Embed Size (px)
Citation preview

Oikos Editorial Office
The Genus Pseudocalliergon in Northern EuropeAuthor(s): Lars HedenäsSource: Lindbergia, Vol. 16, No. 3 (1990), pp. 80-99Published by: Oikos Editorial OfficeStable URL: http://www.jstor.org/stable/20149763 .
Accessed: 14/06/2014 01:13
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Oikos Editorial Office is collaborating with JSTOR to digitize, preserve and extend access to Lindbergia.
http://www.jstor.org
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

LINDBERGIA 16: 80-99. Copenhagen 1990
The genus Pseudocalliergon in northern Europe
Lars Heden?s
9
Heden?s, L. 1992. The genus Pseudocalliergon in northern Europe. -
Lindbergia 16: 80-99.
The genus Pseudocalliergon (Limpr.) Loeske is revised for northern Europe. The
genus is interpreted as consisting of P. angustifolium Heden?s, sp. nov., P. lycopo dioides (Brid.) Heden?s, comb, nov., P. brevifolium (Lindb.) Fleden?s, comb, nov., P. turgescens (T. Jens.) Loeske and P. trifarium (Web. & Mohr) Loeske. The genus is defined by, i.a., the typical colour and gloss of the species, the structure of the alar cell group, the structure and colour of the axillary hairs and the ecology of the
species. All the species grow in lime-rich and, except for P. trifarium, small and often shallow and periodically dry wetland habitats (sometimes the habitats are flushed
with ?lime-rich water). In northern Europe, P. angustifolium grows in the Scandina vian mountain range and in the more eastern areas also in the Northern Boreal zone. P. lycopodioides reaches north to Iceland and S Norway in the west and approxi mately to the Middle Boreal zone in the east. P. brevifolium occurs only in the arctic areas. P. turgescens is found in the Scandinavian mountain range, north-eastwards to north-westernmost Finland and to Troms in Norway, and occurs also in (probably) relict localities around the Baltic Sea. P. trifarium occurs throughout the area. The taxonomic position of the genus is discussed.
L. Heden?s, Dept of Botany, Univ. of Stockholm, S-106 91 Stockholm, Sweden
(present address: Dept of Cryptogamic Botany, Swedish Museum of Natural History, S-104 05 Stockholm, Sweden).
Poa Pseudocalliergon (Limpr.) Loeske nepecMOTpeH othoc CeBepHOM EBponu. IlHTepnpeTauHJi pOAa: oh coctomt m3 P. angustifolium Heden?s, hob. BHA, P. lycopodioides (Brid.) Heden?s, hob. kom6.,P. brevifolium
(Lindb.) Heden?s, hob. kom6., P. turgescens (T. Jens.) Loeske h P.
trifarium (Web. & Mohr) Loeske. Poa onpeAejweTca, cp. np., tm?imhhbim
mseTOM m 6jiecKOM BHAa, crpyicrypoH na3yiiiHOH rpynnu kjictok, crpyicr ypoM h uBeTOM oceBbix BOJioc h OKOJiorneH BHAa. Bee bhau, 3a hckjhohc uneM P. trifarium, pacryT b 6oraTux M3BecTbio v3Khx h nacro mcjiko
BOAHbix, nepnoAHMecKH Aaace cyxnx aaGojioneHUbix Mecrax o6nTannfl
(HuorAa Mecra oGHTaima 3ajmBaioTOi +/- 6oraTo?? H3Becrbio boao??). B Ce?epHOH E?pone P. angustifolium pacr?T Ha CKaiiAHHaBCKOM ropiiOM KtaccHBe h b 6ojiee boctohhux TeppnTopnax, a TamKe b CeBepuo?? 6opea jii>hoh 30He. P. lycopodioides AOCTHraeT Ha ceBepe McjiaHAHH, Ha 3anaAe IO^iioh HopBerHH h npMMepiio CpeAHen 6opeajn>HOH 30iibi Ha boctokc
P. brevifolium BcrpenaeTca jiHiiib b apKTHnecKHx TeppHxopH?X. P. tur
gescens BcrpeiaeTCJi tia CKaiiAHHaBCKOM ropiiOM MaccMBe, Ha cesepo BocroKe h ce?epo-3anaAe Ohhji?ihahh h b o6jiacTH TpoMC b HopBernn, oh BCTpenaeTCii (BepO?THo) Tamse b pejiMKTOBux Mecrax BOKpyr BajiTH ??cKoro Mop)i. P. trifarium BCTpenaeTCfl noBCWAy b AanHOM pernone.
06cy;KAaeTC? TaKCOHOMHnecKoe nojio^enwe poAa.
The section Pseudocalliergon of the genus Hypnum was erected by Limpricht (1989) for the species Loeskyp num badium (Hartm.) Paul, Calliergon trifarium (Web. & Mohr) Kindb. and Pseudocalliergon turgescens (T. Jens.) Loeske. Loeske (1907) used the name Pseudocal
liergon at the generic level for C. trifarium, P. turgescens and Campylium longicuspis (Lindb. & H. Arn.) Hede n?d. Paul (1924) treated these species, except L. ba
Accepted 26 August 1991
? LINDBERGIA
80
dium, in Scorpidium, where the species Drepanocladus
lycopodioides (Brid.) Warnst., D. brevifolius (Lindb.) Warnst., D. latifolius (Lindb. & H. Arn.) Warnst, and
Scorpidium scorpioides (Hedw.) Limpr. were also in
cluded. Except for Campy Hum longicuspis, Tuomikoski and Koponen (1979) included the same species in Scor
pidium and added the species Hamatocaulis vernicosus
(Mitt.) Heden?s and H. lapponicus (Norrl.) Heden?s to
LINDBERGIA 16:3(1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

this genus. While most authors of today (e.g., Smith
1978, Crum and Anderson 1981) do not recognize any
taxon with the name Pseudocalliergon, Nyholm (1965) treats Calliergon trifarium in Calliergon sect. Pseudocal
liergon (Limpr.) Broth, and Karczmarz (1971) treats C.
trifarium and P. turgescens in the same section. Smir
nova (1962) treats Drepanocladus lycopodioides, D.
brevifolius and D. latifolius in Drepanocladus sect. Tur
gidus. In a revision of Scorpidium (s.l.) and Limprichtia,
Heden?s (1989c) suggested that Calliergon trifarium, Pseudocalliergon turgescens and the three Drepanocla
dus species mentioned above are closely related and
that they should be treated in the same genus, Pseudo
calliergon. In the present paper the genus Pseudocal
liergon is taxonomically revised for nothern Europe.
The area covered is the same as treated by Nyholm
(1987).
Material and methods
This revision is based mainly on herbarium material,
but all species except Drepanocladus brevifolius (incl. D. latifolius) were studied in the field as well. All to
gether, more than 1600 specimens of the species recog
nized in Pseudocalliergon were studied. The specimens are located in C, GB, H, ICEL, LD, O, OULU, S,
TRH, TUR, UME and UPS. A few specimens were
also borrowed from LE and from herb. K. Dierssen.
Type material was requested from the following herb
aria (in addition to some of those just mentioned): B, BM, BP, BR, C, FH, G, GL, HBG, JE, KRA, L, LY,
M, MICH, NY, PC, POZ and W. I tried to check the types for names of species, sub
species and varieties suspected to belong to Pseudocal
liergon and reported from Europe. In addition, I tried to check the types of extraeuropean taxa which were
suspected (from information in the protologue or from
earlier synonymizations) to be of importance for the
nomenclature of European species. As regards taxa de
scribed by Sanio, I have mostly followed the judge ments of taxonomic level in Wijk et al. (1964).
From now on I will use the nomenclature suggested in
this paper within Pseudocalliergon to facilitate the un
derstanding of the discussion. Names of vegetation zones follow Ahti et al. (1968: Fig. 9).
Taxonomically important characters
Stem. The stem is unbranched or slightly and irregularly branched in Pseudocalliergon trifarium, slightly and ir
regularly branched in P. turgescens and usually in P.
brevifolium and slightly to more distinctly, but usually irreguarly, pinnately branched in P. lycopodioides and
LINDBERGIA 16:3 (1990)
P. angustifolium. The stem has got a central strand and
the cortex consists of l-3(?4) layers of incrassate cells
(Figs IL, 3L, 5M, 6K, 8L). Stem leaves. The measurements and cross-sections of
the stem leaves were made as described in Heden?s
(1989a, c). In the species with straight, rounded or
apiculate leaves (Pseudocalliergon trifarium, P. turges
cens) both leaf length and width were measured. In the
species with falcate leaves which gradually or ? sud
denly narrow to an acuminate apex (P. augustifolium, P.
lycopodioides, P. brevifolium) only leaf width was mea sured. In all species except P. angustifolium (Fig. 1C) the leaves are normally very broad in relation to their
length (Figs 3C, 5C, D, 6C, 8C) and more or less
strongly concave. These features, together, with the
rather crowded insertion of the leaves, often make the
shoots turgid (again with the exception of P. angustifo
lium). The leaves of species of Pseudocalliergon have
sometimes (P. trifarium) or often a characteristic golden
gloss when dry. This is not found in other species of the
Calliergon-Scorpidium-Drepanocladus (CSD) complex. In addition, the Pseudocalliergon species are often de
veloping a distinct brownish yellow colour which is
rarely parallelled in other CSD complex species (most close are some phenotypes of Drepanocladus sendtneri
(Schimp. ex H. M?ll.) Warnst.). In all five species, the alar cells and those just above
them, the supra-alar cells, are indistinctly delimited
from each other and from the surrounding cells (Figs.
U, 3J, 5K, 6J, 8J). The alar cells are quadrate, rectan
gular or longly rectangular and usually more or less
inflated. The walls may be thin but are mostly thick,
yellow and ?porose. In most specimens the alar cells
form an indistinct, transversely triangular group, but
sometimes they form a single row of cells or are appar
ently undifferentiated. The supra-alar cells are rectan
gular, qradrate or transversely rectangular and are
found in one or several rows along the leaf margin above the alar cells. Sometimes the supra-alar cells are
apparently not differentiated. The ontogeny of the alar
cells is of a modified Drepanocladus aduncus type (He den?s 1987a).
The leaf margin is distinctly denticulate (at least
partly) in Pseudocalliergon angustifolium, smooth or
often partly finely denticulate in P. lycopodioides and smooth or sometimes very finely denticulate in P. brevi
folium and P. turgescens. In P. trifarium the leaf margin is ?smooth.
Axillary hairs. In the genus Pseudocalliergon the axil
lary hairs have 1?2(?3) upper cells. The apical cells tend to be long or very long in comparison with most
other taxa of pleurocarpous mosses (Heden?s 1989d).
There is some variation in this last feature (cf. Figs IM, 3M, 5N, 6L, 8M), but in most specimens the majority of the hairs have got long apical cells. The cell walls of the
upper cells of the axillary hairs are getting yellowish or brownish early, i.e. close to the shoot apex and ? as
soon as they have their mature shape. I have only stud
81
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

