Embed Size (px)
Citation preview

The Effects of Lactobacillus rhamnosus GR-1 on
Cytokines/Chemokines and Prostaglandins in Human Amnion Cells
by
Rebecca Jean Elizabeth Koscik
A thesis submitted in conformity with the requirements
for the degree of Masters of Science
Department of Physiology
University of Toronto
© Copyright by Rebecca Jean Elizabeth Koscik (2011)

ii
The Effects of Lactobacillus rhamnosus GR-1 on
Cytokines/Chemokines and Prostaglandins in Human Amnion Cells
Rebecca Koscik
Masters of Science
Department of Physiology
University of Toronto
2011
Abstract
The incidence of preterm labor has risen over recent decades and preventative treatments are
ineffective. Associated with a 40% increased risk of preterm birth, bacterial vaginosis is
characterized by a decrease in lactobacilli and an increase in pathogenic bacteria in the vaginal
microbiota. Ascent of bacterial products to the intrauterine environment stimulates cytokine and
prostaglandin secretion from invading immune cells and gestational tissues. Probiotic
lactobacilli modulate the immune responses in mouse macrophages and human placental
trophoblast cells. The focus of this thesis was to determine the influence of Lactobacillus
rhamnosus GR-1 (GR-1) on cytokines and prostaglandins which are part of the activated
pathway in infection and/or inflammation-mediated preterm labour. Pro-inflammatory cytokines
were decreased by GR-1. The release of several chemokines and prostaglandin E2 were elevated
by GR-1. It is possible that GR-1 may enhance the host defense barriers of the amnion to
pathogenic bacteria.

iii
“When is it that nature does anything in vain?”
Paraphrased from Sir Isaac Newton

iv
Dedicated to those who guided me with their years of wisdom, intellectual creativity and passion
for academia; to those who insisted on lunch breaks and offered constant encouragement and
reassurance during my midnight trips to the laboratory; to wiped tears, shared happiness and
laughter; and to my family whose love and support has made me capable of overcoming
challenges and of celebrating successes.

v
Table of Contents
Abstract .......................................................................................................................................... ii
Table of Contents ...........................................................................................................................v
List of Abbreviations ................................................................................................................... ix
List of Figures .............................................................................................................................. xii
Chapter 1 ........................................................................................................................................1
General Introduction .....................................................................................................................2
1.1 Human Parturition ............................................................................................................2
1.1.1 Anatomy ..................................................................................................................4
1.1.2 Mechanisms of Human Parturition ......................................................................9
1.2 Preterm Birth ...................................................................................................................12
1.2.1 Epidemiology ........................................................................................................12
1.2.2 Etiology .................................................................................................................13
1.2.3 Current Diagnostic Strategies and Treatments for Preterm Birth .................13
1.3 Infection and Inflammation ............................................................................................15
1.3.1 Vaginal Microbiota ..............................................................................................16
1.3.2 Routes of Intrauterine Infection .........................................................................17
1.3.3 Bacterial Vaginosis...............................................................................................18
1.3.4 Current Diagnostic Strategies and Treatments for Bacterial Vaginosis ........19
1.4 Probiotics ..........................................................................................................................22
1.4.1 Lactobacillus Species as Candidate Probiotics for use During Pregnancy. ....23
1.4.2 The Potential Mechanisms of Probiotics............................................................26
1.5 Cytokines and Chemokines .............................................................................................27
1.5.1 Pro-inflammatory Cytokines ..............................................................................28
1.5.2 CXC and CC Chemokines...................................................................................30

vi
1.5.3 Immune Cell Activating Cytokines ....................................................................31
1.6 Prostaglandins ..................................................................................................................34
1.6.1 Prostaglandin Production ...................................................................................34
1.6.2 The Role of Amnion Produced Prostaglandins in Preterm Labour ................39
1.7 Summary ...........................................................................................................................41
Chapter 2 ......................................................................................................................................42
Hypotheses, Rationale and Specific Aims ..................................................................................43
2.1 Overall Hypothesis ...........................................................................................................43
2.1.1 Rationale ...............................................................................................................43
2.2 Chapter 3: Effect of Lactobacillus rhamnosus GR-1 on Cytokines and
Chemokines in Human Amnion Cells at Term .............................................................44
2.3 Chapter 4: Effect of Lactobacillus rhamnosus GR-1 on Prostaglandin Output
and Prostaglandin Metabolizing Enzymes in Human Amnion Cells at Term ...........45
Chapter 3 ......................................................................................................................................46
Effect of Lactobacillus rhamnosus GR-1 on Cytokines and Chemokines in Human
Amnion Cells at Term .................................................................................................................47
3.1 Introduction ......................................................................................................................47
3.2 Methodology .....................................................................................................................48
3.2.1 Placenta Collection...............................................................................................48
3.2.2 Mixed Amnion Epithelial and Mesenchymal Cell Culture ..............................48
3.2.3 Lactobacilli Preparation ......................................................................................49
3.2.4 Treatment Protocol ..............................................................................................50
3.2.5 Lactate Dehydrogenase Assay ............................................................................50
3.2.6 Cytokine and Chemokine Measurement ...........................................................51
3.2.7 Statistical Analysis ...............................................................................................52
3.3 Results ...............................................................................................................................52
3.4 Comment ...........................................................................................................................57

vii
Chapter 4 ......................................................................................................................................70
Effect of Lactobacillus rhamnosus GR-1 on Prostaglandin Output and Prostaglandin
Metabolizing Enzymes in Human Amnion Cells at Term .......................................................71
4.1 Introduction ......................................................................................................................71
4.2 Methodology .....................................................................................................................72
4.2.1 Placenta Collection...............................................................................................72
4.2.2 Amnion Mixed Epithelial and Mesenchymal Cell Culture ..............................72
4.2.3 Lactobacilli Preparation ......................................................................................73
4.2.4 Treatment Protocol ..............................................................................................73
4.2.5 Lactate Dehydrogenase Assay ............................................................................73
4.2.6 Prostaglandin E2 Measurement ..........................................................................73
4.2.7 Prostaglandin Metabolizing Enzyme Expression Measurement .....................74
4.2.8 Placental Trophoblast Cell Culture ...................................................................75
4.2.9 Statistical Analysis ...............................................................................................76
4.3 Results ...............................................................................................................................76
4.4 Comment ...........................................................................................................................78
Chapter 5 ......................................................................................................................................91
Overall Conclusions and Future Directions ..............................................................................92
5.1 Overall Conclusions .........................................................................................................92
5.1.1 Potential Increases in TNFα and IL-1β Antagonists ........................................92
5.1.2 Recruitment of Immune Cells .............................................................................93
5.1.3 Modulation of Immune Cell Activity .................................................................93
5.1.4 Stimulation of Prostaglandin E2 .........................................................................95
5.1.5 Additional Effects of Prostaglandins on the Amnion .......................................97
5.1.6 Lactobacillus rhamnosus GR-1 Supernatant Metabolite(s) .............................98

viii
5.1.7 Lactobacillus rhamnosus GR-1 and Prevention of Preterm Birth
Associated with Infection and/or Inflammation................................................99
5.1.8 Limitations ..........................................................................................................100
5.2 Future Directions ...........................................................................................................102
References ...................................................................................................................................105

ix
List of Abbreviations
ACTH Adrenocorticotropic Hormone
BSA Bovine Serum Albumin
cAMP cyclic Adenosine Monophosphate
CAP Contraction Associated Protein
CCB Calcium Channel Blocker
cGMP cyclic Guanosine Monophosphate
CRH Corticotropin-Releasing Hormone
DMEM Dulbecco Modified Eagle Medium
ECL Enhanced Chemiluminescence
EIA Enzyme Immuno-Assay
ELISA Enzyme-Linked ImmunoSorbent Assay
ER Estrogen Receptor
FBS Fetal Bovine Serum
G-CSF Granulocyte Colony Stimulating Factor
GM-CSF Granulocyte-Macrophage Colony Stimulating Factor
HPA Hypothalamic-Pituitary-Adrenal
HSD Hydroxysteroid Dehydrogenase
IFNγ Interferon γ
IL Interleukin

x
LDH Lactate Dehydrogenase
LTA Lipoteichoic Acid
LPS Lipopolysaccharide
MAMP Microbe-Associated Molecular Pattern
MCP Monocyte Chemotactic Protein
MgSO4 Magnesium Sulphate
MLCK Myosin Light Chain Kinase
MMP Metalloproteinase
MRS de Man, Rogosa, and Sharpe media
NCS Newborn Calf Serum
NS Not significant
NSAID Non-Steroidal Anti-Inflammatory Drug
PAMP Pathogen-Associated Molecular Pattern
PBS Phosphate Buffered Saline
PBST Phosphate Buffered Saline + Tween
PG Prostaglandin
PGDH 15-Hydroxyprostaglandin Dehydrogenase
PGES Prostaglandin E Synthase
PGHS Prostaglandin endoperoxidase H Synthase
PR Progesterone Receptor

xi
PROM Premature Rupture Of the Membranes
ra receptor antagonist
RANTES Regulated on Activation, Normal T Cell Expressed and Secreted
RIPA RadioImmunoPrecipitation Assay
ROD Relative Optical Density
SEM Standard Error of the Mean
SNP Single Nucleotide Polymorphism
TLR Toll-Like Receptor
TNFα Tumor Necrosis Factor α

xii
List of Figures
Figure 1.1: The layers of the fetal membranes: the amnion and chorion ...................................... 8
Figure 1.2: Potential sites of bacterial infection amongst the intrauterine tissues. ...................... 21
Figure 1.3: Proposed pathway of infection and/or inflammation-mediated preterm birth .......... 33
Figure 1.4: Effects of stimulants on the prostaglandin production pathway in the amnion. ....... 38
Figure 3.1: Experimental design .................................................................................................. 60
Figure 3.2: The effect of treatments on amnion cell toxicity ....................................................... 61
Figure 3.3: Lipoteichoic and lipopolysaccharide dose optimization ........................................... 62
Figure 3.4: Time course responses of Interleukin-6 and Interluekin-8 ........................................ 63
Figure 3.5: Lactobacillus rhamnosus GR-1 dilution optimization .............................................. 64
Figure 3.6: Correlation between ELISA and bioplex assay methodologies ................................ 65
Figure 3.7: The effects of Lactobacillus rhamnosus GR-1 on Tumor Necrosis Factor - α
concentration in human amnion cells ........................................................................................... 66
Figure 3.8: The effects of Lactobacillus rhamnosus GR-1 on Interleukin-6 concentration in
human amnion cells ...................................................................................................................... 67
Figure 3.9: The effect of Lactobacillus rhamnosus GR-1 on chemokine concentrations in human
amnion cells .................................................................................................................................. 68
Figure 3.10: The effects of Lactobacillus rhamnosus GR-1 on immune cell activating cytokines
....................................................................................................................................................... 69
Figure 4.1: Experimental design .................................................................................................. 83
Figure 4.2: The effect of IL-1β, dexamethasone (Dex), lipopolysaccharide (LPS) and de Man,
Rogosa, and Sharpe (MRS) on prostaglandin production in human amnion cells ....................... 84

xiii
Figure 4.3: The effect of Lactobacillus rhamnosus GR-1 on Prostaglandin E2 concentration in
medium from cultured human amnion cells ................................................................................. 85
Figure 4.4: The effects of Lactobacillus rhamnosus GR-1 on PGHS-2 expression in human
amnion cells .................................................................................................................................. 86
Figure 4.5: The effect of Lactobacillus rhamnosus GR-1 on cytosolic PGES expression in
human amnion cells ...................................................................................................................... 87
Figure 4.6: The effect of Lactobacillus rhamnosus GR-1 on microsomal PGES1 expression in
human amnion cells ...................................................................................................................... 88
Figure 4.7: The effects of Lactobacillus rhamnosus GR-1 on microsomal PGES2 expression in
human amnion cells ...................................................................................................................... 89
Figure 4.8: Correlation between PGES expression and PGE2 production in human amnion cells
....................................................................................................................................................... 90
Figure 5.1: The effects of Lactobacillus rhamnosus GR-1 on inflammatory mediators in human
amnion cells ................................................................................................................................ 101

1
Chapter 1
General Introduction

2
Chapter 1
General Introduction
Preterm birth accounts for up to 8% and 13% of all births in Canada and the United States of
America respectively (Goldenberg et al., 2008; Public Health Agency of Canada, 2008). Thirty
percent of all preterm births are associated with infection and/or inflammation. Despite ongoing
studies of the mechanisms underlying infection and/or inflammation-mediated preterm birth, no
effective preventative treatment exists. Preterm birth results in neonatal morbidities that last into
adulthood, cost an estimated $26 billion in medical care annually in the United States of America
and place an emotional and psychological burden on families that is immeasurable by any
monetary means. Probiotics are increasingly being investigated as preventative therapy due to
their ability to modulate immune responses across many tissues and systems. However, the
effects of probiotics during pregnancy are largely uninvestigated. Therefore, the focus of this
thesis was to determine the possibility of Lactobacillus rhamnosus GR-1 as a potential
preventative treatment by measuring its effects on cytokines, chemokines and prostaglandins that
are known to be elevated in association with preterm delivery in human amnion cells.
1.1 Human Parturition
Human parturition occurs as the result of a series of feed-forward paracrine, autocrine and
endocrine events resulting in myometrial contractions, remodeling of the cervix and weakening
and rupture of the fetal membranes, ultimately leading to expulsion of the fetus from the uterus
(Challie et al., 2002). During pregnancy, the uterus and cervix work together to promote a
quiescent state in order to maintain gestation. Throughout gestation, the myometrium transitions
through several phases: quiescence, activation, stimulation and expulsion (Challis et al, 2002).
During these phases, multiple factors are recruited and interact to promote synchronous uterine
activity. These factors include uterine stretch due to fetal growth and biochemical factors
associated with an increase in the activation of the fetal hypothalamic-pituitary-adrenal (HPA)
axis that occurs in late gestation.
A quiescent state is maintained for approximately 95% of gestation by high levels of circulating
progesterone, locally produced prostacyclin, relaxin, nitric oxide and parathyroid hormone-
related protein that maintain myometrial inactivity (Challis et al., 2002). Many of these agents

3
inhibit muscle contractions by activating downstream pathways that increase cytosolic cAMP
and/or cGMP. Consequentially, increases in cAMP and/or cGMP cause intracellular calcium to
be sequestered and the inactivation of myosin light chain kinase (MLCK). Without the activity of
MLCK, myosin is not phosphorylated and the actin-myosin cross-bridges in the myometrial
smooth muscle remain in the relaxed state (Challis et al., 2000). Progesterone inhibits the
actions of estrogen by preventing expression of the estrogen receptor, (ER)-α to also maintain
quiescence (Mesiano et al., 2002).
As gestation progresses the uterine myometrium transitions to an active state. Throughout this
transition, uterine stretch caused by the growing fetus leads to up-regulation of contraction-
associated protein (CAP) genes (Challis et al, 2002). As a result, the myometrium becomes
capable of responding to uterotonins, or stimulants of myometrial contractility.
Uterotonins are molecules that alter the myometrial phenotype to an active state making it
capable of producing synchronous contractions. Oxytocin, prostaglandins and corticotropin-
releasing hormone (CRH) promote myometrial contractility and their concentrations rise
throughout gestation. As gestation progresses, the HPA axis of the fetus becomes increasingly
active resulting in increased adrenal cortisol production. Cortisol promotes the activity of
estrogen and prostaglandins while decreasing the sensitivity of the myometrium to progesterone.
The final phase of expulsion occurs as a result of powerful myometrial co-ordinated contractions
that are capable of expelling the fetus and placenta from the uterus.
As the myometrium progresses through different phenotypic states, other gestation tissues,
namely the cervix and the fetal membranes, undergo alterations in preparation for labour.
During pregnancy the cervix remains closed while the fetal membranes maintain the volume of
the amniotic fluid and provide mechanical strength necessary in pregnancy. As gestation
progresses, prostaglandins increase metalloproteinase (MMP) expression in the amnion. MMPs
degrade collagen molecules that constitute a large proportion of the extracellular matrix of both
the cervix and the fetal membranes. This results in remodeling of the cervix and weakening of
the fetal membranes (Menon et al., 2004; Yoshida et al., 2002).
Therefore, parturition results from the interaction of two interdependent pathways. The first is
mechanical stretch of the uterus and the other is biochemical factors and endocrine activities

4
associated with increased activation of the HPA axis of the fetus. In cases of membrane rupture
or infection, the amnion contributes a large pool of prostaglandins that may result in premature
myometrial activation, cervical ripening and membrane weakening.
1.1.1 Anatomy
1.1.1.1 Uterine Myometrium
The uterine myometrium is the strong, densely packed smooth muscle layer of the uterus that
gives much of the bulk to this organ. In the non-pregnant uterus, this layer contains blood and
lymphatic vessels as well as cholinergic, sympathetic and peptidergic nerve innervations. The
myocytes of the uterus, like other smooth muscle cells, are highly plastic and are capable of
transitioning through several phenotypes as gestation progresses. These include the proliferative,
synthetic and contractile stages leading to preparation for expulsion of the fetus during
parturition (Shynlova et al., 2009). Throughout pregnancy the control of the myometrial
phenotype switches from a global autonomic control system with the loss of cholinergic,
peptidergic and sympathetic nerves from the myometrium to one of humoral or local control by
prostaglandins, oxytocin and steroid hormones (Riemer et al., 1998).
The first phase of uterine growth during pregnancy is due to myometrial hyperplasia and is under
the control of estrogen and progesterone. Estrogen and progesterone simultaneously
hyperpolarize myometrial cells making them less responsive to contractile agonists (Riemer et
al., 1998; Shynlova et al., 2004; Shynlova et al., 2005). Myometrial quiescence is partially
maintained by high levels of oxytocinase and 15-hydroxyprostaglandin dehydrogenase (PGDH)
expression. These proteins metabolize oxytocin and prostaglandins, respectively (Mitchell et al.,
1995). Studies in pregnant rats have characterized the phenotypic changes that occur across
pregnancy (Shynlova et al., 2009). As uterine tension increases, stretch and progesterone act in
concert to induce growth and remodeling resulting in hypertrophy and matrix changes conducive
to the contractile phase of myometrial activity (Shynlova et al., 2009). The switch to the
contractile phenotype is characterized by increased expression of basement membrane proteins
such as fibronectin, laminin β2 and collagen IV within myometrial cells. Near the end of
gestation, uterine growth ceases while fetal growth continues, resulting in biomechanical
distension of the uterus. This distension is necessary, but not sufficient to cause up-regulation of
CAP gene expression as demonstrated by the lack of up-regulation of these genes in the absence

5
of progesterone withdrawal (Shynlova et al., 2009). Therefore, the phenotype of the uterus is
dependent upon both the tension of the myometrium and the endocrine environment that is
altered throughout pregnancy and at the onset of parturition.
1.1.1.2 Fetal Membranes/Amnion
The fetal membranes consist of the chorion and the amnion. The fetal membranes derive from
fetal origins and thus have the same chromosomal sex of the fetus. The amnion begins to form
eight days after conception from the ectodermal cell nest in the dorsal aspects of the zygote
(Calvin et al., 2007). The amniotic cavity rapidly increases in size as amniotic fluid begins to
surround the developing fetus. By 10-12 weeks of gestation, the amnion comes into contact with
the chorion forming the chorioamnion membrane. As the amniotic cavity further expands, the
fetal membranes contact the decidua along the uterine walls essentially sealing the endometrial
cavity by 16 weeks gestation (Calvin et al., 2007). Between the two fetal membranes, a layer of
phospholipids running parallel to the two tissues provides a surfactant-like effect to prevent
shearing of the membranes during movement. By the end of gestation, the fetal membranes
encompass an area of 1000-1200 cm2, resistant to the increased pressure placed upon them by
increased volumes of amniotic fluid and by fetal movements that become more vigorous as
gestation progresses (Bryant-Greenwood, 1998; Myatt et al., 2010).
The amnion has no blood supply or nerve innervation and as such relies on amniotic fluid and
the chorion for nutrient supply and waste transport (Parry et al., 1998). It is a largely an acellular
tissue in comparison to the chorion and provides the tensile strength and elasticity required to
withhold the amniotic fluid (Bryant-Greenwood, 1998). The amnion consists of two major
portions. The first is the placental amnion, defined as the portion of the amnion in direct contact
with the chorion of the placental chorion plate and the second portion consists of the remaining
amnion, termed the membranous amnion (Calvin et al., 2007).
The amnion consists of five layers starting from the visceral side or that in contact with the
amniotic fluid, fetus, and umbilical cord to the outer layer, or that in contact with the chorion: 1)
Epithelium, 2) Basement Membrane, 3) Compact Layer, 4) Fibroblast Layer, and 5) Spongy
Layer or Zona spongiosa (Figure 1.1, page 8; Bourne, 1962; Calvin et al., 2007; Parry et al.,
1998).

6
The epithelial layer of the amnion consists of cuboidal cells that are typically mononuclear with
dense granular cytoplasm with many vacuoles and few small mitochondria (Bourne, 1962). The
cells have a slight convex shape on the most apical surface in contact with the amniotic fluid and
the edges form a brush border of microvilli (Bourne, 1962). Many vacuoles are found in amnion
epithelial cells that are connected by fine channels close to adjacent cell membranes (Bourne,
1962). These tunnels and channels form an intricate network that allows amnion epithelial cells
to communicate with the extracellular matrix (Bourne, 1962). Additionally, both murine and
human amnion epithelial cells contain tight junctions (Kobayashi et al., 2009; Kobayashi et al.,
2010a). Amnion epithelial cells produce MMP-1, -2, -9, and collagens I, III, and IV, as well as
the non-collagenase proteins laminin, fibronectin and nidogen that compose the other amnion
layers (Aplin et al., 1985; Bourne, 1962; Parry et al., 1998). Epithelial cells attach to a basement
membrane composed of a network of collagen III, IV and V that act as a scaffold for the
assembly of laminin, fibronectin, heparin sulphate proteoglycan and nidogen (Bryant-
Greenwood, 1998; Parry et al., 1998). The epithelial cells attach to the basement membrane
through digitations of blunt processes from both the reticular network of the second layer, and
the first layer cells (Bourne, 1962).
The third layer of the amnion is the compact layer, a dense acellular network of reticular fibres.
It is composed of fibronectin, collagen I, III, and IV with low levels of collagens V, VI, and VII
(Bryant-Greenwood, 1998; Parry et al., 1998). Collagens I and II form large parallel bundles
that sustain mechanical integrity and give the amnion much of its tensile strength. The compact
layer is relatively permeable to chemotaxis of macrophages and rarely has antigen presenting
cells present unless they are actively phagocytic (Bourne, 1962).
The thickest layer of the amnion, the fibroblast layer is composed of the second major cell type
constituting the amnion, the mesenchymal cells. This layer is relatively weak compared to the
compact layer and is composed of a loose network of non-collagenous glycoproteins, reticulin,
collagens I, III, and IV, lamninin, nidogen, and fibronectin and scattered mesenchymal cells
(Bourne, 1962; Parry et al., 1998). This layer contains Hofbauer cells, macrophages of placental
origin where they exist as residential cells capable of phagocytosis (Bourne, 1962).
The outer most layer of the amnion, the spongy layer is in direct contact with the chorion. It is
composed of loose bundles of reticulin composed of collagens I, III, and IV as well as hydrated

7
proteoglycans covered by mucin (Bourne, 1962; Parry et al., 1998). This layer is prone to edema
as proteoglycans absorb water causing increases in the thickness of the amnion (Bourne, 1962).
The presence of mucin and the hydrophilic characteristics of proteoglycans allow the amnion to
move along the chorion while absorbing physical stress (Bryant-Greenwood, 1998). This layer
also contains resident Hofbauer cells (Bourne, 1962).
The amnion is responsive to many physiologic factors including cytokines, growth factors,
bacterial endotoxins, and glucocorticoids. It is equipped with receptors necessary to respond to
these factors including cytokine receptors such as CXCR2, bacterial endotoxins, namely toll-like
receptor (TLR) 2, TLR4, the family of prostaglandin receptors EP1, EP1, EP3, EP4, and F, as
well as the glucocorticoid receptor (Kallapur et al., 2009; Sun et al., 1996; Unlugedik et al.,
2010). Progesterone receptors are absent or expressed in very low levels in the amnion as
mRNA for both progesterone receptor (PR)-A and PR-B are undetectable (Merlino et al., 2009).
The amnion expresses natural antimicrobials and innate immune molecules that have anti-
bacterial, anti-viral, and anti-fungal activity. These include human α-defensins 1, 2 and 3,
human β-defensins 1, 2 and 3 as well as elafin and secretory leukocyte protease inhibitor (King
et al., 2007). As important components of the innate immune system, these molecules are part of
the immune defense against uterine infection during menstruation, pregnancy and labour (King
et al., 2007). The amnion produces pro-inflammatory and anti-inflammatory cytokines as well as
chemokines and steroids. However, a major product of the amnion is prostaglandin, in particular
prostaglandin (PG) E2 and to a lesser degree, PGF2α.
The amnion is in an ideal anatomical position to respond to many different molecules allowing
for signaling between the maternal and fetal compartments during pregnancy. Direct contact
between the amnion and the amniotic fluid provides not only a pathway for molecules secreted
from the fetus, but also foreign bacterial products during microbial invasion of the amniotic
cavity to influence the release of cytokines, chemokines and prostaglandins from the epithelial
layer of the amnion. In addition, the amnion relies on the chorion for nutrients and waste
elimination. Consequentially, ascending bacteria and bacterial products may influence the
mesenchymal cells of the amnion in close proximity to the chorion. Therefore, an intimate
relationship between the amnion, fetal environment and chorion exists which differentially
dictate the response and role of the amnion throughout gestation and parturition.

8
Figure 1.1: The layers of the fetal membranes: the amnion and chorion
The inner layer of the fetal membranes, the amnion is an avascular tissue composed of five layers: 1)
Epithelium, 2) Basement membrane, 3) Compact Layer, 4) Fibroblast Layer, 5) Spongy Layer of Zona
Spongiosa.
[The image has been modified from American Journal of Obstetrics and Gynecology, Vol 79. Bourne
GL. Microscopic anatomy of human amnion and chorion, 1070-1073. Copyright Elsevier (1960) and
appears here with the permission of the journal.]
2 1
3
4
5

9
1.1.2 Mechanisms of Human Parturition
1.1.2.1 Uterine Stretch
Stretch induces contractions in many muscle systems including the myocardium and the
myometrium (Riemer et al., 1998). Ultimately, the ability of the uterine myometrium to contract
is regulated by the availability of intracellular calcium. The myometrium consists of smooth
muscle cells that contain actin-myosin bridges. As intracellular calcium levels increase, MLCK
is activated resulting in increased adenosine triphosphatase activity of the myosin head which
becomes phosphorylated and the muscle contracts. Agents that decrease intracellular calcium
levels and/or increase cyclic adenosine monophosphate (cAMP) or cyclic guanosine
monophosphate (cGMP) levels that sequester intracellular calcium, result in relaxation of the
uterine myometrium. In contrast, agents that increase intracellular calcium result in uterine
myometrial contraction.
As gestation progresses, the growth of the fetus causes biomechanical distension of the uterus
leading to both hypertrophy and hyperplasia of the myometrial smooth muscle cells in the
presence of progesterone (Challis et al., 2005). Throughout gestation, circulating progesterone
maintains uterine quiescence through nuclear PR-B expressed on both the decidua and the nuclei
of myometrial cells (Merlino et al., 2007). Progesterone maintains quiescence by restricting
transcription of CAPs genes which translate into proteins involved in myometrial stimulation and
include the oxytocin receptor, progesterone receptors, the gap junction protein Cx43, increasing
expression of PGDH, an enzyme responsible for degradation of PGE2 in the chorion and
myometrium and inducing phosphokinase A signaling that promotes smooth muscle relaxation
by inhibiting phospholipase C activity (Merlino et al., 2007). Unlike other mammalian species,
circulating progesterone levels in the human do not decrease and estrogen levels do not increase
abruptly near the end of gestation (Challis et al., 2001). However, a functional withdrawal of
progesterone accompanied by functional estrogen activation occurs at term. This results in a
conversion of the myometrial phenotype from quiescence to activation. Throughout gestation,
PR-A expression increases. PR-A resides in the nucleus of myometrial cells and thus is ideally
located to inhibit the expression of PR-B (Merlino et al., 2007). As a result, the effects of PR-B
that maintain uterine quiescence until late gestation are withdrawn, including the inhibitory
effects on estrogen receptor (ER)-α expression (Merlino et al., 2007; Mesiano et al., 2002).

10
Circulating estrogen subsequently increases uterine contractility through the up-regulation of
CAP genes as the inhibitory actions of progesterone are functionally withdrawn (Kamel, 2010;
Mesiano et al., 2002). Estrogen, in conjunction with mechanical distension works in concert to
activate the myometrium, thus allowing for high amplitude, high-frequency contractions to occur
in the next phenotypic state of the myometrium.
Up-regulation of CAP genes results in increased expression of three major groups of proteins
necessary for activation and subsequent contraction of the myometrium: 1) Ion channels, 2) Gap
junction proteins, and 3) Contractile agonist receptors (Challis et al., 2000; Challis et al., 2002).
Ion channels determine the resting membrane potential of myocytes and thus determine the
contractile response to contractile agonists. Gap junction proteins, specifically C43, synchronize
longitudinal co-ordinated contractions over the entire uterine myometrium during labor (Merlino
et al., 2007; Riemer et al., 1998). Contractile agonist receptors, including the oxytocin receptor
as well as the prostaglandin receptors F, EP1 and EP3 increase the sensitivity of the myometrium
to oxytocin and prostaglandins that act as uterotonins during myometrial activation. Stretch also
causes increases in myometrial prostaglandin H synthase (PGHS)-2 expression (Riemer et al.,
1998). Experiments in unilateral pregnant rats demonstrate the inability of experimentally
induced stretch to increase the expression of CAP genes in the non-gravid horn, unlike results in
the gravid horn (Ou et al., 1998). This suggests that CAP gene expression is dependent on
stretch as well as the endocrine environment, which is largely governed by the fetal genes and
activation of the HPA axis (Shynlova et al., 2009).
1.1.2.2 Hypothalamic-pituitary-adrenal Axis
A key mechanism for parturition is the endocrine action of increased glucocorticoids and their
influence on the gestational tissues. Across many species, including humans, the fetal HPA axis
matures near the end of gestation. Glucocorticoids are vital in the maturation of organs
necessary for survival outside of the womb and also play a crucial role in determining the length
of gestation (Smith et al., 2007). Experiments in sheep have characterized the mechanisms in the
fetal brain that account for an increase in HPA activity. Initially, CRH mRNA levels increase in
the paraventricular nucleus of the hypothalamus as adrenocorticotrophic hormone (ACTH)
concentrations in the fetal circulation rise (Challis et al., 2000). The fetal sheep adrenal gland
increases expression of enzymes involved in cortisol production as ACTH receptor expression is

11
up-regulated (Holloway et al., 2000). In this species, the net result of the maturing fetal HPA
axis is increased sensitivity of the fetal adrenal gland to ACTH, and greater production of
cortisol (Challis et al., 2001). In primates, similar increases in cortisol occur in late gestation. In
addition to increased adrenal cortisol output in the human fetus, glucocorticoids influence human
gestational tissue (Karalis et al., 1996). In the human placenta, prostaglandins down-regulate the
expression of 11β-hydroxysteroid dehydrogenase (HSD) 2 and increase the expression of 11 β -
HSD1 thereby decreasing conversion of cortisol to cortisone (Alfaidy et al., 2001). Similarly, in
the fetal membranes cortisone is produced through the activity of 11β-HSD1 (Sun et al., 2003;
Myatt et al., 2010). Near the end of term maternal blood concentrations of CRH increase and
CRH-binding protein levels decrease. Altogether, the influence of CRH and glucocorticoids
increase near the end of term and activate downstream cascades that lead to parturition onset.
This activation includes a feed-forward loop initiated by increased glucocorticoid production and
results in increased prostaglandins.
Elevated CRH levels stimulate prostaglandin production by modulating the levels of
prostaglandin metabolizing enzymes expressed in the amnion, chorion and placenta trophoblast
cells. PGHS-2 expression in the fetal membranes and placenta is increased by CRH (Alvi et al.,
1999; Challis et al., 2002). Simultaneously, CRH and cortisol decrease the expression of PGDH
in chorionic trophoblast cells. As a result, prostaglandins act on the fetal membranes to increase
the activity of 11β–HSD1 in chorionic trophoblasts and decrease expression of 11β-HSD2 in
placental trophoblasts (Alfaidy et al., 2001; Challis et al., 2001). The activity of 11β-HSD1
converts inactive cortisone to cortisol whereas activity of 11β-HSD2 inactivates cortisol to
cortisone. Elevated CRH levels promote different effects based on the regionalization of the
cortisol receptor subtypes throughout the gestational tissues (Hillhouse et al., 2002). In the
fundal region, CRH acts through CRH receptor subtype 1 to increase prostaglandin production
through increased PGHS-2 and decreased PGDH (Challis et al., 2000). As a result of varying
receptor expression, the fundal region of the uterus relaxes allowing for regionalization of
activity at term (Cong et al., 2009; Stevens et al., 1998). As well, the surge of CRH in late
pregnancy is associated with increased estrogen resulting in altered progesterone to estrogen
ratios indicative of a role for CRH in myometrial activation (Smith et al., 2009). Elevated
prostaglandins induce expression of MMP-1, 2, and 9 in the amnion that begin to degrade the
collagen present in the epithelium, compact, fibroblast and spongy layers that constitute the

12
tissue. Therefore, increased glucocorticoid levels result in a positive feed-forward loop between
cortisol, CRH and prostaglandins that is necessary for activation of the myometrium, cervical
ripening and membrane weakening.
1.2 Preterm Birth
Both autocrine and paracrine signals mediated by uterine stretch and activation of the HPA axis
interact to promote phenotypic changes in the gestational tissues leading to the onset of labour at
term. However, the phenotypic state of the gestational tissues can be altered earlier in gestation
if similar autocrine and paracrine signals are up-regulated. During infection and/or inflammation
cytokines and prostaglandins levels become elevated that subsequently activate pathways
involved in modifying the phenotype of the myometrium, cervix and fetal membranes.
Ultimately, the ability of these signals to modify and activate downstream pathways indicates a
potential mechanism leading to preterm delivery and highlights potential molecules to be
targeted in preventative treatment of infection and/or inflammation-mediated preterm birth.
1.2.1 Epidemiology
Preterm birth is a major obstetrical concern that occurs in 8% and 13% of all pregnancies in
Canada and the United States of America, respectively (Goldenberg et al., 2008; Public Health
Agency of Canada, 2008). Defined as labour occurring between 20 and 37 weeks gestation,
preterm birth accounts for 75-85% of all neonatal morbidities and mortalities (Challis et al.,
2002). Many of these morbidities, including pulmonary disorders, blindness, deafness, cerebral
palsy, and neurodevelopmental delays linger into adulthood causing poor health later in life. The
cost of caring for preterm infants has a large economic impact on the healthcare system. In 2005,
it was estimated that $26.2 billion in the United States of America was spent on medical care for
preterm neonates (Institute of Medicine Report, 2006). An impact that cannot be measured
through monetary means is the emotional and psychological burden placed upon the families
caring for infants born prematurely.

