Embed Size (px)
Citation preview

The emergence and importance of compartment models
Eörs Szathmáry
Collegium Budapest AND Eötvös University

Early origin-of-life models were compartmental
• Oparin and coacervates• Fox and microspheres• The idea of naked replicators became
fashionable after the discovery of DNA structure
• Even though DNA never self-replicates!• Standard forms of the RNA world today
usually assume naked replicators

A lot of confusion arises when units of evolution and units of life are
taken to be identical
• The problem of the virus• Gánti’s analogy: the virus is the living cells
as a self-replicating programme is to the computer
• Neither the virus nor the programme do not do anything alone
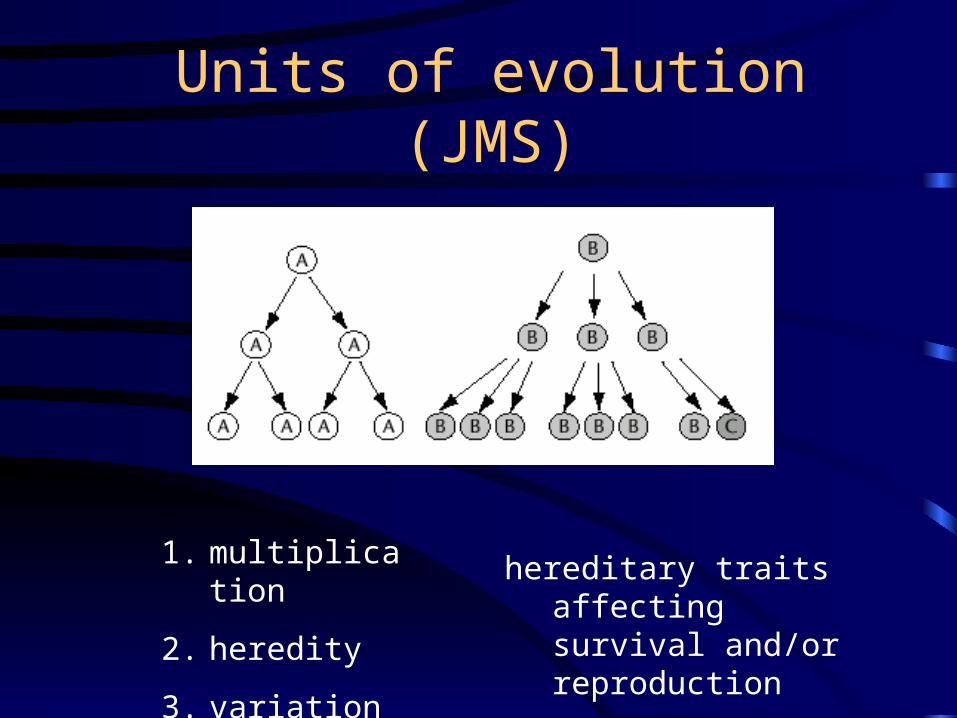
Units of evolution (JMS)
hereditary traits affecting survival and/or reproduction
1. multiplication
2. heredity
3. variation