ied this feature in herbarium specimens and not in fresh
material. The colouration may thus be due to prolonged
drying of the material, but, in any case, most other
pleurocarpous species do not have early coloured walls
of the upper axillary hair cells even when studied in
herbarium specimens. Perichaetial leaves. I have not seen mature perichae
tia in Pseudocalliergon angustifolium. In the other spe
cies the inner perichaetical leaves are concave and pli cate. In P. lycopodioides they are ?ovate and gradually or suddenly narrowed to an acuminate point (Fig. 3N). In P. brevifolium the perichaetial leaves are rather simi
lar, but slightly more shortly acuminate (Fig. 50). In P.
turgescens these leaves are even more shortly acuminate
(Fig. 6M) and in P. trifarium they are ?suddenly nar
rowed to an acute or obtuse apex (Fig. 8N).
Sexuality. All species of Pseudocalliergon are dioi
cous.
Features of the proximal branch leaves and of the
rhizoids seem to be of little value in the taxomony of
these species. However, rhizoids are rare and I have not
seen them in Pseudocalliergon brevifolium and P. tur
gescens.
Capsule. I have not seen sporophytes in Pseudocal
liergon angustifolium. In the other species, the capsule is of Amblystegium type (cf. Heden?s 1987b, 1989b). It is approximately horizontal in P. lycopodioides (Fig. 30) and P. brevifolium (Fig. 5P) whereas it is horizontal to inclined in P. turgescens (Fig. 6N) and P. trifarium (Fig, 80). It should be noted that I have only seen one
mature capsule of P. brevifolium. The exostome, and
especially the endostome processes are slightly nar
rowed in P. turgescens. and P. trifarium (as compared with the other species; Figs 6R, 8S), a condition which is most probably due to the often inclined capsules in
these species. The exothecial cells were studied in the
position indicated by Heden?s (1989a, c). Other features of the sporophyte were not found to
be of great value in separating the species.
Ecology. In northern Europe, all species of the genus
Psedocalliergon except P. trifarium tend to grow most
commonly in shallow, lime-rich wetland habitats. The
habitats are often small or very small in size (sometimes
only a few m2) and they commonly dry out perodically, or sometimes the habitats are flushed with ? lime-rich
water. Although P. trifarium differs somewhat from the
other species in its habitat preferences, it is still a spe cies of calcium-rich, but usually deeper and larger wet
lands and it is often growing close to the surface of the habitat. The genus is thus rather homogeneous as re
gards at least some aspects of the habitat preferences of
the included species.
82
Delimitation and phylogenetic considerations
As discussed by Heden?s (1989b, c; see also Heden?s
1988), it seems unlikely that the species of Pseudocal
liergon are closely related to the Warnstorfia-Calliergon
species of Tuomikoski and Koponen (1979) or to the other species which have been included in Scorpidium at one time or another (i.e. those of Scorpidium and
Hamatocaulis as interpreted by Heden?s (1989c) or
Campylium longicuspis (Heden?s 1988)). Characters
supporting a treatment of the Pseudocalliergon species in a different taxonomic position than the Scorpidium and Hamatocaulis species (as interpreted by Heden?s
1989c) are: (1) The absence of red colours, but, instead,
the presence of a characteristic brownish yellow or yel low-brown colour, often with a distinct golden gloss in
Pseudocalliergon; (2) The differences in the structure of the alar cell group; (3) The different kinds of leaf curva ture in Pseudocalliergon, Scorpidium and Hamatocaulis
(in species with curved leaves); (4) The axillary hairs, which have l-2(?3), often yellowish or brownish upper cells in Pseudocalliergon and 2-9 (?11) hyaline upper cells in Scorpidium and Hamatocaulis; (5) The well
developed cortex of incrassate cells, without hyaloder mis in the stems of Pseudocalliergon species as com
pared with the well developed cortex with hyalodermis in Scorpidium species and the weakly developed cortex
without hyalodermis in those of Hamatocaulis; (6) The lower outer peristomial layer is basically cross-striolate
(with occasional irregularities) in Pseudocalliergon whereas it is commonly or always (depending on which
species is considered) dotted or dotted-striate (partly or
entirely) in Scorpidium and Hamatocaulis; (7) The
largest spores tend to be somewhat smaller in Pseudo
calliergon (spore size range: 10.2-17.8 (-19.0) urn) than in Scorpidium (12.0-21.0 urn)) or Hamatocaulis
(10.5-24.5 pui). Features which support the treatment of the Pseudo
calliergon species in the same genus are: (1) The charac
teristic colouration and gloss; (2) The typical structure of the alar cell group (to me it seems very unlikely that
as complicated structures as the alar cell groups should
have developed as similar as in the Pseudocalliergon
species due to chance only); (3) The characteristic axil
lary hairs; (4) The similarities in habitat preferences. It is quite clear that the Pseusocalliergon species be
long to "group IV" (including the Amblystegiaceae, parts of the Hypnaceae and parts of the Leskeaceae/
Thuidiaceae) of Heden?s (1989b), but the exact posi tion within this group is difficult to assess. One may speculate on a relationship between the species of Pseu
docalliergon and those of Drepanocladus s. str. (i.e. the
species around D. aduncus (Hedw.) Warnst.), but at
present I am not aware of any synapomorphy joining
Pseudocalliergon with Drepanocladus s. str. with cer
tainty. The axillary hairs have 1-2 (-3) upper cells in both genera, but this character state is found also in
LINDBERGIA 16:3 (1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

other taxa usually referred to the Amblystegiaceae (He
den?s 1989d). Features in which Drepanocladus species
differ from those of Pseudocalliergon are: (1) The alar cells are different (usually distinctly more inflated and
more distinctly delimited in Drepanocladus than in
Pseudocalliergon) and their ontogeny is not quite simi
lar in the two genera (Heden?s 1987a); (2) The two
genera differ in the coloration of the plants. The Drepa
nocladus species have not got the golden gloss of the
Pseudocalliergon species and (with the exception of D.
sendtneri) are rarely having brownish yellow or yellow brown colours; (3) The axillary hairs are hyaline and have usually not got markedly elongate cells in Drepa
nocladus; (A) The Drepanocladus species are often
growing in nutrient-rich habitats (rich in nitrogen/phos
phorus) whereas their need for calcium seems to be less
expressed than in the Pseudocalliergon species (again with the exception of Drepanocladus sendtneri, which
seems to be almost as extreme as the Pseudocalliergon
species as regards the calcium demands).
Since I have not been able to find a reasonably prob
able sister group of the Pseudocalliergon species, any
phylogenetic consideration is very uncertain. However,
intuitively one would guess that P. angustifolium, which
is most similar to most species of e.g. Drepanocladus s.
str., Warnstorfia and Campy Hum in leaf shape, is the
more primitive of the Pseudocalliergon species treated
here, and that the broader (P. lycopodioides) and later shorter (P. brevifolium), rounded-apiculate (P. turges
cens) and at last entirely rounded (P. trifarium) leaves have evolved later, as adaptations to different habitats.
In parallel, the capsules have envolved from horizontal
and with more perfect peristomes (P. lycopodioides, P.
brevifolius) to often inclined and with a tendency to
have slightly reduced peristomes (P. turgescens and P.
trifarium). It seems less likely that the ovate leaves,
gradually narrowed to an acuminate apex, should have
evolved from broadly ovate rounded leaves, and that
the perfect peristomes should have developed from the less perfect ones, than that the opposite should have
happened. Although speculative, I do not think that it is
possible to get a more firmly based hypothesis on the
relationships between the Pseudocalliergon species at
present.
Taxonomy
Pseudocalliergon (Limpr.) Loeske
Hedwigia 46: 311. 1907. -
Hypnum sect. Pseudocal
liergon Limpr., Laubm. Deutschl. 3: 547. 1899. -
Dre
panocladus sect. Pseudocalliergon (Limpr.) Broth.,
Nat. Pfl. 1(3): 1035. 1908. - Calliergon sect. Pseudocal
liergon (Limpr.) Broth., Laubm. Fennosk. 486. 1923. -
Lectotype (nov.): Pseudocalliergon turgescens (T. Jens.) Loeske, Hedwigia 46: 311. 1907.
- Hypnum tur
LINDBERGIA 16:3 (1990)
gescens T. Jens., Vid. Medd. Naturh. For. Kj?benh. 1858 (\-A)\ 63. 1858.
Calliergon sect. Trifaria C. Jens., Bot. Not. 1921: 30.
1921. - Lectotype (nov.): Calliergon trifarium (Web. &
Mohr) Kindb., Canad. Rec. Sc. 6(2): 72. 1894. -
Hyp num trifarium Web. & Mohr, Naturh. Reise Schwedens
177. 2f 2a-d. 1804.
Drepanocladus sect. Turgidus Z. Smirn., Not. Syst.
Sect. Crypt. Inst. Bot. Komarov. Ac. Se. USSR 15: 181.
1962. - Lectotype (nov.): Drepanocladus lycopodioides
(Brid.) Warnst., Beih. Bot. Centralbl. 14: 401, 413.
1903. -
Hypnum lycopodioides Brid., Spec. Musc. 2:
227. 1812.
Dioicous. Plants pleurocarpous, medium-sized to ro
bust, sometimes turgid, green, brown-green, yellowish or yellow-brown, often with golden gloss when dry, unbranched to irregularly pinnately branched ?in one
plane. Stem leaves slightly to strongly concave, with
ovate-lanceolate to broadly ovate base, at apex broadly
rounded, suddenly narrowed to apiculate point or grad
ually to ?suddenly narrowed to shortly or longly acum
inate point, straight, and then mostly imbricate, or fal
cate, not plicate, nondecurrent or longly decurrent;
margin smooth to distinctly denticulate, plane; nerve
double and short or single and long, sometimes varying in the same species, in cross-section (when single) bi
convex or plano-convex; cells in mid-leaf linear, thin
walled and eporose or incrassate and por?se, epapil
lose, basal cells wider and sometimes shorter, more
strongly incrassate and more strongly por?se, cells near
leaf apex epapillose; alar cells inflated, slightly inflated or (rarely) hardly inflated, thin-walled or incrassate to
strongly incrassate and then often somewhat por?se,
with yellow walls when mature, in approximately trans
versely triangular group or sometimes in single trans
verse basal row or apparently undifferentiated, always
indistinctly delimited from surrounding cells, ontogeny of modified aduncus type, supra-alar cells in one to
several rows along leaf margin above alar cells, some
times apparently lacking or very indistinct, indistinctly delimited from surrounding cells; branch leaves often
slightly smaller and sometimes narrower than the stem
leaves, proximal branch leaves broad, with broadly rounded or apiculate point, sometimes acuminate.
Pseudoparaphyllia foli?se, broad. Paraphyllia lacking. Rhizoids rare, smooth, reddish brown, slightly
branched and inserted on the stem at or just below the leaf insertion. Stem with central strand (sometimes nar
row) and a usually well developed cortex of thick-walled cells. Axillary hairs with 1?2(?3) upper, early yellowish or brownish cells, well developed and usually with long apical cell, abundant. Inner perichaetial leaves straight
and erect, ovate, plicate; margin plane, not or indis
tinctly bordered at shoulder, smooth or partly dentic
ulate below, more strongly denticulate or with single
83
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

3
Fig. 1. Pseudocalliergon angustifolium (holotype, S).
A: Shoot. B: Shoot apex. C: Stem leaves. D: Branch leaves. E: Proximal branch
^ leaves. F: Pseudopara phyllium. G: Mid-leaf cells
(of different stem leaves). H: Basal cells near nerve.
I: Leaf margin (mid-leaf). J: Alar cells. K: Cross sections of stem leaves. L:
Cross-section of stem. M:
Axillary hairs. - Scales: a: A. b: B. c: C, D, E. d: F, K. e: G, H, I, J, L, M.
teeth at shoulder; nerve single or branched, ending in
mid-leaf or above; cells smooth; vaginula hairy. Perigo nial leaves from broad basal part suddenly or gradually narrowed to broadly acute or acuminate apex. Calyptra
smooth, cucullate, naked. Seta long, smooth, when dry
dextrorsely twisted below and sinistrorsely twisted
above, sometimes untwisted below or above or dextror
sely twisted above, with central strand and a cortex of
1-3 layers of incrassate cells. Capsule cylindrical,
curved, horizontal or inclined: lid conical; exothecial
84
cells quadrate to longly rectangular, thin-walled or in
crassate (especially longitudinal walls), just below mouth (1?)2-6 rows of isodiametric or transversely
rectangular cells; stomata long-pored, near base of cap
sule; annulus separating. Exostome brownish yellow, well developed; outer peristomial layer in lower part
normally cross-striolate, in upper part papillose and
weakly dentate; transverse ridges of primary peristo mial layer normally developed above; border widened
at zone of transition in outer peristomial layer pattern.
LINDBERGIA 16:3 (1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

