Embed Size (px)
Citation preview

The Effects of Developmental Nicotine Exposureon Hypoglossal Motoneuron Primary Dendrite
and Soma Development in the Neonatal Rat
Item Type text; Electronic Thesis
Authors Gaddy, Joshua L.
Publisher The University of Arizona.
Rights Copyright © is held by the author. Digital access to this materialis made possible by the University Libraries, University of Arizona.Further transmission, reproduction or presentation (such aspublic display or performance) of protected items is prohibitedexcept with permission of the author.
Download date 07/05/2018 23:15:47
Link to Item http://hdl.handle.net/10150/621005

1
THE EFFECTS OF DEVELOPMENTAL NICOTINE EXPOSURE ON HYPOGLOSSAL
MOTONEURON PRIMARY DENDRITE AND SOMA DEVELOPMENT IN THE
NEONATAL RAT
By
JOSHUA GADDY
_________________________
A Thesis Submitted to the Faculty of the
DEPARTMENT OF CELLULAR AND MOLECULAR MEDICINE
In Partial Fulfillment of the Requirements
For the Degree of
MASTER OF SCIENCE
In the Graduate College
THE UNIVERSITY OF ARIZONA
2016

2
STATEMENT BY AUTHOR
The thesis titled THE EFFECTS OF DEVELOPMENTAL NICOTINE EXPOSURE ON
HYPOGLOSSAL MOTONEURON PRIMARY DENDRITE AND SOMA DEVELOPMENT
IN THE NEONATAL RAT prepared by JOSHUA GADDY has been submitted in partial
fulfillment of requirements for a master’s degree at the University of Arizona and is deposited in
the University Library to be made available to borrowers under rules of the Library. Brief
quotations from this thesis are allowable without special permission, provided that an accurate
acknowledgement of the source is made. Requests for permission for extended quotation from or
reproduction of this manuscript in whole or in part may be granted by the head of the major
department or the Dean of the Graduate College when in his or her judgment the proposed use of
the material is in the interests of scholarship. In all other instances, however, permission must be
obtained from the author.
SIGNED:_______ _____ _____________
APPROVAL BY THESIS DIRECTOR
This thesis has been approved on the date shown below:
______________________________ __________________
Ralph Fregosi Date
Professor of Physiology

3
ACKNOWLEDGEMENT
To start I would like to thank Dr. Ralph Fregosi and Dr. Richard Levine, for their
patience and guidance throughout my graduate education. It was an honor to work with them on
my thesis project. Additionally, I would like to thank my committee members Dr. Paul St. John
and Dr. Helen Amerongen for supporting my education. I would like to acknowledge, Drs.
Marina Cholanian and Andrew Hill, Lila Wollman for helpful critics and feedback on my
practice presentations. I would like to thank Sowmiya Murali and Drs. Gregory Powell and Fei
Xu for allowing me to contribute to their work and for their help. Thank you to Seres Cross for
outstanding technical assistance. I would like to acknowledge Dr. Naomi Rance’s lab for their
help and equipment in analyzing our motoneuron tracings. I would like to thank the Cellular and
Molecular Medicine graduate program, including faculty, staff, and students.
I would like to thank Dr. Maria Velez for funding and always being there when I or any
student needed help. I would also like to thank Donna Treloar for her help and hard work. To my
family, thank you for your support and positive encouragement throughout my education.
Lastly I would like to thank my wonderful wife, Valerisa. You have always supported
me, helped me study and encouraged me to go beyond what I thought I was capable of. You
always believed in me when I doubted myself. You are the hardest working, kindest and the most
altruistic person I know and I am beyond grateful to have you in my life.

4
DEDICATION
I would like to dedicate this thesis to my wife, who provided me with her love and
extraordinary support throughout my educational endeavors.

5
TABLE OF CONTENTS
LIST OF FIGURES ......................................................................................................... 6
LIST OF TABLES ........................................................................................................... 7
ABSTRACT ...................................................................................................................... 8
INTRODUCTION ............................................................................................................ 9
MATERIALS AND METHODS .................................................................................... 13
RESULTS .......................................................................................................................... 20
DISCUSSION ................................................................................................................... 25
CONCLUSION ................................................................................................................ 30
REFERENCES ................................................................................................................. 32

6
LIST OF FIGURES
MATERIALS AND METHODS:
FIGURE 1: FILLED AND TRACED CONTROL AND DNE HYPOGLOSSAL
MOTONEURONS .............................................................................................................. 15
FIGURE 2: BRAINSTEM SLICES CONTAINING THE HYPOGLOSSAL
NUCLEUS (RED ARROWHEAD) AND THE FOURTH VENTRICLE (WHITE
ARROWHEAD) ................................................................................................................. 16
RESULTS:
FIGURE 3: COMPARING TREATMENT AND LOCATION EFFECTS ON
MAXIMUM AND MINIMUM SOMA DIMETER AND THE NUMBER OF PRIMARY
NODES ............................................................................................................................... 24
DISCUSSION:
FIGURE 4: ILLUSTRATION OF XIIMN ORGANIZATION WITHIN THE NUCLEUS
AND THE INNERVATION OF TONGUE MUSCLES.................................................... 28

7
LIST OF TABLES
RESULTS:
TABLE 1: THE EFFECTS OF DNE ON XIIMN MORPHOLOGY PROPERTIES ........ 22
TABLE 2: THE EFFECTS OF DNE ON XIIMN MORPHOLOGY PROPERTIES IN
CAUDAL AND ROSTRAL LOCATIONS ....................................................................... 23

8
Abstract
Nicotine from smoking or from other products containing nicotine has adverse effects on
the fetus during pregnancy, such as respiratory problems. Our laboratory has previously shown
that exposure to nicotine during development (DNE) alters hypoglossal motor neuron (XII MN)
function, including decreased excitatory synaptic input, desensitized nicotinic acetylcholine
receptors, increased input resistance, and differences in the precision and reliability of spike
timing in XIIMNs. Evidence of DNE effects on XIIMN function prompted us to test the
hypothesis that DNE will affect the development of primary dendrites and the soma. Brainstem
slices were collected from neonates and motoneurons were filled with neurobiotin via whole-cell
patch clamp. Filled cells were visualized with heavy metal intensified-3,3'-Diaminobenzidine
(DAB) reaction. DAB-stained cells were analyzed using Neurolucida hardware and software. On
average, the maximum soma diameter of more rostral XIIMNs was larger than that in more
caudal cells. Also, caudal XIIMNs had more primary nodes than rostral XIIMNs, and there was
a significant treatment effect on minimum soma diameter (Control, 13.76 ± 0.71 µm; DNE,
18.09 ± 1.22 µm). The results from this study uncovered potential effects of nicotine on XIIMNs
found in rostral and caudal regions of the hypoglossal nucleus.

