Embed Size (px)
Citation preview

511U C coUS.AFSAM-TR-89-18 AD-A229 022
THE EFFECTS OF ATROPINE AND PYRIDOSTIGMVINEON THERMVOREGULATION AND WORK TOLERANCE INTHE PATAS MONKEY
Reynaido S. Elizondo; Ph.D.
The Unlvmsit of Texa at E3 PasoEa Paso, TX 7g968-0509
OTICS LECTE
n"m Rs.,Ort %x Pvmiod M" IN6S - September 1968
EAppproied for public release; distribution is unlimited.
PV "are forUSAF SCHOOL OF AEROSPACE MEDCINE"to.u Sstems Dmvton (AFSC)Srooft Air Forc Buss, TX 78235-530

NOTICES
This final report was submitted by The University of Texas atEl Paso, El Paso, Texas, under contract F33615-87-C-0615, job order2729-04-22, with the USAF School of Aerospace Medicine, HumanSystems Division, AFSC, Brooks Air Force Base, Texas. Dr. Stefan H.Constable (USAFSAM/VNC) was the Laboratory Project Scientist-in-Charge.
When Government drawings, specifications, or other data areused for any purpose other than in connection with a definitelyGovernment-related procurement, the United States Government incursno responsibility or any obligation whatsoever. The fact that theGovernment may have formulated or in any way supplied the saiddrawings, specifications, or other data, is not to be regarded byimplication, or otherwise in any manner construed, as licensing theholder or any other person or corporation; or as conveying anyrights or permission to manufacture, use, or sell any patentedinvention that may in any way be related thereto.
The animals involved in this study were procured, maintained,and used in accordance with the Animal Welfare Act and the "Guidefor the Care and Use of Laboratory Animals" prepared by theInstitute of Laboratory Animal Resources - National ResearchCouncil.
The Office of Public Affairs has reviewed this report, and itis releasable to the National Technical Information Service, whereit will be available to the general public, including foreignnationals.
This report has been roviewed and is approved for publication.
STEF CONSTABLE, Ph.D. F. WES-Y BAUMGARDNER, Ph.D.
Project Scientist Superv sor
GEO (, EESCHWE' E, Colonel, USAF, MC, CFSCommander

UNCLASSIFIEDSECIISIli CiLASS.;'CAT!CN 09 Tl-ýS VAcE
REPORT DOCUMENTATION PAGE OA48 No. 0704- 0 1
I&. AEP-IT SIC~RiTy CL.ASS1I$AT4iO 1b RESTRICTIVE MAAXiNGSUnclassified
2&. $&Ct.AI7Y Ci.ASSiF1CATIONj AiiT~q0IrY . iSTjti&UTIONIAAiLA&LMTY Of REPORT'
2b. :)CASFAODVhA~N SCEDL A';"-oved fcr puhlic releo3se distributionjis unlim'ited.
4. PERFORMING ORCIAAiZATION RMPOR1 .. ,SS S M#O~iTORIiN OAtGAtiZATiON REPORT hkMSER(S)
USArSAM-TR-89-18
G& K&A04 Of PIRMOAMIN4G ORAMI4"T)0N 60. Of.CE syliiot 7&. %"AI OF VONiTOLRiG ORC 1ANQ.A70?4The Urniversitv of Texas at USAF School of Aterospac- M4i, inc(VwEl Paso. College-of SCienro6L ADOO4IS ýOy State &Wi7P COW) 71ý ADORE~ SSCiy Sltot. AM ZLPCO
El Paso, Texas 79968-0509 Hum~an Svstems Division (AF3C)Brooks Air Force Base, ',X 781Q-510l
S&. i4AI OfFvhi~S~t0W OFK s~o Sysmaot. I c1KmjRUephr iNST0IN MiN' O4TFICA1bO4N iVtA
9L &DME SS(CAY.S(StO MV Z COW)0 SOURCE Of SjNO~NG t4$QPtOGRAM 0001QECT 'AA OKi
_______________________ 2 202F 272', ' 22
77TITh (kimo Soiwm7 GCaw~kapwiThe Effects of Atropine and Pvrllr stziine -n' L~roc~iii ii-'
ia. PtR$OPAL "Ti4OA(S.z,,ndn). iR' 'n~ildo S.
i3.. "'Pt Of REAO"T 13o.ME COI1t0 jia )Art 0(004T roo, tv Z,47i 5-A
16 S M4fjItvTA*Y NOTATWOpe
'1 (O)SA" COO)I$ 10 00"$~C 1~ ~ C41o 7.1 I
F4I. 1Ir '4-C10 rit 4 nheit x,'., 1,;C iix it i'!! I ' t -
lie effirt If a 1~i In, r1-i, rjl.r atrnn ife n leci:t ion (0.i) 01 ~ it rb.
