Embed Size (px)
Citation preview

Proceedings of the National Academy of SciencesVol. 68, No. 3, pp. 672-676, March 1971
The Gramicidin A Transmembrane Channel: A Proposed 7r(L,D) Helix
D. W. URRY
Division of Molecular Biophysics, Laboratory of Molecular Biology, University of Alabama MedicalCenter, 1919 Seventh Avenue South, Birmingham Ala. 35233
Communicated by Henry Eyring, January 4, 1971
ABSTRACT A lipophilic, left-handed helical structureis proposed for gramicidin A in which the C-O bonds alter-nately point toward the amino and carboxyl ends; it is ahybrid of the 4.314 and 4.416 helices. The C-O groups point-ing toward the carboxyl end form part of 16-memberedhydrogen-bonded rings, whereas the C-O moieties point-ing toward the amino end form 14-membered hydrogen-bonded rings. The proposed structure is based on con-formational analysis combined with requirements forthe gramicidin A transmembrane channel. Two helicescombine to form the channel. The alternating C-O direc-tions allow hydrogen-bonded dimerization by the uniquepossibilities of head-to-head and tail-to-tail attachment.The formyl group at the amino end allows for a favorablehead-to-head attachment with no loss of structural con-tinuity. Unpublished studies. by M. C. Goodall on thelipid bilayer conductance of deformyl gramicidin Astrongly argue for head-to-head attachment. Such hydro-gen-bonded association is not possible with previouslydescribed helices, as the C-O groups all point in the samedirection. In relation to possible 7r(LD) helices in mammal-ian systems, it should be noted that glycines would fillthe role of D residues.The conformation can undergo ion-induced relaxations,
which provide approximate tetrahedral coordination forthe ion, with facile shifting of coordinations. The readyexchange of coordinations provides the mechanism formovement of the ion along the channel. Conceivably,such transmembrane channels could have application asmodels for ion transport across biological membranes-anapplication which may be as great as, or greater than, thatof carriers such as valinomycin and nonactin. Specifically,biogenic amines and drugs containing aromatic groupscould control access to the channel by interactions withthe two tryptophan residues at the ethanolamine end andwith the negative region provided by the three oxygens.
Two established mechanisms of achieving ion flux across lipidbilayer membranes are by neutral carriers such as valinomycin(1) and nonactin (2), and by the formation of transmembranestructures as occur with nystatin, amphotericin B, tyrocidins(3), and gramicidin A. One example of a transmembranestructure would be the pores, 7-11 A in diameter, formed byamphotericin B (4-5) as evidenced by hydrodynamic flow(6). However, a transmembrane structure need not be a poreof fixed and specific dimension. Instead the structure couldaccommodate ion flux by local relaxations of conformation.The local relaxations would be initiated by particular ionicspecies in such a way that the coordinations of the ion could besatisfied during transit in the channel. This would be quiteanalogous to, and a generalization of, the mechanism by whichthe neutral carrier nonactin successively displaces the hydra-tion shell of an ion by acyl oxygens (7).
Thus, the requirements of such a transmembrane channelfor lipid bilayers would be (a) a lipophilic exterior; (b) a lowenergy conformation that is sufficiently flexible to allow localion-induced relaxations of conformation; (c) a length suffi-cient to span the lipid portion of the bilayer and reflect mem-brane thickness-dependent conductances; and (d) the capacityto interact with ions without the perfect sequestering observedin the carrier mechanism (8-10), that is, the structure shouldbe one in which the interacted ion can readily, in a directionalmanner, exchange coordinations and thereby allow flow alongthe transmembrane channel.
