisconsin, Madison, WI 53706

36 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1

2002 THE GREAT LAKES ENTOMOLOGIST 37
1Department of Entomology and Center for Integrated Plant Systems, Michigan State University, East Lansing, MI 48824 2 Entomology & Nematology Department, University of Florida, Gainsville, FL 32611
ATTRACTION OF APPLE MAGGOT FLIES (DIPTERA: TEPHRITIDAE) TO SYNTHETIC FRUIT VOLATILE COMPOUNDS AND FOOD
ATTRACTANTS IN MICHIGAN APPLE ORCHARDS
Lukasz L. Stenliski1 and Ocar E. Liburd2
ABSTRACT The apple maggot, Rhagoletis pomonella (Walsh), is a serious pest of
apples in the United States, requiring reliable monitoring and control programs. Various synthetic apple volatile lures with and without protein hydrolysate, ammonium acetate, or ammonium carbonate were evaluated from 1998-2000 for their attractiveness to R. pomonella adults with red sticky-sphere (9 cm diam.) monitoring traps. A blend consisting of butyl butanoate (10%), propyl hexano-ate (4%), butyl hexanoate (37%), hexyl butanoate (44%), and pentyl hexanoate (5%) was the most effective lure tested for attracting both sexes of R. pomonella adults during all three field seasons. The addition of protein hydrolysate or am-monium compounds to spheres baited with a commercial attractant (BioLure) consisting of plastic dispensers containing butyl hexanoate, did not significantly increase apple maggot fly captures. Spheres baited with the blend or with butyl hexanoate in polyethylene vials and spheres baited with BioLure dispensers were highly selective in capturing R. pomonella flies relative to non-target insects. However, spheres baited with ammonium compounds with or without synthetic apple lures were non-selective with respect to apple maggot captures. Protein hydrolysate alone was ineffective for monitoring R. pomonella flies. We provide further evidence that baiting red-sticky sphere traps with the volatile blend without ammonium bait additives creates a highly effective and selec-tive device for capturing apple maggot flies. The blend could be an important addition to current monitoring and control programs for apple maggot flies in Michigan orchards and other important apple growing regions.
____________________Michigan ranks among the top states in apple production value in the U.S.
Apples are grown commercially on more than 23,400 ha and more apples are produced by volume than all other Michigan fruits combined (Michigan Agricul-tural Statistics1999-2000). The apple maggot, Rhagoletis pomonella (Walsh), is an important fruit pest infesting commercially grown apples in Michigan and throughout the eastern U. S. There is zero tolerance for maggot infestation in apples bound for commercial sales. High numbers of apple maggot flies move from wild hosts into commercial orchards in Michigan, responding to visual and olfactory cues of host fruit, where mating and oviposition occurs (Liburd and Stelinski 1999). Sensitive and reliable monitoring techniques for the apple maggot fly are of extreme importance for making accurate and responsible control decisions.
Apple maggot flies are attracted to apple odors in the field (Prokopy et al.1973, Reissig 1974). Location of appropriate mating and oviposition sites by apple maggot flies is mediated in part by odors, which serve as primary cues for host site loca-tion and acceptance in fruit-infesting insects (Frey and Bush 1990). Among apple volatiles isolated from two apple varieties (Red Delicious and Red Astrachan), a blend consisting of a series of short chain carbon esters, including butyl hexanoate, is attractive to apple maggot flies (Fein et al. 1982). A comparison of visual stimuli showed that unbaited, 8 cm diam. red spheres captured significantly more apple maggot flies than unbaited, 10 cm diam. red spheres (Duan and Prokopy 1992).

38 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Furthermore, sticky spheres baited with vials releasing butyl hexanoate captured significantly more apple maggot flies than unbaited spheres, although there was no difference in catch with one, two, or four vials/sphere. A combination treatment containing one vial of ammonium carbonate and one vial of butyl hexanoate also significantly increased the capture of apple maggot flies compared to capture on unbaited spheres.
In addition to synthetic fruit volatiles, other odor baits are used in teph-ritid monitoring programs. Attraction of the Queensland fruit fly, Dacus tryoni Froggatt, to several hydrolysed proteinaceous baits is mainly due to the am-monia they release (Bateman and Morton 1981). Ammonia is a by-product of the bacterial decay of several adult tephritid food sources (Prokopy and Roitberg 1984) and is commonly used as an attractant in tephritid monitoring programs. Ammonia-producing baits may be attractive because flies are seeking a protein source important for egg maturation (Prokopy and Roitberg 1989, Prokopy 1993, and Prokopy et al. 1994).
The production of synthetic semiochemicals has led to the development of effective attractant-baited traps for monitoring and trapping of insects in agricultural production systems (Gut et al. in press). The approach is used to base management decisions on adult catches rather than taking a prophylactic calendar-based approach. Control sprays are typically applied only if catches exceed a predetermined level. Red sticky spheres baited with apple volatiles are often used to monitor apple maggot flies and alert pest managers for the need to apply control measures (Agnello et al. 1990, Stanley et al. 1987). An action threshold of 8 flies per trap was developed, which allowed for a 70% reduction in sprays with acceptable levels of control (Agnello et al. 1990).
More recently, Reynolds and Prokopy (1997) evaluated synthetic odor lures (butyl hexanoate, ammonium carbonate, or butyl hexanoate plus ammonium carbonate on red sticky spheres) for their attractiveness to apple maggot flies. Butyl hexanoate plus ammonium carbonate was more attractive than either odor alone. Finally, a recent study has shown that a lure consisting of a blend of synthetic apple volatiles, including butyl butanoate, propyl hexanoate, butyl hexanoate, hexyl butanoate, and pentyl hexanoate combined in specific propor-tions, was significantly more attractive to adult apple maggot flies than lures containing butyl hexanoate alone (Zhang et al. 1999).
Although a single published study showed evidence of increased attraction using the same blend we investigated, (Zhang et al. 1999), more research was needed to document information regarding the performance of this newly identi-fied blend under different environmental conditions where apple maggot pressure is derived from within orchard plots and from surrounding abandoned fields. Furthermore, the selectivity of this blend to apple maggot flies as compared with other contemporary (butyl hexanoate, Biolure) lures and conventional (ammonium compounds) baits has not been documented. This information is relevant when developing monitoring programs for key pests since Drummond et al. (1984) and Liburd et al. (2000) have previously outlined the problems associated with trap-ping non-target insects on monitoring traps for Rhagoletis species.
The first objective of this research was to further investigate the attractive-ness and selectivity of the newly identified blend. The second was to compare the attractiveness of the blend with contemporary butyl hexanoate lures and conven-tional ammonium baits. Our overall goal was to establish a sensitive, reliable, and selective monitoring device for apple maggot flies that could be implemented by apple growers in Michigan as well as other apple producing regions.
MATERIALS AND METHODS.Research was conducted in apple orchards in Van Buren Co., Michigan during
the 1998, 1999, and 2000 field seasons. Each treatment consisted of a 9 cm diam.,

2002 THE GREAT LAKES ENTOMOLOGIST 39
red sphere coated with approximately 13 g of Tangle Trap (Great Lakes IPM, Vestaburg, MI). Spheres were hung approximately 25 m apart and 30 m between 1 ha blocks of Red Delicious and Golden Delicious. Treatments were arranged in a randomized complete block design (blocked by apple variety) with four replica-tions. Trap positions were re-randomized weekly and old traps were replaced with new ones every 3 weeks. Apple maggot flies caught in traps were identified by sex, counted and removed biweekly. In addition, all non-target insect species captured on traps were counted and removed biweekly in 1998 and 2000. The non-target insects were separated taxonomically, and were used as a whole to es-timate the proportion of apple maggot flies captured compared with non-targets.
1998. To evaluate the importance of protein hydrolysate and ammonium baits, we thoroughly mixed 2.0 g of ammonium acetate or ammonium carbonate and 0.5 g of protein hydrolysate into ca. 13 g of Tangle Trap before application to spheres. The experiment evaluated nine treatments that included red spheres coated with various synthetic protein, ammonia, and apple volatile sources (Table 1). BioLure dispensers (Consep. Inc., Bend, OR) containing a 1.8 g load rate of butyl hexanoate were also used in some treatments.
In a second study, we examined the attractiveness of different types of apple volatile blends with and without ammonium acetate (Table 2). In some
Table 1. Attraction of apple maggot flies to synthetic fruit volatiles and baits, Michi-gan, (1998).
Treatment Mean±SEMfliespertrap 25 June—9 August
Volatile mix blend polyethylene vial 175.0 ± 35.5 aVolatile mix blend polyethylene vial + ammonium acetate 60.5 ± 25.4 bBiolure® dispenser 48.0 ± 2.9 bBiolure® dispenser + ammonium acetate 55.8 ± 16.8 bButyl hexanoate polyethylene vial 75.3 ± 10.7 bButyl hexanoate polyethylene vial + ammonium acetate 71.3 ± 22.1 b
Means followed by the same letter are not significantly different (P = 0.05, LSD test).
Table 2. Attraction of apple maggot flies to synthetic fruit volatiles with and without ammonium-odor and protenaceous baits, Michigan, (1998).
Treatment Mean±SEMfliespertrap 3 July—19 August
Ammonium acetate + protein hydrolysate + Biolure® dispenser 175.3 ± 39.6 aAmmonium acetate + protein hydrolysate 143.8 ± 29.3 aAmmonium acetate 215.5 ± 54.2 aAmmonium carbonate + protein hydrolysate + Biolure® dispenser 174.3 ± 43.8 aAmmonium carbonate + protein hydrolysate 156.3 ± 23.3 aAmmonium carbonate 171.8 ± 35.9 aProtein hydrolysate 34.3 ± 9.6 bBiolure® dispenser 200.5 ± 53.5 aUnbaited (control) 133.0 ± 26.1 a
Means followed by the same letter are not significantly different (P = 0.05, LSD test).

40 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
treatments, we used 5 ml polyethylene vials (Great Lakes IPM, Vestaburg, MI) containing 1.8 ml of butyl hexanoate or 1.8 ml of an apple volatile blend (1 g/ ml solution concentrations). The blend contained butyl butanoate (10%), pro-pyl hexanoate (4%), butyl hexanoate (37%), hexyl butanoate (44%), and pentyl hexanoate (5%) as described in Zhang et al. (1999).
1999 and 2000. For our second and third field seasons, we chose four of the treatments evaluated in 1998 and compared them with unbaited (control) spheres (Table 3). The location of our study, experimental design, and sampling regime were the same as described for 1998.
Statistical analysis. This study was conducted using a randomized com-plete block design and all analyses were conducted using analysis of variance (ANOVA) (PROC GLM, SAS Institute 1989). Blocks in the experimental design were based upon known variability including tree cultivar differences. Data were square root transformed (√x + 0.5) to stabilize the variances. This was followed by mean separation using the least significant difference (LSD) test (SAS Institute 1989). Differences in captures between male and female flies were compared using t-test. Selectivity to apple maggot fly captures was calcu-lated by determining the proportion of apple maggot flies captured relative to all non-target insect captures per trap. Percentage of apple maggot fly captures was also transformed (arcsine [square root (y)]) before analysis.
RESULTS1998. Significantly (F = 4.8; df = 5,15; P < 0.01) more R. pomonella flies
were captured on spheres baited with the blend compared with all other treat-ments tested (Table 1). There were no significant differences in total captures between spheres baited with a mixture of butyl hexanoate and ammonium acetate compared to catches with spheres baited with either compound alone (Table 1). Spheres having ammonium acetate bait with and without synthetic fruit volatiles captured significantly (P < 0.05) more females than males early in the season. This difference was not observed with spheres baited with synthetic fruit volatiles alone (Figure 1).
Spheres baited with protein hydrolysate captured significantly (F = 4.1; df = 8,24; P = 0.03) fewer R. pomonella flies in comparison to spheres baited with other lures (Table 2). With the exception of protein hydrolysate, there were no significant differences among catches with the other baits and lures evaluated in experiment 1. Spheres baited with BioLure (n = 15) alone were highly selective and captured an average of 70 ± 8.6 % apple maggot flies relative to non-target insects. Alternatively, spheres baited with ammonium carbonate and ammonium acetate were non-selective capturing on average 38 ± 3.6 % apple maggot flies, as well as many non-target insects.
1999. As observed in 1998, significantly (F = 3.2; df = 7,12; P = 0.04) more R. pomonella flies were captured on spheres baited with the blend compared
Table 3. Attraction of apple maggot flies to synthetic fruit volatiles and baits, Michi-gan, (1999).
Treatment Mean±SEMfliespertrap
Volatile mix blend polyethylene dispenser 326.5 ± 67.2 aBiolure® dispenser (Butyl hexanoate) 111.5 ± 13.8 bAmmonium Acetate 164.3 ± 51.7 bButyl hexanoate polyethylene vial 150.3 ± 23.7 bUnbaited (control) 105.5 ± 9.0 b
Means followed by the same letter are not significantly different (P = 0.05, LSD test)
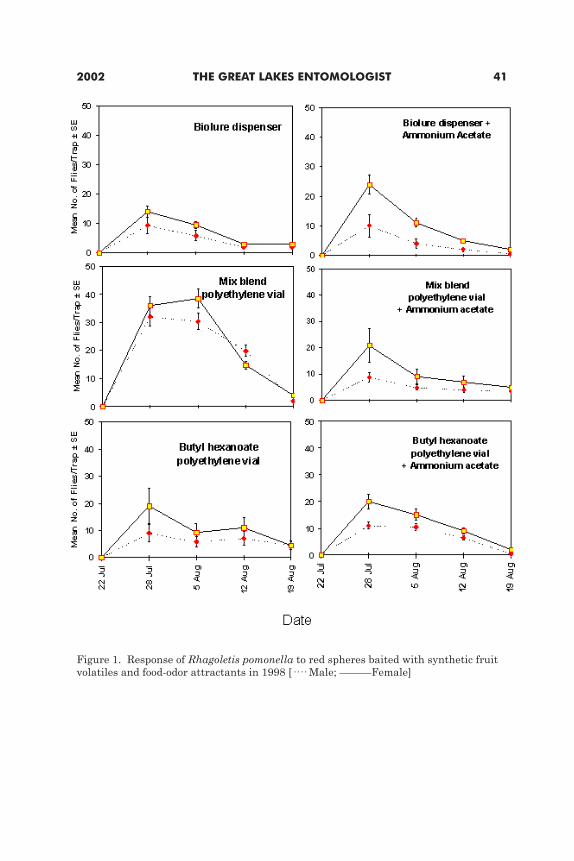
2002 THE GREAT LAKES ENTOMOLOGIST 41
Figure 1. Response of Rhagoletis pomonella to red spheres baited with synthetic fruit volatiles and food-odor attractants in 1998 [···· Male; ———Female]

42 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
with all other treatments (Table 3). In fact, spheres baited with the apple vola-tile blend captured approximately twice as many apple maggot flies as spheres baited with the standard ammonium acetate bait and approximately three times as many apple maggot flies as unbaited (control) spheres. There were no differences in catches observed among the other treatments evaluated (Table 3).
2000. Spheres baited with the blend captured significantly (F = 13.5; df = 7,12; P < 0.01) more apple maggot flies compared with spheres baited with any of the other lures evaluated in 2000 (Table 4). Furthermore, we recorded significantly (F = 13.5; df = 7,12; P < 0.01) more R. pomonella flies captured on spheres baited with butyl hexanoate placed in polyethylene vials compared with spheres baited with the Biolure dispenser and unbaited (control) spheres. We observed a high degree of selectivity to apple maggot capture on spheres baited with the blend (polyethylene vial), butyl hexanoate (polyethylene vial), and the Biolure dispenser, which captured 74.7 ± 5.3 %, 84.2 ± 7.5 %, and 79.5 ± 7.1 %, respectively, of apple maggot flies in relation to non-target insect captures. We observed a moderate degree of selectivity for the unbaited (control) spheres, which captured 60.3 ± 12.9 % apple maggot flies relative to non-target insects. The lowest degree of selectivity was observed on spheres baited with ammonium acetate, which captured 19.8 ± 2.0 % apple maggot flies. Throughout the season, spheres baited with ammonium acetate captured significantly (F = 17.5; df = 3,4; P = 0.05) more R. pomonella females than males, while such a difference was not observed with the other treatments evaluated (Figure 2).
DISCUSSIONOver the course of three field seasons, the apple volatile blend was the most
effective lure evaluated for attracting apple maggot flies. The apple maggot fly is known to use chemical stimuli in long-range orientation to their host plants. Chemical cues are also used to discriminate between host and non-host fruit (Prokopy and Roitberg 1984). When apple maggot flies detect an appropriate blend of odors, such as a synthetic fruit volatile lure, they move upwind in a series of flights that eventually culminate in the arrival at the odor source (Aluja and Prokopy 1992). The blend, as a fruit odor source, was significantly more attractive to apple maggot flies than butyl hexanoate alone. It is possible that the apple volatile blend may have produced a distinct, attractive odor due to a synergistic relationship among the individual compounds that elicit a greater response than individual compounds. Alternatively, differing release rates of compounds from vials loaded with the blend versus butyl hexanoate alone may have impacted trap attractiveness. More research is needed to further investi-gate this hypothesis.
The results from our first experiment in 1998 indicate that spheres baited with protein hydrolysate alone are not an effective means of monitoring R. po-monella fly populations. Spheres baited in this fashion were less effective than spheres baited with various other treatments tested, as well as unbaited spheres.
Table 4. Attraction of apple maggot flies to synthetic fruit volatiles and baits, Michi-gan, (2000).
Treatment Mean±SEMfliespertrap
Volatile mix blend polyethylene dispenser 983.8 ± 203.5 aBiolure® dispenser (Butyl hexanoate) 421.0 ± 32.1 cdAmmonium Acetate 580.0 ± 60.8 bcButyl hexanoate polyethylene vial 610.5 ± 143.4 bUnbaited (control) 336.3 ± 49.1 d
Means followed by the same letter are not significantly different (P = 0.05, LSD test).

2002 THE GREAT LAKES ENTOMOLOGIST 43
Figure 2. Response of Rhagoletis pomonella to red spheres baited with synthetic fruit volatiles and food-odor attractants in 2000 [···· Male; ——— Female]. Significant (p<0.05) differences between pairs of means are indicated by *.

