Embed Size (px)
Citation preview

Accepted Manuscript
The evolution of photosynthesis and aerobic respiration in the cyanobacteria
Rochelle M. Soo, James Hemp, Philip Hugenholtz
PII: S0891-5849(18)32300-1
DOI: https://doi.org/10.1016/j.freeradbiomed.2019.03.029
Reference: FRB 14214
To appear in: Free Radical Biology and Medicine
Received Date: 30 October 2018
Revised Date: 5 March 2019
Accepted Date: 26 March 2019
Please cite this article as: R.M. Soo, J. Hemp, P. Hugenholtz, The evolution of photosynthesis andaerobic respiration in the cyanobacteria, Free Radical Biology and Medicine (2019), doi: https://doi.org/10.1016/j.freeradbiomed.2019.03.029.
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service toour customers we are providing this early version of the manuscript. The manuscript will undergocopyediting, typesetting, and review of the resulting proof before it is published in its final form. Pleasenote that during the production process errors may be discovered which could affect the content, and alllegal disclaimers that apply to the journal pertain.
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
x
Calvin cycle (Rubisco & PRK)Phototropy (PSI & PSII)
Loss of ETC
Complex III
bccomplexes
b6f
bc-related
bc1
ACIII
Vampirovibrionia (Melainabacteria)
Sericytochromatia (Blackallbacteria)
Obscuribacterales
SHAS531
V201-46
S15B-MN24
GL2-53
Vampirovibrio
Genus Order Class
Vampirovibrionales2-02-FULL-34-122-02-FULL-35-15
Cyanobacteriia (Oxyphotobacteria)
GCA-2770975UBA7694
UBA4093 UBA8530
Caenarcanum
Gastranaerophilales
Caenarcaniphilales
Obscuribacterales
S15B-MN24 UBA7694 (GL2-53)
Complex IV
High oxygenadapted
Low oxygenadapted
A-family oxygenreductase
C-family oxygenreductase
bd oxidase
Gloeobacterales Eurycoccales Pseudanabaenales Gloeomargaritales
Synechococcales_A
Limnotrichales Synechococcales Synechococcales_B Thermosynechococcales PCC-7407 Neosynechococcales Leptolyngbyales Psueudophormidiales Phormidesmiales
Cyanobacteriales
Gre
at O
xida
tion
Even
t (~2
.3 G
a)
1.00.4 0.5 0.6 0.7 0.8 0.90.30.2
Relative evolutionary divergence
≥90% ≥70% ≥50%

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
The evolution of photosynthesis and aerobic respiration in the
Cyanobacteria
Rochelle M. Sooa, James Hempb, Philip Hugenholtza aAustralian Centre for Ecogenomics, School of Chemistry and Molecular
Biosciences, University of Queensland, St Lucia, Queensland, Australia 5
bDepartment of Internal Medicine, University of Utah, Salt Lake City, UT, USA
*Corresponding author: Rochelle M. Soo
Highlights 10
• The recent discovery of two major non-photosynthetic bacterial lineages (classes)
specifically related to oxygenic photosynthetic Cyanobacteria has reinvigorated
debate on the evolution of oxygenic photosynthesis.
• The three cyanobacterial classes use different sets of proteins to perform aerobic 15
respiration, suggesting that this trait was independently acquired in each class.
• Independent acquisition of aerobic respiration is consistent with oxygenic
photosynthesis and the rise of oxygen occurring after divergence of the primary
cyanobacterial lines of descent, although alternative scenarios cannot be ruled out 20
based on the currently available data.
Abstract
For well over a hundred years, members of the bacterial phylum Cyanobacteria have 25
been considered strictly photosynthetic microorganisms, reflected in their
classification as “blue-green algae” in the botanical code. Recently, genomes
recovered from environmental sequencing surveys representing two major
uncultured basal lineages (classes) of Cyanobacteria have been found to completely
lack photosynthetic and CO2 fixation genes. The most likely explanation for this 30
finding is that oxygenic photosynthesis was not an ancestral feature of the
Cyanobacteria, and rather originated following divergence of the primary lines of

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
descent. Here we describe recent findings on the evolution of aerobic respiration in
the non-photosynthetic cyanobacterial classes, and how this has been interpreted by
researchers interested in the evolution of oxygenic photosynthesis. 35
Abbreviations
MAGs, metagenome assembled genomes; ETC, electron transport chain; ACIII,
Alternative complex III, RCs, Reaction centres; HGT, horizontal gene transfer; Ga,
billion years ago 40
Keywords
Cyanobacteria; oxygenic photosynthesis; aerobic respiration
1. Introduction 45
The origin of oxygenic photosynthesis was a key biological event in Earth’s history,
leading to the oxygenation of its surface environment and giving rise to complex
Eukaryotes [1]. Traditionally Cyanobacteria were thought to be strictly
photosynthetic, however this dogma was challenged when environmental 16S
ribosomal RNA gene surveys revealed at least two major basal cyanobacterial 50
lineages, 4C0d-2 and ML635J-21, in a range of aphotic habitats, including rumen [2],
drains [3], termite guts [4], human guts [5], groundwater [6], a hot-spring [7], rice
paddy soil [8] and a glacier [9].
The recent ability to extract draft genomes of individual microbial populations from 55
metagenomic datasets (so called metagenome-assembled genomes or MAGs) has
provided us with a new and exciting opportunity to examine the metabolic potential of
as-yet uncultured organisms [10], including basal cyanobacterial lineages. The first
non-photosynthetic Cyanobacteria MAG was reported in 2012 from an aphotic
anaerobic microbial community decomposing poplar wood chips. This MAG encoded 60
the ability for aerobic respiration, however it contained no genes associated with
photosynthesis. Unfortunately, the authors did not address its evolutionary
importance [11]. In 2013, Di Rienzi and colleagues reported the first MAG
representatives of 4C0d-2, which they obtained from human faeces and a
subsurface aquifer [12]. It was suggested that these MAGs represent a new 65