9
Introduction
Worldwide, approximately 5 million deaths occur per year due to direct use of tobacco,
with smoking being the most common method (WHO, 2015), with approximately 480,000
deaths per year in the United States (U.S. Department of Health and Human Services, 2014). The
number of women who smoke while they are pregnant and/or immediately after child birth is a
cause for major public health concern. According to Tong et al., (2013) 23% of women smoked
before pregnancy, 54% quit during pregnancy, 16% began smoking again after delivery, and
11% of women smoked throughout their pregnancy in 2010. Looking at these percentages it is up
to clinicians to provide information on smoking cessation practices. According to one study one-
fifth of pregnant women received information about smoking cessation methods from clinicians,
while one-fourth of pregnant women did not (Kapaya, et al., 2015). These numbers are a cause
for concern not only for the mother, but also for the unborn child.
Out of the thousands of chemicals found in a cigarette, nicotine is among the most
clinically relevant. Pauly and Slotkin (2008) found the effects of nicotine exposure and cigarette
smoke exposure in utero had the same effect on neurogenesis in hippocampal neurons which
exemplifies the effects of nicotine alone. The focus on nicotine is because nicotine replacement
therapy (NRT), such as nicotine patches and nicotine gum, is prescribed to pregnant smokers;
also electronic nicotine delivery systems (ENDS), such as e-cigarettes, are readily available in
the market (Kapaya, et al., 2015; Berlin, et al., 2014; Kralikova, et al., 2013). One of the most
devastating attributes of nicotine is that it can cross the placental barrier in pregnant women and
can cross the fetal blood brain barrier (Benowitz et al., 2009; Slotkin, 1998). Within the fetus
nicotine can bind to acetylcholine (ACh) receptors, which are expressed early in development,
but normally bind endogenous ACh rather than nicotine (Gotti et al., 2006; Zoli et al., 1995).

10
Acetylcholine receptors (AChR) are crucial in regulating fetal brain development, and
overstimulation by nicotine can lead to critical periods in development being disrupted
(Muhammad et al., 2012; Dwyer et al., 2009). Increased risk for sudden infant death syndrome
(SIDS), neurocognitive defects, breathing problems, underweight births and an increased risk of
stillbirth have been attributed to nicotine exposure in utero (Aoyama et al., 2015; England et al.,
2015; U.S. Department of Health and Human Services, 2014; Lavezzi et al, 2010; Button et al.,
2007; Raymond et al., 2005; Alm et al., 1998). Animal models have shown that developmental
nicotine exposure (DNE) is associated with cell damage, including reduced cell number and size,
impaired synaptic activity, and a premature change from cell replication to differentiation
(Aoyama et al., 2015; England, et al., 2015; Pilarski et al., 2011; Dwyer et al., 2009, 2008).
Among the most devastating effects of smoking are those related to respiratory function.
Many neuronal structures in the brainstem contribute to the normal respiration. For example the
preBӧtzinger Complex (preBӧtC) generates the inspiratory rhythm during breathing and
influences the respiratory final motor output to the muscles of breathing, including the tongue
muscles, which are innervated by hypoglossal motoneurons (XIIMNs). The tongue muscles
participate in essential motor functions such as breathing, mastication, swallowing, and
vocalization (Fregosi and Ludlow, 2014; Fregosi, 2011; Bailey and Fregosi, 2004; Berger et al.
1996; Lowe et al., 1980). Previous studies have shown DNE effects on brainstem neurons that
control respiration, including desensitization of nicotinic AChRs on XIIMNs (Pilarski et al.,
2012), reduced excitatory synaptic input and increased excitability (Jaiswal et al., 2013; Pilarski
et al., 2011), increased expression of glutamatergic receptor subunits in preBӧtC and XIIMNs
(Jaiswal et al., 2013), and altered the precision and reliability of spike timing in XIIMNs (Powell
et al., 2015). Continuous bombardment of nicotine and ACh on AChRs at the postsynaptic,

11
presynaptic and axon terminal locations during development can lead to motoneurons being
hyperexcitable (Paterson and Norberg, 2000). This hyperexcitability may be affecting the way
these motoneurons develop, and how they receive and transmit information (Pilarski et al.,
2011).
The current study stemmed from a previous study that demonstrated nicotine exposure
stunted the dendritic growth of higher branch orders in DNE XIIMNs compared to control
XIIMN dendrites (Powell, et al., 2016). Other DNE neuronal morphology studies demonstrated
effects on potentially important features of the XIIMNs, such as the branching points (node), the
base diameter of dendrites, and where these XIIMNs are located in the nucleus (Jaiswal et al.,
2013; Muhammad et al., 2012; Pilarski et al., 2011; Núñez-Abades, 1995; Roy and Sabherwal,
1994). Obtaining a complete image of XIIMN morphology may shed light on how nicotine may
affect the development of the motoneurons and dendrites in the hypoglossal nucleus of neonatal
rats. In this study we focused on how DNE affects primary dendrite and soma development, but
also how DNE affects XIIMN development in the rostral and caudal ends of the hypoglossal
nucleus. There are many studies looking at how nicotine affects the overall XIIMN population,
but no studies have investigated the effects of nicotine on XIIMNs located in different regions of
the hypoglossal nucleus, such as rostral and caudal ends. Expanding the knowledge of DNE
effects on XIIMNs in various regions of the hypoglossal nucleus is crucial because the different
areas of the nucleus innervate specific muscles of the tongue, for example the ventrolateral
subnucleus in the caudal end of the nucleus innervates the geniohyoid muscle (Slotkin, 1993).
Dendrites are important cellular processes that help the neuron receive and integrate the
signals from other neurons. Dendrites are essential features of neurons due to the fact that the
majority of a neuron’s surface area exists in the dendrites (Sholl, 1955, 1953). A failure in