Gramicidin A is a linear pentadecapeptide antibiotic con-taining alternating L and D residues (11-14) (see Fig. 1). Theend groups are blocked by formylation of the amino terminusand by amide-bond linkage to ethanolamine at the carboxyterminus (11, 12). Conductance measurements on gramicidinA-mediated ion flux across lipid bilayer membranes exhibitsteps of uniform height, which suggests that transmembranechannels are formed and that each channel contributes aspecific conductance (15), which tends toward space chargesaturation with increasing electrolyte concentration. Studiesof the effect of antibiotic concentration on equilibrium con-
ductances indicate that two molecules of gramicidin A arerequired per channel (16, 17, 3). Since the length of the lipidportion of the bilayer is 40-50 A, the two molecules of grami-cidin A must combine in such a way as to span this dimension.Because the bilayer thickness can vary, it is of interest to see
what effect this variable has on gramicidin A-mediated con-ductances. In this connection Goodall has observed an interest-ing lipid specificity (3). When the lipid is negatively charged,as in the case of phosphatidyl serine or phosphatidyl inositol,the initial rate of conductance due to gramicidinA is decreased.This may be understood in terms of repulsion of negativecharges on the surfaces of the bilayer. The charge repulsionresults in membrane thickening (Goodall, M. C., personalcommunication). The inference is that gramicidin A is a bettermediator for thinner membranes. This view is further sup-ported by the autocatalytic effect exhibited with phos-phatidyl serine bilayers (17, 3). As the concentration ofgramicidinA channels is increased, thenegative charge becomes
HCO-L-Vol-Gly-L-AIG-D-Leu-L-Alo-D-VOI-L-VOI -D - Vol- L- Try -
o- Lou- L-Try-D-Leu- L-Try- D- LOu- L-Try- NHCH2CH20H
FIG. 1. Primary-structure of gramicidin A (11).
672
Dow
nloa
ded
by g
uest
on
Dec
embe
r 28
, 202
1
Gramicidin A Transmembrane Structure 673
neutralized by positive ions. This removes the negative chargerepulsion and allows thinning of the membrane, with resultantincrease in rate of conductance development. The initial ratealso increases with increased ionic strength (Goodall, utn-1)Lblished) and with application of a transmembrane l)otell-tial. Counter-ion clouds would neutralize the charge rep)ulsiolland result in thinning of the inemibrane (Goodall, personalcommunication). Application of a trausmembrane potentialwould also result in a decreased thickness of the membrane.Thus, for the gramicidin A transmembrane channel we havetwo specific conditions: (a) two molecules are required to forma, channel spanning the lipid region, and (b) the length of thechannel is closer to 40 A than to 50 A. In this connection itmay be recalled that, when the fatty acid moieties of the lipidhave aln average chain length of 18, the thickness of the lipidlayer would be approxinately 45 A (18). The above are usefulconsiderations in arriving at a possible transmembrane chan-nel for gramicidin A.
Conformational analysis of the grainicidin Alipophilic structure
All of the side chains of gramicidin A are lipophilic (see Fig. 1).Only the peptide moieties and the terminal ethanolaminegroup may be considered polar. In general. therefore, one isseeking a conformation in which the side chains are in the lipidmedium and the required polar interactions of the peptides aresatisfied by intra or intermolecular hydrogen bonding and byinteraction at the polar surface of the membrane. We will firstconsider the interchain hydrogen bonding conformations.That a stable and regular structure exists can be seen from
3
2
0qt
x
-I
-2
-3
I % b
/I II
GRAMICIDIN A
a. Dioxone 570 C
b. Trimethylphosphate27° C
\ b- , /
II IIl//\l//
II
\\ II
\\ I
I22 240200 220 240
X (nm)
FIG. 2. Circular dichroismn spectrum of gramicidin A indioxane at 570C and trimethyl phosphate at 270C. The largenegative ellipticities are similar in magnitude to a-helical polypep-tides and proteins. Such large negative ellipticities imply an
ordered structure even at elevated temperatures.