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
candidate phylum sibling to the Cyanobacteria, for which they proposed the name
Melainabacteria, after the Greek nymph of dark waters [12]. Metabolic inference
indicated that the Melainabacteria were markedly different to the classical
Cyanobacteria in that they entirely lack the ability to perform photosynthesis, aerobic
respiration and CO2 fixation [12]. In 2014, additional MAGs belonging to this lineage 70
were obtained from koala and human faeces, an anaerobic sludge blanket and an
enhanced biological phosphorous removal reactor [13]. Based on this analysis, it
was proposed that the Melainabacteria should be reclassified as a class within the
phylum Cyanobacteria, due to robust monophyly with photosynthetic Cyanobacteria
and shared (inferred) ancestral traits, such as cell envelope components and 75
circadian rhythm and light-response regulators [13]. The latter classification has
been recently supported by a normalised taxonomy based on genome phylogeny
[14], and four orders are currently defined in GTDB release 03-RS86 (Fig. 1),
although MAGs representing at least two additional orders have been reported [15].
The 2014 study also found that MAG representatives of the Melainabacteria lack 80
genes for photosynthesis and CO2 fixation, but identified genes for aerobic
respiration in representatives of the Obscuribacterales, specifically Complex III and
IV [13].
In 2015, the first cultured representative of the Melainabacteria was identified 85
serendipitously, Vampirovibrio chlorellavorus [16], an obligate predator of the
microalga Chlorella vulgaris [17]. V. chlorellavorus had been isolated in co-culture
with its host in 1972, but erroneously classified as a member of the
Deltaproteobacteria [18]. Many years later, its 16S rRNA gene was sequenced by
the American Type Culture Collection as part of the Living Tree Project [19], alerting 90
researchers to its true identity. Shotgun sequencing of V. chlorellavorus directly from
lyophilised cells confirmed its phylogenetic affiliation (the sole representative of the
order Vampirovibrionales, Fig. 1) and revealed a genome devoid of photosynthetic
and CO2 fixation genes, consistent with all other known members of the
Melainabacteria. However, it had a complete set of electron transport chain (ETC) 95
genes, including a terminal oxidase (Complex IV), confirming its known ability to
aerobically respire based on cultivar studies [18], [20], [21].

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
The recovery of three additional representatives of the Caenarcaniphilales in 2016
[22] identified genes encoding Complexes III and IV, leaving the Gastranaerophilales 100
as the only order lacking evidence for aerobic respiration. Recently Utami and
colleagues identified 16S rRNA genes belonging to multiple Gastranaerophilales
populations (Fig. 1) in the gut of a number of termite and cockroach species, where
they are estimated to represent up to 1.9% of the gut community [23]. A partially-
complete single-cell genome from one of the Gastranaerophilales populations was 105
obtained which lacked genes required for photosynthesis, CO2 fixation, and
respiratory metabolism. While habitat and physiology supports the inference of an
absence of these metabolic traits, it should be noted that the estimated genome
completeness was 61% meaning that the presence of these traits cannot be entirely
ruled out based on genomic information alone. 110
In 2017 [15], MAG representatives of the second basal cyanobacterial class
identified in 16S rRNA gene surveys (ML635J-21), were obtained from a coal bed
methane well [24], an algae-associated biofilm from a lab-scale bioreactor and
subsurface groundwater [25]. Comparative genome analysis confirmed their 115
affiliation with the Cyanobacteria in a lineage distinct from both classical
photosynthetic cyanobacteria and Melainabacteria, and absence of genes for
photosynthesis and CO2 fixation. As with the Melainabacteria, some members of the
group had genes for aerobic respiration that were inferred to have been acquired
well after the divergence of the primary lines of descent (Fig. 1 and see below). For 120
this reason the class Sericytochromatia was proposed, meaning “late cytochromes”
[15]. Two orders belonging to the Sericytochromatia are currently recognised in
GTDB release 03-RS86; S15B-MN24 and UBA7694 (also called GL2-53; [15]) (Fig
1). Recently, three additional MAGs belonging to a single species (>95% ANI identity
between genomes) in the order UBA7694, were reported from a deep terrestrial 125
aquifer, for which a new candidate phylum name was proposed, Blackallbacteria
[26].
Given the current taxonomic uncertainty of the Cyanobacteria (including the non-
trivial complication that they are still classified under the Botanical Code), we are 130
following the GTDB classification which uses a normalised genome-based
phylogenetic framework [14], combined with a recent proposal to formalise the rank

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
of phylum [27]. Consequently, the phylum Cyanobacteria becomes Cyanobacteriota
[28], encompassing the classes Cyanobacteriia comprising all oxygenic phototrophs
(previously called Oxyphotobacteria, [13], [15]), Vampirovibrionia after the first 135
cultured representative and replacing the Candidatus name Melainabacteria, and
Candidatus Sericytochromatia proposed by Soo et al., [15], which predates
Candidatus Blackallbacteria [26], noting however, that both names lack a
nomenclature type which will need to be assigned [29] (Fig. 1).
140
The availability of basal Cyanobacteriota provides an opportunity to re-evaluate the
origin and evolution of oxygen associated metabolisms, oxygenic photosynthesis
and aerobic respiration, in this phylum. We first address the evolution of aerobic
respiration in these lineages and then discuss the ongoing debate on the evolution of
oxygenic photosynthesis within the phylum. 145
2. Evolution of aerobic respiration in Cyanobacteriota
ETC complexes III and IV can be used to provide insights into the evolution of
aerobic respiration in the Cyanobacteriota, as genes encoding these complexes are
present in all three classes (Fig. 1, Table 1). 150
2.1. Cyanobacteriia
All members of the Cyanobacteriia share a unique cytochrome b6f complex in
addition to photosystem I, photosystem II and an A-family oxygen reductase.
Approximately 11% of genomes in this class also contain one or more bd oxidases 155
used in low oxygen habitats (Fig. 1, Table 1). Phylogenetic analyses suggest that
the cytochrome b6f complex, the A-family oxygen reductase and potentially the bd
oxidase were acquired by the ancestor of the Cyanobacteriia after diverging from
Vampirovibrionia and Sericytochromatia. Following acquisition, the b6f gene has
been predominantly vertically inherited, whereas both the A-family oxygen reductase 160
and bd oxidases appear to have been extensively, but exclusively, transferred
horizontally within the Cyanobacteriia [15]. More recently, a C-family oxygen
reductase was acquired in the genus Synechococcus (Fig. 1, [30]) suggesting that
while aerobic respiration is well established and conserved in the Cyanobacteriia,
innovations are still occurring. 165