3' C 'in the ther1miireijii.itrrrv fm i t " aIn' exer' i ýv rji';- j "t ~ ~i'.sr.. nVe'tJVatced. It primate' vr-d~2i i.~ w-ii4 !r- t r~lii , eitithe dr;igsz 4, tne Pxprs ip toleranc-e I i'-e Nectal t4'.;-t-illirv (Tri, .u- 'reirt rilto 1W0ýwere crrnt ifluiiuly futnitored bv ; tei,C-i tTY qqt-401 while s~iter l'-q wi'r 4 ,ý i - tt .- , A tweight 41 f fevr en-* hp fe 're ari 9 a' t v r s xvr iI , e . At ro v jne 4,f f ct % wie '~r r -w ,ervo PTTA, : 1,I 33C am indl¶ ifsed hy ;I ;iinif aint ri'diitii'n in waitor (rrq 4 ri.) wht.,h w.,ý i i' )diiWith 4n 4Vev.ne 'Mo'rr i16e timp If h) min 1,,iq thain the contts'1 Yvfiii,. heo f hii HR inid tri'r Pq,1- n(kt thr~e Itr,,Tlinp vxperimentq wort,' qignitj4carntv iic7.yi~i IbIVC the COiitrri
,V-1~e 1rr'~ ,",:mItie qj'inf iI antlv in, r'i*-zef wa-ter jou. (61I.)wlhi, ! w-%% a''iated with4n 4vvrakvp err.A , p tilinefiA0dmin I rnv:er .tlmn t hi r 0t,11 r vaii ii. The- fi nal HR ind rri'reQipr'nP werp noit qtivnifi'.a;nt~ eiffv tod Ti the pyrdt-111ýtiqffll treaitment.
40 )1T3SYi*Q'OPrAVA#LAgiqii 00 Ali$ii" 'iAvs~ C.,qitV CLASWC~A~iO4
~ Aeu&~ee~'~ ''I CJ~ rAM OPT ), * ;t~ I n a " I I i'd
_ _ _ _~h~ I00Oform 1473. JUN354 PIV-0 0000M tw__44419__ýI
- --- ---
I
i i Ii

!.¶CLA-SSTF1Efl-=
SECU04TY CLASSIF4CATION Of T"?S PCAGE
A continuous flow indirett calorimetry system was developed consisting of a holuirg cagein which the monke, r':.ided. Calculation of metabolic parameters was done trom measure-ments of 02, CO2, PH20' Pa. F ou'and temperature using the algorithms of Brown (3). As
oa addition to the munkev exercising wheel, an airtight enclosure was designed whicher~abl2d the use of the rLmerLerv s' -,em fcr measurement of metaboliJ" rates duringexercise. Tpica] metd'- ... rates durý.;A davlight hours were 40 to 53 watts per metersquare (W/m') at rest ana 70 to 80 ý,/m~ during exeicise. It is highly significant thatthe Respiratory Quotient (RQ) and aetabolic rate measured with our new calorimetrysystem are the same as those we previously reported in chair-restrained animals. This newtechnology will be very useful as we ccntinue to examine the short- and long-term effectsof neuroactive aoents .ýn crimate thermoregulation, metabolisa, and exercise tolerance,
8 y . . ......
~C LA
~~~~~1 T_____________

TABLE OF CONTENTS
Page
INTRODUCTION .................. ........................ I
METHODS AND MATERIALS ................... .................... 1
Design of the Exercise Device ............. .............. 1
Exercise Program Operation .............. ............... 2
Pyridostigmine Study ................. .................. 2
Treatment of Data ................... .................... 4
RESULTS ..................... ............................ 4
DISCUSSION ... ............ ........... ................. 10
REFERENCES .................... ......................... 12
APPENDIX: Report on the Measurement of Energy Expenditure byIndirect Calorimetry in Patas Monkeys at Rest andDuring Exercise ................ ................. 15
List of Figures
Fig.No.
1. Primate Exercise Wheel (PEW) ........... ............ 3
2. Effects of Atropine and Exercise on Heart Rate at Taof 25 'C .................... ...................... 5
3. Effects of Atropine and Exercise on Rectal Tempera-ture at Ta of 25 .C ...... ................... 6
4. Effects of Atropine and Exercise o-i Heart Rate at Taof 35 C .................................. 8
5. Effects of Atropine on Rectal Temperature during ex-ercise at Ta of 35 .C ....... ............... 9
A-1. Monkey Indirect Xtlorimetry System ...... ......... 1.
lii

List of TablesTableINo. Page
1. Summary of Atropine Results During Exercise at25 'C and 35 'C .............. ................... 4
2. Summary of Pyridostigmine Effects During Exerciseat 35 *C ................. ...................... 7
A-i. Calorimetry Data Average Daily 02 and CO 2 and Min-ute Metabolic Rate ........... ................ 16
Iv

THE EFFECTS OF ATROPINE AND PYRIDOSTIGXINEON THERMOREGULATION AND WORK TOLERANCE IN THE PATAS MONKEY
INTRODUCTION
Atropine, the most common antidote for anticholinesterasepoisoning (13), suppresses thermoregulatory sweating, and even-tually, evaporative heat loss through its anticholinergic activity(2) resulting in increased net heat storage (1, 15), decreased heattolerance, and reduced exercise performance (4, 6, 7, 9).
Pyridostigmine is used in conjunction with atropine as aprophylactic against anticholinesterase poisoning by reversiblyinhibiting cholinesterase. Depending on the degree of enzymeinhibition and state of ordination, these drugs affectthermoregulation and exercise performance in a negative (11) orpositive way (12).
In a previous study, the physiological effects of 2 neuro-genic drugs, atropine and pyridostigmine, on the thermoregulatoryeffector system of the patas monkeys were evaluated at rest. Itwas concluded that this species was an appropriate animpl model tostudy the effects of neuroactive drugs on temperature regulationand thermoregulatory capacity (1, 10). This study expands theprevious work by testing the effects of atropine and pyridos-tigmine on the thermoregulatory capacity of patas monkeys duringexercise in the heat. Prolonged exercise in the heat represents acondition where tha metabolic and environmental heat load may beconsiderable and provides the body with maximum strain to chal-lenge the thermoregulatory control system. Since atropine andpyridostigmine interfere with the heat dissipation mechanisms, thetesting of these neuroactive agents during prolonged exercise inthe heat provides maximum challenge to the thermoregulatory systemand gives information about the thermoregulatory capacity ofprimates, and perhaps, man.
