Embed Size (px)
Citation preview

The similarity between Drosophila andC. elegans early embryos extends beyondthe parallels between hunchback and glp-1translational regulation. Both embryos con-tain cytoplasmic "granules" in the posteriorregion of the zygote that are segregated ulti-mately to germ cell precursors during em-bryogenesis-the polar granules of Dro-sophila (14) and the P granules of C. elegans(15) (see figure). In Drosophila, maternalnanos RNA is associated with polar gran-ules; perhaps in C. elegans, a homolog ofnanos is associated with P granules.
What about vertebrates? Does transla-tional repression in the posterior cyto-plasm establish embryonic polarity in these"higher" animals? A hint that this mecha-nism may indeed function in vertebratescomes from the identification of a maternaltranscript that encodes a nanos-like proteincalled Xcat-2 in Xenopus embryos (16).Although the function of Xcat-2 is un-known, its location at the vegetal pole sug-gests a role in early pattern formation. Fur-thermore, a "germ plasm" exists in the veg-etal cytoplasm of amphibian embryos thatmay be analogous to P granules and polargranules of worms and flies (17). Over thepast decade, a handful of molecular mecha-nisms have been implicated in the pattern-ing of Drosophila, C. elegans, and Xenopusembryos (1, 18, 19). On the basis of the di-versity of these mechanisms, the prevail-ing view has been that each embryo hasdifferentially employed a handful of com-mon molecular mechanisms to create itsown coordinate system. For example, local-ized transcriptional activators are utilizedfor patterning of both C. elegans and Dro-sophila early embryos (20-23), but themechanisms for localization, types of DNAbinding protein, and specified fates are notobviously similar.
By contrast, the molecular parallels be-tween hunchback and glp-1 regulation sug-gest the existence of an ancient mechanismfor creating asymmetric patterns of geneexpression in early embryos (see figure).This mechanism is predicted to depend ona trans-acting regulator similar to nanosand to act through cis-acting sequencessimilar to NREs in the 3'UTRs of mater-nal transcripts. If this molecular machineryregulates polarity in embryos as diverse asworms, flies, and frogs, it becomes plausiblethat it influences axis formation in all ani-mal embryos, including mammals. "Molecu-lar tinkering" (24) may then come intoplay to reinforce this primitive patterningcontrol and to derive other axes from it.
Research in Drosophila has pioneeredour understanding of the molecular mecha-nisms that can establish the body axes inan early embryo. Now, phylogenetic com-parisons will tell us which mechanisms areprimitive and which have evolved to rein-
578
force, modify, or extend the underlyingmap. Are the controls that localize transla-tional repression conserved? Are polargranules the ancient seat of pattern gover-nance? What links the early controls ofaxis formation to the later controls ofhomeobox genes, a highly conserved sys-tem that specifies individual regions alongthe anterior-posterior axis of all knownmetazoa (25)? The hunchback protein is atranscriptional regulator that resides at thetop of a cascade of transcriptional regula-tors, whereas the glp-1 protein is a mem-brane receptor that directs inductive inter-actions. Clearly, these distinct modes ofregulation must converge to control expres-sion of homeobox genes. How similar arethe mechanisms of convergence? Answersto these questions, among the most funda-mental of all developmental biology, maybe waiting around the corner.
References
1. D. St. Johnston and C. NOsslein-Volhard, Cell 68,201 (1992).
2. T. C. Evans etal., ibid. 77, 183 (1994).
3. S. C. Morris, Nature 361, 219 (1993).4. A. Sidow and W. K. Thomas, Curr. Biol. 4, 596
(1994).5. J. E. Sulston etal., Dev. Biol. 100, 64 (1983).6. J. R. Priess, H. Schnabel, R. Schnabel, Cell 51,
601 (1987).7. C. C. Mello, B. W. Draper, J. R. Priess, ibid. 77, 95
(1994).8. H. Hutter and R. Schnabel, Development 120,
2051 (1994).9. S. E. Mango et al., ibid., p. 2305.