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
2.2. Vampirovibrionia
Originally, it was thought that Vampirovibrionia were strictly fermentative as the first
set of MAGs described for this class lacked ETC genes [12]. However, subsequently
a number of Vampirovibrionia lineages were predicted to be capable of aerobic
respiration, based on the presence of near complete gene sets for Complexes III and 170
IV [15]. Of the four described Vampirovibrionia orders, three have representatives
predicted to be capable of aerobic respiration; Vampirovibrionales,
Obscuribacterales and Caenarcaniphilales (Fig. 1). It was originally thought that the
Caenarcaniphilales had lost the ability for aerobic respiration as they adapted to
anoxic environments [15] but the recovery of two additional MAGs [22] have 175
indicated that this ability has been retained in at least one member of the
Caenarcaniphilales, extracted from a well under high O2 conditions. The
Gastranaerophilales are found primarily in animal gastrointestinal tracts [12], [13]
and are inferred to gain energy strictly via fermentation. All aerobic Vampirovibrionia
have a unique fused complex III-IV operon consisting of a C-family oxygen reductase 180
(cbb3-type) and two cytochrome bc-related proteins, which are inferred to be
ancestral to known members of the class and subsequently lost in the
Gastranaerophilales and some members of the Caenarcaniphilales. The
Obscuribacterales has an additional fused complex III-IV operon consisting of a
cytochrome bc complex and a bd-like oxidase with a cytochrome c fused to the 185
periplasmic side. An unfused bd-like oxidase appears to have been independently
acquired in the Vampirovibrionales. As yet, no high oxygen-adapted reductases (A-
family oxygen reductases) have been identified in the Vampirovibrionia suggesting
that the aerobic members of the group are adapted to low-oxygen conditions [15].
However, it should be stressed that these inferences will need to be reassessed as 190
more MAG representatives are discovered, and as representatives are cultivated
and experimentally characterised, as activity of these partial and predicted
complexes has yet to be confirmed.
2.3. Sericytochromatia 195
Presently, the Sericytochromatia are represented by only seven MAGs but they
display a remarkable diversity of respiratory proteins compared to other
Cyanobacteriota, with an inferred ability to function in both high and low-oxygen
conditions (Fig. 1; [15]). Members of UBA8530 have a fused Complex III-IV operon

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
comprised of a cytochrome bc1 and a highly modified A-family oxygen reductase that 200
is missing its proton channels, suggesting it is unable to pump protons [15]. This
genus also contains a second cytochrome bc1 and a second (unmodified) A-family
oxygen reductase only distantly related to the first, and an Alternative Complex III
(ACIII) and C- family oxygen reductase. All of these respiratory genes appear to
have been acquired after UBA4093 diverged from the family UBA8530 (Fig. 1), 205
although more representatives of these lineages are needed to refine this inference.
In the other major recognised branch of the class, order UBA7694, all members have
an ACIII and an A- and C-family oxygen reductase distinct from those found in
UBA8530. The genus GCA-002770975 also has an additional phylogenetically
distinct A-family oxygen reductase. The presence of numerous complex III and IV 210
genes in the Sericytochromatia with distantly related homologues in each gene
family (Fig. 1) points to multiple independent acquisitions of aerobic respiration in
this class [15]. Given the extremely limited genomic sampling of this class to date, it
is likely that we have only glimpsed the tip of the respiratory iceberg in the
Sericytochromatia. 215
2.4. Comparison of Cyanobacteriota ETCs
Comparison of potential for aerobic respiration in the Cyanobacteriia,
Vampirovibrionia and Sericytochromatia indicate that phylogenetically distinct
respiratory genes are used in each class, often involving novel combinations, 220
particularly instances of fused Complex III and IV genes (Fig. 1). The most
parsimonious interpretation of these data is that the ancestor of the Cyanobacteriota
was not capable of aerobic respiration and that this capability was independently
acquired in all three classes on multiple occasions following, and likely permitted by,
the rise of atmospheric oxygen [15]. However, other hypotheses have been 225
presented in the literature following the genomic characterisation of Vampirovibrionia
and Sericytochromatia. These are summarised in the next section.
3. The evolution of oxygenic photosynthesis in Cyanobacteriota
There is an ongoing debate concerning the timing and mechanism of the origin of 230
oxygenic photosynthesis [31]. Some groups propose an early origin (3.0-3.8 Ga) that
potentially preceded the radiation of extant bacteria [32], [33], whereas others argue

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
that the origin directly preceded the Great Oxygenation event at 2.35 Ga [15], [30],
[34], [35]. Cyanobacteriota is the only known phylum with members capable of
oxygenic photosynthesis via coupling of type 1 and type 2 reaction centres. These 235
reaction centres are found individually in anoxygenic phototrophic members of seven
other lineages, RC1 in the Chlorobi, Firmicutes and Acidobacteria, and RC2 in the
Proteobacteria, Chloroflexi, Gemmatimonadetes and most recently in candidate
phylum WPS-2 [36], [37].
240
3.1. Current theories
There are three classes of theories concerning the evolution of oxygenic
photosynthesis in the Cyanobacteriia; the fusion, selective loss and cyanobacterial
export models [1], [31]. The fusion model first proposed in 1990 [38] suggests that
RC1 and RC2 were obtained by a non-photosynthetic cyanobacterial ancestor via 245
horizontal gene transfer (HGT) from two different anoxyphototrophic bacteria,
although there is no compelling evidence for which lineages developed the original
reaction centres [1]. The selective loss model posits that both RCs were present in a
single unknown photosynthetic ancestor and thereafter all photosynthetic lineages
inheriting those genes either vertically or horizontally, lost either RC1 or RC2 with the 250
exception of the Cyanobacteriia [39], [40], [41]. The export hypothesis was first
proposed by Mulkidjanian and colleagues in 2006 [42]. They compared 15 complete
cyanobacterial genome sequences (all members of the class Cyanobacteriia)
revealing over 1,000 protein families that were core to at least 14 of them. Only a few
components of the photosynthetic machinery were represented in the anoxygenic 255
phototrophs, suggesting that photosynthesis originated in the cyanobacterial lineage.
In this hypothesis, anoxygenic photosynthetic lineages acquired their reaction
centres via HGT from anoxygenic ancestors of the extant cyanobacteria referred to
as “procyanobacteria”. Magnabosco et al. (2018) [43] used molecular clock analyses
to explore the different scenarios using what they consider to be the three most 260
ancient phototrophic groups, the Cyanobacteriota, Chloroflexi and Chlorobi. They
concluded that the stem Cyanobacteriota are unlikely to be the recipient of an RC1
from Chlorobi but could still have been either the donor or recipient of RC2 proteins
from the Chloroflexi prior to the rise of O2. Another molecular clock analysis
concluded that phototrophy was acquired in Chloroflexi significantly after the Great 265