MENTODS AND MATERIALS
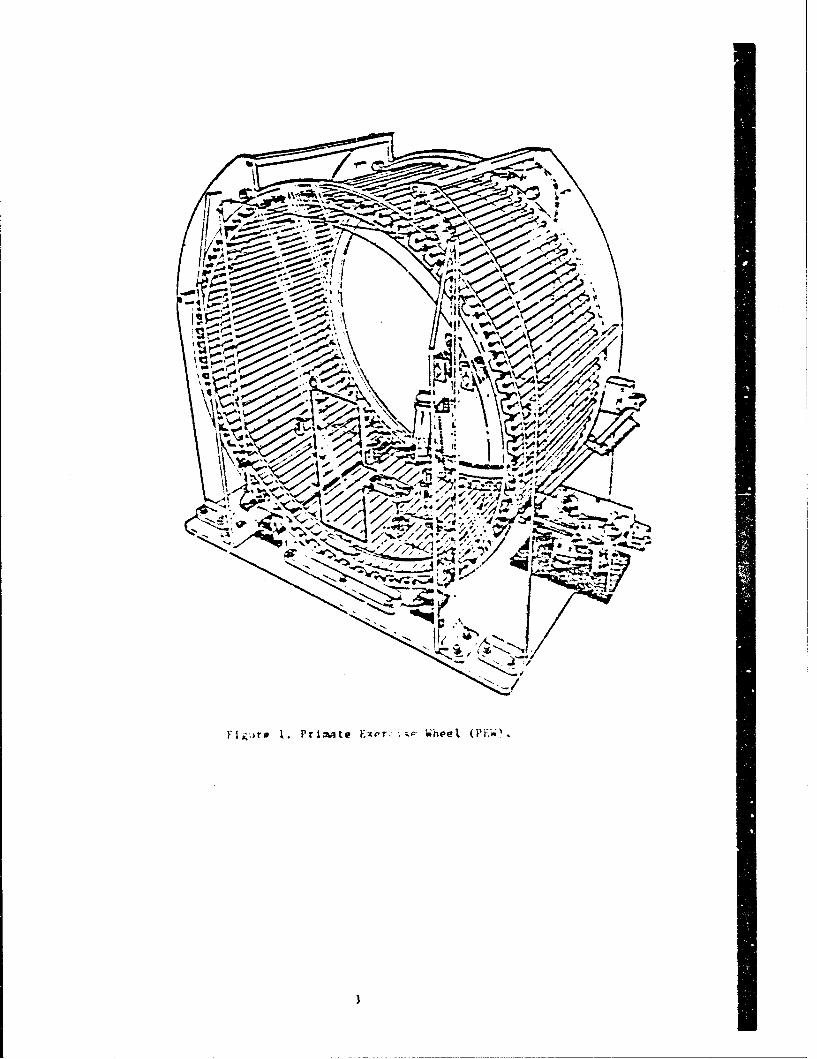
The apparatus used to evaluate the exercise tolerance of thepAtau monkey was a treadmill wheel (Fig. 1) specificallyconstructed to exercise nonhuman primates (8) (Arpendix). Thewhqel, consisting of two 122-cm (48 in.) diameter Lucito rings and120 aluminum bars, forms a circular cage aich rotated freely onfour bearings. Another ring, affixed to the outside of each Lucitering, was connoctad to alternate alluminum bar's to provideelectrical mJmulation for conditioning the animal. The activity

wheel included: 1) a magnetic tachometer pickup to quantitatespeed; 2) an automatic control panel interconnected with thetachometer to allow the setting of upper and lower speed limiCswhich the animal had to maintain in order to avoid a sequence ofvisual or electrical stimuli; 3) a micrcswitch at the bottom of thecontrol panel which counted each revoluticn of the wheel andcalculated distance traveled; and 4) a brake to stabilize the wheelduring the rest periods as well as to prevent the animal fromcperating the wheel at higher than predetermined rates.
Inercise Progran Operation
Five patas monkeys were trained to operate the treadmill wheelunt'l they learned to run at a minimum rate of 2 miles/hour (mph)for 60 min (20).
At the end of a 20-week training period. each animal wascapable of completing at ieast 1 h of exercise while control valuesof exercising heart rate (HR) and rectal temperature (Tre) weremeasured every 15 nin by a noninvasive telemetry system. The systemconsisted of an AMF Quantum XL transmitter for measuring the HR anda Mini-Mitter rectal probe transmitter for measuring Tre.HR was recorded by an AMF Quantum digital watch receiver inbeats/min (bpm) while Tre was recorded by a President AX52 FMreceiver and converted to 0C using calibration curves validated inthis laboratory. Total distance covered in miles, average speed iný-.les/hour, and total exercise time were also recorded. Water losswas estimated from weight differonce measured before and after theexercAsf test.
The criteria used to establish the exercise tolerance of theanina1 .s and to terminate the standard exercise test were any one ofth" following: heart rates that approach maximal heart rates forthi" species, approximately 300 bpm; a Tre :iqher than 40 C toF.rý,toct the animals from heat injiury, or going J times underspoed(2 -p•h) in each of 2 consecutive 15-rin periods.
The standard axorcirip teots were then repeated followingit rr~pmne treatmenort anrJ 3 monthn later after pyrtdotigminetreatment.