44 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Gow (1954) evaluated various protein hydrolysates and found that although they were initially very attractive to the oriental fruit fly, D. dorsalis Hendel, their attractiveness decreased when the protein hydrolysates experienced bacterial decay. Other studies have shown that hydrolysates are also attractive to apple maggot flies (Dean and Chapman 1973, Reissig 1974). We noticed that when protein hydrolysate was mixed into the Tangle-Trap and applied to a red sphere, the sphere became discolored, assuming a whitish appearance. Spheres baited in this fashion were possibly visually unattractive to R. pomonella flies. In ad-dition, when spheres baited with protein hydrolysate were left in the field for two weeks, they began to lose their initial whitish color as the bait progressively dissipated. These aged spheres (2 wk. exposure) baited with protein hydrolysate, gradually caught increasing numbers of apple maggot flies as they began to regain their original dark red color.
Drummond et al. (1984) found that spheres baited with ammonium com-pounds were not selective for apple maggot flies. Instead, the traps used in their study attracted a wide variety of Dipteran (beneficial) insects, which decreased trap effectiveness due to the reduced trapping area caused by non-target insect captures. We recorded similar results in our study; spheres baited with ammo-nium compounds became saturated with insects after 7-10 d during all three field seasons. We observed that the average life span of a sphere baited with ammonium acetate/carbonate was at most three weeks, depending on fly pres-sure. After three weeks the decomposing insects caused a detectable odor that may have competed with the odor given off by the ammonia baits. Bateman and Morton (1981) studied the role of ammonium compounds and concluded that both ammonia and amino acids were essential to attract tephritid (D. tryoni) species, the former for olfactory stimulation and the latter for feeding response. Additional studies revealed that although ammonia is the major attractant, an accompanying increase in the pH level of the bait compound further increases its attractiveness. They concluded that at this increased pH level, additional unidentified volatiles may be produced, resulting in increased attractiveness to D. tryoni flies. These variables may have affected the results of our study; however, we did not measure these factors.
The addition of protein hydrolysate or ammonium compounds to spheres baited with BioLure did not significantly increase the attractiveness of these spheres to apple maggot flies. Similar results demonstrating limited effectiveness of red sphere traps baited with ammonium compounds have been documented previously (Reynolds and Prokopy 1997). In fact, the only difference that was recorded by combining an ammonium bait with an apple volatile lure in com-parison to using the apple volatile alone, was an increased capture of female R. pomonella relative to males in early season. However, unbaited spheres and spheres baited only with BioLure, butyl hexanoate, or the blend, were far more selective in capturing R. pomonella flies than lures containing ammonium com-pounds. Such spheres containing no ammonia or protein bait, impregnated in the Tangle-Trap, maintained the original deep red color of the plastic. Spheres that were treated with baits impregnated into the Tangle-Trap tended to become discolored (lighter in color, whitish appearance).
The attractiveness of unbaited spheres observed in both 1998 and 2000 may be indicative of the visual stimulus, which mimics apples in both size and color (Prokopy 1968). The addition of apple volatiles to these spheres increased their attractiveness to apple maggot flies, while at the same time preserving their selectivity. The principal component of BioLure is butyl hexanoate, a fruit odor that attracts sexually mature apple maggot flies for mating and oviposition (Reynolds and Prokopy 1997). The possible reason why we did not observe an increased total trap capture by combining ammonium baits with BioLure, may be the decreased selectivity associated with this type of trap baiting system in comparison to using the synthetic apple volatile alone. Also, apple maggot flies immigrating into commercial orchards are sexually mature and therefore exhibit

2002 THE GREAT LAKES ENTOMOLOGIST 45
a greater response to host-fruit volatiles than to food-odor attractants (Rull and Prokopy 2000). Therefore, we feel that using synthetic apple volatiles without ammonium baits may produce more accurate and sensitive monitoring of both resident and immigrant populations of R. pomonella flies in Michigan com-mercial apple orchards.
We found that by using synthetic apple volatiles (apple volatile blend, butyl hexanoate, BioLure), the effectiveness of red, sticky sphere traps used in apple maggot monitoring programs could be extended beyond the three week life span characterizing monitoring traps baited with ammonium attractants. The combination of attractiveness and selectivity associated with using the blend may provide a high degree of effectiveness for monitoring both sexes of apple maggot flies in commercial apple orchards. Furthermore, the use of the apple volatile blend may produce the best possible results in apple maggot fly mass trapping programs.
ACKNOWLEDGMENTSWe thank two anonymous owners of Michigan apple orchards for allow-
ing us to conduct our study. We thank Dan Young for his technical assistance with treatment preparation, sampling, and fieldwork. We also thank Dr. John Wise, Dr. Larry Gut, and the staff at the Trevor Nichols Research Complex for their valuable assistance on this project. We thank Noah Ressa for reviewing a previous version of the manuscript. The research reported here was supported by USDA Special Fruit Grants 61-4058 and 61-4415.
LITERATURE CITEDAgnello, A. M., S. M. Spangler, and W. H. Reissig. 1990. Development and evaluation of
a more efficient monitoring system for apple maggot (Diptera: Tephritidae). J. Econ. Entomol. 83: 539-546.
Aluja, M. and R. J. Prokopy. 1992. Host search behavior of Rhagoletis pomonella flies: inter-tree movement patterns in response to wind-borne fruit volatiles under field conditions. Physiol. Entomol. 17: 1-8.
Bateman, M. A. and T. C. Morton. 1981. The importance of ammonia in proteinaceous attractants for fruit flies (Family: Tephritidae). Aust. J. Agric. Res. 32: 883 - 903.
Dean, R. W. and P. J. Chapman. 1973. Bionomics of the apple maggot in Eastern New York. New York State Agric. Entomol. Geneva 3(10): 62 pp.
Drummond, F., E. Groden, and R. J. Prokopy. 1984. Comparative efficacy and optimal positioning of traps for monitoring apple maggot flies (Diptera: Tephritidae) Environ. Entomol. 13: 232-235.
Duan, J. J. and R. J. Prokopy. 1992. Visual and odor stimuli influencing effectiveness of sticky spheres for trapping apple maggot flies Rhagoletis pomonella (Walsh) (Dipt., Tephritidae). J. Appl. Entomol. 113: 271-279.
Fein, B. L., W. H. Reissig, and W. L. Roelofs. 1982. Identification of apple volatiles at-tractive to the apple maggot, Rhagoletis pomonella. J. Chem. Ecol. 8: 1473-1487.
Frey, J. E. and G. L. Bush. 1990. Rhagoletis sibling species and host races differ in host odor recognition. Entomol. Exp. Appl. 80: 163-165.
Gow, P. L. 1954. Proteinaceous baits for the oriental fruit fly. J. Econ. Entomol. 47: 153-160.Gut, L. J., L. L. Stelinski, D. R. Thomson, and J. R. Miller. 2003. Behavior modifying
chemicals: prospects and constraints. Chapter 2 In: Koul, O. and G. S. Dhaliwal (eds.) Integrated Pest Management: Potential, Constraints, and Challenges. CABI Press. Wallingford, UK.
Liburd, O. E. and L. L. Stelinski. 1999. Apple maggot fly and its sibling species: physi-ological and environmental status. MSU CAT Alert Ext. Bull. 14: 3-4.

46 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Liburd, O. E., L. L. Stelinski, L. J. Gut, and G. Thornton. 2000. Performance of various trap types for monitoring populations of cherry fruit fly species (Diptera: Tephritidae). Environ. Entomol. 30: 82-88.
Prokopy, R. J. 1968. Visual responses of apple maggot flies, Rhagoletis pomonella (Dip-tera: Tephritidae): orchard studies. Entomol. Exp. Appl. 11: 403-422.
Prokopy, R. J. 1993. Levels of quantitative investigation of tephritid fly foraging behav-ior, 165-171. In M. Aluja & P. Liedo [eds.], Fruit flies: biology and management. Springer, New York.
Prokopy, R. J., S. S. Cooley, J. J. Prokopy, Q. Quan, and J. P. Buonaccorsi. 1994. Interac-tive effects of resource abundance and state of adults on residence of apple maggot (Diptera: Tephritidae) flies in host tree patches. Environ. Entomol. 23: 304-315.
Prokopy, R. J., V. Moericke, and G. L. Bush. 1973. Attraction of apple maggot flies to odor of apples. Environ. Entomol. 2: 743-749.
Prokopy, R. J. and B. D. Roitberg. 1984. Resource foraging behavior of true fruit flies. Am. Sci. 72: 41-49.
Prokopy, R. J. and B. D. Roitberg. 1989. Fruit fly foraging behavior, pp. 293-306. In A. S. Robinson and G. Hooper (eds.) Fruit flies: their biology, natural enemies and control. Elsevier, Amsterdam.
Reissig, W. H. 1974. Field tests of the response of Rhagoletis pomonella to apples. Envi-ron. Entomol. 3: 733-736.
Reynolds, A. H. and R. J. Prokopy. 1997. Evaluation of odor lures for use with red sticky spheres to trap apple maggot flies. J. Econ. Entomol. 90: 1655-1660.
Rull, J. and R. J. Prokopy 2000. Attraction of apple maggot flies, Rhagoletis pomonella (Diptera: Tephritidae) of different physiological states to odour-baited traps in the presence and absence of food. B. Entomol. Res. 90: 77-88.
SAS Institute. 1989. SAS/STAT user’s guide, version 6, 4th ed., vol. 1. SAS Institute, Cary, NC.
Stanley, B. H., W. H. Reissig, W. L. Roelofs, M. R. Schwartz, and C. A. Shoemaker. 1987. Timing treatments for apple maggot (Diptera: tephritidae) control using sticky sphere traps baited with synthetic apple volatiles. J. Econ. Entomol. 80: 1057-1063.
Stelinski, L. L., O. E. Liburd, S. E. Wright, R. J. Prokopy, R. Behle, and M. R. McGuire. 2001. Comparison of neonicotinoid insecticides for use with biodegradable and wooden spheres for control of key Rhagoletis species (Diptera: Tephritidae). J. Econ. Entomol. 94: 1142-1150.
Zhang, A., C. Linn Jr., S. Wright, R. Prokopy, W. Reissig, and W. Roelofs. 1999. Identi-fication of a new blend of apple volatiles attractive to the apple maggot, Rhagoletis pomonella. J. Chem. Ecol. 25: 1221-1232.

2002 THE GREAT LAKES ENTOMOLOGIST 47
A PRESERVATIVE-FREE EMERGENT TRAP FOR THE ISOTOPIC AND ELEMENTAL ANALYSIS OF EMERGENT INSECTS
FROM A WETLAND SYSTEM
Richard A. MacKenzie1 , 2 and Jerry L. Kaster 1
ABSTRACTThis study reports a cost-effective, live emergent trap designed for the
preservative-free use in both biogeochemical and ecological analyses of emerging insects. The trap proved to be advantageous in several ways. First, the simple design made the trap time-efficient since it was easy to set-up, change, and maintain. Second, live sampling not only provided uncontaminated organisms for elemental and stable isotopic analyses, it minimized disfigurement. This resulted in rapid and easy handling, as well as identification, of adult insects. Finally, trap avoidance by ephemeropterans and odonates, a common problem encountered in the literature, was minimal and organisms from both insect orders were successfully collected.
____________________Aquatic and semi-aquatic invertebrates are known to be an important link
between primary production and higher trophic levels in wetlands (Mitch and Gosselink 1993, Batzer and Wissinger 1996). The large amount of emerging insect biomass and abundances reported from wetland systems (Welch et al. 1988, McLaughlin and Harris 1990, Leeper and Taylor 1998, Stagliano et al. 1998) suggest that they might play a significant role in the export of nutrients when they leave the system for dispersion or reproductive reasons (Davies 1984), or as food for anadromous fish or migratory waterfowl (Herdendorf et al. 1986, Hanson and Riggs 1995). A popular tool for examining these trophic interactions (Rau and Anderson 1981, Peterson and Fry 1987, Fry 1991, Ke-ough et al. 1996) as well as the nutrient budgets of aquatic systems (Quay et al. 1986, Garman and Macko 1998) is stable isotopic analysis. However, many modern emerging insect traps include collecting jars filled with preservatives (e.g., Davies 1984, Merrit and Cummins 1996) that can contaminate samples and ultimately affect the carbon and nitrogen content of the captured insects. Thus, in order to further our quantitative understanding of the role emerging insects play in trophic dynamics and mass balances of wetland systems, it was essential to design an emergent insect sampling device suitable for stable iso-topic and elemental analysis.
We have designed a cost-effective, live emergent trap that eliminated the use of preservatives. The traps were relatively simple to make and maintain, and were highly field-usable. This method allowed for easy identification of insects since adults were brought back intact and alive to the lab reducing disfigurement (e.g., wing venation) resulting from transfer or storage in liquid preservative over an extended period of time.
MATERIALS AND METHODSTrap design. The main body of the trap (Fig. 1) was constructed of 1.0-
mm mesh mosquito netting sewn into a 136-cm high, 80-cm diameter cylinder. The base of the netting was hemmed around a drawstring 310 cm long allowing the trap to be easily cinched shut. The frame of the trap was made from an 1The Center for Great Lakes Studies at the WATER Institute, University of Wisconsin-Milwaukee, 600 E. Greenfield Ave., Milwaukee, WI, 53204.2Present address: The Wells National Estuarine Research Reserve, 342 Laudholm Farm Rd., Wells, ME, 04090.

48 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
80-cm diameter children’s plastic “hula hoop” filled with sand as ballast and a capped, 2.75-cm diameter PVC pipe of varying height depending upon the depth of the water.
The trap was deployed by clamping the hemmed base of the netting around the hoop using pieces of garden hose 10 cm long, slit down the middle. After the PVC pipe was driven into the substrate, the clamped netting and hoop assemblage were lowered over the PVC pipe until the hoop-base was slightly submerged. The edge of the open end at the top of the trap was bundled to-gether and sealed off using a polyurethane cable-tie to prevent any insects from escaping the trap. The trap was then secured by staking the hoop-base into the wetland. The entire assemblage resembled a tee-pee and sampled a 0.5-m2 area.
Retrieval of the net entailed removing the garden hose clamps from the hoop/net base while the hoop was still submerged. Tightening the drawstring around the PVC pipe closed the base of the net underwater. By cinching the trap shut underwater, any insects that had fallen back onto the water’s surface or that were in the process of ovipositing were also captured. The closed net was slid up over the PVC pipe, removing any insects resting there as well. The net samples were placed in a 0oC freezer for 24 hours. Insects were then removed from the traps by gently shaking the nets, and whole, intact, frozen specimens were keyed down to family using Lehmkuhl (1979) and Merrit and Cummins (1996). Nets were then inspected for holes, darned, and cleaned in a washing machine.
To illustrate the effectiveness of this trap, a data set from the Peshtigo wetland is presented where insect densities reported represent 5% or more of the total number of insects collected. For a more detailed description of the Peshtigo wetland, the study design, and the data collected, see MacKenzie (2001). Traps were deployed at four stations designated as riverine stations (0-5 m from the wetland-riverine interface) and five wetland interior stations (stations greater than 70 m from the wetland-riverine interface) located within the Peshtigo riverine wetland during the first week of June in 1997 and were
Figure 1. Exploded view and dimensions of emergent trap.

2002 THE GREAT LAKES ENTOMOLOGIST 49
changed every other week until November 1997. Riverine stations were char-acterized by sparse emergent macrophytes that included Typha sp., Carex sp., Sagittaria sp., Scirpus sp., and/or Lythrum salicaria as well as submergent vegetation such as Potamogeton sp. and Myriophyllum sp. Water column depth was > 1 m with an average dissolved oxygen concentration of 7.3 ± 0.6 mg.L-1 and an average temperature of 19.8 ± 1.3 oC. Wetland interior stations were characterized by dense stands of Carex sp., Sparganium sp., Typha sp., and/or Phalaris arundinacea while submergent vegetation was generally absent. Water column depth ranged from 0.1 to 0.4 m with an average dissolved oxygen concentration of 4.0 ± 0.5 mg.L-1 and an average temperature of 18.3 ± 1.5 oC.
Statistical analyses were performed using SYSTAT (version 8, SPSS Inc., Chicago). Emerging insect densities were first normalized using log-transformations (Elliot 1977) and then tested for significant differences among dominant taxa between riverine and wetland stations using a 1-way ANOVA.
RESULTS AND DISCUSSIONCommunity composition of insects. Our trap design was effective
enough to reveal some differences in the community composition at the family level as well as differences in densities of insects emerging from riverine and the wetland interior stations (Table 1), with total densities significantly greater at riverine stations (P = 0.01). The presence of immature forms in kick net samples collected from the same stations (MacKenzie 2001) also revealed the ability of our trap design at capturing organisms relevant to the sampling area. Dipterans were the most abundant insects captured from both sampling areas and were represented largely by 3 families: Chironomidae, Dolichopodidae, and Ephydri-dae. Densities of Chironomidae and Ephydridae were significantly greater at the riverine stations (P = 0.019 and P = 0.04 respectively). Ephemeropterans, largely Leptophlebiidae, and trichopterans, largely Hydropsychidae and Molannidae, also exhibited higher densities at riverine stations, although only Molannidae caddisfly densities were significant (P < 0.05). The absence of emerging lepto-phlebiid adults from the riverine stations was likely due to their migration from the adjacent river to more stagnant waters (i.e., the wetland interior) where they tend to emerge (Unzicker and Carlson 1982). This observation was supported by the sudden appearance of late instar leptophlebiid mayfly nymphs in kick net samples prior to their emergence (MacKenzie 2001). Odonates were also collected and were largely represented by the damselfly family, Coenagrionidae, with densities significantly greater at riverine stations (P = 0.003).
The traps, which cost around eight dollars each, have been used in areas ranging from minimal or no surface water to heavily inundated areas in both fresh and saltwater systems. They were excellent alternatives to traps that used preservatives and or plastic coverings. Not only did they allow for the elemental and isotopic analyses of emerging insects, they also allowed for the qualitative and quantitative determination of densities and biomass due to minimal trap avoidance exhibited by certain insect orders. This also allowed us to document differences in community composition at the family level (e.g., leptophlebiid mayflies). Since mosquito netting was used, the problems of condensation or temperature increase within the traps were not significant, as reported when polyurethane traps have been utilized (Davies 1984, Stagliano et al. 1998). The only major problem that may have affected our sampling was shading caused by the netting. However, organisms in the nets coincided with the immature, benthic populations, which suggested this was not a significant problem (R. A. MacKenzie, unpublished data). Finally, since nets were frozen to kill the trapped adults, their carapaces became fragile and brittle. This resulted in damage to some of the adults, particularly the odonates, when they were removed from the traps. This was quickly resolved by simply employing care and finesse in the removal of the remaining adults from the traps.

50 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
ACKNOWLEDGEMENTSWe thank Donald C. Szmania, Jeremy J. Solin, and Tracy L. Sykes for their
many hours spent setting and retrieving nets in the Peshtigo wetland. This work was funded by the University of Wisconsin Sea Grant Institute under grants from the National Sea Grant College Program, National Oceanic and Atmospheric Administration, U. S. Department of Commerce, and from the State of Wisconsin. Federal Grant number NA46RG0481, project number R/EC - 2.
LITERATURE CITEDBatzer, D. P. and S. A. Wissinger. 1996. Ecology of insect communities in nontidal
wetlands. Ann. Rev. of Entom. 41:75-100.Davies, I.J. 1984. Sampling aquatic insect emergence. Pages 161-227 in J.A. Downing
and F.H. Riegler (editors). A manual on methods for the assessment of secondary productivity in fresh waters. IBP Handbook 17, Blackwell Scientific Publications, Oxford, UK.
Elliot, J. M. 1977. Some methods for the statistical analysis of samples of benthic inver-tebrates. Freshwater Biological Assoc., Scientific Publication no. 25. 159 pp.
Fry, B. 1991. Stable isotope diagrams of freshwater food webs. Ecology 72:2293 -2297.Garman, G. C. and S. A. Macko. 1999. Contribution of marine-derived organic matter to
an Atlantic coast, freshwater, tidal stream by anadromous clupeid fishes. J. N. Am. Benthol. Sci. 17:277-285.
Table 1. Total emerging numbers (±1 SE) over the 5-month sampling period from the riverine and wetland interior stations. All insect orders and families represent at least 5% of the total insect densities collected. Densities reported for each order may include individuals that could not be keyed down to family. Statistical comparisons were made between riverine samples (n=4) and wetland interior samples (n=5) using an unbal-anced, one-way ANOVA (* P < 0.05).
Density (no.m-2) WetlandTaxon Riverine interior F-ratio P-value
Diptera 324 ± 76 81 ± 30 9.30 0.019* Chironomidae 238 ± 87 33 ± 17 7.99 0.025* Dolichopodidae 25 ± 13 22 ± 21 1.94 0.207 Ephydridae 42 ± 14 7 ± 3 6.07 0.043*
Ephemeroptera 6± 1 21 ± 16 0.01 0.953 Leptophlebidae 0 ± 0 17 ± 17 0.78 0.407 Siphlonuridae 6 ± 1 4 ± 2 3.417 0.074
Odonata 29 ± 1 9 ± 3 9.48 0.022* Libellulidae 1 ± 1 5 ± 2 5.20 0.057 Coenagrionidae 27 ± 1 3 ± 3 19.43 0.003*
Trichoptera 45 ± 13 3 ± 4 20.15 0.003* Hydropsychidae 10 ± 10 0 ± 0 1.30 0.292 Limnephilidae 4 ± 3 2 ± 2 0.45 0.523 Molannidae 12 ± 7 0 ± 0 6.63 0.037*
Total 413 ± 71 122 ± 27 11.73 0.011*

2002 THE GREAT LAKES ENTOMOLOGIST 51
Hanson, M. A. and M.R. Riggs. 1995. Potential effects of fish predation on wetland in-vertebrates: a comparison of wetlands with and without fathead minnows. Wetlands 15:167-175.
Herdendorf, C.E., C.N. Rapheal and E. Jaworski. 1986. The ecology of Lake St. Claire wet-lands: a community profile. U.S. Fish and Wildlife Service Biological Report No. 85.
Keough, J.R., M.E. Sierszen and C.A. Hagley. 1996. Analysis of a Lake Superior coastal food web with stable isotope techniques. Limnol.Oceanogr. 41:136-146.
Leeper, D.A. and B. E. Taylor. 1998. Insect emergence from a South Carolina (USA) temporary wetland pond, with emphasis on the Chironomidae (Diptera). J. N. Am. Benthol. Soc. 17:54 -72.
Lehmkuhl, D.M. 1979. How to know the aquatic insects. Wm. C. Brown, Dubuque, Iowa.MacKenzie, R.M. 2001. Great Lakes coastal wetland-estuarine systems: Invertebrate
communities, biogeochemical patterns, and sediment particle dynamics. Ph.D. thesis University of Wisconsin –Milwaukee, Milwaukee, Wisconsin.
Mclaughlin, D. B. and H. J. Harris. 1990. Aquatic insect emergence in two Great Lakes marshes. Wet. Ecol. Manage. 1:111-121.
Merritt, R.W. and K.W. Cummins. 1996. An introduction to the aquatic insects of North America. 3rd ed. Kendall/Hunt, Dubuque, Iowa.
Mitsch, W.J. and J.G. Gosselink. 1993. Wetlands. 2nd edition. Van Nostrand Renhold, New York.
Peterson, B.J. and B. Fry. 1987. Stable isotopes in ecosystem studies. Ann. Rev. Ecol. Syst. 18:293-320.
Quay, P.D., S.R. Emerson, B.M. Quay and A.H. Devol. 1986. The carbon cycle for Lake Washington - A stable isotope study. Limnol. Oceanogr. 31:596-611.
Rau, G.H. and N.H. Anderson. 1981. Use of 13C/12C to trace dissolved and particulate or-ganic matter utilization by populations of an aquatic invertebrate. Oecologia 48:19-21.
Stagliano, D. M., A.C. Benke and D.H. Anderson. 1998. Emergence of aquatic insects from 2 habitats in a small wetland of the southeastern USA: temporal patterns of numbers and biomass. J. N. Am. Benthol. Soc. 17:37-53.
Unzicker, J.D. and P.H. Carlson. 1982. Ephemeroptera. Pages 3.1 to 3.97 in A.R. Brigham, W.U. Brigham, and A. Gnilka (editors). Aquatic insects and oligochaetes of North and South Carolina. Midwest aquatic enterprises, Mahomet, Illinois.
Welch, H.E., J.K Jorgenson and M.F. Curtis. 1988. Emergence of Chironomidae (Dip-tera) in fertilized and natural lakes at Saqvaqjuac, N.W.T. Can. J. Fish. Aqua. Sci. 45:731-737.