10. I. P. G. Moskowitz, S. B. Gendreau, J. H.Rothman, ibid., p. 3325.
11. M. Wickens, Ed., Semin. Dev. Biol. 3 (1992).12. R. P. Wharton and G. Struhl, Cell67, 955 (1991).13. C. Wang and R. Lehmann, ibid. 66, 637 (1991).14. A. P. Mahowald, J. Exp. Zool. 176, 329 (1971).15. S. Strome and W. B. Wood, Cell 35, 15 (1983).16. L. Mosquera et al., Development 117, 377 (1993).17. A. W. Blackler, Dev. Biol. 14, 330 (1966).18. W. B. Wood and L. G. Edgar, Trends Genet. 10,
49 (1994).19. D. A. Melton, Science 252, 234 (1991).20. W. Driever and C. NOsslein-Volhard, Cell 54, 83
(1988).21. _, Nature 337, 138 (1989).22. B. Bowerman et al., Cell 74, 443 (1993).23. B. Bowerman, B. A. Eaton, J. R. Priess, ibid. 68,
1061 (1992).24. F. Jacob, The Possible and the Actual. The Jessie
and John Danz Lectures (Pantheon, New York,1982).
25. C. Kenyon, Cell 78,175 (1994).
The Embryonic Vertebrate Forebrain:The Prosomeric Model
John L. R. Rubenstein, Salvador Martinez,Kenji Shimamura, Luis Puelles
The mammalian forebrain-the cerebralcortex, basal ganglia, hypothalamus, andthalamus-is the seat of higher cognitivefunctions. How much of forebrain develop-ment and structure is controlled by a ge-netic program? Although at the later stagesof development incoming synaptic infor-mation from the thalamus has been shownto influence patterning in the neocortex(1), at early embryological stages a specificset of newly discovered genes pattern thebrain into a highly organized structure-be-fore synaptic influences are present. Fur-thermore, the primordia of major structuralelements, such as the thalamus, are segre-gated by cellular boundaries that are alignedparallel to the topologically transverse andlongitudinal axes of the neural tube. Spe-cific combinations of genes that are ex-
J. L. R. Rubenstein and K. Shimamura are in the NinaIreland Laboratory of Developmental Neurobiology,Center for Psychiatry and Neurobiology, Programs inNeuroscience and Developmental Biology, 401Parnassus Avenue, University of California, San Fran-cisco, CA 94143-0984, USA. S. Martinez and L.Puelles are in the Department of Morphological Sci-ences, Faculty of Medicine, University of Murcia,Murcia, Spain 30100.
SCIENCE * VOL. 266 * 28 OCTOBER 1994
pressed in these domains direct the uniquedevelopment of each region. Finally, theorganization of the forebrain indicates thatit is a segmental structure.
The basic morphogenetic unit of em-bryonic insects is a transverse domain, orsegment (2). The identity of each segmentis determined by its position along theanterior-posterior axis and is controlledby the expression of the homeobox seg-ment identity genes (3). These genes en-code transcriptional regulators of specificsets of target genes, which define theunique developmental pathway of eachindividual segment.
It is widely held that this paradigm mayapply to the organization of the somiticmesoderm (the vertebral column), therhombencephalon (hindbrain), and thebranchial arches of vertebrates. This view isbased on the existence of homologs of thehomeobox segment identity genes in verte-brates (the Hox genes) (3) as well as themetameric (segmental) morphological andhistological features of these structures.This hypothesis has been confirmed in partby the use of genetic manipulations that al-
on
Nov
embe
r 7, 2
012
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fro
m

ter the expression of homeobox genes. Forinstance, null mutations in Hox-a2, Hox-b4, Hox-c8, and Hox-d3 and ectopic expres-sion of Hox-a7 and Hox-d4 change theidentity of vertebral and cranial bones(homeotic transformations) (4). In thecentral nervous system, overt morphologi-cal segmentation of the hindbrain is tran-siently apparent during embryogenesiswhen it is subdivided into seven or eightdiscrete units (rhombomeres) by constric-tions in the wall of the neural tube (5).These constrictions correspond to bound-aries that partially restrict the inter-segmental mixing of neuroepithelial cells(6) as well as the intercellular movement ofmolecules whose molecular mass is greaterthan approximately 300 daltons (7). As inthe Drosophila embryo, the expression ofhomeobox and other candidate regulatorygenes is delimited by segment boundaries(8). Moreover, a null mutation in one ofthese genes (Hox-al) alters the develop-ment of the anteriormost segments inwhich it is expressed (9, 10).
