Embed Size (px)
Citation preview

Journal of Pineal Research 2:13-19 (1985)
The Effects of Photoperiod and Melatonin on Serum Prolactin Levels of Mink During the
Autumn Molt
Jack Rose, Fredrick Stormshak, James Oldfield, and John Adair Department of Animal Science, Oregon State University, Corvallis
An experiment was conducted to determine the effects of a reduced daily photoperiod and exogenous melatonin on serum prolactin levels of mink during the autumn molt and growth of the winter pelage. During the last week of June, adult standard dark female mink (Mustela vison) were exposed to natural changes in daylength (controls), a reduced photoperiod of 6 h light: 18 h dark (6L: 18D) or exposure to natural changes in daylength and treated with melatonin (10 nig) in a Silastic implant inserted subcutaneously over the scapular area. Beginning July 2 , and continuing through October 22, blood samples were collected at nine biweekly intervals, and serum prolactin concentrations were quantified by a heterologous double antibody radioimmunoassay. Both reduced photoperiod and exogenous melatonin caused serum prolactin levels to decline rapidly after mid- July, resulting in concentrations that were significantly lower than those o f controls 6 to 8 wk earlier. These data suggest that growth of the winter pelage of mink is strongly associated with declining prolactin levels. I t appelrs that part o f the photoperiodic-induced effects on fur growth of the mink are mediated through melatonin and its effects on prolactin synthesis and/or secretion.
Key words: mink, pelage, photoperiod, pituitary, pineal, prolactin, melatonin
Received September 24, 1984; accepted November 9, 1984.
Technical Paper No. 7279, Oregon Agr. Exp. Station
Address reprint requests to Dr. F. Stormshak, Department of Animal Science, Oregon State University, Corvallis, OR 97331.
0 1985 Alan R. Liss, lnc.

14 Rose etal.
INTRODUCTION
Mink exhibit a seasonal fur growth cycle, which has been shown to be regulated by changes in photoperiod [Bissonnette and Wilson, 1939; Ham- mond, 1954; Duby and Travis, 19721. Decreasing daylength after the summer solstice stimulates molting and growth of the winter pelage, while increasing daylength following the spring equinox induces molting and growth of the summer fur. Exposing mink to an artificially reduced daily photoperiod (6L: 18D), beginning at the end of June, stimulates molting of the summer fur and growth of fully prime winter pelage by mid-October, 6 wk earlier than normal [Weiss et al., 1980; Rose et al., 19841.
The mechanism by which photoperiod influences fur growth cycles is unknown; however, considerable evidence indicates that both the pituitary and pineal gland are involved. Mink that are hypophysectomized, either during winter or summer pelage, exhibit a series of asynchronous molts followed by regrowth of a subnormal winter fur [Rust et al., 19651. Prolactin levels have been shown to be influenced by photoperiodic changes in mink [Martinet et al., 1981, 192131 and a variety of other species [Buttle, 1974; Bex et al., 1978; Peters and Tucker, 19781. It is possible, therefore, that this pituitary hormone is necessary for normal fur growth cycles. Furthermore, in mink, spring and autumn molts are correlated with increasing and decreas- ing plasma prolactin concentrations, respectively [Allain et al., 1981 ; Marti- net et al., 1981, 19831. It has also been demonstrated that the administration of melatonin to mink [Allain and Rougeot, 1980; Rose et al., 19841 and weasels [Rust and Meyer, 19691 in summer accelerates molting and growth of the winter fur much earlier than occurs naturally. These observations implicate the pineal gland in the regulation of fur growth cycles. Because melatonin secretion normally increases during darkness [Reiter, 19831, it is possible that the seasonal pattern of melatonin secretion in the mink is diametrically opposed to that of prolactin. Thus, it is conceivable that a regulatory mechanism exists between these two glands in the control of fur growth cycles.
The objectives of the present experiment were to determine the effects of a short photoperiod and exogenous melatonin on serum prolactin levels of adult standard dark female mink during the autumn molt and growth of the winter fur.
MATERIALS AND METHODS Animals and Blood Collection
On June 26, 1980, 30 standard dark adult female mink (MusteZa vison) were assigned randomly to an experiment of 3 X 9 factorial design. There were three treatments consisting of exposure to (1) natural changes in day- length (controls), (2) reduced photoperiod of 6 h light: 18 h dark (6L: 18D), and (3) natural changes in daylength with each animal receiving a Silastic (Dow Corning, Midland, MI) implant containing 10 mg melatonin (Sigma, St. Louis, MO) inserted subcutaneously over the scapular area. Implants were constructed according to the procedures of Moore [ 19811. The effects of the

Pineal-Pituitary Interactions 15
above treatments on molting and winter fur growth of mink have been previously reported [Rose et al., 19841.