12
normal dendritic development may compromise the normal synaptic connectivity and the overall
function of these motoneurons (Powell et al., 2016; Powell et al., 2015; Pilarski et al., 2011;
Jaiswal et al., 2013; Puram et al., 2011). Multiple studies have demonstrated effects of DNE on
the dendritic morphology in various brain regions, including the hypoglossal nucleus (Powell, et
al., 2016; Powell et al., 2015; Jalili et al., 2014; Jaiswal et al., 2013; Mychasiuk et al., 2013;
Muhammad et al., 2012; Pilarski et al., 2011; Roy and Sabherwal, 1994). Investigating the
effects of teratogens, such as nicotine, on XIIMN development can potentially help understand
respiratory diseases associated with smoking, for example sleep apnea (Kahn et al., 1994). The
influence of DNE on soma size, morphology, and other cellular features in other regions of the
brain prompts us to hypothesize that DNE will have a negative effect on XIIMN primary
dendrite and soma development, and that the influences of DNE in rostral and caudal XIIMNs
will differ.
Our hypothesis was tested using a dark stain to obtain detailed morphologic
characteristics of XIIMNs. XIIMNs used in the study came from neonatal rats with an age range
of postnatal day one (P1) to postnatal day four (P4). In an in vivo study, Huang et al. (2010,
2004) demonstrated that perinatal nicotine exposure affected breathing in the days just after
birth. XIIMN soma area and primary dendrite morphology (number of primary dendrites,
dendritic length and surface area) was previously investigated by Powell at al. (2016). For this
study we analyzed these features, but used different analytical methods to analyze the XIIMN
morphology. We measured (1) the number of primary nodes, (2) maximum and minimum soma
diameter, (3) dendritic base diameter, and (4) average branch diameter. Our results found no
treatment and developmental effects on the basic features of XIIMN primary dendrite and soma
maximum and minimum diameter. Treatment and hypoglossal nucleus location had no effect on

13
primary dendrite features, except on the number of branch points on dendrites originating
directly from the soma (primary dendrite) and on maximum soma diameter. The minimum soma
diameter in DNE caudal XIIMNs was larger on average than control caudal XIIMN (DNE, 18.09
± 1.22 µm; Control, 13.76 ± 0.71 µm).
Materials and Methods
Animal Usage and Treatments
All animal care/use and procedures were approved by the Institutional Animal Care and
Use Committee at the University of Arizona. Data was collected from 71 neonatal Sprague-
Dawley rats, ranging in age from postnatal day (P) 1 to P4. The 71 motoneurons that were traced
and measured were separated into two age groups (P1+P2 and P3+P4) and two treatment groups
(control and DNE), as follows: 20 control/P1+P2 individuals; 23 DNE/P1+P2 individuals; and
14 P3+P4/control individuals; 14 P3+P4/DNE individuals. In another analysis we separated
hypoglossal motoneurons (XIIMNs) into rostral versus caudal positions (33 rostral XIIMNs and
34 caudal XIIMNs). Rostral-caudal separation was accomplished by confirming the transition of
the spinal central canal into the 4th
ventricle using images and tracings of the XIIMNs and the
Atlas of the Newborn Rat Medulla oblongata for reference (Figure 2; Ruangkittisakul, et al.).
Pregnant Sprague-Dawley rats were implanted with an osmotic mini-pump (Alzet,
Cupertino, CA) subcutaneously between the shoulder blades on gestational day 5 (Jaiswal et al.
2013; Pilarski et al. 2011; Huang et al. 2004, 2010; Pilarski and Fregosi 2009; Luo et al. 2004,
2007). Rat mothers and pups were exposed to nicotine bitartrate (6 mg/kg/day) or physiologic
saline for 28 days after implantation. After birth the neonatal rats continued to receive nicotine
through the mother’s milk. Treated animals included an unexposed treatment group that did not

14
undergo surgery or pump implantation. There were no electrophysiological differences between
the unexposed and saline-exposed animals, the data collected from both of these treatment
groups were combined into a single control group, and will be referred to as such throughout the
thesis.

15
DNE
A. B.
Control
C. D.
Figure 1. Filled and traced control and DNE hypoglossal motoneurons. (A) An example of a DNE
motoneuron that was filled with neurobiotin (magnification 40x), underwent DAB staining, and (B)
traced with the Neurolucida tracing system. (C) A filled control and (D) reconstructed XIIMN. Scale bars
represent 25 µm.
40x
40x

16
A.
B.
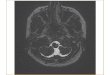
Figure 2. Brainstem slices containing the
hypoglossal nucleus (red arrowhead) and
the fourth ventricle (white arrowhead). These brainstem slices were used as a
reference to divide the XIIMNs into rostral
versus caudal, where: (A) caudal region of
the hypoglossal nucleus, (B) central canal is
opening into the fourth ventricle, marking
the obex, which we used to demarcate the
transition from rostral to caudal regions of
the hypoglossal nucleus, (C) shows the fully
opened fourth ventricle (Ruangkittisakul et
al.). All XIIMNs located below the obex,
are considered caudal, while those above the
obex are defined as rostral. The other
arrowheads label various structures in the
medulla, for example the dark green
arrowhead labels the medial inferior olive,
gray arrowhead labels the dorsal motor
nucleus of the vagus nerve, dark blue
arrowhead labels the dorsal inferior olive,
light blue arrowhead labels principal inferior
olive.
C.

17
Brainstem Slice Preparation and Electrophysiology
Rat pups of either sex were randomly collected from their litters and weighed. The rat
pups were anesthetized with ice until they were unresponsive to paw pinch, and quickly
decerebrated at the coronal suture. Evisceration ensued to expose the surrounding vertebral
column and ribcage and transferred to a dish containing chilled (4°C–8°C) and oxygenated
(95%O2/5%CO2) artificial cerebrospinal fluid (aCSF) solution (in mM: 120 NaCl, 26 NaHCO3,
30 glucose, 1 MgSO4, 3 KCl, 1.25 NaH2PO4, 1.2 CaCl2, pH 7. 4). The medulla and upper
spinal cord were extracted and pinned to a cutting block, with the rostral surface up, for serial
microsection slicing using a Vibratome™ (VT1000, Leica), as described previously (Jaiswal et
al. 2013). Transverse medullary slices were taken until the rostral inferior olive and the most
rostral hypoglossal nerve rootlet were near the surface. 400 µm slices were collected to capture
the majority of the hypoglossal motor nucleus. Preparations were then transferred to a recording
chamber, which was continuously perfused with oxygenated and modified room temperature
aCSF (in mM: 120 NaCl, 26 NaHCO3, 30 glucose, 1 MgSO4, 9 KCl, 1.25 NaH2PO4, 1.2
CaCl2, pH 7. 4).
Staining Protocol
Electrophysiological recordings were conducted on XIIMNs from the brain stem slices
that were collected, as described previously (Pilarski et al., 2011). After electrophysiological
experiments, hypoglossal motoneurons (XIIMNs) were slowly filled with the tracer Neurobiotin
(Vector Labs, Burlingame, CA) to acquire detailed morphology. Visualization of filled XIIMNs
was obtained using a protocol developed by McMullen and deVenecia (1993). Briefly,
Neurobiotin-filled cells were incubated with a primary goat anti-biotin antibody (1:10,000,
Sigma), a secondary biotinylated rabbit antigoat IgG (1:250; Vector Labs, Burlingame, CA), the