The simplest cells today are bacterial
• THUS we want to explain the origin of some primitive bacterium-like cell
• Even present-day bacteria are far too complex• The main problem is the genetic code
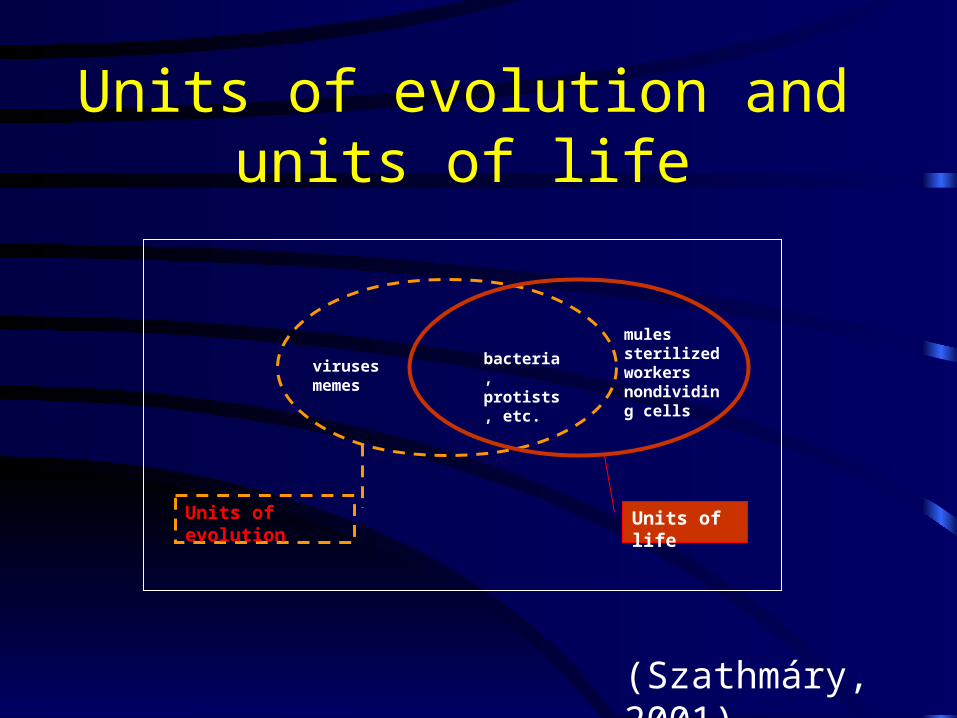
Units of evolution
Units of evolution and units of life
virusesmemes
mulessterilized workersnondividing cells
bacteria,protists, etc.
Units of life
(Szathmáry, 2001)

We need a modern picture of compartmentation
• Molecular biology
• Cell biology
• Biochemistry

Gánti’s chemoton model (1974)
ALL THREE SUBSYSTEMS ARE AUTOCATALYTIC
template copying
metabolism
membrane growth

The fission of the chemoton
• Membrane surface doubled• Quantity of internal materials doubled• Assume spherical shape• Concentration cannot be kept with a
growing sphere: volume increases with the cubic of the radius
• Volume of sphere with a surface are doubled would be more than doubled

Compartments are important, because
• They prohibit constituents diffusing away• Can increase local concentration• Create a special microenvironment, e.g. by
keeping many molecules out• For example, imagine when a poison cannot
enter the compartment• The problem of the origin of life is
essentially that of metabolite channelling!• Last but not least: a higher-level unit of
evolution

Mineral surfaces are a poor man’s form of compartmentation (?)
• A passive form of localisation (limited diffusion in 2D)
• Thermodynamic effect (when leaving group also leaves the surface)
• Kinetic effects: surface catalysis (cf. enzymes)• How general and diverse are these effects?• Good for polymerisation, not good for metabolism
(Orgel)• What about catalysis by the inner surface of the
bilayer (composomes)?

Population dynamics on surfaces
• Reaction-diffusion on the surface (following Hogeweg and Boerlijst, 1991)
• One tends to interact with one’s neighbours
• This is important, because lesson from theoretical ecology indicates that such conditions promote coexistence of competitors
• Important effect on the dynamics of the primordial genome (cf. Eigen’s paradox)
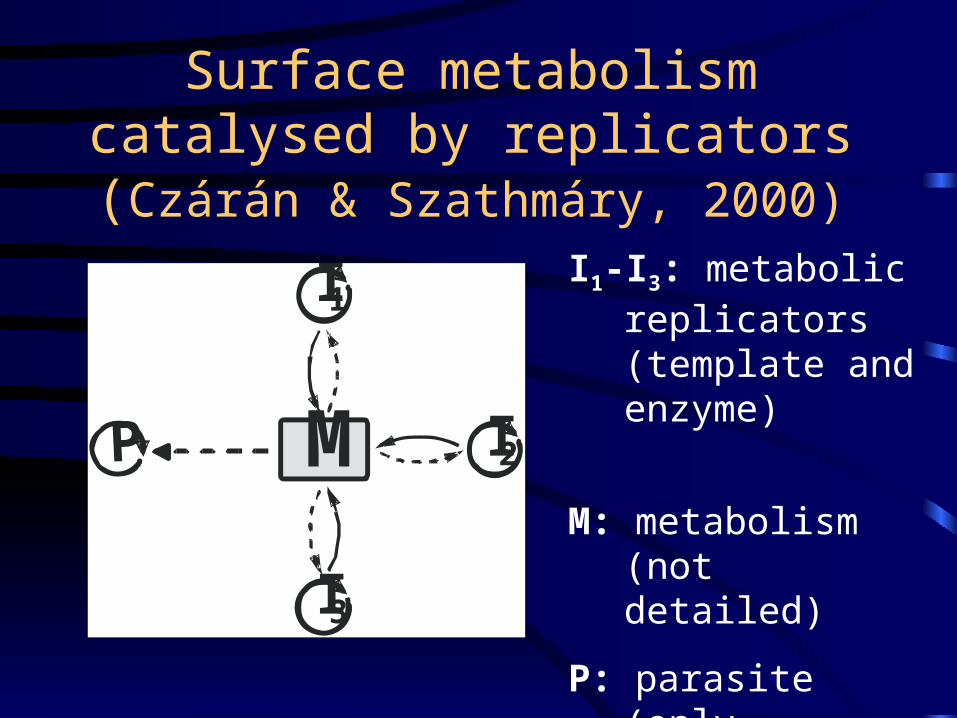
Surface metabolism catalysed by replicators (Czárán & Szathmáry, 2000)
I1-I3: metabolic replicators(template and enzyme)
M: metabolism (not detailed)
P: parasite (only template)
M
I1
I2
I3
P