Beginning July 2 and continuing through October 22, mink were bled by cardiac puncture nine times at biweekly intervals, under ketamine hydro- chloride anesthesia. Blood samples taken between 1300 and 1500 h were allowed to clot for 24 h at 4"C, prior to harvesting serum, which was subsequently stored at -15°C until assayed. As a result of using cardiac puncture to obtain blood samples, several mink in each group died. Only serum from those animals that survived the experiment were analyzed for prolactin concentrations (samples sizes are indicated in the legend of Fig. 2).
Radioiodinations
Porcine prolactin' (PPRL) utilized in the radioimmunoassays was iodi- nated using the chloramine-T method of Greenwood et al. [ 19631 as modified by Kraeling et al. [1982]. At each iodination 25 p1 of 0.5 M phosphate buffered saline (PBS, pH 7.5) were added to the reaction vial containing the pPRL (5 pg in 5 pl 0.01 M ammonium bicarbonate, pH 8.6), followed by 1 mCi Na'251 (IMS.30; 100 mCi/ml, Amersham Corp., Arlington Heights, IL) and 30 pg chloramine-T (Sigma) in 0.05 M phosphate buffer (PH 7.5). The mixture was allowed to react for 15 s, with gentle agitation, and the reaction was stopped by the addition of 125 pg sodium metabisulfite in 0.05 M phosphate buffer. The reaction mixture was then diluted with 100 p1 of 1 % potassium iodide in 16% sucrose, and the entire contents were layered on an 8 X 120 mm column of Sephadex (Sigma) G-75-120, previously equili- brated with 0.05 M phosphate buffer @H 7.5). The reaction vial was then rinsed with 100 pl of 2.5% BSA-Frac-V (Sigma) in PBS, followed by 70 p1 of 1% potassium iodide in 8% sucrose, and both rinses were added to the column. Thirty O.5-ml fractions were collected into tubes containing 0.5 ml PBS-1% BSA (assay buffer). Two peaks of radioactivity were observed with the '251-pPRL eluting in the first peak (fractions 5-10). Further purification was achieved by passing 1-ml fractions containing the iodinated hormone through a column (0.9 x 40.0 cm) of Sephadex (Sigma) G-100-120, previ- ously equilibrated with 0.05 M phosphate buffer (PH 7.0). Approximately forty 1-ml fractions were collected into tubes, each containing 1 ml of assay buffer. Three peaks of radioactivity were obtained with material from the second peak (fractions 26-27) containing the immunoreactive 251-pPRL.
PRL Radioimmunoassay
Mink serum prolactin concentrations were measured by a heterologous double antibody radioimmunoassay based on the procedures developed by Niswender et al. [ 19691. To test for assay accuracy, mink serum (200 p1) plus 100 p1 of assay buffer or varying amounts of standard pPRL (0.039 to 40.0 ng) in 300 p1 assay buffer were added to 12 x 75-mm glass culture tubes. To
'Porcine prolactin for radioiodinations (USDA-pPRL-I- 1 ) and for use as standards (USDA-pPRL- B-I) was generously provided by Dr. D. Bolt of the USDA Reproduction Laboratory, Beltsville, MD.

16 R o s e e t al.
each tube was added 100-pl goat anti-pPRL serum (Research Products Inter- national [RPI], Mt. Prospect, IL, Lot No. 10333) diluted 1: 165,000 in 0.05 M EDTA-PBS (PH 7.0). Nonspecific binding tubes received 100 pl of 1:200 normal goat serum (RPI, Lo t No. 22980) in 0.05 M EDTA-PBS (PH 7.0) , in place of the first antibody. All tubes were gently vortexed and incubated at 4°C for 24 h. Following incubation, 100 pl of 1251-pPRL (14,000 cpm) were added to all tubes, vortexed, and incubated an additional 24 h at 4°C. Subsequently, 200 pl of rabbit anti-goat IgG serum (RPI, Lot No. 2439 8 L ) diluted 1: 12 in assay buffer were added to all tubes, vortexed, and incubated another 72 h at 4°C. The reaction was then terminated by adding 3 ml PBS (PH 7.0) to all tubes and centrifuging at 1,500g for 30 min at 4°C. The resulting supernatant was decanted, and the pellets counted in a Packard Gamma Counter with 60% counting efficiency.