18
Elite ABC reagent (Avidin-Biotin Complex; Vector Labs, Burlingame, CA), and heavy metal
intensified 3,3’-diaminobenzidine (DAB; Sigma). This staining protocol produced a dark brown
reaction product and clearly stained the cell body and all its processes. Stained medullary slices
were mounted on brass rings with dimethyl sulfoxide solution (DMSO, Sigma-Aldrich, St.
Louis, MO).
Neuron Tracing
A detailed method of XIIMN tracing was previously described in Powell et al. (2016). A
light microscope (Nikon) and Neurolucida® software/equipment (MBF Bioscience, Williston,
VT) were used to capture and measure stained motoneurons. Briefly, Neurolucida was used to
trace the medullary slice and the edges of the fourth ventricle at 5x magnification. Stained cells
were located and magnified to 63x magnification with an oil immersion lens (Zeiss). The soma
(cell body) was labeled and its edges were traced. Dendrites and their branches were traced to
completion and labelled as “normal” or “incomplete”. Normal endings were those dendritic
branches that had a clear ending, whereas incomplete endings were those dendritic branches that
disappeared into the tissue or were cut at the surface of the tissue. Dendrite tracing was repeated
for all dendritic trees that sprouted off the soma.
Complete dendrites (dendrites labeled as “normal” ending during the Neurolucida
tracing) were selected for analysis to avoid errors that would result from including cut or faded
processes in the quantification. Some spines were traced and labeled, but not all XIIMNs had
spines. Dendritic length, surface area, volume, number of primary dendrites, number of branch
points (nodes) and soma size (maximum and minimum diameter) were the measurements
selected for further analysis (Table 1-3).

19
Neurolucida Explorer®
Branched Structure Analysis
Traced XIIMNs were analyzed using the Neurolucida Explorer (MBF Bisoscience,
Williston, VT), a companion software to the Neurolucida tracing software. Cells underwent an
analysis that quantified basic cellular structures, such as number of branches, branch endings,
branch order, branch length, branch surface area, soma maximum/minimum diameter, and other
properties. Examples of filled and reconstructed XIIMNs for each treatment condition can be
seen in Figure 1.
Statistical Analysis
Cellular properties that were measured in the Branched Structure Analysis included the
number of primary dendrites, dendritic total length and surface area, soma maximum/minimum
diameter, mean base diameter, mean average branch diameter and the number of branching
points. Neurolucida measures the maximum and minimum soma diameter as two independent
measurements, and the two diameter measures do not have to be at 90o angles to one another.
Any unbranched primary dendrites with a total length less than 10 µm were not used in this
study. Excluding short unbranched primary dendrites was implemented to control for potential
spines on the cell body. Spines originating from the cell body were excluded because the
majority of XIIMNs traced did not possess spine-like features.
Prism 6.05 (Graphpad, San Diego, CA, USA) was used to conduct a 2-way ANOVA with
age (P1+P2 and P3+P4) and treatment (Control and DNE) as the two main factors, and the
dependent variable being each of the cellular properties measured. A univariate 3-way ANOVA
was conducted using SPSS version 23 (IBM, Corp., Armonk, NY, USA) to further investigate
the interactions between all three main factors (age x treatment x position) on XIIMN maximum

20
and minimum soma diameter, and the number of nodes. After each significant ANOVA test, post
hoc analyses were done with the Tukey’s multiple comparison test to compare mean values , for
example DNE:P1+P2 and control:P1+P2 or DNE:P3+P4.
The data included in this report were derived from staining and analysis that I performed
on 68 motoneurons that were recorded in separate experiments by Drs. Fei Xu and Gregory
Powell.
Results
Branched Structure Analysis of XIIMNs
The number of primary dendrites, complete primary dendritic length and surface area,
average base diameter, mean average branch diameter, and soma maximum/minimum diameter
were quantified in Branched Structure analysis using Neurolucida. The standard error of the
means and P-values were analyzed with a 2-way ANOVA, where age and treatment or XIIMN
location and treatment were the two factors and each morphological property was the dependent
variable. There were no significant treatment or age effects on XIIMN morphology (Table 1; P >
0.05). Interestingly, there were significant treatment and location effects on the number of
primary nodes (P = 0.04), maximum and minimum soma diameter (P = 0.04 and P = 0.02
respectively; Table 2). Tukey’s multiple comparisons test found smaller soma minimum
diameter in Control:Caudal (Mean = 13.76 ± 0.71 µm) compared to Nicotine:Caudal (Mean =
18.09 ± 1.22 µm). Tukey’s multiple comparisons test could not find exactly where the
significant differences occurred in the number of primary nodes and maximum soma diameter,
which indicates that the data are too variable, or the differences are too small to provide
significant statistical power. Cellular features with significant differences are graphed in Figure
3.

21
Maximum and minimum soma diameter and number of nodes in primary dendrites were
further investigated using a 3-way ANOVA, where age, treatment, and location were the 3
factors. The 3-way ANOVA found no significant treatment, age, or location effects on maximum
or minimum soma diameter. The location factor had significant effects on the number of nodes
(P = 0.02), but Tukey’s multiple comparisons test could not find exactly where the differences
occurred because the interaction terms were not statistically significant.