Are the more anterior parts of the verte-brate central nervous system also segmen-tally organized? The complex morphologyand histology of the forebrain have led todivergent views about its embryologic orga-nization. A century-old school of neuro-embryology has postulated that segmenta-tion contributes to subdividing functionallydistinct domains of the central nervous sys-tem (1 1). However, the generally acceptedanatomical viewpoint is based on an alter-native model of forebrain organization-the "columnar model" of Herrick andKuhlenbeck [see (11) for a comparison ofthe neuromeric and columnar models]. Re-cently, a number of publications have re-vived efforts to elucidate the organizationof the embryonic forebrain. This renewedinterest was stimulated in part by the dis-covery of a large number of regulatorygenes that are expressed in regionally re-stricted patterns in the forebrain. Amongthese are at least 30 homeobox genes, someof which-such as members of the Dlx,Emx, and Otx families-are related to theDrosophila genes Dll, Ems, and Otd (12).These genes direct the pattern of head de-velopment in Drosophila (13).
Various studies have concluded that atleast part of the forebrain is segmentally or-ganized (11, 12, 14-20). Our conclusionthat the forebrain is made up of segments isbased on morphological considerations (forexample, the presence of transverse con-strictions in the wall of the neural tube inseveral species), on the expression patternsof candidate regulatory genes in mouse andchicken embryos (14, 21), and on experi-mental embryological results (22, 23).We have proposed (11, 12, 14) that the
forebrain is subdivided into six transverse
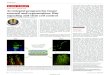
Prosomeres defined by gene expression. The expression of six genes-Dlx-2 14, Gbx-2 14, Nkx-2.119, Nkx-2.2 19, Otx-2 18, and sonic hedgehog (sonic hh) (31)-in the neural plate (E8.5) and theneural tube (E10.5 and E12.5) of the embryonic mouse brain. The fate map of the neural plate isbased on the studies of other workers [see references in (11)], and its relation to the expressionpatterns is hypothetical. The provisional transverse and longitudinal boundaries are indicated asthin black lines. D, diencephalon; E, eyes; H, rhombencephalon-hindbrain; I, isthmus; M, mesen-cephalon-midbrain; os, optic stalk; p, prosomere; r, rhombomere; sc, spinal cord; SP, secondaryprosencephalon. [Data adapted from ( 11, 14, 21)]
domains (forebrain segments) namedprosomeres [by the terminology of Vaage(24)] (see figure). The prosomeres can begrouped into two large transverse subdivi-sions: The diencephalon (which includesprosomeres p1 to p3) and the secondaryprosencephalon (p4 to p6). The ventral re-gion of the secondary prosencephalon con-sists of the hypothalamus; the telencephalicvesicles constitute its dorsal aspect.
Furthermore, according to this "pro-someric model" the forebrain is also orga-nized into longitudinal domains, nonover-lapping regions parallel to the longitudinalaxis of the neural tube. These domains areanalogous to the roof, alar, basal, and floorplates of the spinal cord, and each of theprosomeres is subdivided by them. Themodel rests on our definition of the longi-tudinal axis of the forebrain, which follows
SCIENCE * VOL. 266 * 28 OCTOBER 1994
the ventral and the dorsal midlines. Theprosomeric and columnar models largelydiffer in their definitions of the longitudi-nal axis (11, 25).
This model has been tested by examin-ing the expression of some 30 differentgenes in mouse and chicken embryos atvarious stages of embryogenesis (11, 14,21) (see figure). Each of the transverse(neuromeric) subdivisions coincides withthe expression boundaries of several genes,some of which are shown in the figure (forexample, Gbx-2 is expressed in most of thealar plate of p2). Several genes are ex-pressed in cells located in specific trans-verse boundary zones (sonic hedgehog in thep2-p3 boundary). In addition, the expres-sion patterns have defined a number of lon-gitudinal domains that extend across sev-eral or all of the brain segments (such as
579
E 8.5
,i s~la4
pltl _1 +_
Prechordalfloor regionFloor plate M
Otx-2 ESonic hh aNkx-2.1 ENkx-2.2a
E10.5Basal plate
Floor plateRoof plate
E12.5
2Dlx-2,Otx-2Sonic hhNkx-2.1laNkx-2.2 a Dlx-2 0 Gbx-2 M Otx-2 M Sonic hhf* Nkx-2.1 M
LIE
on
Nov
embe
r 7, 2
012
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fro
m

Nkx-2.2). (The position and number ofsubdivisions is tentative, particularly withinthe secondary prosencephalon.)