52 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1

2002 THE GREAT LAKES ENTOMOLOGIST 53
MOTHS OF THE DOUGLAS LAKE REGION (EMMET AND CHEBOYGAN COUNTIES), MICHIGAN: VI.
MISCELLANEOUS SMALL FAMILIES (LEPIDOPTERA)
Edward G. Voss1
ABSTRACTForty-seven species in nine families of Lepidoptera (Hepialidae, Psychidae,
Alucitidae, Sesiidae, Cossidae, Limacodidae, Thyrididae, Pterophoridae, Epiplemi-dae) are listed with earliest and latest recorded flight dates in Emmet and Cheboy-gan counties, which share the northern tip of the Lower Peninsula of Michigan. The records are from the principal institutional and private collections of Michigan moths and continue the documented listing of Lepidoptera in the region.
____________________Emmet and Cheboygan counties share the northern tip of the Lower
Peninsula of Michigan, the former bordered on the west by Lake Michigan and the latter, on the east by Lake Huron. The Straits of Mackinac connect these two Great Lakes; the village of Mackinaw City, on the Straits, is bisected by the county line. About 15 miles to the south, the University of Michigan Biological Station has its campus on the southeast shore of Douglas Lake, which is entirely in Cheboygan County although the west shore is very near the Emmet County line. Workers at the Biological Station (which has held regular summer teach-ing and research sessions since 1909) have generally taken the two counties to constitute the local “region” for many studies, although the boundaries are only partly natural.
Having spent at least a portion of each summer (save one) since 1930 in Mackinaw City and taught botany courses at the Biological Station for 35 seasons, my own interests have naturally also centered in the same region. My Lepidoptera collection (and library), which formally commenced in 1944, is in fact kept at Mackinaw City.
Checklists inevitably become soon out of date. But they do provide fixed records of information at a given time, and thus are useful as reference stan-dards even long after various details are considered obsolete. (One must also remember that the very latest taxonomic opinions may not always be the best or most enduring ones.) Unlike previous installments of the present annotated list (Voss 1969, 1983, 1984, 1991; Scholtens 1996), this one does not include a single family or a block of families usually classified near one another in any checklist. (But the sequence of families presented thus far is no longer in ac-cord with modern systems for the Lepidoptera anyway.) Nevertheless, the families presented below are in the same sequence as in the “MONA” (Moths of North America) checklist (Hodges et al. 1983) and within each family the spe-cies are similarly listed, preceded by the MONA number, for ease in checking synonymy. Scientific names and their authors, too, generally follow that list (any exceptions noted). Plant names are those employed in my Michigan Flora (Voss 1972, 1985, 1996).
Moths of the traditional “macrolepidoptera” were treated in the first four installments of this list and totaled 619 species (bringing to 700 the Douglas Lake region “macros” including 81 butterflies as itemized in Voss 1954 and Voss and Wagner 1956). The “macro” families appeared in the sequence of the then-familiar McDunnough checklist of 1938. Scholtens (1996) recorded 187
1University of Michigan Herbarium, 3600 Varsity Dr., Ann Arbor, Michigan 48108-2287

54 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
species of Crambidae and Pyralidae. (All of these numbers are now slightly superseded.) The present installment lists 47 species in nine small families traditionally (except for Epiplemidae) included in the “microlepidoptera.” Some of these are considered to be among the most primitive Lepidoptera and overall they include generally larger, conspicuous, and/or distinctive species — “macro micros” one might say: Hepialidae, Psychidae, Alucitidae, Sesiidae, Cossidae, Limacodidae, Thyrididae, Pterophoridae, and Epiplemidae. The families of “micros” that still await coverage can doubtless add a great many more species (and mostly of very small moths). Slightly over half of the pyraloids listed for Michigan (Nielsen 1998) are known from the Douglas Lake region (Scholtens 1996). Extrapolating to the remaining “micro” families, over 400 additional species may be expected, bringing the predicted total documented lepidopteran fauna to upwards of 1300 species for this two-county region.
For each species listed below, following the county or counties from which it has been recorded, are the earliest and latest dates for collection or obser-vation of adults. For apparently rare species (known generally from a single collection) the year and more complete details, if known, are given. When only other collectors are mentioned, the species is not in my collection from the region (unless otherwise indicated).
Since Moore (1955) omitted all microlepidoptera in his list of Michigan moths and Welch (1915) listed only five species (three of them questionable) in the families treated here, there are very few previously published records of these families from the Douglas Lake region. All species included are listed for Michigan by Nielsen (1998) except for four Pterophoridae (6091.1, Capperia sp., 6149, and 6214). Of the listed species, 83% are in the collection of Brian G. Scholtens, 60% in my collection, 40% in the collection of the University of Michigan Biological Station [UMBS], 26% in the University of Michigan Museum of Zoology [UMMZ], and 11% in the Entomology Museum at Michigan State University [MSU]. These are the bases for the present account and institutional collections are cited by abbreviations as indicated in brackets.
Family HepialidaeThese primitive and interesting Lepidoptera are rarely seen (except at
exactly the times of their nuptial flights), despite their large size.18. Sthenopis argenteomaculatus (Harr.). Cheboygan: 25 June–2 July. This
species is known to fly about a month earlier than the next. Although Forbes (1923, p. 67) states that species of this genus do not come to light or sugar, Rings and Metzler (1990) list S. argenteomaculatus as collected at mercury vapor light. My specimen was collected by Gary R. Williams at carbon arc light at the Biological Station in 1964. Scholtens collected a specimen at light at the Station 2 July 1990.
Dirig (1993) reported females at light and described the brief synchronized evening flight near alders (Alnus), in the roots of which the larvae are said to bore. Handfield (1999) also discussed the habits of this ghostly species (and the next), including its life cycle, crepuscular flight, and attraction to light.
19. Sthenopis purpurascens (Pack.). Emmet, Cheboygan: 14–22 July. My only specimen was resting on a street lamp-post in Mackinaw City on a morning in 1945, presumably attracted to the incandescent light during the night; it is a female and laid 27 eggs in captivity before being killed. There are three specimens at UMBS, presumably taken at light at Douglas Lake, in 1937, 1958, and 1959. One taken in Cheboygan County by J. H. Newman in 1943 is at MSU. All the specimens are the salmon form long separated as S. quadriguttatus (Grt.) (see E. S. Nielsen et al. 2000). I have four specimens (including both color forms) from the Thunder Bay and Algoma districts of

2002 THE GREAT LAKES ENTOMOLOGIST 55
Ontario, where two of them were at gasoline lantern at the Neys Provincial Park campground and one on a post near windows in the town of White River; the years are 1967, 1972, 1981, and 1993. Schmidt and Lawrie (2000) comment on the biennial habit of this species, of which most Alberta adults occurred in odd-numbered years. The larvae bore in the roots of Salicaceae (poplars and willows) and take two years to mature. Handfield (1999) noted that (in Quebec) adults are found at light in all years but slightly more in even-numbered ones. The five Michigan specimens in UMMZ are from Alger, Mackinac, Marquette, Montmorency, and Schoolcraft counties, confirming the northern distribution of this species in the state and giving a somewhat longer range of flight dates (14 July–9 August). M. C. Nielsen collected both this species and Hepialus gracilis Grt. on 15 July 2000 in Otsego County (immediately south of Cheboygan Co.) (Ferge 2001).
Family Psychidae437. Psyche casta (Pallas). Emmet, Cheboygan: 18 June–3 August. First noted
in 1983 by David Cowan at the Biological Station. Collected in 1984 and identification confirmed by Donald Davis. Then new to Michigan, but now known from a number of counties, locally abundant at times. Needle frag-ments of white pine (Pinus strobus) seem to be favored in construction of cases, which may be found on diverse kinds of plants as well as man-made substrates. As noted by Davis (1964), this is an Old World species first dis-covered in North America in 1931 (Massachusetts). It apparently continues to spread westward.
438. Apterona helix (Siebold). Emmet. Cases only found by Scholtens on a building at the Pellston airport in June 2000. Another Eurasian species, periodically introduced at various places in North America (Davis 1964).
449. Hyaloscotes pithopoera (Dyar). Cheboygan: emerged 13 July 1990. Reared by Scholtens from a case found on a downed pine log near the Biological Station. A native species known from scattered localities in North America.
Family Alucitidae2313. Alucita hexadactyla L. Emmet, Cheboygan: 30 July–13 August. My only
specimen was given me by R. B. Schwab, who collected it 13 August 1965, at light at the Biological Station. A specimen in UMBS is labeled 7/30/35 E.N.W. [= Elton N. Woodbury] with no further data. The only Emmet County record is a specimen seen clearly by Gary R. Williams at light in Mackinaw City in late July, 2000; it evaded capture. The name used here applies to a European species. The American species are different, and a thorough study of the genus is in preparation (B. Landry, pers. comm.). Specimens of Alucita are seldom collected in Michigan. In the Timiskaming District of northern Ontario, it was reported as common in 1995, occurring as early as 1 April and as late as 30 October (Layberry 1996). In 1993 it was reported there 2–8 May and 1 August–21 October (Layberry 1994). These dates would indicate two broods or one brood that over-winters, as would dates on specimens in UMMZ and MSU (R. Beebe) from Montmorency County (immediately south of Cheboygan County), 13 & 15 May 1946 and 14 July 1943. The only other Michigan specimens known to me are from Dickinson County 20 May and 13 June 1983 (Adam Porter, MSU) and from Berrien County (E. Liljeblad 7 July 1906, UMMZ).
Family SesiidaeThree comprehensive treatments, well illustrated in color, of this fam-
ily (clearwing moths or clearwing borers, also known as Aegeriidae) in North

56 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
America have been published (Beutenmüller 1901, Engelhardt 1946, Eichlin and Duckworth 1988), but particularly useful for this region is the work of Taft et al. (1991). Mr. Taft has determined the specimens in my own collection as well as those at MSU.
The larvae of these moths are all borers, in a great diversity of trees, shrubs, and herbaceous plants, attacking stems, bark, or underground parts. Some are serious pests of agricultural and ornamental plants. The works cited above include information on reported hosts, which are usually restricted to certain plant families or even genera. Adults of many species have relatively large transparent areas in the wings and mimic bees or wasps — or perhaps even large flies. Furthermore, the adults are diurnal, so the mimicry is thor-ough. Nevertheless, some do come to light, the source of most records noted here. Within the past three decades, much use has been made of synthetic pheromones (sex attractants) to attract adult males, but so far as I am aware, traps thus baited have seldom been employed in the Douglas Lake region. In July 1978, I saw a small sesiid working the flowers of a blunt-leaf orchid (Ha-benaria obtusata) at Grass Bay, east of Cheboygan. It bore two pollinia on its head, but eluded capture and hence identification (cf. Voss and Riefner 1983, where this observation was overlooked).2513. Pennisetia marginata (Harr.). Cheboygan: 16 August 1962. One speci-
men from Mullett Creek, collected by R. E. Beer (UMBS). The larvae of this species, the raspberry crown borer, bore in the canes of blackberries, raspberries, and other species of Rubus.
2524. Paranthrene tabaniformis (Rott.). Cheboygan: 2–30 July. Collected on low dunes at Grass Bay, east of Cheboygan, both by me (in 1982) and by David Cowan (in 1988). Scholtens collected the species in 1989 abut three miles west, in similar terrain near the Lake Huron shore. Willows (Salix) are the reported preferred larval host, and there are several species of this genus at Grass Bay.
2543. Sesia tibialis (Harr.). Emmet, Cheboygan: 3–30 July. Drawn to a phero-mone trap and collected 3–5 July 1991 by Scholtens on the Wildwood jack pine plains south of Indian River. Apparently not common. Two additional specimens are in UMBS and one in MSU. The larval host, aspen (Populus), is abundant in the region — unlike the moth. Welch (1915) listed this spe-cies and an old, undated specimen in UMMZ labeled merely “Douglas Lake” is presumably his. Besides this species, Welch also listed S. apiformis (Cl.), which is not otherwise known from Michigan; if not a complete misidentifi-cation, the insect would, at best, more likely have been S. spartani Eichlin & Taft, recently described from the central Lower Peninsula of Michigan. Welch (1915) also listed “Sesia rutilans Hy. Edw. (?)”; this name is now placed in the synonymy of Synanthedon bibionopennis (Bdv.), but as that species is not known from Michigan or elsewhere in eastern North America and the identification was questioned, the report would seem safely ignored.
2549. Synanthedon scitula (Harr.). Emmet, Cheboygan: 30 June–17 August. The larvae have been reported from a wide range of hosts in eastern North America.
2550. Synanthedon pictipes (G. & R.). Emmet, Cheboygan: 30 June–22 July. The larvae attack species of Prunus (cherry, plum), of which there are sev-eral in the region.
2554. Synanthedon acerni (Clem.). Emmet, Cheboygan: 17 June–8 August. As noted by Covell (1984) and others, this is the only species in the family commonly attracted to light. Purrington and Horn (1997) reported a con-siderable number, of both sexes, at UV light in southern Ohio; Eichlin and Duckworth (1988) mention capture “regularly at black light.” I have also taken it at mercury vapor, carbon arc, and incandescent light. Maple (Acer)

2002 THE GREAT LAKES ENTOMOLOGIST 57
is the larval host, with red maple (Acer rubrum) apparently preferred — a common species in the Douglas Lake region.
2565. Synanthedon pyri (Harr.). Emmet: 14 July 1992. Collected by Scholtens at the Sturgeon Bay dunes. The larvae bore under the bark of woody plants in the family Rosaceae subfamily Maloideae (e.g., apple, pear, serviceberry, hawthorn).
2572. Synanthedon proxima (Hy. Edw.). Cheboygan: 23 July 1995. Collected by Scholtens at Grass Bay. The larvae bore in willows.
2583. Synanthedon exitiosa (Say). Emmet, Cheboygan: 26 June–9 August. Another species collected only by Scholtens, at several sites; some attracted by pheromone lure GPTB. This is the notorious “peachtree borer” (noted also for attacking other species of Prunus).
2585. Synanthedon pini (Kellicott). Emmet: 25 June 1991. Collected by Schol-tens, using pheromone lure ZZOH/EZOH, near Van (north of Pellston). The larvae bore in pines and spruces.
2592. Carmenta anthracipennis (Bdv.). Cheboygan: 4 July 1991. Collected by Scholtens on the Wildwood jack pine plains south of Indian River — the only site known in the two-county region where there grows any native species of blazing-star (Liatris cylindracea), the genus in the underground corm of which the larvae of this species bore. Nielsen has collected the species 2–7 August in Otsego County, about 30 miles due south of the Cheboygan County site and in an area where L. scariosa occurs.
2608. Carmenta pyralidiformis (Wlk.). Emmet, Cheboygan: 30 June–10 August. One of my specimens was taken at the light of a gasoline lantern. An old undated specimen in UMMZ, labeled merely “Douglas Lake,” was presum-ably collected by Welch although he did not list this species (1915). The larva is reported as boring in the roots of the common boneset (Eupatorium perfoliatum) and allied species.
2623. Alcathoe caudata (Harr.). Emmet: 31 July–August. Two specimens in UMBS, the August one without day, the July one taken in 1958 three miles west of Carp Lake. The larvae are known to attack only Clematis vines. Males of this species are easily recognized by the long yellow “tail” abdominal appendage.
Family Cossidae2675. Acossus centerensis (Lint.). Cheboygan: 21 June–8 August. Males are
much more frequent than females at light, where the species is occasional, including incandescent, mercury vapor, ultraviolet, and carbon arc light. The larvae are reported elsewhere as borers principally in poplar (presumably, in our area, the abundant aspens: Populus grandidentata and P. tremuloides). Surprisingly, this relatively large moth, conspicuous (though scarcely com-mon) at lights, receives no mention by Covell (1984).
2693. Prionoxystus robiniae (Peck). Emmet, Cheboygan: 17 June–17 July. In contrast to the preceding, females are more frequent than males at light, including incandescent and carbon arc. However, Munro and Fox (1934) observed that in North Dakota the “male is very active in flight,” while the female “appears to be very limited in her powers of flight,” remaining on the trunk of the tree where she emerged from the pupa, mating and oviposit-ing the same evening. Holland (1903) noted the species to be “exceedingly abundant” locally at electric lights.
2694. Prionoxystus macmurtrei (Guér.). 17 June 1991. One female taken at ultraviolet light at the Biological Station by David Cowan and in the Schol-tens collection.

58 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Family Limacodidae4652. Tortricidia testacea Pack. Emmet, Cheboygan: 30 May–12 July. Taken
at incandescent, ultraviolet, and mercury vapor light.4654. Tortricidia flexuosa (Grt.). Emmet, Cheboygan: 21 June–27 July. Taken
at incandescent, carbon arc, ultraviolet, and mercury vapor light. Apparently more common than the preceding, and often on the wing with it.
4659. Packardia geminata (Pack.). Cheboygan: 22 June–4 July. Apparently rare, taken at mercury vapor and ultraviolet light.
4665. Lithacodes fasciola (H.-S.). Cheboygan: 30 June–30 July. Not common; collected at fluorescent and ultraviolet light.
4669. Apoda biguttata (Pack.). Cheboygan: 24 June–20 July. Taken at carbon arc, mercury vapor, and ultraviolet light.
4697. Euclea delphinii (Bdv.). Cheboygan: 18 June–30 July. Taken at both mercury vapor and ultraviolet light.
Family Thyrididae6076. Thyris maculata Harr. Emmet, Cheboygan: 26 June–20 July. This dis-
tinctive moth with black wings, spotted both white and yellow and scalloped along the margins, is generally considered uncommon. Scholtens has taken specimens mostly visiting wild parsnip (Pastinaca sativa) along the Hebron Mail Route Road in Hebron Township east of Dingman Marsh. Specimens in UMBS are from the Topinabee area. The Emmet County record is a single individual observed (but not captured) by Scholtens and Voss 20 July 2001; it was visiting fresh, black, glistening, amorphous dung (possibly bear) at Cecil Bay. Clematis, the presumed larval food plant here, is however quite widespread in the region.
Family PterophoridaeThe identifications in this difficult family, in which he has specialized,
have all been checked (or made) by Reed A. Watkins, whose labors are deeply appreciated and were facilitated at the National Museum of Natural History by M. Alma Solis and Jon Lewis. While the numbers (with two intercalations) are from the MONA checklist, the generic assignments follow the determinations of Watkins, based on the work of Gielis (1993).6091.1. Geina sheppardi B. Landry. Cheboygan: 29 June–11 July. Reared
from Vitis riparia (river-bank grape) by Scholtens and also taken by him at UV light at the Biological Station. This recently described species (Landry 1989) has long masqueraded as G. periscelidactyla (Fitch). No specimen of the latter has been found to support the listing by Welch (1915, as Oxyptilus periscelidactyla), reported with a question-mark, from the Douglas Lake region.
6092. Geina tenuidactyla (Fitch). Cheboygan: 23 June–14 August. Sometimes stirred up in fields.
[———]. Capperia sp. Cheboygan: 29 June 1956, Emmet: 4 August 1949. Two female specimens appearing to represent a species in this genus have not yet been definitely placed. The Cheboygan County specimen (June, Scholtens) is from the vicinity of Reese’s Bog near the Biological Station.
6102. Dejongia lobidactyla (Fitch). Cheboygan: 15 July–3 August. At UV and mercury vapor light. The larva is reported (Barnes & Lindsey 1921) to feed on Solidago (goldenrod), of which there is no shortage in the region.
6105. Cnaemidophorus rhododactylus (D. & S.). Cheboygan: 30 June–21 July.