Elements of the model• A cellular automaton model
simulating replication and dispersal in 2D
• ALL genes must be present in a limited METABOLIC neighbourhood for replication to occur
• Replication needs a template next door
• Replication probability proportional to rate constant (allowing for replication)
• Diffusion
X
i - 2 i - 1 i i + 1 i + 2
j -2
j - 1
j
j + 1
j + 2
S

Robust conclusions• Protected polymorphism of competitive
replicators (cost of commonness and advantage of rarity)
• This does NOT depend on mesoscopic structures (such as spirals, etc.)
• Parasites cannot drive the system to extinction• Unless the neighbourhood is too large (approaches
a well-stirred system)• Parasites can evolve into metabolic replicators• System survives perturbation (e.g. when death
rates are different in adjacent cells), exactly because no mesocopic structure is needed.

An interesting twist
• This system survives with arbitrary diffusion rates
• But metabolic neighbourhood size must remain small
• Why does excessive dispersal not ruin the system?
• Because it convergences to a trait-group model!

The trait group model (Wilson, 1980)
Random dispersal
Harvest
Applied to early coexistence: Szathmáry (1992)
Mixed global pool
Mixed global pool

Why does the trait group work?• It works only for cases when the “red hair
theorem” applies
• People with red hair overestimate the frequency of people with red hair, essentially because they know this about themselves
• “average subjective frequency”
• In short, molecules must be able to scratch their own back!