13
1.2.2 Etiology
The incidence of preterm birth has been on the rise since the 1980s (Challis et al., 2002). Much
of this trend is accounted for by increased use of reproductive assisted techniques and the
associated rise in multi-fetal pregnancies. Other predisposing factors include advanced or young
maternal age, black race, low socioeconomic status of either or both the father and mother, single
parenthood, type of employment, smoking, alcohol and substance abuse as well as carrying a
male fetus (Goldenberg et al., 2003; Meis et al., 1995; Robinson et al., 2001). Certain aspects of
the mother’s medical history also increase the risk of preterm birth. Such medical conditions
include periodontal disease, lung disease, chronic hypertension, proteinuria or bacteriuria, a
history of a previous preterm delivery, uterine malformation or short cervix. Reproductive tract
infection, pelvic infection, vaginal microbiota alterations and infections including bacterial
vaginosis, Neisseria gonorrhoeae, Chlamydia trachomatis, group B Streptococcus, Ureaplasma
urealyticum, and Trichomonas vaginalis as well as obstetrical complications during pregnancy
such as early pregnancy bleeding, placental abruption, placenta praevia, hydramnios,
preeclampsia, premature rupture of the membranes (PROM), cervical surgery or a previously
induced abortion are all associated with increased preterm birth (Goldenberg et al., 1998;
Goldenberg et al., 2003; Meis et al., 1995; Norwitz et al., 1999; Robinson et al., 2001; Romero et
al., 2002). Preterm birth can be categorized into three groups: 1) Idiopathic (50%), 2) Indicated ,
those induced by a physician as more deleterious to prolong to either the health of the fetus or the
mother due to medical complications (20-30%), and 3) Infection and/or inflammation-mediated
(30%). Early preterm deliveries tend to be associated more so with the presence of histologic
chorioamnionitis and inflammation compared to later preterm deliveries (Goldenberg et al.,
2000; Goldenberg et al., 2003; Vogel et al., 2005). However, despite ongoing research, the
etiology of spontaneous preterm birth remains largely unknown.
1.2.3 Current Diagnostic Strategies and Treatments for Preterm Birth
The diagnosis and prevention of preterm birth remains problematic despite ongoing research.
Commonly, pregnant women are informed by their physician of how to recognize signs of labour
including contractions, pelvic pressure, vaginal discharge and back pain (Denney et al., 2008).
However, only one third of women showing signs of preterm labour give birth within 24–48

14
hours (Bocking et al., 1999; Katz et al., 1999). Despite ongoing research, the onset of labour is
only delayed up to 72 hours using tocolytic therapy. This allows for glucocorticoid treatment to
mature fetal lungs; however the other developmental consequences associated with preterm birth
cannot be corrected within this relatively short prolongation of gestation. Therefore, it is
important to develop effective preventative therapy.
Assessment of a woman’s medical history plays an important role in determining the individual
risk she possesses of undergoing preterm delivery. A history of a previous spontaneous preterm
birth is one of the highest predictive indicators of an increased risk of preterm birth (Goldenberg
et al., 2003). The preterm prediction study by the Maternal Fetal Medicine Network, which
included 3000 pregnancies, indicated the three most effective predictive assessments for preterm
delivery: 1) A positive cervical or vaginal fluid fetal fibronectin test, 2) A cervical length of less
than or equal to 25 mm, and 3) Elevated serum levels of α-fetoprotein, alkaline phosphatase and
granulocyte-colony stimulating factor (G-CSF; Goldenberg et al., 2003). As well, amniotic fluid
samples with elevated levels of interleukin (IL)-6, IL-8, or Ureaplasma urealyticum are
associated with high levels of preterm delivery (Vogel et al., 2005). Within maternal serum or
vaginal and cervical fluids many molecules have been identified as potential predictive markers
of preterm birth. These include low levels of ferritin, folate, zinc and high levels of C-reactive
protein, cytokines, MMPs as well as altered levels of activin, relaxin, collagenase, non-
phosphorylated insulin-like growth factor and CRH (Andrews et al., 2003; Norwitz et al., 1999;
Vogel et al., 2005). Recently, single nucleotide polymorphisms (SNPs) in the promoter region of
cytokines that up-regulate their transcription levels have been correlated with an increased risk of
preterm birth (Harper et al., 2011).
It is important for physicians to determine whether or not prolonging gestation may be more
detrimental to the fetus in a hostile intrauterine environment exposed to hypoxia or infection
and/or to the mother as is the case in preeclampsia. In these cases, preterm delivery may be the
most beneficial option compared to allowing gestation to progress. Tocolytics are agents that
ameliorate uterine contractions and thus prevent expulsion of the fetus from the uterine
environment for 24-72 hours (Norwitz et al., 1999). Ethanol was the first effective tocolytic
identified (Fuchs et al., 1981). However, due to severe fetal side effects, its use was abandoned.
Four routine tocolytic treatments are used in the United States of America: 1) β-mimetics, 2)

15
Magnesium sulphate, 3) Non-steroidal anti-inflammatory drugs, and 4) Calcium channel
blockers (Giles et al., 2007; Katz et al., 1999; Meis et al., 2003; Vercauteren et al., 2009).
However, like early tocolytic drugs, many of these options are associated with adverse side
effects, including myocardial ischemia, tachycardia and arrhythmias as well as severe
consequences to the health of the fetus including intraventricular hemorrhage and premature
closure of the ductus arteriosus (Giles et al., 2007; Katz et al., 1999; Vercauteren et al., 2009).
Therefore, the benefits of tocolytic drugs have been questioned and their use abandoned in many
cases for the prevention of preterm delivery.
After the diagnosis of a pregnancy at high risk of undergoing preterm delivery, it is essential to
provide effective preventative treatment if prolongation of gestation is not deleterious to the
health of the fetus or the mother. However, currently no effective preventative treatment exists.
A common method once used to prevent preterm birth was the use of antibiotics. Although
bacterial vaginosis and Trichomonas vaginalis are largely asymptomatic, these alterations to the
healthy vaginal microbiota are associated with an increased risk of preterm birth (Romero et al.,
2002). When these conditions are diagnosed, antibiotics such as metronidazole, erythromycin,
clindamycin and/or ampicillin, are administered in an attempt to replenish the altered microbiota
by eliminating pathogenic bacteria. However, numerous meta-analyses, cohort studies, and
randomized trials indicate that antibiotic treatment is ineffective at decreasing the risk of preterm
birth (Andrews et al., 2003; Carey et al., 2000; Kenyon et al., 2001; Leitich et al., 2003;
McDonald et al., 2007; Okun et al., 2005). In some cases, the use of metronidazole treatment
actually increases the risk of preterm birth (Andrews et al., 2003; Okun et al., 2005). Antibiotics
may be ineffective at decreasing the risk of preterm birth for several reasons: 1) Bacteria may
have already ascended into the upper genital tract, 2) Antibiotics are unable to eradicate biofilms,
3) Antibiotics are unable to inactivate sialidases and, 4) Antibiotics can eradicate bacteria that
are necessary for host defense mechanisms (Reid et al., 2003). Also, prolonged use of antibiotics
has led to increasingly resistant bacterial strains and promotes an alkaline environment in the
vagina that encourages pathogenic bacterial growth (Locksmith et al., 2001).
1.3 Infection and Inflammation
Infection and/or inflammation accounts for 30% of preterm births. Inflammation acts as a first
line of defense against pathogenic threats by recruiting and activating immune cells while

16
forming a physical barrier to stop the spread of infection. In many cases, the signs and
symptoms of infection and/or inflammation during pregnancy are undetectable unless invasive
procedures such as amniocentesis are performed. The prevalence of chorioamnionitis, an
inflammation of the chorioamnion membranes has been found in up to 50% of pregnancies
ending prematurely. Many of the organisms found in the upper genital tract are of vaginal origin
and bacterial vaginosis, an alteration to the vaginal microbiota where lactobacilli levels decrease,
increases the risk of preterm delivery by 1.4 to 3.8 fold (Andrews et al., 2000). Treatment of
bacterial vaginosis with antibiotics is associated with a high incidence of re-occurrence.
Consequentially, current research has shifted to determine the use of candidate probiotic strains
of bacteria including lactobacilli, to reinstate a healthy vaginal microbiota. Many Lactobacillus
strains exert immunomodulatory effects across various systems; however the role Lactobacillus
strains may exert on the inflammatory processes in the gestational tissues that are associated with
infection and/or inflammation preterm delivery remain largely uninvestigated.
1.3.1 Vaginal Microbiota
The fetal vagina is a sterile environment which is first colonized by bacteria endogenous to the
mother’s vaginal birth canal, the skin of caretakers and the infant’s faeces upon birth (Reid et al.,
2011; Spiegel, 1991). It is not until after menarche, when estrogen levels increase that the
childhood vaginal microbiota characterized by intestinal and cutaneous bacterial species
becomes dominated by Lactobacillus species, the hallmark species of a healthy adult vaginal
microbiota (Forsum et al., 2005; Spiegel, 1991). Estrogen levels at puberty promotes the
production of glycogen in the vaginal epithelial cells necessary for the survival of lactobacilli as
it provides a source of glucose through fermentation (Forsum et al., 2005; Spiegel, 1991). The
bi-product of fermentative metabolism, lactic acid creates an acidic vaginal environment with a
pH of 3.8-4.2 which promotes self-growth of Lactobacillus species within a healthy microbiota.
Three or four species, including Lactobacillus crispatus, Lactobacillus iners, Lactobacillus
gasseri, and Lactobacillus jensenii are present in much higher levels in the vaginal microbiota
(Antonio et al., 1999; Burton et al., 2002; Forsum et al., 2005; Lamont et al., 2011; Pavlova et
al., 2002; Song et al., 1999; Vasquez et al., 2002; Yamamoto et al., 2009). As well, over twenty
species of Lactobacillus, including Lactobacillus acidophilus, Lactobacillus fermentum,
Lactobacillus plantarum, Lactobacillus oris, Lactobacillus ruminis, Lactobacillus reuteri, and

17
Lactobacillus rhamnosus have been isolated from the vaginal microbiota (Antonio et al., 1999;
Forsum et al., 2005; Pavlova et al., 2002). Gram-negative bacteria, including Streptococcus
species, Atopobioum vaginae, Prevotella spp. and Gardnerella vaginalis are present in low
levels in healthy vaginal microbiota (Yamamoto et al., 2009). Using 16S RNA gene sequencing
analysis, Gardnerella vaginalis was identified as a core component in the vaginal microbiota of
the entire female population sampled (Hummelen et al., 2010). Therefore, many bacteria that
may become pathogenic if levels are not maintained are present in healthy vaginal flora at
detectable levels.
The vaginal microbiota provides a barrier to exterior influences and actively participates in
adaptive immunity, the portion of the immune system that is specific and non-inherited (Forsum
et al., 2005). However, the vaginal microbiota is susceptible to alterations resulting in bacterial
vaginosis, Vulvovaginal candidiasis, and Trichomonas vaginalis. Accordingly, women with
altered vaginal microbiota are more susceptible to the transmission of sexually transmitted
diseases including HIV and gonorrhoea, pelvic inflammatory disease, urinary tract infections,
post-operative infections, and upper genital tract infections including chorioamnionitis (Antonio
et al., 1999; Deb et al., 2004; Forsum et al., 2005). Pathogenic bacteria are equipped with
molecules, including proteases, mucinases, and sialidases that promote self-growth through
degradation of the cervicovaginal mucous lining. Beyond the vaginal environment, bacterial
collagenases, phospholipases and endotoxins directly or through stimulated production of
downstream molecules result in remodeling of the collagen structuring of the fetal membranes
and cervix and initiate inflammatory cascades in the gestational tissues of the intrauterine
environment (Goldenberg et al., 2000).
1.3.2 Routes of Intrauterine Infection
Microbial invasion of the amniotic cavity is associated with adverse outcomes including PROM
and preterm birth. Intrauterine infection may result from four different routes of bacterial access:
1) Retrograde migration from the abdominal cavity through the fallopian tubes, 2)
Haematogenous spread from the maternal blood through the placenta, 3) Iatrogenic introduction
through invasive procedures, such as amniocentesis, and 4) Ascent from the vagina and cervix
through the fetal membranes (Figure 1.2, page 21; Goldenberg et al., 2000). The latter of these, is
thought to be the most common route for bacteria to reach the uterine cavity. Evidence for this

18
includes the fact that histologic chorioamnionitis is more severe at the location of membrane
rupture than other sites including the chorionic plate or the site of umbilical cord attachment
(Benirschke, 1960; Romero et al., 1989d). As well, microbes found in congenital infections are
similar to those isolated from the maternal vagina (Benirschke, 1960). The most common
microorganisms found in amniotic fluid samples are Ureaplasma urealyticum, Gardnerella
vaginalis, Bacteroides spp. and Mycoplasma hominis (Goldenberg et al., 2000). The
morphotypes of these bacteria are used by the Nugent scoring system to diagnose bacterial
vaginosis (Goldenberg et al., 2000; Nugent et al., 1991). During twin gestations, twins are
commonly oriented one over the other, such that the membranes of only one twin comes come in
contact with the cervical os, while the other twin is located superior to this twin. Histologic
chorioamnionitis is more frequent in the fetal membranes directly over the cervix. Bacteria
present in these membranes are commonly of vaginal origin indicative of their ascent from the
lower genital tract (Benirschke, 1960). Together, this evidence indicates the most common route
of infection of the intrauterine environment is ascending from the lower genital tract.
1.3.3 Bacterial Vaginosis
1.3.3.1 Epidemiology
Bacterial vaginosis accounts for approximately 80% of all diagnosed vaginal microbiota
alterations and is present in 15% - 20% of pregnant women (Hillier et al., 1995; Imseis et al.,
1997; Romero et al., 2004). Bacterial vaginosis is associated with many factors including: single
marital status, African American ethnicity, low socioeconomic status, previous delivery of a low-
birth-weight infant, douching practices, spermicide and antimicrobial use, smoking, absence of
barrier birth control, and sexual activity (Hawes et al., 1996; Hillier et al., 1995; Reid, 2008).
Bacterial vaginosis increases the incidence of pelvic inflammatory disease, post-operative
infections, post-cesarean endometritis, PROM, histologic chorioamnionitis, and subsequent
preterm delivery as the infection may reach the intra-uterine environment (Hawes et al., 1996;
Imseis et al., 1997; MacPhee et al., 2010; Nugent et al., 1991; Romero et al., 2004; Soper, 1993;
Sweet, 1995).
1.3.3.2 Etiology
Bacterial vaginosis is an alteration to the endogenous vaginal microbiota where levels of
Lactobacillus species decrease and anaerobic bacteria begin to dominate. It is unknown if the

19
decrease of lactobacilli precedes the growth of pathogenic bacteria or overgrowth of harmful
bacteria causes a decrease in Lactobacillus; however, the total concentration of bacteria in the
vagina increases by 100 to 1000 times that of the healthy microbiota (Forsum et al., 2005).
Characteristic bacteria that dominate the microbiota in bacterial vaginosis include Gardnerella
vaginalis, Bacteriodes spp., Mycoplasma hominis, Mobiluncus spp., Prevotella spp., and
Atopobium vaginae; many of which reside in low levels in a healthy vaginal microbiota (Falagas
et al., 2007; Hummelen et al., 2010; Nugent et al., 1991). The microbial profiles of women with
bacterial vaginosis show large inter-patient variability and greater diversity when compared to
profiles of women with a healthy vaginal microbiota. In agreement, no singular microbe has been
associated with the etiology of bacterial vaginosis and diagnosis is dependent on symptoms and
signs as well as a proportional analysis of bacterial species present in characteristically healthy
versus altered vaginal microbiota.
1.3.4 Current Diagnostic Strategies and Treatments for Bacterial Vaginosis
Currently, the diagnosis of bacterial vaginosis combines both clinical syndrome observation with
Nugent scoring according to the following four hallmark indicators: 1) A vaginal pH greater than
4.5, 2) An amine fishy odour when vaginal fluid is mixed with potassium chloride, 3) The
presence of clue cells - vaginal epithelial cells covered in Gardnerella vaginalis making them
appear rough edged under a microscope in a vaginal fluid sample, and 4) A Nugent score greater
than 6. Nugent scoring is based on a weighted representation of the bacterial species visualized
by gram-staining that summate to a score from 0 to 10. Bacteria scored include: Lactobacillus
(gram-positive rods), Gardnerella vaginalis (small gram-variable rods), Bacteroides spp. (small
gram-negative rods), and Mycoplasma spp. (curved gram-variable rods; Nugent et al., 1991).
The values 0-3 represent a normal microbiota, with high proportions of Lactobacillus spp. and
few of the other morphotypes; 4-6 represents an intermediate microbiota with higher proportions
of pathogenic morphotypes, and a score of 7-10 represents bacterial vaginosis, with low
lactobacilli morphotypes present. Inaccuracies in diagnosis persist due to the lack of symptoms
in up to an estimated 50% of women with bacterial vaginosis and the acuity required to identify
associated symptoms (Nugent et al., 1991). Other diagnostic techniques once used for bacterial
vaginosis include laboratory cultures positive for Gardnerella vaginalis growth, gas
chromatography for products of pathogenic vaginal bacteria and a proline amniopeptidase test

20
(Nugent et al., 1991). However, finding Gardnerella vaginalis is no longer deemed a definitive
diagnosis. Gram-staining is the most inexpensive, efficient and reproducible; therefore, gram-
staining and Nugent scoring is the method of choice for bacterial vaginosis diagnosis.
As bacterial vaginosis is a disruption to the vaginal eubiosis, treatment has largely been focused
on the use of antibiotics, administered both orally and vaginally. Despite evidence of cure after
initial treatment, a high reoccurrence rate exists in many patients regardless of the route and/or
type of antibiotic administration. Repetitive antibiotic use is avoided as it may encourage the
development of resistant bacterial strains. In addition, antibiotic treatment of bacterial vaginosis
in pregnant women has not shown any decrease in the incidence of preterm birth.
The association of bacterial vaginosis with preterm delivery has led to the study of adjunctive
probiotic and antibacterial treatment, or probiotic supplementation independently. Despite
relatively low endogenous levels in the vaginal microbiota, Lactobacillus rhamnosus GR-1 and
Lactobacillus reuteri RC-14 are the most effective at restoring and maintaining a normal vaginal
microbiota (Reid et al., 2011). These two strains of lactobacilli persist in the vagina after vaginal
insertion and also alter the host immune defenses (Cadieux et al., 2002; Gardiner et al., 2002).
Lactobacillus rhamnosus GR-1 in particular can populate the vagina, reduce pathogenic bacteria
from reaching the vagina from the anal-rectal area, inhibit growth, adhesion and biofilm
formation of gram-negative bacteria as well as inhibit the persistence of Candida albicans
(Cadieux et al., 2002; Reid, et al., 2003; Reid, 2008).
Bacteria associated with bacterial vaginosis are capable of releasing proteases and endotoxins
that free arachidonic acid from phospholipid membranes and subsequently increase
prostaglandin production in the amnion, chorion and decidua. Increased prostaglandins elevate
the risk of preterm labour by inducing collagen remodeling in the fetal membranes. Antibiotic
treatment is ineffective at preventing preterm birth. This may be a consequence of the inability of
antibiotics to alter downstream pathways that have been activated prior to their administration.
There is little research on the role lactobacilli may play at preventing the production of cytokines
and prostaglandins in the amnion, a major contributor to the amniotic fluid cytokine pool and
prostaglandins. However, due to the known immunoregulatory and other beneficial roles of
Lactobacillus species, it is possible that these bacteria may act as a probiotic to decrease the risk
of preterm birth associated with infection and/or inflammation.

21
Figure 1.2: Potential sites of bacterial infection amongst the intrauterine tissues.
Potential routes for microbes to gain access to the intrauterine environment: 1) Retrograde
migration from the abdominal cavity through the fallopian tubes, 2) Haematogenous spread from
the maternal blood through the placenta, 3) Iatrogenic introduction through invasive procedures,
such as amniocentesis, and 4) Ascent from the vagina and cervix through the fetal membranes.
[The image is modified from New England Journal of Medicine. Vol 342.20. Goldenberg RL,
Hauth JC, Andrews WW. Intrauterine infection and preterm delivery. 1500-1507 (2000) and
appears here with the permission of the journal.]
1
2
3
4 4

22
1.4 Probiotics
Probiotics are defined as, “live microorganisms which when administered in adequate amounts
confer a health benefit on the host” (FAO and WHO, 2001). Probiotics have been used
therapeutically and have shown beneficial effects across a wide spectrum of medical conditions.
These effects include modulation of immunity, lowering cholesterol, treatment of rheumatoid
arthritis, prevention of cancer, improvement of lactose intolerance, prevention of diarrhea and
constipation, reduction or prevention of acute dermatitis and treatment of urinary tract infections
(Kligler et al., 2007; Lee do et al., 2011; Ohara et al., 2010; Ojetti et al., 2010; Reid, 1999; So et
al., 2011; Verna et al., 2010). Although the use of lactobacilli as probiotics have shown promise
in restoring vaginal microbiota for the treatment of vaginal infections and microbiota alterations
during pregnancy, there are currently insufficient studies to assess the impact probiotics have on
pregnancies at risk of undergoing preterm birth associated with infection and/or inflammation
(Othman et al., 2007).
Although the exact mechanism by which probiotics exert their beneficial effects remains
unknown, various types of probiotic bacteria are known to benefit their host through competitive
exclusion of bacterial adherence, bacteriocin and butyrate production, maintenance of the
epithelial barrier, modulation of immune responses through NFΚB transcriptional activity
modification and immune cell recruitment and activation as well as increased IgA production
(Ahn et al., 1990; Falagas et al., 2007; Floch, 2010; Jijon et al., 2004; Lara-Villoslada et al.,
2007; Madsen et al., 2001; Moorthy et al., 2009; Reid, 2008; Yan et al., 2002). Pathogenic
bacteria possess pathogen-associated molecular patterns (PAMPs) on their cellular membrane
that are recognized by the body’s immune system through activation of TLRs. Unlike probiotic
bacteria that instead harbor microbe-associated molecular patterns (MAMPs), recognition of
PAMPs initiate an immune response to defend against pathogenic bacterial threats (Servin,
2004). Additionally, the signaling pathway activated by TLR9 has been shown to be necessary
for immunostimulation by bacterial CpG DNA sequences that play a role in the anti-
inflammatory effects exerted by various probiotics (Rachmilewitz et al., 2004). Therefore, an
opportunity exists for probiotics and/or their byproducts to be used as treatment for many
infectious and autoimmune conditions.

23
1.4.1 Lactobacillus Species as Candidate Probiotics for use During Pregnancy.
The criteria for defining a probiotic candidate has been established including: 1) Of no threat to
the host, therefore they are non-carcinogenic, non-pathogenic, and non-invasive, 2) Able to
persist and multiply, 3) Resistant to vaginal microbicides, 4) Capable of co-aggregation and
forming a normal eubiosis, 5) Capable of adherence to cells, 6) Capable of exclusion or
reduction of pathogenic adherence, and 7) Capable of antagonistic growth through the
production of acids, hydrogen peroxide and bacteriocins (Reid, 1999). In addition, when
identifying a potential probiotic strain of bacteria, it is important to consider the commensal
bacterial microbiota as well as the intrinsic resistance of the probiotic to antibiotics (Servin,
2004). In the case of the vaginal microbiota, loss of the endogenous dominant species of a
healthy vagina, lactobacilli, is associated with pathologies including bacterial vaginosis.
Bacterial vaginosis in pregnant women increases risk of preterm delivery; therefore, a strain of
Lactobacillus may exert beneficial effects during pregnancy. In particular, Lactobacillus
rhamnosus GR-1 possesses many of the characteristics required to define it as a probiotic
bacteria for use in the urogenital tract.
1.4.1.1 Safety of lactobacilli administration
Lactobacillus species are safe for use in humans with low intrinsic immunogenicity as they are
neither carcinogenic nor pathogenic (Cadieux et al., 2002; Servin, 2004). In fact, Lactobacillus
rhamnosus GR-1 injected directly into the bladder of human patients failed to induce an infection
(Hagberg et al., 1989). In addition, a meta-analysis of randomized controlled trials involving
lactobacilli and bifidobacterium during pregnancy indicated that either bacterium do not alter the
incidence of caesarean section, birth weight, malformations, or gestational age at birth in
pregnant women (Dugoua et al., 2009). Therefore, the safety of Lactobacilli rhamnosus GR-1 in
humans is established and has a low or negligible chance of threatening the health of a mother
and her fetus throughout pregnancy.
1.4.1.2 Ability of lactobacilli to colonize the vagina
Studies have shown that after instillation, Lactobacillus rhamnosus GR-1 and Lactobacillus
reuteri RC-14 persist in the vaginal environment for 19 days compared to only 5 days for
Lactobacillus rhamnosus GG (Cadieux et al., 2002). Another study showed Lactobacillus

24
rhamnosus GR-1 present in the vaginal microbiota up to 7 weeks after instillation (Reid et al.,
1994). Importantly, Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri RC-14 are capable
of restoring and maintaining a healthy vaginal microbiota (Reid et al., 2001a, Reid et al., 2003).
Passive forces including electrostatic interactions, hydrophobic steric forces and steric
hinderence via lipoteichoic acid (LTA) allow Lactobacillus to interact with the mucosal layer of
urogenital cells (Servin, 2004). Lactobacillus rhamnosus strains have a high capacity for
adherence to urogenital cells compared to other lactobacilli strains (Reid et al., 1987; Servin,
2004). Since adherence to epithelium plays an important role in initiating the pathogenesis
involved with many urogenital tract infections, the ability of lactobacilli to compete, inhibit,
and/or prevent colonization of the urogenital tract by pathogenic bacteria is significant (Servin,
2004).
1.4.1.3 Pathogenic bacterial defense mechanisms of lactobacilli
Lactobacilli promote exclusion or reduction of pathogenic bacteria adherence through the
production and actions of biosurfactants, hydrogen peroxide, lactic acid and bacteriocins.
Together these molecules limit the adherence of pathogenic bacteria to urogenital cells and
minimize their ability to colonize and flourish within the vaginal environment.
Biosurfactants are surface-active compounds that help restore homeostasis by exclusion or
reduction of pathogenic bacteria (Reid et al., 2011). Biosurfactants produced from vaginal
lactobacilli strains are composed of lipids, proteins and carbohydrates that bind to collagen III
and VI as well as fibronectin which are major components of the extracellular matrices of the
vagina and other intrauterine tissues (Howard et al., 2000; Reid et al., 2011). Lactobacilli
rhamnosus GR-1 and/or Lactobacillus reuteri RC-14 effectively disrupt Escherichia coli,
Enterococcus faucalis, Gardnerella vaginalis, Shigella dysenteria, Candida albicans,
Streptrococcuss aureus, and other bacteria biofilms and/or adherence that are associated with
bacterial vaginosis, urinary tract infections and yeast vaginitis (Howard et al., 2000; Moorthy et
al., 2009; Reid et al., 1995; Reid, 2008; Saunders et al., 2007; Velraeds et al., 1996; Velraeds et
al., 1998; Xu et al., 2008).
Hydrogen peroxide is a non-specific oxidizing agent that promotes the production of free
radicals and activates epithelial cell responsiveness to inflammatory stimuli (Voltan et al., 2008).

25
Gardnerella vaginalis is a non-catalase producing bacteria and therefore, hydrogen peroxide
inhibits its growth (MacPhee et al., 2010). Women with hydrogen peroxide-producing
lactobacilli present in their vaginal microbiota have lower incidence of bacterial vaginosis
(Eschenbach et al., 1989; Hillier et al., 1992; Hillier et al., 1993). Although Lactobacillus
rhamnosus GR-1 does not produce much hydrogen peroxide, it produces lactic acid through the
fermentation of glycogen from the vaginal epithelium (Saunders et al., 2007). Bacteriocins have
a narrow killing spectrum in comparison to hydrogen peroxide and lactic acid as they damage the
cytoplasmic membranes of bacteria (Reid et al., 2011; Servin, 2004).
1.4.1.4 Immunomodulatory properties of lactobacilli
Various strains of lactobacilli have been shown to modulate the immune response in immune cell
lines, gut epithelial and placental trophoblast cells. DC429, a strain of Lactobacillus rhamnosus
increased chemotaxis of polymorphonuclear immune cells, activated phagocytosis in the
recruited cells and increased cytokine production in murine air pouch models (Kotzamanidis et
al., 2010). The cytokine profile stimulated by DC429 was characterized by increased production
of IFNγ, IL-5, IL-6, and IL-10 in the gut mucosal lining of mice. The authors of this study
suggested that DC429 helped to maintain a physiologic state of inflammation crucial to the
defense of the gut epithelial layer (Kotzamanidis et al., 2010). Heat-killed Lactobacillus
rhamnosus GG stimulated IL-4, IL-10 and urocortin release from human placental trophoblast
cells (Bloise et al., 2010). Similar to results shown with the supernatant of Lactobacillus
rhamnosus GR-1, heat-killed Lactobacillus rhamnosus GG prevents LPS-stimulated TNFα
release from primary placental trophoblast cultures (Bloise et al., 2010; Yeganegi et al., 2010).
Additionally, Lactobacillus rhamnosus GR-1 supernatant increased IL-10 output in primary
cultures of human placental trophoblast cells (Yeganegi et al., 2010). Interestingly,
Lactobacillus rhamnosus GR-1 supernatant is capable of altering levels of PGDH, PGHS-2 and
G-CSF in a sex-dependent manner (Yeganegi et al., 2009; Yeganegi et al., 2011). These results
suggest that the supernatant of Lactobacillus rhamnosus GR-1 contains metabolites capable of
exerting the beneficial effects regardless of the presence of live or heat-killed bacteria.
Therefore, Lactobacillus rhamnosus strains and/or supernatant metabolites are capable of
modifying both immune cell chemotaxis and activity levels as well as the cytokine and
prostaglandin metabolizing enzymes in a gestational tissue.

26
1.4.2 The Potential Mechanisms of Probiotics
The exact mechanism by which lactobacilli exert their beneficial effects on the host remains
elusive. However, studies have shown that the supernatant of some Lactobacillus species can be
beneficial. After separating the supernatant from Lactobacillus plantarum 10hk2 into protein
and polysaccharide portions and subsequent analysis of fractions based on molecular weight, an
active protein fraction of 8.7 kDa was identified. This protein fraction was capable of inducing
IL-10 production from a murine macrophage cell line and eliminates activation of key molecules
in the NFκB-cytokine transcription pathway (Chon et al., 2010). Similarly, a soluble protein
factor eluted from a gastrointestinal probiotic reduced Tumor Necrosis Factor α (TNFα) and
Interferon γ (IFNγ) secretion from T84 monolayers (Madsen et al., 2001). The active product in
two other lactic acid bacterium family members, Bifidobacterium breve and Streptococcus
thermophilus have active enzymes resistant to digestive enzymes with a molecular mass of < 3
kDa in culture medium capable of inhibiting lipopolysaccharide (LPS)-induced TNFα in
intestinal cell monolayers. This inhibitory effect was preserved after transepithelial transport
across the intestinal cell monolayer (Menard et al., 2004). Additionally, medium collected from
cultured Lactobacillus rhamnosus GG was sufficient in preventing cytokine-induced apoptosis in
intestinal epithelial cells (Yan et al., 2002). Together, these studies indicate the possibility that
probiotic metabolites are sufficient to exert their effects without the presence of live or heat-
killed bacteria. Although no such evidence has been found regarding the active molecule in
Lactobacilli rhamnosus GR-1, it stands that the culture medium from these bacteria may also be
capable of exerting effects independent of the presence of live bacteria.
The experiments in this thesis were designed to further determine the effects of probiotics on
gestational tissues during pregnancy. Taken together, the above evidence identifies Lactobacillus
rhamnosus GR-1 is a safe candidate for use in pregnancy. Lactobacillus rhamnosus GR-1
displays many anti-pathogenic characteristics that may prevent pathogenic bacteria from
dominating the vaginal flora. If access to the gestational tissues is possible, it is more likely that
a small protein byproduct or metabolite would be capable of accessing the intrauterine
environment than live probiotic bacteria. Currently, however, there is little research focused on
the effects of probiotics and their metabolites to access the intrauterine tissues during pregnancy,
or on the effects this interaction may have on the other gestational tissues including the amnion.