2002 THE GREAT LAKES ENTOMOLOGIST 59
Taken only by Scholtens at UV light.6107. Gillmeria pallidactyla (Haw.). Emmet, Cheboygan: 18 June–30 July.
Clearly our commonest species at light, including incandescent, UV, and gasoline lantern.
6109. Platyptilia carduidactyla (Riley). Cheboygan: 4–21 July. Collected only by Scholtens at UV light and reared from thistle by a Biological Station ecology class.
6149. Stenoptilia coloradensis Fern. Emmet: 28 June 1946. Two specimens (male & female) at incandescent light at Mackinaw City.
6157. Adaina montana (Wlsm.). Emmet: 5–6 August 1945. Two specimens at incandescent light at Mackinaw City.
6163. Oidaematophorus cretidactylus (Fitch). Emmet, Cheboygan: 30 June–13 August. Another rather common species, taken at incandescent light and gasoline lantern.
6168. Oidaematophorus eupatorii (Fern.). Cheboygan: 6–9 August. One female taken in 1996 by Scholtens at Colonial Point on Burt Lake and another fe-male (UMMZ) by Peet at Burt Lake in 1932. The larvae are gregarious on Eupatorium (Barnes & Lindsey 1921), of which two species are abundant in the region, but this moth seems extremely rare here.
6203. Hellinsia homodactyla (Wlk.). Emmet, Cheboygan: 6 July–14 August. Taken at incandescent light, UV, and gasoline lantern.
6204 Hellinsia elliottii (Fern.). Emmet, Cheboygan: 31 July–19 August. One male taken (by me in 1950) at gasoline lantern light and one female net-ted (by Scholtens in 1994), both near the Biological Station. Also a female (UMMZ) taken at Burt Lake by Max M. Peet in 1932.
6205. Hellinsia pectodactyla (Stgr.). Emmet, Cheboygan: 5–14 August. This spe-cies includes what was formerly recognized as Oidaematophorus stramineus.
6214. Hellinsia glenni (Cashatt). Cheboygan: 29 June 1956. One specimen taken by Scholtens in the vicinity of Reese’s Bog near the Biological Station.
6234. Emmelina monodactyla (L.). Emmet, Cheboygan: 18 June–18 July. Two specimens, both female, the former (June, Emmet) at incandescent light at Mackinaw City and the latter at mercury vapor light at the Biological Station.
Family Epiplemidae7650. Callizzia amorata Pack. Emmet, Cheboygan: 26 June–7 July. The only
collections from the Douglas Lake region are by Scholtens, at ultraviolet light. The only Michigan specimens in UMMZ are from neighboring coun-ties, Mackinac (St. Ignace) and Charlevoix (Beaver Islands) — as reported by Moore (1955). The only Michigan specimen in MSU is from Isle Royale (in 1957). Apparently a rare species in the state.
7653. Calledapteryx dryopterata Grt. Cheboygan: 5 July 1997. One specimen, taken by Scholtens at ultraviolet light at Colonial Point, on Burt Lake. An-other rare (or overlooked) species. The only Michigan specimens in MSU are two from Livingston County (1966) and there are none in UMMZ.
ACKNOWLEDGMENTSBrian Scholtens has been of tremendous help, especially with determina-
tions of Sesiidae, suggestions, making available the UMBS collection, and sup-plying all records from his own collection. Without the immense help of Reed A. Watkins, who examined all Pterophoridae in the Scholtens, Voss, UMMZ, and UMBS collections, that family could have been treated only poorly if at all.

60 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Mark O’Brien made available the UMMZ collection and facilitated loans. M.C. Nielsen did likewise for the MSU collections; he also read an early version of the manuscript, as did George Balogh, but neither could add any records from their own collections. William Taft examined my Sesiidae many years ago and Bernard Landry clarified the present situation with Alucitidae. My collecting at Mackinaw City over more than half a century has been encouraged by many friends, neighbors, and family members. I leave the remaining groups of mi-crolepidoptera to others to write up.
LITERATURE CITEDBarnes, W., and A. W. Lindsey. 1921. The Pterophoridae of America, north of Mexico.
Contr. Nat. Hist. Lepid. N. Am. 4: 279–428 + pl. 41–54 + [4] pp. index.Beutenmüller, W. 1901. Monograph of the Sesiidae of America, North of Mexico. Mem.
Am. Mus. Nat. Hist. 1(6): 215–352 + pl. 29–36.Covell, C. V., Jr. 1984. A Field Guide to the Moths of Eastern North America. Houghton
Mifflin, Boston. 496 pp.Davis, D. R. 1964. Bagworm moths of the Western Hemisphere (Lepidoptera: Psychidae).
U. S. Natl. Mus. Bull. 244. 233 pp.Dirig, R. 1993. McLean’s mysterious ghost moths. News Lepid. Soc. 1993: 102–103.Eichlin, T. D., and W. D. Duckworth. 1988. Sesioidea Sesiidae. Moths Am. N. Mex.
Fasc. 5.1. 176 pp.Engelhardt, G. P. 1946. North American clear-wing moths of the family Aegeriidae. U.
S. Natl. Mus. Bull. 190. 222 pp. + 32 pl.Ferge, L. A. 2001. Zone 8 Midwest. News Lepid. Soc. 43 (suppl. 1): 41–50.Forbes, W. T. M. 1923. Lepidoptera of New York and neighboring states. Primitive forms,
Microlepidoptera, Pyraloids, Bombyces. Cornell Agric. Exp. Sta. Mem. 68. 729 pp.Gielis, C. 1993. Generic revision of the superfamily Pterophoroidea (Lepidoptera). Zo-
ologische Verhandelingen 290. 139 pp.Handfield, L. 1999. Le Guide des Papillons du Québec. Vol. 1. Brocquet, Boucherville.
982 pp. + 125 pl. + map + corrections.Hodges, R. W., T. Dominick, D. R. Davis, D. C. Ferguson, J. G. Franclemont, E. G. Munroe,
and J. A. Powell. 1983. Checklist of the Lepidoptera of America North of Mexico. E. W. Classey, London. 284 pp.
Holland, W. J. 1903. The Moth Book. Doubleday, New York. 479 pp. + 48 pl. [Reprinted 1968 by Dover Publ., New York.]
Landry, B. 1989. Geina sheppardi, a new species of grape-feeding Platyptylinae (Lepi-doptera: Pterophoridae) from eastern North America, with notes on related species. Can. Entomol. 121: 763–770.
Layberry, R. A. 1994. Zone 7: Ontario, Quebec. News Lepid. Soc. 1994(2): 37.Layberry, R. A. 1996. Zone 7 Ontario/Quebec. News Lepid. Soc. 38(3): 41–43.McDunnough, J. 1938. Check list of the Lepidoptera of Canada and the United States of
America. Part 1 Macrolepidoptera. Mem. So. Calif. Acad. Sci. 1. 272 pp.Moore, S. 1955. An annotated list of the moths of Michigan exclusive of the Tineoidea
(Lepidoptera). Misc. Publ. Mus. Zool. Univ. Mich. 88. 87 pp.Munro, J. A., and A. C. Fox. 1934. Carpenter worm biology and control. North Dakota
Agric. Exp. Sta. Bull. 278. 23 pp.Nielsen, E. S., G. S. Robinson, and D. L. Wagner. 2000. Ghost-moths of the world: a
global inventory and bibliography of the Exoporia (Mnesarchaeoidea and Hepialoidea) (Lepidoptera). J. Nat. Hist. 34: 823–878.

2002 THE GREAT LAKES ENTOMOLOGIST 61
Nielsen, M. C. 1998. Preliminary list of Michigan moths: the Microlepidoptera. Newsl. Michigan Entomol. Soc. 43(4): 1, 4–14.
Purrington, F. F., and D. J. Horn. 1997. Clearwing moths captured by ultraviolet light traps in southern Ohio (Lepidoptera: Sesiidae). Great Lakes Entomol. 29: 191–193.
Rings, R. W., and E. H. Metzler. 1990. The Lepidoptera of Fowler Woods State Nature Preserve, Richland County, Ohio. Great Lakes Entomol. 23: 43–56.
Schmidt, B. C., and D. D. Lawrie. 2000. Notes on the genus Sthenopis (Hepialidae) in Alberta, Canada. Jour. Lepid. Soc. 53: 127–129.
Scholtens, B. 1996. Moths of the Douglas Lake Region (Emmet and Cheboygan Coun-ties), Michigan: V. Crambidae and Pyralidae (Lepidoptera). Great Lakes Entomol. 29: 141–160.
Taft, W. H., D. Smitley, and J. W. Snow. 1991. A guide to the clearwing borers (Sesiidae) of the north central United States. USDA Forest Service North Central Regional Publ. 394. 30 pp.
Voss, E. G. 1954. The butterflies of Emmet and Cheboygan counties, Michigan, with other notes on northern Michigan butterflies. Am. Midl. Nat. 51: 87–104.
Voss, E. G., and W. H. Wagner, Jr. 1956. Notes on Pieris virginiensis and Erora laeta — two butterflies hitherto unreported from Michigan. Lepid. News 10: 18–24. 1956.
Voss, E. G. 1969. Moths of the Douglas Lake region (Emmet and Cheboygan Counties), Michigan: I. Sphingidae–Ctenuchidae (Lepidoptera). Newsl. Michigan Entomol. Soc. 2: 48–54.
Voss, E. G. 1972. Michigan Flora. A Guide to the Identification and Occurrence of the Native and Naturalized Seed-plants of the State. Part I Gymnosperms and Mono-cots. Cranbrook Inst. Sci. Bull. 55 & Univ. Michigan Herbarium. xviii + 488 pp.
Voss, E. G. 1981. Moths of the Douglas Lake region (Emmet and Cheboygan counties), Michigan: II. Noctuidae (Lepidoptera). Great Lakes Entomol. 14: 87–101.
Voss, E. G., and R. E. Riefner, Jr. 1983. A Pyralid moth (Lepidoptera) as pollinator of blunt-leaf orchid. Great Lakes Entomol. 16: 57–60.
Voss, E. G. 1983. Moths of the Douglas Lake region (Emmet and Cheboygan counties), Michigan: III. Thyatiridae, Drepanidae, Lasiocampidae, Notodontidae, Lymantriidae (Lepidoptera). Great Lakes Entomol. 16: 131–137.
Voss, E. G. 1985. Michigan Flora. ... Part II Dicots (Saururaceae – Cornaceae). Cranbrook Inst. Sci. Bull. 59 & Univ. Michigan Herbarium. xx + 724 pp.
Voss, E. G. 1991. Moths of the Douglas Lake Region (Emmet and Cheboygan Counties), Michigan: IV. Geometridae (Lepidoptera). Great Lakes Entomol. 24: 187–201.
Voss, E. G. 1996. Michigan Flora. ... Part III Dicots (Pyrolaceae – Compositae). Cran-brook Inst. Sci. Bull. 61 & Univ. Michigan Herbarium. xii + 622 pp.
Welch, P. S. 1915. The Lepidoptera of the Douglas Lake region, northern Michigan. Entomol. News 36: 115–119.

62 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1

2002 THE GREAT LAKES ENTOMOLOGIST 63
TARSAL ANOMALY FOUND IN AN ADULT AGABUS SERIATUS (COLEOPTERA: DYTISCIDAE)
J.L. Snitgen1
ABSTRACTAn adult specimen of Agabus seriatus, possessing two complete sets
of tarsal claws on the left mesotarsus, was collected from a small stream in northeastern Wisconsin.
____________________ An adult specimen of Agabus seriatus (Say) was collected in a benthos
sample from Silver Creek, a second order stream in Brown County, Wisconsin. The sample was collected during a 2000 baseline qualitative study conducted by Oneida Tribe of Indians Water Resources staff. The anomaly, located on the left mesotarsus (Fig. 1) was discovered while identifying the specimen. Although deformities of chironomid menta due to heavy metals and other contaminants are well documented (Diggins and Stewart 1993, Warwick 1988) and deformities and arrested development have been produced in terrestrial insects due to exposures with pesticides (Arthur 2001), a literature search produced no published records describing this type of anomaly. Personal communications with coleopteran workers indicated this type of anomaly has not been encountered previously.
This species is common in the southern third of Wisconsin, uncommon in the northern two-thirds. They are collected most often from lotic habitats, especially small, spring-fed streams (Hilsenhoff 1993). The beetles usually occur on mineral substrates such as clay, sand or gravel but may also be in clumps or mats of vegetation in or at the edge of flowing water. Beetles seldom occur in depositional areas or on silty or peaty substrates (Larson et al. 2000). Both larvae and adults within this genus are predators, mainly on other aquatic arthropods; adults are also scavengers (Hilsenhoff, 1995).
Sampling of aquatic invertebrates was performed at the site as part of an Oneida Reservation-wide baseline study of various streams and lakes to even-tually develop biocriteria for Tribal Water Quality Standards. Sampling was performed using an aquatic dip net (traveling kicks and sweeps of aquatic vegeta-tion and undercut banks), and hand held examination and scrubbing of stones and partially decayed wood within the sample area. Substrates were sampled so that aquatic invertebrates from all of the significant microhabitats within the sample were represented in the composite sample of the site (Oneida Tribe of Indians SOP BI002). Organisms were identified to the lowest possible taxa. Adults of Agabus seriatus are identified by their lotic habitat, relatively large size and elongate shape, their almost uniform black color, and small meshes on the elytra that often contain micropunctures of varying sizes (Hilsenhoff 1993).
ACKNOWLEDGMENTSThanks are due to K.L. Schmude for his confirmation of my determination
of Agabus seriatus and for review of the manuscript. Thanks are also due to two anonymous reviewers for their comments and to Stewart Cogswell for the use of the camera equipment.
1Environmental, Health and Safety, Oneida Tribe of Indians of Wisconsin, Oneida, WI 54155.

64 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
LITERATURE CITEDArthur, A. H. 2001. Efficacy of a volatile formulation of hydroprene (Pointsource™) to
control Tribolium castaneum and Tribolium confusum (Coleoptera: Tenebrionidae). Journal of Stored Products Research 39: 205-212.
Diggins, T.P., and K.M. Stewart. 1993. Deformities of aquatic larval midges (Chiron-omidae: Diptera) in the sediments of the Buffalo River, New York. J. Great Lakes Res. 19:648-659.
Hilsenhoff, W.L. 1993. Dytiscidae and Noteridae of Wisconsin (Coleoptera). IV. Dis-tribution, habitat, life cycle, and identification of species of Agabini (Colymbetinae). Great Lakes Entomol. 26:173-197.
Hilsenhoff, W.L. 1995. Aquatic Insects of Wisconsin. Keys to Wisconsin genera and notes on biology, distribution and species. Natural Hist. Museums Council, Univ. Wisconsin-Madison, Publ. No. 3.
Larson, D.J., Alarie, Y., and Roughley, R.E. 2000. Predaceous diving Beetles (Coleoptera:Dytscidae) of the Neartic Region, with emphasis on the fauna of Canada and Alaska. NRC Research Press, Ottawa, Ontario, Canada. 982 pp.
Oneida Tribe of Indians of Wisconsin. 2001. Standard Operating Procedure for the Qualitative Sampling of Streams for Benthic Invertebrates (BI002). Oneida Water Resources Program.
Warwick, W.F. 1988. Morphological deformities in Chironomidae (Diptera) larvae as well as biological indicators of toxic stress, pp. 281-320. In M.S. Evans (ed.) Toxic contaminants and ecosystem health: A Great Lakes focus. Advances in Environmental Science and Technology. Vol. 21. John Wiley and Sons, New York, NY. 602 pp.
Figure 1. Photograph of the left mesotarsus of a specimen of A. seriatus, illustrating tarsal anomaly.

2002 THE GREAT LAKES ENTOMOLOGIST 65
OCCURRENCE OF TWO SPECIES OF OLD WORLD BEES, ANTHIDIUM MANICATUM AND A. OBLONGATUM (APOIDEA: MEGACHILIDAE), IN NORTHERN OHIO
AND SOUTHERN MICHIGAN
Shane R. Miller1, Robert Gaebel1, Randall J. Mitchell1, and Mike Arduser2
ABSTRACTAnthidium manicatum and A. oblongatum are two European bees species
that have recently established themselves in North America. Anthidium mani-catum has previously been documented in New York and Ontario, Canada, and A. oblongatum has been documented in New York, New Jersey, Maryland, and eastern Pennsylvania. We surveyed a number of sites in Ohio, Michigan, and Indiana for these species in 2000 and 2001, and found both bee species to have extended their ranges into northern Ohio, and A. manicatum to have moved into southern Michigan. We present a key identifying the four Anthidium species now known from northeastern North America.
____________________A recent report estimates that approximately 4,500 species of non-in-
digenous organisms (“exotics”) are now established in the United States (U.S. Congress 1993). Insects, including several bees, make up a significant part of this total (U.S. Congress 1993, Ascher 2001). The potential impacts of some of these exotic bees on native bees, and on the wild and cultivated plants that depend on native bees for pollination, may be cause for concern (Buchmann and Nabhan 1996). As part of an effort to track the movements and distribu-tion of nonindigenous species, we report here the first occurrence in Ohio and Michigan of two Old World bees, Anthidium (Anthidium) manicatum (L.) and A. (Proanthidium) oblongatum (Illiger) (Megachilidae, Anthidiini).
The Wool-Carder Bee (A. manicatum), a robust hymenopteran native to Europe, is renowned for its highly aggressive territorial behavior of the males (Severinghaus et al. 1981, Wirtz et al. 1988, Starks and Reeve 1999). This cavity-nesting bee was introduced to New York State sometime before 1963 (Jaycox 1967), and by 1990 had spread to Ontario, Canada (Smith 1991). This species has also invaded various other locales outside its native range, including Brazil, Argentina, Uruguay, and the Canary Islands (Hoebeke and Wheeler 1999).
We first collected A. manicatum in the Akron, Ohio area (Summit County) in 1996, and in the following years noticed that this species was common in many urban and suburban gardens near Akron. Therefore, during summer 2000 and 2001 we searched and collected throughout northern Ohio and adjacent areas in Michigan and Indiana (June - September) to better understand the current dis-tribution of this invader. We chose collecting sites based on convenience, access and permission, and on presence of known or suspected food plants (primarily Lamiaceae). We spent at least two hours at each site, and visited many sites multiple times on separate days. Voucher specimens are deposited at the Ento-mology collection, University of Akron, and at the Enns Entomological Museum, University of Missouri - Columbia. We found A. manicatum to be common in urban and rural gardens throughout northern Ohio and at one site in southern Michigan (Table 1), but we did not find it in any of the several parks and natural areas we studied. Notably, we were unable to find any A. manicatum at our original 1996
1Department of Biology, Program in Integrative Biology, University of Akron, Akron, OH 44325-39082 Missouri Department of Conservation, 2360 Highway D, St. Charles, MO 63304