Endostome well developed or with somewhat narrow
processes, yellowish or brownish, with high basal mem
brane, processes not or narrowly perforate, cilia 1-4,
well developed, nodose. Spores finely papillose, mature
in summer.
Chromosome numbers (known only for P. turgescens): n
= 11, 11 + lace. (Fritsch 1982).
Key to the species in northern Europe 1. Stem leaves falcate, from basal leaf portion gradually
or ? suddenly narrowed to shortly to longly acum
inate apex. 2 -
Stem leaves ?straight, from basal leaf portion sud
denly narrowed to apiculate point or leaf apex
broadly rounded. 4
2. Southern or mountainous species. Stem leaves long and longly and sometimes narrowly acuminate (Figs
1C, 3C); nerve single, 3-5(?6)-stratose and (31.5?)
38.5-90(-100) um wide near base. 3 -
Arctic species. Stem leaves comparatively short and
more shortly acuminate (Fig. 5C, D); nerve single
(sometimes branched) or double, when single 2-3
stratose and 21?53 urn wide near base.
. 3. P. brevifolium 3. Medium-sized, not turgid, mainly mountainous spe
cies (distributed in the mountains and in the North ern Boreal zone). Stem leaves narrow at base (Fig.
1C), (0.45)0.56-1.12(-1.23) mm wide at widest
part; margin partly denticulate or finely denticulate
. I. P. angustifolium -
Usually robust, somewhat turgid, southern species
(approximately reaching the Middle Boreal zone). Stem leaves broad at base (Fig. 3C), (0.68-)0.90 1.68 mm wide at widest part; margin sometimes
partly finely denticulate. 2. P. lycopodioides 4. Stem leaves apiculate.4. P. turgescens -
Stem leaves with broadly rounded apex. . 5. P. trifarium
1. Pseudocalliergon augustifolium Heden?s, sp. nov. (Fig. 1)
P. lycopodioidi (Brid.) Heden?s affinis sed debilior, foliis caulis angustioribus et marginibus folii magis valde
denticulatis.
Etymology: The specific epithet refers to the stem
leaves, which are narrower (in relation to their length), than in the other species of the genus.
Holotype: "Sweden: J?mtland, Frostviken, the valley between Mt. Gerven?kko's E and W peaks, 14 Aug 1988, Late snow-bed tow. SW. C. 980m a.s.l., L. Hede
n?s (J88-559)", in S!, isotypes in FH!, LE!.
LINDBERGIA 16:3 (1990)
Rather than raising the varietal name abbreviatus,
which may cause confusion with Drepanocladus abbre
viatus Card, et Broth., to species level, the species is
here described as new.
Synonyms
Drepanocladus sendneri f. pseudorevolvens Wint. &
Monk, in Wint., Hedwigia 49: 384: 1910. - Isotypes:
"Hypnum Sendtneri forma pseudorevolvens mit revolv
ens, Verte! [--] Central-Norwegen, Dovrefjeld, Jerkin,
im Hochmoor an der [--] nach Kongsvold, lOOOMtr.,
August 1908, leg. Dr. Winter", in herb. Winter in JE!;
"Drepanocladus Sendtneri-Wilsoni forma pseudorevol vens Wint. et. M?nkem. [-] Central-Norwegen, Dov
refjeld, Jerkin, 1000M, August 1908, leg, Dr. Winter", in herb. Winter in JE!; another isotype exists in HBG
according to Walther and Martienssen (1976).
Drepanocladus lycopodioides var. abbreviatus Monk.,
Laubm. Eur. 769. 177b. 1927. -
Lectotype (nov.):
"Hypnum lycopodioides Schwgr., fo. (var.) abbreviata
Moenkem. Finmarken, Vads?, in S?mpfen, Juli 1904,
leg. D. Winter, No 3", in HBG!.
Apparently dioicous. Plants green, yellow-brown or
brownish yellow, usually with golden gloss when dry,
medium-sized, not turgid, slightly or more strongly (but
usually irregularly) pinnately branched ?in one plane. Stem leaves slightly concave, not plicate, from erecto
patent to spreading and ovate-lanceolate to rather
broadly ovate base gradually narrowed to falcate or
? strongly falcate and longly and (often narrowly) acuminate, channelled or almost tubular apex, (0.45-)
0.56-1.12(-1.23)mm wide at widest part, not or hardly
decurrent; margin distinctly denticulate or finely dentic
ulate, often partly smooth, plane; nerve single, ending in acumen, (31.5?)38.5-70(?73.5) urn wide near base,
in cross-section plano-convex or biconvex, 3-5-stratose,
with adaxial (thin nerve) or central (thick nerve) cells
widest; mid-leaf cells (28-)31.5-87.5(-110.2) x (3.5-) 4.2?7(?8.5) urn, thin-walled and eporose or sligtly in
crassate and por?se, straight or slightly flexuose, with
longitudinal walls ? parallel except at the cell ends which are square, rounded or shortly fusiformly nar
rowed, basal leaf cells wider and usually shorter, more
strongly incrassate and por?se, cells near leaf apex
smooth; alar cells rectangular, rarely quadrate, not or
usually slightly inflated, thin-walled or incrassate and
then ?porose and with yellow walls when mature, in
approcximately transversely triangular or indistinct
group, indistinctly delimited from surrounding cells, su
pra-alar cells quadrate or shortly rectangular, in 1-7
marginal rows of cells (up to 5(-9) cells long) along leaf
margin above alar cells, indistinctly delimited from sur
rounding cells; branch leaves similar to stem leaves but
85
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

Fig. 2. Distribution of Pseudocalliergon angustifolium in north ern Europe, based on studied specimens. Open symbols repre sent inexact localities.
narrower, proximal branch leaves broad and rounded,
apiculate or acute to acuminate at apex. Pseudopa
raphyllia foli?se, broad. Rhizoids rare (seen in one
plant), smooth, reddish brown, slightly branched, in
serted at or just below the central parts of the leaf insertion. Stem in cross-section round or flattened, with
central strand and a cortex of 2-3 layers of incrassate
cells. Axillary hairs with 1?2(?3) upper, early yellowish cells, 7-12.5 urn wide, apical cell often long. Mature,
fertilized perichaetia not seen. Perigonial leaves (seen in one plant) from broad base ? suddenly or gradualy
narrowed to acuminate apex; margin smooth or almost
so, distinctly bordered at shoulder. Calyptra and spo
rophyte not seen.
Pseudocalliergon angustifolium is most similar to P. ly
copodioides, but is distinctly weaker and is not turgid, has not narrower leaves (Figs 1C and 3C, respectively) and has got more strongly denticulate leaf margins. The
differences between P. angustifolium and P. brevifolium are discussed under the latter.
Habitat: I have studied Pseudocalliergon angustifolium in the field in the mountains of northernmost J?mtland, central Sweden, during the summers 1988 and 1989. In
this area the species grows in shallow, wet depressions on lime-rich ground, often close to late snow-bed vege tation in the low-alpine region. In the warm summer
1988 some of the depressions with P. angustifolium were almost dry in the middle of August. On the other hand, in the snow-rich year 1989 a large proportion of the
occurrences of the species were still snow-covered in the
beginning of August. I do not know anything about the
86
habitat of the species in the Northern Boreal zone in the eastern part of the area studied.
Distribution: Pseudocalliergon angustifolium is wide
spread in the Scandinavian mountain range and is also
known from scattered localities in the Northern Boreal
zone in more eastern areas (Fig. 2). I have not seen
material from other parts of the world, but it seems
likely that the species occurs also further to the east in
Eurasia. It should be looked for also in northern North
America and in mountain ranges further to the south.
Studied material (excepting types): Sweden: J?mtland, Frostviken, ca. 1 km E of lake at 951 m a.s.l., SE of
Sipmeke, 880-1020 m a.s.l., 13 Aug 1988, L. Heden?s
(J88-473), CANM, G, S. J?mtland, Frostviken, NW
slope of Sipmekjeppe, 880-920 m a.s.l., 13 Aug 1988, L. Heden?s (J88-412), C, S; (J88-438), H, S; (J88 425), S; (J88-436), S; (J88-443), S. J?mtland, Frost
viken, E part of Gerven?kko's top plateau, 1000-1120
m a.s.l., 14 Aug 1988, L. Heden?s (J88-547), S. J?m
tland, Frostviken, S part of Mt. Gerven?kko, 2 Aug 1989, L. Heden?s, S. J?mtland, Frostviken, Jorm,
Brackfj?llet Jul 1926, A. H?lphers, S. ?sele Lappmark, Daunatj?kko, birch reg., 650 m a.s.l., 10 Oct 1931, T.
Arwidson, S. ?sele Lappmark, Dorotea, N. Borgaf
jallen, Genjegetjene, 10 Aug. 1922, C. Stenholm, S.
Lycksele Lappmark, T?rna, Strismasund, 10 Jul 1946, A. H?lphers, S, UME. Lycksele Lappmark, T?rna,
Stora Umevatten, Langfj?llb?cken (251), among dry Salix shrubberies, 16 Jul 1963, O. M?rtensson and G.
Wass?n, UPS. Lycksele Lappmark, Tarna, Abelvattnet,
reg. subalp., the strait betw. the lakes, 15-18 Aug 1967,
O. M?rtensson, UPS. Lule Lappmark, Jokkmokk, Ro
venjaure, 13 Jul 1937, A. H?lphers, S. Lule Lappmark,
Jokkmokk, R?vejaure, 12 Jul 1937, A. H?lphers, S. Lule Lappmark, Jokkmokk, regio Sarjekensis, Unna
Rissavare, vr., 9 Aug 1902, C. Jensen and H. W. Ar
nell, S. Lule Lappmark, G?llivare, Mt. Pierka, 750-945
m a.s.l., 23 Jul 1934, T. G. Halle, S. Torne Lappmark,
Jukkasj?rvi, Abiskojokk, in paludib. reg. betul., 17 Jul
1917, E. J?derholm, HBG, S, UPS. Torne Lappmark,
Jukkasj?rvi, ad lacum Abiskojaure, reg. betul., 8 Aug
1917, E. J?derholm, S, UPS. Torne Lappmark, Jukkas
j?rvi, Abisko Nat. P., P?t j ovare, along "j?kk" from NW to Lake Abiskojaure, reg. alp., 23 Jul 1945, H. Persson
and O. M?rtensson, S, UPS. Torne Lappmark, Jukkas
j?rvi, Tornetr?sk area, K?rsavagge, S of and close to
Lake "681", 16 Aug 1951, O. M?rtensson, UPS. Torne
Lappmark, Jukkasj?rvi, Abisko Nat. P., 1944, G. E.
DuRietz, UPS. Torne Lappmark, Jukkasj?rvi, P?sist
j?rok, reg. alp., 31 Jul 1914, E. J?derholm, UPS. Nor
way: S?r-Tr?ndelag, Dovre, Knudsh?e, 29. 7. 1879, N.
C. Kindberg, S. S?r-Tr?ndelag, Kongsvold/Dovre, n?rdl. Knuds?e, 13-1400 m a.s.l., Aug 1908, Winter,
JE. S?r-Tr?ndelag, Kongsvold, Knudsh?e, Aug 1910, Winter, JE. Finmark, Vads?, Jul 1904, Winter, No. 2,
HBG (syntype of Drepanocladus lycopodioides var. ab
LINDBERGIA 16:3 (1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions
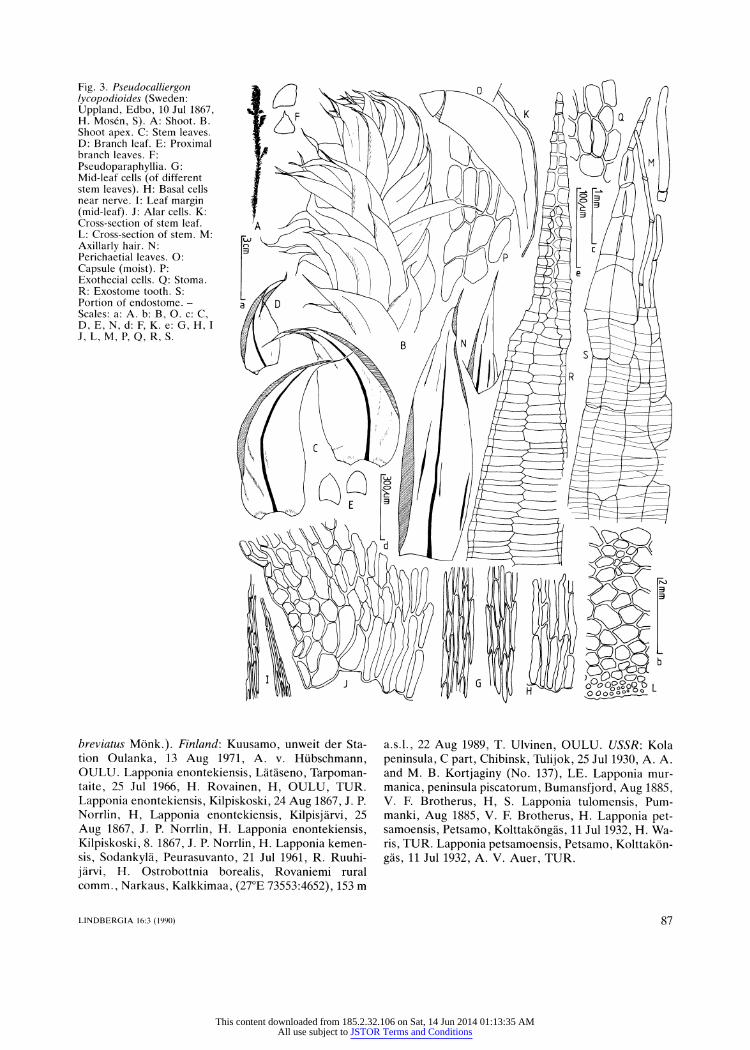
Fig. 3. Pseudocalliergon lycopodioides (Sweden:
Uppland, Edbo, 10 Jul 1867, H. Mos?n, S). A: Shoot. B. Shoot apex. C: Stem leaves. D: Branch leaf. E: Proximal branch leaves. F:
Pseudoparaphyllia. G: Mid-leaf cells (of different
stem leaves). H: Basal cells near nerve. I: Leaf margin (mid-leaf). J: Alar cells. K:
Cross-section of stem leaf. L: Cross-section of stem. M:
Axillarly hair. N:
Perichaetial leaves. O:
Capsule (moist). P:
Exothecial cells. Q: Stoma. R: Exostome tooth. S: Portion of endostome. -
Scales: a: A. b: B, O. c: C, D, E, N, d: F, K. e: G, H, I
J, L, M, P, O, R, S.
3
breviatus M?nk.). Finland: Kuusamo, unweit der Sta
tion Oulanka, 13 Aug 1971, A. v. H?bschmann, OULU. Lapponia enontekiensis, L?t?seno, Tarpoman
taite, 25 Jul 1966, H. Rovainen, H, OULU, TUR.
Lapponia enontekiensis, Kilpiskoski, 24 Aug 1867, J. P.
Norrlin, H, Lapponia enontekiensis, Kilpisj?rvi, 25
Aug 1867, J. P. Norrlin, H. Lapponia enontekiensis,
Kilpiskoski, 8. 1867, J. P. Norrlin, H. Lapponia kemen
sis, Sodankyl?, Peurasuvanto, 21 Jul 1961, R. Ruuhi
j?rvi, H. Ostrobottnia borealis, Rovaniemi rural
comm., Narkaus, Kalkkimaa, (27?E 73553:4652), 153 m
LINDBERGIA 16:3 (1990)
a.s.l., 22 Aug 1989, T. Ulvinen, OULU. USSR: Kola
peninsula, C part, Chibinsk, Tulijok, 25 Jul 1930, A. A. and M. B. Kortjaginy (No. 137), LE. Lapponia mur
manica, peninsula piscatorum, Bumansfjord, Aug 1885, V. F. Brotherus, H, S. Lapponia tulomensis, Pum
manki, Aug 1885, V. F. Brotherus, H. Lapponia pet
samoensis, Petsamo, Kolttak?ng?s, 11 Jul 1932, H. Wa
ris, TUR. Lapponia petsamoensis, Petsamo, Kolttak?n
g?s, 11 Jul 1932, A. V. Auer, TUR.
87
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