27
1.5 Cytokines and Chemokines
Preterm labour associated with infection is accompanied by an inflammatory cascade activated
by invasive bacterial products that stimulate increased secretion of cytokines from invading
immune cells and gestational tissues (Figure 1.3, page 33). Elevated cytokine levels stimulate
prostaglandin production in the amnion, chorion and decidua activating downstream pathways
that result in myometrial contractility, membrane rupture and cervical ripening mimicking
parturition events. Evidently, cytokines and chemokines play crucial roles in the mechanisms
involved in preterm labour associated with infection and/or inflammation. Activated neutrophils
are capable of entering into the compact, mesenchymal and spongy layers of the amnion and may
enter the amniotic cavity between the amnion epithelial cells while stimulating cytokine and
chemokine production in the fetal membranes (Saji et al., 2000). Intra-amniotic infection,
microbial invasion of the intrauterine environment and/or chorioamnionitis have been associated
with elevated levels of LPS, IL-6 and IL-8 as well as IL-1, TNFα, monocyte chemotactic protein
(MCP)-1, Regulated on Activation, Normal T cell Expressed and Secreted (RANTES) and G-
CSF in amniotic fluid compared to women with uncomplicated pregnancies (Andrews et al.,
1995; Coultrip et al., 1994; Esplin et al., 2005; Hillier et al., 1993; Romero et al., 1988b; Romero
et al., 1989a; Romero et al., 1989c). In many cases, however, amniotic fluid is sterile despite
elevated cytokine levels indicating that the presence of bacteria is not necessary for
inflammatory markers to be present (Seong et al., 2006; Smulian et al., 1999). Elevated maternal
serum levels of IL-8 and cervical or amniotic fluid levels of IL-6 are indicative of
chorioamnionitis and subsequent increased risk of preterm delivery (Andrews et al., 1995;
Jacobsson et al., 2005). Cytokine levels are increased in the gestational tissues and bodily fluids
indicative of an ongoing inflammatory response.
The amnion is composed of epithelial and mesenchymal cells that work in concert to produce
cytokines and chemokines. Many cytokines and chemokines are produced and secreted from
primary cultures of human amnion including: IL-2, IL-4, IL-6, IL-8, IL-15, IL-1 receptor
antagonist (IL-1ra), MCP-1, macrophage inflammatory protein-1α, RANTES, and TNFα.
Production and secretion of cytokines and chemokines from the amnion and chorion decreases
when the two portions of the fetal membranes are cultured independently compared to explants
of intact membrane (Menon et al., 2004). This indicates that coordinated interaction exists
between the amnion and chorion in vivo. It is hypothesized that shuttling of mRNA as well as

28
bidirectional communication and cooperation occurs between the amnion and chorion (Zaga et
al., 2004; Zaga-Clavellina et al., 2007).
During infection, bacterial products such as LTA and LPS activate pathogen recognition
receptors that activate the inflammatory cascade. LTA and LPS are wall-component proteins of
gram positive and gram-negative bacteria respectively. LPS activates TLR4 by binding to LPS-
binding protein. TLR4 co-localizes with CD14 resulting in activation of downstream MAP
kinase pathways and transcription of NFΚB genes including IL-1β, TNFα, IL-6 and IL-8. The
amnion is responsive to both bacterial endotoxin and cytokine stimulation as it expresses many
cytokine receptors as well as TLR2 and TLR4. Amnion cells maintained in vitro and as
membrane explants, secrete cytokines and chemokines after stimulation with pro-inflammatory
molecules to levels similar to those found in amniotic fluid samples from women with
pregnancies complicated by infection (Fortunato et al., 1996).
1.5.1 Pro-inflammatory Cytokines
Cytokines are small proteoglycan molecules that are secreted from immune cells and other
tissues that modulate the immune response through up-regulation of other cytokines and
chemokines and direct activation of immune cells and other aspects of immunity. Pro-
inflammatory cytokines are elevated in the amniotic fluid during preterm labour associated with
infection and/or inflammation. Pro-inflammatory cytokines induce the production of other
cytokines and chemokines, growth promotion and inhibition, angiogenesis, cytotoxicity and
other mechanisms to defend against pathogenic threats. The major cascade of pro-inflammatory
cytokines involves the first response cytokines, TNFα and IL-1β as well as IL-6. IL-6 has been
shown to play a more immunoregulatory role than its counterparts in the triad. In the female
reproductive system and throughout pregnancy, TNFα plays a role in follicular development,
ovulation, menstruation and endometrial function regulation (Argiles et al., 1997). However, in
preterm labour associated with infection and/or inflammation TNFα, IL-1β and IL-6 stimulate
prostaglandin production in the amnion. Elevated prostaglandins initiate the onset of parturition
and if levels are increased due to infection, pro-inflammatory cytokines may promote early onset
of labour.
TNFα and IL-1β are the first cytokines found in serum in response to acute infection (Petersen et
al., 2006). In LPS-induced fetal loss in pregnant mice, both TNFα and IL-1β levels are increased

29
(Silver et al., 1994; Silver et al., 1997). Unlike IL-1β levels that become detectable in the third
trimester of pregnancy, TNFα is undetectable in amniotic fluid unless an infection is present
(Romero et al., 1989c). The amnion does not produce IL-1β and its mRNA is not localized to
the amnion. However, IL-1β mRNA is found in both the chorion and decidua and it may be
shuttled and therefore account for secretion found on the fetal side of in vitro membrane explants
(Menon et al., 1995; Thiex et al., 2009; Zaga et al., 2004). IL-1β and TNFα both increase the
availability of phospholipids, the substrate for prostaglandin production, phospholipase protein
levels as well as the expression of PGHS-2, the rate-limiting step in prostaglandin production in
the amnion (Blumenstein et al., 2000; Bry et al., 1992; Pollard et al., 1993; Romero et al., 1989b;
Romero et al., 1989c). As well, IL-1β and TNFα also decrease PGDH expression in chorion
cells resulting in decreased prostaglandin metabolism (Brown et al., 1998; Pomini et al., 1999).
Therefore, pro-inflammatory cytokines cause a net increase in prostaglandin production from the
gestational tissues (Kent et al., 1993; Pollard et al., 1993).
Levels of IL-6 are increased in vaginal fluid at term and preterm labour, in cervical fluid
obtained from women with intrauterine infection and in the amniotic fluid of women with
chorioamnionitis (Imseis et al., 1997). Importantly, IL-6 increases amnion prostaglandin
production in cultured amnion cells to levels similar to those found in the amniotic fluid of
women with intrauterine infections and preterm labour (Kent et al., 1993; Mitchell et al., 1991).
IL-6 is part of the pro-inflammatory cytokine triad alongside TNFα and IL-1β. Despite its pro-
inflammatory initiation of the acute phase response from the liver in response to acute infection,
IL-6 is defined as a pleiotropic immunomodulatory cytokine. In LPS-stimulated IL-6 deficient
knock-out mice, levels of both TNFα and neutrophilia were three times higher than in wild-type
counterparts (Xing et al., 1998). These effects were abolished with treatment of recombinant IL-
6 (Xing et al., 1998). Furthermore, in cancer patients receiving recombinant IL-6 intravenous
therapy, serum levels of both IL-1ra and sTNFα receptor, the antagonists to TNFα and IL-1β,
were increased (Tilg et al., 1994). IL-6 treatment increases levels of the anti-inflammatory
cytokine, IL-10 (Steensberg et al., 2003). Negative feedback is provided by IL-6 against TNFα
and IL-1β which may help maintain and facilitate the decrease and resolution of pro-
inflammatory immune responses (Steensberg et al., 2003).

30
1.5.2 CXC and CC Chemokines
Chemokine release is stimulated by pro-inflammatory molecules from immune cells and other
immune responsive tissues. Classified by patterns of amino acids in their amino terminus,
chemokines are separated into two families: CXC or CC. Chemokines are responsible for
immune cell recruitment, chemotaxis, activation and motility. Levels of chemokines that recruit
immune cells are elevated at term and prior to the onset of labour. However, the levels of many
chemokines increase throughout gestation and in association with infection during pregnancy
and many of their specific roles remain unknown.
IL-8 or CXCL8 is a member of the CX chemokines and plays a role in endothelial cell
proliferation and survival as well as inducing directional motility and activation of neutrophils
and T-lymphocytes (Li et al., 2003). As such, elevated concentrations of IL-8 in amniotic fluid
are associated with microbial infection, increased neutrophil count, membrane rupture, and
cervical ripening (Cherouny et al., 1993; Hsu et al., 1998). IL-8 is constitutively expressed in the
amnion and as mechanical tension increases towards the end of the mid-trimester, the expression
of the stretch-responsive IL-8 gene is increased and subsequently levels of IL-8 in the amniotic
fluid rise (Kendal-Wright, 2007; Romero et al., 1991). It is thought that increased IL-8 levels
associated with intrauterine infection recruit neutrophils into the fetal membranes and placenta
(Cherouny et al., 1993). Unlike TNFα, IL-1β, and IL-6, IL-8 does not stimulate amnion
prostaglandin production. However when the amnion is stimulated with endotoxin, amnion cells
respond with increased IL-8 to levels similar to those found in women with intra-amniotic
infection.
The family of CC chemokines includes MCP-1 and RANTES which recruit immune cells that
express the receptor, CCR5 including monocytes, basophils and eosinophils. MCP-1 is released
from ovarian follicles, endometrium and the choriodecidua as well as the amnion in the third
trimester (Arici et al., 1995; Arici et al., 1997; Denison et al., 1998). MCP-1 levels in amniotic
fluid are increased during preterm labour with chorioamnionitis and with preterm labour with or
without infection (Esplin et al., 2005). RANTES is released by the endometrium, first trimester
trophoblast cells and the amnion (Denison et al., 1998; Hornung et al., 1997; Svinarich et al.,
1996). RANTES levels in amniotic fluid decrease with gestational age and increase with
preterm labour associated with microbial invasion as well as at term (Athayde et al., 1999).

31
RANTES attracts monocytes and lymphocytes and increases immune cell adhesion. CC
chemokines activate immune cells. The important role of CC chemokines is demonstrated by the
inability of MCP-1 deficient knock-out mice to resist microbial infection and an inability to clear
bacteria (Yadav et al., 2010).
1.5.3 Immune Cell Activating Cytokines
Both IFNγ and granulocyte-macrophage colony-stimulating factor (GM-CSF) play important
roles in the adaptive immune system by participating in the activation of recruited immune cells
at the site of infection. IFNγ is an immunomodulatory cytokine crucial in the adaptive immune
system for defense against viral and intracellular bacterial infections. IFNγ increases antigen
presentation through up-regulation of MHC I and MHC II complexes, release of reactive oxygen
species in macrophages and promotes adhesion and binding of leukocytes during migration.
IFNγ plays a role in maternal recognition of pregnancy as it is involved in implantation and
blastocyst attachment (Murphy et al., 2009). In the decidua, IFNγ receptor expression decreases
in both term and preterm placentae (Hanna et al., 2004). IFNγ decreases PGHS-2 expression and
consequentially the production of PGE2 in placental explants in vitro (Hanna et al., 2004). It is
suggested that IFNγ receptor down-regulation occurs at term. With a loss of IFNγ receptor
expression, IFNγ is no longer able to repress PGHS-2 expression in the placenta resulting in an
increase in prostaglandin production (Hanna et al., 2004). IFNγ has not been identified in the
amniotic fluid; however, IFNγ is expressed in amnion tissue and shows no changes as labour
progresses (Veith et al., 1999).
Similarly, GM-CSF promotes proliferation of monocyte, neutrophils and eosinophils, GM-CSF
also activates phagocytosis, chemotaxis and adhesion as well as promoting cell growth and
survival (Gasson, 1991). During early pregnancy, GM-CSF is involved in murine normal
placental growth and differentiation as well as embryonic implantation and cytotrophoblast
differentiation (Bry et al., 1997; Garcia-Lloret et al., 1994; Robertson et al., 1992). Transgenic
mice with overexpression of GM-CSF show high levels of macrophage proliferation and
activation (Metcalf et al., 1988). Levels of GM-CSF increase throughout gestation and women
with PROM show higher levels compared to those with intact membranes during pregnancy (Bry
et al., 1997). GM-CSF is produced in the amnion, trophoblasts and decidua. Both TNFα and IL-
1β stimulate GM-CSF release from amnion cells (Bry et al., 1995). Expression of GM-CSF in

32
the amnion and levels of GM-CSF in the amniotic fluid are elevated during chorioamnionitis and
intrauterine infection respectively (Keelan et al., 2003; Stallmach et al., 1995). It is suggested
that GM-CSF may amplify the immune response in the amnion while increasing host resistance
to infection.
The interaction between cytokines and chemokines plays a crucial role in the immune response
against pathogenic threats. However, during pregnancy an imbalance of TH1 to TH2 cytokines
results in increased risk of preterm delivery through subsequent increases in prostaglandin
production. Infection of the amniotic cavity and the accompanying inflammatory processes are
harmful to the fetus because it may lead to Fetal Inflammatory Response Syndrome associated
with cerebral white matter damage and development of cerebral palsy (Yoon et al., 2003).
Although the fetal membranes act as a barrier to cytokines produced in the decidua, both the
chorion and the amnion are responsive to bacterial endotoxin and cytokine stimulation (Kent et
al., 1994; Menon et al., 2004). Therefore, the contribution of the amnion to the cytokine pool in
response to bacterial endotoxin and cytokines in infection and inflammation may play a major
role in determining the onset of parturition and outcome of labour. By controlling the level of
cytokines and chemokine stimulation during pregnancy, it may be possible to eliminate
activation of downstream pathways leading to preterm delivery. Therefore, this has led to the
investigation of the ability of Lactobacillus rhamnosus GR-1 to modulate the cytokine and
chemokine production from the amnion.

33
Figure 1.3: Proposed pathway of infection and/or inflammation-mediated preterm birth
[The image is taken from New England Journal of Medicine. Vol 342.20. Goldenberg RL, Hauth
JC, Andrews WW. Intrauterine infection and preterm delivery. 1500-1507 (2000) and appears
here with the permission of the journal.]

34
1.6 Prostaglandins
The family of biologically active lipid molecules consists of 20 carbon fatty acids that
categorized into leukotrienes, lipoxins, and prostaglandins (Olson, 2003). Prostaglandins are
produced by all cells of the body and act as local hormones initiating downstream cascades in a
paracrine manner. Prostaglandin production and metabolizing pathways are well characterized
and many of the enzymes involved are mediated by cytokines, bacterial products and endotoxins,
growth factors and glucocorticoids that increase prostaglandin production in states of infection
and/or inflammation. The biosynthesis of prostaglandins begins with the cleavage of the lipid
bilayer membrane either directly by members of the phospholipase A2 family or indirectly
through the actions of phospholipase C (Olson, 2003). As a result, arachidonic acid is freed from
the phospholipid membrane into the cytosol where it is readily converted through varying
enzymatic pathways to form lipoxins, leukotrienes, or prostaglandins. In early gestation,
metabolism leading to lipoxygenase products is favoured; however, as term approaches
metabolism is shifted towards production of prostaglandins dominating prior to the onset of
labour (Figure 1.4, page 38; Mitchell et al., 1995).
1.6.1 Prostaglandin Production
1.6.1.1 Phospholipases
Prostaglandin production begins with the cleavage of arachidonic acid from phospholipid
membranes by phospholipase enzymes. Amnion cell membranes are rich in arachidonate,
therefore cleavage of amnion cell membranes are ideal residues for prostaglandin substrate
(Curbelo et al., 1981). The family of phospholipase A2 enzymes consists of up to 15 isoforms.
Type IV, cytosolic phospholipase A2 has been shown to have a prominent role in the amnion;
however, many isoforms are also expressed in the fetal membranes (Myatt et al., 2010).
Cytosolic phospholipase 2 translocates to the cell membrane in response to agonist stimulation,
including the cytokines, IL-1 and IL-6 as well as bacterial endotoxins (Challis et al., 2002).
Therefore, much like PGHS, the enzyme involved in the rate-limiting step in prostaglandin
production, levels of phospholipase enzymes can be up-regulated in a state of infection and or
inflammation in the amnion.

35
1.6.1.2 Prostaglandin H Synthase
Freed arachidonic acid is converted into PGH2 by two subsequent reactions involving PGHS.
The first reaction is a cyclooxygenase reaction that combines two free oxygen molecules with
arachidonic acid to form PGG2, a relatively unstable intermediate. A peroxidase reaction follows
between PGG2 and PGHS to form PGH2. Two major forms of PGHS, PGHS-1 and PGHS-2, are
present in the amnion, similar to most cell types including various gestational tissues throughout
pregnancy (Hirst et al., 1995; Rose et al., 1990; Teixeira et al., 1994). PGHS enzymes
commonly reside near the membrane and within lipid bodies in various cell types ideally situated
for conversion of freed arachidonic acid substrate (Dvorak et al., 1994; Smith et al., 1981).
PGHS-1 and PGHS-2 share considerable sequence homology and up to 60-65% cDNA sequence
homology despite being the products of two separate genes (Blumenstein et al., 2000; Mitchell et
al., 1995; Xu et al., 1995). Both enzymes are homodimers and have equal affinity and capacity
to convert arachidonic acid to PGG2 and subsequently PGH2 with equal relative efficiency
(Olson, 2003). The major difference in the role of the two enzymes is their differential
expression and regulation. PGHS-2 is inducible as its promoter consists of many binding
elements that can influence its expression pattern: TATA box, two NFκB, NF-IL6, GRE, CRE,
and AP-2, SP-1, and ETS-1 binding sites (Inoue et al., 1995; Tazawa et al., 1994). The
expression of PGHS-2 is regulated and altered by cytokines, oxytocin, mechanical stretch,
glucocorticoids, bacterial endotoxins, growth factors, activators of protein kinase C and factors
that increase intracellular calcium concentrations such as CRH (Bennett et al., 1987;
Blumenstein et al., 2000; Economopoulos et al., 1996; Masferrer et al., 1992; Mohan et al., 2007;
Molnar et al., 1999; Smieja et al., 1993; Zakar et al., 1994; Zakar et al., 1995). Due to the ability
of PGHS-2 mRNA levels, protein expression and activity to be modulated by external stimuli,
conversion of arachidonic acid to PGH2 is a likely candidate for the rate-limiting step in
prostaglandin production. Amnion PGHS-2 mRNA levels, protein expression and activity levels
increase throughout gestation and further at term and with the onset of labour in an NFΚB
dependent manner (Fuentes et al., 1996; Gibb et al., 1996; Hirst et al., 1995; Johnson et al., 2006;
Mijovic et al., 1999; Mitchell et al., 1995; Slater et al., 1998; Teixeira et al., 1994). TNFα, IL-1β
and bacterial endotoxin increase PGHS-2 expression levels up to 80-fold compared to a 2 to 3
fold increase seen in PGHS-1 expression (Blumenstein et al., 2000; Mitchell et al., 1993; Pollard
et al., 1993). Therefore, by altering PGHS-2, cytokines and bacterial products associated with

36
infection and/or inflammation during labour have an ability to largely affect amnion
prostaglandin production.
1.6.1.3 Prostaglandin Synthases
The last step in prostaglandin production is performed by the prostaglandin synthase enzymes.
PGH2 is converted by a member of the prostaglandin synthase enzyme family to either
prostaglandin E2, F2α, D2 and I2. The major prostaglandin product formed in the amnion is
prostaglandin E2 and the minor product PGF2α due to the enzymatic activity of prostaglandin E
synthase (PGES) and prostaglandin F2α synthase, respectively. Three isoforms of PGES are
expressed in the amnion: microsomal (m)PGES1, mPGES2, and cytosolic (c)PGES also known as
mPGES3 (Ackerman et al., 2008). cPGES is constitutively expressed and its activity is thought
to be coupled to PGHS-1 (Han et al., 2002; Myatt et al., 2010). cPGES is localized to the
cytosol, in particular at lipid bodies of amnion epithelium as well as in fibroblasts whereas
mPGES1 is localized only to amnion epithelium membranes and lipid bodies (Meadows et al.,
2003). mPGES1 is the most catalytic efficient form of PGES above both other isoforms (Thoren
et al., 2003). Expression of mPGES1 is induced by various cytokines such as IL-1β, LPS and
other pro-inflammatory stimuli (Forsberg et al., 2000; Jakobsson et al., 1999; Mancini et al.,
2001). As well, mPGES1 is coupled to expression of PGHS-2 and is thought to be regulated
through NFΚB transcriptional activity (Kudo et al., 1999; Thoren et al., 2000). The third isoform
of PGES is mPGES2 which is formed as a golgi-membrane bound protein that translocates to the
cytosol to perform its enzymatic function (Murakami et al., 2003). mPGES2 is coupled to both
PGHS-1 and PGHS-2 and is constitutively expressed (Murakami et al., 2003). No change in
either cPGES or mPGES2 is seen in the fetal membranes at term or preterm with or without
labour; however mPGES1 can be induced by pro-inflammatory cytokines (Meadows et al.,
2003). The exact influence alterations to PGES expression has on labour with and without
infection is unknown (Premyslova et al., 2003).
1.6.1.4 15-Hydroxyprostaglandin Dehydrogenase
PGDH is a catabolic enzyme for PGE2 and PGF2α that plays an important role in minimizing the
spread of prostaglandins produced in the amnion from reaching the other gestational tissues
(Sangha et al., 1994). Unlike the amnion where expression is absent, chorion trophoblast cells
express high levels of PGDH (Cheung et al., 1990). This enables the chorion to act as a

37
metabolic barrier to amnion produced prostaglandins from acting on the other gestational tissues.
Chorionic PGDH activity decreases at term and in idiopathic preterm deliveries allowing amnion
produced prostaglandins to influence the other intrauterine tissues (Sangha et al., 1994; Van Meir
et al., 1996). Cytokines have been shown to decrease the expression of PGDH in the chorion. As
well, inconsistencies of PGDH expression throughout the chorion can be induced during
infection and inflammation (Cheung et al., 1990; Sangha et al., 1994; Van Meir et al., 1996; Van
Meir et al., 1997). Therefore, changes in PGDH throughout the chorion may grant passageway
of amnion produced prostaglandins access to the uterine myometrium and cervix ultimately
leading to activation and ripening respectively and a subsequent increased risk of preterm
delivery (Challis et al., 2002).

38
Figure 1.4: Effects of stimulants on the prostaglandin production pathway in the amnion.
Molecules that stimulate prostaglandin production are in blue font at the top of the diagram. Red
font indicates enzymes involved in prostaglandin production. Hatched lines indicate
metabolizing enzymatic activity and solid lines indicate synthesizing enzyme activity. The major
product of the amnion is PGE2 with a minor product of PGF2α.
Prostaglandin H
Arachidonic Acid
Prostaglandin E2 Prostaglandin F
2α
Phospholipids
Phospholipase A2
Prostaglandin G2
Prostaglandin H Synthase -1 and 2 Cyclooxygenase Reaction +2O2
Peroxidase Reaction -2e-
Cytosolic and microsomal
Prostaglandin E Synthase-1,
2
Prostaglandin F Synthase
Prostaglandin H Synthase-1 and 2
Bacterial Products, LPS, TNFα, IL-1β, IL-6, IL-10, Oxytocin, Glucocorticoids
15- hydroxyprostaglandin dehydrogenase

39
1.6.2 The Role of Amnion Produced Prostaglandins in Preterm Labour
At term, elevated amniotic fluid prostaglandins are paralleled with increased levels of
prostaglandin producing enzymes and activity in amnion epithelial and fibroblast cells. Amnion
produced prostaglandins play a role in fluid or ion balance as well as mediation of
transmembrane ion flow (Challis et al., 2002; Saunders-Kirkwood et al., 1993). Although the
amnion, chorion and decidua produce increasing amounts of prostaglandins throughout gestation,
only the amnion and chorion show increased prostaglandin production prior to the onset of
labour (Olson et al., 1983; Skinner et al., 1985). Increased prostaglandin levels initiate a set of
events necessary for parturition onset and successful completion of labor including: myometrial
activation, cervical ripening, up-regulation of MMPs leading to membrane rupture, fetal
adaptations to labour including decreased breathing and movements, increases in fetal HPA axis
activity and changes in uterine and placental blood flow (Kitterman, 1987; Rankin, 1976; Sastry
et al., 1997; So, 1993; Thorburn, 1992). Elevated levels of prostaglandins can lead to preterm
labour as is demonstrated by administration of prostaglandins in mammalian species that result
in induction of labour or abortion at any stage of gestation (Embrey, 1971).
The two cell types of the amnion, epithelial and mesenchymal cells respond differently to stimuli
when cultured separately in vitro. It is suggested that the two cell types of the amnion exert
paracrine actions on one another to form a regulation loop for prostaglandin production (Whittle
et al., 2001). Mesenchymal cells produced greater per cell quantities of prostaglandins; however
since epithelial cells outnumber amnion fibroblast cells 10 000:1, the overall contribution of
epithelial cells to the prostaglandin pool is greater (Whittle et al., 2000). Many cytokines
including IL-1α, IL-1β, TNFα, IL-6, MIP-1α increase prostaglandin production from the amnion
(Dudley et al., 1996; Furuta et al., 2000; Mitchell et al., 1991; Romero et al., 1989b; Romero et
al., 1989c). Additionally, gram-negative bacteria, LPS and arachidonic acid increase
prostaglandin production in the amnion (Reisenberger et al., 1998; Romero et al., 1988a; Romero
et al., 1988b). In contrast to the typical anti-inflammatory properties of IL-10 across many
tissues, the amnion shows increases prostaglandin production in response to IL-10 treatment
(Economopoulos et al., 1996; Gibb et al., 1990; Mitchell et al., 2004; Potestio et al., 1988). As
well, glucocorticoids stimulated prostaglandin production in amnion mesenchymal cells contrary
to its well-established anti-inflammatory effects in many other tissues (Whittle et al., 2000).

40
In cultured human placenta trophoblast cells, the supernatant of Lactobacillus rhamnosus GR-1
modulates expression levels of enzymes involved in prostaglandin production. Pre-treatment of
Lactobacillus rhamnosus GR-1 prevents LPS-induced PGHS-2 expression in placentae derived
from pregnancies carrying a male fetus (Yeganegi et al., 2009). As well, in placentae derived
from pregnancies carrying a female fetus, Lactobacillus rhamnosus GR-1 supernatant increases
PGDH expression to a higher degree than in placentae derived from pregnancies carrying a male
fetus (Yeganegi et al., 2009). Although the exact mechanism for the decreased expression of
PGDH is unknown, however these results may give a molecular basis for the higher incidence of
preterm labour that occurs in pregnancies carrying a male fetus compared to a female.
Comparatively, another strain, Lactobacillus rhamnosus GG induces PGHS-2 expression in a
time and dose-dependent manner in T84 colon epithelial cells (Korhonen et al., 2004).
Therefore, the effects of Lactobacillus species are dependent on the strain as well as the cell type
and gender of the host tissue. Many stimuli associated with infection and/or inflammation are
capable of stimulating prostaglandin production in the amnion. In addition, molecules that
typically decrease prostaglandin production across tissues, such as cortisol, also up-regulate
prostaglandin production in the amnion. The amnion has been identified as the main site of
prostaglandin production at the end of gestation and the onset of labour (Challis et al., 1997). If
the amnion is stimulated early in gestation by molecules associated with infection and/or
inflammation, events normally associated with term labour may be activated leading to an
increased risk of onset of preterm delivery.

41
1.7 Summary
Bacterial vaginosis, which is associated with increased risk of preterm labour by 40%, is
characterized by the colonization of pathogenic bacteria in the vaginal tract and decreased levels
of endogenous Lactobacillus species (Hillier et al., 1995). Bacterial endotoxins and bacterial
products stimulate cytokine, chemokine and prostaglandin production that may activate
downstream pathways leading to preterm labour (Figure 1.3, page 33). Interfering with the
upstream stimulants of these pathways may allow the prevention of preterm delivery mediated
by infection and/or inflammation. The particular interest of this thesis was on the
immunomodulatory role of Lactobacillus species in the amnion. Lactobacillus species have been
previously shown to modulate the inflammatory response in LPS-stimulated placental
trophoblasts and mouse macrophages and are capable of colonization and restoration of the
vaginal microbiota. However, the effect of lactobacilli on the amnion remains unknown. In
order to help determine its ability to be a potential preventative treatment, this thesis examined
the role of Lactobacillus rhamnosus GR-1 in regulating mediators of inflammation and/or
infection associated with preterm birth release from the human amnion. In particular, the effects
of Lactobacillus on cytokines, chemokines, prostaglandins and prostaglandin metabolizing
enzymes in human amnion were investigated.

42
Chapter 2
Rationale, Hypothesis and Specific Aims

43
Chapter 2
Hypotheses, Rationale and Specific Aims
2.1 Overall Hypothesis
Lactobacillus rhamnosus GR-1 will prevent elevation of cytokines, chemokines and
prostaglandins associated with infection and/or inflammation-mediated preterm birth from the
human amnion.
2.1.1 Rationale
Infection-mediated preterm birth is associated with stimulation of an exaggerated inflammatory
response in both the gestational tissues and recruited immune cells. Bacteria and bacterial
products stimulate cytokines and chemokines from the gestational tissues. During
chorioamnionitis and microbial invasion of the amniotic cavity, the cytokine pool in the amniotic
fluid is characterized by increases in pro-inflammatory cytokines and chemokines. As many
cytokines produced in the decidua are unable to cross the fetal membranes, the amnion is thought
to be the prominent producer of elevated cytokines during an inflammatory response associated
with infection. The amnion also produces prostaglandin E2 (PGE2) that may exert paracrine
actions on both the myometrium and cervix to cause activation and remodeling respectively. As
well, PGE2 exerts paracrine effects on the fetal membranes resulting in collagen degradation and
weakening. Probiotics, namely Lactobacillus species have known immunomodulatory roles in
murine macrophages, monocytic cell lines and human placental trophoblasts which promote an
anti-inflammatory cytokine profile and increased chemokine production. Additionally,
Lactobacillus species modulate prostaglandin-metabolizing enzymes. Therefore, the effects of
Lactobacillus rhamnosus GR-1 on cytokines/chemokines and prostaglandin production from the
amnion was determined in these experiments.
Although numerous strains of lactobacilli have been shown to exert immunomodulatory effects,
Lactobacillus rhamnosus GR-1 was used to treat amnion cells in the current study. Due to its
ability to colonize and replenish the vaginal flora when administered orally and ability to alter
the immune response in human placental trophoblasts and other epithelial cell types,
Lactobacillus rhamnosus GR-1 was assessed in its effects to possibly exert similar effects in

44
human amnion cells. Although the exact route of orally administered probiotics to the vaginal
microbiota is unknown, Lactobacillus species have been shown to be bile and acid resistant and
have been suggested to migrate from the rectum to the vagina after travelling through the
gastrointestinal tract (Reid et al, 2001b).
The intrauterine tissues during healthy pregnancies are sterile; therefore we chose to use treat
amnion cells with the supernatant of Lactobacillus rhamnosus GR-1 as opposed to live or freeze
dried bacteria. It is currently unknown whether endogenous lactobacilli are capable of
interacting with the fetal membranes; however, recent research has identified that small protein
molecules are capable of exerting the effects of other probiotics, including strains of
Lactobacillus. It may be that a small molecule is more capable of accessing the amnion
compared to live or heat killed bacteria.
During bacterial vaginosis, gram-negative bacteria, such as Bacteroides spp. and gram-
positive/gram-variable bacteria, such as Gardnerella vaginalis and Mycoplasma spp. levels are
elevated. Therefore, both LTA, a wall-component of gram-positive bacteria and LPS, a wall-
component of gram-negative bacteria were used to stimulate amnion cells in this study.
Importantly, Lactobacillus rhamnosus GR-1 is a gram-positive bacterium; therefore, LTA
treatment acts as an internal control to separate the effects of Lactobacillus rhamnosus GR-1
metabolites present in the supernatant used from the effects of the characteristic gram-positive
bacterial endotoxin, LTA.
2.2 Chapter 3: Effect of Lactobacillus rhamnosus GR-1 on cytokines and chemokines in human amnion cells at term
Hypothesis:
Lactobacillus rhamnosus GR-1 supernatant will ameliorate increases in LTA and/or LPS-
stimulated pro-inflammatory cytokines and chemokines in human amnion cells in order to
prevent activation of downstream pathways associated with infection and/or inflammation -
mediated preterm birth
Specific Aims:
1. To determine the optimal dose and length of exposure to LTA and LPS on cytokine and
chemokine release in medium collected from cultured human amnion cells at term.

45
2. To determine the effects of Lactobacillus rhamnosus GR-1 on the release of interleukin
(IL)-6, IL-8 and tumor necrosis factor α (TNFα) in medium collected from LPS and LTA
treated human amnion cells at term.
3. To determine the effects of Lactobacillus rhamnosus GR-1 on global cytokine and
chemokine release in medium collected from LTA and LPS treated human amnion cells
at term.
2.3 Chapter 4: Effect of Lactobacillus rhamnosus GR-1 on Prostaglandin Output and Prostaglandin Metabolizing Enzymes in Human Amnion Cells at Term
Hypothesis:
Lactobacillus rhamnosus GR-1 supernatant will ameliorate increases in LTA and/or LPS-
stimulated prostaglandin production in human amnion cells in order to prevent propagation of
downstream pathways associated with infection and/or inflammation -mediated preterm birth
Specific Aims:
1. To determine the effects of Lactobacillus rhamnosus GR-1 on prostaglandin E2
output in LPS and LTA treated human amnion cells at term.
2. To determine expression profiles of prostaglandin metabolic and synthetic
enzymes (Prostaglandin H Synthase-2, cytosolic Prostaglandin E Synthase,
microsomal Prostaglandin E Synthase 1, microsomal Prostaglandin E Synthase 2)
that may account for any changes in PGE2 release caused by LTA, LPS and
Lactobacillus rhamnosus GR-1 in human amnion cells at term.