Two dayz ifter each animal completed a standard exercise test,,t 2r) "C, anothe." excrcse test was carviod cut immediatelyfollowirg a 0.03 mq/qk sinqla intramuncular (i.m.) atropinein)oction: a doro f-apibles of establishing blood lpvtls of atropir.eisa ,,,ious to t hat rproir)IOw-' in himanq following an i.m. in) jction of
nrl (14, 17). Tha siamp proci'dure wan r-pented 3 1;ayn later at
Pyrldogtiqaine Otudy
(,fe •day aft i!r a t dard arall tyst w)n carriet out atj' 0 ach animal1 wat; t iepatpl' oral I y wit h I sepairate doses of
4'
ISI
\~ vt
145~
,or~ ',
Vi ilt 1. Pr imate Exer, -,;ae (PEoO.

solid 0.4 mg/kg pyridostigmine in a piece of banana. The afternoonof the next day, the animal repeated the exercise test afterreceiving 2 doses of 0.4 mg/kg pyridcstigmine- 1 in the morning and1 an hour prior to exercise, making a total of 5 doses. The 0.4mg/kg pyridostigmine dosage is capable of reducing cholinesteraseplasma levels by 25 to 30% of normal immediately after the seconddose and keeps the levels down througnout the treatment period (1).
Treatment sf Da•
The results are presented as means and standard deviation.They were analyzed by a two-way (condition x time) analysis ofvariance (ANOVA) with repeated measures on both factors. Turkey'stest of critical differences was also' used where appropriate. Allsignificant differences are reported at p < .05, unless otherwisenoted.
RESULTS
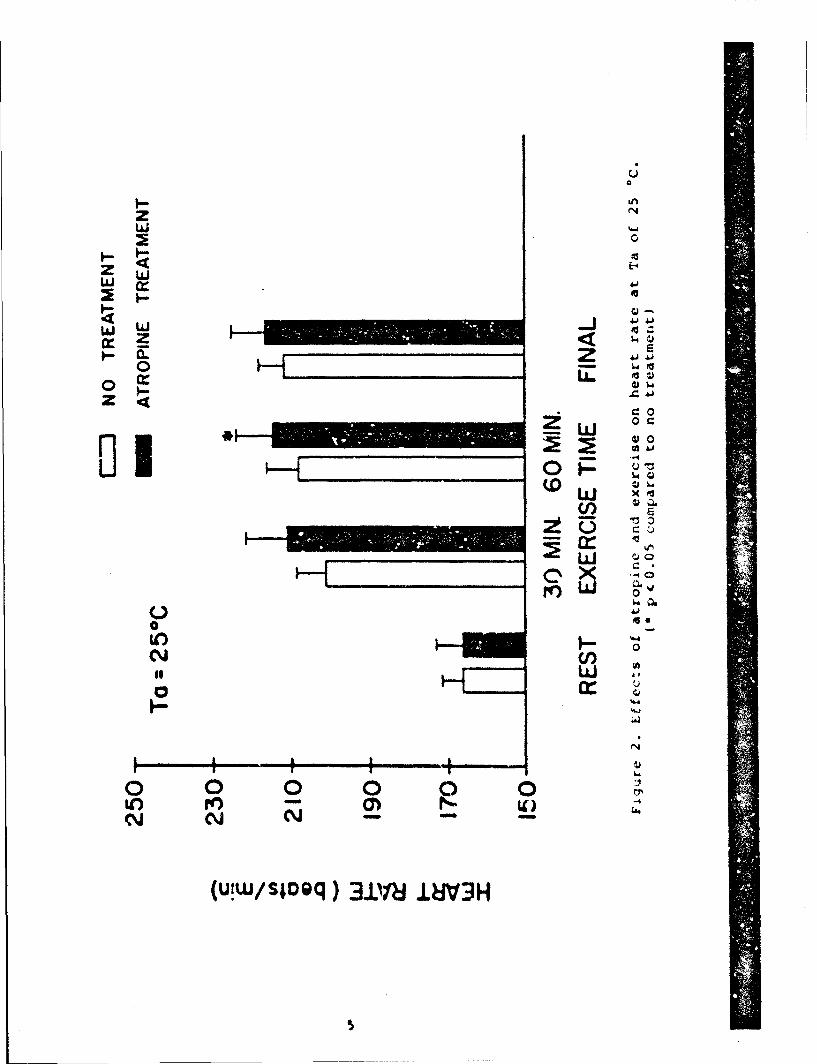
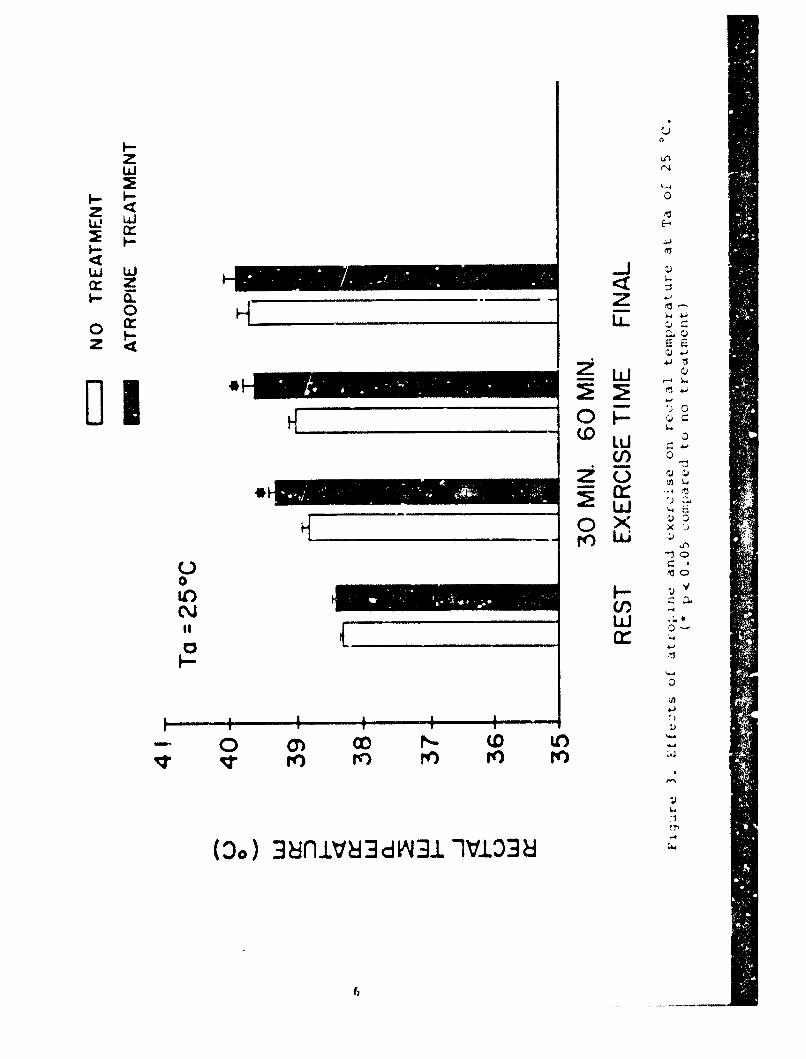
The effects of atropine at 25 IC are shown in Table 1 andFigures 2 and 3.