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Oxygenation Event and therefore the Chloroflexi could not have donated
photosynthetic genes to the ancestor of the Cyanobacteriia [44].
3.2. Recent debate in light of discovery of basal non-photosynthetic
cyanobacterial lineages 270
The recent discovery of basal non-photosynthetic Cyanobacteriota adds to the long-
standing debate on how oxygenic photosynthesis evolved in this phylum by providing
potential new time constraints. The complete absence of photosynthetic and CO2
fixation genes in the currently available Vampirovibrionia and Sericytochromatia
genomes point to at least three possible scenarios (Fig. 2). The first posits that RCs 275
were acquired by the immediate ancestor of the class Cyanobacteriia after diverging
from the other two classes (Fig. 2a). This implies that the Vampirovibrionia and
Sericytochromatia were never photosynthetic. Shih et al. [35] estimated that the
Cyanobacteriia and Vampirovibrionia diverged approximately 2.5 to 2.6 billion years
ago (Ga), before the rise of oxygen estimated at ~2.3 Ga, and that the crown group 280
Cyanobacteriia postdate the rise of oxygen, diverging ~2.0 Ga. Matheus-Carnevali et
al. [45] noted that two uncultured candidate phyla robustly affiliated with the
Cyanobacteriota, the Margulisbacteria (previously known as RBX-1 or ZB3) and
Saganbacteria (WOR-1), also completely lack photosynthetic and CO2 fixation
genes, which they suggested further supports ancestral absence of these traits in the 285
Cyanobacteriota.
By contrast, Martin et al. [46] suggest that the Cyanobacteriota ancestor was
chlorophototrophic and that the classes Vampirovibrionia and Sericytochromatia lost
their photosynthetic capability (scenario Fig. 2c) akin to the subsequent inferred loss 290
of aerobic respiration in some Vampirovibrionia lineages, notably the order
Gastranaerophilales. Thiel et al. [47] also suggested that it is equally possible that
the Vampirovibrionia and Sericytochromatia lost their photosynthetic capacity and
enzymes for CO2 fixation following divergence from the Cyanobacteriia, although this
scenario requires independent losses of these genes in the two basal classes (Fig. 295
2c). A third and intermediate possibility is that the ancestor of the Cyanobacteriia
and Vampirovibrionia acquired RCs after diverging from the Sericytochromatia
followed by a subsequent gene loss in only the Vampirovibrionia (scenario Fig. 2b).
The complete absence of RC and CO2 fixation genes in the available representatives

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
of the basal cyanobacterial classes means that we cannot presently rule out any of 300
these scenarios, or even more complex variations, highlighting the uncertainty of
these ancient events. If no photosynthetic genes or gene remnants are discovered in
the non-photosynthetic classes we may never be able to deconvolute this aspect of
the history of photosynthesis in the Cyanobacteriota. It is, however, unequivocal that
aerobic respiration evolved independently in each of the three classes presumably in 305
response to rising atmospheric oxygen (Fig. 1).
4. Conclusion
The rapid genomic elucidation of uncultured microbial lineages through high
throughput metagenomics is providing fresh perspectives on long debated topics 310
such as the origin of oxygenic photosynthesis and aerobic respiration. While the
current dataset does not definitively resolve the timing and evolutionary history of
these traits in the Cyanobacteriota, it does provide additional constraints which can
help to refine hypotheses. Additional cyanobacterial MAGs will certainly refine our
emerging picture of aerobic respiration in the Cyanobacteriota and other parts of the 315
bacterial domain, however, the origins of oxygenic photosynthesis may never be
resolved via this route, unless photosynthetic members of the basal cyanobacterial
lineages come to light.
Acknowledgements 320
We thank Maria Chuvochina for etymological advice. This work was supported by an
Australian Laureate Fellowship (FL150100038) from the Australian Research
Council.
References 325
[1] M.F. Hohmann-Marriott, R. E. Blankenship
Evolution of photosynthesis
Annu. Rev. Plant Biol., 62 (1) (2011), pp. 515-548
[2] K. Tajima, A. Shozo, K. Ogata, T. Nagamine, H. Matsui, M. Nakmura, R. I.
Aminov, Y. Benno 330
Rumen Bacterial Community Transition During Adaptation to High-grain Diet.
Anaerobe, 6 (2000), pp. 273-284

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
[3] B. Y. Smith, S. J. Turner, K. A. Rodgers
Opal-A and associated microbes from Wairakei, New Zealand: the first 300
days 335
Mineral. Mag., 67 (3) (2003), pp. 563-579
[4] F. Warnecke, P. Luginbühl, N. Ivanova, M. Ghassemian, T. H. Richardson, J.
T. Stege, M. Cayouette, A. C. McHardy, G. Djordjevic, N. Aboushadi, R.
Sorek, S. G. Tringe, M. Podar, H. G. Martin, V. Kunin, D. Dalevi, J. Madejska,
E. Kirton, D. Platt, E. Szeto, A. Salamov, K. Barry, N. Mikhailova, N. C. 340
Kyrpides, E. G. Matson, E. A. Ottesen, X. Zhang, M. Hernádes, Murillo, C., L.
G. Acosta, I, Rigoutsos, G. Tamayo, B. D. Green, C. Chang, E. M. Rubin, E.
J. Mathur, D. E. Robertson, P. Hugenholtz, J. R. Leadbetter
Metagenomic and functional analysis of hindgut microbiota of a wood-feeding
higher termite 345
Nature, 450 (7169) (2007), pp. 560-565
[5] R. E. Ley, F. Bäckhed, P. Turnbaugh, C. A. Lozupone, R. D. Knight, J. I.
Gordon
Obesity alters gut microbial ecology
PNAS, 102 (31) (2005), pp. 11070-11075 350
[6] J. M. Yagi, E. F. Neuhauser, J. A. Ripp, D. M. Mauro, E. L. Madsen
Subsurface ecosystem resilience: long-term attenuation of subsurface
contaminants supports a dynamic microbial community
ISME J., 4(1) (2010), pp. 131-143
[7] C. Takashima, A. Kano, T. Naganuma, K. Tazaki 355
Laminated Iron Texture by Iron-Oxidizing Bacteria in a Calcite Travertine
Geomicrobiol. J, 25 (3-4) (2008), pp. 193-202
[8] S. Ishii, M. Yamamoto, M. Kikuchi, K. Oshima, M. Hattori, S. Otsuka, K.
Senoo
Microbial Populations Responsive to Denitrification-Inducing Conditions in 360
Rice Paddy Soil, as Revealed by Comparative 16S rRNA Gene Analysis
Appl. Environ. Microbiol, 75 (22) (2009), pp. 7070-7078
[9] S. Pradhan, T. N. Srinivas, P. K. Pindi, K. H. Kishore, Z. Begum, P. K. Singh,
A. K. Singh, M. S. Pratibha, A. K. Ysala, G. S. Reddy, S. Shivaji
Bacterial biodiversity from Roopkund Glacier, Himalayan mountain ranges, 365
India