2. Pseudocalliergon lycopodioides (Brid.) Heden?s, comb. nov. (Fig. 3)
Hypnum lycopodioides Brid., Spec. Musc. 2: 227. 1812.
-Stereodon lycopodioides (Brid.) Brid., Bryol. Univ. 2: 824. 1827.
- Hypnum aduncum var. lycopodioides
(Brid.) Dub., Bot. Gall. 2: 561. 1830 (nom. illeg. incl. var. prior.
= Hypnum aduncum var. rugosum Hook. &
Tayl.; see below). -
Amblystegium lycopodioides
(Brid.) DeNot., Cronac. Briol. Ital. 2: 23. 1867. -Hyp num vernicosum var. lycopodioides (Brid.) Vent.,
Nuov. Giorn. Bot. Ital. 17: 173. 1885. -
Harpidium
lycopodioides (Brid.) Lange & C. Jens., Medd. Groen
land 3: 336. 1887. -
Drepanocladus lycopodioides
(Brid.) Warnst., Beih. Bot. Centralbl. 14: 401, 413.
1903. - Hypnum aduncum ssp. lycopodioides (Brid.)
Ren., Rev. Bryol. 33: 91. 1906. -
Scorpidium lycopo diodes (Brid.) Paul, Bryol. Zeitschr. 1: 154. 1918. -
Lectotype (Heden?s 1989c): "Hypnum rugosum Linn.,
Turner, Smith, Web. et Mohr. -
scorpioides Schultz et
alin. autor., In pratis paludosis-turfosis Megapolitanae
copi?se fructic cum Hypno trifario, 1799 zuerst von mir bei Malchin aufgenommen, Musci Frond, assioc. F. IV.
a. 193. [two rows of text which I cannot decipher] H. Stereodon lycopodioides Bryol. Univ.", in herb. Bridel
in B!.
Hypnum aduncum var. rugosum Hook. & Tayl., Muse.
Brit. 111. 1818. (Hypnum rugosum Sm., Fl. Brit. 3:
1325. 1805. (horn, illeg. - non H. rugosum Hedw.,
Spec. Muse. 923. 1801. = Rhytidium rugosum (Hedw.)
Kindb.)). -
Lectotype (nov.): "H. rugosum Fl. Br. non
Hedw. aquo omnio distinctum est. Ex. Anglia.
Hooker", in herb. Swartz (No. 2447) in S!.
Hypnum lycopodioides var. permagnum Limpr., Laubm. Deutschl. 3: 399. 1898.
- Drepanocladus lyco
podioides var. permagnus (Limpr.) Warnst., Beih. Bot.
Centralbl. 13: 415. 1903. -Lectotype (nov.): "Hypnum
lycopodioides var. permagnum Limpr., 27 Mai 1870,
Auf [uninterpretable] Begnatshauser Torfmoor. Im Sa
lem. Oberbaden. Jack. ", in herb. Jack in G!, probable
isolectotypes in H-SOL!, S!.
Drepanocladus wilsoni var. platyphyllus Roth, Hedwi
gia 48: 158, 6 f 5. 1908. - Isotype: "Drepanocladus
Wilsoni var. platyphyllus Roth. Westfalen: Moorgr?ben bei Rheine, 1908, leg. H. Brockhausen, com. Rth.", in
BM!.
Dioicous. Plants green, brown-green or yellow-brown, often with golden glass when dry, robust, somewhat
turgid, slightly or more strongly, but irregularly pin
nately branched ?in one plane. Stem leaves concave,
not plicate, from erecto-patent to almost spreading and
ovate to very broadly ovate base ? gradually narrowed
to falcate or strongly falcate and (acuminate or) longly acuminate, channeled or almost tubular apex, (0.68-)
88
0.90-1.68mm wide at widest part, not or hardly decur
rent; margin smooth or often partly finely denticulate,
plane; nerve single, ending in acumen, (37.0?)42.0
90.0(?100.0) urn wide near base, in cross-section bicon
vex or almost plano-convex, 3-5(?6)-stratose, with
adaxial (thin nerve) or central (thick nerve) cells widest; mid-leaf cells (27.0-)31.5-166 X 5.2-9 urn, thin-walled
and eporose or incrassate and por?se, straight or some
times slightly flexuose, with longitudinal walls ?parallel except at the cell ends which are fusiformly narrowed, rounded or, more rarely, square, basal leaf cells wider
and usually shorter, more strongly incrassate and po
r?se, cells near leaf apex smooth; alar cells rectangular or shortly rectangular, inflated, slightly or strongly in
crassate, somewhat por?se and with yellow walls when
mature, in approximately transversely triangular group
(rarely apparently undifferentiated), indistinctly delim ited from surrounding cells, supra-alar cells shortly rect
angular, quadrate or transversely rectangular, in one or
a few marginal rows of cells (up to ca. 8 cells long) along leaf margin above alar cells (sometimes apparently un
differentiated), indistinctly delimited from surrounding cells; branch leaves similar to stem leaves but smaller,
proximal branch leaves broad and rounded or apiculate at apex. Pseudoparaphyllia foli?se, broad. Rhizoids
rare, smooth, reddish brown, slightly branched, in
serted at or just below the leaf nerve insertion. Stem in
cross-section round or flattened, with central strand and
cortex of 2-3(?4) layers of incrassate cells. Axillary hairs with l-2(?3) upper, early yellowish cells, 8.8-14
urn wide, apical cell often long. Inner perichaetial leaves 2.44-4.40 mm long, concave, ovate and gradually
or suddenly narrowed to acuminate point, plicate; mar
gin plane, not or indistinctly bordered at shoulder, smooth or denticulate below, more coarsely denticulate
or with single teeth at shoulder; nerve single, ending below acumen; cells smooth; vaginula hairy. Perigonial leaves from broad base gradually or suddenly narrowed
to acuminate apex; margin smooth or partly denticulate
above, bordered at shoulder, sometimes indistinctly so.
Calyptra cucullate, smooth, naked. Seta reddish, long,
dextrorsely twisted below and sinistrorsely twisted
above or dextrorsely twisted throughout when dry,
smooth, with central strand and a cortex of 2-3 layers of
incrassate cells. Capsule cylindrical, curved, horizontal
or almost so; lid conical; exothecial cells 23-97 x 15.3
40.8 urn, quadrate to longly rectangular, incrassate, es
pecially longitudinal walls, just below mouth 3-5 rows of transversely rectangular or isodiametric cells; sto
mata long-pored, near base of capsule; annulus separat
ing, of 2-4 rows of cells. Exostome brownish yellow, well developed; outer peristomial layer cross-striolate
below, papillose and weakly dentate above; transverse
ridges of primary peristomial layer normally developed above; border distinctly widened at zone of transition in outer peristomial layer pattern. Endostome well devel
oped, yellowish, smooth or papillose above, smooth
below, with high basal membrane, processes not or
LINDBERGIA 16:3 (1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

Fig. 4. Distribution of Pseudocalliergon lycopodioides in north
ern Europe, based on studied specimens. Four localities in
Sweden and five in the province Karelia onegensis (USSR) are
not included on the map since I was unable to localize them.
narrowly perforate, cilia 1-3, well developed, nodose.
Spores (10.5)11.5-17.5(?19.0) urn, finely papillose, mature in summer.
Pseudocalliergon lycopodioides is the most robust of the
Pseudocalliergon species with acuminate leaves. The
most important differences between this species and P.
angustifolium and P. brevifolium are given under the
latter two species.
Habitat: Pseudocalliergon lycopodioides grows mainly
in small lime-rich wetland habitats. It is often found in
shallow depressions on flat limestone rocks (often on
alvar), on sea or lake shores or in man-made hollows
and ditches. The last kind of habitat was probably more common before modern agricultural methods were in
troduced. Most of the habitats are subject to periodic desiccation.
Distribution: Besides two finds in Iceland (Dierssen 1973; vouchers seen by me, herb. Dierssen), Pseudocal
liergon lycopodioides has a markedly southern and east
ern distribution in norther Europe (Fig. 4). In the west,
the species reaches northwards only to the Oslo area in
Norway. Further to the east, the species occurs further
to the north. Thus, in eastern Sweden P. lycopodioides
reaches the province G?strikland and in Finland it oc
curs northwards to the provinces Ostrobottnia borealis,
Lapponia kemensis and Kuusamo. The report of this
species from the Kola peninsula (USSR) by Schljakov and Konstantinova (1982) refers to P. angustifolium.
The species is widely distributed in Europe and has also been reported from the Asiatic parts of USSR
LINDBERGIA 16:3 (1990)
(Nyholm 1965, Wijk et al. 1962) and from North Amer ica (Wijk et al. 1962). However, I have not seen mate
rial from other areas than Europe and Janssens (1983)
only reports Pseudocalliergon brevifolium from North
America. The occurrences outside Europe are in need
of confirmation.
The material chosen as lectotype of Hypnum aduncum
var. rugosum Hook. & Tayl. is the only possible speci men which I have seen. Since Swartz died in 1818 (Veg
ter 1986), the specimen in his herbarium cannot have
been collected by Hooker after the description of the taxon (Hooker and Taylor 1818).
In BP, where Limprichts herbarium should be located
according to Sayre (1977), no type material of Hypnum lycopodioides var. permagnum Limpr. exists. However,
in G, where Jack's (the collector) herbarium is located
two syntypes are present. One of these is selected as
lectotype of the name.
Exiccata specimens studied (totally 475 collections
seen): E. Bauer, Musci europaei exsiccati No. 1413
1417, S. V. F. Brotherus, Bryotheca Fennica No. 189a,
S; 189b, H, S. F. Gravet, Bryotheca B?lgica No. 247, S.
Husnot, Musci Galliae No. 940A, S, Mikutowicz,
Bryotheca B?ltica No. 629 + 629a-g, S. R. Ochyra, Musci Poloniae Exsiccati No. 363, S. Rabenhorst,
Bryotheca Europaea No. 752, 752b. 914, 1200, S. Sil
l?n, Musci Frondosi Scandinaviae Exsiccati No. 160,
GB, UPS. Bryophyta D?nica Exsiccata No. 326,
OULU, S, UPS; 472 OULU. Bryotheca Polonica No.
1216, S. Cryptogamae exsuccatae editae a Museo Hist.
Natur. Vindobonensi No. 4866, S. Flora exsiccata Ba
varica: Bryophyta No. 31, S.
3. Pseudocalliergon brevifolium (Lindb.)
Heden?s, comb. nov. (Fig. 5)
Hypnum brevifolium Lindb., Oefv. K. Vet. Ak. Foerh.
23: 541. 1867. (H. brevifolium Schleich, in Brid., Spec. Muse. 2: 149. 1812 (nom. nud. in synon.
= Hetero
cladium dimorphum (Brid.) B. S. & G., cf. Bridel
(1812))). -
Harpidium brevifolium (Lindb.) Lange & C.
Jens., Medd. Groenland 3: 325. 1887. -
Amblystegium
brevifolium (Lindb.) C. Jens, in Luetken, Dijmphna Togt. Zool. Bot. Udbytte 69. 1887.
- Hypnum lycopo
dioides var. brevifolium (Lindb.) Ren. in Husn., Muse.
Gall. 395. 1894 (horn illeg., non Hypnum lycopodioides var. brevifolium Berggr., K. Svensk. Vet. Ak. Handl.
13: 83. 1875; cf. below). -
Drepanocladus brevifolius
(Lindb.) Warnst., Beih. Bot. Centralbl. 13: 401, 416.
1903. - Scorpidium brevifolium (Lindb.) Paul, Bryol.
Zeitschr. 1: 154. 1918. -Lectotype (nov.): "H. brevifo
lium Lindb., Spitsb. Lomme bay. 1861. Malmgren", in
H-SOL!.
Hypnum lycopodioides var. brevifolium Berggr., K.
89
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions
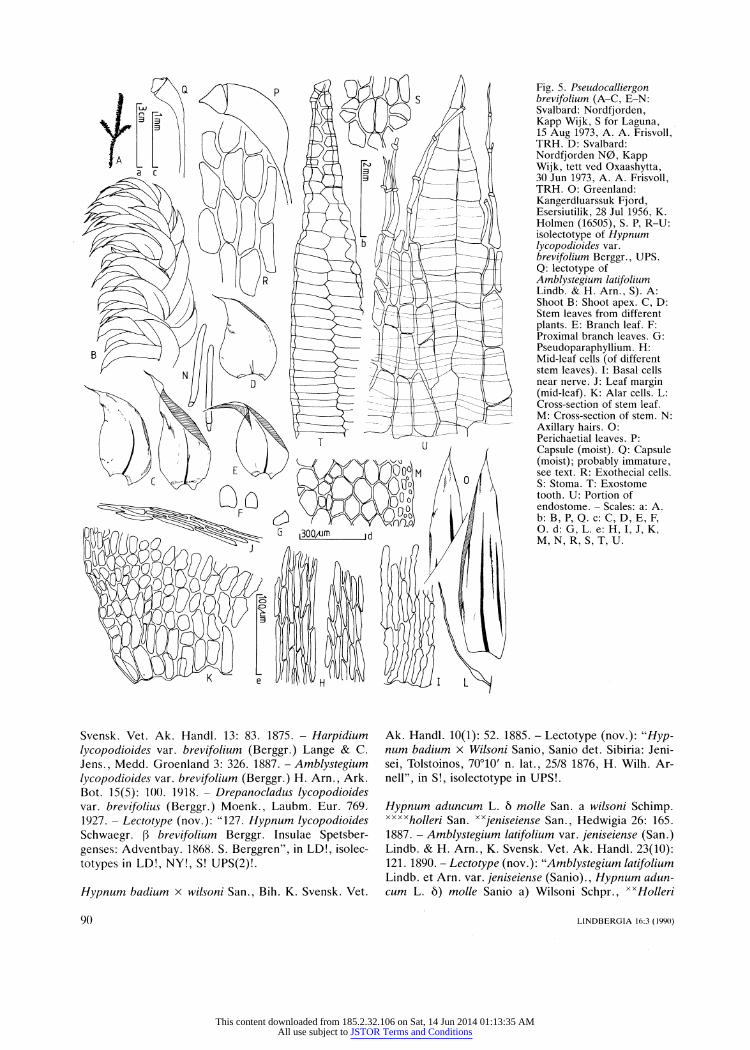
3
Fig. 5. Pseudocalliergon brevifolium (A-C, E-N: Svalbard: Nordfjorden,
Kapp Wijk, S for Laguna, 15 Aug 1973, A. A. Frisvoll,
TRH. D: Svalbard:
Nordfjorden N0, Kapp Wijk, tett ved Oxaashytta,
30 Jun 1973, A. A. Frisvoll, TRH. O: Greenland:
Kangerdluarssuk Fjord, Esersiutilik, 28 Jul 1956, K. Holmen (16505), S. P, R-U:
isolectotype of Hypnum lycopodioides var.
brevifolium Berggr., UPS.
Q: lectotype of
Amblystegium latifolium Lindb. & H. Arn., S). A: Shoot B: Shoot apex. C, D: Stem leaves from different
plants. E: Branch leaf. F:
Proximal branch leaves. G:
PseudoparaphyIlium. H: Mid-leaf cells (of different
stem leaves). I: Basal cells near nerve. J: Leaf margin (mid-leaf). K: Alar cells. L:
Cross-section of stem leaf. M: Cross-section of stem. N:
Axillary hairs. O: Perichaetial leaves. P:
Capsule (moist). Q: Capsule (moist); probably immature, see text. R: Exothecial cells. S: Stoma. T: Exostome tooth. U: Portion of endostome. - Scales: a: A. b: B, P, Q. c: C, D, E, F, O. d: G, L. e: H, I, J, K, M, N, R, S, T, U.
Svensk. Vet. Ak. Handl. 13: 83. 1875. -
Harpidium
lycopodioides var. brevifolium (Berggr.) Lange & C.
Jens., Medd. Groenland 3: 326. 1887. -
Amblystegium
lycopodioides var. brevifolium (Berggr.) H. Arn., Ark.
Bot. 15(5): 100. 1918. - Drepanocladus lycopodioides
var. brevifolius (Berggr.) Moenk., Laubm. Eur. 769,
1927. -
Lectotype (nov.): "127. Hypnum lycopodioides
Schwaegr. ? brevifolium Berggr. Insulae Spetsber
genses: Adventbay. 1868. S. Berggren", in LD!, isolec
totypes in LD!, NY!, S! UPS(2)!.
Hypnum badium x wilsoni San., Bih. K. Svensk. Vet.
90
Ak. Handl. 10(1): 52. 1885. - Lectotype (nov.): "Hyp
num badium x Wilsoni Sanio, Sanio det. Sibiria: Jeni
sei, Tolstoinos, 70?10' n. lat., 25/8 1876, H. Wilh. Ar
nell", in S!, isolectotype in UPS!.
Hypnum aduncum L. ? molle San. a wilsoni Schimp. xxxxholleri San. xxjeniseiense San., Hedwigia 26: 165.
1887. - Amblystegium latifolium var. jeniseiense (San.)
Lindb. & H. Arn., K. Svensk. Vet. Ak. Handl. 23(10): 121. 1890. - Lectotype (nov.): "Amblystegium latifolium
Lindb. et Arn. var. jeniseiense (Sanio)., Hypnum adun
cum L. ?) molle Sanio a) Wilsoni Schpr., xxHolleri
LINDBERGIA 16:3 (1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