TABLE 1. SUMMARY OF ATROPINE RESULTS DURING EXERCISE AT25 *C AND 35 *C
Water loss Time Distance Speed(g/min) (min) (mi) (mph)
' -i - 25 0C
Cortrol 1.2 177.0 4.8 1.6+_1.7 +_39.7 +1.2 10.05
Atropine 1.3 146.8' 4.0 1.6411.6 *_30.5 *0.9 t0.1
Ta - 35 0C
Crn t ro1 2. 149.6 3.6 1.5t2.1 43'7.1 40.8 ±O.1
At rop i ni 1 .8 M 84.00"' 2. 5t) 1.7'*2. ] *-.9 40.5 •0.1
" p " .05"M p OR
Atropine sir;niflcAntiy Incroaesd fAtique And shortened theaxprciqo tolerAnce t imo by ApprnximAtoly 30 min. No uhjqniri-antlift-r-nco wn% rounl in the amo(unt or wimtr lons, distance triveolodor the anima1n' nv.raqo nvpo(I. rlqurops 2 And I show that mean HP
4
z
c-. zzL z
0 mULL
0
;~0 0C.)
Lin
~~WW
'S ~w
0 0 0 0 0 0
(utuw/s40q) 31118 18~V3H
z Lw
II-. ~0z
I-
a. z
0LJ
00LCL)N U)
00
a), TV) TV
(3)JnfiV8dLN~i 1=38J
and Tre responses increased from rest to exercise. While HR duringexercise increased slightly over time in the control experiments,atropine administration caused a further increase in HR and wassignificantly greater at 60 min when compared to the no treatmentcondition. In addition, atropine was associated with a significantincrease in Tre by the end of the first 30 min of exercise andremained 0.5 *C higher at 60 min of exercise, significantlydifferent compared to the respective no treatment period. Meanfinal HR (F-HR) and Tre(F-Tre) values were not significantlydifferent from the respective values in the control experiments.
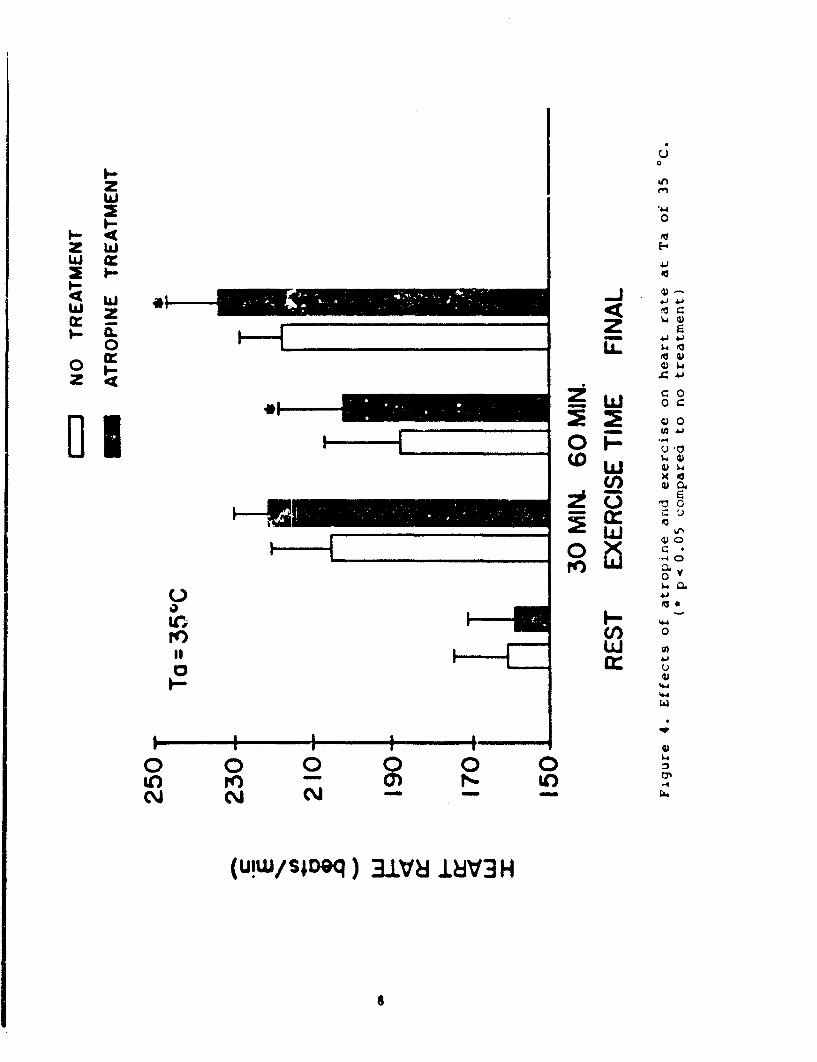
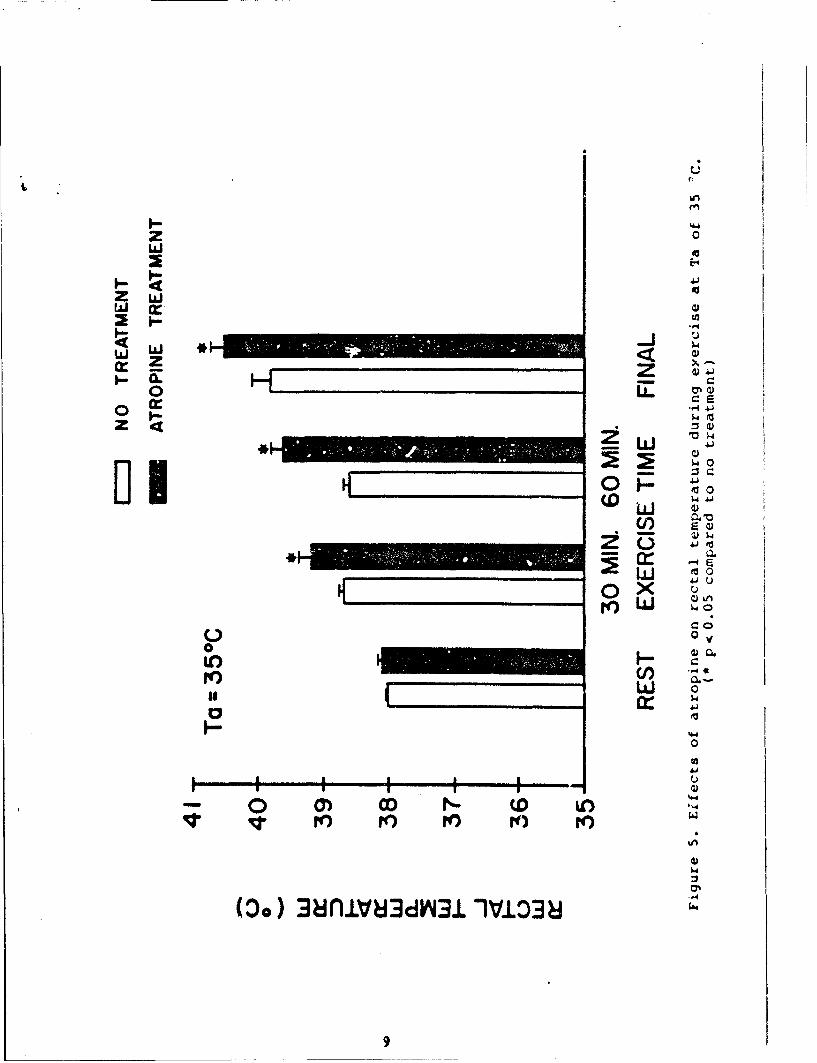