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Extremophiles, 14 (4) (2010), pp. 377-395
[10] D. H. Parks, C. Rinke, M. Chuvochina, P-A. Chaumeil, B. J. Woodcroft, P. N.
Evans, P. Hugenholtz, G. W. Tyson
Recovery of nearly 8,000 metagenome-assembled genomes substantially 370
revises the tree of life
Nat. Biotech., 2 (11) (2017), pp. 1533-1542
[11] D. van der Lelie, S. Taghavi, S. M. McCorkle, L-L. Li, S. A. Malfatii, D.
Monteleone, B. S. Donohoe, S-Y. Ding, W. S. Adney, M. E. Himmel, S. G.
Tringe 375
The Metagenome of an Anaerobic Microbial Community Decomposing Poplar
Wood Chips
PLoS ONE, 75 (5) (2012), e36740
[12] S. C. Di Rienzi, I. Sharon, K. C. Wrighton, O. Koren, L. A. Hug, B. C. Thomas,
J. K. Goodrich, J. T. Bell, T. D. Spector, J. F. Banfield, R. E. Ley 380
The human gut and groundwater harbour non-photosynthetic bacteria
belonging to a new candidate phylum sibling to Cyanobacteria
eLife, 2 (2013), e01102
[13] R. M. Soo, C. T. Skennerton, Y. Sekiguchi, M. Imelfort, S. J. Paech, P. G.
Dennis, J. A. Steen, D. H. Parks, G. W. Tyson, P. Hugenholtz 385
An Expanded Genomic Representation of the Phylum Cyanobacteria
Genome Biol. Evol., 6 (5) (2014), pp. 1031-1045
[14] D. H. Parks, M. Chuvochina, D. W. Waite, C. Rinke, A. Skarshewski, P-A.
Chaumeil, P. Hugenholtz
A standardized bacterial taxonomy based on genome phylogeny substantially 390
revises the tree of life
Nat. Biotech., 36 (2018), pp. 996-1004
[15] R. M. Soo, J. Hemp, D. H. Parks, W. W. Fischer, P. Hugenholtz
On the origins of oxygenic photosynthesis and aerobic respiration in
Cyanobacteria 395
Science, 355 (6332) (2017), pp. 1436-1440
[16] R. M. Soo, B. J. Woodcroft, D. H. Parks, G. W. Tyson, P. Hugenholtz
Back from the dead; the curious tale of the predatory cyanobacterium
Vampirovibrio chlorellavorus
PeerJ, 3 (2015), e968 400

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
[17] D. Coder, M. Starr
Antagonistic association of the chlorellavorus bacterium (“Bdellovibrio
chlorellavorus”) with Chlorella vulgaris
Curr. Microbiol., 1 (1978), pp. 59-64
[18] B. V. Gromov, K. A. Mamkaeva 405
Electron microscopic study of parasitism by Bdellovibrio chlorellavorus
bacteria on cells of the green alga Chlorella vulgaris
Tsitologiia, 14 (1972), pp. 256-260
[19] P. Yarza, C. Spröer, J. Swiderski, N. Mrotzek, S. Spring, B. J. Tindall, S.
Gronow, R. Pukall, H-P Klenk, E. Lang, S. Verbarg, A. Crouch, T. Lilburn, B. 410
Beck, C. Unosson, S. Cardew, E. R. B. Moore, M. Gomila, Y. Nakagawa, D.
Janssens, P. De Vos, J. Peiren, T. Suttels, D. Clermont, C. Bizet, M.
Sakamoto, T. Iida, T. Kudo, Y. Kosako, Y. Oshida, M. Ohkuma, D. R. Arahal,
E. Spieck, A. P. Roeser, M. Figge, D. Park, P. Buchanan, A. Cifuentes, R.
Munoz, J. P. Euzéby, K-H. Schleifer, W. Ludwig, R. Amann, F. O. Glöckner, 415
R. Rosselló-Móra
Sequencing orphan species initiative (SOS): Filling the gaps in the 16S rRNA
gene sequence database for all species with validly published names
Systematic and Applied Microbiol., 36 (1) (2013), pp. 69-73
[20] D. Coder, M. Starr 420
Antagonistic association of the chlorellavorus bacterium (“Bdellovibrio
chlorellavorus”)
Journ. of Phycol. 22 (1986), pp. 543-546
[21] D. M. Coder, L. J. Goff
The host range of the Chlorellavorous bacterium (“Bdellovibrio 425
chlorellavorus“) with Chlorella vulgaris
Current Microbiol., 1 (1978), pp. 59-64
[22] K. Anantharaman, C. T. Brown, L. A. Hug, I. Sharon, C. J. Castelle, A. J.
Probst, B. C. Thomas, A. Singh, M. J. Wilkins, U. Karaoz, E. L. Brodie, K. H.
Williams, S. S. Hubbard, J. F. Banfield 430
Thousands of microbial genomes shed light on interconnected
biogeochemical processes in an aquifer system
Nat. Comm., 7: 13219 (2016)