Sanio, Sanio det., Sibiria: Jenisei, Saostrovskij ostrov,
69?40' n. lat., 10/9 1876, H. Wilh. Arnell", in UPS!,
isolectotypes in S(3)!, UPS!.
Amblystegium latifolium Lindb. & H. Arn., K. Svensk.
Vet. Ak. Handl. 23(10): 120. 1890. -
Drepanocladus
latifolius (Lindb. & H. Arn.) Warst., Beih. Bot. Cen
trabl. 13: 401, 415. 1903. - Hypnum latifolium (Lindb.
& H. Arn.) Bryhn, Rep. 2 Norw. Arct. Exp. Fram
1898-1902, 11: 132. 1902 (hom. illeg., non H. latifolium Tayl., London J. Bot. 7: 196. 1848. =
Ectropothecium
latifolium (Tayl.) Jaeg.; cf. Wijk et al. (1964)). -
Scorpi dium latifolium (Lindb. & H. Arn.) Paul, Bryol. Zeitschr. 1: 154. 1918.
- Lectotype (nov.): "Amblyste
gium latifolium Lindb. et Arn., Hypnum aduncum L. ?) molle Sanio a) Wilsoni Schpr., Sanio det., Sibiria: Jeni
sei, Tolstoinos, 70?10' n. lat., 29. 8. 1876, H. Wilh.
Arnell" in S!, isolectotypes in NY!, S!, UPS!.
Diocous. Plants green, brown-green or yellow-brown, often with golden gloss when dry, medium-sized, often
somewhat turgid, usually slightly and irregularly branched ?in one plane. Stem leaves concave, not pli
cate, from erecto-patent to patent and ovate to very
broadly ovate base gradually or ? suddenly narrowed to
falcate or strongly falcate and (shortly acuminate,) acuminate or longly acuminate, channeled or almost
tubular apex, (0.44-)0.60-1.08 mm wide at widest part, not or hardly decurrent; margin smooth or sometimes
partly very finely denticulate, plane; nerve either single and extending 60-75% way up leaf, 21-53 urn wide near
base, in cross-section biconvex, 2-3-stratose and con
sisting of homogeneous cells or with adaxial cells
widest, or nerve double and extending 30-40(-50)%
way up leaf; mid-leaf cells (17.5-)22-79(-87) x 5-10.2
(-11.5) urn, thin-walled and eporose to strongly in
crassate and por?se, straight or slightly flexuose, with
longitudinal walls ? parallel except at the cell ends which are square, rounded or, more rarely, rather
shortly fusiformly narrowed, basal leaf cells wider and
sometimes shorter, more strongly incrassate and po
r?se, cells near leaf apex smooth; alar cells rectangular or longly rectangular, inflated, slightly or strongly in
crassate, partly por?se and with yellow walls when ma
ture, in apparently transversely triangular group, indis
tinctly delimited from surrounding cells, supra-alar cells
rectangular or quadrate (more rarely transversely rect
angular), in three to several rows of cells (up to ca. 10
cells long) along leaf margin above alar cells, indis
tinctly delimited from surrounding cells; branch leaves similar to stem leaves but smaller and sometimes nar
rower, proximal branch leaves broad and rounded or
apiculate at apex. Pseudoparaphyllia foli?se, broad.
Rhizoids not seen. Stem in cross-section round or flat
tened, with central strand (sometimes narrow) and cor
tex of 1-2 layers of incrassate cells. Axillary hairs with
1?2(?3) upper, early yellowish cells, 6.5-12.7 urn wide,
apical cell often long. Inner perichaetial leaves 2.30
LINDBERGIA 16:3 (1990)
3.80 mm long, concave, ?straight and erect, ovate and
gradually to ? suddenly narrowed to acuminate or
shortly acuminate point, plicate; margin plane, unbor
dered or indistinctly bordered at shoulder, smooth or
indistinctly and irregularly denticulate, sometimes with
single teeth at shoulder; nerve single, ending in mid-leaf
or above; cells smooth, vaginula hairy. Perigonial leaves
from broad base gradually or ? suddenly narrowed to
acuminate point; margin smooth or almost so, indis
tinctly bordered or, more rarely, unbordered. Calyptra
smooth, naked (only young seen). Seta reddish, at least
near base, long, untwisted or dextrorsely twisted below
and sinistrorsely twisted above when dry, smooth, with
central strand and a cortex of 2-3 layers of incrassate
cells. Capsule (one ?mature seen) cylindrical, curved, almost horizontal; lid conical; exothecial cells 42-87.5
(?98) x 12.2-24.5 urn, rectangular, incrassate, just be
low mouth 3-5 rows of isodiametric or transversely rect
angular cells; stomata long-pored, near base of capsule; annulus separating, of 2-3 rows of cells. Exostome
brownish yellow, cross-striolate below, papillose and
weakly dentate above; transverse ridges of primary pe ristomial layer normally developed above; border dis
tinctly widened at zone of transition in outer peristomial
layer pattern. Endostome well developed, yellowish,
finely papillose above, very finely papillose below, with
high basal membrane, processes not or narrowly perfo
rate, cilia 1-4, well developed, nodose. Spores 10.5-14
urn, finely papillose, mature in summer.
Pseudocalliergon brevifolium has the most shortly acuminate leaves of the Pseudocalliergon species with
acuminate leaves. The other nothern species of the ge
nus, P. angustifolium, is easily separated from P. brevi
folium by its more longly and more narrowly acuminate
leaves (cf. Figs 1C and 5C, D), its normally single and
stronger nerve and by its more strongly denticulate leaf
margins. P. brevifolium is often distinctly turgid, this not being the case in P. angustifolium. P. lycopodioides differs from P. brevifolium, i. a., in its wider (0.90-1.68 vs 0.60-1.08 mm wide), more longly acuminate leaves
(cf. Figs 3C and 5C, D, respectively) and in its usually single and stronger nerve. There are also differences
between the three species in the length of the mid-leaf
cells, but the overlap is rather great in this character.
The sporophyte characters described by Lindberg and Arnell (1890) are probably based on poor and imma
tured sporophytes (cf. Fig. 5Q; from the lectotype of
Amblystegium latifolium). Lindberg and Arnell (1890) describe the peristome as imperfect and this agrees well
with what I found in the lectotype of Amblystegium latifolium.
Habitat: As I have not seen Pseudocalliergon brevifo lium in the field, the description of its habitat is based on Arnell and M?rtensson (1959) and Brassard (1971). The species is arctic and grows in calcium-rich habitats,
91
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

^ AH^'^? ?it
Fig. 6. Pseudocalliergon turgescens (Estonian SSR:
Kingissepa, Saarema, L?o, 13 Jun 1989, L. Heden?s
(E-21), S). A: Shoot. B: Shoot apex. C: Stem leaves.
D: Branch leaf. E: Proximal branch leaves. F:
Pseudoparaphyllia. G: Mid-leaf cells (from different stem leaves). H: Basal cells near nerve. I: Leaf margin (mid-leaf). J: Alar cells. K: Cross-section of stem. L: Axillary hairs.
M: Perichaetial leaves. N:
Capsule (moist). O: Exothecial cells. P: Stoma.
Q: Exostome tooth. R: Portion of endostome. -
Scales: a: A. b: B, N. c: C, D, E, M. d: F. e: G, H, I, J, K, L, O, P, Q, R.
in rich fens, on moist excavated soil and in percolation areas. It is often found together with P. turgescens..
Distribution: Within the studied area, Pseudocalliergon
brevifolium occurs only on Svalbard.
The species distribution is circumarctic and I have
seen material from several localities in the arctic parts of
USSR, Alaska, Canada and Greenland.
It should be noted that Hypnum lycopodioides var.
brevifolium Berggr. is not a nomenclatoric synonym of
H. brevifolium Lindb. The latter taxon is also treated in
92
the paper where Berggren described his taxon (Berg
gren 1875). In LD, where Beggrens herbarium is lo
cated, there exists an envelope with two collections
inside ("a)" and "b)") and with the joint label "Hypnum brevifolium Berggr. nova spec. S. Berggren" (no local
ity or date given on this label). The two collections both
belong to Pseudocalliergon brevifolium, but differ in
appearance and have both been distributed as "Musci
Spetsbergens. exsicc. No. 127". An exsiccate specimen with a portion of the collection which agrees best with
the protologue is here chosen as lectotype of Berggrens
(1875) name.
LINDBERGIA 16:3 (1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