H
N
oC_xI
R'
I 0 - H*-0H'vNH /
C/ ~~~~~~N0 ~0
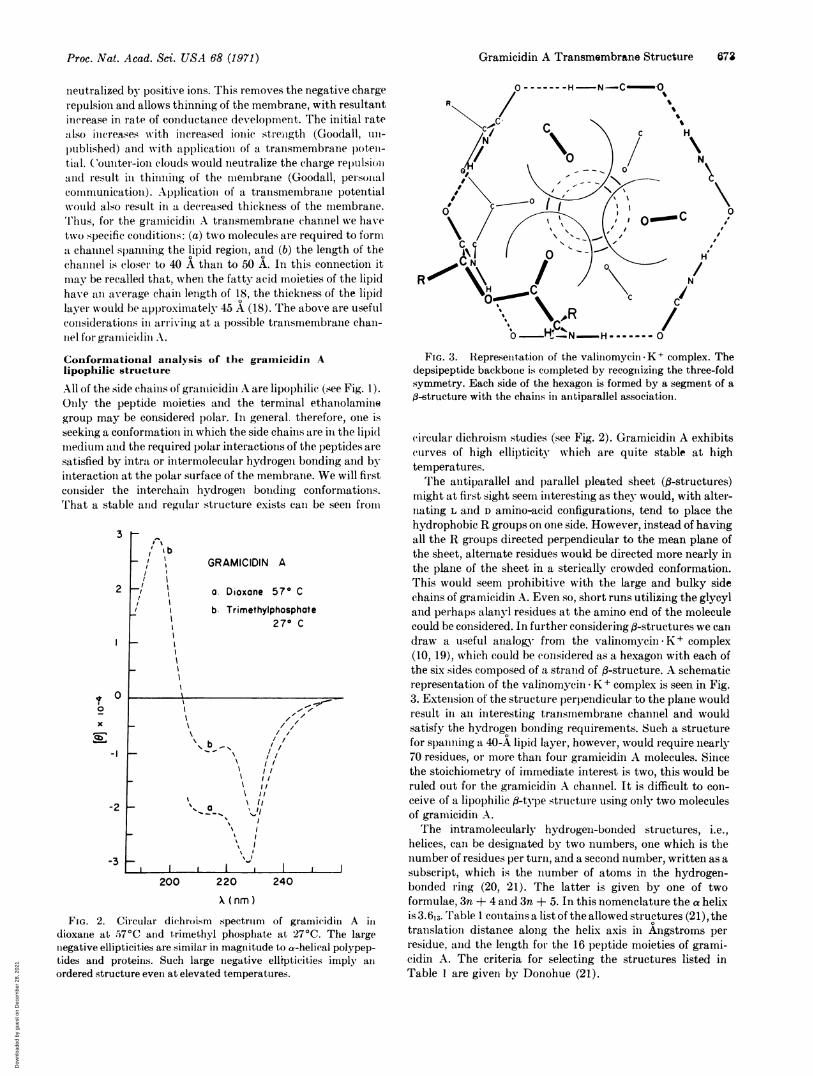
FIG. 3. Represenitationi of the valiniomycin-K+ complex. Thedepsipeptide backbone is completed by recognizing the three-foldsymmetry. Each side of the hexagon is formed by a segment of a--structure with the chains in antiparallel association.
circular dichroism studies (see Fig. 2). Gramicidin A exhibitscurves of high ellipticity which are quite stable at hightemperatures.The antiparallel and parallel pleated sheet (,8-structures)
might at first sight seem interesting as they would, with alter-nating L and D amino-acid configurations, tend to place thehydrophobic R groups on one side. However, instead of havingall the R groups directed perpendicular to the mean plane ofthe sheet, alternate residues would be directed more nearly inthe plane of the sheet in a sterically crowded conformation.This would seem prohibitive with the large and bulky sidechains of gramicidin A. Even so, short runs utilizing the glycyland perhaps alanyl residues at the amino end of the moleculecould be considered. In further considering (-structures we calldraw a useful analogy from the valinomycin K + complex(10, 19), which could be considered as a hexagon with each ofthe six sides composed of a strand of f-structure. A schematicrepresentation of the valinomycin K+ complex is seen in Fig.3. Extension of the structure perpendicular to the plane wouldresult in an interesting transmembrane channel and wouldsatisfy the hydrogen bonding requirements. Such a structurefor spanning a 40-A lipid layer, however, would require nearly70 residues, or more than four gramicidin A molecules. Sincethe stoichiometry of immediate interest is two, this would beruled out for the gramicidin A channel. It is difficult to con-ceive of a lipophilic fl-type structure using only two moleculesof gramicidin A.The intramolecularly hydrogen-bonded structures, i.e.,
helices, can be designated by two numbers, one which is thenumber of residues per turn, and a second number, written as asubscript, which is the number of atoms in the hydrogen-bonded ring (20, 21). The latter is given by one of twoformulae, 3n + 4 and 3n + 5. In this nomenclature the a helixis 3.613. Table 1 contains a list of the allowed structures (21), thetranslation distance alone the helix axis in Angstroms perresidue, and the length for the 16 peptide moieties of grami-cidin A. The criteria for selecting the structures listed inTable I are given by Donohue (21).