66 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1Ta
ble
1. R
esul
ts o
f sea
rch
for A
nthi
dium
spp
. in
nort
hern
Ohi
o an
d ad
jace
nt a
reas
, 199
7-20
01. U
R (u
rban
resi
dent
ial),
UP
(urb
an p
ark)
, RR
(rur
al re
side
ntia
l), a
nd R
P (r
ural
par
k).
Stat
e C
ount
y P
lace
H
abit
at
Coo
rdin
ates
A
nthi
dium
sp.
N
umbe
r D
ate
Pla
nts
visi
ted
pres
ent
Col
lect
ed O
H
Sand
usky
Ba
llvill
e RR
Gar
den
N 4
1.31
806
W 8
3.14
910
A. m
anic
atum
1
6/30
/200
0 St
achy
s by
zant
ina
OH
Sa
ndus
ky
Frem
ont
UR
Gar
den
N 4
1.34
890
W 8
3.09
548
A. m
anic
atum
3
6/23
/200
0 St
achy
s by
zant
ina
OH
Sa
ndus
ky
Gre
en S
prin
gs
UR
Gar
den
N 4
1.25
949
W 8
3.04
696
A. m
anic
atum
1
7/29
/200
1 St
achy
s by
zant
ina
OH
Se
neca
Cl
yde
RR G
arde
n N
41.
2265
3 W
83.
0255
4 A
. man
icat
um
6 6/
30/2
000
Stac
hys
byza
ntin
a
7
/7/2
000
7/15
/200
0 O
H
Sum
mit
Fran
klin
Tw
p U
R G
arde
n N
40.
930
40 W
81.
5427
2 A
. man
icat
um,
3 7/
23/2
000
Sene
cio
cine
rari
a
8/
4/20
00
A. o
blon
gatu
m
2 7/
28/2
000
OH
Su
mm
it U
nive
rsity
of
Ak
ron,
Akr
on
UP
Gar
den
N 4
1.07
679
W 8
1.51
106
A. m
anic
atum
6
8/5/
1998
Pe
rovs
kia
atri
plic
ifolia
8/13
/199
8
8/
19/1
998
8/26
/199
8 O
H
Sum
mit
Wes
t Akr
on
UR
Gar
den
N 4
1.06
646
W 8
1.32
030
A. m
anic
atum
4
7/3/
1997
St
achy
s by
zant
ina,
6/28
/200
0 P
enst
emon
dig
italis
OH
Su
mm
it Ba
th T
owns
hip
Wild
Par
k N
41.
1055
1 W
81.
3856
1 Ab
sent
O
H
Sum
mit
Bos
ton
Mill
s Si
te 1
W
ild P
ark
N 4
1.25
580
W 8
1.53
907
Abse
nt
OH
Su
mm
it Bo
ston
Mill
s Si
te 2
W
ild P
ark
N 4
1.25
927
W 8
1.52
192
Abse
nt
OH
Su
mm
it Bo
ston
Mill
s Si
te 3
W
ild P
ark
N 4
1.25
758
W 8
1.54
059
Abse
nt
OH
Su
mm
it Bo
ston
Mill
s Si
te 4
W
ild P
ark
N 4
1.25
580
W 8
1.53
907
Abse
nt
OH
Su
mm
it Bo
ss P
ark,
Akr
on
UP
Gar
den
N 4
1.06
834
W 8
1.51
503
Abse
nt
OH
Lu
cas
Tole
do
UR
Gar
den
N 4
1.89
850
W 8
3.22
721
A. m
anic
atum
1
7/27
/200
1 St
achy
s by
zant
ina
O
riga
num
her
acle
oti-
cum
IN
LaG
rang
e Sh
ipsh
ewan
a U
R G
arde
n N
41.
6785
1 W
85.
5795
2 Ab
sent
M
I Le
naw
ee
Tipt
on
Rura
l Gar
den
N 4
2.02
927
W 8
4.11
032
A. m
anic
atum
1
7/28
/200
1 St
achy
s by
zant
ina

2002 THE GREAT LAKES ENTOMOLOGIST 67
collection site (Boston Mills Site 1 in Table 1). We did not find A. manicatum at our one Indiana site, perhaps indicating the edge of the range expansion to date.
In agreement with other published reports (Severinghaus et al. 1981), we often found A. manicatum females associated with plants with downy leaf pubescence, such as cultivated Stachys byzantina K. Koch ex Scheele (Lamb’s Ears, Lamiaceae). Female bees use trichomes from this plant as nest-building material (Smith 1991). We most commonly observed A. manicatum visiting flowers and leaves of Lamb’s Ears, and occasionally flowers of Penstemon digi-talis Nutt. ex Sims. (Foxglove Beardtongue, Scrophulariaceae) and Perovskia atriplicifolia Benth. (Russian Sage, Lamiaceae) and leaves of Senecio cineraria DC. (Dusty Miller, Asteraceae).
Both male and female A. manicatum were very abundant wherever we found them; several individuals were often visible at once. Males defended territories near floral resources at all sites (see also Starks and Reeve 1999), and we frequently observed them aggressively defend these territories against other bee species, including honey bees (Apis mellifera (L.)) and bumble bees (Bombus spp.). Many male Anthidium species have prominent spines on the apical margin of tergum 7 which they use in territorial defense, in the case of A. manicatum often killing or disabling honey bees and other bee species (Wirtz et al. 1988, Starks and Reeve 1999).
A second species of Old World Anthidium, A. oblongatum, was discovered in North America (eastern Pennsylvania) in 1995, and has since been found in several other eastern states (New York, New Jersey, and Maryland) (Hoebeke and Wheeler 1999). We only collected this species at the Franklin Township site (Summit County). Here females were gathering trichomes from Dusty Miller (Senecio cineraria), in the same areas where we also caught A. manicatum. We also found a single specimen of A. oblongatum in the University of Akron student Entomology collection, from southern Stark County (Massillon; collected in 1999).
These new records extend the US ranges of A. manicatum and A. oblon-gatum into the upper midwest, where they have not been previously reported (Hoebeke and Wheeler 1999). It seems possible that one or both of these “hitch-hiking” species may be established in areas further west, such as Chicago and Indianapolis. Bee surveys in these and other urban areas are needed. Casual examinations of likely sites in southern Wisconsin and St. Louis, Missouri, how-ever, have not revealed either species (RJM and MSA, personal observations).
Continued spread of these two adventive species, which seems likely, will bring them into the ranges of some of our native Anthidium. Most North American Anthidium occur in the western states, but two native species are known from parts of the northeastern quarter of North America (Hurd 1979). Anthidium (Anthidium) psoraleae Robertson, primarily an oligolege of Fabaceae found in central US prairies and prairie-like areas, occurs as far east as south-western Michigan in remnant prairie communities (M. Arduser, unpublished data). Anthidium (A.) maculifrons Smith, also an apparent oligolege of Fabaceae, is a species of the southern US, but has been found as far north as southern Illinois (although not recently; John Marlin, in litt.), and in southern Missouri (M. Arduser, unpublished data). Both native species seem dependent on natural communities and native plants, and are unlikely to be found in disturbed sites, urban areas, or manicured gardens of non-native plants, habitats apparently favored by the two adventive species (Hoebeke and Wheeler 1999, this study).
As a guide to the identification of the four species of Anthidium now known from the northeastern quarter of North America, we offer the following key [detailed descriptions and illustrations of the two native species (Anthidium psoraleae and A. maculifrons) can be found in Mitchell (1962), of A. oblongatum in Hoebeke and Wheeler (1999); Jaycox (1967) provides brief descriptive notes on A. manicatum].

68 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Key to Anthidium species in northeastern North America:
1. Females (scopa and sting present) ................................................................................ 2Males (scopa and sting absent) ..................................................................................... 5
2. (Females): posterior margin of tergum 6 with a conspicuous projection medially, laterally with flattened, triangular teeth (one on each side) ............................................................................psoraleae Robertson Posterior margin of tergum 6 rounded, without conspicuous projections (though very small teeth or denticles may be present laterally) ............................................................................................................ 3
3. Pronotal lobe divided by a thin, raised plate (lamella) into dorsal and anterior halves; basal mandibular tooth noticeably larger than apical mandibular tooth .........................................................oblongatum (Illiger)Pronotal lobe rounded, not divided by a lamella; basal mandibular tooth similar in size to apical tooth .......................................................... 4
4. Clypeal margin convex and denticulate in dorsal view, produced slightly beyond lower ends of eyes; hairs on surface of clypeus (@ 60x) dense and hooked apically .........................manicatum (L.)Clypeal margin essentially truncate (margin somewhat thickened, with irregular crenulations), not produced beyond lower ends of eyes; hairs on surface of clypeus dense but straight throughout, not hooked or bent apically ........................... maculifrons Smith
5. (Males): posterior margin of tergum 7 bi-lobed, without a median spine; posterior margin of tergum 6 with a small pointed projection medially ...........................................................................................oblongatum (Illiger)Posterior margin of tergum 7 with a distinct spine between the two lateral lobes; posterior margin of tergum 6 entire, without a projection medially. ....................................................................................... 6
6. Lateral margins of tergites 1-5 with conspicuous curly hair tufts, brown to pale brown in color, to some extent obscuring integument; clypeal margin apically with a pair of small teeth lateral to midline, best seen in oblique lateral view. ............................manicatum (L.)Lateral margins of tergites 1-5 without conspicuous curly hair tufts; clypeal margin apically simple, without small teeth ......................................... 7
7. Scape wholly maculated with yellow or whitish-yellow; tergum 6 punctures separated by 1-3 puncture widths; mandibles robust, all three teeth similar in size ............................ maculifrons SmithScape wholly dark; tergum 6 punctures fine, dense, many contiguous or nearly so; mandibles slender, tapering, basal tooth larger than other teeth ..........................................................psoraleae Robertson
ACKNOWLEDGEMENTSWe thank the Cuyahoga Valley National Park; MetroParks serving Sum-
mit County; Bath Field Station and Bath Township; Sandy and Robert Kessler; John and Joy Lin; Will, Mary, and Jessie Miller; Lisa Nye; and the Gaebel Estate for permission to collect and search for bees on their land. We also thank the Biology Department of the University of Akron for support and materials, Dr. Robert Sites and Kristin Simpson, curator and collection manager, respectively, of the Enns Entomological Museum at the University of Missouri - Columbia, for permission to search their holdings for anthidiine bees, Sarah Popoca for aiding in the field study, and the helpful suggestions of E. Richard Hoebeke and two anonymous reviewers.

2002 THE GREAT LAKES ENTOMOLOGIST 69
LITERATURE CITED Ascher, J. S. 2001. Hylaeus hyalinatus Smith, a European bee new to North America,
with notes on other adventive bees (Hymenoptera: Apoidea). Proceedings of the Entomological Society of Washington 103:184-190.
Buchmann, S.L. and G.P. Nabhan. 1996. The Forgotten Pollinators. Island Press, Wash-ington, DC. 292 p.
Hoebeke, E.R. and A.G. Wheeler, Jr. 1999. Anthidium oblongatum (Illiger): an Old World bee (Hymenoptera: Megachilidae) new to North America, and new North American records for another adventive species, A. manicatum (L.). The University of Kansas Natural History Museum Special Publication 24: 21-24.
Hurd, P.D., Jr. 1979. Family Megachilidae, Pp. 1981-2081. In K.V. Krombein, P.D. Hurd, Jr., D.R. Smith, and B.D. Burks (eds.), Catalog of Hymenoptera in America North of Mexico. Volume 2: Apocrita (Aculeata). Smithsonian Institution Press, Washington, DC.
Jaycox, E.R. 1967. An adventive Anthidium in New York State (Hymenoptera: Megach-ilidae). Journal of the Kansas Entomological Society 40:124-126.
Mitchell, T.B. 1962. Bees of the Eastern United States, Volume 2. North Carolina Agri-cultural Experiment Station, Raleigh Technical Bulletin 152. 557 p.
Severinghaus L.L., B.H. Kurtak and G.C. Eickwort. 1981. The reproductive behavior of Anthidium manicatum (Hymenoptera: Megachilidae) and the significance of size for territorial males. Behavioral Ecology and Sociobiology 9:51-58.
Smith, I. 1991. Anthidium manicatum (Hymenoptera: Megachilidae), an interesting new Canadian record. Proceeding of Entomological Society of Ontario 122:105-108.
Starks, P.T. and H.K. Reeve. 1999. Condition-based alternative reproductive tactics in the wool-carder bee, Anthidium manicatum. Ethology Ecology and Evolution 11: 71-75.
U.S. Congress, Office of Technology Assessment. 1993. Harmful Non-Indigenous Species in the United States, OTA-F-565 (Washington, DC: U.S. Government Printing Office, September 1993). 391 p.
Wirtz, P., M. Szabados, H. Pethig, and J. Plant. 1988. An extreme case of interspecific territoriality: male Anthidium manicatum (Hymenoptera: Megachilidae) wound and kill intruders. Ethology 78:159-167.

70 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1

2002 THE GREAT LAKES ENTOMOLOGIST 71
TRAP RESPONSE OF MICHIGAN SOCIAL WASPS (HYMENOPTERA: VESPIDAE) TO THE FEEDING ATTRACTANTS
ACETIC ACID, ISOBUTANOL, AND HEPTYL BUTYRATE.
H. C. Reed 1 and P. J. Landolt2
ABSTRACTNine species of social wasps were captured in traps baited with acetic acid,
isobutanol, heptyl butyrate and combinations of acetic acid and either isobuta-nol or heptyl butyrate. Three yellowjacket species in the Vespula rufa species group were captured in traps (Vespula acadica (Sladen), Vespula consobrina (Saussure), Vespula vidua (Saussure)). They responded similarly, with attrac-tion only to heptyl butyrate. Three yellowjacket species in the Vespula vulgaris species group were also captured in traps (Vespula vulgaris (L.), Vespula flavo-pilosa Jacobson, Vespula maculifrons (Buyyson)). They responded similarly, with attraction primarily to the combination of acetic acid and isobutanol. The bald-faced hornet, Dolichovespula maculata (L.), was attracted to acetic acid and was more strongly attracted to the combination of acetic acid and isobuta-nol. The aerial yellowjacket, Dolichovespula arenaria (Fabr.), was attracted to isobutanol, and was more strongly attracted to the combination of acetic acid and isobutanol. These results add to our understanding of how to target various species of social wasps with chemical lures.
____________________Several chemicals have been identified as attractants for workers of
various species of social wasps. Heptyl butyrate and structurally-related compounds are strongly attractive to the yellowjackets Vespula pensylvanica (Saussure) and Vespula atropilosa, (Sladen) in the western U.S. (Davis et al. 1969, MacDonald et al. 1973), and somewhat attractive to Vespula squamosa (Drury) in the southeastern U.S. (Sharp and James 1979, Landolt et al. 2003). In these studies, small numbers of other species of wasps were trapped, but it is not clear if they were attracted to the chemicals tested. The combination of acetic acid with isobutanol is attractive to a number of species of yellowjackets as well as the bald-faced and European hornets and several paper wasps in the genus Polistes (Landolt 1998, 1999, Landolt et al. 2000). These chemicals have been evaluated and developed as attractants for trapping of nuisance wasps.
Despite these studies, there are large gaps in our knowledge of wasp responses to these chemicals. Results of studies by Grothaus et al. (1973) and Howell et al. (1974) have led to the conclusion that heptyl butyrate and related compounds are unattractive or are weakly attractive to species of wasps in the eastern U.S. (Akre et al. 1980, Sharp and James 1979). However, controlled, replicated experiments have yet to be done to evaluate attraction responses of some social wasp species to heptyl butyrate, which is in commercial use as a lure for wasps (i.e., Sterling International Inc., Spokane, WA). Similarly, tests with acetic acid and isobutanol have not been made in areas possessing popula-tions of some wasp species. A comparison was made of wasp responses to these two attractants in the state of Washington for Vespula germanica (Fabr.), V. pensylvanica, and Polistes aurifer (Fabr.) (Landolt 1998, 1999), which helped assess the relative attractiveness of the lures to these wasps.
1Dept. Biological Science, Oral Roberts University, 7777 S. Lewis Drive, Tulsa, OK 741712 USDA, ARS, Yakima Agricultural Research Laboratory, 5230 Konnowac Pass Road, Wapato, WA 98951

72 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
In an effort to add to this knowledge base, we conducted a comparison of chemical attractants for wasps in northern Michigan, in a forested area possess-ing a number of species not previously targeted for studies with these chemical attractants. The objectives of this study were to (1) determine if the species of wasps known to be present are attracted to heptyl butyrate or are attracted to the combination of acetic acid and isobutanol, and to (2) gain some comparative insights into relative strengths of the attractants, where they are attractive to the same species of wasp.
MATERIALS AND METHODSFive different chemical treatments were compared as attractants for trap-
ping species of social wasps. These chemical treatments were 1) acetic acid, 2) isobutanol, 3) acetic acid and isobutanol, 4) heptyl butyrate, and 5) acetic acid and heptyl butyrate. All chemicals were dispensed from 8 ml polypropylene vials with 3 mm holes in the lid, using the system described by Landolt and Alfaro (2001) for trapping noctuid moths with acetic acid and 3-methyl-1-butanol. Five ml of a chemical were pipetted onto 2 cotton balls in the bottom of each vial. Traps were the Agrisense Trappit® (or Dome trap), which is a wet trap design that is opaque yellow below and clear above, with the trap entrance in the invaginated trap bottom. Traps contained 200 ml of a dilute soap solu-tion (0.01% Dawn dishwashing liquid soap) to drown trapped wasps. The vial chemical dispensers were wired in place inside the top of the trap. Where two chemicals were tested together as one treatment, two vials were placed side by side within the trap.
Traps were hung in shrubbery and on tree branches in a forested area at the Au Sable Institute of Environmental Studies, 14 km southeast of Mance-lona, Kalkaska County, Michigan. A randomized complete block design was used, with 5 replicate blocks of 6 treatments: the 5 chemical treatments plus an unbaited trap. Traps were placed about 10 m apart within blocks. The thirty traps were set out on 20 June 2001 and were checked weekly until 15 August 2001. During weekly checks, the drowning solution was changed, all captured insects were removed, and trap positions within blocks were randomized to provide additional replicates. A total of 40 replicates were obtained over the 8 weeks period. Lures were changed after 4 weeks.
For each species, weekly trap catch data were subjected to an analysis of variance (ANOVA), with means separated by Tukey’s test following a significant F value in the ANOVA (DataMost 1995). For each species, block replicates that provided zero trap catches for all treatments for a week were not included in the analyses. Thus, for each species, the total number of replicates varied.
RESULTSNine different species of social wasps were captured in this study, in three
different genera. Trap capture data for these 9 species is summarized in Table 1. Species of social wasps trapped were identified using Akre et al. (1980) and Jacobson et al. (1978). For all 9 species, numbers of worker wasps captured were sufficient to permit statistical analysis of the data. Wasps in the genera Vespula and Dolichovespula captured were generally workers, and P. fuscatus captured were females.
Numbers of P. fuscatus paper wasps captured in traps baited with the combination of acetic acid and isobutanol were significantly greater than in the unbaited traps. All other chemical lure treatments did not increase captures of that wasp in baited traps compared to control traps.
Numbers of Dolichovespula maculata (L.) captured were significant (greater than controls) for traps baited with acetic acid, acetic acid plus heptyl butyrate, and acetic acid plus isobutanol. The combination of isobutanol and

2002 THE GREAT LAKES ENTOMOLOGIST 73Ta
ble
1. T
otal
num
bers
of w
asps
capt
ured
20
June
- 15
Aug
ust 2
001
in a
ll tr
aps,
and
mea
n +
SE n
umbe
rs o
f was
ps p
er tr
ap p
er w
eek
for
chem
ical
trea
tmen
ts u
sed
as lu
res
in tr
aps.
AA
is a
cetic
aci
d, IB
is is
obut
anol
, AAI
B is
ace
tic a
cid
with
isob
utan
ol, H
B is
hep
tyl b
utyr
ate,
an
d AA
HB
is a
cetic
aci
d w
ith h
epty
l but
yrat
e.
Spec
ies
N
Con
trol
A
A
IB
AA
IB
HB
A
AH
B
Dol
icho
vesp
ula
aren
aria
10
To
tals
0
3 7
11
1 0
Mea
ns 0
.00
+ 0.
00a
0.30
+ 0
.21a
0
.70
+ 0.
30b
1.10
+ 0
.30c
0.
10 +
0.1
0a
0.00
+ 0
.00a
Dol
icho
vesp
ula
mac
ulat
a 35
To
tals
2
161
4 34
0 2
88
M
ean
0.05
+ 0
.05a
4.
60 +
2.1
8b
0.10
+ 0
.07a
9.
70 +
2.6
4c
0.05
+ 0
.05a
2.
50 +
1.0
6bPo
liste
s fu
scat
us
3 To
tal
0 1
0 4
0 0
Mea
n 0.
00 +
0.0
0a
0.33
+ 0
.33a
0.
00 +
0.0
0a
1.33
+ 0
.33b
0.
00 +
0.0
0a
0.00
+ 0
.00a
Vesp
ula
acad
ica
17
Tota
l 0
1 0
0 52
33
Mea
n 0.
00 +
0.0
0a
0.06
+ 0
.06a
0.
00 +
0.0
0a
0.00
+ 0
.00a
3.
06 +
076
c 1.
94 +
0.6
2b
Vesp
ula
cons
obri
na
23
Tota
l 0
0 0
0 47
49
Mea
n 0.
00 +
0.0
0a
0.00
+ 0
.00a
0.
00 +
0.0
0a
0.00
+ 0
.00a
2.
04 +
0.4
9b
2.13
+ 0
.72b
Vesp
ula
vidu
a 27
To
tal
0 1
0 0
97
70
M
ean
0.00
+ 0
.00a
0.
04 +
0.0
4a
0.00
+ 0
.00a
0.
00 +
0.0
0a
3.59
+ 0
.94c
2.
59 +
0.6
7bVe
spul
a fla
vopi
losa
21
To
tal
4 23
9
180
8 15
Mea
n 0.
19 +
0.1
1a
1.10
+ 0
.47a
0.
43 +
0.1
6a
8.57
+ 2
.99b
0.
29 +
0.1
1a
0.57
+ 0
.18a
Vesp
ula
mac
ulifr
ons
20
Tota
l 8
4 21
78
4
8
M
ean
0.40
+ 0
.17a
0.
20 +
0.1
2a
1.05
+ 0
.31b
3.
90 +
0.8
0c
0.20
+ 0
.12a
0.
40 +
0.1
8aVe
spul
a vu
lgar
is
25
Tota
l 3
1 5
74
1 0
Mea
n 0.
12 +
0.0
6a
0.04
+ 0
.04a
0.
20 +
0.0
8a
2.96
+ 0
.69b
0.
04 +
0.0
4a
0.00
+ 0
.00a
* M
eans
in a
row
follo
wed
by
the
sam
e le
tter
are
not
sig
nific
antly
diff
eren
t at p
> 0
.05
by T
ukey
’s te
st.
** N
= n
umbe
r of r
eplic
ates
. Fi
ve re
plic
ates
wer
e se
t out
eac
h w
eek
for 8
wee
ks.
For e
ach
spec
ies,
repl
icat
es in
whi
ch a
ll tr
eatm
ents
pro
-vi
ded
zero
trap
catc
hes
wer
e ex
clud
ed.