Nature 420, 360-363 (2002).
• 2D Reaction-diffusion system: molecules are bound to the surface, and can only interact with their neighbours
• Replicating molecules have template and replicase functions (auto- and heterocatalytic effects)
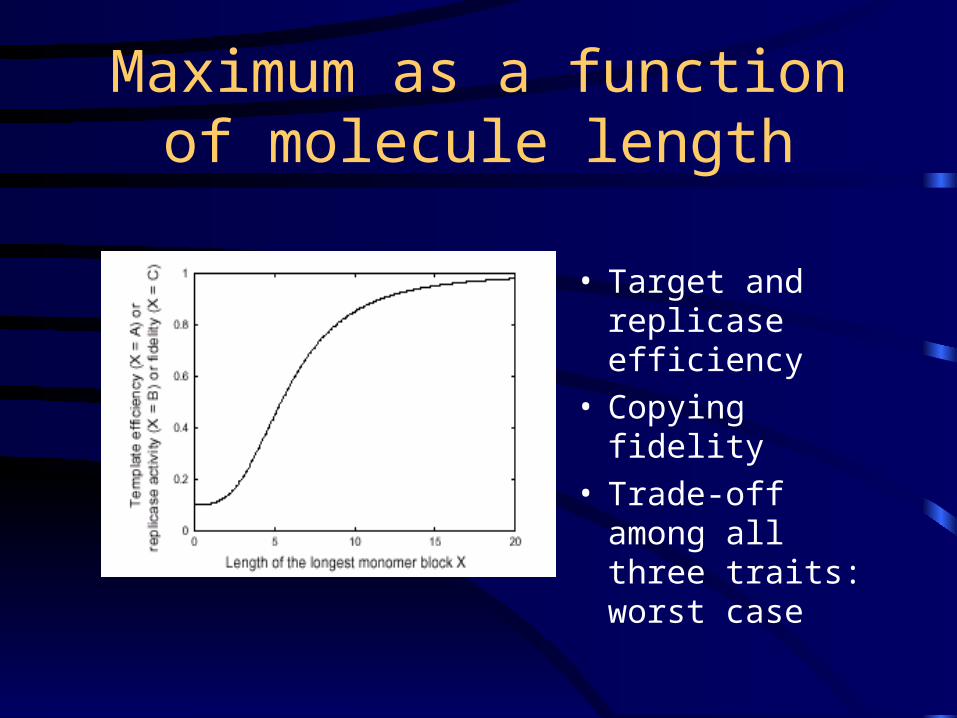
Maximum as a function of molecule length
• Target and replicase efficiency
• Copying fidelity• Trade-off among
all three traits: worst case
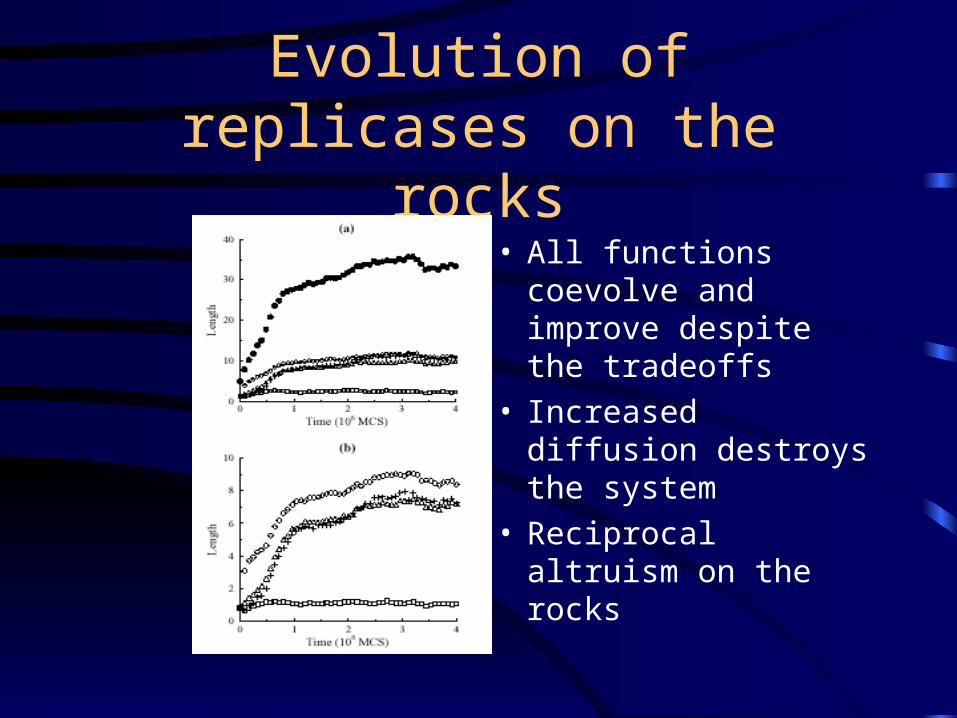
Evolution of replicases on the rocks
• All functions coevolve and improve despite the tradeoffs
• Increased diffusion destroys the system