46
Chapter 3
Effect of Lactobacillus rhamnosus GR-1 on Cytokines and
Chemokines in Human Amnion Cells at Term

47
Chapter 3
Effect of Lactobacillus rhamnosus GR-1 on Cytokines and Chemokines in Human Amnion Cells at Term
3.1 Introduction
Approximately half of all preterm births are idiopathic. However, 30% of preterm births are
associated with infection and/or an activated inflammatory process (Goldenberg et al., 2008;
Hillier et al., 1995). This inflammatory process is characterized by bacterial endotoxin
stimulation of amnion cytokine and chemokine production. Despite ongoing research, current
efforts at prevention are largely ineffective and consequentially, preterm birth remains a major
obstetrical concern.
The presence of bacteria triggers a response by the body’s innate immune system which is
characterized by increased levels of cytokines and chemokines. During pregnancy, intrauterine
infections are associated with increased levels of cytokines, including IL-1β, TNFα, IL-6, IL-8,
RANTES, and MCP-1 in the amniotic fluid (Andrews et al., 1995; Coultrip et al., 1994; Esplin et
al., 2005; Hillier et al., 1993; Romero et al., 1989c). Many cytokines produced in the decidua are
not capable of passing through the membranes. Therefore, the amnion and chorion play a major
role in contributing to the pool of cytokines and chemokines in the amniotic fluid. Elevated
levels of cytokines and chemokines recruit immune cells and increase prostaglandin production
in the amnion, chorion, and decidua (Challis et al., 1997). The net result is activation of
downstream pathways leading to increased metalloproteinase expression and subsequent cervical
ripening and membrane weakening as well as myometrial activation.
Antibiotics, such as metronidazole and clindamycin have been used to prevent infection and/or
inflammation-associated preterm birth. However, in some cases antibiotics may increase the
incidence of preterm labour (Andrews et al., 2003; Okun et al., 2005). Probiotics, defined as,
“live microorganisms which when administered in adequate amounts confer a health benefit on
the host,” are candidates for prevention of preterm labour (FAO and WHO, 2001; Reid, 1999).
Bacterial vaginosis is present in approximately 20% of pregnant women and increases the risk of
undergoing preterm birth by 40% (Hillier et al., 1995). The hallmark characteristic of bacterial
vaginosis is a decline in endogenous Lactobacillus species and a rise in pathogenic bacteria

48
resulting in an alteration to the vaginal microbiota. Due to the ability of Lactobacillus rhamnosus
GR-1 to colonize and restore the vaginal microbiota and exert immunomodulatory effects in
murine macrophages and placental trophoblasts, probiotics stand as a candidate preventative
treatment for infection and/or inflammation-mediated preterm birth (Reid et al., 2003; Yeganegi
et al., 2010; Yeganegi et al., 2011).
The effects of Lactobacillus rhamnosus GR-1 on the human amnion have not been previously
investigated. The amnion is responsive to both bacterial products and cytokines and therefore
plays an important role during infection and/or inflammation. Therefore, Lactobacillus
rhamnosus GR-1 may enable the amnion to resist bacterial endotoxin stimulation of cytokine and
chemokine production that occurs in infection and/or inflammation-mediated preterm birth.
3.2 Methodology
3.2.1 Placenta Collection
All studies were approved by the Faculty of Medicine, University of Toronto and Mount Sinai
Hospital (Toronto, Canada) ethics review boards in accordance with the Canadian Tri-Council
Policy Statements on Human Ethics Reviews (institutional review board #04-0018-U). Placentae
were collected and informed consent was obtained by the Biobank program of the Research
Centre for Woman`s and Infant`s Health at Mount Sinai Hospital (http://biobank.lunenfeld.ca)
from women undergoing elective caesarean sections at term (>37 weeks) at Mount Sinai
Hospital (Toronto, Canada). Inclusion criteria included: 1) No complications during pregnancy,
2) No abnormal antenatal test results (including laboratory results and ultrasounds), 3) An
unremarkable maternal medical history, free of any conditions that may significantly impact
placental pathology (i.e. renal disease, diabetes, hypertension, cardiovascular disease), 4) No
clinical signs of infection or labour 5) Free from any infections during pregnancy including HIV,
Hepatitis B and group B streptococcus. Indications for delivery by caesarean section included:
abnormal presentation of the fetus, cephalopelvic disproportion, or previous caesarean sections.
Patients were not screened for bacterial vaginosis.
3.2.2 Mixed Amnion Epithelial and Mesenchymal Cell Culture
Amnion membranes were stripped from the underlying choriodecidual layer and washed in
saline until the majority of blood was removed. The amnion was suspended in forceps and cut in

49
strips by cutting towards the forceps at 1.5 cm intervals. The amnion was then digested for 45
mins in Dulbecco Modified Eagle Medium (DMEM; GIBCO Invitrogen; Burlington, Ontario,
Canada) + 0.1% trypsin (GIBCO Invitrogen) to obtain amnion epithelial cells in a 37°C
incubator. Tissue was then filtered using a 0.8 nm gauze filter. Filtered DMEM was treated with
filtered newborn calf serum (NCS; Wisent Incorporated; St. Bruno, Quebec, Canada) to inhibit
trypsin activity. Tissue was cut into fragments of approximately 1.0 cm in length and further
digested for 45 mins in DMEM + 0.1% trypsin; filtered and treated with filtered NCS to inhibit
trypsin activity. Epithelial cells from the first and second digestion were pelleted by
centrifugation at 1250 g for 5 mins. Excess medium was discarded and epithelial cells were re-
suspended in culture medium DMEM-F12 (GIBCO Invitrogen) + 10% fetal bovine serum (FBS;
Wisent Incorporated) + 1% antibiotics. The remaining tissue was cut into approximately 0.5 cm
pieces and digested for 1 hr in DMEM + 0.1% collagenase (Roche Diagnostics; Mannheim
Germany) in order to obtain amnion mesenchymal cells. After digestion, mesenchymal cells
were centrifuged for 10 mins at 1500 g. Total mesenchymal cells obtained were re-suspended in
culture medium and mixed with total epithelial cells from previous digestions, despite in vivo
proportions. Mixed amnion cells were counted using a Trypan Blue exclusion assay to ensure
greater than 98% viability. 20 µl of mixed amnion cell culture was diluted in 180 µl of culture
medium. 40 µl of diluted cultured cells were mixed with 40 µl Trypan Blue. Cells were them
viewed under 40x magnification and counted on a hemocytometre. Blue cells representing cell
death were assessed in the viewing field and cultures with dispersed cells displaying greater than
2% cell death were discarded. Mixed amnion epithelial and mesenchymal cells were plated to a
density of 0.8x106 cells per well in a 24-well culture plate.
3.2.3 Lactobacilli Preparation
Lactobacillus rhamnosus GR-1 was grown anaerobically for 48 hours to reach the stationary
growth phase in de Man, Rogosa, and Sharpe media (MRS) by colleagues in Dr. S.O. Kim’s
laboratory at the University of Western Ontario. The culture medium was then collected and
centrifuged at 6000 g for 10 minutes at 4⁰C. Residual bacteria were removed by filtration of the
supernatant through a 0.22 μm pore size filter. Supernatant was aliquoted and frozen at -80⁰C
until used.

50
3.2.4 Treatment Protocol
Cells were cultured for 48 hr at 37°C in 5% CO2 and 95% room air and washed with room
temperature phosphate buffered saline (PBS) without calcium and magnesium. Culture medium
was replaced and cell confluency was reached after an additional 48 hr incubation. Endotoxin
dose, time and Lactobacillus rhamnosus GR-1 supernatant optimization experiments were
performed. Both LTA (Invivogen; San Diego, California, United States of America) and LPS
(Sigma-Aldrich; St Louis, Missouri, United States of America) at doses 0.1, 1, 10, 100, and 1000
ng/ml were tested for a time period of 12 hr after starvation in serum-free conditions. Medium
was collected for LTA (10 ng/ml) and LPS (100 ng/ml) at time 4 hr, 8 hr, 12 hr and 24 hr post-
treatment. The effects of various Lactobacillus rhamnosus GR-1 supernatant dilutions, 1:10,
1:20, 1:50, and 1:100 were assessed at 12 hr post-treatment. After conditions were optimized,
cell cultures were serum starved for 12 hr. Lactobacillus rhamnosus GR-1 supernatant was
thawed and diluted in DMEM at a ratio of 1:20 and incubated for 12 hr. Excess supernatant was
discarded after each experiment. Fresh batches of supernatant were received every two months to
maintain similar levels of enzymatic activity across experiments. Cells were then treated with
LTA (10 ng/ml) or LPS (100 ng/ml) for 12 hr in serum-free DMEM and GR-1 treatment was
washed off and discontinued prior to the addition of the endotoxins. Aliquots of medium (100
µl) were collected and frozen at -80°C (Figure 3.1, page 60).
3.2.5 Lactate Dehydrogenase Assay
Cultured amnion cells were plated to a density of 0.8x106 cells per well in a 24-well culture
plate. After incubation, cells were treated with 1% Triton X (Sangon Limited; Mississauga,
Ontario, Canada), control, GR-1 (1:20 dilution), LTA (10 ng/ml), or LPS (100 ng/ml). Medium
was collected after a 12 hr incubation period and 100 µl was transferred into an optically clear
96-well plate. Reaction mixture was prepared to a ratio of catalyst solution (BioVision; Mountain
View, California, United States of America) 1:4 to dye solution (BioVision) and 100 µl was
immediately added to corresponding wells. The plate was incubated in the dark at room
temperature for 30 mins and then was read using a spectrophotometer at 495 nm with a reference
wavelength of 600 nm. Lactate dehydrogenase (LDH) was measured in collected medium from
each treatment group in relation to Triton X treatment groups representing one hundred percent
LDH release.

51
3.2.6 Cytokine and Chemokine Measurement
3.2.6.1 Enzyme-Linked ImmunoSorbent Assay
Levels of IL-1β, IL-4, IL-6, IL-8, IL-10, TNFα, and GM-CSF were measured in culture media in
commercially available Enzyme-Linked ImmunoSorbet Assay (ELISA) kits (GM-CSF from R
and D Systems, Burlington, Ontario, Canada; All other cytokines from EBioscience; San Diego,
California, United States of America). A 96-well plate was coated with 100 µl/well capture
antibody diluted in coating buffer to a dilution of 1:250. The plates were incubated overnight at
4°C and were then washed with 300 µl of wash buffer (PBS+0.5% Tween 20) 5 times for 1
minute incubation periods each. Between each wash, plates were blotted on absorbent paper.
Assay diluent diluted at a ratio of 1:1 in Millipore filtered water was added to each well at a
volume of 200 µl and incubated at room temperature for 1 hr. After 5 washes, prepared standard
sets in 1:2 dilutions were added to the plate at a volume of 100 µl/well. Samples were added in
1:10 dilutions for IL-6 and IL-8 kits and 1:1 dilution for IL-1β, IL-4, GM-CSF and TNFα kits at
a volume of 100 µl to each well. Plates were incubated overnight at 4°C after which the plates
were washed an additional 5 times. 100 µl to each well of detection antibody diluted to 1:250 in
assay diluent was added followed by an incubation of 1 hr at room temperature. Plates were
washed 5 times and avidin-horseradish peroxidase diluted to 1:250 in assay diluent was added
100 µl to each well and incubated in the dark at room temperature for 30 mins. Plates were
washed 7 times before addition of 100 µl of substrate solution to each well for 15 mins at room
temperature. 50 µl of stop solution (H2SO4) was added to each well before reading the plate in a
spectrophotometer at 450 nm using 570 nm as a reference wavelength. The minimum detection
limits for IL-1β, IL-4, IL-6, IL-8, IL-10, GM-CSF, and TNFα kits were 4.0 pg/ml, 2.0 pg/ml, 2.0
pg/ml, 4.0 pg/ml, 4.0 pg/ml, 7.8 pg/ml, and 2.0 pg/ml respectively.
3.2.6.2 Bioplex Pro Human 27-Cytokine/Chemokine Assay
A Bioplex Pro Human 27-Cytokine/Chemokine Assay (BioRad Laboratories Incorporated;
Mississauga, Ontario, Canada) was performed to measure the levels of the following cytokines
and chemokines with minimum detection limits as indicated: Platelet-Derived Growth Factor
(1.33pg/ml), IL-1ra (2.96 pg/ml), IL-4 (0.21 pg/ml), IL-6 (1.54 pg/ml), IL-8 (1.53 pg/ml), IL-9
(1.58 pg/ml), IL-10 (1.48 pg/ml), IL-12 (2.19 pg/ml), IL-15 (1.75pg/ml), IL-17 (1.36 pg/ml),
Eotaxin (1.51 pg/ml), Fibroblast growth factor (0.66 pg/ml), G-CSF (1.76 pg/ml), GM-CSF (0.84

52
pg/ml), IFNγ (1.44 pg/ml), IFN-inducible Protein (2.51 pg/ml), MCP-1 (1.22 pg/ml), MIP-1α
(1.02 pg/ml), MIP-1β (0.92 pg/ml), RANTES (1.20 pg/ml), TNFα (5.72 pg/ml), Vascular
Endothelial Growth Factor (1.93 pg/ml), IL-1β (1.92 pg/ml), IL-2 (1.1 pg/ml), IL-5 (1.79 pg/ml),
IL-7 (1.67 pg/ml), and IL-13 (2.2 pg/ml). Samples were prepared in a total volume of 100 µl
with 0.5% bovine serum albumin (BSA; Sigma-Aldrich). After addition of BSA, samples were
vortexed and centrifuged at 1000 g for 10 mins and stored on ice. Stock standard solution was
reconstituted and 96 µl were added to 183 µl culture medium to a total volume of 275 µl. 128 µl
of each the reconstituted standard were mixed with 72 µl of standard diluent to prepare standard
1. Serial dilutions of 1:2 were prepared to create standards 2-8. Next, 5 µl of beads were added
to each well in a 50 µl of assay buffer. Excess fluid was aspirated and the plate was washed 2
times with wash buffer. Standards and samples were mixed on a vortex shaker, added to wells at
a volume of 50 µl and incubated on a shaker covered with foil at room temperature at 1000 g for
30 mins. The plate was washed 3 times and 25 µl of detection antibody, prepared to a 1:10
dilution in detection antibody diluent, were added to each well. The plate was then incubated on
a shaker covered with foil at room temperature at 1000 g for 30 mins. The plate was washed 3
times and 50 µl streptavidin-PE, prepared to a 1:100 dilution in assay buffer, were added to each
well. The plate was incubated on a shaker with foil for 10 mins, washed 3 times and 125 µl of
assay buffer were added to each well. The plate was shaken at 1 100 rpm for 30 secs before it
was read in a Bio-Plex reader according to manufacturer`s software.
3.2.7 Statistical Analysis
Data are presented as mean values ± standard error of the mean (SEM) where each n represents
individual amnion cultures from placentas received from patients meeting inclusion and
exclusion criteria as state previously. Statistical significance between groups was determined
using a One-way Anova statistical test if data was normally distributed followed by Student-
Newman-Keuls post-hoc analysis or using a non-parametric Kruskal-Wallis statistical analysis if
data did not have normality or equal variation using STAT32 version 2.0 statistical software.
Statistical significance was accepted at p<0.05.
3.3 Results
There were no significant differences in measured LDH levels between Lactobacillus rhamnosus
GR-1 (1:20) dilution, LTA (10 ng/ml) and LPS (100 ng/ml) and control (DMEM only; Figure

53
3.2, page 61). Triton X treatment, representing maximal cell death confirmed the validity of the
LDH assay (n=5). Therefore, treatments do not cause significant amounts of amnion cell death.
After 12 hr stimulation, all subsequent treatments were performed in serum-free DMEM.
Previous studies have shown that the presence of serum in medium increased basal levels of
prostaglandin output in cultured human amnion cells (Whittle et al., 1994). Thereby eliminating
the presence of serum, any confounding stimulants of either cytokines or prostaglandins was
prevented.
Throughout experiments in this study, amnion cells were treated with an LTA dose of 10 ng/ml
or an LPS dose of 100 ng/ml and medium was collected after 12 hours incubation with or
without pre-treatment of Lactobacillus rhamnosus GR-1 supernatant diluted to a ratio of 1:20
with DMEM. These values were chosen based on IL-6 and IL-8 time course and dose
optimization studies. LPS increased levels of IL-6 to maximal stimulation at 100 ng/ml where
IL-8 peaked at a dose of 10 ng/ml. The concentration of IL-8 was significantly increased by an
LPS dose of 100 ng/ml compared to control (Figure 3.3, page 62). Therefore, an LPS dose of 100
ng/ml was used for subsequent experiments. IL-6 concentrations showed two peaks at an LTA
dose of 10 ng/ml and 1000 ng/ml. However, in contrast, IL-8 concentrations appeared to peak at
an LTA dose of 10 ng/ml which then decreased with higher doses of endotoxin (Figure 3.3, page
63). Therefore, based on preliminary data, a dose of 10 ng/ml was used for LTA treatment.
To assess time course response, medium was collected and analyzed 4, 8, 12 and 24 hours after
LTA (10 ng/ml) or LPS (100 ng/ml) treatment (Figure 3.4, page 63). At 12 hr, both IL-6 and IL-
8 showed significant increases compared to 4 hr control by LPS stimulation. LTA also showed
increased IL-6 and IL-8 at 12 hr compared to 4 hr and 8 hr control. Although LTA significantly
increased IL-6 at 24 hr and LPS increased IL-8 at 24 hr compared to 4 hr and 8 hr controls, the
24 hr control levels of both cytokines appear to be elevated above the other control groups.
Therefore, a time point of 12 hr was chosen due to the elevated basal cytokine concentrations
observed at 24 hr and to the significantly increase in both IL-6 and IL-8 that was stimulated by
LTA and LPS. Additionally, numerous cytokines and chemokines showed peak stimulation at 12
hrs as assessed by Bioplex analysis including: MCP-1, RANTES, and IFNγ (data not shown).

54
Based on analysis of medium collected from amnion cells treated with various dilutions of
Lactobacillus rhamnosus GR-1 supernatant, the greatest differences in IL-6 concentration
compared to control were observed at a Lactobacillus rhamnosus GR-1 supernatant dilution of
1:20 (Figure 3.5, page 64). Lactobacillus rhamnosus GR-1 significantly increased IL-8
concentration compared to control, 1:50 and 1:100 treatments. Therefore, we chose a 1:20
dilution for all experiments. The same dilution of supernatant, 1:20 was used in previous studies
in studies on primary cultures of placental trophoblast and mouse macrophages (Kim et al., 2006;
Yeganegi et al., 2009; Yeganegi et al., 2010; Yeganegi et al., 2011). The lack of response for a
1:10 dilution make indicate a possible dose-dependency or threshold level required for the
unknown metabolite or metabolites in Lactobacillus rhamnosus GR-1 supernatant to exert its
effects on the unknown receptor present on the amnion. In addition, the bell curve shape
obtained suggests that the unknown signalling molecule present in Lactobacillus rhamnosus GR-
1 supernatant requires optimal dosage to exert its effects on target tissues. Competition may arise
if the concentration of this molecule is too high or homodimerization may occur preventing
interaction with the appropriate receptors. However, due to the unknown identity of the
molecule responsible for signalling in probiotic strains, these suggestions are merely speculation.
Two different methods were used to analyze cytokines and chemokines in these studies. To
ensure results were comparable between both methodologies, the medium collected from both a
pregnancy carrying a male fetus and one carrying a female fetus whose levels, as previously
measured by ELISA ,where closest to the preliminary mean were assessed once again in ELISA
and simultaneously in a Bioplex assay for IL-6 (Figure 3.6, page 65). Measured IL-6 levels were
highly correlated between the two methodologies (R2 –value = 0.8728).
Both LTA (10 ng/ml) and LPS (100 ng/ml) caused statistically significant increases in TNFα
concentrations in cultured amnion cell medium compared to control (Figure 3.7, page 66). There
was no significant change in TNFα with treatment of GR-1 (1:20) compared to control. The
increase in TNFα with either LPS or LTA treatment was attenuated when cultured amnion cells
were pre-treated with GR-1 in GR-1+LTA and GR-1+LPS treatment groups. No significant
changes were observed with other treatment groups. IL-1β (N=8), IL-10 (N=8) and IL-4 (N=6)
levels were undetectable across all treatment groups with lowest detectable limits (data not
shown).

55
Both LTA (10 ng/ml) and LPS (100 ng/ml) caused significant increases in IL-6 concentrations in
cultured amnion cell medium compared to control (Figure 3.8, page 67). GR-1 (1:20) caused a
significant increase in IL-6 concentration compared to control. GR-1+LTA and GR-1+LPS
treatments caused a significant increase in IL-6 concentration compared to control. No other
significant changes were observed across all other treatment groups.
Both LTA (10 ng/ml) and LPS (100 ng/ml) caused significant increases in IL-8 concentrations in
cultured amnion cell medium compared to control (Figure 3.9A, page 68). GR-1 (1:20) caused a
significant increase compared to control. GR-1+LTA and GR-1+LPS treatments caused a
significant increase compared to control group. RANTES concentrations were increased by GR-
1 compared to control (Figure 3.9B, page 68). As well, GR-1+LTA significantly increased
RANTES compared to control. GR-1+ LPS treatment significantly increased RANTES
compared to control. No other significant changes were observed between treatment groups.
MCP-1 was increased by GR-1 compared to control (Figure 3.9C, page 68). GR-1+LTA
significantly increased MCP-1 compared to control, LTA and LPS alone. As well, GR-1+LPS
treatment showed significant increases of MCP-1 compared to control, LTA and LPS alone. No
other significant changes were observed between treatment groups.
There was no significant change in IFNγ concentrations in medium collected from cultured
human amnion cells with either LTA (10 ng/ml) or LPs (100 ng/ml; Figure 3.10A, page 69). GR-
1 (1:20) significantly increased IFNγ concentration compared to control, LTA and LPS alone.
GR-1+LTA significantly increased IFNγ concentration compared to control, LTA and LPS
alone. GR-1+LPS significantly increased IFNγ concentration compared to control, LTA and
LPS alone. Neither GR-1 nor LTA and LPS caused significant increases in GM-CSF
concentration compared to control (Figure 3.10B, page 69). GR-1+LTA significantly increased
GM-CSF concentration compared to control, GR-1 and LTA alone. There was no significant
change in GM-CSF concentration with either LPS or GR-1+LPS treatments.
All data were additionally analyzed according to the gender of the fetus carried in the pregnancy
from which tissue was received. For each cytokine measured, the numbers for tissues derived
from a pregnancy carrying a female fetus versus a male fetus were as follows: TNFα (2 female: 3
male), IL-6 (4 female: 9 female), IL-8 (5 female: 6 female), RANTES (3 female: 5 male), MCP-
1 (3 female; 5 male), IFNγ (4 female: 6 male), and GM-CSF (3 female: 5 male). No obvious

56
trends were observed based on the gender of the fetus carried in pregnancies from which amnion
tissue was derived (data not shown).

57
3.4 Comment
The current study focused on the effects of Lactobacillus rhamnosus GR-1 on LTA and LPS
stimulated human amnion cells. In accordance with previous results, we have shown that
Lactobacillus rhamnosus GR-1 attenuated LTA and LPS-stimulated TNFα release from the
amnion similar to experiments performed on human placental trophoblasts and murine
macrophages (Yeganegi et al., 2010; Kim et al., 2006). Additionally, we demonstrated the
ability of Lactobacillus rhamnosus GR-1 to increase chemokine production, namely IL-8, MCP-
1 and RANTES as well as cytokines capable of activating recruited immune cells, IFNγ and GM-
CSF. Together, these results suggest that Lactobacillus rhamnosus GR-1 promotes a strong
chemotactic response in the amnion while minimizing the potential for an exaggerated pro-
inflammatory immune response to enable the amnion to defend against bacterial endotoxin
entrance into the fetal environment.
TNFα is a member of the pro-inflammatory triad alongside IL-1β and IL-6. As expected in
accordance with previous literature, IL-1β was not found above detectable limits in our culture
system. However, Lactobacillus rhamnosus GR-1 attenuated both LTA and LPS stimulated
TNFα production and increased IL-6 production to a similar level of LTA and LPS alone. TNFα
is only detectable in the amniotic fluid in the presence of an infection and/or inflammatory
response and is therefore indicative of an underlying infection (Romero et al., 1989c). The
influence of TNFα on the gestational tissues results in the activation of downstream pathways
leading to infection and/or inflammation-mediated preterm delivery. Alongside IL-1β, TNFα
elevates prostaglandin production in the gestational tissues leading to metalloproteinase
stimulation and subsequent cervical ripening and fetal weakening as collagen is degraded as well
as myometrial activation (Blumenstein et al., 2000; Bry et al., 1992; Pollard et al., 1993; Romero
et al, 1989b; Romero et al., 1989c). These events normally occur in parturition at term and early
activation by uterotonins, such as prostaglandins can lead to preterm delivery. By decreasing the
release of LTA and LPS-stimulated TNFα from the human amnion, Lactobacillus rhamnosus
GR-1 may significantly interfere with activation of infection and/or inflammation-mediated
processes associated with preterm labour in the amnion.
The amnion responds to Lactobacillus rhamnosus GR-1 with increased release of IL-6, IFNγ and
GM-CSF prior to LTA and LPS stimulation. IL-6, IFNγ, and GM-CSF all mediate the extent of

58
the pro-inflammatory response initiated by TNFα and IL-1β. These cytokines increase the
production of the antagonists to TNFα and IL-1β, namely the soluble TNFα receptor and IL-1
receptor antagonist (Aderka et al., 1989; Schindler et al., 1990; Tilg et al., 1994; Xing et al.,
1998). The soluble TNFα receptor binds and neutralizes TNFα. Similarly, IL-1 receptor
antagonist binds to IL-1 receptor preventing attachment of its agonist, IL-1β. Therefore,
Lactobacillus rhamnosus GR-1 may indirectly attenuate the effects of TNFα and IL-1β by
increasing cytokines that may stimulate the production of pro-inflammatory cytokine
antagonists.
The fetal membranes are situated between the amniotic fluid and the maternal gestational tissue.
As such, they act as a defensive barrier for pathogens, bacterial products and endotoxin from
reaching the fetal environment. We have demonstrated that Lactobacillus rhamnosus GR-1
stimulates a cytokine profile in the human amnion that may be capable of recruiting and
activating immune cells to the fetal membranes. Immune cells of monocyte and neutrophil origin
are localized to both the amnion and the chorion and neutrophilia in these tissues increases at
term (Witkin et al., 2011). Our studies show that Lactobacillus rhamnosus GR-1 increases
chemokines of both the CXC and CC family, namely IL-8 and MCP-1 and RANTES. IL-8 is
responsible for the recruitment and activation of neutrophils whereas MCP-1 and RANTES play
a role in the chemotaxis of monocytes. In addition, Lactobacillus rhamnosus GR-1 enhanced
MCP-1 production when challenged with LTA and LPS compared to endotoxin stimulation
alone. Therefore, Lactobacillus rhamnosus GR-1 may permit chemotaxis of immune cells,
especially monocytes to the amnion to allow for increased defense against bacterial endotoxin.
The activation state of monocytes/macrophages is dependent on the immunological signals
present in the microenvironment to which they are recruited. Although classically defined as
taking either an M1 or an M2 phenotypic state of activation, recent research has shown that the
activity of the macrophage is much more continuous. The M1 phenotype is characterized by
strong pro-inflammatory properties that encourage potent bacterial death and clearance (Mehta et
al., 2004; Sierra-Filardi et al., 2011). In comparison, the M2 phenotype encourages attenuated
pro-inflammatory responses and elevated anti-inflammatory responses (Mehta et al., 2004). In
this study Lactobacillus rhamnosus GR-1 increased the production of IFNγ and when pre-treated
prior to LTA and LPS stimulation, increased both IFNγ and GM-CSF release from amnion cells.

59
In conjunction with TNFα, IFNγ and GM-CSF promote the activation of macrophages to an M1
phenotype. However, Lactobacillus rhamnosus GR-1 attenuated LTA and LPS-stimulated TNFα
production in amnion cells. When stimulated with IFNγ in the absence of TNFα, immune cells
are activated to a phenotype characterized by its ability to phagocytose and produce anti-
microbial products with a less severe pro-inflammatory response (Mosser et al., 2008).
Therefore, Lactobacillus rhamnosus GR-1 promotes the amnion to produce cytokines capable of
activating immune cells to defend against pathogenic threats while minimizing the associated
pro-inflammatory response that may be deleterious to the prolongation of gestation.
In summary, Lactobacillus rhamnosus GR-1 increases the ability of the amnion to defend against
bacteria and bacterial products while minimizing the extent of the accompanying pro-
inflammatory response. However, caution must be taken in interpretation of these results for an
exaggerated immune cell response may also be deleterious to the prolongation of gestation if not
restricted. Chorioamnionitis is associated with preterm delivery that is characterized by increased
neutrophilia in the membranes. Therefore, further studies are necessary to determine whether or
not the regulated activation of monocytes/macrophages provided by cytokine and chemokines
up-regulated by Lactobacillus rhamnosus GR-1 in the amnion is conducive to defense against
pathogens as well as the prolongation of gestation. In addition, IL-6 has been shown to stimulate
prostaglandin production in the amnion (Kent et al., 1993; Mitchell et al., 1991). IL-6 is also
elevated in conjunction with chorioamnionitis and at term in amniotic fluid (Imseis et al., 1997).
As mentioned previously, prostaglandin elevation is associated with infection and/or
inflammation-mediated preterm birth. Lactobacillus rhamnosus GR-1 increases IL-6 to a level
comparable to LTA and LPS stimulation alone. Therefore, further experiments are necessary to
determine if a controlled immune cell response and IL-6 stimulation of prostaglandin production
is deleterious.

60
48
ho
urs
4
8 h
ou
rs
12
ho
urs
1
2 h
ou
rs
12
ho
urs
Pri
ma
ry
Cu
ltu
re
Med
ium
Co
llec
tion
Wash
Cu
ltu
re u
nti
l co
nfl
uen
t
(≤ 9
6 h
ou
rs)
Sta
rve
GR
-1 (
1:2
0)
LT
A
(10 n
g/m
l)
LP
S
(100 n
g/m
l)
Fig
ure
3.
1:
Exp
erim
enta
l d
esig
n
Pri
mar
y a
mnio
n c
ult
ure
s w
ere
gro
wn f
or
48 h
ours
and t
hen
was
hed
. A
fter
anoth
er 4
8 h
our
when
cel
ls r
each
ed c
onfl
uen
cy,
cult
ure
s w
ere
seru
m s
tarv
ed f
or
12 h
ours
. C
ells
wer
e pre
-tre
ated
wit
h L
act
obaci
llus
rham
nosu
s G
R-1
super
nat
ant
(GR
-1)
for
12 h
ours
. A
fter
12 h
ours
of
LT
A o
r L
PS
tre
atm
ent,
med
ium
was
coll
ecte
d.

61
Figure 3.2: The effect of treatments on amnion cell toxicity
Lactate dehydrogenase measured in the media of cultured human amnion cells treated with
control (DMEM), Lactobacillus rhamnosus GR-1 supernatant (GR-1, 1:20 dilution), lipoteichoic
acid (LTA, 10 ng/ml), and lipopolysaccharide (LPS, 100 ng/ml). Results are expressed as mean
values ± SEM relative to LDH release by Triton X-treated cell medium. Statistical significance
between groups is denoted with a different letter as assessed by One-way Anova followed by a
Student-Newman-Keuls post-hoc analysis (N=5, p<0.05).
a
b
b b b
0
20
40
60
80
100
120
Triton X control GR-1 LTA LPS
LDH
Lev
el (
%)

62
Figure 3.3: Lipoteichoic and lipopolysaccharide dose optimization
(A) IL-6 concentration (pg/ml; N=4, NS) and (B) IL-8 concentration (pg/ml; N=7, p<0.05) in media from amnion
cell cultures after treatment with control (DMEM), LTA or LPS doses of 0.1, 1, 10, 100, and 1000 ng/ml for 12
hours. Results are expressed as mean values ± SEM. Statistical significance within groups is denoted by different
letters as assessed by Kruskal-Wallis statistical test based on ranks followed by a Dunn’s post-hoc analysis. Circled
points show results at dosages chosen for subsequent experiments.
0
50
100
150
200
250
300
350
400
450
500
control 0.1 1 10 100 1000
IL-6
(p
g/m
l)
LTA
LPS
A
B
a
a
a
a b b
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
control 0.1 1 10 100 1000
IL-8
(p
g/m
l)
Endotoxin Dose (ng/ml)
LTA
LPS

63
Figure 3.4: Time course responses of Interleukin-6 and Interluekin-8
Histogram displaying (A) IL-6 concentration (pg/ml) and (B) IL-8 (pg/ml) in media from amnion cell cultures after
treatment with control (DMEM), LTA (10 ng/ml) or LPS (100 ng/ml) for periods of 4, 8, 12, and 24 hours. Results
are expressed as mean values ± SEM. Significance was determined using a Kruskal-Wallis statistical test based on
ranks followed by a Dunn’s post-hoc analysis. * denotes significance to 4h control, + denotes significance to 8h
control, ** denotes significance to 12h control, ^ denotes significance to 4h LTA (N=6, p<0.05).
* ^
* +
** ^ *
0
500
1000
1500
2000
2500
3000
3500
4000
4h 8h 12h 24h
IL-6
(p
g/m
l)
control
LTA
LPS
* +
* +
*
* + ^
^
* + ^
* +
0
500
1000
1500
2000
2500
3000
3500
4000
4500
4h 8h 12h 24h
IL-8
(p
g/m
l)
control
LTA
LPS
A
B

64
Figure 3.5: Lactobacillus rhamnosus GR-1 dilution optimization
(A) IL-6 concentration (pg/ml) and (B) IL-8 concentration (pg/ml) in media from amnion cell cultures
after treatment with control (DMEM), Lactobacillus rhamnosus GR-1 supernatant (GR-1; 1:10, 1:20,
1:50, 1:100 dilutions). Results are expressed as mean values ± SEM. Statistical significance within groups
is denoted with a different letter as assessed by Kruskal-Wallis statistical test based on ranks followed by
a Dunn’s post-hoc analysis (N=3, p<0.05).
0
200
400
600
800
1000
1200
control 1:10 GR-1 1:20 GR-1 1:50 GR-1 1:100 GR-1
IL-6
(p
g/m
l)
a
b,c c
b
b
0
500
1000
1500
2000
2500
3000
3500
4000
4500
control 1:10 GR-1 1:20 GR-1 1:50 GR-1 1:100 GR-1
IL-8
(p
g/m
l)
A
B

65
Figure 3.6: Correlation between ELISA and Bioplex assay methodologies
Correlation between ELISA measurements and Bioplex measurements for Interleukin-6 in
human amnion cultures. (n=2, R2=0.8728).
R² = 0.8728
0
500
1000
1500
2000
2500
3000
3500
0 500 1000 1500 2000 2500 3000 3500 4000 4500
ELIS
A IL
-6 (
pg/
ml)
Bioplex IL-6 (pg/ml)

66
Figure 3.7: The effects of Lactobacillus rhamnosus GR-1 on Tumor Necrosis Factor - α
concentration in human amnion cells
Histogram showing TNFα (pg/ml) concentrations in media from human amnion cell cultures.
Control (DMEM), GR-1 (1:20), LTA (10 ng/ml), LPS (100 ng/ml). Results are expressed as
mean values ± SEM. Statistical significance between groups is denoted with a different letter as
assessed by One-way Anova followed by a Student-Newman-Keuls post-hoc analysis (N=5,
p<0.05).
a a
b
a
c
a
0
50
100
150
200
250
300
350
400
450
500
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
TNFα
(p
g/m
l)

67
Figure 3.8: The effects of Lactobacillus rhamnosus GR-1 on Interleukin-6 concentration in
human amnion cells
Histogram showing IL-6 (pg/ml) concentrations in media from human amnion cell cultures.
Control (DMEM), GR-1 (1:20), LTA (10 ng/ml), LPS (100 ng/ml). Results are expressed as
mean values ± SEM. Statistical significance between groups is denoted with a different letter as
assessed by One-way Anova followed by a Student-Newman-Keuls post-hoc analysis (N=13,
p<0.05).
a
b
b b b
b
0
500
1000
1500
2000
2500
3000
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
IL-6
(p
g/m
l)

68
Figure 3.9: The effect of Lactobacillus rhamnosus GR-1 on chemokine concentrations in
human amnion cells
Histograms showing (A) IL-8 (pg/ml) (B) RANTES (pg/ml) and (C) MCP-1 (pg/ml) concentrations in media from
human amnion cell cultures. Control (DMEM), GR-1 (1:20), LTA (10 ng/ml), LPS (100 ng/ml). Results are
expressed as mean values ± SEM. Statistical significance between groups is denoted with a different letter as
assessed by One-way Anova followed by a Student-Newman-Keuls post-hoc analysis (N=11 for IL-8, N=8 for
RANTES, N=8 for MCP-1, p<0.05).
a
b
b
b
b
b
0
1000
2000
3000
4000
5000
6000
7000
8000
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
IL-8
(p
g/m
l)
a
b
a,b
b
a,b
b
0
20
40
60
80
100
120
140
160
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
RA
NTE
S (p
g/m
l)
a
b
a
b
a
b
0
500
1000
1500
2000
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
MC
P-1
(p
g/m
l)
A
B
C