22
P1+P2 P3+P4 P-
Value
N
(Control/DNE) Control DNE Control DNE
Number of
Primary
dendrites
per neuron
4.1±0.37 4.8±0.43 3.92±0.37 4.0±0.33 P >
0.05 33/34
Complete
primary
total
dendritic
length (µm)
213.14±31.5
4
265.86±44.4
9 203±50.71
171.71±29.9
1
P >
0.05 33/34
Complete
primary
total
dendritic
surface
area (µm2)
732.14±112.
96
1273.32±232
.48
832.65±170.
57
749.91±156.
94
P >
0.05 33/34
Average
Primary
Base
Diameter
(µm)
1.86±0.16 1.77±0.18 2.36±0.36 2.80±0.96 P >
0.05 33/34
Mean
Primary
Average
Branch
Diameter
(µm)
1.56±0.13 1.66±0.13 2.08±0.27 2.32±0.68 P >
0.05 33/34
Total
number of
Primary
Nodes
2.90±0.25 3.10±0.36 2.61±0.37 2.79±0.38 P >
0.05 33/34
Soma
maximum
diameter
(µm)
24.16±1.17 29.40±1.97 27.43±2.60 27.09±1.88 P >
0.05 33/34
Soma
minimum
diameter
(µm)
14.08±0.74 17.24±0.93 14.68±0.93 15.89±1.75 P >
0.05 33/34
Table 1. The effects of DNE on XIIMN morphology properties. Developmental nicotine exposure
(DNE) had no treatment or age effect on the morphological properties. Values represent the mean ± the
standard error of the mean (SEM). P-values were obtained using a 2-way ANOVA, with P ≤ 0.05
considered statistically significant. Units, μm = micron and μm2 = microns squared.

23
Rostral Caudal
P-Value N
(Rostral/Caudal) Control DNE Control DNE
Number of
Primary
dendrites
per neuron
4.31±0.44 4.71±0.47 3.82±0.30 4.24±0.35 P > 0.05 33/34
Complete
primary
total
dendritic
length (µm)
253.71±44.8
2
235.69±40.8
4
167.37±29.4
7
218.49±44.2
2 P > 0.05 33/34
Complete
primary
total
dendritic
surface
area (µm2)
889.01±166.
18
1035.09±22
5.52
661.37±94.1
7
1080.52±22
2.17 P > 0.05 33/34
Average
Primary
Base
Diameter
(µm)
2.05±0.26 1.54±0.20 2.07±0.24 2.85±0.78 P > 0.05 33/34
Mean
Primary
Average
Branch
Diameter
(µm)
1.74±0.19 1.45±0.15 1.81±0.20 2.42±0.54 P > 0.05 33/34
Total
number of
Primary
Nodes
2.69±0.31 2.41±0.37 2.88±0.28 3.53±0.32 P = 0.04 33/34
Soma
maximum
diameter
(µm)
26.69±2.19 30.86±0.2.0
3 24.24±1.26 25.96±0.78 P = 0.04 33/34
Soma
minimum
diameter
(µm)
14.91±0.91 15.4±1.22 13.77±0.71 18.09±1.22 P = 0.02 33/34
Table 2. The effects of DNE on XIIMN morphology properties in caudal and rostral locations.
Developmental nicotine exposure (DNE) had no treatment or location effect on the morphological
properties, except on the number of nodes, max and min soma diameter. Values represent the mean ± the
standard error of the mean (SEM). P-values were obtained using a 2-way ANOVA, with P ≤ 0.05
considered statistically significant. Units, μm = micron and μm2 = microns squared.

24
A.
B.
.
B.
C.
.
B.
Figure 3. Comparing
treatment and location
effects on maximum and
minimum soma dimeter and
the number of primary
nodes. A.) 2-way ANOVA
found locational effects on
maximum soma diameter; on
average rostral max soma
diameters were larger than
caudal max soma diameters,
but no DNE effect. B.) 2-way
ANOVA found significant
treatment effect on minimum
soma diameter; on average the
soma minimum diameter in
Control:Caudal (Mean = 13.76
µm) was smaller compared to
Nicotine:Caudal (Mean =
18.09 µm). C.) 2-way
ANOVA found location
effects on number of primary
nodes; on average caudal
XIIMNs had more primary
nodes than rostral primary
nodes, but no statistical
significant DNE effect.

25
Discussion
Tracing programs like Neurolucida collect and provide a wave of information about
complex morphologic structures like those found on neurons. Morphology analysis can consist
of counting the total number of dendrites, number of branching points, measuring total dendritic
length, conducting Sholl analysis, or convex hull analysis (Kawa, et al., 1998; Sholl, 1953).
These morphometrics alone cannot provide a full understanding of the dendritic field, but
collectively can provide ample information about dendritic arborization (Langhammer et al.,
2010). The data collected and presented here show that overall treatment and age had no effect
on the total number of XIIMN primary dendrites, primary dendritic length, primary dendritic
surface area, average base diameter, mean average branch diameter, and soma maximum and
minimum diameter. Treatment and location affected XIIMN maximum and minimum soma
diameter and the number of primary nodes. Statistical analyses found that maximum soma
diameter and number of nodes varied with rostral and caudal locations (Figure 3). Treatment
affected caudal XIIMN minimum soma diameter, where DNE caudal XIIMNs had larger
minimum soma diameters compared to control minimum soma diameter (Figure 3).
This study was conducted to test the hypothesis that DNE would affect primary dendrite
and soma morphometric development in XIIMNs and have an effect on motoneurons located in
rostral and caudal ends of the hypoglossal nucleus. Prenatal nicotine exposure studies have found
disturbances to neuronal growth and proliferation, axonogenesis and synaptogenesis, apoptosis
and migration, which are all important events in nervous system development (Slotkin, 2008).
The cell body has important functions, such as protein synthesis, and if DNE affected the size in
anyway, this may alter the overall functionality of the neuron. Results from previous studies
found dendritic branching and soma size development were affected by DNE (Mychasiuk et al.,