• Reciprocal altruism on the rocks
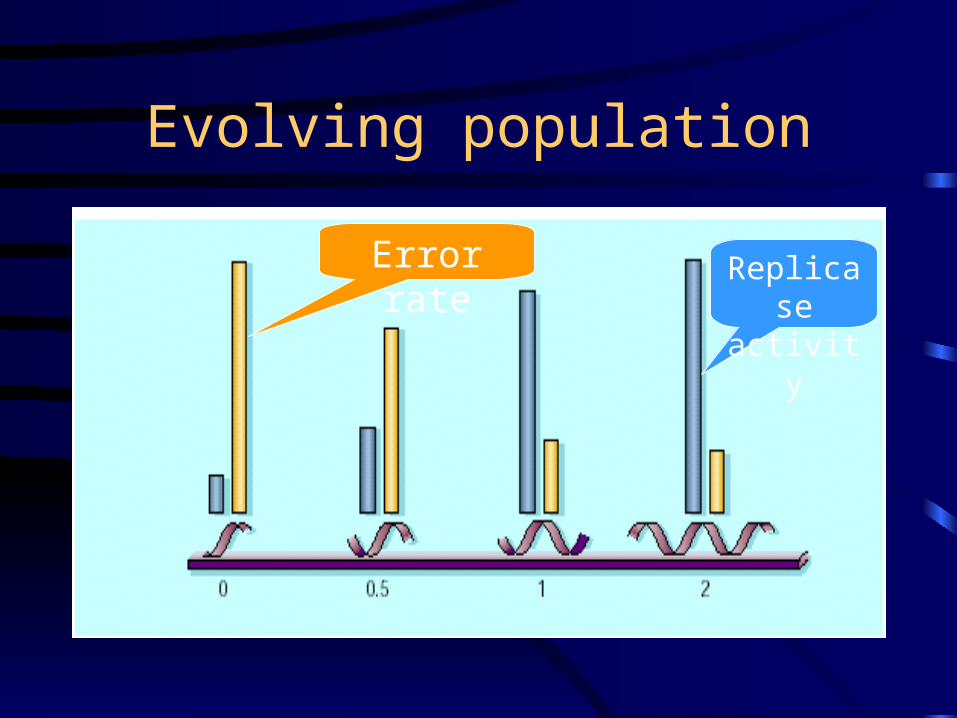
Evolving population
Error rate Replicase activity
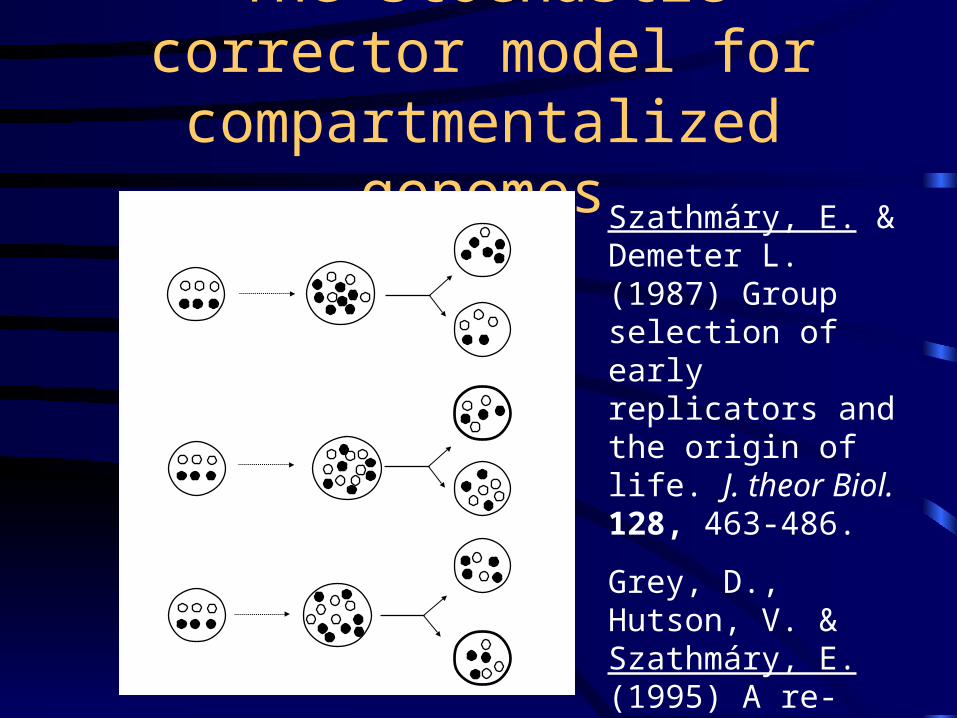
The stochastic corrector model for compartmentalized genomes
Szathmáry, E. & Demeter L. (1987) Group selection of early replicators and the origin of life. J. theor Biol. 128, 463-486.
Grey, D., Hutson, V. & Szathmáry, E. (1995) A re-examination of the stochastic corrector model. Proc. R. Soc. Lond. B 262, 29-35.
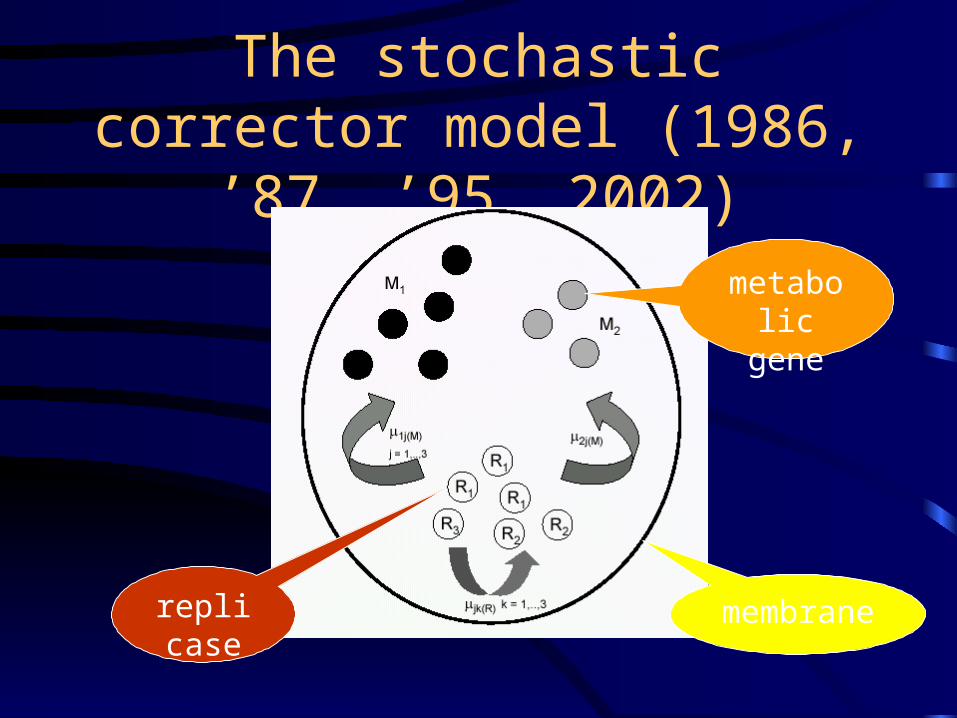
The stochastic corrector model (1986, ’87, ’95, 2002)
metabolic gene
replicase
membrane