The effects of atropine at 35 0C appear in Table 2 and Figures4 and 5. At this environmental temperature, atropine produced areduction in water loss rates (22%) and a reduction in totalexercise time of 65 min less than ;he control value. Consequently,the distance covered in miles was also less (p < .06). It was alsointeresting that average speed in mph was slightly butsignificantly faster in the atropine experiments as compared to thecontrols. As illustrated in Figure 4, the mean HR responseincreased from rest to exercise and at the end of exercise. Themean HR response at 60 min of exercise decreased slightly from thatat 30 min of exercise due to the fact that 2 animals reached theirexercise tolerance before the first hour of exercise and only themean HR response for 3 animals is shown. The mean HR response inthe atropine experiments was higher at 60 min of exercise andbefore the end of exercise compared to the respective controlexperimental periods. Figure 5 shows the mean Tre responses in theheat continued to increase throughout the exercise time. However,atropine administration caused further increase in the rate of heatstorage, compared to the respective no treatment exercise period.
Table 2 shows pyridostigmine treatment tended to increasewater loss (11%) and was associated with an average exercise timeof 61 min longer than the control value and consequently longerdistance traveled in miles. Average speed in mph was slightly butsigniticantly higher in the pyridostigmine experiments compared tothe controls. Final heart rate (F-HR) and final Tre (F-Tre)responses were not significantly different from those in thecontrol experiments.