69
Figure 3.10: The effects of Lactobacillus rhamnosus GR-1 on immune cell activating
cytokines
Histograms showing (A) IFNγ (pg/ml) and (B) GM-CSF (pg/ml) concentrations in media from
human amnion cell cultures. Control (DMEM), GR-1 (1:20), LTA (10 ng/ml), LPS (100 ng/ml).
Results are expressed as mean values ± SEM. Statistical significance between groups is denoted
with a different letter as assessed by One-way Anova followed by a Student-Newman-Keuls
post-hoc analysis (N=10, for IFNγ, N=8 for GM-CSF, p<0.05).
a
b
a
b
a
b
0
5
10
15
20
25
30
35
40
45
50
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
IFN
γ (p
g/m
l)
a a
a,b
b
a,b
a,b
0
20
40
60
80
100
120
140
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
GM
-CSF
(p
g/m
l)
A
B

70
Chapter 4
Effect of Lactobacillus rhamnosus GR-1 on Prostaglandin Output
and Prostaglandin Metabolizing Enzymes in Human Amnion Cells
at Term

71
Chapter 4
Effect of Lactobacillus rhamnosus GR-1 on Prostaglandin Output and Prostaglandin Metabolizing Enzymes in Human Amnion Cells at Term
4.1 Introduction
Preterm delivery is a major obstetrical concern as it accounts for up to 80% of all neonate
morbidities and mortalities. Its incidence has been rising over the past few decades and over
50% of all preterm births remain idiopathic despite advances in understanding its multifactorial
etiology. It is estimated that approximately 30% of all pre-term births are due to an underlying
infection and/or inflammation; however, the prevention of preterm birth by antibiotic treatment
is largely ineffective and in some cases increase its risk (Andrews et al., 2003; Okun et al.,
2005).
Bacterial vaginosis is an alteration to the endogenous vaginal microbiota where Lactobacillus
species decrease and pathogenic anaerobes increase and is known to increase the risk of preterm
birth by 40% (Hillier et al., 1995). Intrauterine infection can result from bacteria and/or bacterial
products gaining access to the intrauterine environment by ascent from the vagina through the
cervix (Goldenberg et al., 2000). The majority of microbes found in amniotic fluid samples
associated with intrauterine infections are of vaginal origin, leading us to study the effect of
lactobacilli on the amnion (Benirschke, 1960).
Infectious and/or inflammatory processes present in the intrauterine tissues are known to activate
a downstream cascade increasing prostaglandin production from the amnion, chorion and
decidua. In a state of infection, the chorionic 15-hydroxyprostaglandin dehydrogenase (PGDH)
metabolic barrier to prostaglandins produced in the amnion is decreased due to bacterial
endotoxin and cytokine release (Cheung et al., 1990; Sangha et al., 1994; Van Meir et al., 1996).
Bacterial endotoxin, bacterial products, cytokines, and chemokines increase net prostaglandin
output by up-regulating both phospholipase and prostaglandin H synthase (PGHS) -2 expression
in amnion cells (Kent et al., 1993; Mitchell et al., 1991; Pollard et al., 1993; Romero et al.,
1988a; Romero et al., 1989a). A net increase of prostaglandin production accompanies infection
and/or inflammation due to decreased metabolism and increased production. Prostaglandins

72
activate downstream pathways that result in a myometrial activation, cervical ripening and
membrane weakening.
Although numerous investigators have studied the role of antibiotics in preventing preterm birth,
their use has been shown to be largely ineffective and in some cases increases the risk of preterm
labour (Andrews et al., 2003; Okun et al., 2005). Probiotics are defined as, “live microorganisms
which when administered at adequate amounts confer a health benefit on the host” and have been
identified as a potential treatment alternative (FAO and WHO, 2001; Reid, 1999). Lactobacillus
rhamnosus GR-1 is capable of colonizing and restoring the microbiota of the vagina and
modulation of prostaglandin production enzymes in the gestational tissues (Reid et al., 2003;
Yeganegi et al., 2009).
In placental trophoblasts, Lactobacillus rhamnosus GR-1 increased expression of the
prostaglandin metabolizing enzyme, PGDH in pregnancies carrying a female but not male fetus
(Yeganegi et al., 2009). Also, Lactobacillus rhamnosus GR-1 reduced the LPS-induced increase
in PGHS-2 in placental trophoblast cells from pregnancies carrying a male but not female fetus
(Yeganegi et al., 2009). One of the major products of the amnion is prostaglandins. In a state of
infection and/inflammation, where metabolic barriers become less effective, the amnion
contribution to prostaglandin elevation may be detrimental to the maintenance of gestation.
Therefore, we hypothesized that Lactobacillus rhamnosus GR-1 will prevent alterations to
prostaglandin production and key prostaglandin metabolizing enzymes in amnion cells in
response to bacterial endotoxin stimulation.
4.2 Methodology
4.2.1 Placenta Collection
Placenta collection was performed as described in Chapter 3 (Page 48)
4.2.2 Amnion Mixed Epithelial and Mesenchymal Cell Culture
Amnion mixed epithelial and mesenchymal cell cultures were performed as described in Chapter
3 (Page 48-49). Cells were plated to a density of 8x106 cells per well in 60 mm culture dishes.
Protein was collected from cell lysates in RIPA lysis buffer + protease inhibitors (Roche
Diagnostics) and frozen at -80°C.

73
4.2.3 Lactobacilli Preparation
Lactobacillus rhamnosus GR-1 was grown as described in Chapter 3 (Page 49).
4.2.4 Treatment Protocol
Treatment was performed as described in Chapter 3 (Page 50; Figure 4.1, page 83).
4.2.5 Lactate Dehydrogenase Assay
LDH assay was performed as described in Chapter 3 (Page 50).
4.2.6 Prostaglandin E2 Measurement
4.2.6.1 Enzyme Immuno-Assay
Levels of PGE2 were measured using commercially available quantitative enzyme immuno-assay
(EIA) kits (Cayman Chemical; Ann Arbor, Michigan, United States of America). EIA buffer was
added to non-specific binding wells and maximum binding wells to a volume of 100 µl and 50 µl
respectively. Standards 2-8 were prepared in 1:2 dilutions in EIA buffer and standard 1 of 1000
pg/ml, and 50 µof each were added to wells. Samples were added in a dilution of 1:10 in EIA
buffer to a final volume of 50 µl to wells. Prostaglandin E2 AchE Tracer was reconstituted in 6
ml EIA buffer and 50 µl were added to all wells except Total Activity and Blank wells.
Prostaglandin E2 Monoclonal Antibody was reconstituted in 6ml EIA Buffer and 50 µl were
added to each well except Total Activity, Non-Specific Binding wells, and blank wells. Plates
were incubated overnight at 4°C. Plates were washed 5 times in 300 µl of Wash Buffer. Ellman’s
reagent colour developing solution was reconstituted in 20 ml of Millipore-filtered water and 200
µl were added to each well to develop colour reaction. Prior to incubation at room temperature
in the dark, 5 µl of Prostaglandin E2 Acetylcholine Esterase Tracer was added to the Total
Activity well. The plate was read when the 405 nm absorbance of the Maximum Binding wells
minus the 450 nm absorbance of the Blank wells was greater than 0.3, a time ranging from 60 to
90 minutes in a spectrophotometer (Tecan Infinite M200). The minimum detection limit for
PGE2 was 80% B/Bo of 15 pg/ml.

74
4.2.7 Prostaglandin Metabolizing Enzyme Expression Measurement
4.2.7.1 Western Blot Analysis
Protein samples in RIPA were thawed and immediately mixed on a vortex shaker before
incubation for 20 mins on ice. This was repeated 3 times followed by centrifugation at 4°C for
30 mins at 1000 g. Supernatant was then collected. Protein concentration was calculated using a
Bio-Rad Protein Assay with BSA serial dilutions as a standard. Samples were frozen at -80°C
until further analysis. Thirty micrograms of protein extract were mixed with specific volumes of
RIPA lysis buffer and 5µl of loading dye to a total volume of 25 µl.
Protein samples were run on a 10% sodium dodecyl sulphate poly-acrylamide gel electrophoresis
alongside a Pageruler pre-stained protein ladder (Fermentas; Burlington, Ontario, Canada). After
electrophoresis, proteins were transferred to a nitrocellulose membrane at 100 V for 90 mins.
Protein transfer was visually assessed using Ponceau S dye followed by 3 washes of 5 mins each
in PBS with 0.1 Tween 20 (PBST, Sigma-Aldrich). Membranes were blocked overnight at 4°C
in a blocking solution consisting of 5% non-fat skim milk powder (BioShop Canada
Incorporated; Burlington, Ontario, Canada) dissolved in PBST.
Membranes were incubated separately on a rocking platform in primary antibody diluted in
blocking solution for 1 hr at room temperature at the following dilutions: β-actin monoclonal
antibody 1:1000 (Sigma-Aldrich), TLR4 monoclonal antibody 1:250 (Cayman Chemicals),
PGHS-2 monoclonal antibody 1:500 (Sigma-Aldrich), PGDH monoclonal antibody 1:500
(Sigma-Aldrich), cPGES polyclonal antibody 1:500 (Cayman Chemicals), mPGES1 polyclonal
antibody 1:500 (Cayman Chemicals), and mPGES2 polyclonal antibody 1: 500 (Cayman
Chemicals). Membranes were washed 5 times for 5 mins in PBST. Secondary anti-mouse IgG
(anti-PGHS-2, anti-β-actin; GE Healthcare; Little Chalfont, Buckinghamshire, United Kingdom)
and anti-rabbit IgG (anti-cPGES, anti-mPGES1, anti-mPGES2; GE Healthcare), horseradish
peroxidase linked whole antibody 1:4000 dilution incubation on a rocking platform was
performed for 1 hr at room temperature at a dilution of 1:4000 for 1 hr. Membranes were
washed 6 times for 5 mins in PBST. Membranes were incubated in enhanced
chemiluminescence (ECL; Perkin Elmer, Waltham Massachusetts, United State of America)
substrate for 1 min or in ECL Advance (Perkin Elmer) for 2 mins and were exposed to Kodak X-
OMAT Blue XB film for 30 secs, 1 mins, 5 mins, 10 mins and 15 mins. Optical densities were

75
measured using Scion Imaging Software. Optical densities of proteins were divided by optical
density of β-actin to standardize for protein loading. These relative densities were then used for
statistical analysis using Scion Image computer analysis software version Beta 4.0.2.
4.2.8 Placental Trophoblast Cell Culture
Placental trophoblast cells were isolated based on previously established primary culture
protocols (Sun et al., 1997). Briefly, placental tissue was separated from fetal membranes and
washed in saline. Approximately 60 g of villous placental tissue was cut and digested in a 37°C
incubator with 0.125% trypsin and 0.02% deoxyribonuclease-I (Sigma-Aldrich) in DMEM for
30 mins. Supernatant was them removed and discarded as tissue was incubated in 0.125% trypsin
in a 37°C incubator two more times for 30 mins. Supernatant was collected after the second and
third digestion and centrifuged at 1700g for 10 mins and resuspended in DMEM. The pooled
supernatant was then filtered through a 200 μm pore size nylon gauze filter. Cells were then
purified using a Percoll gradient (35, 40, 65, 70 increments) and centrifuged for 20 mins at
2500g. Placental trophoblasts were removed from the appropriate Percoll band and resuspended
in culture medium. Cells were plated to a density of 106 in 60 mm culture dishes.
4.2.8.1 Treatment
Cells were incubated for 72 hr in 37°C 5% CO2 and 95% air in order to form a syncytium after
which cells were serum starved for 12 hr. Cells were then treated with Lactobacillus rhamnosus
GR-1 supernatant for 12 hours prior to LPS (100 ng/ml) incubation for 8 hr. Protein was then
collected from cell lysates in RIPA + protease incubator and frozen until use at -80°C.
4.2.8.2 Western Blot Analysis
Proteins were extracted and run on a Western Blot Analysis as previously described (Chapter 4,
page 73). 25 µg of protein was loaded as previously described (Chapter 4, page 73). Primary
antibodies were used to the following dilutions: β-actin monoclonal antibody 1:1000 and PGHS-
2 monoclonal antibody 1:500 (Sigma-Aldrich). Secondary antibodies were used to the following
dilutions: anti-mouse IgG (anti-PGHS-2, anti-β-actin) horseradish peroxidase linked whole
antibody 1:4000 dilution.

76
4.2.9 Statistical Analysis
PGE2 data are presented as average values ± SEM. Protein data are presented as average values ±
SEM relative to control. Statistical significance between groups was determined using a Kruskal-
Wallis statistical test based on ranks followed by a Dunn’s post-hoc analysis using STAT32
version 2.0 statistical software. A non-parametric Kruskal-Wallis statistical analysis was
performed based on the lack of normality and equal variation in the data of Chapter 4 as is
required for parametric One-way Anova statistical analysis. Statistical significance was accepted
at p<0.05.
4.3 Results
Out of the stimulants tested and at the amounts stated, only IL-1β (100 ng/ml) produced a
significant increases in PGE2 release compared to control treatment of DMEM alone in human
amnion cells (Figure 4.2, page 84). No significant differences in PGE2 production between LPS
of 1000 ng/ml, LPS of 100 ng/ml compared to control were observed. Dexamethasone had no
effect on PGE2 production. Both IL-1β and dexamethasone, in mesenchymal cells only, at 100
ng/ml have previously been shown to stimulate prostaglandin production in cultured human
amnion cells at term (Romero et al., 1989b, Whittle et al., 2000). MRS (1:20 dilution), the
medium in which Lactobacillus rhamnosus GR-1 was cultured was tested as a control for
supernatant treatment and did not cause a significant change in PGE2 release.
Placental trophoblast cell cultures were performed to confirm the duplication of previously
established results on LPS stimulation of PGHS-2 expression (data not shown). This was
performed to not only confirm culture technique but also to confirm the ability of LPS to modify
prostaglandin metabolizing and synthesizing enzyme expression. Placental trophoblasts were
cultured and then treated according to the following groups: control (DMEM), GR-1 (1:20
dilution), LPS (100 ng/ml), and GR-1+LPS. Final results were similar to findings by Yeganegi et
al. 2009. Although significance was not assessed due to an n-value of 2, similar trends were
observed as in experiments performed by Yeganegi et al., namely LPS increased PGHS-2
expression that was prevented by pre-treatment with GR-1.
PGE2 was significantly increased by GR-1 (1:20) alone and when pre-treated prior to endotoxin
stimulation with both LTA (10 ng/ml) and LPS (100 ng/ml) compared to control (DMEM)

77
treatment (Figure 4.3, page 85). In addition, GR-1+LTA stimulation caused a significant increase
in PGE2 concentration compared to control and LTA. GR-1+LPS stimulation caused a
significant increase in PGE2 concentration compared to control and LTA as well as a significant
decrease to GR-1. However, without pre-treatment with GR-1, neither LTA nor LPS caused
significant changes in PGE2 concentration compared to control.
To determine whether changes in PGE2 output were due to prostaglandin enzyme expression
modification, PGDH, PGHS-2, cPGES, mPGES1, and mPGES2 levels were measured across all
treatment groups. PGDH was found at very low levels across all treatment groups as was
expected from previously performed experiments indicating little or no PGDH is expressed in
the human amnion (N=6; data not shown). No significant differences in PGHS-2, cPGES or
mPGES1 levels were observed between treatment groups: control (DMEM only), GR-1 (1:20
dilution), LTA (10 ng/ml), GR- 1+LTA, LPS (100 ng/ml), and GR-1+LPS (Figure 4.4, 4.5, 4.6,
pages 86-88). Interestingly, a significant increase in mPGES2 expression was observed between
GR-1 and GR-1+LTA to control treatment groups (Figure 4.7, page 89). No other significant
changes were observed between all other treatment groups.
Expression of the different isoforms of PGES, cPGES, mPGES1, and mPGES2 were correlated
with PGE2 release from human amnion cells (Figure 4.8, page 90). An R2 –value of 0.6321,
0.6249, and 0.8214 exist for cPGES, mPGES1, and mPGES2 with PGE2 respectively.
All data were additionally analyzed according to the gender of the fetus carried in the pregnancy
from which tissue was received. For PGE2 and enzymes measured, the numbers for tissues
derived from a pregnancy carrying a female fetus versus a male fetus were as follows: PGE2 (4
female: 8 male), PGHS-2 (2 female: 4 male), cPGES (2 female: 5 male), mPGES1 (3 female: 5
male), mPGES2 (3 female: 6 male). No obvious trends were observed based on the gender of the
fetus carried in pregnancies from which amnion tissue was derived (data not shown).

78
4.4 Comment
The focus of this study was to determine the influence of Lactobacillus rhamnosus GR-1
supernatant on amnion prostaglandin output and modulation of the enzymes involved in their
metabolism. We have shown that Lactobacillus rhamnosus GR-1 significantly increased PGE2
release in human amnion cells. This response appears to be slightly attenuated in amnion cells
stimulated with either LTA or LPS after Lactobacillus rhamnosus GR-1 pre-treatment compared
to Lactobacillus rhamnosus GR-1 alone. From these results, we suggest that enhanced
prostaglandin production caused by Lactobacillus rhamnosus GR-1 permits the amnion to: 1)
Control the extent of a cytokine-induced immune cell response and 2) Provide a means of
pregnancy termination if the threat of bacterial endotoxin exposure becomes detrimental to the
health of the fetus.
Surprisingly, in our culture system we were unable to demonstrate enhanced prostaglandin
release by LPS stimulation in amnion cells as has been shown previously (Bennett, et al., 1987;
Reisenberger et al., 1998; Romero et al., 1988a; Shon et al., 2002). Consequentially, we
performed several subsequent experiments to verify our results. These included verification of
the viability of our cell culture as well as the activity of LPS and expression of its receptor,
TLR4. As well, we verified the ability to produce and secrete prostaglandins and performed
parallel experiments in a different cell culture, placental trophoblast cells.
Cells release prostaglandins during apoptosis. If a high proportion of cultured cells were in an
apoptotic phenotype at the time our samples were taken, a change in prostaglandin production by
LPS may be masked by elevated levels at baseline prior to stimulation. However, an LDH assay
indicated little cell death compared to those cells treated with Triton X, a compound that
solubilizes cell membranes to release into the cell medium a maximal level of LDH, an enzyme
regularly bound to the mitochondrial membrane. In this assessment, the proportion of LDH in the
medium collected across treatment groups is representative of the proportion of cell death. All
treatment groups including LPS at a dose of 100 ng/ml exhibited low LDH levels indicating little
cell death. Therefore, the lack of prostaglandin production stimulated by LPS was not due to an
elevated baseline release of prostaglandins due to cell death.
One potential cause for the lack of LPS-stimulated prostaglandin release in our cultured human
amnion cells may be a lack of biological activity of our LPS. It is well established that LPS

79
treatment of cultured human amnion cells increases the production of many cytokines. In our
system, we demonstrated that in amnion IL-6, IL-8 and TNFα release were stimulated by LPS at
a dose similar to previous experiments of 100 ng/ml. Therefore, the activity of LPS used in our
experiments shows biological activity capable of stimulating increased release similar to that
previously shown by other investigators.
LPS is a bacterial endotoxin that is recognized by cell membrane receptors of the body known as
TLRs. The main receptor for LPS is TLR4 that is expressed on many immune cells and tissues,
including the amnion. We also determined that TLR4 is expressed in our culture system
verifying the presence of the receptor for LPS on amnion cells present in our cell culture system.
Prostaglandin release from the amnion is stimulated by many factors including bacterial products
such as endotoxins, pro-inflammatory cytokines as well as glucocorticoids (Pollard et al., 1993;
Romero et al., 1988a; Romero et al., 1989b; Whittle et al, 2000; Zaga et al., 2004). Studies have
shown that an LPS dose within a range of 10 ng/ml to 1000 ng/ml stimulated prostaglandin
production and release from human amnion cells cultured with a similar method, incubation
period and treatment time as used in our experiments (Romero et al., 1988a, Thiex et al., 2009).
However, here both 100 ng/ml and 1000 ng/ml of LPS were unable to increase prostaglandin
production significantly compared to control groups. Similarly, dexamethasone at 100 ng/ml was
unable to increase prostaglandin production. It has been previously demonstrated that
mesenchymal amnion cells produce greater amounts of PGE2 compared to epithelial amnion
cells (Whittle et al., 2000). Dexamethasone may not have shown stimulation in mixed amnion
cell cultures in this study if the proportion of epithelial cells were much greater than
mesenchymal cells. The proportion of epithelial and mesenchymal cells obtained from the
culture techniques previously described, were not mixed to a ratio similar to that which exists in
vivo. Therefore a greater proportion of epithelial cells may account for the lack of
dexamethasone stimulation of PGE2 observed. Previous studies indicate that IL-1β increases
prostaglandin production and release in cultured human amnion cells through increases in
phospholipase and PGHS-2 expression at a dose of 100 ng/ml (Blumenstein et al., 2000; Bry et
al., 1992; Pollard et al., 1993; Romero et al., 1989a; Romero et al., 1989b). Similarly, we showed
that IL-1β at a dose of 100 ng/ml increase prostaglandin release from our cultured human amnion
cells (Romero et al., 1989b). Therefore, we have shown that in our system, prostaglandin release
can be stimulated by exogenous factors as was previously reported.

80
Placental trophoblasts are another gestational tissue responsive to LPS. Previously, our
laboratory has demonstrated the ability of LPS to modulate enzymes involved in prostaglandin
metabolism in cultured human placental trophoblast cells (Yeganegi et al., 2009). To verify not
only the activity of LPS by attempting to parallel previously established results, we cultured
placental trophoblast cells to verify culture and treatment technique. Previously, it had been
shown that LPS-stimulated PGHS-2 expression that was minimized by Lactobacillus rhamnosus
GR-1 pre-treatment (Yeganegi et al., 2009). These results were duplicated demonstrating that the
LPS used in amnion cell experiments is biologically active and verified culture techniques.
In human placental trophoblasts, Lactobacillus rhamnosus GR-1 supernatant prevented changes
in PGHS-2 expression caused by LPS in placentae derived from pregnancies carrying a male
fetus (Yeganegi et al., 2009). As well, in placentae derived from pregnancies carrying a female
fetus, Lactobacillus rhamnosus GR-1 supernatant increases PGDH expression to a higher degree
than in placentae derived from pregnancies carrying a male fetus (Yeganegi et al., 2009). In
contrast Lactobacillus rhamnosus GR-1 stimulated PGE2 release from amnion cells in this study.
In the amnion Lactobacillus rhamnosus GR-1 significantly increased mPGES2 expression alone
and when pre-treated prior to LTA stimulation. Together, with the trends observed in the
expression profiles of cPGES and mPGES1, the changes observed in mPGES2 may partly
account for the effects of Lactobacillus rhamnosus GR-1 on human amnion prostaglandin
production. Placental trophoblasts and the amnion have diverse roles throughout pregnancy and
are situated at anatomically distinct locations. Therefore it is not surprising that Lactobacillus
rhamnosus GR-1 exerts different effects on prostaglandin production and prostaglandin
metabolizing and synthesizing enzymes in placental trophoblasts and the amnion based on the
role of each tissue in gestation.
The amnion is the most biologically active producer of prostaglandins amongst the gestational
tissues at the onset of labour (Kinoshitak, 1984). Not surprisingly, numerous stimulants of
prostaglandin release from human amnion cells have been identified. These include but are not
limited to 1) bacterial products and endotoxins including phospholipases, LTA and LPS, 2) Pro-
inflammatory cytokines such as IL-1α, IL-1β, TNFα, and IL-6, 3) Chemokines, such as MIP-1α,
4) Anti-inflammatory cytokines, including IL-10 and IL-1ra, and 5) Glucocorticoids in
mesenchymal cells (Dudley et al., 1996; Furuta et al., 2000; Mitchell et al., 1991; Reisenberger
et al., 1998; Romero et al., 1989b; Romero et al., 1989c). Three of these stimulants stand out as

81
having paradoxically stimulatory roles in the amnion compared to their well-established anti-
inflammatory roles in other tissues: IL-1ra, IL-10, glucocorticoids (Economopoulos et al., 1996;
Gibb et al., 1990; Mitchell et al., 2004; Potestio et al., 1988). We have demonstrated that
Lactobacillus rhamnosus GR-1 can be added to this list of prostaglandin stimulants in cultured
human amnion cells.
The amnion increases prostaglandin production in response to stimulants, such as glucocorticoids
that normally prevent their production in other tissues. Therefore, one might speculate that
prostaglandins may play an important function in the role of the amnion, possibly to enhance the
barrier provided by the membranes against pathogenic bacteria and their products from reaching
the amniotic fluid. It is well-established that elevated levels of prostaglandins stimulate increased
expression of metalloproteinases that cause weakening of the fetal membranes and cervical
ripening ultimately leading to preterm delivery. Therefore, uncontrolled elevated levels of
prostaglandins stimulated by bacterial endotoxins and cytokines may lead to an elevated risk of
preterm birth. However, low or absent prostaglandin levels have been associated with
exaggerated immune responses due to the immunomodulatory role this family of eicosanoids
exert (Goodwin et al., 1983).
Arachidonic acid metabolites, including prostaglandins have significant roles in suppressing or
terminating immune responses (Chizzolini et al., 2009). An established function of PGE2 in
particular, is modulation of cytokine production in macrophages (Nataraj et al., 2001). PGE2 acts
to shift the activated macrophage phenotype from a strong pro-inflammatory response to a more
anti-inflammatory response (Harris et al., 2002). This shift in phenotype is characterized by
prevention of LPS stimulated release of type 1 immune response cytokines such as TNFα and IL-
12 while elevating the release of type two response cytokines such as IL-4 and IL-10 (Betz et al.,
1991, Katamura et al., 1995, Scales et al., 1989, Shinomiya et al., 2001, van der Pouw Kraan et
al., 1995). A similar role of PGE2 on the amnion or any other tissue has not been shown, but this
effect is modulated through EP4 (Nataraj et al., 2001). EP4 is expressed on the amnion; therefore
a similar modulation of cytokine response may occur due to the actions of PGE2 in the amnion. It
is important to consider that a metabolic barrier in the chorion expressing high levels of PGDH
may be maintained allowing the deleterious effects of increased prostaglandin production to be
minimized amongst the other gestational tissues while providing the immunoregulatory effects at
the amnion. Therefore, Lactobacillus rhamnosus GR-1 stimulation of prostaglandins in the

82
amnion tissue may aid in maintaining the anti-microbial defenses properties inherent to this layer
of the fetal membrane.
The amnion is the layer of tissue in closest proximity to the fetal environment. Therefore, the
ability of the amnion to defend against pathogenic threats may be the deciding factor if bacteria
and bacterial endotoxin reach the fetal environment causing developmental damage. The amnion
possesses inherent anti-microbial, anti-bacterial and anti-viral properties that prevent infection
during various obstetrical events. Although elevated levels can lead to activation of downstream
pathways associated with preterm labor, the production of prostaglandins may be crucial in the
defense provided by the amnion against pathogenic threats. Prostaglandins modulate the immune
response by minimizing pro-inflammatory cytokines from recruited immune cells. Therefore,
increased prostaglandin release caused by Lactobacillus rhamnosus GR-1 may contribute to the
overall ability of the amnion to defend against infections by controlling the extent of the
inflammatory response if levels of prostaglandins are also controlled and maintained.
Conversely, increased prostaglandin production by Lactobacillus rhamnosus GR-1 may provide
the amnion with the ability to terminate the pregnancy through increased prostaglandin
production in order to protect the fetus from exposure to bacterial endotoxin given that the threat
of deleterious effects associated with infection may be too great to allow for prolonged gestation.
Further experiments are required to differentiate between these potential effects of Lactobacillus
rhamnosus GR-1 on amnion production of prostaglandins.

83
Fig
ure
4.
1:
Exp
erim
enta
l d
esig
n
Pri
mar
y a
mnio
n c
ult
ure
s w
ere
gro
wn
for
48 h
ou
rs a
nd t
hen
was
hed
. A
fter
anoth
er 4
8 h
our
wh
en
cell
s re
ached
con
fluen
cy,
cult
ure
s w
ere
seru
m s
tarv
ed f
or
12 h
ours
. C
ells
wer
e pre
-tre
ated
wit
h
Lact
obaci
llus
rham
nosu
s G
R-1
super
nat
ant
(GR
-1)
for
12 h
ours
. A
fter
12 h
ours
of
LT
A o
r L
PS
trea
tmen
t, p
rote
in f
rom
cel
l ly
sate
s w
as c
oll
ecte
d.
48
hou
rs
48
ho
urs
1
2 h
ou
rs
12
ho
urs
1
2 h
ou
rs
Pri
ma
ry
Cu
ltu
re
Pro
tein
Co
llec
tion
fro
m C
ell
Ly
sate
s
Wash
Cu
ltu
re u
nti
l co
nfl
uen
t
(≤ 9
6 h
ou
rs)
Sta
rve
GR
-1 (
1:2
0)
LT
A
(10
ng/m
l)
LP
S
(10
0 n
g/m
l)

84
Figure 4.2: The effect of IL-1β, dexamethasone (Dex), lipopolysaccharide (LPS) and de
Man, Rogosa, and Sharpe (MRS) on prostaglandin production in human amnion cells
Histogram showing PGE2 output in media from human amnion cell cultures by various treatments.
Control (DMEM), IL-1β (100 ng/ml), Dexamethasone (100 ng/ml), Lipopolysaccharide (100 and 1000
ng/ml), and de Man, Rogosa, and Sharpe (1:20 dilution). Results are mean values ± SEM. Statistical
significance within groups is denoted with a different letter as assessed by Kruskal-Wallis statistical test
based on ranks followed by a Dunn’s post-hoc analysis (N=4, p<0.05).
a
b
a
a a
a
0
200
400
600
800
1000
1200
1400
1600
control Il-1β Dex LPS100 LPS1000 MRS
PG
E 2 (
pg/
ml)

85
Figure 4.3: The effect of Lactobacillus rhamnosus GR-1 on Prostaglandin E2 concentration
in medium from cultured human amnion cells
Histogram showing PGE2 in media collected from human amnion cell cultures. Control
(DMEM), LTA (10 ng/ml), LPS (100 ng/ml), GR-1 (1:20). Results are expressed as mean
values ± SEM. Statistical significance within groups is denoted with a different letter as assessed
by Kruskal-Wallis statistical test based on ranks followed by a Dunn’s post-hoc analysis (N=12,
p<0.05).
a
b
a
b
a
a,b
0
200
400
600
800
1000
1200
1400
1600
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
PG
E 2 (
pg
/ml)

86
PGHS-2 72 kDa
β-actin 42 kDa
Figure 4.4: The effects of Lactobacillus rhamnosus GR-1 on PGHS-2 expression in human
amnion cells
Histogram showing (A) PGHS-2 expression fold increase relative to control in human amnion
cell cultures. Control (DMEM), LTA (10 ng/ml), LPS (100 ng/ml), GR-1 (1:20). Results are
expressed as mean values ± SEM. Statistical significance assessed using a Kruskal-Wallis
statistical test based on ranks followed by a Dunn’s post-hoc analysis (B) Relative Optical
Density (ROD) of PGHS-2/β-actin western blot (N=6, NS).
A
B
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
PG
HS-
2/β
-act
in O
D (
rela
tive
to
co
ntr
ol)
Co
ntr
ol
GR
-1
LTA
GR
-1+
LTA
LPS
GR
-1+L
PS
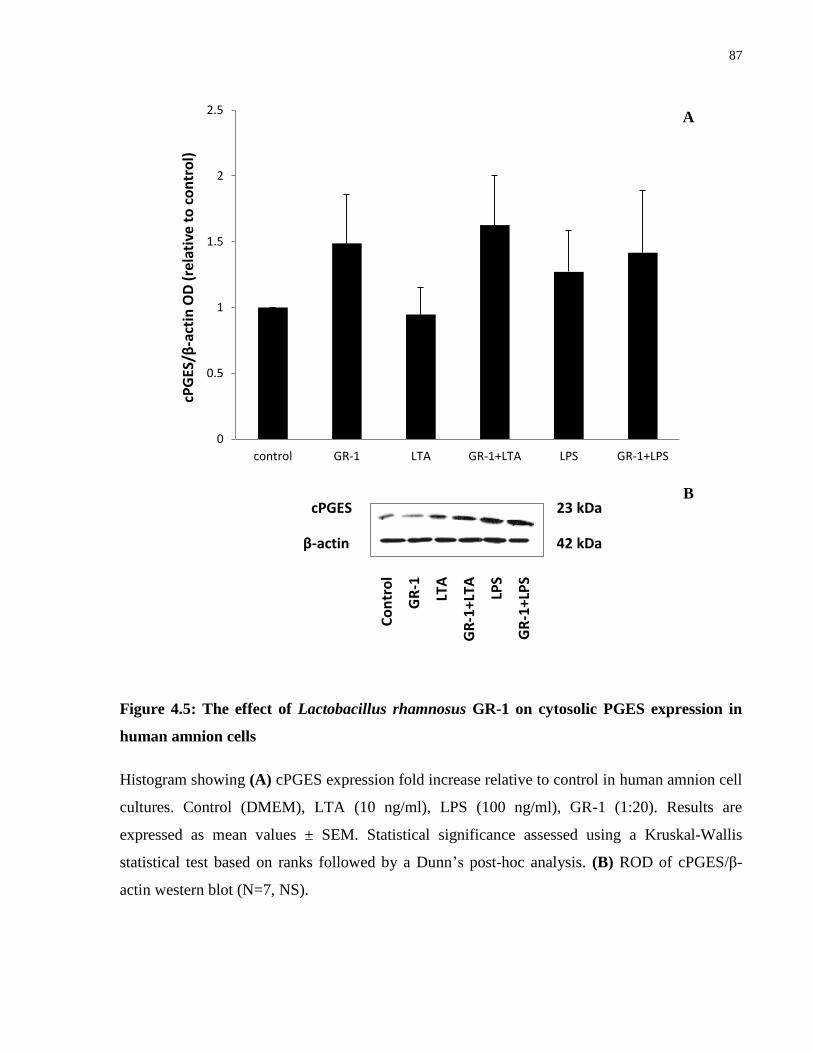
87
cPGES 23 kDa
β-actin 42 kDa
Figure 4.5: The effect of Lactobacillus rhamnosus GR-1 on cytosolic PGES expression in
human amnion cells
Histogram showing (A) cPGES expression fold increase relative to control in human amnion cell
cultures. Control (DMEM), LTA (10 ng/ml), LPS (100 ng/ml), GR-1 (1:20). Results are
expressed as mean values ± SEM. Statistical significance assessed using a Kruskal-Wallis
statistical test based on ranks followed by a Dunn’s post-hoc analysis. (B) ROD of cPGES/β-
actin western blot (N=7, NS).
0
0.5
1
1.5
2
2.5
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
cPG
ES/β
-act
in O
D (
rela
tive
to
co
ntr
ol)
A
B
Co
ntr
ol
GR
-1
LTA
GR
-1+
LTA
LPS
GR
-1+
LPS