Selected collections studied (totally 49 collections seen):
Svalbard: Ny-?lesund, 7 Jul 1985, J. Ekman, S. Wijde Bay, Ostfjorden, 8 Aug 1899, T. Wulff, S, UPS. Kings Bay, 1868, S. Berggren, S. Liefde Bay, 1868, S. Berg
gren, S, UPS. Prins Charles Forelands Sund, 1868, S.
Berggren, S, UPS. Nordfjorden, 1868, S. Berggren, S,
UPS. Bear Island, Mount Misery, 1868, S. Berggren,
UPS. Kings Bay Distr., the slope of Zeppelinfjeilet "556" above Ny-?lesund, 22 Jul 1956, S. Arnell and O.
M?rtensson, UPS. Kings Bay Distr., Ny-?lesund, rich
fen beside pools immed. E of Gluudneset, 25 Jul 1956, S. Arnell and O. M?rtensson, UPS. Br?ggerhalv?ya,
ved vestlige Huklagunene, 25 Jul 1974, A. A. Frisvoll, TRH. Kongsfjorden, Ny-?lesund, Storv?hut, 11 Jul
1974, A. A. Frisvoll, TRH. Nordfjorden, Kapp Wijk, S for laguna, 8 Jul 1973, A. A. Frisvoll, TRH. Back
fjorden, Trollkjeldene, 9 Aug 1974, A. A. Frisvoll, TRH. Kapp Linn?, inn for radiostasjonen, 1 Jul 1974,
A. A. Frisvoll, TRH. Kongsfj orden, Ny-?lesund, Sol
vatnet, 7 Jul 1974, A. A. Frisvoll, TRH. Br?ggerhal
v?ya, Kvadehuken inn for Huklagunene (vestligst), 25
Jul 1974, A. A. Frisvoll, TRH. Kongsfjorden, Gluud
neset, dam tett ?st for neset, 16 Jul 1974, A.A. Frisvoll,
TRH. Adventfjorden N, Advent City, strand, 23 Jul
1977, A. A. Frisvoll, TRH. Mitrahalv?ya, p? sletta
mellom Collinsodden og Willeberget, 23 Jul 1974, A.
A. Frisvoll, TRH. Reinsdyrflya, litt vest for Worsley
hamna, 24 Aug 1974, A. A. Frisvoll, TRH. Dicksonf
jorden SO, litt S for Myggdalen p? skr?ningen mot
fjorden, 14 Aug 1973, A. A. Frisvoll, TRH.
4. Pseudocalliergon turgescens (T. Jens.) Loeske
(Fig. 6)
Hedwigia 46: 311. 1907. -
Hypnum turgescens T. Jens.,
Vid. Medd. Naturh. For. Kjoebenh. 1858(1-4): 63.
1858. -
Hypnum molle ssp. turgescens (T. Jens.) C.
Hartm., Handb. Skand. Fl. ed. 9, 2: 6. 1864. -
Ster
eodon turgescens (T. Jens.) Mitt., J. Linn. Soc. Bot. 8:
42. 1865 (according to Wijk et al. 1967, but I could not
find the combination there). -
Amblystegium turgescens
(T. Jens.) Lindb., Musci Scand. 33. 1879. -
Hypnum
aduncum ssp. ? molle var. b) turgescens (T. Jens.) San.,
Bih. K. Svensk. Vet. Ak. Handl. 10(1): 38. 1885. -
Calliergon turgescens (T. Jens.) Kindb., Canad. Rec.
Sc. 6(2): 72. 1894. -
Scorpidium turgescens (T. Jens.)
Loeske, Verh. Bot. Ver. Brandenburg 46: 199. 1905. -
Hypnum molle x turgescens C. Hartm. ex Par., Ind.
Bryol. ed. 2, 3: 102. 1905 (nom. inval. err. pro. H. molle
ssp. turgescens (T. Jens.) C. Hartm.). -
Drepanocladus
turgescens (T. Jens.) Broth., Nat. Pfl. 1(3): 1035. 1908. -
Scorpidium scorpioides var. turgescens (T. Jens.)
Moenk., S?sswasserfl. 154. 1914. -
Lectotype (Heden?s
1989c): "Kongsvold, Dovre, 7/56, Th. Jensen, Hypnum
turgescens Th. Jensen in Vidensk. Medd. [etc.]", in C!,
isolectotype in H!.
LINDBERGIA 16:3 (1990)
Hypnum turgescens var. uliginosum Lindb., Oefv. K.
Vet. Ak. Foerh. 23: 539. 1867. -
Calliergon turgescens var. uliginosum (Lindb.) Warnst., Hedwigia 54: 137.
1913. -
Lectotype (nov.): "Hypnum turgescens var. uli
ginosum (19 Febr. 67), Spitsbergen, ad Treurenberg
bay, 1861, leg, Malmgren", in H-SOL!.
Hypnum turgescens var. tenue Berggr., K. Svensk. Vet.
Ak. Handl. 13(7): 91. 1875. - Amblystegium turgescens
var. tenue (Berggr.) C. Jens., Medd. Groenland 15:
433. 1898. -
Calliergon turgescens var. tenue (Berggr.)
Karcz., Monographiae Botanicae 34: 169. 1971. - Lec
totype (nov.): "151b. Hypnum turgescens Seh. ? tenue.
Beeren Eiland: Nordhamnen, 1868, S. Berggren", in
LD!, isolectotypes in FH!, H!, NY!, S(2)!, UPS!, prob able isolectotypes in LD!, NY!,S!.
f Hygrohypnum szaferi Podp. in Szafr., Starunia (Pol ska Akademja Umiejetnosci, Krakow) 1:10. 6. 1934. -
Probable holotype (slide): "Hygrohypnum szaferi Podp.", in KRA!.
Dioicous. Plants green, brown-green or yellow-brown, often with golden gloss when dry, robust, rarely
weaker, turgid, slightly and irregularly branched ?in one plane; shoot apices often falling of and propagating the species vegetatively. Stem leaves strongly concave,
imbricate or sometimes ?spreading, not plicate, ovate
or broadly ovate, upwards suddenly narrowed to short
apiculus, (0.84-)1.52-2.88 mm long, (0.67-)0.80-1.46 mm wide at widest part, not or hardly decurrent; margin smooth or partly very finely denticulate, plane; nerve
mostly double and extending 20-40% way up leaf, in scattered leaves single (then often ill developed and sometimes branched) and extending 60-65% way up leaf; mid-leaf cells (17.9-)21-107(-117) x (5.2-)6.4 11.5 urn, slightly to strongly incrassate, ?por?se,
straight or slightly flexuose, with longitudinal walls
?parallel except at the cell ends which are square,
rounded or ?shortly fusiformly narrowed, basal leaf
cells wider and often shorter, more strongly incrassate
and por?se, cells near leaf apex smooth; alar cells quad rate to rectangular, slightly inflated, incrassate or
strongly incrassate, slightly por?se and with yellow walls
when mature, in approximately transversely triangular
group, indistinctly delimited from surrounding cells, su
pra-alar cells transversely rectangular, quadrate or
shortly rectangular, sometimes very indistinctly delim
ited from both the alar cells and from the lamina cells further up, sometimes more distinct (but still indis
tinctly delimited from surrounding cells) and then in four to several rows of cells (up to ca. 15 cells long) along leaf margin above alar cells; branch leaves similar
to stem leaves or smaller, proximal branch leaves broad
and rounded or sometimes apiculate at apex. Pseudopa
raphyllia foli?se, broad, rare. Rhizoids not seen. Stem
in cross-section round or flattened, with central strand
93
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

Fig. 7.Distribution of Pseudocalliergon turgescens in northern
Europe, based on studied spedimens and one literature report
(square; Ahti and Roivainen 1968; not possible to locate in H
according to P. Isoviita in litt 1989). Open symbols represent inexact localities. One locality in the Swedish province Torne
Lappmark is not included on the map since I was unable to
localize it.
(sometimes narrow) and cortex of (1?)2-3 layers of
incrassate cells. Axillary hairs with 1?2(?3) upper,
early yellowish cells, 8.9-14 urn wide, apical cell often
long. Inner perichaetical leaves 2.20-3.80 mm long,
concave, ?straight and erect, ovate and gradually or
rather suddenly narrowed to shortly acuminate point,
plicate; margin plane, indistinctly bordered at shoulder,
smooth or denticulate below, denticulate or coarsely denticulate at shoulder; nerve single or branched, end
ing in mid-leaf or above; cells smooth; vaginula hairy.
Perigonial leaves from broad base ?suddenly narrowed
to apiculate or acuminate point; margin smooth or
slightly denticulate near apex, bordered at shoulder,
sometimes indistinctly so. Calyptra cucullate, smooth,
naked. Seta reddish, long, dextrorsely twisted below
and untwisted or sinistrorsely twisted above when dry,
smooth, with central strand and a cortex of (1?)2-3
layers of incrassate cells. Capsule cylindrical, curved,
horizontal to inclined; lid conical; exothecial cells 20.4
88.0 x (7.6-) 11.5-35.6 urn, (quadrate or) shortly to
longly rectangular, thin-walled or with longitudinal walls incrassate, just below mouth 2-6 rows of isodia
metric or transversely rectangular cells; stomata long
pored, near base of capsule; annulus separating, of 2-3
(-4) rows of cells. Exostome brownish yellow, well
developed; outer peristomial layer cross-striolate below
(in single teeth partly irregularly so or with small por tions (0?10(?25)%) irregularly dotted), finely papillose and weakly dentate above; transverse ridges of primary
peristomial layer normally developed above; border dis
94
tinctly widened at zone of transition in outer peristomial
layer pattern. Endos tome well developed but processes sometimes rather narrow, yellowish or brownish, papil lose above, smooth or finely papillose below, with high
basal membrane, processes narrowly perforate, cilia
1-4, well developed, nodose. Spores 10.2-17.0 urn,
finely papillose, mature in summer.
Pseudocalliergon turgescens is easily known by its rather
robust and usually weakly branched shoots with
straight, imbricate or sometimes slightly spreading, ovate or broadly ovate, apiculate and strongly concave
leaves. The species is sometimes propagated vegeta
tively by means of buds which fall of from the shoots (cf. Miller 1985, and references therein).
Habitat. Pseudocalliergon turgescens grows in lime-rich
areas, and mainly in two kinds of habitats. Firstly, it is
found in small wetland habitats, often in shallow de
pressions in flat limestone rocks (often on alvar in the
southern localities). Secondly, it may be found along rills or on rocks flushed with ?lime-rich water. More
rarely, it grows submerged in small lakes or pools.
Distribution. Pseudocalliergon turgescens is widely dis
tributed in the Scandinavian mountain range, from the
sea level on the Norwegian coast to the alpine region. In
addition, the species occurs in numerous localities
around the Baltic Sea and in the Swedish province V?s
terg?tland (Fig. 7). The southern localities are inter
preted as late-glacial relict occurrences by Albertson
(1940). A striking feature of the species distribution
pattern is its apparent absence from the Norwegian
province Finnmark and from the Kola peninsula in
USSR (cf. Schljakov and Konstantinova 1982), except for one locality reported by Karczmarz (1971; material
not seen by me). Outside the study area, Pseudocalliergon turgescens
occurs in scattered areas throughout the Holartic re
gion, in South America, New Guinea (Crum and An
derson 1981) and in Africa (Karczmarz 1971).
Karczmarz (1971) states that the type of Hypnum tur
gescens var. uliginosum Lindb. was collected in the
Swedish province ?land and that the holotype is in L. This must be due to some mistake since Lindberg (1867) described this taxon on material from Svalbard and
Linberg's herbarium is located in H-SOL.
According to Karczmarz (1971), the holotype o? Hyp num turgescens var. tenue Berggr. exists in LD. How
ever, in LD there is no material which with certaintly can be regarded as the holotype. One specimen from
Berggren's herbarium which could possibly be the holo
type has no indication that it is the same material as is
distributed as "Musci Spetsbergens. exsicc. No. 151b."
Thus, the name must be lectotypified on an exsiccate
specimen and the specimen in LD was chosen.
Wijk et al. (1962) state that Hygrohypnum szaferi
LINDBERGIA 16:3 (1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

Fig. 8. Pseudocalliergon trifarium (Sweden: Sm?land,
J?nk?ping, Munksj?n, Aug 1905, E. L. Ekman, UPS).
A: Shoot. B. Shoot apices. C: Stem leaves. D: Branch leaves. E: Proximal branch leaves. F:
Pseudoparaphyllia. G. Mid-leaf cells (stem leaf). H: Basal cells near nerve. I: Leaf margin (mid-leaf). J: Alar cells. K: Cross-section of stem leaf. L: Cross section of stem. M: Axillary hairs. N: Perichaetial leaves.
O: Capsules (moist). P: Exothecial cells. Q: Stoma. R: Exostome tooth. S: Portion of endostome. -
Scales: a: A. b: B, O. c: C, D, E, N. d: F, K. e: G, H, I, J, L, M, P, Q, R, S.
3
Podp. is an extant taxon. This is, however, not the case,
but the taxon is included in the present revision (despite that it is a fossil one) to clarify the situation. Since none
of the leaves on the slide in KRA can with certainly be said to be the same as the one illustrated in Szafran
(1934), I regard the KRA slide a probable holotype only.
Exsiccata speciments studied (totally 435 collections
seens): E. Bauer, Musci europaei exsiccati No. 1518?
1521, S. Husnot, Musci Galliae No. 788, S, UPS. Miku
LINDBERGIA 16:3 (1990)
towicz, Bryotheca B?ltica No. 142, 142a-c, S. R.
Ochyra, Musci Poloniae Exsiccati No. 669, S. Raben
horst, Bryoyheca Europaea No. 490, S. Bryophyta Sval
bardensia Exsiccata No. 51, S. Cryptogamae exsiccatae
editae a Museo Hist. Natur. Vindobonensi No. 2599a,
b, S. Flora Exsiccata Austro-Hungarica No. 711, S.
95
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