TABLE 2. SUMMARY OF PYRIDOSTIGMINE EFFECTS DURING EXERCISE AT35 0C
R-HR F-HR R-Tre 7-Tre Water Time Distance SpeedLoss
(bRnM) (Mp ) OC) (-i-C) (aliin)(mm) 12 (4m01
Control 160.4 223.0 37.9 39.3 1.9 115.0 3.0 1.5t10.9 +11.0 +0.1 t0.2 +3.0 +29.0 t0.8 ±0.1
Pyrido- 160.6 214.2 37.7 39.0 2.1 175.0* 5.0* 1.7'stigmine t10.7 +11.8 +0.2 +0.3 +3.0 +29.0 +U.9 +0.1
* p < .05R-RestingF-Final
7
zwW0
wzwCLJ
Ir-
0- O
Q 0(A )
0 I.M .Uu
w 0 1
".4
0~~ 0 0 0
i()cf 0)10
a)
zU
cr.4
1-0
Cz z
Z41
(00*-41 w 0
C/) a0v
W0a
41 uX u I
I - 0)
U)
0)
W4
9A

DISCUSSION
The results of these studies showed that the physiologitcaleffects of atropine in exercising patas monkeys were similar tothose reported for humans. The suppressed sweating capacity of theatropine significantly affected the heat loss mechanisms of theexercising animals in the heat as indicated by the 43% reduction inwater loss. On the contrary, during exercise at thermoneutralenvironment, the amount of water loss was not significantlydifferent between the atropine and c'.,ntrol experiments. Duringexercise, there is enhanced release of epinephrine by the adrenalmedulla which accounts for increased sweating (24); a fact which atneutral environment seems to completely counteract the negativeeffects of atropine on sweating. The suppressed sweating inducedby atropine generally resulted in increased heat storage causingconsiderable ther"al strain on the animals. It is speculated toaccount for the reduced time that the animals could exercise. Inaddition, exercise in the heat caused much faster rise in the rateof neat storage on the animal's body with potentially greaterperformance decrements and increased risks for heat injuries. Forinstance, exercise in the heat, for all animals but one, wasterminated by the investigator when the Tre was in excess of 40 OC.Conversely, at the thermoneutral environment, the animals tooklonger to reach exercise tolerance. The time that elapsed betweenthe first signs of discomfort and the time they reached thecriteria for termination of exercise (see Methods and Materials)was also much longer. At this cnvironmertal temperature, allanimals but one were stopped from exercising because they fulfilledthe criterion concerning the speed limit that should be sustained.Although mean F-Tre response at 25 0C was not significantlydifferent in the control and atropine exercise experiments, thesedata revealed that animals with low exercise tolerance (exercisetime < 2.30 h) demonstrated much higher F-Tre values in tteatropine experiments (40.2 "C vs control value of 39.5 CC). This mayhave to do with the pharmocokinetics of atropine in the plasma(half-life 2.5 h) and the fact that the exercise time of theanimals varied between 1.30 and 5.0 hours.
Mahoney (22) investiqated the response of a single patasmonkey while running in the heat and reported that at any giventemperature the onset of exer.ise caused a threefold increase !nconductance reqardless of the particular speed. Some of thisincreave could be associated with increased blood flow to theworking muscles, and previous work from this laboratory in restingAnimals showed that atropine induced peripheral vasodilation asindicated by increased whole-body conductance (1). Similarincreases in ronductance were also found in exercising humansubjects after atropine (9, 19). Therafore, it was speculated thatincreased blood pooling in the peL phery to compensate for thedecreased heat dissipation from sweating would have aqgravated theproblem by compromising blood flow to the exercising muscles andwould havy reduced their exercise capacity.
10

Although atropine blood levels were not measured in thisstudy, results show that the time course for the physiologicaleffects of atropine on heart rate and heat loss mechanisms, asindicated by increase in Tre, was between 30 to 60 min post-injection. This finding agrees with the reslts found by otherinvestigators involving a variety of atropine dosages injected inhuman subjects (5, 6, 21, 23, 25). Furthermore, Craig (4) reportedthat 2 mg of atropine, a dose analogous to the one administered inthe present study, resulted in nearly maximal vagal inhibition. Inthis study, mean final heart rate increased by !6 bpm and meanfinpl rectal temperature increased by 0.7 0C under the influence ofatropine during exercise in the heat. Various results, ccn.,erningthe amount of increase in HR and Tre, have been found in humanstudies (4, 21, 25). Direct comparisons between the studies aretenuous because of differences in environmental conditions, subjectvariability, exercise mode, and protocols used.
The effects of pyridostigmine treatment have been reported inrats but not in primates. Francesconi et al. (11) reported that inrats, pharmacological acute doses of pyridostigmine qreat.&,affected exercise performance, thermoregulation, and chemicalindices of heat/exercise injury following 64% cholinesterase inhi-bition. In another study, Francesconi et al. (12) reported thatmoderate cholinesterase inhibition of 23% and 39% attenuatedseveral of the acute responses and had no debilitating effects onphysical performance and thermoregulation during exercise in theheat. They reached the conclusion that a rather narrow range ofcholinesterase inhibition must be achieved before the physiologicaland protective effects of pyridostigmine are observed. Theirfindings agzee with the results found in this study. We found thatoral administration of 0.4 mg/kg pyridostigmine improvedthermoregulatory function and was associated with a 60 min longeraverage exercise time than the control value.
11

REYERENCIS
1. Avlonitou, E., and Elizondo, R. S. Effects of atropine andpyridostig.mine in heat-stressed patas monkeys. Aviat SpaceEnviron Med 59:544-548 (1988).
2. Batra, G., Traystman, R., Rudnick, H., and Menkes, H. Effects,of body position and cholinergic blockade on mechanisms ofcollateral ventilation. J Appl Physiol 50:358-360 (1981).
3. Brown, E. Analysis of gaseous exchange in open circuit calo-rimetry. Medical Biological Engineering and Computing.22:333-338 (1984).
4. Craig, F. N. Effects of atropine, work and heat on heart rateand sweat production in man. J Appl Physiol 4:309-319 (1952).
5. Craig, F. N., and Cummings, E. Dehydration and muscular work.J Appl Physiol 21:670-674 (1966).
6. Cullumbine, H., McKee, W. H. E., and Creasey, N. H. Theeffects of atropine sulfate upon healthy male subjects. Q JExp Physiol 40:309-319 (1955).