88
mPGES1 16 kDa
β-actin 42 kDa
Figure 4.6: The effect of Lactobacillus rhamnosus GR-1 on microsomal PGES1 expression
in human amnion cells
Histogram showing (A) mPGES1 expression fold increase relative to control in human amnion
cell cultures. Control (DMEM), LTA (10 ng/ml), LPS (100 ng/ml), GR-1 (1:20). Results are
expressed as mean values ± SEM. Statistical significance assessed using a Kruskal-Wallis
statistical test based on ranks followed by a Dunn’s post-hoc analysis. (B) ROD of mPGES1/ β-
actin (N=8, NS).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
mP
GES
1/β
-act
in (
OD
rel
ativ
e to
co
ntr
ol)
B
A
Co
ntr
ol
GR
-1
LTA
GR
-1+
LTA
LPS
GR
-1+L
PS

89
mPGES2 33 kDa
β-actin 42 kDa
Figure 4.7: The effects of Lactobacillus rhamnosus GR-1 on microsomal PGES2 expression
in human amnion cells
Histogram showing (A) mPGES2 expression fold increase relative to control in human amnion
cell cultures. Control (DMEM), LTA (10 ng/ml), LPS (100 ng/ml), GR-1 (1:20). Results are
expressed as mean values ± SEM. Statistical significance within groups denoted with a different
letter as assessed by Kruskal-Wallis statistical test based on ranks followed by a Dunn’s post-hoc
analysis. (B) ROD of mPGES2/β-actin western blot (N=9, p<0.05).
a
b
a,b
b
a,b
a,b
0
1
2
3
4
5
6
control GR-1 LTA GR-1+LTA LPS GR-1+LPS
mP
GES
2/β
-act
in (
rela
tive
to
co
ntr
ol)
B
A
Co
ntr
ol
GR
-1
LTA
GR
-1+
LTA
LPS
GR
-1+
LPS

90
Figure 4.8: Correlation between PGES expression and PGE2 production in human amnion
cells
Correlation plot between (A) cPGES, (B) mPGES1 and (C) mPGES2 expression and PGE2
production in human amnion cell cultures (n=4, R2=0.6321 for cPGES, R
2=0.6249 for mPGES1,
and R2=0.8214 for mPGES2).
R² = 0.6321
0
0.5
1
1.5
2
0 200 400 600 800 1000 1200
cPG
ES/β
-act
ion
OD
(r
ela
tive
to
co
ntr
ol)
PGE2 (pg/ml)
R² = 0.6249
0
0.2
0.4
0.6
0.8
1
1.2
0 200 400 600 800 1000 1200
mP
GES
1/β
-act
ion
OD
(r
ela
tive
to
co
ntr
ol)
PGE2 (pg/ml)
R² = 0.8214
0
0.5
1
1.5
2
2.5
3
3.5
4
0 200 400 600 800 1000 1200
mP
GES
2/β
-act
in O
D
(re
lati
ve t
o c
otn
rol)
PGE2 (pg/ml)
A
B
C

91
Chapter 5
Overall Conclusions and Future Directions

92
5.1 Overall Conclusions
Infection and/or inflammation-mediated preterm birth is characterized by increased pro-
inflammatory actions amongst the gestational tissues. In the amnion specifically this includes
induction of a milieu of pro-inflammatory markers that are increased in the amniotic fluid during
intrauterine infection. In vitro studies have shown that many of these markers are stimulated by
LPS, a gram-negative bacterial endotoxin. This work has described the effects of Lactobacillus
rhamnosus GR-1 on LTA and LPS treated amnion cells and the modulation of cytokines,
chemokines, prostaglandins and prostaglandin metabolizing and synthesizing enzymes. The
effects of Lactobacillus rhamnosus GR-1 can be grouped accordingly as molecules that may
cause: 1) Increases in TNFα and IL-1β antagonists, 2) Recruitment of immune cells, 3)
Modulation of immune cell activity and, 4) Stimulation of PGE2.
5.1.1 Potential Increases in TNFα and IL-1β Antagonists
LPS induces TNFα from the amnion and IL-1β from intact fetal membranes. Both TNFα and IL-
1β are strong TH1 pro-inflammatory cytokines that alongside LPS and other bacterial products,
induce transcription of other cytokines through NκFB control as well as increased prostaglandin
production through modification of prostaglandin metabolizing enzymes. We have shown that
IL-1β is not detectable from our culture system as it was not detected above lowest acceptable
limit of the assay. However, similar to previous results shown in placental trophoblast cells and
murine macrophages, Lactobacillus rhamnosus GR-1 minimizes LTA and LPS stimulated TNFα
release from our human amnion culture system. Release of TNFα may also be decreased by the
effects that other cytokines stimulated by Lactobacillus rhamnosus GR-1 cause. Our studies
demonstrated that IL-6, IFNγ, and GM-CSF release from the amnion is increased by
Lactobacillus rhamnosus GR-1. Each of these cytokines has been shown across various systems
to increase the production of the soluble TNFα receptor and IL-1 receptor antagonist, the
antagonists to TNFα and IL-1β respectively (Aderka et al., 1989; Schindler et al., 1990; Tilg et
al., 1994; Xing et al., 1998). Essentially, IL-6, IFNγ and GM-CSF together provide the potential
to minimize TNFα and IL-1β stimulation of prostaglandins in the amnion. Therefore, by up-
regulating IL-6, IFNγ and GM-CSF, Lactobacillus rhamnosus GR-1 in addition to the direct
effects observed by supernatant alone prevents the actions of pro-inflammatory cytokines

93
responsible for initiation of downstream pathways leading to infection and/or inflammation-
mediated preterm birth.
Lactobacillus rhamnosus GR-1 increased IFNγ release from amnion cells compared to control
and endotoxin stimulation. In placental trophoblast cells, IFNγ reduced PGHS-2 expression and
prostaglandin production (Hanna et al., 2004). At term, this protective mechanism is lost as
IFNγ receptor expression decreases. The result is a net increase in prostaglandin production in
placental trophoblasts at term (Hanna et al., 2004). It is unknown if similar effects on expression
of amnion PGHS-2 are caused by IFNγ. However, an increase in IFNγ caused by Lactobacillus
rhamnosus GR-1 release form the amnion may act in a paracrine manner to help maintain low
expression levels of PGHS-2 in placental trophoblasts to help minimize prostaglandin
production. Consequentially, the cytokines elevated by Lactobacillus rhamnosus GR-1 may be
able to interact with the other gestational tissues to promote additional beneficial effects beyond
the amnion.
5.1.2 Recruitment of Immune Cells
The chemokines MCP-1, RANTES, and IL-8 are involved in chemotaxis of both neutrophils and
macrophages. Lactobacillus rhamnosus GR-1 increases production of these chemokines
compared to control and endotoxin treatments. Consequentially, increased chemotaxis may occur
in the amnion with the influence of Lactobacillus rhamnosus GR-1. Histological analysis of the
amnion shows the presence of immune cells of Hofbauer cells of placental origin in the compact
layer and as residential cells amongst the mesenchymal cells (Bourne, 1962). Therefore,
infiltration and activation of immune cells can occur in the amnion.
5.1.3 Modulation of Immune Cell Activity
Macrophages act as a primary defense against invading pathogenic threats, including bacteria.
The microenvironment to which immune cells are recruited dictate their phenotype and activity.
Bacterial toxins, cytokines, and chemokines all influence the activity of immune cells. The M1
macrophage phenotype is induced by TNFα, IFNγ, GM-CSF and LPS resulting in macrophages
that exhibit strong pro-inflammatory properties and encourage potent bacterial death (Mehta et
al., 2004; Sierra-Filardi et al., 2011). In contrast, the M2 macrophage phenotype is stimulated by

94
IL-4 and IL-13 resulting in macrophages that encourage attenuated pro-inflammatory responses
and elevated anti-inflammatory responses (Mehta et al., 2004). Recently, macrophage activation
has been described as more fluid than simply having two polar phenotypes. Studies have shown
that dependent on the cytokines, endotoxins and other stimulants present in the
microenvironment, macrophages can display characteristics along a wide spectrum between the
M1 and M2 phenotypes (Mosser et al., 2008).
Lactobacillus rhamnosus GR-1 increased IFNγ release from the amnion and Lactobacillus
rhamnosus GR-1 prior to LTA or LPS treatment increases GM-CSF compared to control
treatments. Therefore, Lactobacillus rhamnosus GR-1 increased two of the cytokines involved in
activation of monocytes/macrophages to a highly active pro-inflammatory state capable of
defending against intercellular pathogens. However, studies have shown that optimal activation
of an M1 phenotype is dependent on the activities of TNFα and IFNγ. Monocytes/macrophages
influenced by IFNγ without the presence of TNFα show a phenotype capable of phagocytosis
and production of anti-microbial molecules with a less severe pro-inflammatory phenotype
compared to those activated by both IFNγ and TNFα (Mosser et al., 2008). Lactobacillus
rhamnosus GR-1 attenuated LTA and LPS-stimulated TNFα and increased IFNγ release from the
amnion. Therefore, by the effect of Lactobacillus rhamnosus GR-1 in the amnion, it is expected
that immune cells are recruited and activated to a phenotype capable of bacteria and bacterial
product defense accompanied by a less severe pro-inflammatory responses.
Other studies have demonstrated that prostaglandins and other lipid mediators also influence the
activity of recruited immune cells. Lactobacillus rhamnosus GR-1 increased the release of PGE2
from amnion cells. Amongst all prostaglandins, PGE2 shows the greatest effect on modulation of
macrophage activity (Chizzolini et al., 2009). The role of PGE2 on macrophages is described as
immunosuppressive. In particular, PGE2 modifies the activity of macrophages towards a M2
phenotype as strong pro-inflammatory responses are minimized as anti-inflammatory responses
dominate (Harris et al., 2002). This shift in phenotype is characterized by prevention of LPS
stimulated release of type 1 immune response cytokines such as TNFα and IL-12 while elevating
the release of type two response cytokines such as IL-4 and IL-10 (Betz et al., 1991, Katamura et
al., 1995, Scales et al., 1989, Shinomiya et al., 2001, van der Pouw Kraan et al., 1995). Other

95
studies demonstrate an immunomodulatory role for Lactobacillus rhamnosus GR-1 in mouse
macrophages. In particular, LPS-treated mouse macrophages pretreated with supernatant show
increased levels of G-CSF production, an anti-inflammatory cytokine (Kim et al., 2006). G-CSF
is also increased by Lactobacillus rhamnosus GR-1 supernatant in a fetal sex-dependent manner
in LPS-stimulated placental trophoblast cells (Yeganegi et al., 2011). Therefore, Lactobacillus
rhamnosus GR-1 increases many molecules involved in the activation of immune cells that
appear to promote defense against bacteria and their products while minimizing pro-
inflammatory responses.
Therefore, Lactobacillus rhamnosus GR-1 increased chemokines that may recruit immune cells
while also increasing chemokines, cytokines and prostaglandins that influence the activity of
activated macrophages. The recruitment and activated phenotype of immune cells is difficult to
interpret from the given data and future experiments are necessary to determine this. However,
these results suggest it may be possible that Lactobacillus rhamnosus GR-1 causes immune cells
to be recruited and activated to a state capable of defending against pathogens while the pro-
inflammatory actions that accompany this response are minimized.
5.1.4 Stimulation of Prostaglandin E2
The supernatant of Lactobacillus rhamnosus GR-1 increases cytokines and immune cell activity
that have the potential to minimize LPS and pro-inflammatory prostaglandin stimulation. In
parallel, Lactobacillus rhamnosus GR-1 directly enhances prostaglandin production in amnion
cells. This is similar to the effects of Lactobacillus rhamnosus GG demonstrated to increase
PGHS-2 expression in T84 colon epithelial cells (Korhonen et al., 2004). As well, Lactobacillus
rhamnosus GR-1 increased IL-6 in amnion cells. IL-6 has been shown to stimulate prostaglandin
production in the amnion (Kent et al., 1993; Mitchell et al., 1991). These results suggest that
prostaglandins are stimulated in a more controlled manner, independent of LPS and pro-
inflammatory stimulation in the presence of Lactobacillus rhamnosus GR-1. Amnion produced
prostaglandins may exert stimulatory actions on the myometrium, and cause structural changes
to the fetal membranes and cervix in conjunction with preterm birth when produced in
uncontrolled levels. Lactobacillus rhamnosus GR-1 attenuates LPS stimulation of prostaglandins
perhaps minimizing the harmful effects of deregulation caused in infection and/or inflammation-

96
mediated preterm birth. However, prostaglandins may be produced both directly and indirectly
by actions exerted by Lactobacillus rhamnosus GR-1 in the amnion.
Various physiological stimuli have been shown to increase prostaglandin production in the
amnion including glucocorticoids and IL-10 (Economopoulos et al., 1996; Gibb et al., 1990;
Mitchell et al., 2004; Potestio et al., 1988). Both glucocorticoids and IL-10 exert typically anti-
inflammatory actions in a wide variety of other systems. This indicates that the role of amnion
prostaglandins may be crucial to amnion function. The amnion displays anti-bacterial, anti-
fungal, and anti-viral properties. These include the expression of natural antimicrobials and
innate immune molecules including human α-defensins 1, 2 and 3, human β-defensins 1, 2 and 3
as well as elafin and secretory leukocyte protease inhibitor (King et al., 2007). The innate
expression of such a variety of components of the innate immune system suggests a major role of
the amnion is to prevent uterine infection during pregnancy and labour (King et al., 2007).
Prostaglandins are released from macrophages, NK cells, mast cells, basophils and eosinophils as
part of the immediate innate immune response. Prostaglandins act to perpetuate the
inflammatory response by modulating endothelial cells to allow for entrance of additional
immune cells including leukocytes and are capable of degrading membrane phospholipids of
pathogenic bacteria. Therefore a basal level of prostaglandin production stimulated by various
physiological stimuli may help maintain the important anti-microbial role innate to the amnion.
Consequentially, glucocorticoids, IL-10 and Lactobacillus rhamnosus GR-1 stimulation of
prostaglandins may act to enhance the protective mechanism of the amnion to bacterial and
endotoxin exposure to the fetal environment.
During infection and/or inflammation-mediated preterm birth, bacteria, bacterial products and
pro-inflammatory cytokines, TNFα and IL-1β stimulate amnion prostaglandin production. As the
chorionic PGDH barrier is decreased by similar stimulants, elevated prostaglandin levels lead to
myometrial activation and increased metalloproteinase expression and subsequent cervical
remodeling and membrane weakening. We suggest that Lactobacillus rhamnosus GR-1 prevents
this uncontrolled pathway of prostaglandin production in the amnion and replaces it with a more
controlled probiotic and immunomodulatory cytokine induced pathway. This pathway is possibly
characterized by direct stimulation of prostaglandin production by Lactobacillus rhamnosus GR-

97
1 metabolites or through indirect consequences of Lactobacillus rhamnosus GR-1 induced IL-6
production (Figure 5.1, page 101). The abundance of stimulants of prostaglandin production in
the amnion including the paradoxical actions of IL-10, glucocorticoids and Lactobacillus
rhamnosus GR-1, suggest that prostaglandins play an essential role in the innate immune
functions of the amnion.
5.1.5 Additional Effects of Prostaglandins on the Amnion
The amnion is the closest layer of tissue to the fetal environment and as such is an important
barrier for entrance into the fetal environment. Part of the barrier function the amnion plays is
dictated by the permeability of its epithelial layer dictated by the sealing of the intercellular
space by part of the intercellular complex, tight junctions. In states of infection, bacteria,
bacterial products, cytokines and prostaglandins have been shown to alter tight junction integrity,
structure, and function in various different epithelial layers (Bruewer et al., 2003; Lecuit et al.,
2000; McNamara et al., 2001; Nusrat et al., 2001; Prasad et al., 2005; Talavera et al., 2004;
Tanaka et al., 2008). Disruption to the epithelial barrier is a common step in the inflammatory
response and causes initiation of a mucosal immune response that can then stimulate a stronger
pro-inflammatory immune response that may harm instead of protect the host.
Commensual strains of bacteria in the gastrointestinal tract, including lactobacilli can prevent
changes in tight junction function and structure induced by pathogenic bacteria. Lactobacillus
rhamnosus GG, a strain that adheres to the intestinal cell wall, prevents alterations to tight
junction structure and function caused by Escherichia coli O157:H7 in MDCK-I cell line
(Johnson-Henry et al., 2008). Lactobacillus rhamnosus GG is capable of preventing alterations
caused by Escherichia coli and thus may prevent increased permeability in the gut thereby
preventing entrance of the bacteria into the interstitial space. Lactobacillus plantarum can
prevent similar changes caused by Escherichia coli 0124:NM to both tight junction structure and
function in intestinal epithelial cells (Qin et al., 2009). Recently tight junctions have been
identified in both murine and human amnion (Kobayashi et al., 2009; Kobayashi et al., 2010a).
Inflammatory markers such as TNFα, LPS, and PGE2 have been shown to disrupt the amniotic
barrier through induction of apoptosis and alterations to tight junction structure (Kobayashi et al.,
2010b). Therefore, Lactobacillus rhamnosus GR-1 may also exert protective effects on human

98
amnion tight junction structure and function to help maintain their innate role in a state of
infection and/or inflammation-mediated preterm birth.
Lactobacillus rhamnosus GR-1 administered both orally and through vaginal instillation has
been shown to colonize and replenish the vaginal microbiota. As well, when injected directly
into the bladder of patients, this strain of lactobacilli does not result in any pathogenic responses.
Therefore, the use of therapeutic Lactobacillus rhamnosus GR-1 will most likely be safe for use
in pregnant women. In addition, in vitro studies have shown that both primary amnion and
placental trophoblast cultures, Lactobacillus rhamnosus GR-1 attenuated release of the pro-
inflammatory cytokine, TNFα demonstrating a beneficial immunomodulatory role when the
tissues are challenged with either LTA or LPS. This prevention of pro-inflammatory cytokine
release may act in ameliorating the activation of downstream pathways associated with infection
and/or inflammation-mediated preterm delivery. Additionally, in the amnion, Lactobacillus
rhamnosus GR-1 increases the defensive capabilities of the fetal membranes by recruiting and
activating immune cells capable of defending against pathogenic threats at the fetal environment
interface. Therefore, given the well-established safety of lactobacilli, the ability to colonize the
vagina and the beneficial immunomodulatory effects on the gestational tissues, the results of this
thesis have further demonstrated the potential for probiotic use in pregnancy to prevent infection
and/or inflammation-mediated preterm birth.
5.1.6 Lactobacillus rhamnosus GR-1 Supernatant Metabolite(s)
Although the exact signaling mechanism and molecule or molecules responsible for the effects
caused by the supernatant collected from Lactobacillus rhamnosus GR-1 is unknown, studies on
other probiotics have been performed. Previously, probiotic supernatant was shown to exert pro-
inflammatory actions through polysaccharide molecules and anti-inflammatory molecules
extracted from collected media (Chon et al., 2010). As well, probiotics have been shown to
signal through the MAPK and JAK/STAT pathways as JNK and JAK/STAT molecules are
phosphorylated through unknown metabolites present in the supernatant of various probiotics
(Kim et al., 2008; Yeganegi et al., 2010). Interestingly, the current study demonstrates a
threshold dilution of Lactobacillus rhamnosus GR-1 to increase IL-6 and IL-8 concentrations in
human amnion cells of 1:20 compared to 1:50 and 1:100 where no stimulation was shown.

99
Therefore, the metabolite or metabolites present in the Lactobacillus rhamnosus GR-1
supernatant may only exert its effects if it is present above a certain threshold at optimal
concentrations. As suggested by previously performed experiments, the unknown metabolite
may interact with TLR2 or TLR4, or unknown receptors. This may occur either through
antagonistic activity to the ligands of a given receptor by either blocking the ligands interaction
with the receptor of by leading to downstream signaling that may interfere with those activated
by pathogenic ligands such as LTA or LPS or by activating downstream signaling independent of
TLR2 and TLR4 that leads to an anti-inflammatory response. In addition, the current study
demonstrated that a 1:10 dilution of Lactobacillus rhamnosus GR-1 supernatant does not exert
the same effects on IL-6 and IL-8 as 1:20. This may suggest that homodimerization may occur
between the metabolite or competition between various metabolites may occur to block the
ability to activate associated receptors. Therefore, the effects exerted by probiotics are dependent
on both the strain of bacteria and the tissue and cell type of the host as well as the concentration
of metabolites produced by the bacteria.
5.1.7 Lactobacillus rhamnosus GR-1 and Prevention of Preterm Birth Associated with Infection and/or Inflammation
When considering the use of Lactobacillus rhamnosus GR-1 as a preventative treatment, the
effect of its supernatant on the individual gestational tissue must be considered. In placental
trophoblasts, Lactobacillus rhamnosus GR-1 exerts anti-inflammatory effects characterized by
increased IL-10 and G-CSF and decreased LPS-stimulated TNFα release (Yeganegi et al., 2009;
Yeganegi et al., 2010; Yeganegi et al., 2011). This prevention of LPS-stimulated TNFα release
is paralleled in the amnion; however it is accompanied by a strong chemotactic response capable
of modulating immune cell activity and increasing prostaglandin production. Although further
studies are required to determine if the in vivo effects Lactobacillus rhamnosus GR-1 on immune
cell recruitment and activation to the amnion are beneficial or detrimental to the prolongation of
gestation, the effect of this probiotic on up-regulation of PGE2 must be taken into consideration
when determining the route of administration for preventative treatment for infection and/or
inflammation mediated preterm birth. As preventative therapy, the most user-friendly method
would be oral administration possibly as part of a daily supplement for pregnancy women.
Orally administrated probiotics have been shown capable of reaching the vaginal flora and

100
despite the sterility of the amnion in healthy pregnancies, bacteria or their metabolites may be
capable of reaching the fetal membranes in vivo. Therefore, the dose and the potential
interactions with the inner layer of the fetal membranes, the amnion must be taken into
consideration in a more representative model than in vitro cell cultures to determine the best
method of preventative treatment for pregnancy women from undergoing infection and/or
inflammation mediated preterm birth.
5.1.8 Limitations
Several limitations are of importance in regards to the experiments and conclusions presented in
this study. Many experiments performed on the amnion are done on either intact membranes
together with the chorion or on dispersed cells separated into epithelial and mesenchymal cell
types. As the response of the amnion varies dependent on the presence of absence of not only the
other tissue of the membranes, the chorion as well as the partner cell type, epithelial or
mesenchymal cells the results of this study are limited in their in vivo representation. In addition,
cultured cells were not mixed in proportion to in vivo amnion tissue thus over or under
representation of either cell type may be possible.
When intact membranes are tested in a transwell system, shuttling of cytokine mRNA is thought
to occur between the amnion and chorion. Due to the lack of the chorion in the system used in
this study, both IL1β and IL-10 were not detected from mixed amnion cell cultures. IL-1β is a
strong pro-inflammatory cytokine that is thought to play a role in initiation of infection and/or
inflammation mediated preterm birth whereas IL-10 is an anti-inflammatory cytokine that
counteracts many pro-inflammatory actions and is compatible with the TH2 pregnancy cytokine
profile. Therefore, the effects of Lactobacillus rhamnosus GR-1 on IL-1β and IL-10 from intact
membranes cannot be determined using the system of mixed amnion cell cultures used in this
study.

101
Am
nio
tic
Flu
id
Am
nio
n
Lact
ob
aci
llus
rha
mn
osu
s G
R-1
met
abo
lite
(s)
+ LT
A o
r LP
S
Asc
en
din
g b
acte
rial
en
do
toxi
ns
LTA
or
LPS
TNFα
LTA
LPS
Me
mb
ran
e w
eak
enin
g
MM
P-2
, -9
PG
E 2
TNFα
IL-6
IFNγ
GM
-CSF
IL-8
MC
P-1
RA
NT
ES
Rec
ruit
men
t A
ctiv
atio
n
LTA
LPS
LPS
LTA
Me
mb
ran
e w
eak
enin
g
MM
P-2
, -9
PG
E 2
A
B
C
Fig
ure
5.1
: T
he
effe
cts
of
La
ctob
aci
llu
s rh
am
no
sus
GR
-1 o
n h
um
an
am
nio
n c
ells
. (A
) U
ntr
eate
d s
tate
of
infe
ctio
n a
nd/o
r in
flam
mat
ion
. T
NF
α l
evel
s in
crea
se P
GE
2
pro
du
ctio
n l
ead
ing t
o c
oll
agen
deg
rad
atio
n,
mem
bra
ne
wea
ken
ing a
nd
en
tran
ce o
f b
acte
rial
en
do
toxin
to t
he
amn
ioti
c fl
uid
and
th
e on
set
of
pre
term
lab
ou
r. (
B)
Eff
ect
of
La
ctob
aci
llu
s rh
am
no
sus
GR
-1 t
reat
men
t o
n i
mm
un
e ce
lls.
Tre
atm
ent
pri
or
to L
TA
or
LP
S s
tim
ula
tion
res
ult
s in
att
enu
atio
n o
f T
NF
α a
nd
in
crea
se c
hem
ota
xis
and
acti
vat
ion
of
imm
un
e ce
lls
to a
ph
eno
typ
e ca
pab
le o
f d
efen
din
g a
gai
nst
bac
teri
a an
d b
acte
rial
pro
du
cts
wh
ile
min
imiz
ing p
ro-i
nfl
amm
ato
ry r
esp
on
ses.
(C
) E
ffec
t o
f
La
ctob
aci
llu
s rh
am
no
sus
GR
-1 o
n p
rost
agla
nd
ins.
In
crea
sed
PG
E2 p
rod
uct
ion
by L
act
ob
aci
llu
s rh
am
no
sus
GR
-1 m
ay a
ct a
s a
pro
tect
ive
def
ense
agai
nst
pat
hogen
s
reac
hin
g t
he
feta
l en
vir
on
men
t b
y t
erm
inat
ing p
regn
ancy
if
bac
teri
al t
hre
ats
bec
om
e d
etri
men
tal
to t
he
hea
lth
of
the
fetu
s.
Ep
ith
elia
l ce
lls
Mes
ench
ym
al c
ells
Imm
un
e ce
lls

102
5.2 Future Directions
Currently, there is no effective treatment to prevent preterm delivery that occurs in
approximately 10% of all pregnancies. Consequentially, preterm neonates account for 80% of all
neonate morbidities and mortalities. The ultimate goal of the research presented in this thesis was
to aid in developing an effective preventative treatment for infection and/or inflammation
mediated preterm birth. The effects of Lactobacillus rhamnosus GR-1 on cytokine, chemokine
and prostaglandin production in the amnion described in this thesis hopefully bring researchers
one step closer to developing a treatment to prevent preterm birth and its associated
consequences.
The future directions of the current study include further investigation into the effects of
Lactobacillus rhamnosus GR-1 on products of the human amnion. Arachidonic acid is
metabolized into several different end products including leukotrienes, lipoxins and eicosanoids.
Both leukotrienes and lipoxins influence inflammation and immune cell responses. Another
molecular target that may be of interest is the metalloproteinases and TIMPs that are
differentially expressed under LPS stimulation. These enzymes are responsible for degradation
or prevention of degradation of collagen during membrane weakening and cervical remodeling.
They are up-regulated by prostaglandins and are part of the downstream pathways associated
with infection and/or inflammation-mediated preterm labour. Therefore, the effect of
Lactobacillus rhamnosus GR-1 on leukotrienes, lipoxins, MMPs and TIMPs may further
illustrate the effects of probiotic treatment on the immune response from the amnion.
The cytokine and chemokine profile produced in response to Lactobacillus rhamnosus GR-1
described in this thesis suggests an effect on the recruitment and activation of immune cells.
However, many of the cytokines and chemokines mentioned in this study have well-defined roles
amongst the other gestational tissues that have not yet been established in the amnion.
Therefore, before the conclusion of immune cell recruitment and activation can be confirmed,
determining the precise effects of cytokines and chemokines on the amnion is crucial. Of
particular interest are the effects of IFNγ on PGHS-2 expression and prostaglandin production as
well as GM-CSF and IL-6 on production of sTNFα receptor and IL-1ra in the amnion. In

103
addition, it will be important to determine if immune cell recruitment can occur in vivo and if this
reaction provides additional defenses or is deleterious to the prolongation of gestation.
In order to better understand the mechanism of action of the metabolites of Lactobacillus
rhamnosus GR-1, studies identifying the downstream signaling associated with probiotic
treatment should be determined. Previously, Lactobacillus rhamnosus GR-1 was shown to
modify the phosphorylation state of enzymes involved in MAP kinase and NFκB signaling
pathways. Probiotic enhancement of G-CSF in mouse macrophages is caused by a JNK-
dependent mechanism (Kim et al., 2006). Determining the effects of Lactobacillus rhamnosus
GR-1 on common pathways known to modulate cytokine transcription and translation will
provide insight into the signaling pathway.
By determining the effects of Lactobacillus rhamnosus GR-1 on the type of molecules produced
in the amnion that are associated with infection and/or inflammation-mediated preterm birth, a
better understanding of the effects of probiotic treatment on this tissue should be determined.
Since Lactobacillus rhamnosus GR-1 may cause both recruitment and activation of immune cells
and increased prostaglandin production from the amnion the use of its supernatant on pregnancy
may be two-fold. Firstly, by administering lactobacilli in a route that minimizes contact and
interaction with the amnion, pro-inflammatory responses in placental trophoblasts and amnion
may be attenuated and therefore prevent infection and/or inflammation preterm labour.
Secondly, it may be possible that administration of Lactobacillus rhamnosus GR-1 in a route that
allows for direct interaction of the supernatant with the amnion may be used to promote
protection from bacterial endotoxin entrance to the fetal environment for use in idiopathic or
induced labour where prolongation of gestation is deemed deleterious to the health of the fetus or
mother. However, these options may be dependent on the dose effects of Lactobacillus
rhamnosus GR-1 on the amnion which were not assessed in this study. Therefore, various routes
of supernatant administration including oral, vaginal or directly into the cervix may result in
different interaction of supernatant with the amnion and consequentially different effects on
infection and/or inflammation-mediated preterm birth.

104
The goal of the research presented in this thesis was to aid in the development of an effective
preventative treatment for infection and/or inflammation-mediated preterm birth. Hopefully, the
presented work has not only answered apparent questions but also opened routes and areas of
interest for future research. By answering these questions, an effective treatment will help
prevent the devastating consequences of preterm birth and lead to greater overall health and
success of high-risk pregnancies.

105
References
n.a. (2006). Preterm Birth: Causes, Consequences, and Prevention. Institute of Medicine of the
National Academies. Available at: http://www.iom.edu/~/media/Files/Report%20Files/2006/Pre
term-Birth-Causes-Consequences-and prevention/Preterm%20Birth%202006%20Report%20Bri
ef.pdf
n.a. Public Health Agency of Canada. (2008). Canadian Perinatal Health Report - 2008 Edition.
Available at: http://www.phac-aspc.gc.ca/publicat/2008/cphr-rspc/pdf/cphr-rspc08-eng.pdf
n.a. Food and Agriculture Organization of the United Nations (FAO) and World Health
Organization (WHO). (2001). Health and nutritional properties of probiotics in food including
powder milk with live lactic acid bacteria. FAO and WHO Joint Expert Committee Report,
2001. Available at: http://www.who.int/foodsafety/publicatons/fs_management/en/probiotics.pdf
Ackerman, W. E.,4th, Summerfield, T. L., Vandre, D. D., Robinson, J. M., & Kniss, D. A.
(2008). Nuclear factor-kappa B regulates inducible prostaglandin E synthase expression in
human amnion mesenchymal cells. Biology of Reproduction. 78, 68-76.
Aderka, D., Le, J. M., & Vilcek, J. (1989). IL-6 inhibits lipopolysaccharide-induced tumor
necrosis factor production in cultured human monocytes, U937 cells, and in mice. Journal of
Immunology (Baltimore, Md.: 1950). 143, 3517-3523.
Ahn, C., & Stiles, M. E. (1990). Antibacterial activity of lactic acid bacteria isolated from
vacuum-packaged meats. Journal of Applied Bacteriology. 69, 302-310.

106
Alfaidy, N., Xiong, Z. G., Myatt, L., Lye, S. J., MacDonald, J. F., & Challis, J. R. (2001).
Prostaglandin F2alpha potentiates cortisol production by stimulating 11beta-hydroxysteroid
dehydrogenase 1: A novel feedback loop that may contribute to human labor. Journal of Clinical
Endocrinology and Metabolism. 86, 5585-5592.
Alvi, S.A., Brown, N.L., Bennett, P.R., Elder, M.G., Sullivan, M.H. (1999). Corticotrophin-
releasing hormone and platelet-activating factor induce transcription of the type-2 cyclo-
oxygenase gene in human fetal membranes. Molecular Human Reproduction. 5, 476-480.
Andrews, W.W., Hauth J.C., Goldenberg, R.L. (2000). Infection and preterm birth. American
Journal of Perinatology. 17(7), 357-365.
Andrews, W. W., Hauth, J. C., Goldenberg, R. L., Gomez, R., Romero, R., & Cassell, G. H.
(1995). Amniotic fluid interleukin-6: Correlation with upper genital tract microbial colonization
and gestational age in women delivered after spontaneous labor versus indicated delivery.
American Journal of Obstetrics and Gynecology. 173, 606-612.
Andrews, W. W., Sibai, B. M., Thom, E. A., Dudley, D., Ernest, J. M., McNellis, D., Leveno,
K. J., Wapner, R., Moawad, A., O'Sullivan, M. J., Caritis, S. N., Iams, J. D., Langer, O.,
Miodovnik, M., Dombrowski, M., & National Institute of Child Health & Human
Development Maternal-Fetal Medicine Units Network. (2003). Randomized clinical trial of
metronidazole plus erythromycin to prevent spontaneous preterm delivery in fetal fibronectin-
positive women. Obstetrics and Gynecology. 101, 847-855.