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
[23] Y. D. Utami, H. Kuwahara, T. Murakami, T. Morikawa, K. Sugaya, K. Kihara,
M. Yuki, N. Lo, P. Deevong, S. Hasin, W. Boonriam, T. Inoue, A. Yamada, M. 435
Ohkuma, Y. Hongoh
Phylogenetic Diversity and Single-Cell Genome Analysis of “Melainabacteria”,
a Non-Photosynthetic Cyanobacterial Group, in the Termite Gut
Microbes Environ., 33 (1) (2018), pp. 50-57
[24] D. An, S. M. Caffrey, J. Soh, A. Agrawal, D. Brown, K. Budwill, X. Dong, P. F. 440
Dunfield, J. Foght, L. M. Gieg, S. J. Hallam , N. W. Hanson, Z. He, T. R. Jack,
J. Klassen, K. M. Konwar, E. Kuatsjah, C. Li, S. Larter, V. Leopatra, C. L.
NesbØ, T. Oldenburg, A. P. Pagé, E. Ramos-Padron, F. F. Rochman, A.
Saidi-Mehrabad, C. W. Sensen, P. Sipahimalani, Y. C. Song, S. Wilson, G.
Wolbring, M. L. Wong, G. Voordouw 445
Metagenomics of hydrocarbon resource environments indicates aerobic taxa
and genes to be unexpectedly common
Environ. Sci. Technol., 47 (2013), pp. 10708-10717
[25] L. A. Hug, B. C. Thomas, C. T. Brown, K. R. Frischkorn, K. H. Williams, S. G.
Tringe, J. F. Banfield. L 450
Aquifer environment selects for microbial species cohorts in sediment and
groundwater
ISME J., 9 (2015), pp. 1846-1856
[26] A. J. Probst, B. Ladd, J. K. Jarett, D. E. Geller-McGrath, C. M. K. Sieber, J. B.
Emerson, K. Anantharaman, B. C. Thomas, R. R. Malmstrom, M. Stieglmeier, 455
A. Klingl, T. Woyke, M. C. Ryan, J. F. Banfield
Differential depth distribution of microbial function and putative symbionts
through sediment-hosted aquifers in the deep terrestrial subsurface
Nat. Microbiol. 3 (2018), pp. 328-336
[27] A. Oren, M. S. da Costa, G. M. Garrity, F. A. Rainey, R. Rosselló-Móra, B. 460
Schink, I. Sutcliffe, M. E. Trujilllo, W. B. Whitman
Proposal to include the rank of phylum in the International Code of
Nomenclature of Prokaryotes
Int. J. Syst. Evol. Microbiol.(2015), pp. 4284-4287
[28] W. B. Whitman, A. Oren, M. Chuvochina, M. S. da Costa, G. M. Garrity, F. A. 465
Rainey, R. Rossello-Mora, B. Schink, I. Sutcliffe, M. E. Trujillo, S. Ventura

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Proposal of the suffix –ota to denote phyla. Addendum to ‘Proposal to include
the rank of phylum in the International Code of Nomenclature of Prokaryotes’
Int. J. Syst. Evol. Microbiol. 68 (2018), pp. 967-969
[29] M. Chuvochina, C. Rinke, D. H. Parks, M. S Rappé, G. W. Tyson, P. Yilmaz, 470
W. B. Whitman, P. Hugenholtz
The importance of designating type material for uncultured taxa
Sys. Appl. Microbiol. 42 (1) (2019), pp. 15-21
[30] G. Schmetterer
The Respiratory Terminal Oxidases (RTOs) of Cyanobacteria 475
Cytochrome Complexes: Evolution, Structures, Energy Transduction, and Signalling,
(2016), pp. 331-355[31] W. W. Fischer, J. Hemp, J. E. Johnson
Evolution of Oxygenic Photosynthesis
Annu. Rev. Earth Planet. Sci., 44 (2016), pp. 647-683
[32] S. A. Crowe, L. N. Døssing, N. J. Beukes, M. Bau, S. J. Kruger, R. Frei, D. 480
E. Canfield
Atmospheric oxygenation three billion years ago
Nature, 501 (2013), pp. 535-538
[33] T. Cardona, P. Sanchez-Baracaldo, W. W. Rutherford, A. Larkum
Molecular evidence for the early evolution of photosynthetic water oxidation 485
BioRxiv, 109447
[34] L. M. Ward, J. L. Kirschvink, W. W. Fischer
Timescales of Oxygenation Following the Evolution of Oxygenic
Photosynthesis
Orig. Life Evol. Biosp., 46 (1) (2016), pp. 51-65 490
[35] P. M. Shih, J. Hemp, L. M. Ward, N. J. Matzke, W. W. Fischer
Crown group Oxyphotobacteria postdate the rise of oxygen
Geobiology, 15 (1) (2017), pp. 19-29
[36] T. Cardona
A fresh look at the evolution and diversification of photochemical reaction 495
centers
Photosyn. Res., 126 (1) (2015), pp. 111-134
[37] H. Holland-Moritz, J. Stuart, L. R. Lewis, S. Miller, M. C. Mack, S. F.
McDaniel, N. Fierer
Novel bacterial lineages associated with boreal moss species 500

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Environ. Microbiol., 20 (7) (2018), pp. 2625-2638
[38] P. Mathis
Compared structure of plant and bacterial photosynthetic reaction centers
Evolutionary implications
Biochim. Biophys. Acta, 1018 (1990), pp. 163-167 505
[39] J. M. Olson
The Evolution of Photosynthesis
Science, 168 (3930) (1970), pp. 438-446
[40] J. M. Olson, B. K. Pierson
Origin and evolution of photosynthetic reaction centres 510
Origins of Life, 17 (1987), pp. 419-430
[41] T. Cardona
Photosystem II is a Chimera of Reaction Centers
J. Mol. Evol, 84 (2017), pp. 149-151
[42] A. Y. Mulkidjanian, E. V. Koonin, K. S. Makarova, S. L. Mekhedov, A. Sorokin, 515
Y. I. Wolf, A. Dufresne, F. Partensky, H. Burd, D. Kaznadzey, R. Haselkorn,
M. Y. Galperin
The cyanobacterial genome core and the origin of photosynthesis
PNAS, 103(35) (2006), pp. 13126-13131
[43] C. Magnabosco, K. R. Moore, J. M. Wolfe, G. P. Fournier 520
Dating phototrophic microbial lineages with reticulate gene histories
Geobio., 16 (2) (2018), pp. 179-189
[44] P. M. Shih, W. M. Ward, W. W. Fischer
Evolution of the 3-hydroxypropionate bicycle and recent transfer of
anoxygenic photosynthesis into the Chloroflexi 525
PNAS, 114 (40) (2017), pp. 10749-10754
[45] P. B. Matheus-Carnevali, F. Schulz, C. J. Castelle, R. Kantor, P. Shih, I.
Sharon, J. Santini, M. Olm, Y. Amano, B. C. Thomas, K. Anantharaman, D.
Burstain, E. D. Becraft, R. Stepanauskas, T. Woyke, J. F. Banfield
Hydrogen-based metabolism as an ancestral trait in lineages sibling to the 530
Cyanobacteria
Nat. Comm. 10, 463 (2019)
[46] W. F. Martin, D. A. Bryant, J. T. Beatty
A physiological perspective on the origin and evolution of photosynthesis