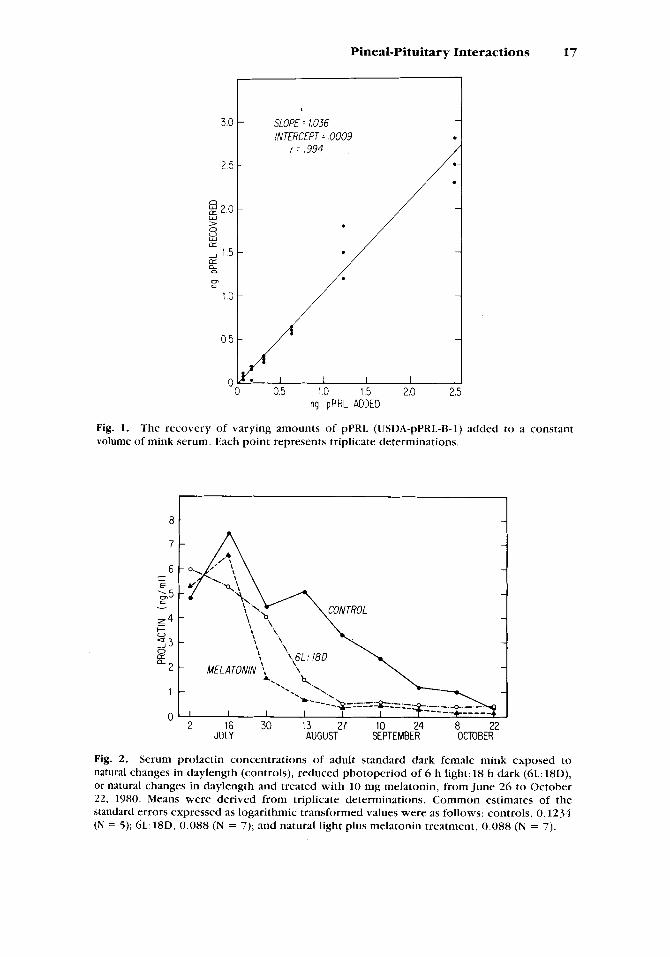
The recovery of various amounts of standard pPRL (.078 to 2.5 ng) added to 100 p1 of pooled mink serum is depicted in Figure 1. Regression analysis revealed a slope of 1.036, an intercept of 0.0009, and a correlation coefficient of 0.994. Dose response-curves for mink serum and for increasing concentrations of the standard pPRL, were parallel, indicating no nonspecific interference. Sensitivity of the assay was 0.13 ng/tube, which compares favorably with the porcine PRL assay of Kraeling et al. [1982]. The intraassay coefficient of variation was 9.6%. No interassay coefficient of variation was calculated because all samples were analyzed in a single assay.
Kraeling et al. [1982] have evaluated the hormonal specificity of the primary antisera, for measuring pPRL, and found that the cross-reactivity with other pituitary hormones (FSH, LH, GH) even in large quantities (2- 2,000 ng) to be less than 1 % . However, because purified mink hormones are unavailable, we cannot rule out the possibility of cross-reactivity with endog- enous hormones, especially GH.
Statistical Analysis
Because of heterogeneity of variance, data on serum PRL levels were subjected to natural logarithmic transformation. The transformed data were then analyzed by the use of split-plot analysis of variance. Differences be- tween specific means were tested using unpaired Student's t-tests. Nontrans- formed data are utilized in graphically depicting the changes in serum PRL concentrations in control and treated mink over time.
RESULTS AND DISCUSSION
Serum prolactin concentrations in mink were markedly affected by treatments (treatment X time interaction, P < 0.0001). Prolactin levels in animals maintained under natural daylength conditions (controls) peaked in mid-July and then declined throughout the study (Fig. 2). However, both reduced light and exogenous melatonin caused PRL levels to drop precipi- tously after July 16 and were reduced to basal levels (< 1.5 ng/ml) 6 and 8 wk, respectively, earlier than those of controls (P < 0.05). In rats, and primates, plasma prolactin levels are low during the day and elevated at night [Dunn et al., 1972; Sassin et at., 1972; Kizer et al., 1975; Brainard et al.,
Pineal-Pituitary Interactions 17
3.0
2 5
n g 2 0
:: 1 1.5 a,
W > 0
LL
LL
cn L:
10
0 5
SLOPE = 1.036 lNTERCEPT ,0009
ng pPRL ADDED
Fig. 1. volume of mink serum. Each point represents triplicate determinations.