107
Antonio, M. A., Hawes, S. E., & Hillier, S. L. (1999). The identification of vaginal
lactobacillus species and the demographic and microbiologic characteristics of women colonized
by these species. Journal of Infectious Diseases. 180, 1950-1956.
Aplin, J. D., Campbell, S., & Allen, T. D. (1985). The extracellular matrix of human amniotic
epithelium: Ultrastructure, composition and deposition. Journal of Cell Science. 79, 119-136.
Argiles, J. M., Carbo, N., & Lopez-Soriano, F. J. (1997). TNF and pregnancy: The paradigm
of a complex interaction. Cytokine & Growth Factor Reviews. 8, 181-188.
Arici, A., MacDonald, P. C., & Casey, M. L. (1995). Regulation of monocyte chemotactic
protein-1 gene expression in human endometrial cells in cultures. Molecular and Cellular
Endocrinology. 107, 189-197.
Arici, A., Oral, E., Bukulmez, O., Buradagunta, S., Bahtiyar, O., & Jones, E. E. (1997).
Monocyte chemotactic protein-1 expression in human preovulatory follicles and ovarian cells.
Journal of Reproductive Immunology. 32, 201-219.
Athayde, N., Romero, R., Maymon, E., Gomez, R., Pacora, P., Araneda, H., & Yoon, B.H.
(1999). A Role for the novel cytokine RANTES in pregnancy and parturition. American Journal
of Obstetrics and Gynecology. 181, 989-994.
Benirschke, K. (1960). Routes and types of infection in the fetus and the newborn. A.M.A.
Journal of Diseases of Children. 99, 714-721.

108
Bennett, P. R., Rose, M. P., Myatt, L., & Elder, M. G. (1987). Preterm labor: Stimulation of
arachidonic acid metabolism in human amnion cells by bacterial products. American Journal of
Obstetrics and Gynecology. 156, 649-655.
Betz, M., Fox, B.S. (1991). Prostaglandin E2 inhibits production of Th1 lymphokines but not of
Th2 lymphokines. Journal of Immunology. 146(1), 108-113.
Bloise, E., Torricelli, M., Novembri, R., Borges, L. E., Carrarelli, P., Reis, F. M., Petraglia,
F. (2010). Heat-killed Lactobacillus rhamnosus GG Modulates Urocortin and Cytokine Release
in Primary Trophoblast Cells. Placenta. 31, 867-872.
Blumenstein, M., Hansen, W. R., Deval, D., & Mitchell, M. D. (2000). Differential regulation
in human amnion epithelial and fibroblast cells of prostaglandin E(2) production and
prostaglandin H synthase-2 mRNA expression by dexamethasone but not tumour necrosis factor-
alpha. Placenta. 21, 210-217.
Bocking, A. D., Challis, J. R., & Korebrits, C. (1999). New approaches to the diagnosis of
preterm labor. American Journal of Obstetrics and Gynecology. 180, S247-8.
Bourne, G. (1962). The foetal membranes. A review of the anatomy of normal amnion and
chorion and some aspects of their function. Postgraduate Medical Journal. 38, 193-201.
Brown, N. L., Alvi, S. A., Elder, M. G., Bennett, P. R., & Sullivan, M. H. (1998). Interleukin-
1beta and bacterial endotoxin change the metabolism of prostaglandins E2 and F2 alpha in intact
term fetal membranes. Placenta. 19, 625-630.

109
Bry, K., & Hallman, M. (1992). Transforming growth factor-beta opposes the stimulatory
effects of interleukin-1 and tumor necrosis factor on amnion cell prostaglandin E2 production:
Implication for preterm labor. American Journal of Obstetrics and Gynecology. 167, 222-226.
Bry, K., Hallman, M., & Lappalainen, U. (1995). Production of granulocyte-macrophage
colony-stimulating factor (GM-CSF) by amnion cells and its regulation by proinflammatory
cytokines. Journal of the Society for Gynecologic Investigation. 2, 167-167.
Bry, K., Hallman, M., Teramo, K., Waffarn, F., & Lappalainen, U. (1997). Granulocyte-
macrophage colony-stimulating factor in amniotic fluid and in airway specimens of newborn
infants. Pediatric Research. 41, 105-109.
Bryant-Greenwood, G. D. (1998). The extracellular matrix of the human fetal membranes:
Structure and function. Placenta. 19, 1-11.
Burton, J.P., Reid, G. (2002). Evaluation of the bacterial vaginal flora of 20 postmenopausal
women by direct (Nugent score) and molecular (polymerase chain reaction and denaturing
gradient gel electrophoresis) techniques. Journal of infectious diseases. 186, 1770-1780.
Cadieux, P., Burton, J., Gardiner, G., Braunstein, I., Bruce, A. W., Kang, C. Y., & Reid, G.
(2002). Lactobacillus strains and vaginal ecology. Journal of the American Medical Association.
287, 1940-1941.
Calvin, S. E., & Oyen, M. L. (2007). Microstructure and mechanics of the chorioamnion
membrane with an emphasis on fracture properties. Annals of the New York Academy of
Sciences. 1101, 166-185.

110
Carey, J. C., Klebanoff, M. A., Hauth, J. C., Hillier, S. L., Thom, E. A., Ernest, J. M.,
Heine, R. P., Nugent, R. P., Fischer, M. L., Leveno, K. J., Wapner, R., & Varner, M. (2000).
Metronidazole to prevent preterm delivery in pregnant women with asymptomatic bacterial
vaginosis. national institute of child health and human development network of maternal-fetal
medicine units. New England Journal of Medicine. 342, 534-540.
Challis, J. R., Bloomfield, F. H., Bocking, A. D., Casciani, V., Chisaka, H., Connor, K.,
Dong, X., Gluckman, P., Harding, J. E., Johnstone, J., Li, W., Lye, S., Okamura, K., &
Premyslova, M. (2005). Fetal signals and parturition. Journal of Obstetrics and Gynaecology
Research. 31, 492-499.
Challis, J. R., Lye, S. J., & Gibb, W. (1997). Prostaglandins and parturition. Annals of the New
York Academy of Sciences. 828, 254-267.
Challis, J. R., Sloboda, D., Matthews, S. G., Holloway, A., Alfaidy, N., Patel, F. A., Whittle,
W., Fraser, M., Moss, T. J., & Newnham, J. (2001). The fetal placental hypothalamic-
pituitary-adrenal (HPA) axis, parturition and post natal health. Molecular and Cellular
Endocrinology. 185, 135-144.
Challis, J. R., Sloboda, D. M., Alfaidy, N., Lye, S. J., Gibb, W., Patel, F. A., Whittle, W. L.,
& Newnham, J. P. (2002). Prostaglandins and mechanisms of preterm birth. Reproduction
(Cambridge, England). 124, 1-17.
Challis, J. R. G., Matthews, S. G., Gibb, W., & Lye, S. J. (2000). Endocrine and paracrine
regulation of birth at term and preterm. Endocrine Reviews. 21, 514-550.

111
Cherouny, P. H., Pankuch, G. A., Romero, R., Botti, J. J., Kuhn, D. C., Demers, L. M., &
Appelbaum, P. C. (1993). Neutrophil attractant/activating peptide-1/interleukin-8: Association
with histologic chorioamnionitis, preterm delivery, and bioactive amniotic fluid leukoattractants.
American Journal of Obstetrics and Gynecology. 169, 1299-1303.
Cheung, P. Y., Walton, J. C., Tai, H. H., Riley, S. C., & Challis, J. R. (1990).
Immunocytochemical distribution and localization of 15-hydroxyprostaglandin dehydrogenase in
human fetal membranes, decidua, and placenta. American Journal of Obstetrics and Gynecology.
163, 1445-1449.
Chizzolini, C., Brembilla, N.C. (2009). Prostaglandin E2: igniting the fire. Immunology and
cell biology. 87(7), 510-511.
Chon, H., Choi, B., Jeong, G., Lee, E., & Lee, S. (2010). Suppression of proinflammatory
cytokine production by specific metabolites of lactobacillus plantarum 10hk2 via inhibiting NF-
kappaB and p38 MAPK expressions. Comparative Immunology, Microbiology and Infectious
Diseases. 33, e41-9.
Cong B., Zhang L., Gao L., Ni X. (2009). Reduced expression of CRH receptor type 1 in upper
segment human myometrium during labour. Reproductive Biology Endocrinology. 7, 43-51.
Coultrip, L. L., Lien, J. M., Gomez, R., Kapernick, P., Khoury, A., & Grossman, J. H.
(1994). The value of amniotic fluid interleukin-6 determination in patients with preterm labor
and intact membranes in the detection of microbial invasion of the amniotic cavity. American
Journal of Obstetrics and Gynecology. 171, 901-911.

112
Curbelo, V., Bejar, R., Benirschke, K., & Gluck, L. (1981). Premature labor. I. prostaglandin
precursors in human placental membranes. Obstetrics and Gynecology. 57, 473-478.
Deb, K., Chaturvedi, M. M., & Jaiswal, Y. K. (2004). Comprehending the role of LPS in
gram-negative bacterial vaginosis: Ogling into the causes of unfulfilled child-wish. Archives of
Gynecology and Obstetrics. 270, 133-146.
Denison, F. C., Kelly, R. W., Calder, A. A., & Riley, S. C. (1998). Cytokine secretion by
human fetal membranes, decidua and placenta at term. Human Reproduction (Oxford, England).
13, 3560-3565.
Denney, J. M., Culhane, J. F., & Goldenberg, R. L. (2008). Prevention of preterm birth.
Women's Health (London, England). 4, 625-638.
Dudley, D. J., Edwin, S. S., & Mitchell, M. D. (1996). Macrophage inflammatory protein-I
alpha regulates prostaglandin E2 and interleukin-6 production by human gestational tissues in
vitro. Journal of the Society for Gynecologic Investigation. 3, 12-16.
Dugoua, J. J., Machado, M., Zhu, X., Chen, X., Koren, G., & Einarson, T. R. (2009).
Probiotic safety in pregnancy: A systematic review and meta-analysis of randomized controlled
trials of Lactobacillus, Bifidobacterium, and Saccharomyces spp. Journal of Obstetrics and
Gynaecology Canada. 31, 542-552.
Dvorak, A. M., Morgan, E. S., Tzizik, D. M., & Weller, P. F. (1994). Prostaglandin
endoperoxide synthase (cyclooxygenase): Ultrastructural localization to nonmembrane-bound

113
cytoplasmic lipid bodies in human eosinophils and 3T3 fibroblasts. International Archives of
Allergy and Immunology. 105, 245-250.
Economopoulos, P., Sun, M., Purgina, B., & Gibb, W. (1996). Glucocorticoids stimulate
prostaglandin H synthase type-2 (PGHS-2) in the fibroblast cells in human amnion cultures.
Molecular and Cellular Endocrinology. 117, 141-147.
Embrey, M. (1971). PGE compounds for induction of labour and abortion. Annals of the New
York Academy of Sciences. 180, 518-523.
Eschenbach, D. A., Davick, P. R., Williams, B. L., Klebanoff, S. J., Young-Smith, K.,
Critchlow, C. M., & Holmes, K. K. (1989). Prevalence of hydrogen peroxide-producing
lactobacillus species in normal women and women with bacterial vaginosis. Journal of Clinical
Microbiology. 27, 251-256.
Esplin, M. S., Romero, R., Chaiworapongsa, T., Kim, Y. M., Edwin, S., Gomez, R., Mazor,
M., & Adashi, E. Y. (2005). Monocyte chemotactic protein-1 is increased in the amniotic fluid
of women who deliver preterm in the presence or absence of intra-amniotic infection. The
Journal of Maternal-Fetal & Neonatal Medicine. 17, 365-373.
Falagas, M. E., Betsi, G. I., & Athanasiou, S. (2007). Probiotics for the treatment of women
with bacterial vaginosis. Clinical Microbiology and Infection. 13, 657-664.
Floch, M. H. (2010). The effect of probiotics on host metabolism: The microbiota and
fermentation. Journal of Clinical Gastroenterology. 44 Suppl 1, S19-21.

114
Forsberg, L., Leeb, L., Thoren, S., Morgenstern, R., & Jakobsson, P. (2000). Human
glutathione dependent prostaglandin E synthase: Gene structure and regulation. FEBS Letters.
471, 78-82.
Forsum, U., Holst, E., Larsson, P. G., Vasquez, A., Jakobsson, T., & Mattsby-Baltzer, I.
(2005). Bacterial vaginosis--a microbiological and immunological enigma. APMIS : Acta
Pathologica, Microbiologica, Et Immunologica Scandinavica. 113, 81-90.
Fortunato, S. J., Menon, R. P., Swan, K. F., & Menon, R. (1996). Inflammatory cytokine
(interleukins 1, 6 and 8 and tumor necrosis factor-alpha) release from cultured human fetal
membranes in response to endotoxic lipopolysaccharide mirrors amniotic fluid concentrations.
American Journal of Obstetrics and Gynecology. 174, 1855-61; discussion 1861-2.
Fuchs, A. R., & Fuchs, F. (1981). Ethanol for prevention of preterm birth. Seminars in
Perinatology. 5, 236-251.
Fuentes, A., Spaziani, E. P., & O'Brien, W. F. (1996). The expression of cyclooxygenase-2
(COX-2) in amnion and decidua following spontaneous labor. Prostaglandins. 52, 261-267.
Furuta, I., Yamada, H., Sagawa, T., & Fujimoto, S. (2000). Effects of inflammatory cytokines
on prostaglandin E(2) production from human amnion cells cultured in serum-free condition.
Gynecologic and Obstetric Investigation. 49, 93-97.
Garcia-Lloret, M. I., Morrish, D. W., Wegmann, T. G., Honore, L., Turner, A. R., &
Guilbert, L. J. (1994). Demonstration of functional cytokine-placental interactions: CSF-1 and

115
GM-CSF stimulate human cytotrophoblast differentiation and peptide hormone secretion.
Experimental Cell Research. 214, 46-54.
Gardiner, G. E., Heinemann, C., Bruce, A. W., Beuerman, D., & Reid, G. (2002).
Persistence of Lactobacillus fermentum RC-14 and Lactobacillus rhamnosus GR-1 but not L.
rhamnosus GG in the human vagina as demonstrated by randomly amplified polymorphic DNA.
Clinical and Diagnostic Laboratory Immunology. 9, 92-96.
Gasson, J. C. (1991). Molecular physiology of granulocyte-macrophage colony-stimulating
factor. Blood. 77, 1131-1145.
Gibb, W., & Lavoie, J. C. (1990). Effects of glucocorticoids on prostaglandin formation by
human amnion. Canadian Journal of Physiology and Pharmacology. 68, 671-676.
Gibb, W., & Sun, M. (1996). Localization of prostaglandin H synthase type 2 protein and
mRNA in term human fetal membranes and decidua. Journal of Endocrinology. 150, 497-503.
Giles, W., & Bisits, A. (2007). Preterm labour. the present and future of tocolysis. Best Practice
& Research.Clinical Obstetrics & Gynaecology. 21, 857-868.
Goldenberg, R. L., Culhane, J. F., Iams, J. D., & Romero, R. (2008). Epidemiology and
causes of preterm birth. Lancet. 371, 75-84.
Goldenberg, R. L., Hauth, J. C., & Andrews, W. W. (2000). Intrauterine infection and preterm
delivery. New England Journal of Medicine. 342, 1500-1507.

116
Goldenberg, R. L., Iams, J. D., Mercer, B. M., Meis, P., Moawad, A., Das, A., Copper, R.,
Johnson, F., & National Institute of Child Health and Human Development Maternal-Fetal
Medicine Units Network. (2003). What we have learned about the predictors of preterm birth.
Seminars in Perinatology. 27, 185-193.
Goldenberg, R. L., & Rouse, D. J. (1998). Prevention of premature birth. New England Journal
of Medicine. 339, 313-320.
Goodwin, J.S., Ceuppens, J. (1983). Regulation of the immune response by prostaglandins.
Journal of Clinical Immunology. 3(4), 295-315.
Hagberg, L.A., Bruce, A.W., Reid, G., Svanborg Eden, C., Lincoln, K., Lidin-Janson G.
(1989). Colonization of the urinary tract with live bacteria from the normal fecal and urethral
flora in patients with recurrent symptomatic urinary tract infections. University of Chicago Press.
194-197.
Han, R., Tsui, S., & Smith, T. J. (2002). Up-regulation of prostaglandin E2 synthesis by
interleukin-1beta in human orbital fibroblasts involves coordinate induction of prostaglandin-
endoperoxide H synthase-2 and glutathione-dependent prostaglandin E2 synthase expression.
Journal of Biological Chemistry. 277, 16355-16364.
Hanna, N., Bonifacio, L., Reddy, P., Hanna, I., Weinberger, B., Murphy, S., Laskin, D., &
Sharma, S. (2004). IFN-gamma-mediated inhibition of COX-2 expression in the placenta from
term and preterm labor pregnancies. American Journal of Reproductive Immunology (New
York, N.Y.: 1989). 51, 311-318.

117
Harper, M., Zheng, S.L., Thom, E., Klebanoff, M.A., Thorp, J. Jr., Sorokin, Y., Varner,
M.W., Iams, J.D., Dinsmoor, M., Mercer, B.M., Rouse, D.J., Ramin, S.M., Anderson, G.D.
(2011) Cytokine gene polymorphisms and length of gestation. Obstetrics and Gynecology.
117(1), 125-130.
Harris, S.G., Padilla, J., Koumas, L., Ray, D., Phipps, R.P. (2002). Prostaglandins as
modulators of immunity. Trends in Immunology. 23(3), 144-150.
Hawes, S. E., Hillier, S. L., Benedetti, J., Stevens, C. E., Koutsky, L. A., Wolner-Hanssen,
P., & Holmes, K. K. (1996). Hydrogen peroxide-producing lactobacilli and acquisition of
vaginal infections. Journal of Infectious Diseases. 174, 1058-1063.
Hillhouse, E.W., Grammatopoulos, D.K. (2002). Role of stress peptides during human
pregnancy and labour. Reproduction. 124, 323-329.
Hillier, S. L., Krohn, M. A., Klebanoff, S. J., & Eschenbach, D. A. (1992). The relationship of
hydrogen peroxide-producing lactobacilli to bacterial vaginosis and genital microflora in
pregnant women. Obstetrics and Gynecology. 79, 369-373.
Hillier, S. L., Krohn, M. A., Rabe, L. K., Klebanoff, S. J., & Eschenbach, D. A. (1993). The
normal vaginal flora, H2O2-producing lactobacilli, and bacterial vaginosis in pregnant women.
Clinical Infectious Diseases. 16, S273-81.
Hillier, S. L., Nugent, R. P., Eschenbach, D. A., Krohn, M. A., Gibbs, R. S., Martin, D. H.,
Cotch, M. F., Edelman, R., Pastorek, J. G.,2nd, & Rao, A. V. (1995). Association between

118
bacterial vaginosis and preterm delivery of a low-birth-weight infant. the vaginal infections and
prematurity study group. New England Journal of Medicine. 333, 1737-1742.
Hillier, S. L., Witkin, S. S., Krohn, M. A., Watts, D. H., Kiviat, N. B., & Eschenbach, D. A.
(1993). The relationship of amniotic fluid cytokines and preterm delivery, amniotic fluid
infection, histologic chorioamnionitis, and chorioamnion infection. Obstetrics and Gynecology.
81, 941-948.
Hirst, J. J., Teixeira, F. J., Zakar, T., & Olson, D. M. (1995). Prostaglandin endoperoxide-H
synthase-1 and -2 messenger ribonucleic acid levels in human amnion with spontaneous labor
onset. Journal of Clinical Endocrinology and Metabolism. 80, 517-523.
Holloway, A.C., Gyomorey S., Challis J.R. (2000). Effects of labor on pituitary expression of
proopiomelanocortin, prohormone convertase (PC)-1, PC-2, and glucocorticoid receptor mRNA
in fetal sheep. Endocrine. 13, 17-23.
Hornung, D., Ryan, I. P., Chao, V. A., Vigne, J. L., Schriock, E. D., & Taylor, R. N. (1997).
Immunolocalization and regulation of the chemokine RANTES in human endometrial and
endometriosis tissues and cells. Journal of Clinical Endocrinology and Metabolism. 82, 1621-
1628.
Howard, J. C., Heinemann, C., Thatcher, B. J., Martin, B., Gan, B. S., & Reid, G. (2000).
Identification of collagen-binding proteins in lactobacillus spp. with surface-enhanced laser
desorption/ionization-time of flight ProteinChip technology. Applied and Environmental
Microbiology. 66, 4396-4400.

119
Hsu, C. D., Meaddough, E., Aversa, K., & Copel, J. A. (1998). The role of amniotic fluid L-
selectin, GRO-alpha, and interleukin-8 in the pathogenesis of intraamniotic infection. American
Journal of Obstetrics and Gynecology. 178, 428-432.
Hummelen, R., Fernandes, A.D., Macklaim, J.M., Dickson, R.J., Chanqalucha, J., Gloor,
G.B., Reid, G. (2010). Deep sequencing of vaginal microbiota of women with HIV. PLoS One.
5, 1-9.
Imseis, H. M., Greig, P. C., Livengood, C. H.,3rd, Shunior, E., Durda, P., & Erikson, M.
(1997). Characterization of the inflammatory cytokines in the vagina during pregnancy and labor
and with bacterial vaginosis. Journal of the Society for Gynecologic Investigation. 4, 90-94.
Inoue, H., Kosaka, T., Miyata, A., Hara, S., Yokoyama, C., Nanayama, T., & Tanabe, T.
(1995). Structure and expression of the human prostaglandin endoperoxide synthase 2 gene.
Advances in Prostaglandin, Thromboxane, and Leukotriene Research. 23, 109-111.
Jacobsson, B., Mattsby-Baltzer, I., & Hagberg, H. (2005). Interleukin-6 and interleukin-8 in
cervical and amniotic fluid: Relationship to microbial invasion of the chorioamniotic
membranes. Journal of Obstetrics and Gynaecology. 112, 719-724.
Jakobsson, P. J., Thoren, S., Morgenstern, R., & Samuelsson, B. (1999). Identification of
human prostaglandin E synthase: A microsomal, glutathione-dependent, inducible enzyme,
constituting a potential novel drug target. Proceedings of the National Academy of Sciences of
the United States of America. 96, 7220-7225.

120
Jijon, H., Backer, J., Diaz, H., Yeung, H., Thiel, D., McKaigney, C., De Simone, C., &
Madsen, K. (2004). DNA from probiotic bacteria modulates murine and human epithelial and
immune function. Gastroenterology. 126, 1358-1373.
Johnson, R. F., Mitchell, C. M., Giles, W. B., Bisits, A., & Zakar, T. (2006). Mechanisms
regulating prostaglandin H2 synthase-2 mRNA level in the amnion and chorion during
pregnancy. Journal of Endocrinology. 188, 603-610.
Kallapur, S. G., Moss, T. J., Auten, R. L.,Jr, Nitsos, I., Pillow, J. J., Kramer, B. W., Maeda,
D. Y., Newnham, J. P., Ikegami, M., & Jobe, A. H. (2009). IL-8 signaling does not mediate
intra-amniotic LPS-induced inflammation and maturation in preterm fetal lamb lung. American
Journal of Physiology.Lung Cellular and Molecular Physiology. 297, L512-9.
Kamel, R. M. (2010). The onset of human parturition. Archives of Gynecology and Obstetrics.
281, 975-982.
Karalis, K., Goodwin, G., & Majzoub, J. A. (1996). Cortisol blockade of progesterone: A
possible molecular mechanism involved in the initiation of human labor. Nature Medicine. 2,
556-560.
Katamura, K., Shintake, N., Yamauchi, Y., Fukui, T., Ohshima, Y., Mayumi, M., Furusho,
K. (1995). Prostaglandin E2 at priming of naïve CD4+ cells inhibits acquisition of an ability to
produce IFN-gamma and IL-2, but not IL-4 and IL-5. Journal of Immunology. 155, 4604-4612.
Katz, V. L., & Farmer, R. M. (1999). Controversies in tocolytic therapy. Clinical Obstetrics
and Gynecology. 42, 802-819.

121
Keelan, J. A., Blumenstein, M., Helliwell, R. J., Sato, T. A., Marvin, K. W., & Mitchell, M.
D. (2003). Cytokines, prostaglandins and parturition--a review. Placenta. 24 Suppl A, S33-46.
Kendal-Wright, C. E. (2007). Stretching, mechanotransduction, and proinflammatory cytokines
in the fetal membranes. Reproductive Sciences. 14, 35-41.
Kent, A. S., Sullivan, M. H., & Elder, M. G. (1994). Transfer of cytokines through human fetal
membranes. Journal of Reproduction and Fertility. 100, 81-84.
Kent, A. S., Sullivan, M. H., Sun, M. Y., Zosmer, A., & Elder, M. G. (1993). Effects of
interleukin-6 and tumor necrosis factor-alpha on prostaglandin production by cultured human
fetal membranes. Prostaglandins. 46, 351-359.
Kent, A. S., Sun, M. Y., Sullivan, M. H., & Elder, M. G. (1993). The effects of interleukins 1
alpha and 1 beta on prostaglandin production by cultured human fetal membranes.
Prostaglandins. 46, 51-59.
Kenyon, S. L., Taylor, D. J., Tarnow-Mordi, W., & ORACLE Collaborative Group. (2001).
Broad-spectrum antibiotics for spontaneous preterm labour: The ORACLE II randomised trial.
ORACLE collaborative group. Lancet. 357, 989-994.
Kim, S. O., Sheikh, H. I., Ha, S. D., Martins, A., & Reid, G. (2006). G-CSF-mediated
inhibition of JNK is a key mechanism for Lactobacillus rhamnosus-induced suppression of TNF
production in macrophages. Cellular Microbiology. 8, 1958-1971.
King, A. E., Kelly, R. W., Sallenave, J. M., Bocking, A. D., & Challis, J. R. (2007). Innate
immune defences in the human uterus during pregnancy. Placenta. 28, 1099-1106.

122
Kitterman, J. A. (1987). Arachidonic acid metabolites and control of breathing in the fetus and
newborn. Seminars in Perinatology. 11, 43-52.
Kligler, E., Hanaway, P., Cohrssen, A. (2007). Probiotics in children. Pediatric Clinics of
North America. 54, 949-967.
Kobayashi, K., Inai, T., Shibata, Y., & Yasui, M. (2009). Dynamic changes in amniotic tight
junctions during pregnancy. Placenta. 30, 840-847.
Kobayashi, K., Kadohira, I., Tanaka, M., Yoshimura, Y., Ikeda, K., & Yasui, M. (2010a).
Expression and distribution of tight junction proteins in human amnion during late pregnancy.
Placenta. 31, 158-162.
Kobayashi, K., Miwa, H., Yasui, M. (2010b). Inflammatory mediators weaken the amniotic
membrane barrier through disruption of tight junctions. Journal of Physiology. 588, 4859-4869.
Korhonen, R., Kosonen, O., Korpela, R., & Moilanen, E. (2004). The expression of COX2
protein induced by Lactobacillus rhamnosus GG, endotoxin and lipoteichoic acid in T84
epithelial cells. Letters in Applied Microbiology. 39, 19-24.
Kotzamanidis, C., Kourelis, A., Litopoulou-Tzanetaki E., Tzanetakis, N., Yiangou, M.
(2010). Evaluation of adhesion capacity, cell surface traits and immunomodulatory activity of
presumptive probiotic Lactobacillus strains. International Journal of Food Microbiology. 140,
154-163.
Kudo, I., & Murakami, M. (1999). Diverse functional coupling of prostanoid biosynthetic
enzymes in various cell types. Advances in Experimental Medicine and Biology. 469, 29-35.

123
Lamont, R., Sobel, J., Akins, R., Hassan, S., Chaiworapongsa, T., Kusanovic, J., &
Romero, R. (2011). The vaginal microbiome: New information about genital tract flora using
molecular based techniques. BJOG. 118, 533-549
Lara-Villoslada, F., Sierra, S., Boza, J., Xaus, J., & Olivares, M. (2007). Beneficial effects of
consumption of a dairy product containing two probiotic strains, Lactobacillus coryniformis
CECT5711 and Lactobacillus gasseri CECT5714 in healthy children. Nutricion Hospitalaria. 22,
496-502.
Lee do, K., Park, S.Y., Jang, S., Baek, E.H., Kim, M.J., Huh, S.M., Choir, K.S., Chung,
M.J., Kim, J.E., Lee, K.O., Ha. N.J. (2011). The combination of mixed lactic acid bacteria and
dietary fiber lowers serum cholesterol levels and fecal harmful enzyme activities in rats.
Archives of Pharmacology Research. 34, 23-29.
Leitich, H., Brunbauer, M., Bodner-Adler, B., Kaider, A., Egarter, C., & Husslein, P.
(2003). Antibiotic treatment of bacterial vaginosis in pregnancy: A meta-analysis. American
Journal of Obstetrics and Gynecology. 188, 752-758.
Li, A., Dubey, S., Varney, M. L., Dave, B. J., & Singh, R. K. (2003). IL-8 directly enhanced
endothelial cell survival, proliferation, and matrix metalloproteinases production and regulated
angiogenesis. Journal of Immunology (Baltimore, Md.: 1950). 170, 3369-3376.
Livengood, C. H. (2009). Bacterial vaginosis: An overview for 2009. Reviews in Obstetrics and
Gynecology. 2, 28-37.

124
Locksmith, G., & Duff, P. (2001). Infection, antibiotics, and preterm delivery. Seminars in
Perinatology. 25, 295-309.
MacPhee, R. A., Hummelen, R., Bisanz, J. E., Miller, W. L., & Reid, G. (2010). Probiotic
strategies for the treatment and prevention of bacterial vaginosis. Expert Opinion on
Pharmacotherapy. 11, 2985-2995.
Madsen, K., Cornish, A., Soper, P., McKaigney, C., Jijon, H., Yachimec, C., Doyle, J.,
Jewell, L., & De Simone, C. (2001). Probiotic bacteria enhance murine and human intestinal
epithelial barrier function. Gastroenterology. 121, 580-591.
Mancini, J. A., Blood, K., Guay, J., Gordon, R., Claveau, D., Chan, C. C., & Riendeau, D.
(2001). Cloning, expression, and up-regulation of inducible rat prostaglandin e synthase during
lipopolysaccharide-induced pyresis and adjuvant-induced arthritis. Journal of Biological
Chemistry. 276, 4469-4475.
Masferrer, J. L., Seibert, K., Zweifel, B., & Needleman, P. (1992). Endogenous
glucocorticoids regulate an inducible cyclooxygenase enzyme. Proceedings of the National
Academy of Sciences of the United States of America. 89, 3917-3921.
McDonald, H. M., Brocklehurst, P., & Gordon, A. (2007). Antibiotics for treating bacterial
vaginosis in pregnancy. Cochrane Database of Systematic Reviews (Online). (1), CD000262.
Meadows, J. W., Eis, A. L., Brockman, D. E., & Myatt, L. (2003). Expression and localization
of prostaglandin E synthase isoforms in human fetal membranes in term and preterm labor.
Journal of Clinical Endocrinology and Metabolism. 88, 433-439.

125
Mehta, A., Brewington, R., Chatterji, M., Zoubine, M., Kinasewitz, G. T., Peer, G. T.,
Chang, A. C., Taylor, F. B.,Jr, & Shnyra, A. (2004). Infection-induced modulation of m1 and
m2 phenotypes in circulating monocytes: Role in immune monitoring and early prognosis of
sepsis. Shock (Augusta, Ga.). 22, 423-430.
Meis, P. J., Klebanoff, M., Thom, E., Dombrowski, M. P., Sibai, B., Moawad, A. H., Spong,
C. Y., Hauth, J. C., Miodovnik, M., Varner, M. W., Leveno, K. J., Caritis, S. N., Iams, J. D.,
Wapner, R. J., Conway, D., O'Sullivan, M. J., Carpenter, M., Mercer, B., Ramin, S. M.,
Thorp, J. M., Peaceman, A. M., Gabbe, S., & National Institute of Child Health and
Human Development Maternal-Fetal Medicine Units Network. (2003). Prevention of
recurrent preterm delivery by 17 alpha-hydroxyprogesterone caproate. New England Journal of
Medicine. 348, 2379-2385.
Meis, P. J., Michielutte, R., Peters, T. J., Wells, H. B., Sands, R. E., Coles, E. C., & Johns,
K. A. (1995). Factors associated with preterm birth in cardiff, wales. II. indicated and
spontaneous preterm birth. American Journal of Obstetrics and Gynecology. 173, 597-602.
Menard, S., Candalh, C., Bambou, J. C., Terpend, K., Cerf-Bensussan, N., & Heyman, M.
(2004). Lactic acid bacteria secrete metabolites retaining anti-inflammatory properties after
intestinal transport. Gut. 53, 821-828.
Menon, R., & Fortunato, S. J. (2004). Fetal membrane inflammatory cytokines: A switching
mechanism between the preterm premature rupture of the membranes and preterm labor
pathways. Journal of Perinatal Medicine. 32, 391-399.

126
Menon, R., Swan, K. F., Lyden, T. W., Rote, N. S., & Fortunato, S. J. (1995). Expression of
inflammatory cytokines (interleukin-1β and interleukin-6) in amniochorionic membranes.
American Journal of Obstetrics and Gynecology. 172, 493-500.
Merlino, A., Welsh, T., Erdonmez, T., Madsen, G., Zakar, T., Smith, R., Mercer, B., &
Mesiano, S. (2009). Nuclear progesterone receptor expression in the human fetal membranes
and decidua at term before and after labor. Reproductive Sciences. 16, 357-363.
Merlino, A. A., Welsh, T. N., Tan, H., Yi, L. J., Cannon, V., Mercer, B. M., & Mesiano, S.
(2007). Nuclear progesterone receptors in the human pregnancy myometrium: Evidence that
parturition involves functional progesterone withdrawal mediated by increased expression of
progesterone receptor-A. Journal of Clinical Endocrinology and Metabolism. 92, 1927-1933.
Mesiano, S., Chan, E. C., Fitter, J. T., Kwek, K., Yeo, G., & Smith, R. (2002). Progesterone
withdrawal and estrogen activation in human parturition are coordinated by progesterone
receptor A expression in the myometrium. Journal of Clinical Endocrinology and Metabolism.
87, 2924-2930.
Metcalf, D., & Moore, J. G. (1988). Divergent disease patterns in granulocyte-macrophage
colony-stimulating factor transgenic mice associated with different transgene insertion sites.
Proceedings of the National Academy of Sciences of the United States of America. 85, 7767-
7771.
Mijovic, J. E., Zakar, T., Angelova, J., & Olson, D. M. (1999). Prostaglandin endoperoxide H
synthase mRNA expression in the human amnion and decidua during pregnancy and in the
amnion at preterm labour. Molecular Human Reproduction. 5, 182-187.