.
t
Proc. Nat. Acad. Sci. USA 68 (1971)
Dow
nloa
ded
by g
uest
on
Dec
embe
r 28
, 202
1

674 Biochemistry: D. W. Urrv
H
/N~~~~~~~~~~~~~~~~~~~
C---
IA/R
FIG. 4. H1elix axis perspective of the left-halded r(I_ 0) helix.Note the two distinct, hydrogen-bonided rings. The hydrogeni bondat the left-hand side of the figure completes a 14-inembered hydro-gen-bonded ring, while that at the top of the figure forms a 16-membered hydrogent-bonded ring.
Although the 2.27 helix is of a proper length for penetratingthe lipid layer, it does iot, with two molecules, formii a lil)o-philic structure. The 310 helix would be a lipophilic structure,but two molecules attached head to tail would have at lengthof more than 64 A. Such a length would be unnecessary andwould not reflect the membrane thickness dependences. The aor 3.613 helix would be lipophilic, two molecules head to tailwould give a structure about 50 A long, and the same helixsense could conceivablv contain b)oth L and D residues. Thelatter follows fromt the calculations of Ooi et al. (22), whereillonly a small difference in energy between the right- and left-handed helices is found for l)oly(L-valine). This implies thatone helical sense could accommodate both configurations. Thedrawbacks for this structure are that it lies in a sharp coni-formational energy minimum (22), implying a rigid structure,and that the length of 50 A would not explain the membranethinning effects. These are not compelling disadvantages, assome estimates of lipid layer thickness are greater than 50 A(23), and as the alternating L and D amino-acid sequence miaygive flexibility. The 5.117 or -y helix (can be readily excluded asit is too short. The 4.314 and 4.41c (7r) helices are of approxi-mnately the correct length but they have a. prefelellce forhandedness with a given amino acid configuration, whichmeans that they would be unstable with an alternatilln L-I)sequence.There is. however a unique helix obtained by alternating
TABLE 1. Possible helical structures
Helix 2.27* 310 3.613(c) 4.314 4.416(7r) 5.117(Y)
Length perresidue (A) 2.75 2.0) 1.5 1.2 1.15 0.98
Length of 16peptides(A) 44 :32 24 19.2 18.4 15..7
* For explanation of designations of helical structure see text.
the direction of the C-O bond. The result is a hybrid betweenthe 4.314 and 4.416 helices with alternating 14- and 16-mem-hered hydrogen-bonded rings (see Fig. 4). We will refer to thisconformationi as a 7r(L,n) helix. The struictuire is lipl)0)hilic, withthe R groups directed radially into solutioll.The above analysis leaves, the a helix as a marginal possi-
bility and the 7r(LD) helix as an interesting conformation for thegramicidin A transmnembrane channel.
Characteristics of the 7r(L,D) helix
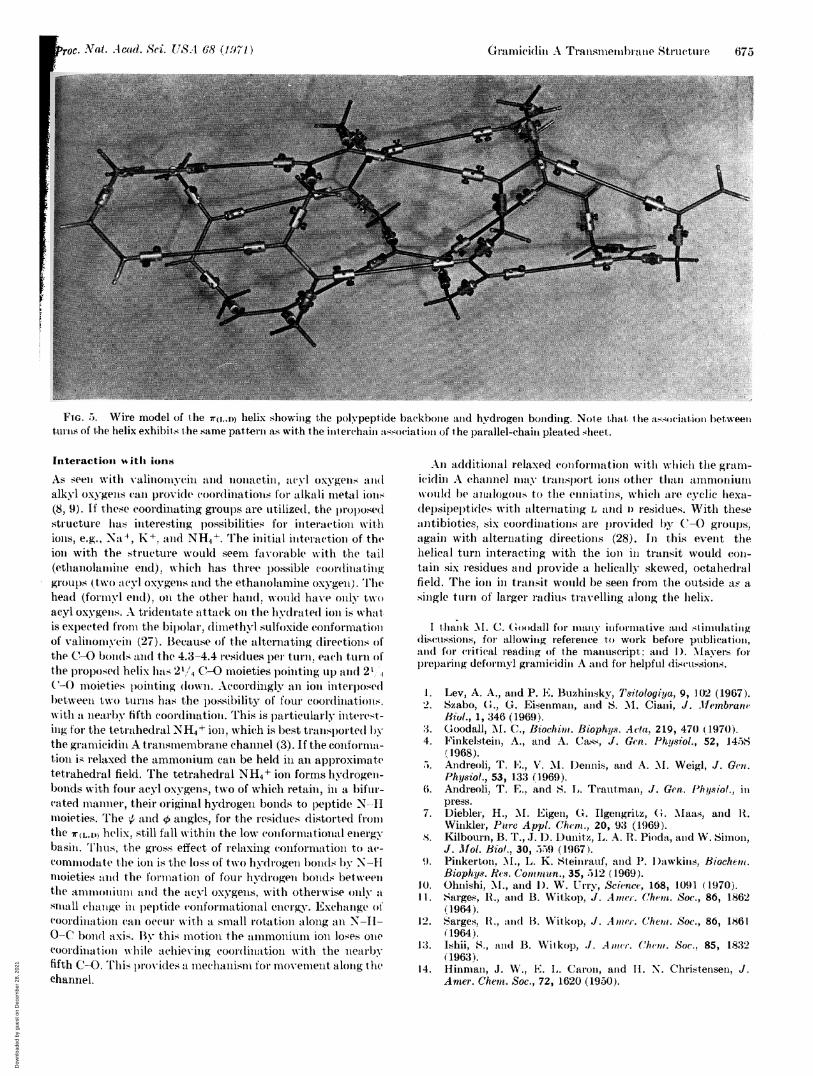