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
FEMS Microbiol. Rev., 42 (2018), pp. 205-231 535
[47] V. Thiel, M. Tank, D. A. Bryant
Diversity of Chlorophototrophic Bacteria Revealed in the Omics Era
Ann. Rev. Plant Biol., 69 (2018), pp. 21-49
[48] M. N. Price, P. S. Dehal, A. P. Arkin
FastTree 2 – Approximately Maximum-Likelihood Trees for Large Alignments 540
PLoS ONE, 5 (3) (2010), e9490
545
550
555
560

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
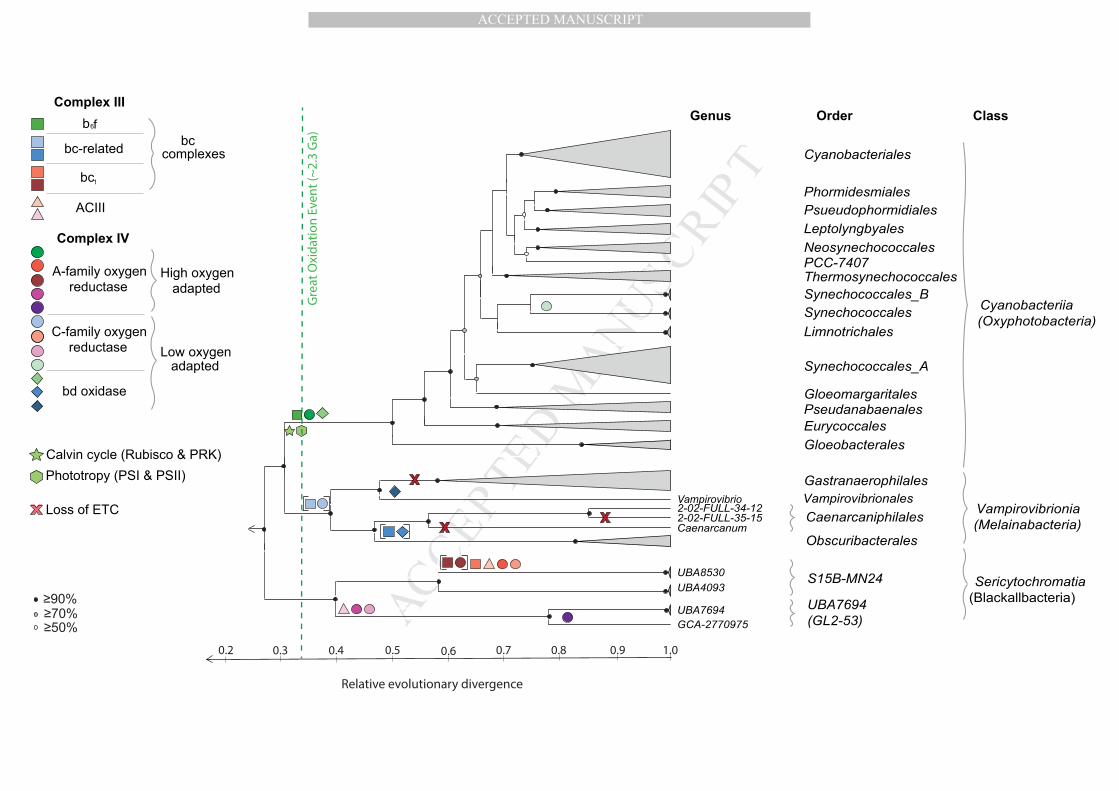
Fig. 1. Evolution of oxygenic photosynthesis and aerobic respiration in the
Cyanobacteriota. A cladogram of the phylum Cyanobacteriota taken from a larger
tree of the bacterial domain based on phylogenetic inference of 120 concatenated
single copy marker proteins scaled according to relative evolutionary divergence 565
(shown at base of figure; 0 = root of bacterial tree, 1 = extant taxa) [15]. Bootstrap
resampling analyses (100 times) with maximum likelihood was performed with
FastTree [48]. Black circles represent interior nodes with ≥90% bootstrap support,
grey circles ≥70% bootstrap support and white circles ≥50% bootstrap support.
Current proposed nomenclature based on GTDB [14] is shown to the right of the 570
cladogram. Previously proposed names [12], [13], [26] for the same groups are
shown in parentheses. Genes for Complexes III & IV, Calvin cycle and phototrophy
are distinguished by shape according to the legend at left. Different colours indicate
phylogenetically distinct versions of a given complex. Square brackets denote
operon fusions, and a red “X” indicates putative loss of ETCs. PRK, 575
phosphoribulokinase. In this representation, we infer that the Cyanobacteriota were
ancestrally nonphototrophic and that the class Cyanobacteriia acquired the ability for
photosynthesis after they diverged from the Vampirovibrionia. However, alternative
scenarios have also been proposed (see Fig. 2). The three Cyanobacteriota classes
likely acquired aerobic respiration independently after the rise of oxygen 580
(atmospheric oxygen is represented by a dashed green line). Figure is adapted from
Soo et al., 2017 [15].