74 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
acetic acid was superior to acetic acid or isobutanol alone. Numbers of D. maculata wasps captured with the combination of heptyl butyrate and acetic acid were not significantly greater than with acetic acid alone.
Although only 22 Dolichovespula arenaria (Fabr.) were captured, numbers were significant for traps baited with isobutanol and with the combination of acetic acid and isobutanol. Additionally, the combination of acetic acid and isobu-tanol attracted more D. arenaria than did either isobutanol or acetic acid alone.
The three species in the Vespula rufa species group (Bequaert 1931), Vespula acadica (Sladen), Vespula consobrina (Saussure), and Vespula vidua (Saussure), all responded significantly to heptly butyrate and the combination of acetic acid with heptyl butyrate and not to other lure treatments (Table 1). Numbers of V. acadica were significantly greater in traps baited with heptyl butyrate in comparison to the combination of acetic acid and heptyl butyrate.
The three species in the Vespula vulgaris species group (Bequaert 1931, Jacobson et al. 1978), Vespula flavopilosa Jacobson, Vespula maculifrons (Buys-son), and Vespula vulgaris (L.), were all significantly attracted to the combina-tion of isobutanol and acetic acid, and were not trapped with heptyl butyrate. Numbers of V. maculifrons captured in traps baited with isobutanol alone were also significantly greater than numbers captured in unbaited traps.
DISCUSSIONThe results of this study clearly demonstrate for the first time the attrac-
tion of several species of social wasps to heptyl butyrate and of several other species of social wasps to the combination of acetic acid and isobutanol. Heptyl butyrate is a strong attractant for the yellowjackets V. pensylvanica and V. atropilosa ((Davis et al. 1969, 1973, MacDonald et al. 1973) and had been con-sidered unattractive or a weak attractant for V. squamosa (Grothaus et al. 1973, Howell et al. 1974). We here demonstrated attraction of V. acadica, V. consobrina and V. vidua to the same compound. These three species in the Vespula rufa species group have been captured before in traps baited with heptyl butyrate (Reierson and Wagner 1978), but these were not controlled experiments that might demonstrate attractiveness to the chemical.
The combination of isobutanol and acetic acid was known previous to this study to be attractive to V. pensylvanica and V. germanica (Landolt 1998), Polistes aurifer Saussure (Landolt 1999), and V. maculifrons, V. squamosa, Vespa crabro (L.), D. maculata, Polistes fuscatus (F.), Polistes metricus (Say), and Polistes dominulus Fabr.(Landolt et al. 2000). The results summarized in this paper support previous evidence of D. maculata, P. fuscatus, and V. maculifrons attraction to this lure (Landolt et al. 2000) and also demonstrate attraction of D. arenaria, V. flavopilosa, and V. vulgaris wasps to acetic acid with isobutanol.
For most of the 9 species of wasps that were trapped, acetic acid alone was not attractive, although in several species acetic acid enhanced responses to isobutanol. The exception was D. maculata, which was captured in traps baited with acetic acid alone. This finding is consistent with the earlier results of trapping experiments in Maryland and Oklahoma (Landolt et al. 2000), demonstrating attraction of D. maculata to acetic acid alone. Vespula pensyl-vanica, V. squamosa, V. maculifrons, P. fuscatus, Polistes perplexus Cresson, and Polistes annularis (L.) were weakly attracted to acetic acid in another study (Landolt et al. 2000).
For D. maculata, D. arenaria, P. fuscatus, V. flavopilosa, V. maculifrons, and V. vulgaris, numbers of wasps trapped with the combination of acetic acid and isobutanol were greater than the sums of the numbers trapped with either compound tested separately. That is, the trap catch data indicate that the two chemicals were enhancing or synergistic in attracting these wasps. This pat-tern is consistent with that seen in previous studies demonstrating attraction

2002 THE GREAT LAKES ENTOMOLOGIST 75
of social wasps to these two chemicals, including D. maculata, P. fuscatus, and V. maculifrons (Landolt et al. 2000).
For those species of wasps attracted to heptyl butyrate, there was no indication of any enhancement or synergism with the combination of heptyl butyrate and acetic acid. For V. acadica, V. consobrina, and V. vidua, responses to the combination of acetic acid and heptyl butyrate appear to be due simply to a response to heptyl butyrate. In the case of D. maculata, the response to the combination of acetic acid and heptyl butyrate appears to be due solely to the response to acetic acid.
With this added information on wasp responses to these lures, taxonomic patterns are more evident. For example, the three species of yellowjackets that are in the Vespula rufa species group, V. acadica, V. consobrina, and V. vidua, responded in a similar fashion to the lures, with significant responses to heptyl butyrate and not to isobutanol and acetic acid. This response was the same as that observed for the related species V. atropilosa (Landolt 1998). For the three species of yellowjackets in the Vespula vulgaris species group, V. flavopilosa, V. maculifrons, and V. vulgaris, responses were also similar, with significant attraction to the combination of acetic acid and isobutanol, and with no response to heptyl butyrate or acetic acid. This attraction pattern is similar to that observed previously for another species in the V. vulgaris spe-cies complex, V. germanica, and confirms previous findings for V. maculifrons. Results with V. pensylvanica however (Landolt 1998) are contrary to this pat-tern. Vespula pensylvanica is in the V. vulgaris species group (Bequaert 1931) but responds well to both types of lures: heptyl butyrate and the combination of acetic acid and isobutanol. Several species of Polistes in the Fuscopolistes subgenus (Richards 1978) are attracted to the combination of acetic acid and isobutanol, including P. fuscatus, P. aurifer, P. metricus (Landolt 1999, Landolt et al. 2000). Species of Polistes do not appear to respond to heptyl butyrate, but may respond weakly to acetic acid (Landolt et al. 2000). As in other studies, the strengths of responses to these chemicals by different wasp species are not determined. For example, it is not known if low, but significant, numbers of wasps in traps are due to a weak response or are due to low populations or low activity at the trapping site. In this study, that is a question particularly for D. arenaria. Although numbers in traps baited with isobutanol and acetic acid with isobutanol were statistically significant, they were a small fraction of the numbers captured of other species. This wasp was not captured in significant numbers in trapping studies conducted in Maryland (Landolt 2000), where it is generally present (Akre et al. 1980).
As a practical matter, it is important to know which species of wasps might be trapped with different chemicals. For example, yellowjackets in the V. vulgaris species group are considerably more pestiferous than yellowjackets in the V. rufa species group, due to differences in nesting phenology, colony size, and foraging behavior. It would not be advantageous to trap species of wasps in the V. rufa group with heptyl butyrate in an area where members of the V. vulgaris group are problematic but are not attracted to heptyl butyrate. However, the results here and elsewhere (Davis et al. 1969; Landolt 1998; Landolt 1999; Landolt et al. 2000, 2003; Day and Jeanne 2001) indicate that most, if not all, North American vespine wasps are attracted either to heptyl butyrate, to the combination of acetic acid and isobutanol, or to both lures.
It is interesting that there were no wasp species in this study that re-sponded to both lures: either they were attracted to heptyl butyrate or they were attracted to acetic acid with isobutanol. This underscores the uniqueness of V. pensylvanica which can be trapped with heptyl butyrate (Davis et al.1969) or acetic acid with isobutanol (Landolt 1998). It is assumed that the attraction responses to these chemicals, evidenced by captures of wasps in traps, are based on differences in wasp food foraging behavior, but there is little information available on the presence of these compounds in materials consumed by wasps.

76 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Acetic acid is a product of microbial fermentation of sugars, while isobutanol is produced in small amounts by the bacteria Citrobacter freundii (DeMilo et al. 1996) and was also found in fermented sugar solutions by Utrio and Eriks-son (1977). Esters similar to heptyl butyrate are often found in fruit odors (i.e. Mattheis et al. 1991 for apple volatiles). While it seems logical to assume that wasp response to these chemicals is a strategy to locate carbohydrates, specifi-cally sugars, it is not yet known what natural sugar sources might be located by orientation to heptyl butyrate or to acetic acid with isobutanol.
ACKNOWLEDGMENTSWe thank J. Alfaro and C. Smithhisler for making the lures for these stud-
ies, and C. Haley, D. Lefever, and R. Walter-Toews, for assistance in maintain-ing trapping experiments. The mention of a commercial or proprietary product does not constitute an endorsement for its use by USDA.
LITERATURE CITEDAkre, R. D., A. Greene, J. F. MacDonald, P. J. Landolt, and H. G. Davis. 1980. Yellow-
jackets of America North of Mexico. U.S. Department of Agriculture, Agriculture Handbook No. 552, 102 pp.
Bequaert, J. 1931. A tentative synopsis of the hornets and yellowjackets of America. Entomol. Americana 12: 71-138.
DataMost. 1995. StatMost statistical analysis and graphics. DataMost, Salt Lake City, UT.Davis, H. G., G. W. Eddy, T. P. McGovern, and M. Beroza. 1969. Heptyl butyrate, a new
synthetic attractant for yellow jackets. J. Econ. Entomol. 62:1245. Davis, H. G., R. W. Zwick, W. M. Rogoff, T. P. McGovern and M. Beroza. 1973. Perimeter
traps baited with synthetic lures for suppression of yellowjackets in fruit orchards. Environ. Entomol. 2: 569-571.
Day, S. E. and R. L. Jeanne. 2001. Food volatiles as attractants for yellowjackets (Hy-menoptera: Vespidae). Environ. Entomol. 30: 157-165.
DeMilo, A. B., C.-J. Lee, D. S. Moreno, and A. J. Martinez. 1996. Identification of volatiles derived from Citrobacter freundii fermentation of a trypticase soy broth. J. Agric. Food Chem. 44: 607-612.
Grothaus, R. H., H. G. Davis, W. M. Rogoff, J. A. Fluno, and J. M. Hirst. 1973. Baits and attractants for east coast yellowjackets, Vespula spp. Environ. Entomol. 2: 717-718.
Howell, J. O., T. P. McGovern, and M. Beroza. 1974. Attractiveness of synthetic compounds to some eastern Vespula species. J. Econ. Entomol. 67:629-630.
Jacobson, R. S., R. W. Matthews, and J. F. MacDonald. 1978. A systematic study of the Vespula vulgaris group with a description of a new yellowjacket species in eastern North America (Hymenoptera: Vespidae). Ann. Entomol. Soc. Amer. 71: 299-312.
Landolt, P. J. 1998. Chemical attractants for trapping yellowjackets Vespula germanica and Vespula pensylvanica (Hymenoptera: Vespidae). Environ. Entomol. 27: 1229-1234.
Landolt, P. J. 1999. A chemical attractant for the golden paper wasp, Polistes aurifer Saussure (Hymenoptera: Vespidae). J. Kansas Entomol. Soc. 71:69-73.
Landolt, P. J. and J. F. Alfaro. 2001. Trapping Lacanobia subjuncta, Xestia c-nigrum, and Mamestra configurata (Lepidoptera: Noctuidae) with acetic acid and 3-methyl-1-butanol in controlled release dispensers. Environ. Entomol. 30: 656-662.
Landolt, P. J., H. C. Reed, J. R. Aldrich, A. L. Antonelli, and C. Dickey. 2000. Social wasps (Hymenoptera: Vespidae) trapped with acetic acid and isobutanol. Florida Entomol. 82: 609-614.

2002 THE GREAT LAKES ENTOMOLOGIST 77
Landolt, P. J., H. C. Reed, and D. J. Ellis. 2003. Trapping yellowjackets (Hymenoptera: Vespidae) with heptyl butyrate emitted from controlled release dispensers. Florida Entomol. (In press).
MacDonald, J. F., R. D. Akre, and W. B. Hill. 1973. Attraction of yellow jackets (Vespula spp) to heptyl butyrate in Washington State (Hymenoptera: Vespidae). Environ. Entomol. 2: 375-379.
Mattheis, J. P., J. K. Fellman, P. M. Chen, and M. E. Patterson. 1991. Changes in head-space volatiles during physiological development of Bisbee Delicious apple fruit. J. Agric. Food Chem. 39: 1902-1906.
Reierson, D. A. and R. E. Wagner. 1978. Trapping to determine the sympatry and seasonal abundance of various yellowjackets. Environ. Entomol. 7: 418-422.
Richards, O. W. 1978. The Social Wasps of the Americas Excluding the Vespinae. British Museum of Natural History. London. 580 pp.
Sharp, J. L. and J. James. 1979. Color preference of Vespula squamosa. Environ. En-tomol. 8: 708-710.
Utrio, P. and K. Eriksson. 1977. Volatile fermentation products as attractants for Mac-rolepidoptera. Ann. Zool. Fennici 14: 98-104.

78 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1

2002 THE GREAT LAKES ENTOMOLOGIST 79
GROUND BEETLES (COLEOPTERA: CARABIDAE) INHABITING STANDS OF REED CANARY GRASS PHALARIS ARUNDINACEA ON
ISLANDS IN THE LOWER CHIPPEWA RIVER, WISCONSIN
Paula K. Kleintjes1, Anita M. Christensen1, William J. Barnes1 and Lori A. Lyons1
ABSTRACTWe used pitfall traps to assess ground beetle diversity (Coleoptera:Carabidae)
on two islands in the lower Chippewa River, Eau Claire County, Wisconsin, with rapidly expanding populations of reed canary grass, Phalaris arundinaceae. We collected 233 individuals belonging to 17 species over four, 3-9 day sampling periods, May-August 1994. All species have been documented in Wisconsin and most are considered habitat generalists. Agonum fidele, A. extensicolle, Aniso-dactylus harrisii and Bembidion quadrimaculatum oppositum comprised 70% of all species collected. Seven species were common to both islands, with 13 species collected on Canarygrass Island and 11 species on Ski Jump Island. Carabid species diversity (Shannon’s H=2.01) was greatest on Canarygrass Island.
____________________Reed canary grass Phalaris arundinacea Linnaeus is an invasive species
in lowland and riparian areas of the Midwest and other regions of the northern hemisphere (Apfelbaum and Sams 1987, Barnes 1999). Both native populations and European ecotypes are thought to exist in Wisconsin, the latter being more aggressive and responsible for displacement of native populations and other wetland plant species (Fassett 1951, Apfelbaum and Sams 1987, Borman et al. 1997, Barnes 1999). In the lower Chippewa River basin of Wisconsin, P. arundinacea has become increasingly conspicuous on islands and floodplains, and its impact on native herbaceous species composition and diversity has been documented (Barnes 1999). Barnes (1999) attributed its expansion, in part, to the establishment of the European ecotype and its adaptability to grow under fluctuating water levels. Based on a literature review we found no citations of epigeal insects inhabiting islands in the Chippewa River, Wisconsin or of epigeal insects associated with P. arundinacea dominated habitats. Our objective was to document the ground beetles that inhabited two P. arundinacea populated islands in the lower Chippewa River, Wisconsin.
METHODS AND MATERIALSWe selected two islands, Ski Jump (SJ) and Canarygrass (CI), both ap-
proximately 5 km downstream of the City of Eau Claire, Wisconsin. Ski Jump Island is approximately 400 m long and 120 m at its widest point and has a maximum elevation of 2 m. The island is thought to have formed between 1934 and 1938 (Barnes 1985). The upstream end of the island is dominated by P. arundinacea. Canarygrass Island is located 1000 m upstream of Ski Jump Island. It is approximately 200 m long by 70 m wide and nearly level with a maximum elevation of 1 m. The island formed at the same time as Ski Jump Island, but it has disappeared and reformed several times, persisting since 1972 (Barnes, personal observation). The entire island is dominated by P. arundinacea. The soil of both islands is riverwash consisting of sand and cobble stone surfaces.
1Department of Biology, University of Wisconsin-Eau Claire, Eau Claire, WI 54702

80 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
We used custom-designed arthropod pitfall traps to sample carabid spe-cies. Each trap consisted of a 295 ml plastic beverage cup placed in the ground with the top rim flush with the ground. A piece of 10 x 10 cm brown linoleum was held in place 2 cm above each cup by two, 7cm long nails to keep out rain and leaves. A 1:1 ratio of 70% ethyl alcohol and water was placed 2 cm deep in the bottom of each cup to kill insects and minimize evaporation. Twenty-five traps were placed within a monotypic stand of P. arundinacea occupying the upstream end of each island. Traps were placed 3 m apart along five transects, each spaced 5 m apart and running the width of each island. Traps were checked, emptied and replaced with new traps at the end of each of four sampling periods during the summer of 1994. Dates of sampling occurred after spring flooding: 20-23 May, 9-16 June and 1-8 July for both islands, 20-26 July for Ski Jump Island, and 25 July-2 August for Canarygrass Island. After trap collection, all arthropods were sorted and identified to morphospecies. All carabid beetles were removed and stored separately by island in vials of 70% ethyl alcohol. Beetles were subsequently pinned or point mounted and species identified by Kirk Larsen, Luther College, Decorah, Iowa. Taxonomy followed Bousquet and Larochelle (1993). Voucher specimens are housed in collections at the University of Wisconsin-Eau Claire and Luther College. Data were summed for each collec-tion period per island and pooled across all samples to obtain the mean number of each species captured/trap. A Shannon’s H diversity index (H=- ∑ piln(pi)) was calculated for each island (Hayek and Buzas 1997).
We sampled vegetation in each pitfall trap area by determining the pres-ence of all herbaceous plant species observed within a 1 m diameter circular plot surrounding each trap. Data were collected during the last pitfall collection period when grasses were in bloom and easily identifiable to species. Frequency of occurrence of each plant species was determined by dividing the total number of plots in which a species was present by the total number of plots sampled (25).
RESULTS AND DISCUSSIONWe collected 233 Carabidae beetles belong to 17 species (Table 1). At the
time of the study, all species had been previously documented in Wisconsin with the exception of Agonum fidele Casey (Bousquet and Larochelle 1993). However, A. fidele has since been documented from southern Wisconsin (Sauk Co.) in 1998 (Purrington et al. 2000). Seven species (41%) were common to both islands, with 13 species (76%) collected on Canarygrass Island and 11 species (65%) on Ski Jump Island. Canarygrass Island had greater beetle abundance and species richness. It also had a higher Shannon’s H Diversity Index (2.01 for CI vs. 1.88 for SJ) out of a maximum possible of 2.83 based on H=ln(17) if all relative abundances of the 17 species were equal (Hayek and Buzas 1997).
Frequency of P. arundinacea occurrence differed little between the two trap areas. All 25 trap quadrats (100%) on Canarygrass Island contained P. arundinacea whereas 88% of the quadrats contained it on Ski Jump Island. Greater than 36% of the Canarygrass Island plots were occupied by three plant species, the only three species observed in the samples (Table 2). In contrast, 13 plant species were observed on Ski Jump Island plots, although most occurred at low (<5%) frequencies (Table 2). We do not know why the P. arundinacea stand on Ski Jump Island had fewer numbers of beetles (51 vs. 182) but we speculate that it was because the island had a greater variety of adjacent habitats to which beetles could disperse rather than being limited to the P. arundinacea monoculture occupying Canarygrass Island.
Carabid species richness in our study was less or nearly equal to that found for specific vegetation cover types, e.g. agricultural crops, old fields, and prairie remnants in the upper Midwest (~10-60 spp.) (Esau and Peters 1975, House and All 1981, Dritschillo and Irwin 1982, Purrington et al. 1989, Ep-stein and Kulman 1990, Weiss et al. 1990, Will et al. 1995, Clark et al. 1997,
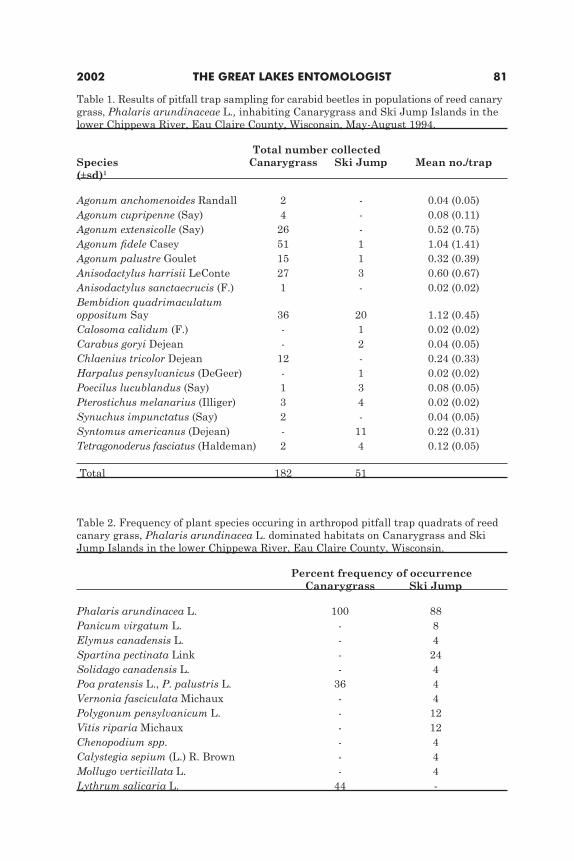
2002 THE GREAT LAKES ENTOMOLOGIST 81
Table 1. Results of pitfall trap sampling for carabid beetles in populations of reed canary grass, Phalaris arundinaceae L., inhabiting Canarygrass and Ski Jump Islands in the lower Chippewa River, Eau Claire County, Wisconsin, May-August 1994.
Total number collectedSpecies Canarygrass Ski Jump Mean no./trap (±sd)1
Agonum anchomenoides Randall 2 - 0.04 (0.05)Agonum cupripenne (Say) 4 - 0.08 (0.11)Agonum extensicolle (Say) 26 - 0.52 (0.75)Agonum fidele Casey 51 1 1.04 (1.41)Agonum palustre Goulet 15 1 0.32 (0.39)Anisodactylus harrisii LeConte 27 3 0.60 (0.67)Anisodactylus sanctaecrucis (F.) 1 - 0.02 (0.02)Bembidion quadrimaculatum oppositum Say 36 20 1.12 (0.45)Calosoma calidum (F.) - 1 0.02 (0.02)Carabus goryi Dejean - 2 0.04 (0.05)Chlaenius tricolor Dejean 12 - 0.24 (0.33)Harpalus pensylvanicus (DeGeer) - 1 0.02 (0.02)Poecilus lucublandus (Say) 1 3 0.08 (0.05)Pterostichus melanarius (Illiger) 3 4 0.02 (0.02)Synuchus impunctatus (Say) 2 - 0.04 (0.05)Syntomus americanus (Dejean) - 11 0.22 (0.31)Tetragonoderus fasciatus (Haldeman) 2 4 0.12 (0.05)
Total 182 51
Table 2. Frequency of plant species occuring in arthropod pitfall trap quadrats of reed canary grass, Phalaris arundinacea L. dominated habitats on Canarygrass and Ski Jump Islands in the lower Chippewa River, Eau Claire County, Wisconsin.
Percent frequency of occurrence Canarygrass Ski Jump
Phalaris arundinacea L. 100 88Panicum virgatum L. - 8Elymus canadensis L. - 4Spartina pectinata Link - 24Solidago canadensis L. - 4Poa pratensis L., P. palustris L. 36 4Vernonia fasciculata Michaux - 4Polygonum pensylvanicum L. - 12Vitis riparia Michaux - 12Chenopodium spp. - 4Calystegia sepium (L.) R. Brown - 4Mollugo verticillata L. - 4Lythrum salicaria L. 44 -