Fig. 9. Distribution of Pseudocalliergon trifarium in northern
Europe, based on studied material and on information from the Swedish wetland survey (small dots). Open symbols repre sent inexact localities. Two localities in Sweden (Pite Lapp
mark, Lule Lappmark) and one in USSR (Lapponia murman
ica) are not included on the map since I was unble to localize them.
5. Pseudocalliergon trifarium (Web. a Mohr) Loeske (Fig. 8)
Hedwigia 46: 301.1907. - Hypnum trifarium Web. &
Mohr, Naturh. Reise Schwedens 177. 2f 2a-d. 1804. -
Hypnum stramineum var. ? H. trifarium (Web. &
Mohr) Schwaegr., Sp. Muse. Suppl. 1(2): 212. 1816. -
Hypnum sarmentosum a trifarium (Web. & Mohr)
Wahlenb., Suppl. Fl. Lapponiae 67. 1826. -
Stereodon
trifarium (Web. & Mohr) Brid., Bryol. Univ. 2: 825. 1827. -
Amblystegium trifarium (Web. & Mohr) De
Not., Cronac. Briol. Ital. 2: 23. 1867. -
Calliergon trifa rium (Web. & Mohr) Kindb., Canad. Rec. Sc. 6(2): 72. 1894.
- Drepanocladus trifarius (Web. & Mohr) Broth,
ex Par., Coll. 10. 1909 (according to Wijk et al. 1962; not seen by me).
- Scorpidium trifarium (Web. & Mohr)
Paul, Bryol. Zeitschr. 1: 154. 1918. -
Acrocladium trifa rium (Web. & Mohr) Richs. & Wall., Trans. Brit.
Bryol. Soc. 1(4): 25. 1950. - Lectotype (Heden?s
1989c): "Hypnum trifarium Weber et Mohr. Upsaliae.
[H. Stereodon trifarius. Bryol. Univ.]" in herb. Bridel in
B!, probable isolectotype in C?.
Dioicous. Plants green, brown-green, yellowish or yel
low-brown, medium-sized (rarely almost robust),
slightly turgid, unbranced or slightly and irregularly branched ?in one plane. Stem leaves concave or
strongly concave, imbricate or ?spreading (more
rarely), not plicate, broadly or very broadly ovate with
broadly rounded apex, 1.08-2.24 mm long, 0.80-1.68
96
mm wide at widest part, longly and broadly decurrent
(often about half way down to leaf below); margin smooth or almost so, plane; nerve single, extending
60-90(-95)% way up leaf, 33-68.8 urn wide near base, in cross-section plano-convex, 2-3-stratose and consist
ing of homogeneous cells or adaxial cells widest; mid
leaf cells (30.6-)33.1-104(-107) x 5.1-10.8 urn, in
crassate and por?se or, more rarely, thin-walled and
eporose, straight or slightly flexuose, with longitudinal walls ? parallel except \at the cell ends which are
(square,) rounded or fusiformly narrowed, basal leaf
cells wider and sometimes shorter, more strongly in
crassate and por?se, cells near apex smooth; alar cells
longly rectangular, inflated, incrassate or strongly in
crassate, somewhat por?se and with yellow walls when
mature, in single transverse row or, more commonly, in
approximately transversely triangular group, indis
tinctly delimited from surrounding cells, supra-alar cells
variable, quadrate to rectangular, sometimes forming an indistinct transition from alar cells to lamina cells
further up, sometimes more distinct (but still indis
tinctly delimited from surrounding cells) and then of four to sveral rows of cells (up to ca. 10 cells long) along leaf margin above alar cells; branch leaves somewhat
smaller than the stem leaves, proximal branch leaves
broad and rounded at apex. Pseudoparaphyllia foli?se, broad. Rhizoids rare, smooth, reddish brown, slightly
branched, inserted at or just below the leaf nerve in
sertion. Stem in cross-section round or flattened, with
central strand and cortex of (1?)2-3 layers of incrassate
cells. Axillary hairs with 1-2 upper, early yellowish cells
(rarely hyaline), 8.9-12.8 urn wide, apical cell often
long. Inner perichaetial leaves 2.32-3.20 mm long, con
cave, straigth and erect, ovate, ?suddenly narrowed to
obtuse or acute apex, plicate; margin plane, unbordered
and smooth or irregularly denticulate, especially at
shoulder; nerve single, ending in upper half of leaf; cells
smooth; vaginula hairy. Perigonial leaves from broad
base gradually or, more rarely, ?suddenly narrowed to
broadly acute to acuminate apex; margin smooth or
partly indistinctly denticulate, not or indistinctly bor dered at shoulder. Calyptra cucullate, smooth, naked.
Seta reddish, long, not or dextrorsely twisted below and
sinistrorsely twisted above when dry, smooth, with cen
tral strand and a cortex of l-2(?3) layers of incrassate
cells. Capsule cylindrical, curved, horizontal to inclined; lid conical; exothecial cells 25.5-63.7 x 12.8-28 urn,
(quadrate or) shortly to longly rectangular, with longi tudinal walls somewhat incrassate, just below mouth
(1?)2-5 rows of isodiametric or transversely rectan
gular cells; stomata long-pored, near base of capsule; annulus separating of 2-4 rows of cells. Exostome brow
nish yellow, well developed (sometimes teeth somewhat
narrow); outer peristomial layer cross-striolate below,
papillose and weakly dentate above; transverse ridges of primary peristomial layer normally developed above; border narrow below, distinctly widened at zone of
transition in outer peristomial layer pattern.
LINDBERGIA 16:3 (1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

Endostome well developed but processess often rather
narrow, yellowish or brownish, papillose above, smooth
or very finely papillose below, with high basal mem
brane, processes not or narrowly perforate, cilia 1-3,
well developed, nodose. Spores 10.4-17.8 urn, finely
papillose, mature in summer.
Pseudocalliergon trifarium is easily known by its un
branched or almost unbranched shoots with straight and
imbricate or more rarely ?spreading, concave and
broadly or very broatly ovate leaves with broadly rounded apices. Sometimes the shoots of P. trifarium are possible to divide into distinct segments, which may correspond with growth seasons. In the lower part of
each segment the leaves are smaller and more strongly imbricate than in the upper portion of the same seg
ment. The transition between the differently sized
leaves is gradually within each segment, whereas the
transition between two segments is rather sudden.
Habitat: Pseudocalliergon trifarium grows in lime-rich,
deep fens. It is often found as single shoots among other
mosses (especially Scorpidium scorpioides). The species occurs from the sea level to the alpine region of the
mountains.
Distribution: Pseudocalliergon trifarium occurs through out the area (Fig. 9), but seems to avoid the coastal
districts in some areas. This may be due to a lack of
suitable habitats. P. trifarium is sometimes considered
to be a rather rare species in northern Europe, but, as
the wetland surveys in parts of northern Sweden have
shown (Fig. 9, small dots) this may be due to lack of
investigations rather than to anything else.
The species has been found in Europe, Asia and
North America. According to Crum and Anderson
(1981) it occurs in Haiti and Venezuela as well.
Exsiccata specimens studied (totally 633 collections
seen): E. Bauer, Musci europaei exsiccati No. 1526, S.
V. F. Brotherus, Bryotheca Fennica No. 83, H, S. V. F.
Brotherus, M. F. E. No. 145, H. Husnot, Musci Galliae
No. 449, 696, S. W. Migula, Kryptogamae Germaniae,
Austriae et Helvetiae exsiccatae No. 175, S. Mikutow
icz, Bryotheca B?ltica No. 144, 144a, S. R. Ochyra, Musci Poloniae Exsiccati No. 169, S. Rabenhorst,
Bryotheca Europaea No. 300, 751, S. Bryotheca Polon
ica No. 86, 896, 1644, S. Flora Exsiccata Austro-Hun
garica No. 1929, S. Flora exsiccata Bavarica: Bryophyta No. 95, S. Hepaticae et Musci URSS Exsiccati No. 50, H, S. Musci Macroregioni Meridionali Poloniae Ex
siccati No. 79, S. Westfalens Laubmoose No. 361, 361b,
S.
LINDBERGIA 16:3 (1990)
Excluded taxa
Hypnum badium Hartm., Handb. Skand. Fl. ed. 5: 332.
1849. -
Drepanocladus badius (Hartm.) Roth, Eur.
Laubm. 2: 569. 1904. -
Loeskypnum badium (Hartm.)
Paul, Bryol. Zeitschr. 1: 155. 1916. -
Lectotype (nov.): "25. Hypnum badium Hn. Lui. 1pm. Ananas. 23.6.48.
HH", in herb. Hartman in UPS!.
In herb. Hartman in UPS there are syntypes from both localities mentioned in the description of Hypnum ba
dium (Hartman 1849). The specimen chosen as lecto
type is the richer one. Hypnum badium was placed in
Hypnum sect. Pseudocalliergon by Limpricht (1899).
Hypnum illecebrum Schultz, Fl. Starg. 318. 1806."
"Hypnum illecebrum Schultz" is stated to be a synonym of Pseudocalliergon trifarium by Karzmarz (1971). However, in Schultz (1806) reference is made to Bridel
(1801: 91), where H. illecebrum Brid. is described. The latter is certainly not a synonym of P. trifarium, but of
Scleropodium tourretii (Brid.) L. Koch (Koch 1949). Thus, Karczmarz's (1971) synonymisation is not cor
rect.
Drepanocladus latinervus Warnst., Beih. Bot. Cen
tralbl. 13: 416. 1903. - Amblystegium latinerve Lindb. in Lindb. & H. Arn., K. Svensk. Vet. Ak. Handl. 23(10): 121. 1890 (nom. nud. in synon.
= A. latifolium var.
jenisseiense (San.) Lindb. & H. Arn.). -
Hypnum lati
nerve H. Arn. in Warnst., Beih. Bot. Centralbl. 13: 416.
1903 (nom. nud. in synonym. =
Drepanocladus latiner
vus Warnst.). -
Holotype: "A. latinerve n. sp., Amblys
tegium n. sp., Sibirien, Jenisei, Tolstoinos, 70?10' n.
lat., 25. 8. 1876, H. With. Arnell", in H-SOL!, isotype in S!.
The type material belongs to Drepanocladus sendtneri
or a taxon close to this species.
Amblystegium longicuspis Lindb. & H. Arn., K.
Svensk. Vet. Ak. Handl. 23(10): 123. 1890. -
Pseudo
calliergon longicuspis (Lindb. & H. Arn.) Loeske, Hed
wigia 46: 311. 1907. - Drepanocladus longicuspis
(Lindb. & H. Arn.) Broth., Nat. Pfl. 1(3): 1035. 1908. -
Scorpidium longicuspis (Lindb. & H. Arn.) Paul, Bryol. Zeitschr. 1: 154. 1918.
- Campylium longicuspis (Lindb.
& H. Arn.) Heden?s, Linbergia 14: 144. 1988 (publ. 1989).
- Lectotype (Heden?s 1988): "Amblystegium lon
gicuspis c. fr.!, Sibiria: Jenisei, Dudinka, 4/8 1876, H.
Wilh. Arnell. Ex HERBARIO S. O. LINDBERGH in S!, isolectotypes in S(3)!, H-SOL, UPS(2)!.
The taxonomic position of Amblystegium longicuspis is discussed by Heden?s (1988).
Hypnum lycopodioides var. genuinum San., Bot. Cen
97
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