As Iloted above, an interesting helix cane be constructed byalternating the direction ill which the (-O points (first towardlthe amino end and then toward the carboxyl end of the helicall)olypeptide) and by utilizing 14- and 16-membered hydrogen-bonded rings. With the exception.s of the ends, all leltides arep1operly hydrogen-bonded anid the R groups are all directe(lradially into solution. This provides a very lipophilic strtu-ture. If we follow the normnenclature of Edsall el al. (24) a1ltake proper cognizance of the alternating L-D configurations,the two dihedral angles specifying the structure are ~ 270'and o - 60°. Reference to dihedral angle conformationalenergy maps (22, 24, 25) shows this contformation to be in atflat low-energy basin. Thus, iol)-induced relaxations of (0ol-formation could readily occur. C(ollsidering the end grou)s, thelength of two inolecules would be 40 A. This length wouldexplain the observed membrane thickness dependence. Twoadditional features of the helix may be noted. Whemi thesequence nuumbers of the L residues are odd, the helix sense isleft-handed, and wvhen the- are exent, it is right-handed. Whenthe structure is rel)resented as an opened cylinder the hydro-geli-bonding lpattelll is that of a l)aallel pleated sheet, (see Fig.5).
Previously described helices have (-O bond vectors iniwhich the comlponents along the helix axis point in the sainedirectioll. A ullique aspect of the 7r(LD) helix is the alternatingdirection of the C-0 bolid. This alternation provides newl)ossibilities for dimerization by hydrogen bonding, thesebeing head-to-head and tail-to-tail coupling. Analysis of thestructure shows that head-to-head attachment would restult illvirtlually uninterrupted structural continuity. This is palrtiell-larlv so vith forinvlation of the N-terminus. On the otherhand, N-acetylation wvith two inethylis occurring at the junc-tion would be less favorable. Head-to-head coupling (couldl)rovide an explanation for the formyl moiety, which is a rareNH2-terminial blocking group (12). With the possibility ofhead-to-head attachment in mind, deformyl gramicidin A was
p)re)ared (26). Identity of the product was verified by imuIr at220 MHz (Glickson, J., and D. W. U{rry, unpublished). Whentested by Goodall for its ion-transport l)roperties it was foundto be inactive. Furthermore, it did not competitively inhibit.the action of gramicidin A, even when deformyl gramicidlin Awas at a concentration 10,000 times that of gramicidin A.These results strongly sulpport head-to-head attachment. and,therefore, the unique structure lproposed.** Another unusual feature of the proposed conformation is thelarge aCH-NH dihedral angle (160-180O). Stuch large dihedralangles result in uniquely large coupling constants, e.g. greaterthan 7 Hz (Jiamachandran, G. N., 1I. Chandrasekarun, andK. D. Kopple, private communication). Preliminary nmr studiesait 220 MIHz show several resolved peptide proton resomlaiices tohalve coupling constants of 8-10 Hz, which are the values expectedolnly for 160-180( dihedral angles.