26
2013; Muhammad et al., 2012; Roy and Sabherwal, 1994). The Sholl analysis used in the Powell
et al. (2016) study found changes in dendritic arborization patterns as a function of distance from
the soma. For example, DNE hypoglossal motoneuron dendrite length and surface area
decreased, but control hypoglossal motoneuron dendrite length and surface area increased as the
animal aged. The Powell et al. (2016) study also looked at primary dendritic length and surface
area, and soma area, but did not examine the effects of location of the cell in the nucleus or the
density of somatic spines.
Investigating where these XIIMNs are located in the nucleus is predicated on the fact that
XIIMNs found in specific regions of the nucleus innervate specific muscles of the tongue
(Kanjhan, et al., 2015; Sokoloff et al., 1993). Significant DNE effects on neuron morphology on
multiple regions of the central nervous system, including the hypoglossal nucleus, and on
XIIMN electrophysiology warranted an in-depth investigation into XIIMN primary dendrite and
soma development (Jalili et al., 2014; Jaiswal et al., 2013; Pilarski et al., 2011; Núñez-Abades,
1995; Roy and Sabherwal, 1994). The primary dendrite features investigated here showed no
significant effects of DNE or age on XIIMN development (Table 1). However, these results do
not counter the results found in Powell et al. (2016), where DNE affected soma size when
comparing treatment and age factors. Utilizing different analytical methods, such as (identifying
potential soma spines), looking at other potentially important dendritic features (base diameter,
average branch diameter, number of nodes), and the multiple ways to measure the soma
(maximum and minimum diameter versus soma area) adds to what was found in Powell et al
(2016). Soma spines were not included in this study due to the majority of the XIIMNs traced not
having spines on the soma, though spines due exist on soma surfaces (Nimchinsky et al., 2002).
Other measured features of the XIIMNs, such as base diameter and average branch diameter,

27
described dendrite morphology. If any changes were detected in base diameter and average
branch diameter it could imply potential effects on the dendrite membrane conductance and
synapses (Powell et al., 2016). Effects on branch points can indicate how complex a neuron is
because branch points (nodes) lead to higher branch orders, which potentially affects the entire
dendritic field.
We analyzed XIIMN cellular structures based on hypoglossal nucleus regions and found
that soma max diameter and the number of primary branch points (nodes) differed in rostral and
caudal regions (Table 2). The hypoglossal nucleus is a complex structure that consists of
subdivisions dedicated to controlling specific muscles in the tongue. The hypoglossal nucleus
can be divided into three subdivisions; the ventral subdivision is found in the caudal 2/3, the
dorsal subdivision in the rostral 2/3 of the nucleus, with overlap in the middle of the nuclear
structure (Sokoloff, 1993; Krammer, et al., 1979). Motoneurons in the dorsal subdivision
innervate the tongue retrusor muscles (hyoglossus and styloglossus); the ventral subdivision
innervates the tongue protrusor muscle (genioglossus); and the ventrolateral subdivision
innervates the suprahyoid muscle and geniohyoid (Figure 4; Kanjhan, et al., 2015; Sokoloff,
1993). Krammer et al (1979) found that XIIMNs innervating the suprahyoid and geniohyoid
muscles were located in the most caudal side of the hypoglossal nucleus, while XIIMNs
innervating the genioglossus are located throughout the length of the hypoglossal nucleus (Figure
4; Krammer et al., 1979).

28
Figure 4. Illustration of XIIMN organization within the nucleus and the innervation of tongue
muscles. (Left) Transverse sections illustrate the differences between subnucleus regions as you move
from the caudal (level 2) to the rostral (level 6) ends of the hypoglossal nucleus. (Right) Diagram shows
what regions of the nucleus innervate which muscles of the tongue. Right diagram also shows what
proportion of the nucleus contributes to what tongue muscle, for example the dorsal subnucleus can be

29
found in the 2/3 rostral end and innervates the styloglossus and hyoglossus muscles. Diagram from
Krammer et al., 1979.
The effects of DNE on XIIMN electrical properties, such as reduced excitatory synaptic
input and increased excitability (Jaiswal et al., 2013; Pilarski et al., 2011), and differences in the
precision and reliability of spike timing in XIIMNs (Powell et al., 2015) may be attributed to the
changes in soma morphology (Jaiswal et al., 2013; Pilarski et al., 2011). Maximum soma
diameter varied with nuclear location, and the minimum soma diameter of XIIMNs in the caudal
region of the hypoglossal nucleus showed DNE effects (Table 2). Our results and previous
evidence show that rostral and caudal XIIMNs show differences in morphology and function
(Kanjhan, et al., 2015; Krammer et al., 1979). In early nervous system development the AChRs
are among the first types of receptors expressed and are expressed first in the spinal cord, and
then continue in the rostral direction (Zoli et al., 1995). The way AChRs develop in the nervous
system (in a spinal cord to rostral direction) can be affected by nicotine exposure. Potentially,
XIIMNs in the caudal regions may be affected by nicotine or exposed to nicotine earlier than the
XIIMNs rostral regions within the hypoglossal nucleus. This mean that the caudal XIIMNs
innervating the suprahyoid and geniohyoid muscles of the tongue, which assist in respiration, are
affected by nicotine exposure during development (Takahashi et al., 2002; Sokoloff, 1993;
Krammer et al., 1979).
Future Methodological Suggestions
Our present study was a step in the right direction for a complete study of the entire XII
motor nucleus, and the effects nicotine may have on XIIMN development and function. Primary
dendrites were the only branch order analyzed because there is high probability of higher branch
orders being truncated during brainstem acquisition. Obviously, XIIMNs have higher branch

30
orders and capturing these branch orders is ideal for complete morphometric analysis of XIIMNs
(Powell et al., 2016; Kanjhan, et al., 2015; Nunez-Abades et al., 1995 and 1994). Obtaining the
majority of neuron tracings can be accomplished by serial sectioning by reducing the number of
higher branch orders being lost, but can still introduce some pieces of dendrites being lost
(Kanjhan, et al., 2015). The maximum and minimum soma diameters were interesting
measurements of the soma, but exactly how the maximum and minimum soma diameters were
calculated in Neurolucida is still unclear. The aspect ratio (how flat the soma can be) and form
factor (a measurement that combines how smooth the perimeter and how compact a cell body is)
could further characterize the XIIMN soma by Neurolucida. In addition to rostral versus caudal
regions the dorsal subdivision, ventral subdivision, and ventrolateral subdivision may be of
interest due to the tongue muscle that they innervate and its contribution to respiratory function
(England et al., 2015; Kanjhan, et al., 2015; Sokoloff, 1993; Krammer et al., 1979). Postnatal day
one to four was used in this study, but investigation of nicotine on days and weeks beyond
postnatal day four, or the influence of stopping nicotine exposure after birth may be interesting to
observe (Carrascal et al., 2005). The addition of these methods in a DNE study may provide
more information on nicotine effects on XIIMNs in all regions of the nucleus, including the long-
term effects on XIIMNs.
Conclusions
In the present study we found the hypoglossal motoneuron features, maximum and
minimum soma diameter and the number of nodes varied based on nuclear locations. The caudal
DNE XIIMNs soma exhibited larger minimum soma diameter compared to caudal control
XIIMNs. Prenatal nicotine exposure studies continue to find nervous system disruptions, for
example respiration in infants is critical for life and infants born to smoking mothers have a

31
higher incidence of sudden infant death syndrome (SIDS) and central and obstructive apneas
(England, et al., 2015). Nicotine replacement therapy continues to be prescribed to pregnant
smokers and exposure to other nicotine sources warrants an investigation into how nicotine
affects the development and function of hypoglossal motoneurons.