The recovery of varying amounts of pPRL (USDA-pPRL-B-1) added to a constant
8
7
6 - - E . c 0 5
z4
93 &? a2
1
+ 0
1
0 2 16 30 13 27 10 24 8 22
JULY AUGUST SEPTEMBER OCTOBER
Fig. 2. Serum prolactin concentrations of adult standard dark female mink exposed to natural changes in daylength (controls), reduced photoperiod of 6 h light: 18 h dark (6L: 18D), or natural changes in daylength and treated with 10 mg melatonin, from June 26 to October 22, 1980. Means were derived from triplicate determinations. Common estimates of the standard errors expressed as logarithmic transformed values were as follows: controls, 0.1234 (N = 5) ; 6 L : 18D, 0.088 (N = 7); and natural light plus melatonin treatment, 0.088 (N = 7).

18 Rose et al.
19811. It is not known whether; a circadian rhythm in prolactin secretion occurs in mink. If such a rhythm does occur in this species, we cannot exclude the possibility that reduced light and/or exogenous melatonin might promote a phase shift that could account in part for the observed reduction in prolactin concentrations.
We have previously reported fur growth patterns for the animals uti- lized in the present study [Rose et al., 19841. It is intriguing that the rapid growth of winter pelage did not begin in any of the three groups until serum PRI, had fallen to basal levels. Rapid growth of fur began shortIy after September 10 in the control group, August 13 in the reduced light group, and slightly earlier in the melatonin-treated group.
Martinet et al. [1983] have demonstrated that mink exposed to a re- duced daily photoperiod or given injections of melatonin, prior to the spring molt, have significantly reduced plasma prolactin concentrations, and that the spring molt is retarded as long as prolactin levels remain low. Further- more, prolactin injections, given 7 days after mating, have been shown to induce the spring molt in mink at an earlier date than that which occurs normally [Martinet et al., 19811. Although the effects of exogenous melatonin on the autumn molt and growth of the winter fur are stimulatory, this effect does not extend to serum prolactin levels. Allain et al. [I9811 found that the administration of melatonin to male pastel mink during late April or early July caused a premature reduction in plasma prolactin concentrations, but in contrast to the effects of melatonin on the spring molt [Martinet et al., 19831, molting and regrowth of the winter fur were enhanced.
Our data support the observations of Allain et al. [198l] and extend these findings to adult standard dark female mink. Furthermore, by maintain- ing animals on a constant reduced daily photoperiod, it appears that part of the photoperiodic-induced effects on molting and growth of the winter fur of mink are mediated through the pineal hormone melatonin, and its effects on pituitary prolactin synthesis and/or secretion. These findings make it clear that reduced serum prolactin levels are associated with the onset of winter fur growth. It is possible that prolactin may exert a stimulatory effect on fur growth during the spring molt, as shown by Martinet et al. [ 19831 and inhibit the autumn molt until hormone concentrations decline to basal levels. Duncan and Goldman [ 19841 have shown that injections of oPRL inhibit the short photoperiod-induced winter molt of the Djungarian hamster. Such experiments have not been conducted thus far with mink.
ACKNOWLEDGMENTS
The authors wish to thank Catherine Gardiner-Laird for technical assis- tance with the radioimmunoassays (RIA). Dr. Robert Kraeling of the USDA Richard B. Russell Agricultural Research Center, Athens, GA, generously supplied the goat anti-pPRL for development of the RIA as well as technical advice. We would like to acknowledge Ron Scott and Cliff Thomson for care and handling of mink and Dr. Ken Rowe, Dept. of Statistics, for assistance with the statistical analysis. Mink were obtained through the Experimental Fur Farm of Oregon State University. This research was sup- ported by the Mink Farmers Research Farmers Research Foundation of America.

Pineal-Pituitary Interactions 19
LITERATURE CITED
Allain, D., L. Martinet, J. Rougeot,(l981) Effect of melatonin implants on changes in the coat, plasma prolactin level and testis cycle in the mink (Mustela vison). In: Photoperiodism and Reproduction. R. Ortavant, J. Pelletier, and J.P. Ravault, eds., INRA Publ., Paris, pp. 263-271.
Allain, D., J. Rougeot (1980) Induction of autumn moult in mink (Mustela vison Peak and Beauvois) with melatonin. Reprod. Nutr. Dev. 20: 197-201.
Bex, F., A. Bartke, B.D. Goldman, S. Dalterio (1978) Prolactin, growth hormone, luteinizing hormone receptors, and seasonal changes in testicular activity in the golden hamster. Endocrinology 103:2069-2080.
Bissonnette, T.H., E. Wilson (1939) Shortening daylight periods between May 15 and Septem- ber 12 and the pelt cycle of the mink. Science 89:418-419.