This model should be generalizable toother vertebrate species; thus, it will be im-portant to determine whether prosomericorganization is conserved in birds, reptiles,amphibians, and fish. Recent clonal expan-sion (15) and earlier morphologic studies(26) are consistent with diencephalic seg-mentation in the chick. In addition, pre-liminary gene expression studies in thechick are consistent with the prosomericmodel (21).
The prosomeric model provides a mor-phological foundation from which one canmake predictions about potential mecha-nisms that pattern forebrain development.Two-dimensional patterning may be coor-dinated by a scaffold of organizing tissuesthat provide morphogenetic information(27). Thus, some of the patterning alongthe anterior-posterior axis may require thepresence of transverse rings of neuro-epithelia that have inductive and boundaryproperties (transverse organizers). The isth-mus and the zona limitans (p2-p3 bound-ary) may be neuroepithelial organizers (22,23). Perhaps all of the neuromeric bound-aries have organizing properties. Pattern-ing along the dorsal-ventral axis, as well asthe anterior-posterior axis, may dependon ventral signals emanating from theaxial mesoderm (the notochord and theprechordal mesoderm) and the floor plateand dorsal signals emanating from theroof plate. These anterior-posterior anddorsal-ventral signals may contribute tothe progressive parcellation of the neuro-epithelium into a grid-like arrangement ofhistogenic fields such as the thalamus.
The histogenic fields are patterned alongthree dimensions. Patterning along themedio-lateral axis involves differential mi-gration of neuronal populations to sequen-tially form deep and superficial nuclei orlayers. Patterning along the anterior-poste-rior and dorsal-ventral axes may be regu-lated by the boundary zones encasing thefields. For instance, the entire dorsal thala-mus is derived from one alar longitudinaldomain in p2 (see figure). That the tha-lamic anlage gives rise to multiple nuclei
demonstrates that this field is not uniformand must be under the further influence ofasymmetric morphogenetic signals. Perhapseach boundary zone, or the other tissuesflanking the four sides of the thalamic an-lage, produce a unique set of signals thatcreate the morphogenetic complexity nec-essary to pattern this structure. Suggestiveevidence for this hypothesis is found in theexpression patterns of Gbx-2 and Wnt-3within the thalamic anlage, where eachgene is expressed in a complex pattern con-sistent with gradients originating from theboundaries (14).
The neocortical anlage, according toavailable evidence, is not overtly parti-tioned into transverse domains at earlystages of embryogenesis. However, the post-natal neocortex shows region-specific geneexpression (28) and histological structure(29). Pattern formation within the neocor-tex may rely on signals from boundaryzones that lead to region-specific gene ex-pression, histogenesis, and innervation pat-terns. Once afferent fibers arrive, activity-dependent processes could refine the orga-nization of the neocortex.
The prosomeric model also provides aframework for studying evolution of theforebrain. Neuromeric structures are wellsuited to allow for the duplication, dele-tion, or respecification of neuronal do-mains. A newly evolved structure such asthe mammalian neocortex could be formedby the transformation of a preexisting tel-encephalic domain or by the formation of anovel domain.
The prosomeric model is a working hy-pothesis that should facilitate future studiesof forebrain development, but that muststill be refined. The model may prove use-ful, for example, in examining whethergrowing axon pathways derive positionalinformation from signals encoded in thescaffold of domains and boundaries (30).However, many key questions remain: Howare the neural plate and neural tube pat-terned? How are the boundaries formed?What cellular interactions are involved?Where are the sources for the patterningsignals and what are the signals? What arethe transcription factors that specify re-gional identity? Many developmentally im-
portant genes have been discovered; nowtheir functions need to be assessed. Suchstudies will not only elucidate the blueprintof forebrain organization but will also ulti-mately aid in efforts to understand abnor-mal neural development.