Proc. Nat. Acad. Sci. USA 68 (1,971)
Dow
nloa
ded
by g
uest
on
Dec
embe
r 28
, 202
1
G.c6(Iamlui(liin ArTransmnemnbl ne Structure 675
FIG. 5. Wire model of the 7r(l,,D) helix showing the polypeptide backbone and hydrogen bonding. Note that. the association betweenturns of the helix exhibits the same pattern as with the inter(chaini association of the pairallel-chain pleated s-heet.
Interactioni 4 itL ions
As seeii with valinoinycin aand nonactin, acyl oxgellns an1alkyl oxygens can provideC C(oldinationks for alkali metal ions(8, 9j). If these coordinating groups are utilized, the prol)osedlstructure has interesting possibilities for interaction withions, e.g., Na 4, K+, anld NH4+. The initial interactioni of theioI with the structure woulid seem favorable with the tail(ethanolain ine etid). which has th iee loss-ible coordinathinggroups (two acyl oxygens and the ethan-olaminie oxygeln). '1Thehead (forimyl end), on the other hand, would have only twoacyl oxygens. A tridentate attack on the hydr-ated ion is whltis expected fronm the hip)olar, d iunethyl sulfoxide conformationof valinonivein (27). Because of the alternating directions olthe C-()bonds and the 4.3-4.4 residues pel turn. each turn ofthe proposed( helix has 2',/4 C-0 moieties pointing lui) aII(I 21',('-O moieties pointinlg do(wn. Accordingly an ion inlterposell)etween two tunrns has the p)ossilility of four coordinatioIns.with a nearIbv fifth coordiination. This is particularly interest-ing for the tetrahedral NH4+ ioln, which is best transpoirte(l bythe gramicidin A tranismembrane channel (3). If the confolrmna-tion is- relaxed the ammon-ium can be held iii anr approximlatetetrahedral field. The tetrahedral NH4+ iOnl forms hydrogen-b)onds with four acyl oxygens, two of which retain, in a bifur-(ated nianier, their origiiial hydrogen b)o1nds to peptide N -Hmioieties. The ; andC angles, for the residues diStortted fro0m1the ?r(LI,[) helix, still fall within the low conformiational energybasini. Thus, the gross effect of relaxing con1forma1.1tion to ac-conmminiodate the ion is the loss of two hvdrogdeni bonids by N-Hmoieties an I the fort nation of fou1- hydirogei bonds l)etweethe aInionliunI anld the acvl oxygens, with otherwise ontly--asnila11 (chalgige inL)ept(le conformiational energy. Exchange ofcoordination can occur with a Small rotation along an N-Il-0-C bond .axis. Bv this motion the amnmonium ion loses on1ecoordination while achievim ig coor(lination with the niearbyfifth C-0. Thi; )r-ovides a mnechianism for imovemient alonig thechannel.
An additional relaxed conformatioi with which the gram-icidin A cha1Innel] nm-ay transl)O-t ijons otiher thi,-ti amillmoliunmwoild be analogolus to the enni:ltilns, which are cyclic hexa-(lepsip)el)tides with alternatilg L and I) residues. With theseaintibiotics, six coordiimations are )rovi(ded(by C-O groups,again with alternating directions (28). In this event thehelical turn interacting with the ion1 inI transit would con-tain six residues atnd provide a, helically skewed, octahedralfield. The ion in transit would be seen from the outside a:F asingle ttirn of larger racius travelling along the helix.
I thlank MI. C(. ()oodall for manya- iniforniative anid stimulatingdiscussions, for allowing reference to work before publication,and for critical reading of the manuscript: and I). Mavers forrep)aring deformyl granicidin A and for helpful discussions.