82 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Ellsbury et al. 1998). Epstein and Kulman (1990) found approximately 20 species in mesic-hydric lowland forest sites and about 10 species in mesic field sites of east-central Minnesota. About 70% our species were also collected in at least one of the studies cited above – Agonum cupripenne (Say), A. extensicolle (Say), A. palustre Goulet, Anisodactylus sanctaecrucis (Fabricius), A. harrisii LeConte, Bembidion quadrimaculatum oppositum Say, Chlaenius tricolor De-jean, Harpalus pennsylvanicus (DeGeer), Poecilis lucublandus (Say), Pterosti-chus melanarius (Illiger), Syntomus americanus (Dejean) and Tetragonoderus fasciatus (Haldeman).
Most of the species we collected are considered habitat generalists com-monly found in gardens, fields, agricultural crops and/or forests (Epstein and Kulman 1990, Clark et al. 1997). Bembidion quadrimaculatum oppositum (Say) was the most commonly collected species on both islands. It is an abundant generalist with high numbers recorded from surveys of agricultural crops and old fields (Esau and Peters 1975). Agonum palustre ranked fifth in abundance on Canarygrass Island and is considered a hydric lowland forest specialist (Ep-stein and Kulman 1990). Thus, our results support its tolerance of wet areas and we suspect it can tolerate periodic flooding. We collected one individual of Synuchus impunctatus (Say) also found by Purrington et al. (1989) in upland forest habitat and 11 individuals of S. americanus, also found in oak (Quercus spp.) forest by Epstein and Kulman (1990). Syntomus americanus was the second most abundant carabid collected on Ski Jump Island which does have a woody vegetation component at higher elevations. Upland oak forest also occurs on neighboring mainland hillsides.
We recognize that additional sampling of P. arundinacea dominated habi-tats and adjacent non-dominated grassland habitats on more islands and along the mainland is needed to determine whether carabid species abundance and richness are affected by the expansion of P. arundinacea. In addition, assess-ment of other insects such as those that are monophagous herbivores or low in vagility, e.g., some lepidoptera larvae, may be better for determining the effects of P. arundinacea upon insect divesity because the insects may be reliant on native plant species that could be outcompeted by P. arundinacea.
ACKNOWLEDGMENTSWe thank Helen Barnes, Pam Berquist, Josh Conner, Chad Hess, Rebecca
van Nelson-Cullen and UWEC Upward Bound Students for field assistance and Kirk Larsen for Carabidae species identification. The research was supported by a UWEC Office of Research and University Sponsored Projects Undergraduate Faculty-Student Collaboration Research Grant.
LITERATURE CITEDApfelbaum, S. I. and C. E. Sams. 1987. Ecology and control of reed canary grass. Nat.
Areas. J. 7:69-74.Barnes, W. J. 1985. Population dynamics of woody plants on a river island. Can. J. Bot.
63:647-655.Barnes, W. J. 1999. The rapid growth of a population of reed canary grass (Phalaris arun-
dinacea L.) and its impact on some riverbottom herbs. J. Torrey Bot. Soc. 126:133-138.Borman, S., R. Korth and J. Temte. 1997.Through the looking glass: A field guide to
Aquatic plants. Wisconsin Department of Natural Resources. Publ. # FH-207-97, Merrill, WI.
Bousquet, Y. and A. Larochelle. 1993. Catalogue of the Geadephaga (Coleoptera: Trachy-pachidae, Rhysodidae, Carabidae including Cicindelini) of America north of Mexico. No. 167. Entomological Society of Canada Memoir, Ottawa, Canada.

2002 THE GREAT LAKES ENTOMOLOGIST 83
Clark, M. S., S. H. Gage, and J. R. Spence. 1997. Habitats and management associated with common ground beetles (Coleoptera: Carabidae) in a Michigan agricultural landscape. Environ. Entomol. 26:519-527.
Dritschillo, W. and T. L. Irwin. 1982. Responses in abundance and diversity of cornfield carabid communities to differences in farm practices. Ecology 63: 900-904.
Ellsbury, M. M., J. E. Powell, F. Forcella, D. W. Woodson, S. A. Clay, W. E. Riedell. 1998. Diversity and dominant species of ground beetle assemblages (Coleoptera: Carabidae) in crop rotation and chemical input systems for the northern Great Plains. Ann. Entomol. Soc. Amer. 91:619-625.
Epstein, M. E. and H. M. Kulman. 1990. Habitat distribution and seasonal occurrence of carabid beetles in east-central Minnesota. Am. Midl. Nat.123: 209-225.
Esau, K. L. and D. C. Peters. 1975. Carabidae collected in pitfall traps in Iowa cornfields, fencerows and prairies. Environ. Entomol. 4:509-513.
Fassett, N. C. 1951. Grasses of Wisconsin. University of Wisconsin Press, Madison, WI. 173pp.
Hayek, L.C. and M. A. Buzas. 1997. Surveying Natural Populations. Columbia University Press. New York, 563p.
House, C. J. and J. N. All. 1981. Carabid beetles in soybean agroecosystems. Environ. Entomol. 10:194-196.
Purrington, F. F., J. E. Bater, M. G. Paoletti and B. J. Stinner. 1989. Ground beetles from a remnant oak-maple-beech forest and its surroundings in northeastern Ohio (Coleoptera: Carabidae). Great Lakes Entomol. 22:105-110.
Purrington, F. F., D. K. Young, K. J. Larsen, and J. C-T. Lee. 2000. New distribution records of ground beetles from the North Central United States. (Coleoptera: Cara-bidae). Great Lakes Entomol. 33:199-204.
Weiss, M. J., E. U. Balsbaugh Jr., E. W. French and B. K. Hoag. 1990. Influence of tillage management and cropping system on ground beetle (Coleoptera: Carabidae) fauna in the northern great plains. Environ. Entomol. 19: 1388-1391.
Will, K. W., F. F. Purrington and D. J. Horn. 1995. Ground beetles of islands in the west-ern basin of Lake Erie and the adjacent mainland (Coleoptera: Carabidae including Cicindelini). Great Lakes Entomol. 28:55-70.

84 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1

2002 THE GREAT LAKES ENTOMOLOGIST 85
DEVELOPMENT AND SEX RATIO OF MELITTOBIA AUSTRALICA AND M. DIGITATA (HYMENOPTERA: EULOPHIDAE) ON
MEGACHILE ROTUNDATA (HYMENOPTERA: MEGACHILIDAE) AND TRYPOXYLON POLITUM (HYMENOPTERA: SPHECIDAE)
Jorge M. González1 and Robert W. Matthews1
ABSTRACTLife history, development time, and sex ratio of Melittobia australica
Girault and M. digitata Dahms on Megachile rotundata (Fabricius) and Try-poxylon politum Say are presented and compared with previous reports. Total development time and sex ratio differs slightly amongst the different hosts but falls within the expected range reported for different hosts and Melittobia species. The slightly longer development times and higher proportion of males recorded for both species on M. rotundata hosts probably reflects poorer host nutritional quality, relative to T. politum. We confirm that the one unusually extended development time reported by Schmieder for Melittobia chalybii Ash-mead collected in New Jersey, can be accounted for as likely having represented a previously unknown species since described as M. femorata Dahms.
____________________Melittobia wasps (Hymenoptera: Eulophidae) are arrhenotokous parasitoid
wasps normally associated with mud dauber wasps (Hymenoptera: Sphecidae) of the genera Trypoxylon and Sceliphron. However, many bees, wasps, and even insects of different orders have been reported to be attacked by these parasitoids (Dahms 1984b, Girault 1912, González and Terán 1996, Howard 1892, Maeta and Yamane 1974, Matthews et al. 1996).
Even though Melittobia spp. can be relatively easily cultured in the labo-ratory on mud dauber wasp prepupae (Trypoxylon spp. and Sceliphron spp.), it is relatively difficult to obtain numerous hosts. Some researchers have used honeybee prepupae as hosts with complete success (Cônsoli and Vinson 2002a, 2002b, Varanda et al. 1984). Others have used puparia of different flies as hosts (Balfour-Browne 1922, Matthews et al. 1996). Some Melittobia spp. can be reared on puparia of flesh flies, Neobellieria bullata (Parker), (Diptera, Sar-cophagidae), which is how the WOWbug (Melittobia digitata) is commercially reared (Matthews et al. 1996, Cônsoli and Vinson 2002a, 2002b).
Even though some Melittobia species have been reported as pests of Mega-chile spp. and especially of the leaf–cutter bee, Megachile rotundata (Fabricius), by various authors in different regions (Edwards and Pengelly 1966, Farkas and Szalay 1985, Peck 1969, Tasei 1978, Woodward 1994) no information ex-ists concerning development time and sex ratio of Melittobia spp. on this host. Furthermore, while the pipe-organ mud dauber wasp, Trypoxylon politum Say, and other species in this genus also have been reported as hosts of a few Melit-tobia species (Buckell 1928, González and Terán 1996, González et al. 1985, Lith 1955, Matthews et al. 1985, Schmieder 1933, 1938) there is only one brief mention in the literature for development time and sex ratio of Melittobia digi-tata Dahms on T. politum (Matthews et al. 1996), and nothing for Melittobia australica Girault. The objective of the present study is to provide information on these aspects of the biology of M. australica and M. digitata reared on M. rotundata and T. politum prepupae under uniform conditions.
1University of Georgia, Department of Entomology, Athens, GA 30602, USA

86 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
MATERIALS AND METHODSMelittobia australica and M. digitata were reared on prepupae of M.
rotundata and T. politum. Trypoxylon politum hosts were collected as diapaus-ing prepupae from the vicinity of Athens, GA and kept refrigerated until use. Megachile rotundata were obtained as diapausing cocoons from Ron Bitner (Pioneer Hybrid International, Namper, ID) and also kept refrigerated until use. For rearing each species of Melittobia a single host (of both host species) was removed from its cocoon and placed in a clear-view polystyrene chamber (15 mm x 20 mm x 45 mm) with 1 , 2 or 3 mated Melittobia females per host. Forty replicates on each host were simultaneously started for each Melittobia species under study. Hosts of both species were taken from the same batches, kept in a refrigerator, and were presumably the same age. Hosts were weighed and only those weighing 0.30 g (T. politum) and 0.03 g (M. rotundata) were used for this study. Cultures were reared at 25° C and 50 % RH in an incubator.
Duration, in days, of development from egg to larvae, from larvae to pupae, and from pupae to imago, was determined through daily observations and reported as ranges. Final progeny counts were used to determine sex ratio.
Data are compared with information from other Melittobia species and hosts that have been previously published.
RESULTS AND DISCUSSIONAs on other prepupae of different wasps and bees, both M. australica and
M. digitata laid clusters of eggs mainly along intersegmental grooves but not restricted to any particular area of the host (Dahms 1984b, González 1994). In both Melittobia species, eggs hatched between 2 and 4 days on both hosts, which is typical for most Melittobia species reared at temperatures between 20 and 25 º C (Table 1). The larvae of M. australica became pupae after 8 – 14 days on M. rotundata, but slightly earlier (7 – 11 days) on Trypoxylon politum. The same trend was found in M. digitata, which became pupae between 9 –12 days on M. rotundata and 7 – 9 days on T. politum. Melittobia australica eclosed as adults after 5 – 8 days as pupae on M. rotundata and 5 – 7 days as pupae on T. politum. A similar overall pattern of slightly faster development on T. politum hosts was exhibited by M. digitata (Table 1).
Most studied Melittobia species have larval stages that last 7 – 15 days before transforming into pupae, except for M. hawaiiensis whose larval devel-opment time varies between 5 – 7 days and M. acasta that can require up to 18 days (Table 1). A curious case is M. chalybii reported by Schmieder (1933) in which the larval stage lasted up to 71 days; we will address this later. The pupal stage lasts between 4 to 8 days in most studied species. However in M. acasta reared at 25 º C some require as few as 3 days and in M. chalybii the pupal stage may last up to 15 days (Table 1).
Cônsoli and Vinson (2002a, 2002b) have suggested that nutritional quality of the host can dramatically affect development time in M. digitata. Specifi-cally, they show that larvae reared primarily on host hemolymph develop faster than those that feed on the remaining host tissues having correspondingly less hemolymph. This finding suggests a basis for the slightly longer development times we found for both Melittobia developing on M. rotundata compared to T. politum. The prepupae of M. rotundata are less than half as large as those of T. politum, and are not nearly as flaccid which probably reflects reduced avail-ability of hemolymph, thereby forcing the developing Melittobia to cope with a less optimal diet. This ultimately results in a greater proportion of the long-wing morph relative to the short-wing (brachypterous) form on M. rotundata and the former require longer to develop (Cônsoli and Vinson 2002).

2002 THE GREAT LAKES ENTOMOLOGIST 87Ta
ble
1: S
umm
ary
of p
ublis
hed
life
hist
ory
deve
lopm
ent t
ime
of d
iffer
ent M
elitt
obia
spe
cies
on
vari
ous
host
s at
diff
eren
t pla
ces
and
tem
pera
-tu
res.
Val
ues
are
aver
ages
or r
ange
s.
Mel
itto
bia
Hos
t Te
mp
Egg
s La
rvae
P
upae
To
tal
Cou
ntry
R
efer
ence
Spec
ies
(°
C)
(Day
s)
(Day
s)
(Day
s)
(Day
s)
aust
ralic
a M
egac
hile
25
2
– 4
8
– 14
5
– 8
15
– 2
6
USA
Th
is s
tudy
ro
tund
ata
aust
ralic
a Tr
ypox
ylon
25
2
– 4
7
– 11
5
– 7
14
– 2
2
USA
Th
is s
tudy
po
litum
digi
tata
M
egac
hile
25
2
– 4
9
– 12
5
– 8
16
– 2
4
USA
Th
is s
tudy
ro
tund
ata
digi
tata
Tr
ypox
ylon
25
2
– 4
7
– 9
5
– 8
14
– 2
1
USA
Th
is s
tudy
po
litum
acas
ta
Scel
iphr
on
20
2 –
3
16 –
18
4
– 6
23
– 2
7
Vene
zuel
a G
onzá
lez,
fistu
lari
um
1994
acas
ta
Scel
iphr
on
25
1 –
3
14 –
16
3
– 5
18
– 2
4
Vene
zuel
a G
onzá
lez,
fis
tula
rium
1
994
acas
ta
Ody
neru
s sp
p.,
18 –
19
3
– 4
11
– 1
5
7 –
8
21 –
27
Fi
nlan
d Li
th, 1
955
Tr
ypox
ylon
spp
. ac
asta
Va
riou
s 26
1
– 2
6
– 13
3
– 10
10
– 2
4
Japa
n M
aeta
, 197
8
Hym
enop
tera
acas
ta
Vari
ous
Dip
tera
?
2 –
7 8
– 9
7 17
– 2
3 En
glan
d Ba
lfour
-
and
Brow
ne, 1
928
H
ymen
opte
raau
stra
lica
Scel
iphr
on
25 –
30
3
– 4
8
– 10
3
– 4
14
– 1
8
Aust
ralia
D
ahm
s,
form
osum
19
84b

88 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1Ta
ble
1: C
ontin
ued
Mel
itto
bia
Hos
t Te
mp
Egg
s La
rvae
P
upae
To
tal
Cou
ntry
R
efer
ence
Spec
ies
(°
C)
(Day
s)
(Day
s)
(Day
s)
(Day
s)
chal
ybii
Tryp
oxyl
on
19 –
25
4
7
– 71
3
– 15
14
– 9
0
USA
Sc
hmie
der,
(2 -
4 sp
p.)*
po
litum
19
33ch
alyb
ii Tr
ypox
ylon
?
3 –
4
11 –
15
7
– 8
21
– 2
7
USA
Bu
ckel
l, (=
digi
tata
) po
litum
19
28di
gita
ta
Tryp
oxyl
on
22 –
30
3 –
4
7 –
10
4 –
7
14 –
21
U
SA
Mat
thew
s et
al.,
po
litum
19
96di
gita
ta
Neo
belli
eria
22
– 3
0
3 –
4
7 –
10
4 –
7
14 –
21
U
SA
Mat
thew
s et
al.,
bu
llata
19
96ha
wai
iens
is
Apis
mel
lifer
a ?
1 –
2
5 –
7
5 –
7
11 –
16
Br
asil
Vara
nda
et a
l.,
19
84 *
Schm
iede
r (19
33) p
rese
nts
draw
ings
of M
. cha
lybi
i, bu
t men
tions
diff
eren
t loc
aliti
es in
clud
ing
the
one
from
whi
ch M
. eva
nsi w
as la
ter
desc
ribe
d. H
e al
so n
otes
the
occu
rren
ce o
f a “d
iapa
usin
g M
elitt
obia
” and
sug
gest
s th
at it
is re
late
d to
“tro
phic
cond
ition
s ob
tain
ed d
urin
g th
e la
rval
gro
wth
”. Pa
rt o
f Sch
mie
der’s
mat
eria
l was
als
o us
ed b
y W
hitin
g (1
947)
, who
men
tions
the
use
of fo
ur M
elitt
obia
spe
cies
and
sho
ws
a ph
otog
raph
of M
. dig
itata
. See
text
for f
urth
er d
etai
ls.