Brainard, G.C., R.H. Asch, R.J. Reiter (1981) Circadian rhythms of serum melatonin and prolactin in the Rhesus monkey (Macaca mulatta). Biomed. Res. 2:291-297.
Buttle, H . L . (1974) Seasonal variation of prolactin in plasma of male goats. J. Reprod. Fertil. 37 :95-99.
Duby, R.T., H.F. Travis (1972) Photoperiodic control of fur growth and reproduction in the mink (Mustela vison). J. Exp. Zool. 182:217-225.
Duncan, M.J., B.D. Goldman (1984) Hormonal regulation of the annual pelage color cycle in the Djungarian hamster (Phodopus sungorus). 11. Role of prolactin. J . Exp. 2001. 230:97- 103.
Dunn, J.D., A. Arimura, L.E. Scheving (1972) Effect of stress on circadian periodicity in serum LH and prolactin concentration. Endocrinology 90:29-33.
Greenwood, F.C., W.M. Hunter, J.S. Glover (1963) The preparation of "'I labeled human growth hormone of high specific radioactivity. Biochem. J. 89: 114-123.
Hammond, J., Jr. (1954) Effects of Artificial Lighting on the Reproductive and Pelt Cycles of Mink. W. Heffer and Sons, Ltd., Cambridge, p. 16.
Kizer, J.S., J.A. Zivin, D.M. Jacobowitz, I.J. Kopin (1975) The nyctohemeral rhythm of plasma prolactin: Effects of ganglionectomy, pinealectomy, constant light, constant darkness or 6-OH-dopamine administration. Endocrinology 96: 1230-1240.
Kraeling, R.R., G.B. Rampacek, N.M. Cox, T.E. Kiser (1982) Prolactin and luteinizing hormone secretion after bromocryptine (CB- 154) treatment in lactating sows and ovariectomized gilts. J. Anim. Sci. 54:1212-1220.
Martinet, L., D. Allain, M. Meunier (1983) Regulation in pregnant mink (Mustela vison) of plasma progesterone and prolactin concentrations and regulation of onset o f the spring moult by daylight ratio and melatonin injections. Can. J. 2001. 61: 1959-1963.
Martinet, L., M. Meunier, D. Allain (1981) Control of delayed implantation and onset of spring moult in the mink (Mustela vison) by daylight ratio, prolactin and melatonin. In: Photoperiodism and Reproduction. R. Ortavant, J. Pelletier, and J.P. Ravault, eds., INRA Publ., Paris, pp. 253-261.
Moore, F.L. (1981) Technique for making small hormone-filled capsules. Gen. Comp. Endocri- nol. 43:409.
Niswender, G.D. , L.E. Reichert, Jr., A.R. Midgley, Jr., A.V. Nalbandov (1969) Radioimmunoas- say for bovine and ovine luteinizing hormone. Endocrinology 84: 1166-1173.
Peters, R.R., H.A. Tucker (1978) Prolactin and growth hormone responses to photoperiod in heifers. Endocrinology 103:229-234.
Reiter, R.J. (1983) The role of light and age in determining melatonin production in the pineal gland. In: The Pineal Gland and Its Endocrine Role. J. Axelrod, F. Fraschini, and G.P. Velo, eds., Plenum Publ. Corp., pp. 227-241.
Rose, J., F. Stormshak, J. Oldfield, J. Adair (1984) Induction of winter fur growth in mink (Mustela vison) with melatonin. J. Anim. Sci. 58:57-61.
Rust, C.C., R.K. Meyer (1969) Hair color, molt, and testis size in male, short-tailed weasels treated with melatonin. Science 165:92 1-922.
Rust, C.C., R.M. Shackelford, R.K. Meyer (1965) Hormonal control of pelage cycles in the mink. J. Mammol. 46:549-565.
Sassin, J.F., A.G. Frantz, E.D. Weitzman, S. Kapen (1972) Human prolactin: 24-hour pattern with increased release during sleep. Science 177: 1205-1207.
Weiss, D.E., L.V. Swanson, J. Adair, J.E. Oldfield, F. Stormshak (1980) Photoperiodic effects on serum glucocorticoids and fur growth in mink. J. Anim. Sci. 51: 1367-1372.




![EARLY FLOWERING3 Redundancy Fine-Tunes Photoperiod … · EARLY FLOWERING3 Redundancy Fine-Tunes Photoperiod Sensitivity1[OPEN] Andrew J.S. Rubenach, Valérie Hecht, Jacqueline K](https://img.pdfslide.us/doc/110x75/5f70a7e86c02c415f04ab3da/early-flowering3-redundancy-fine-tunes-photoperiod-early-flowering3-redundancy-fine-tunes.jpg)