32
References
Alm B, Milerad J, Wennergren G, Skjaervan R, Øyen N, Norvenius G, Daltveit AK,
Helweg Larson, K., Markestad, T., Irgens, L. M. (1998) A case-control study of smoking
and sudden infant death syndrome in Scandinavian countries, 1992 to 1995. Arch Dis
Child, 78:329-334
Aoyama Y, Toriumi K, Mouri A, Hattori T, Ueda E, Shimato A, Sakakibara N, Soh Y,
Mamiya T, Nagai T, Kim HC, Hiramatsu M, Nabeshima T, Yamada K. (2015)
Prenatal Nicotine Exposure Impairs the Proliferation of Neuronal Progenitors, Leading to
Fewer Glutamatergic Neurons in the Medial Prefrontal Cortex. [Online] Available from:
http://www.ncbi.nlm.nih.gov/pubmed/26105135
Bailey EF and Fregosi RF. (2004). Coordination of intrinsic and extrinsic tongue muscles during
spontaneous breathing in the rat. J Appl Physiol. 96(2):440-449.
Benowitz NL, Hukkanen J, and Jacob III, P. (2009) Nicotine Chemistry, Metabolism,
Kinetics and Biomarkers. In J. E. Henningfield, E. D. London, and S. Pogun (Eds.),
Nicotine Psychopharmacology, Handbook of Experimental Pharmacology 192 (pp. 29\
60). Berlin Heidelberg, Germany: Springer-Verlag.
Berger A J, Bayliss DA, Viana F. (1996). Development of hypoglossal motoneurons. J Appl
Physiol. 81(3):1039-48.
Berlin I, Grange G, Jacob N, Tanguy ML. (2014). Nicotine patches in pregnant smokers:
Randomised, placebo controlled, multicentre trial of efficacy. Bmj 348:g1622
Button TM, Maughan B, McGuffin P. (2007). The relationship of maternal smoking to
psychological problems in the offspring. Early Hum Dev. 83:727-32.
Dwyer JB, Broide RS, Leslie FM. (2008). Nicotine and brain development. Birth Defects Res C
Embryo Today, 84: pp. 30–44
Dwyer JB, McQuown SC, Leslie FM. (2009) The Dynamic Effects of Nicotine on the
Developing Brain. Pharmacol Ther. 122(2):125-39.
England LJ, Bunnell R E, Pechacek T F, Tong VT, McAfee TA. (2015) Nicotine and the
Developing Human: A Neglected Element in the Electronic Cigarette Debate. Am J Prev
Med. 49:286-93.
Fregosi RF. (2011). Respiratory related control of hypoglossal motoneurons--knowing what we
do not know. Respir Physiol Neurobiol. 179(1):43-7.
Fregosi RF and Ludlow CL. (2014). Activation of upper airway muscles during breathing and
swallowing. J Appl Physiol. 116(3):291-301.

33
Gotti C, Zoli M, Clementi F. (2006) Brain nicotinic acetylcholine receptors: native subtypes
and their relevance. Trends Pharmacol Sci. 27(9):482-91.
Huang YH, Brown, Costy-Bennett S, Luo Z, Fregosi RF. (2004). Influence of prenatal nicotine
exposure on postnatal development of breathing pattern. Respir Physiol Neurobiol 143:1
-8.
Huang Y H, Brown AR, Cross SJ, Cruz J, Rice A, Jaiswal S, Fregosi RF. (2010). Influence of
prenatal nicotine exposure on development of the ventilatory response to hypoxia and
hypercapnia in neonatal rats. J Appl Physiol. 109:149-58.
Jaiswal SJ, Pilarski JQ, Harrison CM, Fregosi RF. (2013). Developmental Nicotine
Exposure Alters AMPA Neurotransmission in the Hypoglossal Motor Nucleus and Pre
Bötzinger Complex of Neonatal Rats. J Neurosci. 33:2616-25.
Jalili C, Salahshoor MR, Khademi F, Jalili P, Roshankhah SH. (2014). Morphometrical
analysis of the effect of nicotine administration on brain's prefrontal region in male rat.
Int. J. Morphol., 32:761-766, 2014.
Kahn A, Groswasser J, Sottiaux M, Kelmanson I, Rebuffat E, Franco P, Dramaix M, Wayenberg
JL. 1994. Prenatal exposure to cigarettes in infants with obstructive sleep apneas.
Pediatrics 93:778–783.
Kanjhan R, Fogarty MJ, Noakes PG, Bellingham MC. (2015). Developmental changes in the
morphology of mouse hypoglossal motor neurons. Brain Struct Funct. [Epub ahead of
print]
Kapaya M, Tong V, Ding H. (2015). Nicotine replacement therapy and other interventions for
pregnant smokers: Pregnancy Risk Assessment Monitoring System, 2009-2010. Prev
Med. 78:92-100.
Kralikova E, Novak J, West O, Kmetova A, Hajek P. 2013. Do e-cigarettes have the potential to
compete with conventional cigarettes?: A survey of conventional cigarette smokers'
experiences with e-cigarettes. Chest 144:1609–1614.
Krammer EB, Rath T, Lischka MF. (1979). Somatotopic organization of the hypoglossal
nucleus: a HRP study in the rat. Brain Res. 170(3):533-7.
Langhammer CG, Previtera ML, Sweet ES, Sran SS, Chen M, Firestein BL. (2010)
Automated Sholl Analysis of Digitized Neuronal Morphology at Multiple Scales: Whole
Cell Sholl Analysis Versus Sholl Analysis of Arbor Subregions. Cytometry A. 77:1160-
8.