127
Mitchell, B. F., & Wong, S. (1995). Metabolism of oxytocin in human decidua, chorion, and
placenta. Journal of Clinical Endocrinology and Metabolism. 80, 2729-2733.
Mitchell, M. D., Dudley, D. J., Edwin, S. S., & Schiller, S. L. (1991). Interleukin-6 stimulates
prostaglandin production by human amnion and decidual cells. European Journal of
Pharmacology. 192, 189-191.
Mitchell, M. D., Edwin, S. S., Lundin-Schiller, S., Silver, R. M., Smotkin, D., & Trautman,
M. S. (1993). Mechanism of interleukin-1 beta stimulation of human amnion prostaglandin
biosynthesis: Mediation via a novel inducible cyclooxygenase. Placenta. 14, 615-625.
Mitchell, M. D., Romero, R. J., Edwin, S. S., & Trautman, M. S. (1995). Prostaglandins and
parturition. Reproduction, Fertility, and Development. 7, 623-632.
Mitchell, M. D., Simpson, K. L., & Keelan, J. A. (2004). Paradoxical proinflammatory actions
of interleukin-10 in human amnion: Potential roles in term and preterm labour. Journal of
Clinical Endocrinology and Metabolism. 89, 4149-4152.
Mohan, A. R., Sooranna, S. R., Lindstrom, T. M., Johnson, M. R., & Bennett, P. R. (2007).
The effect of mechanical stretch on cyclooxygenase type 2 expression and activator protein-1
and nuclear factor-kappaB activity in human amnion cells. Endocrinology. 148, 1850-1857.
Molnar, M., Rigo, J.,Jr, Romero, R., & Hertelendy, F. (1999). Oxytocin activates mitogen-
activated protein kinase and up-regulates cyclooxygenase-2 and prostaglandin production in
human myometrial cells. American Journal of Obstetrics and Gynecology. 181, 42-49.

128
Moorthy, G., Murali, M. R., & Devaraj, S. N. (2009). Lactobacilli facilitate maintenance of
intestinal membrane integrity during Shigella dysenteriae 1 infection in rats. Nutrition. 25, 350-
358.
Mosser, D.M., Edwards, J.P. (2008). Exploring the full spectrum of macrophage activation.
Nature Reviews: Immunology. 8, 958-969.
Murakami, M., Nakashima, K., Kamei, D., Masuda, S., Ishikawa, Y., Ishii, T., Ohmiya, Y.,
Watanabe, K., & Kudo, I. (2003). Cellular prostaglandin E2 production by membrane-bound
prostaglandin E synthase-2 via both cyclooxygenases-1 and -2. Journal of Biological Chemistry.
278, 37937-37947.
Murphy, S. P., Tayade, C., Ashkar, A. A., Hatta, K., Zhang, J., & Croy, B. A. (2009).
Interferon gamma in successful pregnancies. Biology of Reproduction. 80, 848-859.
Myatt, L., & Sun, K. (2010). Role of fetal membranes in signaling of fetal maturation and
parturition. International Journal of Developmental Biology. 54, 545-553.
Nataraj, C., Thomas, D. W., Tilley, S. L., Nguyen, M. T., Mannon, R., Koller, B.H.,
Coffman, T. M. (2001). Receptors for prostaglandin E(2) that regulate cellular immune
responses in the mouse. Journal of Clinical Investigation. 108(8), 1229-1235.
Norwitz, E. R., Robinson, J. N., & Challis, J. R. (1999). The control of labor. New England
Journal of Medicine. 341, 660-666.

129
Nugent, R. P., Krohn, M. A., & Hillier, S. L. (1991). Reliability of diagnosing bacterial
vaginosis is improved by a standardized method of gram stain interpretation. Journal of Clinical
Microbiology. 29, 297-301.
Ohara, T., Yoshino, K., Kitajima, M. (2010). Possibility of preventing colorectal
carcinogenesis with probiotics. Hepatogastroenterology. 57, 1411-1415.
Ojetti, V., Gigante, G., Gabrielli, M., Ainora, M.E., Mannocci, A., Lauritano, E.C.,
Gasbarrini, G., Gasbarrini, A. (2010). The effect of oral supplementation with Lactobacillus
reuteri or tilactase in lactose intolerant patients: randomized trial. European review for medical
and pharmacological sciences. 14, 163-170.
Okun, N., Gronau, K. A., & Hannah, M. E. (2005). Antibiotics for bacterial vaginosis or
trichomonas vaginalis in pregnancy: A systematic review. Obstetrics and Gynecology. 105, 857-
868.
Olson, D. M. (2003). The role of prostaglandins in the initiation of parturition. Best Practice &
Research. Clinical Obstetrics & Gynaecology. 17, 717-730.
Olson, D. M., Skinner, K., & Challis, J. R. (1983). Prostaglandin output in relation to
parturition by cells dispersed from human intrauterine tissues. Journal of Clinical Endocrinology
and Metabolism. 57, 694-699.
Othman, M., Neilson, J. P., & Alfirevic, Z. (2007). Probiotics for preventing preterm labour.
Cochrane Database of Systematic Reviews (Online). 1, CD005941.

130
Ou, C.W., Chen, Z.Q., Qi, S., Lye, S.J. (1998) Increased expression of the rate myometrial
oxytocin receptor messenger ribonucleic acid during labor requires both mechanical and
hormonal signals. Biology of Reproduction. 59(5), 1055-1061.
Parry, S., & Strauss, J. F.,3rd. (1998). Premature rupture of the fetal membranes. New England
Journal of Medicine. 338, 663-670.
Pavlova, S. I., Kilic, A. O., Kilic, S. S., So, J. S., Nader-Macias, M. E., Simoes, J. A., & Tao,
L. (2002). Genetic diversity of vaginal lactobacilli from women in different countries based on
16S rRNA gene sequences. Journal of Applied Microbiology. 92, 451-459.
Petersen, A. M., & Pedersen, B. K. (2006). The role of IL-6 in mediating the anti-inflammatory
effects of exercise. Journal of Physiology and Pharmacology. 57, 43-51.
Pollard, J. K., & Mitchell, M. D. (1993). Tumor necrosis factor alpha stimulates amnion
prostaglandin biosynthesis primarily via an action on fatty acid cyclooxygenase. Prostaglandins.
46, 499-510.
Pomini, F., Caruso, A., & Challis, J. R. (1999). Interleukin-10 modifies the effects of
interleukin-1beta and tumor necrosis factor-alpha on the activity and expression of prostaglandin
H synthase-2 and the NAD+-dependent 15-hydroxyprostaglandin dehydrogenase in cultured
term human villous trophoblast and chorion trophoblast cells. Journal of Clinical Endocrinology
and Metabolism. 84, 4645-4651.

131
Potestio, F. A., Zakar, T., & Olson, D. M. (1988). Glucocorticoids stimulate prostaglandin
synthesis in human amnion cells by a receptor-mediated mechanism. Journal of Clinical
Endocrinology and Metabolism. 67, 1205-1210.
Premyslova, M., Li, W., Alfaidy, N., Bocking, A. D., Campbell, K., Gibb, W., & Challis, J.
R. (2003). Differential expression and regulation of microsomal prostaglandin E(2) synthase in
human fetal membranes and placenta with infection and in cultured trophoblast cells. Journal of
Clinical Endocrinology and Metabolism. 88, 6040-6047.
Rachmilewitz, D., Katakura, K., Karmeli, F., Hayashi, T., Reinus, C., Rudensky, B., Akira,
S., Takeda, K., Lee, J., Takabayashi, K., & Raz, E. (2004). Toll-like receptor 9 signaling
mediates the anti-inflammatory effects of probiotics in murine experimental colitis.
Gastroenterology. 126, 520-528.
Rankin, J. G. (1976). A role for prostaglandins in the regulation of the placental blood flows.
Prostaglandins. 11, 343-353.
Reid, G. (1999). The scientific basis for probiotic strains of Lactobacillus. Applied and
Environmental Microbiology. 65, 3763-3766.
Reid, G. (2008). Probiotic lactobacilli for urogenital health in women. Journal of Clinical
Gastroenterology. 42, S234-S236.
Reid, G., Beuerman, D., Heinemann, C., & Bruce, A. W. (2001a). Probiotic Lactobacillus
dose required to restore and maintain a normal vaginal flora. FEMS Immunology and Medical
Microbiology. 32, 37-41.

132
Reid, G., & Bocking, A. (2003). The potential for probiotics to prevent bacterial vaginosis and
preterm labor. American Journal of Obstetrics and Gynecology. 189, 1202-1208.
Reid, G., Bruce, A.W., Fraser, N., Heinemann, C., Owen, J., Henning, B. (2001b). Oral
probiotics can resolve urogenital infections. FEMS Immunology and Medical Microbiology. 30,
49-52.
Reid, G., Charbonneau, D., Erb, J., Kochanowski, B., Beuerman, D., Poehner, R., & Bruce,
A. W. (2003). Oral use of Lactobacillus rhamnosus GR-1 and L. fermentum RC-14 significantly
alters vaginal flora: Randomized, placebo-controlled trial in 64 healthy women. FEMS
Immunology and Medical Microbiology. 35, 131-134.
Reid, G., Cook, R. L., & Bruce, A. W. (1987). Examination of strains of lactobacilli for
properties that may influence bacterial interference in the urinary tract. Journal of Urology. 138,
330-335.
Reid, G., Millsap, K., & Bruce, A. W. (1994). Implantation of Lactobacillus casei var
rhamnosus into vagina. Lancet. 344, 1229.
Reid, G., Tieszer, C., & Lam, D. (1995). Influence of lactobacilli on the adhesion of
staphylococcus aureus and candida albicans to fibers and epithelial cells. Journal of Industrial
Microbiology. 15, 248-253.
Reid, G., Younes, J. A., Van der Mei, H. C., Gloor, G. B., Knight, R., & Busscher, H. J.
(2011). Microbiota restoration: Natural and supplemented recovery of human microbial
communities. Nature Reviews. Microbiology. 9, 27-38.

133
Reisenberger, K., Egarter, C., Knofler, M., Schiebel, I., Gregor, H., Hirschl, A. M., Heinze,
G., & Husslein, P. (1998). Cytokine and prostaglandin production by amnion cells in response
to the addition of different bacteria. American Journal of Obstetrics and Gynecology. 178, 50-53.
Riemer, R. K., & Heymann, M. A. (1998). Regulation of uterine smooth muscle function
during gestation. Pediatric Research. 44, 615-627.
Robertson, S. A., Mayrhofer, G., & Seamark, R. F. (1992). Uterine epithelial cells synthesize
granulocyte-macrophage colony-stimulating factor and interleukin-6 in pregnant and non-
pregnant mice. Biology of Reproduction. 46, 1069-1079.
Robinson, J. N., Regan, J. A., & Norwitz, E. R. (2001). The epidemiology of preterm labor.
Seminars in Perinatology. 25, 204-214.
Romero, R., Brody, D. T., Oyaizu, E., Mazor, M., Wu, Y. K., Hobbins, J. C., & Durum, S.
K. (1989a). Infection and labor. III. interleukin-1: A signal for the onset of parturition. American
Journal of Obstetrics and Gynecology. 160, 1117-1123.
Romero, R., Ceska, M., Avila, C., Mazor, M., Behnke, E., & Lindley, I. (1991). Neutrophil
attractant/activating peptide-1/interleukin-8 in term and preterm parturition. American Journal of
Obstetrics and Gynecology. 165, 813-820.
Romero, R., Chaiworapongsa, T., Kuivaniemi, H., & Tromp, G. (2004). Bacterial vaginosis,
the inflammatory response and the risk of preterm birth: A role for genetic epidemiology in the
prevention of preterm birth. American Journal of Obstetrics and Gynecology. 190, 1509-1519.

134
Romero, R., Durum, S., Dinarello, C. A., Oyarzun, E., Hobbins, J. C., & Mitchell, M. D.
(1989b). Interleukin-1 stimulates prostaglandin biosynthesis by human amnion. Prostaglandins.
37, 13-22.
Romero, R., Espinoza, J., Chaiworapongsa, T., & Kalache, K. (2002). Infection and
prematurity and the role of preventive strategies. Seminars in Neonatology. 7, 259-274.
Romero, R., Hobbins, J. C., & Mitchell, M. D. (1988a). Endotoxin stimulates prostaglandin E2
production by human amnion. Obstetrics and Gynecology. 71, 227-228.
Romero, R., Manogue, K. R., Mitchell, M. D., Wu, Y. K., Oyarzun, E., Hobbins, J. C., &
Cerami, A. (1989c). Infection and labor. IV. cachectin-tumor necrosis factor in the amniotic
fluid of women with intraamniotic infection and preterm labor. American Journal of Obstetrics
and Gynecology. 161, 336-341.
Romero, R., Roslansky, P., Oyarzun, E., Wan, M., Emamian, M., Novitsky, T. J., Gould,
M. J., & Hobbins, J. C. (1988b). Labor and infection. II. bacterial endotoxin in amniotic fluid
and its relationship to the onset of preterm labor. American Journal of Obstetrics and
Gynecology. 158, 1044-1049.
Romero, R., Sirtori, M., Oyarzun, E., Avila, C., Mazor, M., Callahan, R., Sabo, V.,
Athanassiadis, A. P., & Hobbins, J. C. (1989d). Infection and labor. V. prevalence,
microbiology, and clinical significance of intraamniotic infection in women with preterm labor
and intact membranes. American Journal of Obstetrics and Gynecology. 161, 817-824.

135
Rose, M. P., Myatt, L., & Elder, M. G. (1990). Pathways of arachidonic acid metabolism in
human amnion cells at term. Prostaglandins, Leukotrienes, and Essential Fatty Acids. 39, 303-
309.
Saji, F., Samejima, Y., Kamiura, S., Sawai, K., Shimoya, K., & Kimura, T. (2000). Cytokine
production in chorioamnionitis. Journal of Reproductive Immunology. 47, 185-196.
Sangha, R. K., Walton, J. C., Ensor, C. M., Tai, H. H., & Challis, J. R. (1994).
Immunohistochemical localization, messenger ribonucleic acid abundance, and activity of 15-
hydroxyprostaglandin dehydrogenase in placenta and fetal membranes during term and preterm
labor. Journal of Clinical Endocrinology and Metabolism. 78, 982-989.
Sastry, B. V., Hemontolor, M. E., Chance, M. B., & Johnson, R. F. (1997). Dual messenger
function for prostaglandin E2 (PGE2) in human placenta. Cellular and Molecular Biology. 43,
417-424.
Saunders, S., Bocking, A., Challis, J., & Reid, G. (2007). Effect of Lactobacillus challenge on
gardnerella vaginalis biofilms. Colloids and Surfaces. B, Biointerfaces. 55, 138-142.
Saunders-Kirkwood, K., Cates, J. A., & Roslyn, J. J. (1993). Prostaglandin E2 stimulates ion
transport in prairie dog gallbladder. Digestive Diseases and Sciences. 38, 167-172.
Scales, W. E., Chensue, S. W., Otterness, I., Kunkel, S. L. (1989). Regulation of monokine
gene expression: prostaglandin E2 suppresses tumor necrosis factor but not interleukin-1 alpha or
beta-mRNA and cell-associated bioactivity. Journal of Leukocyte Biology. 45(5), 416-421.

136
Schindler, R., Mancilla, J., Endres, S., Ghorbani, R., Clark, S. C., & Dinarello, C. A.
(1990). Correlations and interactions in the production of interleukin-6 (IL-6), IL-1, and tumor
necrosis factor (TNF) in human blood mononuclear cells: IL-6 suppresses IL-1 and TNF. Blood.
75, 40-47.
Seong, H., Kang, Y. D., Lee, S. I. E., Shim, S. S., Romero, R., & Yoon, B. H. (2006). Lack of
evidence for microorganisms in most women with clinical chorioamnionitis: A need to revisit the
clinical and microbiologic criteria for one of the most important obstetrical complications.
American Journal of Obstetrics and Gynecology. 195, S73-S73.
Servin, A. L. (2004). Antagonistic activities of lactobacilli and bifidobacteria against microbial
pathogens. FEMS Microbiology Reviews. 28, 405-440.
Shinomiya, S., Naraba, H., Ueno, A., Utsunomiya, I., Maruyama, T., Ohuchida, S.,
Ushikubi, F., Yuki, K., Narumiya, S., Sugimoto, Y., Ichikawa, A., Oh-ishi, S. (2001).
Regulation of TNFalpha and interleukine-10 production by prostaglandins I(2) and E(2): studies
with prostaglandin receptor-deficient mice and prostaglandin E-receptor subtype-selective
synthetic agonists. Biochemical Pharmacology. 61(9), 1153-1160.
Shon, Y.H., Kim, J.H., Nam, K.S. (2002). Effect of Astragali radix extract on
lipopolysaccharide-induced inflammation in human amnion. Biol Pharm Bull. 25, 77-80.
Shynlova, O., Mitchell, J. A., Tsampalieros, A., Langille, B. L., & Lye, S. J. (2004).
Progesterone and gravidity differentially regulate expression of extracellular matrix components
in the pregnant rat myometrium. Biology of Reproduction. 70, 986-992.

137
Shynlova, O., Tsui, P., Dorogin, A., Chow, M., & Lye, S. J. (2005). Expression and
localization of alpha-smooth muscle and gamma-actins in the pregnant rat myometrium. Biology
of Reproduction. 73, 773-780.
Shynlova, O., Tsui, P., Jaffer, S., & Lye, S. J. (2009). Integration of endocrine and mechanical
signals in the regulation of myometrial functions during pregnancy and labour. European Journal
of Obstetrics, Gynecology, and Reproductive Biology. 144 Suppl 1, S2-10.
Sierra-Filardi, E., Puig-Kroger, A., Blanco, F. J., Nieto, C., Bragado, R., Palomero, M. I.,
Bernabeu, C., Vega, M. A., & Corbi, A. L. (2011). Activin A skews macrophage polarization
by promoting a pro-inflammatory phenotype and inhibiting the acquisition of anti-inflammatory
macrophage markers. Blood. epub ahead of print
Silver, R. M., Edwin, S. S., Umar, F., Dudley, D. J., Branch, D. W., & Mitchell, M. D.
(1997). Bacterial lipopolysaccharide-mediated murine fetal death: The role of interleukin-1.
American Journal of Obstetrics and Gynecology. 176, 544-549.
Silver, R. M., Lohner, W. S., Daynes, R. A., Mitchell, M. D., & Branch, D. W. (1994).
Lipopolysaccharide-induced fetal death: The role of tumor-necrosis factor alpha. Biology of
Reproduction. 50, 1108-1112.
Skinner, K. A., & Challis, J. R. (1985). Changes in the synthesis and metabolism of
prostaglandins by human fetal membranes and decidua at labor. American Journal of Obstetrics
and Gynecology. 151, 519-523.

138
Slater, D., Allport, V., & Bennett, P. (1998). Changes in the expression of the type-2 but not
the type-1 cyclo-oxygenase enzyme in chorion-decidua with the onset of labour. British Journal
of Obstetrics and Gynaecology. 105, 745-748.
Smieja, Z., Zakar, T., & Olson, D. M. (1993). Stimulation of cultured amnion cell
prostaglandin endoperoxide H synthase activity by glucocorticoids and phorbol ester. American
Journal of Obstetrics and Gynecology. 169, 653-661.
Smith, R., & Nicholson, R. C. (2007). Corticotrophin releasing hormone and the timing of birth.
Frontiers in Bioscience. 12, 912-918.
Smith, R., Smith, J. I., Shen, X., Engel, P. J., Bowman, M. E., McGrath, S. A., Bisits, A. M.,
McElduff, P., Giles, W. B., & Smith, D. W. (2009). Patterns of plasma corticotropin-releasing
hormone, progesterone, estradiol, and estriol change and the onset of human labor. Journal of
Clinical Endocrinology and Metabolism. 94, 2066-2074.
Smith, W. L., Rollins, T. E., & DeWitt, D. L. (1981). Subcellular localization of prostaglandin
forming enzymes using conventional and monoclonal antibodies. Progress in Lipid Research. 20,
103-110.
Smulian, J. C., Shen-Schwarz, S., Vintzileos, A. M., Lake, M. F., & Ananth, C. V. (1999).
Clinical chorioamnionitis and histologic placental inflammation. Obstetrics and Gynecology. 94,
1000-1005.

139
So, J.S., Song, M.K., Kwon, H.K., Lee, C.G., Chae, C.S., Sahoo, A., Jash, A., Lee, S.H.,
Park, Z.Y., Im, S.H. (2011). Lactobacillus casei enhances type II collagen/glucosamine-
mediated suppression of inflammatory responses in experimental osteoarthritis. 88, 358-366.
So, T. (1993). The role of matrix metalloproteinases for premature rupture of the membranes].
Nippon Sanka Fujinka Gakkai Zasshi. 45, 227-233.
Song, Y. L., Kato, N., Matsumiya, Y., Liu, C. X., Kato, H., & Watanabe, K. (1999).
Identification of and hydrogen peroxide production by fecal and vaginal lactobacilli isolated
from japanese women and newborn infants. Journal of Clinical Microbiology. 37, 3062-3064.
Soper, D. E. (1993). Bacterial vaginosis and postoperative infections. American Journal of
Obstetrics and Gynecology. 169, 467-469.
Spiegel, C. A. (1991). Bacterial vaginosis. Clinical Microbiology Reviews. 4, 485-502.
Stallmach, T., Hebisch, G., Joller, H., Kolditz, P., & Engelmann, M. (1995). Expression
pattern of cytokines in the different compartments of the feto-maternal unit under various
conditions. Reproduction, Fertility, and Development. 7, 1573-1580.
Steele, E. C.,Jr, Kerscher, S., Lyon, M. F., Glenister, P. H., Favor, J., Wang, J., & Church,
R. L. (1997). Identification of a mutation in the MP19 gene, Lim2, in the cataractous mouse
mutant To3. Molecular Vision. 3, 5.
Steensberg, A., Fischer, C. P., & Keller, C. (2003). IL-6 enhances plasma IL-1ra, IL-10, and
cortisol in humans. American Journal of Physiology and Endocrinology Metabolism. 285, 433-7.

140
Stevens, M.Y., Challis, J.R., Lye, S.J. (1998). Corticotropin-releasing hormone receptor
subtype 1 is significantly up-regulated at the time of labor in the human myometrium. Journal of
Clinical Endocrinology and Metabolism. 83, 4107-4115.
Sun, K., Myatt, L. (2003). Enhancement of glucocorticoid-induced 11beta-hydroxysteroid
dehydrogenase type 1 expression by proinflammatory cytokines in cultured human amnion
fibroblasts. Endocrinology. 144, 5568-5677.
Sun M., Ramirez M., Challis J. R., Gibb W. (1996). Immunohistochemical localization of the
glucocorticoid receptor in human fetal membranes and decidua at term and preterm delivery.
Journal of Endocrinology. 149, 243-248.
Svinarich, D. M., Bitonti, O. M., Araneda, H., Romero, R., & Gonik, B. (1996). Induction
and postranslational expression of G-CSF and RANTES in a first trimester trophoblast cell line
by lipopolysaccharide. American Journal of Reproductive Immunology (New York, N.Y.: 1989).
36, 256-259.
Sweet, R. L. (1995). Role of bacterial vaginosis in pelvic inflammatory disease. Clinical
Infectious Diseases. 20, 271-275.
Tazawa, R., Xu, X. M., Wu, K. K., & Wang, L. H. (1994). Characterization of the genomic
structure, chromosomal location and promoter of human prostaglandin H synthase-2 gene.
Biochemical and Biophysical Research Communications. 203, 190-199.
Teixeira, F. J., Zakar, T., Hirst, J. J., Guo, F., Sadowsky, D. W., Machin, G., Demianczuk,
N., Resch, B., & Olson, D. M. (1994). Prostaglandin endoperoxide-H synthase (PGHS) activity

141
and immunoreactive PGHS-1 and PGHS-2 levels in human amnion throughout gestation, at
term, and during labor. Journal of Clinical Endocrinology and Metabolism. 78, 1396-1402.
Thiex, N. W., Chames, M. C., & Loch-Caruso, R. K. (2009). Tissue-specific cytokine release
from human extra-placental membranes stimulated by lipopolysaccharide in a two-compartment
tissue culture system. Reproductive Biology and Endocrinology. 7, 117-127.
Thorburn, G. D. (1992). The placenta, PGE2 and parturition. Early Human Development. 29,
63-73.
Thoren, S., & Jakobsson, P. J. (2000). Coordinate up- and down-regulation of glutathione-
dependent prostaglandin E synthase and cyclooxygenase-2 in A549 cells. inhibition by NS-398
and leukotriene C4. European Journal of Biochemistry. 267, 6428-6434.
Thoren, S., Weinander, R., Saha, S., Jegerschold, C., Pettersson, P. L., Samuelsson, B.,
Hebert, H., Hamberg, M., Morgenstern, R., & Jakobsson, P. J. (2003). Human microsomal
prostaglandin E synthase-1: Purification, functional characterization, and projection structure
determination. Journal of Biological Chemistry. 278, 22199-22209.
Tilg, H., Trehu, E., Atkins, M. B., Dinarello, C. A., & Mier, J. W. (1994). Interleukin-6 (IL-6)
as an anti-inflammatory cytokine: Induction of circulating IL-1 receptor antagonist and soluble
tumor necrosis factor receptor p55. Blood. 83, 113-118.
Tomeczek, L., Reid, G., Cuperus, P. L., McGroarty, J. A., van der Mei, H. C., Bruce, A.
W., Khoury, A. E., & Busscher, H. J. (1992). Correlation between hydrophobicity and

142
resistance to nonoxynol-9 and vancomycin for urogenital isolates of lactobacilli. FEMS
Microbiology Letters. 73, 101-104.
Unlugedik, E., Alfaidy, N., Holloway, A., Lye, S., Bocking, A., Challis, J., & Gibb, W.
(2010). Expression and regulation of prostaglandin receptors in the human placenta and fetal
membranes at term and preterm. Reproduction, Fertility, and Development. 22, 796-807.
van der Pouw Kraan, T.C., Boeije, L.C., Smeenk, R.J., Wijdenes, J., Aarden, L. A. (1995).
Prostaglandin-E2 is a potent inhibitor of human interleukin 12 production. Journal of
Experimental Medicine. 181, 775-779.
Van Meir, C. A., Matthews, S. G., Keirse, M. J., Ramirez, M. M., Bocking, A., & Challis, J.
R. (1997). 15-hydroxyprostaglandin dehydrogenase: Implications in preterm labor with and
without ascending infection. Journal of Clinical Endocrinology and Metabolism. 82, 969-976.
Van Meir, C. A., Sangha, R. K., Walton, J. C., Matthews, S. G., Keirse, M. J., & Challis, J.
R. (1996). Immunoreactive 15-hydroxyprostaglandin dehydrogenase (PGDH) is reduced in fetal
membranes from patients at preterm delivery in the presence of infection. Placenta. 17, 291-297.
Vasquez, A., Jakobsson, T., Ahrne, S., Forsum, U., & Molin, G. (2002). Vaginal
Lactobacillus flora of healthy Swedish women. Journal of Clinical Microbiology. 40, 2746-2749.
Veith, G. L., & Rice, G. E. (1999). Interferon gamma expression during human pregnancy and
in association with labour. Gynecologic and Obstetric Investigation. 48, 163-167.

143
Velraeds, M. M., van de Belt-Gritter, B., van der Mei, H. C., Reid, G., & Busscher, H. J.
(1998). Interference in initial adhesion of uropathogenic bacteria and yeasts to silicone rubber by
a Lactobacillus acidophilus biosurfactant. Journal of Medical Microbiology. 47, 1081-1085.
Velraeds, M. M., van der Mei, H. C., Reid, G., & Busscher, H. J. (1996). Inhibition of initial
adhesion of uropathogenic enterococcus faecalis by biosurfactants from Lactobacillus isolates.
Applied and Environmental Microbiology. 62, 1958-1963.
Vercauteren, M., Palit, S., Soetens, F., Jacquemyn, Y., & Alahuhta, S. (2009).
Anaesthesiological considerations on tocolytic and uterotonic therapy in obstetrics. Acta
Anaesthesiologica Scandinavica. 53, 701-709.
Verna, E.C., Lucak, S. (2010). Use of probiotics in gastrointestinal disorders: what to
recommend? Therapeutic advances in gastroenterology. 3, 307-319.
Vogel, I., Thorsen, P., Curry, A., Sandager, P., & Uldbjerg, N. (2005). Biomarkers for the
prediction of preterm delivery. Acta Obstetricia Et Gynecologica Scandinavica. 84, 516-525.
Voltan, S., Martines, D., Elli, M., Brun, P., Longo, S., Porzionato, A., Macchi, V., D'Inca,
R., Scarpa, M., Palu, G., Sturniolo, G. C., Morelli, L., & Castagliuolo, I. (2008).
Lactobacillus crispatus M247-derived H2O2 acts as a signal transducing molecule activating
peroxisome proliferator activated receptor-gamma in the intestinal mucosa. Gastroenterology.
135, 1216-1227.
Whittle, W.L., Benzie, R.J., Gibb, W. (1994). Influence of culture media on prostaglandin
output by dispersed amnion cells. Prostaglandins Leukot Essent Fatty Acids. 50, 93-96.

144
Whittle, W. L., Gibb, W., & Challis, J. R. (2000). The characterization of human amnion
epithelial and mesenchymal cells: The cellular expression, activity and glucocorticoid regulation
of prostaglandin output. Placenta. 21, 394-401.
Whittle, W. L., Patel, F. A., Alfaidy, N., Holloway, A. C., Fraser, M., Gyomorey, S., Lye, S.
J., Gibb, W., & Challis, J. R. (2001). Glucocorticoid regulation of human and ovine parturition:
The relationship between fetal hypothalamic-pituitary-adrenal axis activation and intrauterine
prostaglandin production. Biology of Reproduction. 64, 1019-1032.
Witkin, S. S., Linhares, I. M., Bongiovanni, A. M., Herway, C., & Skupski, D. (2011).
Unique alterations in infection-induced immune activation during pregnancy. BJOG. 118, 145-
153.
Xing, Z., Gauldie, J., Cox, G., Baumann, H., Jordana, M., Lei, X. F., & Achong, M. K.
(1998). IL-6 is an anti-inflammatory cytokine required for controlling local or systemic acute
inflammatory responses. Journal of Clinical Investigation. 101, 311-320.
Xu, H. Y., Tian, W. H., Wan, C. X., Jia, L. J., Wang, L. Y., Yuan, J., Liu, C. M., Zeng, M.,
& Wei, H. (2008). Antagonistic potential against pathogenic microorganisms and hydrogen
peroxide production of indigenous lactobacilli isolated from vagina of Chinese pregnant women.
Biomedical and Environmental Sciences. 21, 365-371.
Xu, X. M., Hajibeige, A., Tazawa, R., Loose-Mitchell, D., Wang, L. H., & Wu, K. K. (1995).
Characterization of human prostaglandin H synthase genes. Advances in Prostaglandin,
Thromboxane, and Leukotriene Research. 23, 105-107.

145
Yadav, A., Saini, V., & Arora, S. (2010). MCP-1: Chemoattractant with a role beyond
immunity: A review. Clinica Chimica Acta. 411, 1570-1579.
Yamamoto, T., Zhou, X., Williams, C. J., Hochwalt, A., & Forney, L. J. (2009). Bacterial
populations in the vaginas of healthy adolescent women. Journal of Pediatric and Adolescent
Gynecology. 22, 11-18.
Yan, F., & Polk, D. B. (2002). Probiotic bacterium prevents cytokine-induced apoptosis in
intestinal epithelial cells. Journal of Biological Chemistry. 277, 50959-50965.
Yeganegi, M., Leung, C. G., Martins, A., Kim, S. O., Reid, G., Challis, J. R., & Bocking, A.
D. (2010). Lactobacillus rhamnosus GR-1-induced IL-10 production in human placental
trophoblast cells involves activation of JAK/STAT and MAPK pathways. Reproductive
Sciences. 17, 1043-1051.
Yeganegi, M., Leung, C. G., Martins, A., Kim, S. O., Reid, G., Challis, J. R., & Bocking, A.
D. (2011). Lactobacillus rhamnosus GR-1 stimulates colony-stimulating factor 3 (granulocyte)
(CSF3) output in placental trophoblast cells in a fetal sex-dependent manner. Biology of
Reproduction. 84, 18-25.
Yeganegi, M., Watson, C. S., Martins, A., Kim, S. O., Reid, G., Challis, J. R., & Bocking, A.
D. (2009). Effect of Lactobacillus rhamnosus GR-1 supernatant and fetal sex on
lipopolysaccharide-induced cytokine and prostaglandin-regulating enzymes in human placental
trophoblast cells: Implications for treatment of bacterial vaginosis and prevention of preterm
labor. American Journal of Obstetrics and Gynecology. 200, 532.e1-532.e8.

146
Yoon, B. H., Park, C. W., & Chaiworapongsa, T. (2003). Intrauterine infection and the
development of cerebral palsy. BJOG. 110, 124-127.
Yoshida M., Sagawa N., Itoh H., Yura S., Takemura M., Wada Y., Sato T., Ito A., Fujii S.
(2002). Prostaglandin F2α, cytokines and cyclic mechanical stretch augment matrix
metalloproteinase-1 secretion from cultured human uterine cervical fibroblast cells. Molecular
Human Reproduction. 8, 681-687.
Zaga, V., Estrada-Gutierrez, G., Beltran-Montoya, J., Maida-Claros, R., Lopez-Vancell, R.,
& Vadillo-Ortega, F. (2004). Secretions of interleukin-1beta and tumor necrosis factor alpha by
whole fetal membranes depend on initial interactions of amnion or choriodecidua with
lipopolysaccharides or group B streptococci. Biology of Reproduction. 71, 1296-1302.
Zaga-Clavellina, V., Garcia-Lopez, G., Flores-Herrera, H., Espejel-Nunez, A., Flores-
Pliego, A., Soriano-Becerril, D., Maida-Claros, R., Merchant-Larios, H., & Vadillo-Ortega,
F. (2007). In vitro secretion profiles of interleukin (IL)-1beta, IL-6, IL-8, IL-10, and TNF alpha
after selective infection with Escherichia coli in human fetal membranes. Reproductive Biology
and Endocrinology. 5, 46-53.
Zakar, T., Hirst, J. J., Mijovic, J. E., & Olson, D. M. (1995). Glucocorticoids stimulate the
expression of prostaglandin endoperoxide H synthase-2 in amnion cells. Endocrinology. 136,
1610-1619.
Zakar, T., Teixeira, F. J., Hirst, J. J., Guo, F., MacLeod, E. A., & Olson, D. M. (1994).
Regulation of prostaglandin endoperoxide H synthase by glucocorticoids and activators of
protein kinase C in the human amnion. Journal of Reproduction and Fertility. 100, 43-50.





