Dynamics of the SC model• Independently reassorting genes (ribozymes
in compartments)• Selection for optimal gene composition
between compartments• Competition among genes within the same
compartment• Stochasticity in replication and fission
generates variation on which natural selection acts
• A stationary compartment population emerges

The paradox of gene dosage
• Koch, A.L. (1980)
• High dosage: safeguard against stochastic loss
• BUT dilution of beneficial mutations
• If genes are ribozymes, there is a concentration effect on catalysis

Group selection of early replicators: sitting in the same boat
• Many more compartments than templates within any compartment
• No migration (fusion) between compartments
• Each compartment has only one parent
• Group selection is very efficient
• Selection for replication synchrony

The mathematical model• Inside compartments, there are numbers rather
than concentrations• Stochastic kinetics was applied:• Master equations instead of rate equations: P’(n, t)
= ……. Probabilities• Coupling of two timescales: replicator dynamics
and compartment fission• A quasipecies at the compartment level appears• Characterized by gene composition rather than
sequence

Further considerations
• Selection for replication synchrony by clonal selection on compartments
• Start with a small number of genes synchronise replication rates increase gene number
• Selection for chromosomes: only one child
• Genes are certain to find their partners in the same compartment (Maynard Smith & Szathmáry, 1993)

A surprising recent result: increase of the error threshold by intragenic,
intracompartment recombination
• Santos, M., Szathmáry, E. & Zintzaras, E. (2004) J. Mol. Evol. In press.
• Typically, the wild type copies are lost by mutation pressure
• They can be restored by recombination
• A within-compartment analogue to Muller’s ratchet

A forthcoming publication
• E. Szathmáry & M. Santos (2005) Models of compartments. In Walde, P. (ed.) Topics in Current Chemistry, in prep.













![Johann Sebastian Bach BACH COLLEGIUM JAPAN …BIS-SACD-1981-booklet...Johann Sebastian Bach BACH COLLEGIUM JAPAN ... [Tromba da tirarsi in BWV140] ... ruft uns die Stimme. Bach Collegium](https://img.pdfslide.us/doc/110x75/5b18c8407f8b9a19258c1e2d/johann-sebastian-bach-bach-collegium-japan-bis-sacd-1981-bookletjohann-sebastian.jpg)