34
Lavezzi AM, Corna, M., Mingrone, R., Matturri, L. (2010). Study of the human hypoglossal
nucleus: Normal development and morpho-functional alterations in sudden unexplained
late fetal and infant death. Brain Dev. 32:275-84.
Lowe AA. (1980). The neural regulation of tongue movements. Prog Neurobiol. 15(4):295-
344.
Luo Z, Costy-Bennett S, Fregosi RF. (2004). Prenatal nicotine exposure increases the
strength of GABA(A) receptor-mediated inhibition of respiratory rhythm in neonatal rats.
J Physiol. 561:387-93.
Luo Z, McMullen NT, Costy-Bennett S, Fregosi RF. (2007). Prenatal nicotine exposure alters
glycinergic and GABAergic control of respiratory frequency in the neonatal rat
brainstem-spinal cord preparation. Respir Physiol Neurobiol. 157:226-34.
McMullen NT, de Venecia RK. (1993). Thalamocortical patches in auditory neocortex. Brain
Res. 620:317-22.
Muhammad A, Mychasiuk R, Nakahashi A, Hossain SR, Gibb R, Kolb. B. (2012) Prenatal
nicotine exposure alters neuroanatomical organization of the developing brain. Synapse.
66:950-4.
Nimchinsky EA, Sabatini BL, Svoboda K. (2002). Structure and function of dendritic spines.
Annu. Rev. Physiol. 64:313–53
Núñez-Abades PA, He F, Barrionuevo G, Cameron WE. (1994). Morphology of developing rat
genioglossal motoneurons studied in vitro: changes in length, branching pattern, and
spatial distribution of dendrites. J Comp Neurol. 339:401-20.
Núñez-Abades PA and Cameron WE. (1995). Morphology of developing rat genioglossal
motoneurons studied in vitro: relative changes in diameter and surface area of somata and
dendrites. J Comp Neurol. 353:129-42.
Paterson D and Nordberg A. (2000) Neuronal nicotinic receptors in the human brain. Prog
Neurobiol. 61:75-111.
Pauly JR, Slotkin TA. (2008). Maternal tobacco smoking, nicotine replacement and
neurobehavioural development. Acta Paediatr. 97(10):1331-7.
Pilarski JQ and Fregosi RF. (2009). Prenatal nicotine exposure alters medullary nicotinic and
AMPA mediated control of respiratory frequency in vitro. Respir Physiol Neurobiol.
169:1-10.

35
Pilarski JQ, Wakefield HE, Fuglevand AJ, Levine RB, Fregosi R.F. (2011).
Developmental nicotine exposure alters neurotransmission and excitability in
hypoglossal motoneurons. J Neurophysiol. 105: 423-433.
Pilarski JQ, Wakefield HE, Fuglevand AJ, Levine RB, Fregosi RF. (2012). Increased
nicotinic receptor desensitization in hypoglossal motor neurons following chronic
developmental nicotine exposure. J Neurophysiol. 107:257-64.
Powell GL, Levine RB, Frazier AM, Fregosi RF. (2015). Influence of developmental
nicotine exposure on spike-timing precision and reliability in hypoglossal motoneurons. J
Neurophysiol. 113:1862-72.
Powell GL, Gaddy J, Xu F, Fregosi RF, Levine RB. (2016). Developmental Nicotine Exposure
disrupts dendritic arborization patterns of hypoglossal motoneurons in the neonatal rat.
Dev Neurobiol. doi: 10.1002/dneu.22379. [Epub ahead of print]
Puram, SV, Kim, A. H., Ikeuchi, Y., Wilson-Grady, J. T., Merdes, A., Gygi, S. P., Bonni, A.
(2011). A CaMKIIβ signaling pathway at the centrosome regulates dendrite patterning in
the brain. Nat Neurosci. 14:973-83.
Raymond, E. G., Cnattingius, S., Kiely, J. L. (2005) Effects of maternal age, parity, and smoking
on the risk of stillbirth. Br J Obstet Gynaecol. 101:301-6.
Roy, T. S. and Sabherwal, U. (1994). Effects of Prenatal Nicotine Exposure on the
Morphogenesis of Somatosensory Cortex. Neurotoxicol Teratol. 16:411-21.
Sholl, D. A. (1953). Dendritic organization in the neurons of the visual and motor cortices of the
cat. J Anat. 87:387–406
Sholl, D. A. (1955). The organization of the visual cortex in the cat. J Anat. 89:33-46.
Slotkin T. A. (2008). If nicotine is a developmental neurotoxicant in animal studies, dare we
recommend nicotine replacement therapy in pregnant women and adolescents?
Neurotoxicol Teratol. 30(1):1-19.
Slotkin, T. A. (1998). Fetal nicotine or cocaine exposure: which one is worse? J Pharmacol Exp
Ther. 285:931-45.
Sokoloff AJ. (1993). Topographic segregation of genioglossus motoneurons in the neonatal rat.
Neurosci Lett. 155(1):102-6.
Sokoloff, A. J. and Deacon, T. W. (1993). Musculotopic organization of the hypoglossal nucleus
in the cynomolgus monkey, Macaca fascicularis. J Comp Neurol. 324(1):81-93.

36
Takahashi S1, Ono T, Ishiwata Y, Kuroda T. (2002). Breathing modes, body positions, and
suprahyoid muscle activity. J Orthod. 2002 Dec;29(4):307-13.
Tong, V. T.,Dietz, P. M., Morrow, B., D'Angelo, D. V., Farr, S. L., Rockhill, K. M., and
England, L. J. (2013) Trends in Smoking Before, During, and After Pregnancy
Pregnancy Risk Assessment Monitoring System, United States, 40 Sites, 2000-2010.
Morbidity and Mortality Weekly Report; 62(SS06); 1–19.
U.S. Department of Health and Human Services. (2014). 5, Nicotine. In The Health
Consequences of Smoking—50 Years of Progress: A Report of the Surgeon General.
Atlanta: U.S. Department of Health and Human Services, Centers for Disease Control
and Prevention, National Center for Chronic Disease Prevention and Health Promotion,
Office on Smoking and Health.
World Health Organization (2015). Tobacco. [Online] Available from:
http://www.who.int/mediacentre/factsheets/fs339/en/. [Accessed 20 June 2016].
Zoli, M., Le Novère, N., Hill, J. A. Jr, Changeux, J. P. (1995) Developmental regulation of
nicotinic ACh receptor subunit mRNAs in the rat central and peripheral nervous systems.
J Neurosci. 15:1912-39.