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Fig. 2. Three possible scenarios for the evolution of oxygenic photosynthesis
in Cyanobacteriota. A. Acquisition of RCs by the Cyanobacteriia after primary 585
divergence of classes [15, 35]. B. Acquisition of RCs by the ancestor of the
Cyanobacteriia and Vampirovibrionia, with subsequent loss of RCs in the
Vampirovibrionia [46]. C. Acquisition of RCs prior to divergence of all three classes,
with subsequent loss of RCs in the Vampirovibrionia and Sericytochromatia. Green
arrows represent acquisition events and red arrows represent loss [46, 47]. 590
595
600
Vampirovibrionia
Cyanobacteriia
Sericytochromatia
A. Acquisition of RCs
B. Acquisition and loss of RCs
Vampirovibrionia
Cyanobacteriia
Sericytochromatia
C. Loss of RCs
Vampirovibrionia
Cyanobacteriia
Sericytochromatia
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
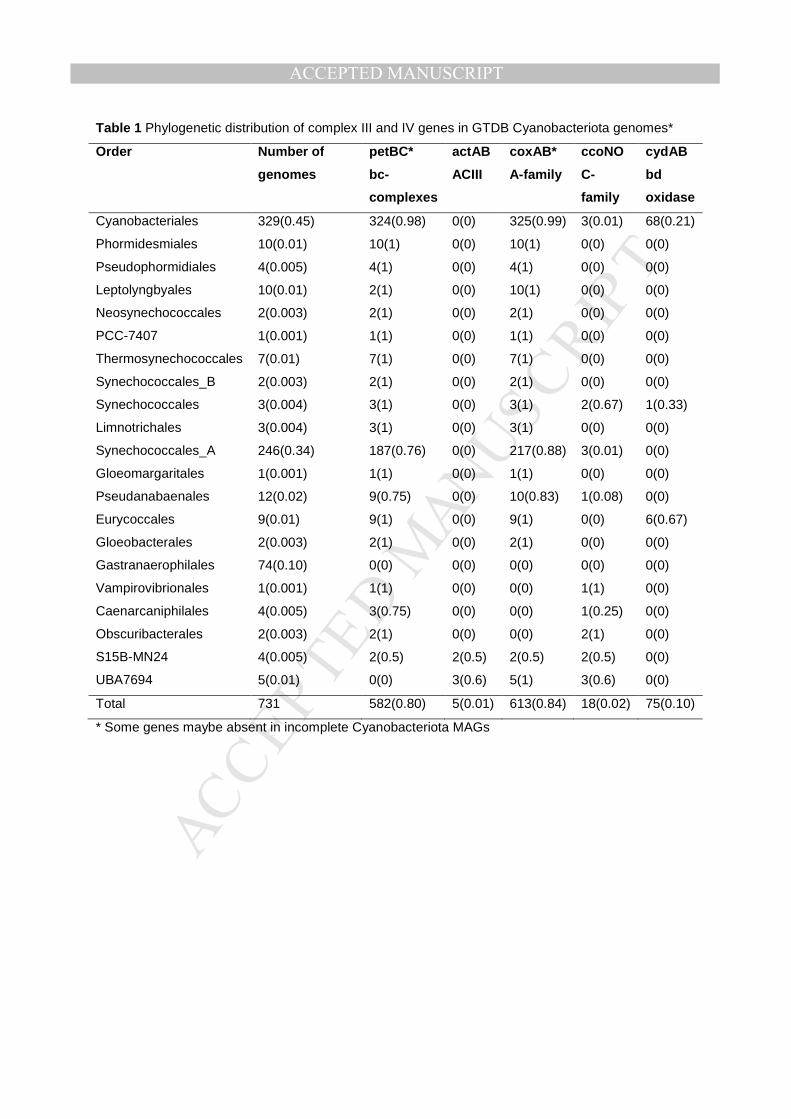
Table 1 Phylogenetic distribution of complex III and IV genes in GTDB Cyanobacteriota genomes*
Order Number of
genomes
petBC* actAB coxAB* ccoNO cydAB
bc-
complexes
ACIII A-family C-
family
bd
oxidase
Cyanobacteriales 329(0.45) 324(0.98) 0(0) 325(0.99) 3(0.01) 68(0.21)
Phormidesmiales 10(0.01) 10(1) 0(0) 10(1) 0(0) 0(0)
Pseudophormidiales 4(0.005) 4(1) 0(0) 4(1) 0(0) 0(0)
Leptolyngbyales 10(0.01) 2(1) 0(0) 10(1) 0(0) 0(0)
Neosynechococcales 2(0.003) 2(1) 0(0) 2(1) 0(0) 0(0)
PCC-7407 1(0.001) 1(1) 0(0) 1(1) 0(0) 0(0)
Thermosynechococcales 7(0.01) 7(1) 0(0) 7(1) 0(0) 0(0)
Synechococcales_B 2(0.003) 2(1) 0(0) 2(1) 0(0) 0(0)
Synechococcales 3(0.004) 3(1) 0(0) 3(1) 2(0.67) 1(0.33)
Limnotrichales 3(0.004) 3(1) 0(0) 3(1) 0(0) 0(0)
Synechococcales_A 246(0.34) 187(0.76) 0(0) 217(0.88) 3(0.01) 0(0)
Gloeomargaritales 1(0.001) 1(1) 0(0) 1(1) 0(0) 0(0)
Pseudanabaenales 12(0.02) 9(0.75) 0(0) 10(0.83) 1(0.08) 0(0)
Eurycoccales 9(0.01) 9(1) 0(0) 9(1) 0(0) 6(0.67)
Gloeobacterales 2(0.003) 2(1) 0(0) 2(1) 0(0) 0(0)
Gastranaerophilales 74(0.10) 0(0) 0(0) 0(0) 0(0) 0(0)
Vampirovibrionales 1(0.001) 1(1) 0(0) 0(0) 1(1) 0(0)
Caenarcaniphilales 4(0.005) 3(0.75) 0(0) 0(0) 1(0.25) 0(0)
Obscuribacterales 2(0.003) 2(1) 0(0) 0(0) 2(1) 0(0)
S15B-MN24 4(0.005) 2(0.5) 2(0.5) 2(0.5) 2(0.5) 0(0)
UBA7694 5(0.01) 0(0) 3(0.6) 5(1) 3(0.6) 0(0)
Total 731 582(0.80) 5(0.01) 613(0.84) 18(0.02) 75(0.10)
* Some genes maybe absent in incomplete Cyanobacteriota MAGs





