1. Lev, A. A., and P. E. Buzhinsky, Tsitologiya, 9, 102 (1967).2. Szabo, G;., G. Eisenmaln, and S. M. Ciani, J. Membrane
Biu,., 1, 346 (1969).:3. (3oodall, M. C., Biochimt. Biophys. Acta, 219, 470 (1970).4. Finkelstein, A., and A. Catss, J. Gen. Physiol., 52, 1458
(1968).5. Andreoli, T. El., V. M. l)ennis, and A. M. Weigi, .J. Gen.
Physiol., 53, 133 (1969).6. Andreoli, T. E., and S. L. Trautman, J. Gen. Physiol., in
press.7. Diebler, H., _M. Eigen, c. Jlgengritz, G. Maas, and It.
Winkler, Pure Appl. (Chem., 20, 93 (1969)..X. Kilbourn, B. T.,.J. 1). DuLn1itZ, L. A. R. Pioda, and WV. Simon,
J. .M1ol. Biol., 30, 559 (1967).9). Pinkerton, M., L. K. Steiiratuf, and P. 1a)awkinis, Biochein.
Biophys. Res. Conitnun., 35, 512 (1969).10. Ohnishi, 'M., and 1). W. Urry, Science, 168, 1091 (1970).ll. Sarges, R.., and B. Witkop, J. Anmer. (hent. Soc., 86, 1862
(1964).12. Sarges, It., andBl. Witkop, J. Amner. C(hem. Soc., 86, 1861
(1964).13. ISHii, S., and B. Witkop, .J. A mur. Chem. Soc., 85, 1832
(1963).14. Hinnian, J. Wi., E. I.. CaOro, and II. N. Christensen, J.
Amer. Chemn. Soc., 72, 1620 (1950).
c. Nat. Acad. 8ri. US-4 68 (19'1"1)
Dow
nloa
ded
by g
uest
on
Dec
embe
r 28
, 202
1

676 Biochemistry: D. W. Urry
15. Hladky, S. B., and D. A. Haydon, Nature (London), 225,451 (1970).
16. Tosteson, D. C., T. E. Andreoli, M. Tieffenberg, and P.Cook, J. Gen. Physiol., 51, 373S (1968).
17. Goodall, M.v. C., Biochim. Biophys. Acta, 219, 28 (1970).18. Hanai, T., D. A. Haydon, and J. Taylor, Proc. Roy. Soc.
Ser. A, 281: 377 (1964).19. Urry, D. W., and M. Ohnishi, in Spectroscopic Approaches
to Biomolecildar Conformation, ed. 1). W. Urry (AmericanMedical Association Press. Chicago, 1970), p. 263.
2-0. Bragg, W. K., J. C. Kendrew, and M\. F. Perutz, Proc. Roy.Soc. Ser., 203, 321 (1950).
21. Donohue, J., Proc. Nat. Acad. Sci. USA, 39, 470 (1953).22. Ooi, T., R. A. Scott, G. Vanderkooi, and H. A. Scheraga,
J. Chem. Phys., 46, 4410 (1967).
Proc. Nat. Acad. Sci. USA 68 (1975)
23. Cherry, R. J., and D. Chapman, J.MoN. Biol., 40, 19 (1969).24. Edsall, J. T., P. J. Flory, J. C. Kendrew, A. M. Liquori,
G. Nemethy, G. N. Ramachandran, and H. A. Scheraga,Biopolymers, 4, 121 (1966); J. Biol. Chem.. 241, 1004 (1966):J. Mot. Rio!., 15, 399 (1966).
25.BgibsonW K.i)., and H. A. Scheraga, Biopolymers, 4, 709(1966).
26. Ishii, S., and B. Witkop, J. Amer. Chem. Soc., 86, 1848(1964).
27. Urry, D. W., in Chemical Dynamics ed. J. 0. Hirschfelderand 1). Henderson (John Wiley & Sons, 1971), p. 581.
28. Ovchinnikov, Y. A., V. T. Ivanov, A. V. Evstratov, V. F.Bystrov, N. D. Abdulaev, E. M. Popov, G. M. ipkind, S.F. Arkhipova, E. S. Efremov, and M. M. Shemyakin,Biochem. Biophys. Res. Commun., 37, 668 (1969).
Dow
nloa
ded
by g
uest
on
Dec
embe
r 28
, 202
1





