tralbl. 4(Beil. 2): 24. 1880. - Amblystegium lycopo
dioides var. genuinum (San.) Lindb. & H. Arn., K.
Svensk. Vet. Ak. Handl. 23(10): 121. 1890. -
Lectotype
(Heden?s 1989c): "83. Hypnum lycopodioides Schw. a
geninum Sanio, Lyck: Rothes Bruch in einigen Torf
l?cher, 5. 5. 1873, C. Sanio", in S!.
Despite the name "genuinum", the type material be
longs to Hamatocaulis vernicosus (Mitt.) Heden?s (He den?s 1989c).
Stereodon vernicosus Mitt., J. Linn. Soc. Bot. 8: 43.
1864. -
Hypnum lycopodioides var. vernicosum (Mitt.)
San., Bot. Centralbl. 4(Beil. 2): 23. 1880. -Hamatocau
lis vernicosus (Mitt.) Heden?s, Lindbergia 15: 27. 1989
(publ. 1990). -
Lectotype (Heden?s 1989c): "Hypnum vernicosum Lindb., c? et c. fr., Sthlm, in palude ad
Carlsbergs park, 20 Julii 1858, leg. S. O. Lindberg", in H-SOL!.
This species is discussed by Heden?s (1989c).
Hypnum sendtneri var. ? wilsonii Schimp., Musci Eur.
Nov. Bryol. Eur. Suppl. fase. 3-4 (Mon.): 3. 3. 1866. -
Hypnum lycopodioides ssp. wilsonii (Schimp.) Ren. in
Husn., Muse. Gall. 375. 107 f. 4a, 5b. 1894.
This is a synonym of Drepanocladus sendtneri (Isoviita and Koponen 1981).
Hypnum lycopodioides ssp. wilsonii var. flageyi Ren. in
Husn., Muse. Gall. 395. 113 f. 3, 4. 1894. -
Lectotype
(nov.): "Hypnum Wilsoni Lindb. var. Flageyi Ren.,
Gallia = Jura: Pontarlier. leg. Flagey", in HBG (ex herb. Renauld)!, isolectotype in S!.
Since no type material seems to exist in PC (herb.
Renauld), a specimen in HBG is chosen as lectotype. The material belongs to Drepanocladus sendtneri.
Hypnum lycopodioides x fluitans San., Bih. K. Svensk.
Vet. Ak. Handl. 10(1): 50. 1885. - Amblystegium lyco
podioides x fluitans (San.) Lindb. & H. Arn., K.
Svensk. Vet. Ak. Handl. 23(10): 122. 1890. -
Lectotype
(nov.): "Hypnum lycopodioides x fluitans Sanio, Sanio
det., Sibiria: Jenisei, Fatjanova, 64?5' n. lat., 10. 7.
1876, H. Wilh. Arnell", in S!, isolectotype in S!.
The material belongs to Warnstorfia fluitans (Hedw.) Loeske.
Doubtful taxa and nomina nuda
Hypnum aduncum var. hamatum B. S. & G., Bryol.
Eur. 6: 112. 606 8. (fase. 57-61 Mon 36. 24, 2 e). 1854. -
Hypnum lycopodioides ssp. wilsonii var. hamatum (B.
S. & G.) Ren. in Husn., Muse. Gall. 370. 1894.
98
No certain type material exists. However, in BM (under the name Hypnum lycopodioides) one collection with
the label "Auf Torfmooren u Mecklenburg. Hth Schultz.
Funk. 41." exists. I regard this as a possible isotype of
the name. It belongs to Scorpidium scorpioides (Hedw.) Limpr.
Hypnum aduncum ssp. ? molle var. xxxbinerve San.,
Bih. K. Svensk. Vet. Ak. Handl. 10(1): 38. 1885.
I have not seen original material of this taxon.
Amblystegium badium x latifolium H. Arn. in Lindb. & H. Arn., K. Svensk. Vet. Ak. Handl. 23(10): 122. 1890.
(nom. nud. in synon. =
Hypnum badium x wilsonii
San.; see under Pseudocalliergon brevifolium).
Hypnum calcareum Lindb. in R. Hartm., Bryac. Scand.
Exs. n. 388. 1868 (nom. nud.). -
Hypnum turgescens var. calcareum Podp., Consp. 584. 1954 (nom. nud. in
synon. =
Pseudocalliergon turgescens fo.).
Hypnum lycopodioides var. falcatum Hobk., Syn. Brit.
Moss 169. 1873.
I have not seen original material of this taxon.
Drepanocladus lycopodioides var. gracilis Jaap. in
Warnst., Krypt. Fl. Brandenburg 2: 1027. 1906 (nom. nud. in synon.
= Pseudocalliergon lycopodioides).
Hypnum lycopodioides x aduncum var. wilsonii San.,
Hedwigia 26: 214. 1887.
I have not seen original material of this taxon.
Hypnum lycopodioides x fluitans var. exannulatus San.,
Hedwigia 26: 212. 1887.
I have not seen original material of this taxon.
Hypnum lycopodioides (lapponicum) x fluitans var. ex
annulatum San., Bih. K. Svensk. Vet. Ak. Handl. 10
(1): 50. 1885.
I have not seen original material of this taxon.
Hypnum lycopodioides x revolvens San., Bot. Cen
trale. 13: 432. 1883.
I have not seen original material of this taxon.
Hypnum spirale Schleich, in Schwaegr., Spec. Muse.
Suppl. 1(2): 212. 1816 (nom. nud. in synon. = Pseudo
calliergon trifarium).
LINDBERGIA 16:3 (1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions

Calliergon trifarium var. giganteum Warnst., Hedwigia
57: 124. 1915.
I have not seen original material of this taxon. Accord
ing to Prof. H. Hertel (in litt. 1989) no original material exists in herbarium Holler (one collector) in M.
Calliergon turgescens var. patens Karcz., Monographiae
Botanicae 34: 167. 1971. -
Holotype: "GFR, Nieder
bayern, Landau a. I., Kiesgrube bei Griepenau, umweit
Bahnhof Pilstig, 10. XII. 1914, leg. H. Paul", in W (not seen by me; according to Dr. U. Passauer (in litt. 1989) the type material cannot be found in W at present).
Hypnum uliginosum Schleich, in Web. & Mohr, Bot. Taschenb. 319. 1807 (nom. nud. in synon.
= Pseudocal
liergon trifarium).
Acknowledgements - I thank the curators of the herbaria men
tioned in this paper for large loans of material and/or invalu
able help with locating type material. W. R. Buck, R. Grolle, R. Ochyra and the staff at BM were helpful with literature
references not available in Stockholm. M. L?froth was very
helpful with some of the fieldwork and also provided informa
tion from the Swedish wetland survey. I thank T. -B. Engel mark, E. Nyholm and N. Lundqvist for valuable discussions
and help with various problems. Part of the fieldwork was
supported by grants from Karin and Axel Binnings foundation
(the Royal Swedish Academy of Sciences).
References
Ahti, T., H?met-Ahti, L. and Jalas, J. 1968. Vegetation zones
and their sections in northwestern Europe. - Ann. Bot.
Fennici 5: 169-211. - and Roivainen, H. 1968. Accessions to the Botanical Mu
seum, University of Helsinki (Helsingfors) in 1967-1968. -
Mem. Soc. Fauna Flora Fennica 44: 248-252.
Albertson, N. 1940. Scorpidium turgescens (Th. Jens.) Moen
kem. En senglacial relikt i nordisk alvarvegetation. - Acta
Phytogeogr. Suecica 13: 7-26.
Arnell, S. and M?rtensson, O. 1959. A contribution to the
knowledge of the bryophyte flora of W. Spitsbergen, and
Kongsfjorden (King's Bay, 79?N) in particular. - Arkiv
Botanik 4: 105-164.
Berggren, S. 1875. Musci et Hepaticae Spetsbergenses. - K.
Svensk. Vet. Ak. Handl. 13: 1-103.
Brassard, G. R. 1971. The mosses of northern Ellesmere Is
land, Arctic Canada. I. Ecology and phytogeography, with an analysis for the Queen Elisabeth Islands. -
Bryologist 74: 233-281.
Bridel, S. E. 1801. Muscologia Recentiorum seu analysis, his
toria et descriptio methodica. - Gothae. - 1812. Muscologiae Recentiorum seu Species Muscorum. II.
- Gothae.
Crum, H. and Andersson, L. E. 1981. Mossses of Eastern
North America, Vol. 2. - Columbia Univ. Press, New
York.
Dierssen, K. 1973. Erg?nzungen zur Moosflora Islands. - Her
zogia 3: 83-109.
Fritsch, R. 1982. Index to plant chromosome numbers - bryo
phyta. -
Regnum Vegetabile 108: I-XIV. 1-268.
Hartman, C. J. 1849. Handbok i Skandivaniens Flora, in
nefattande Sveriges och Norriges Vexter till och med Moss
orna, ed. 5. - Stockholm.
Heden?s, L. 1987a. On the ontogeny of alar cells in Drepano cladus aduncus, D. exannulatus and some other species.
- J.
Bryol. 14: 753-759.
- 1987b. On the taxonomic position of Tomentypnum Loeske. - J. Bryol. 14: 729-736.
- 1988. Amblystegium longicuspis Lindb. & H. Arn., its sta
tus and taxonomic position. -
Lindbergia 14: 142-146. - 1989a. The genus Sanionia (Musci) in Northwestern Eu
rope, a taxonomic revision. - Ann. Bot. Fennici 26: 399
419. - 1989b. Some neglected character distribution patterns
among the pleurocarpous mosses. - Bryologist 92: 157-163.
- 1989c. The genera Scorpidium and Hamatocaulis, gen. nov., in northern Europe.
- Lindbergia 15: 8-36.
- 1989d (1990). Axillary hairs in pleurocarpous mosses - a
comparative study. -
Lindbergia 15: 166-180.
Hooker, W. J. and Taylor, T. 1818. Muscologia Brittanica,
containing the mosses of Great Britain and Ireland, system
atically arranged and described. - London.
Isoviita, P. and Koponen, T. 1981. Typification of Drepanocla dus sendtneri (Musci, Amblystegiaceae).
- Ann. Bot. Fen
nici 18: 207-211.
Jannssens, J. A. 1983. Past and extant distribution of Drepano cladus in North America, with notes on the differentiation
of fossil fragments. - J. Hattori Bot. Lab. 54: 251-298.
Karczmarz, K. 1971. A monograph of the genus Calliergon
(Sull.) Kindb. - Monogr. Bot. 34: 1-209, PI. 1-20.
Koch, L. 1949. The nomenclature of Hypnum illecebrum. -
Rev. Bryol. Lieh. 18: 176-178.
Limpricht, K. G. 1899. Die Laubmoose Deutschlands, Oester
reichs und der Schweiz. Ill Abt.: 513-576. - Leipzig.
Lindberg, S. O. 1867. F?rteckning ?fver mossor, insamlade
under de svenska expeditionerna till Spitsbergen 1858 och
1861. - Oefv. K. Vet. Ak. Foerh. 23: 535-561. - and Arnell, H. W. 1890. Musci Asiae Borealis. - K.
Svensk. Vet. Ak. Handl. 23(10): 1-163.
Loeska, L. 1907. Drepanocladus, eine biologische Mischgat
tung. -
Hedwigia 46: 300-321.
Miller, N. G. 1985. Fossil evidence of the dispersal and estab
lishment of mosses as gametophyte fragments. -
Monogr. Syst. Bot. Missouri Bot. Gard. 11: 71-78.
Nyholm, E. 1965. Illustrated moss flora of Fennoscandia. II.
Musci. Fase. 5. - Lund. - 1987. Illustrated flora of Nordic mosses. Fase. 1. Fissidenta
ceae-Seligeriaceae. - Nordic Bryological Society, Copenha
gen. Paul, H. 1924. Hypnum turgescens T. Jensen. Eine system
atische-geographische Studie. - Kryptogamische Forschun
gen, M?nchen 6: 408-419.
Sayre, G. 1977. Authors of names of Bryophytes and the
present location of their herbaria. - Bryologist 80: 502-521.
Schljakov, R. N. and Konstantinova, N. A. 1982. Konspekt flori mochoobraznich Murmanskoj oblasti. -
Kolskij filial
Akademia Nauk SSSR, Apatity. Schultz, C. F. 1806. Prodromus Florae Stargardiensis.
- Bero lini.
Smirnova, Z. N. 1962. O sektsiyakh roda Drepanocladus (C.
M?ll.) Roth. - Bot. Mater. Otdela Sporovykh Rast. 15:
170-185.
Smith, A. J. E. 1978. The moss flora of Britain and Ireland. -
Cambridge Univ. Press, Cambridge. Szafran, B. 1934. Mchy dyluwjum w Staruni. - Starunia (Pol
ska Akademja Umiejetnosci, Krakow) 1: 1-17, 1 plate. Tuomikoski, R. and Koponen, T. 1979. On the generic taxo
nomy of Calliergon and Drepanocladus (Musci, Amblyste
giaceae). - Ann. Bot. Fennici 16: 213-227.
Vegter, I. H. 1986. Index Herbariorum. Part 11(6). Collectors.
S. - Regnum Vegetabile 114: 805-985.
Walther, K. and Martienssen, G. 1976. Die Laubmoostypen des Herbariums Hamburgense.
- Inst. Allg. Bot. Univ.
Hamburg, Hamburg.
Wijk, R. van der, Margadant, W. D. and Florsch?tz, P. A.
1962. Index Muscorum, Vol. II (D-Hypno). -
Regnum
vegetabile 26: 1-535. - , Margadant, W. D. and Florsch?tz, P. A. 1964. Index
Muscorum, Vol. Ill (Hypnum-O). -
Regnum Vegetabile 33: 1-529.
- , Margadant, W. D. and Florsch?tz, P. A. 1967. Index
Muscorum, Vol. IV (P-S). -
Regnum Vegetabile 48: 1-604. LINDBERGIA 16:3 (1990)
This content downloaded from 185.2.32.106 on Sat, 14 Jun 2014 01:13:35 AMAll use subject to JSTOR Terms and Conditions





