2002 THE GREAT LAKES ENTOMOLOGIST 89
Arrhenotokous insects with extreme inbreeding normally have female-biased ratios (Assem et al. 1982, Dahms 1984b, Hamilton 1967). The sex ratio of different Melittobia species has been reported by many authors as highly skewed toward females, with the percentage of males varying from 1 – 5 % (Table 2) regardless of host used. Unlike some other species of parasitoid wasps (reviewed by Godfray 1994) Melittobia females do not adjust their sex ratio in response to the number of foundress females. Although more individuals were produced in the larger host, sex ratios were basically the same when 1, 2 or 3 female foundresses were used in each Melittobia species on each host. Bra-chypterous females were also produced on both hosts but their number varied (decreasing as the number of foundresses increased). Data on these aspects will be presented elsewhere.
When sex ratio is compared in the two Melittobia species used in this study on the two different hosts the percentage of males was slightly higher when reared on M. rotundata(Table 2) . One possible explanation is that on M. rotundata “late” developing individuals fail to survive due to active or passive interference by their siblings, including facultative siblicidal behavior as shown by Cônsoli and Vinson (2002a). This would disproportionately affect females, since males develop slightly faster, causing the sex ratio to be slightly more male-biased. Adams (2002) has shown that males are laid at a relatively constant daily rate throughout the foundresses’ tenure on the host. It is also possible that the relatively poorer host quality of M. rotundata affected the final sex ratio.
Table 2: Sex ratios of different Melittobia species compared from this study and the literature.
Melittobia Host Male - Female Referencespecies Ratio and range
australica Megachile rotundata 5:95 (4 – 8):(96 – 92) This studyaustralica Trypoxylon politum 2:98 (1 – 3):( 99 – 97) This studydigitata Megachile rotundata 4:96 (3 – 5):(97 – 95) This studydigitata Trypoxylon politum 2:98 (1 –3):(99 – 97) This studyacasta Sceliphron (3 – 5):(97 – 95) González, 1994 fistularium australica Sceliphron (1 – 4):(99 – 96) Dahms, 1984b formosum chalybii Sceliphron sp., 3:97 Schmieder, 1938(= 3 diff. spp.)* Chalybion sp., Trypoxylon spdigitata Trypoxylon politum 5:95 Matthews et al., 1996digitata Trypoxylon politum 2.4:97.6 Adams 2002hawaiiensis Apis mellifera 2:98 Varanda et al., (1 – 3):(99 – 97) 1984* In this paper, Schmieder mentions that he was working with probably three species that when intercrossed did not “…produce any female offspring…” but were “cytologi-cally, like chalybii”. See text for further details.

90 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
Schmieder (1933) reared M. chalybii between 19 – 25 º C and recorded a life cycle of up to 90 days with a larval stage lasting up to 71 days. Dahms (1984b) mentioned that these times seemed excessive, but did not offer an explanation. Schmieder (1939) suspected that he had been working with either two or three dif-ferent species, even though he mentioned them only as M. chalybii. Whiting (1947) mentions that Schmieder was probably rearing as many as four different species. After carefully reviewing Schmieder’s papers (1933, 1938, 1939) and one by Whit-ing (1947) we suspected that he was working with at least three different species, and possibly as many as four (which we believed were M. chalybii, M. evansi, M. digitata, and M. femorata). According to Dahms (1984a), M. chalybii is reportedly common from the New Jersey area where Schmieder worked. Melittobia evansi was described by Dahms (1984a) from specimens that came originally from our labora-tory in Athens, GA, and also from Goshen, N.J. reared by Schmieder. The third species, M. digitata, also described by Dahms (1984a), appears in a photograph in Whiting (1947) credited as from Schmieder’s cultures. The male in the photograph clearly exhibits the scape morphology characteristic of M. digitata.
The fourth species, M. femorata, is a common species in Georgia, Ala-bama, North and South Carolina, but also occurs in New Jersey, and we have also collected it in West Virginia, Virginia, Kentucky, Ohio and Michigan. As yet it is poorly studied. However, we have maintained it in the laboratory for several years. Even at a constant temperature of 25o C, it has an obligatory prepupal diapause on T. politum hosts that lasts for several weeks, and is the only Melittobia species we know with this life history variation. Thus, its total life history could easily last 90 days or longer. Recently, we were able to study voucher specimens from Schmieder’s and Whiting’s research deposited at the Smithsonian Institution (NMNH). These confirmed our suspicions; some speci-mens labeled as M. chalybii actually are M. evansi, M. digitata and M. femorata.
ACKNOWLEDGEMENTSTwo anonymous reviewers provided insightful comments on the origi-
nal manuscript. Eric Grissell and Michael Gates kindly helped us to access Schmieder’s and Whiting’s Melittobia vouchers at the National Museum of Natural History. This research was supported by NSF Grant 0088021, R. W. Matthews, principal investigator.
LITERATURE CITEDAdams, R. F. 2002. Dynamics of daily progeny production in Melittobia digitata Dahms:
Baseline studies. Undergrad. Sci. Bull. 3: 17 –21.Assem, J. van den, H.A.J. in den Bosch, and E. Prooy. 1982. Melittobia courtship behav-
iour: a comparative study of the evolution of a display. Neth. J. Zool. 32(4): 427 – 471.Balfour-Browne, F. 1922. On the life-history of Melittobia acasta Walker, a chalcid parasite
of bees and wasps. Parasitology 14: 349 – 370.Buckell, E.R. 1928. Notes on the life history and habits of Melittobia chalybii Ashmead
(Chalcidoidea: Elachertidae). Pan-Pacific Entomol. 5: 14 – 22.Cônsoli, F.L., and S. B. Vinson. 2002a. Clutch size, development and wing morph differ-
entiation of Melittobia digitata. Entomol. Exp. Appl. 102: 135 – 143.Cônsoli, F.L., and S. B. Vinson. 2002b. Larval development and feeding behavior of the
wing dimorphics of Melittobia digitata Dahms (Hymenoptera: Eulophidae). J. Hym. Res. 11(2): 188 – 196.
Dahms, E. 1984a. Revision of the genus Melittobia (Chalcidoidea; Eulophidae) with the description of seven new species. Mem. Qld. Mus. 21(2): 271 – 336.
Dahms, E. 1984b. A review of the biology of species in the genus Melittobia (Hymenoptera: Eulophidae) with interpretations and additions using observations on Melittobia australica. Mem. Qld. Mus. 21(2): 337 – 360.

2002 THE GREAT LAKES ENTOMOLOGIST 91
Edwards, C.J., and D.H. Pengelly. 1966. Melittobia chalybii Ashmead (Hymenoptera: Eulophidae) parasitizing Bombus fervidus Fabricius (Hymenoptera: Apidae). Proc. Entomol. Soc. Ont. 96: 98 – 99.
Farkas, J., and L. Szalay. 1985. Controlling of insect-parasites of alfalfa leaf cutting bee stock (Megachile rotundata F., Hymenoptera, Megachile). Apidologie 16(2): 171 – 180.
Girault, A.A. 1912. A new Melittobia from Queensland. Psyche, Cambr. 19: 203 – 205.Godfray, H.C.J. 1994. Parasitoids: Behavioral and Evolutionary Ecology. Princeton Uni-
versity Press, N.J. 488 pp.González, J.M. 1994. Taxonomía, biología y comportamiento de avispas parasíticas del
género Melittobia Westwood (Hymenoptera: Eulophidae) en Venezuela. Ph.D. Thesis, Universidad Central de Venezuela, Maracay, Venezuela. 118 pp.
González, J.M., and J.B. Terán. 1996. Parasitoides del género Melittobia Westwood (Hymenoptera: Eulophidae) en Venezuela. Distribución y hospederos. Bol. Entomol. Venez. (N.S.) 11(2): 139 – 147.
González, J.M., R.W. Matthews, and J.R. Matthews. 1985. A sex pheromone in males of Melittobia australica and M. femorata (Hymenoptera: Eulophidae). Fla. Entomol. 68(2): 279 – 286.
Hamilton, W.D. 1967. Extraordinary sex ratios. Science 156: 477 – 488.Howard, L.O. 1892. The habits of Melittobia. Proc. Entomol. Soc. Wash. 2: 244 – 249.Lith, J.P., van. 1955. Biologie van Melittobia acasta Walker (Hymenoptera, Chalcididae).
Tijdschr. Ent. 98; 29 – 42.Maeta, Y., and S. Yamane. 1974. Host records and bionomics of Melittobia japonica. Bull.
Tohoku Nat’l. Agric. Exp. Stn. 47: 115 – 131.Matthews, R.W., J. Yukawa, and J.M. González. 1985. Sex pheromones in male Melittobia
parasitic wasps: female response to conspecific and congeneric males of 3 species. J. Ethol. 3: 59 – 62.
Matthews, R.W., T.R. Koballa, jr., L.R. Flage, and E.J. Pyle. 1996. WOWBugs: New Life for Life Science. Riverview Press, Athens, GA. 318 pp.
Peck, O. 1969. Chalcidoid (Hymenoptera) parasites of the alfalfa leaf-cutter bee, Megachile rotundata in Canada. Can. Entomol. 101: 418 – 422.
Schmieder, R.G. 1933. The polymorphic forms of Melittobia chalybii Ashmead and the determining factors involved in their production, (Hymenoptera: Chalcidoidea, Eu-lophidae). Biol. Bull. Mar. Biol. Lab., Woods Hole 65: 338 – 352.
Schmieder, R.G. 1938. The sex ratio in Melittobia chalybii Ashmead, gametogenesis and cleavage in females and in haploid males (Hymenoptera: Chalcidoidea). Biol. Bull. Mar. Biol. Lab., Woods Hole 74: 256 – 266.
Schmieder, R.G. 1939. The significance of the two types of larvae in Sphecophaga burra (Cresson) and the factors conditioning them (Hymenoptera: Ichneumonidae). Ento-mol. News 50: 125 – 131.
Tasei, J.N. 1978. Action de deux insecticides sur Melittobia acasta Walk. et son hote: Megachile pacifica Pz. (Hymenoptera Eulophidae, Megachilidae). Méthode de préven-tion du parasitisme. Apidologie 9(2): 101 – 110.
Varanda, E.A., C.S. Takahashi, A.E.E. Soares, and L.A. de Oliveira C. 1984. Consider-ations of the courtship and copulation behavior, sex ratio and life cycle of Melittobia hawaiiensis (Hym. Eulophidae). Rev. Brasil. Genet. 7(1): 65 – 72.
Whiting, P.W. 1947. Some experiments with Melittobia and other wasps. J. Hered. 38: 11 – 20.Woodward, D.R. 1994. Predators and parasitoids of Megachile rotundata (F.) (Hymenop-
tera: Megachilidae), in South Australia. J. Aust. Ent. Soc. 33: 13 – 15.

92 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1

2002 THE GREAT LAKES ENTOMOLOGIST 93
FOOD PLANT AND DISTRIBUTION OF MELIGETHES SAEVUS (COLEOPTERA: NITIDULIDAE)
Andrew H. Williams1
ABSTRACTMeligethes saevus (Coleoptera: Nitidulidae) is here reported to be a spe-
cialist on its food plant Onosmodium molle (Boraginaceae). The geographic distributions of these taxa are discussed. This is the first report of M. saevus from Wisconsin.
____________________Beetles in the genus Meligethes (Nitidulidae) are often called “flower” or
“pollen” beetles because the adults and larvae, where known, feed on pollen, and some are restricted to particular food plants (Parsons 1943, Osborne 1965).
Meligethes saevus LeConte adults were found on Onosmodium molle A. Michaux (Boraginaceae) in a Wisconsin survey of the arthropod fauna using this locally rare, herbaceous perennial (Williams 1996). Adults were collected from flower buds, from flowers and often from among damaged unfurling leaves at shoot tips over the periods 7 May to 1 August, 1993-96, at 29 sites in Buf-falo, Dane, Grant, Green, Iowa and Pierce Counties. Many more observations of M. saevus on O. molle were made at these study sites than were vouchered by collections.
Solitary larvae were collected on 2 July 1996 from inside the tubular flow-ers of O. molle at one Wisconsin site where many M. saevus adults were present. These larvae resemble those of M. aeneus Fabricius depicted in Osborne (1965), bearing at the tips of their prothoracic legs setae characteristic of the subfamily Meligethinae (Böving and Craighead 1931), which in the Nearctic is represented only by the genus Meligethes (Parsons 1943).
Criddle (1928) collected M. saevus at Aweme, Manitoba, where he also collected O. molle (his plant specimen is at the Canadian Museum of Nature in Ottawa). In July 1996, on a brief pilgrimage to the Criddle homestead at Aweme, I collected M. saevus adults from O. molle in Bottineau, Foster, Nel-son, Rolette and Towner Counties, North Dakota, and at two sites in southern Manitoba, one of which was only a few miles south of Aweme. My own records of M. saevus on O. molle can be summarized as from 36 sites over a transect of about 600 miles, made on many occasions over several years.
Meligethes saevus has been reported from Illinois and Tennessee west to Missouri, Iowa, Minnesota, Kansas, Nebraska, South Dakota, North Dakota, Manitoba (Aweme), New Mexico, Colorado, Montana and Alberta (Lethbridge) (LeConte 1859, Popenoe 1877, Horn 1879, Leng 1920, Criddle 1928, Criddle 1929, Parsons 1943, Blackwelder and Blackwelder 1948, Easton 1955, Arnett 1975, Kirk and Balsbaugh 1975). I examined historic specimens from Illinois and Tennessee west to Iowa, Minnesota, Kansas, Nebraska, South Dakota, North Dakota, New Mexico and Colorado. Most of these specimens lack plant data, but six specimens at Iowa State University in Ames bear the data: “Ames, Iowa; June 14, 1892; on Onosmodium molle.”
With data from 137 herbaria, I mapped the distribution of O. molle, which grows widely over the entire region delimited above and is the sole member of the genus Onosmodium growing in this region (Williams 1996). This plant grows
1Department of Entomology, University of Wisconsin, Madison, WI, 53706.

94 THE GREAT LAKES ENTOMOLOGIST Vol. 35, No. 1
in gravelly or rocky calcareous prairies, barrens, openings, glades and bluffs (Williams 1996). Though it ranges widely between the Appalachian and Rocky Mountains, it is most common in the central and western parts of it range, and becomes more rare in the east where its preference for sweet well-drained soils and abundant sunlight restrict its distribution. It grows farther south and also farther east than M. saevus has been collected, and the beetle should be sought in these areas wherever O. molle grows so that we might more fully understand the beetle’s distribution.
Two old reports suggest M. saevus may also feed on a closely related plant. Gibson (1919) reported M. saevus on Mertensia paniculata (Aiton) G. Don (Bor-aginaceae) in Manitoba, where, quoting J. B. Wallis, the species, ‘occurred along the line of the Hudson Bay Ry., wherever its food plant grew.’ Gibson (1919) also reported Carr’s 1915 record of M. saevus at Edmonton, Alberta, which is farther north than O. molle occurs (Williams 1996). These more northern reports are probably referable to Meligethes canadensis Easton. This species was described in 1955 from paratypes that included some collected by Carr at Edmonton in 1910 and 1919 and some that had earlier been labelled M. saevus (Easton 1955). The suspected relationship between M. canadensis and Mertensia paniculata requires confirmation in the field.
My specimens have been deposited at the Wisconsin State Herbarium and the Insect Research Collection of the University of Wisconsin - Madison. Plant nomenclature herein follows Gleason and Cronquist (1991). This is the first report of M. saevus larvae and adults using O. molle and of M. saevus from Wisconsin.
ACKNOWLEDGMENTSThis work was part of my MS thesis research which could not have been
completed without the assistance of E. Williams, R. Christoffel, D. Young, R. Kowal and T. Vale. I am grateful to E. Moore, who financed my trip to Manitoba with the admonition, “Follow your beetle,” and to M. Price for bibliographic help. Essential to mapping O. molle were the data shared by curators of 137 herbaria (see Williams 1996). I am also grateful to curators of 65 insect collections: Academy of Natural Sciences, Philadelphia; Agriculture and Agri-Food Canada, Ottawa; American Museum of Natural History, New York; Arizona State University, Tempe; California Academy of Sciences, San Francisco; Carnegie Museum of Natural History, Pittsburgh; Cincinnati Museum of Natural His-tory, Cincinnati; Clemson University, Clemson; Colorado State University, Ft. Collins; Cornell University, Ithaca; Dayton Museum of Natural History, Dayton; Field Museum of Natural History, Chicago; Harvard University, Cambridge; Illinois Natural History Survey, Champaign; Iowa State University, Ames; Kansas State University, Manhattan; Los Angeles County Museum of Natural History, Los Angeles; Louisiana State University, Baton Rouge; Michigan State University, East Lansing; Milwaukee Public Museum, Milwaukee; Mississippi State University, State College; Montana State University, Bozeman; New York State Museum, Albany; North Carolina Dept. of Agriculture, Raleigh; North Carolina State University, Raleigh; North Dakota State University, Fargo; Northern Arizona University, Flagstaff; Ohio State University, Columbus; Oklahoma State University, Stillwater; Oregon Dept. of Agriculture, Salem; Oregon State University, Corvallis; Pennsylvania State University, University Park; Purdue University, West Lafayette; Royal Ontario Museum, Toronto; Rutgers University, New Brunswick; South Dakota State University, Brook-ings; Southern Illinois University, Carbondale; Texas A & M University, College Station; Université du Québec, Chicoutimi; University of Alberta, Edmonton; University of Arizona, Tucson; University of Arkansas, Fayetteville; University of British Columbia, Vancouver; University of California, Berkeley; University

2002 THE GREAT LAKES ENTOMOLOGIST 95
of California, Davis; University of California, Riverside; University of Colorado, Boulder; University of Connecticut, Storrs; University of Delaware, Newark; University of Idaho, Moscow; University of Kansas, Lawrence; University of Louisville, Louisville; University of Minnesota, St. Paul; University of Missouri, Columbia; University of Missouri, St. Louis; University of Nebraska, Lincoln; University of Oklahoma, Norman; University of Vermont, Burlington; University of Wisconsin, Madison; University of Wyoming, Laramie; Utah State University, Logan; Virginia Polytechnic Institute, Blacksburg; Washington State University, Pullman; and the personal collections of Edward G. Riley and Shawn M. Clark.
LITERATURE CITEDArnett, R. H., Jr. 1975. Nitidulidae, Family 78. pp. 1-18. In: R. H. Arnett, Jr. (ed.), Check-
list of the beetles of North and Central America and the West Indies, vol 5, Ladybird beetles and related groups. Flora & Fauna Publications, Gainesville, FL.
Blackwelder, R. E. and R. M. Blackwelder. 1948. Fifth supplement to the Leng catalogue of the Coleoptera of America, North of Mexico. Lane Press, Burlington, VT.
Böving, A. G. and F. C. Craighead. 1931. Illustrated synopsis of the principal larval forms of the order Coleoptera. Brooklyn Entomol. Soc., Brooklyn, NY.
Criddle, N. 1928. Entomological record, 1927. pp. 92-103. In: 58th annual report Entomol. Soc. Ont. Ont. Dept. Agr., Toronto, Ont.
Criddle, N. 1929. Entomological record, 1928. pp. 110-124. In: 59th annual report Entomol. Soc. Ont. Ont. Dept. Agr., Toronto, Ont.
Easton, A. M. 1955. Revision of the Nearctic species of the beetle genus Meligethes (Niti-dulidae). Proc. US Natl. Mus. 104(3339):87-103.
Gibson, A. 1919. Entomological Record, 1918. pp. 97-123. In: 49th annual report Entomol. Soc. Ont. Ont. Dept. Agr., Toronto, Ont.
Gleason, H. A. and A. Cronquist. 1991. Manual of vascular plants of northeastern United States and adjacent Canada. 2nd ed. N. Y. Bot. Garden, Bronx, NY.
Horn, G. H. 1879. Revision of the Nitidulidae of the United States. Trans. Amer. Entomol. Soc. 7:267-336.
Kirk, V. M. and E. U. Balsbaugh, Jr. 1975. List of the beetles of South Dakota. SD St. Univ. Agr. Exp. Sta. Tech. Bull. 42. Brookings, SD.
LeConte, J. L. 1859. Coleoptera of Kansas and eastern New Mexico. Smithson. Contrib. Knowl.11:1-58.
Leng, C. W. 1920. Catalogue of the Coleoptera of America, North of Mexico. Cosmos Press, Cambridge, MA.
Osborne, P. 1965. Morphology of the immature stages of Meligethes aeneus (F.) and M. viridescens (F.) (Coleoptera, Nitidulidae). Bull. Entomol. Res. 55:747-759.
Parsons, C. T. 1943. Revision of Nearctic Nitidulidae (Coleoptera). Bull. Mus. Comp. Zool. 92(3): 1-278, 13 pl.
Popenoe, E. A. 1877. List of Kansas Coleoptera. Trans. Kans. Acad. Sci. 5:21-40.Williams, A. H. 1996. Conservation of the plant Onosmodium molle A. Michaux (Boragi-
naceae) and the beetle Longitarsus subrufus LeConte (Chrysomelidae) in Wisconsin. MS thesis. Univ. of Wisconsin, Madison, WI. Available from: University Microfilms, Ann Arbor, MI; Order #1382083.





