References and Notes
1.
2.
3.
4.5.
6.
7.
8.9.
10.
11.
12.
13.
14.15.
16.
17.
18.
19.20.
21.
22.
23.24.
25.
26.
27.28.
29.30.
31.32.
D. D. M. O'Leary and S. E. Koester, Neuron 10,991 (1993).P. A. Lawrence, The Making of a Fly (Blackwell,London, 1992).W. McGinnis and R. Krumlauf, Cell 68, 283(1992).F. M. Rijli etal., ibid. 75, 1333 (1993).A. Lumsden and R. Keynes, Nature 337, 424(1989).E. Birgbauer and S. E. Fraser, Development 120,1347 (1994).S. Martinez, E. Geijo, M. V. Sanchez-Vives, L.Puelles, R. Gallego, ibid. 116,1069 (1992).R. Krumlauf, Trends Genet. 9, 106 (1993).P. Dolle et al., Proc. Natl. Acad. Sci. U.S.A. 90,7666 (1993).E. M. Carpenter et al., Development 118, 1063(1993).L. Puelles and J. L. R. Rubenstein, TrendsNeurosci. 16, 472 (1993).J. L. R. Rubenstein and L. Puelles, Curr. Top.Dev. Biol. 29, 1 (1994).S. Cohen and G. Jurgens, Trends Genet. 7, 267(1991).A. Bulfone et al., J. Neurosci. 13, 3155 (1993).M. C. Figdor and C. D. Stern, Nature 363, 630(1993).P. C. Salinas and R. Nusse, Mech. Dev. 39, 151(1992).A. Zimmer and A. Zimmer, Development 116, 977(1992).E. Boncinelli, M. Gulisano, V. Broccoli, J.Neurobiol. 24, 1356 (1993).M. Price, ibid., p. 1385.A. Stoykova and P. Gruss, J. Neurosci. 14, 1395(1994).K. Shimamura, S. Martinez, J. L. R. Rubenstein,unpublished results.S. Martinez, M. Wassef, R. M. Alvarado-Mallart,Neuron 6, 971 (1991).F. Marin and L. Puelles, Dev. Biol. 163, 19 (1994).S. Vaage, Ergeb. Anat. Entwicklungsgesch. 41, 1(1969).L. S. Ross, T. Parrett, S. S. Easter, J. Neurosci. 12,467 (1992).L. Puelles, J. A. Amat, M. Martinez-de-la-Torre, J.Comp. Neurol. 266, 247 (1987).A. Ruiz Altaba, Trends Neurosci. 17, 233 (1994).M. Cohen-Tannoudji, C. Babinet, M. Wassef, Na-ture 368, 460 (1994).P. Rakic, Science 241, 170 (1988).S. W. Wilson, M. Placzek, A. J. Furley, TrendsNeurosci. 16, 316 (1993).Y. Echelard et al., Cell 75, 1417 (1993)Supported by grants to J.L.R.R. [March of Dimes,National Alliance for Research on Schizophreniaand the Depressions, the John Merck Fund,Pfizer Pharmaceuticals, and the National Insti-tutes of Health (MH49428-01 and MH01046-01)],S.M. (NATO), and L.P. (DGICYT PB90/0296C01).
SCIENCE * VOL. 266 * 28 OCTOBER 1994
am~ iiimiiiiiiiii'immw~ 'I'llii iii .... ii i,iiiim-
580
on
Nov
embe
r 7, 2
012
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fro
m



![[XLS] of Missed PPOs... · Web viewJOGA RAMAN MALATI PURKAYASTHA Life And Non-Marriage Certificate Not Available SOPHILA GURUNG BENDER LYNGDOH HORINWELL TONGWAH MILTYMAI LALOO MARY](https://img.pdfslide.us/doc/110x75/5ab607607f8b9ab47e8d76f3/xls-of-missed-pposweb-viewjoga-raman-malati-purkayastha-life-and-non-marriage.jpg)