7. Cullumbine, H., and Miles, S. The effects of atropine sulfateon men exposed to warm environments. Q J Exp Physiol 41:162-179 (1956).
8. Curran, C. R., Wiegel, W. R., and Stevens, D. N. Design andoperation of an exercise device for subhuman primates. ArmedForces Radiology Research Institute TN 72-1 Defense NuclearAgency. Bethesda, MD (1972).
9. Davies, C. T. M., Brotherhood, J. R., and Zeidiford, E.Effects of atropine and beta blockade on temperatureregulation and perforvance during prolonged exercise. Eur JAppl Physiol 38:225-2':2 (1978).
10. Ellzondo, R., Primate models to study sweating. Am J Prima-tology 14:265-276 (1988).
11. Francesconi, R., Hubbard, R., and Mager, M. Effects of pyrido-stigmine on ability of rats to work in the heat. J ApplPhysiol 45(4):891-895 (1984).
12. Francesconi, R., Hubbard, R., Matthew, C., Lava N., Young J.,and Pease, V. Oral pyridostigmin. administration in rats:Effects on thermorequlation, clinical chemistry, and per-formance in the heat. Pharmacol Biochem Behav 25:1071-1075(1986).
12

13. Gisolfi, C. F., Sato, K., Wall, P. T., and Satc, F. In vivoand in vitro characteristics of eccrine sweating in patas andrhesus monkeys. J Appl Physiol 56 (3): 425-431 (1982).
14. Harris, L. W., Stitcher, D. L., Heyl, W. C., Lieske C. N.,Lowe, J. R., and Cl'ark, J. H., Broomfield, C. A. The effects ofatropine-oximetherapyon cholinesterase activity and survivalof animals intoxicated with p-nitrodiphenyl-di-n-butylphos-phinate. Toxicol Appl Pharmacol 49:23-29 (1979).
15. Hubbard, R. W., Matthew, C. B., and Francesconi, R. Heatstressed rat: Effects of atropine, desalivation or restraint.J Appl Physiol 53:1171-1174 (1982).
16. Johnson, G. S., and Elizondo, R. S. Temperature regulation inMacaca m : A thermal balance study. J Appl Physiol 46:268-277 (1979).
17. Kalser, S. C., and McLain, P. L. Atropine metabolism in man.Clin Pharmacol Ther 11:214-227 (1970).
18. Kolka, M. A., and Elizondo, R. S. Thermoregulation in Ery-throcebus patas: A thermal balance study. J Appl Physiol55(5):16303-16308 (1983).
19. Kolka, M. A., and Stephenson, L. A. Cutaneous blood flow andlocal sweatinq after systemic atropine administration.Pflugere Anch 410:524-529 (1987).
20. Kramsch, D. M., Aspen, A. S., Abramawitz, B. M., Kreimendahl,T., and Hood, W. R. Reduction of coronary atherosclerosis bymoderate conditioning exercise in monkeys on an atherogenicdiet. N Engl J Med 305(25):1483-1489 (1981).
21. Levine, L., Sawka, M. N., Joyce, B. E., Cadarette, B. S., andPandolf, K. B. Varied and repeated atropine dosages andexercise heat stress. Eur J Appl Physiol 53(l):12-16 (1984).
22. Mahoney, S. A. Cost of locomotioun and heat balance during restand running from 00 to 55 0 C in a patas monkey. J Appl Physiol49:788-800 (1980).
23. Mirahhur, R. K. Comparative study of the effects of oral andi.m. atropine and hyoscine in volunteers. Br J Anaesth 50:591-598 (1987).
24. Robertahaw, D., Taylor, C. R., and Mazzia, L. M. Sweating inprimates: Secretion by the adrenal medulla during exercise.Am J Physiol 224:678-681 (1973).
13

25. Sawka, M. N., Levine, L., Kolka, M. A., Appleton, B. S.,Joyce, B. E., and Pandolf, K. B. Effects of atropine on theexercise - heat performance of man. Fundam Appl Toxicol4:S190-S-194 (1984).
14

APPENDIX
REPORT ON THE MEASUREMENT OF ENERGY EXPENDITUREBY INDIRECT CALORIMETRY IN PATAS MONKEYS
AT REST AND DURING EXERCISE
A continuous flow indirect calorimetry system was developedconsisting of a metabolic cage in which the monkey resided. Freshair was pulled through the cage by a fan and the airflow out wasmeasured by a turbine flow-rate meter. Airstream temperature andwater vapor pressure were measured before a small aliquot of gaswas sampled for oxygen and carbon dioxide content (Fig. A-l).Calculation of metabolic parameters from the measurement of
2 ,CO, 2 ,P,, P,, F ,and temperature was done using the algorithms ofBrown (31', which account for the storage (integration) of gasvolumes in the metabolic cage. As an addition to the monkeyexercising wheel, an airtight enclosure was developed which usedthe calorimetry system for metabolic rate measurement duringexercise.
Five patas monkeys underwent a week of metabolic measurementsin the metabolic cage. These same 5 monkeys then underwent a weekin the metabolic cage combined with 3 exercise periods of 3 h.Typical metabolic rates during daylight hours were 40 W/m2 at restand 70 to 80 W/mz during exercise. Table A-1 illustrates thecalorimetry data for daily oxygen, carbon dioxide, and metabolicrates for each monkey at rest and with exercise. It should be notedthat the W/m2 are on a per minute basis and are averages for anentire day including the dark period in which metabolic rates maygo as low as 16 to 18 W/m2 .
15
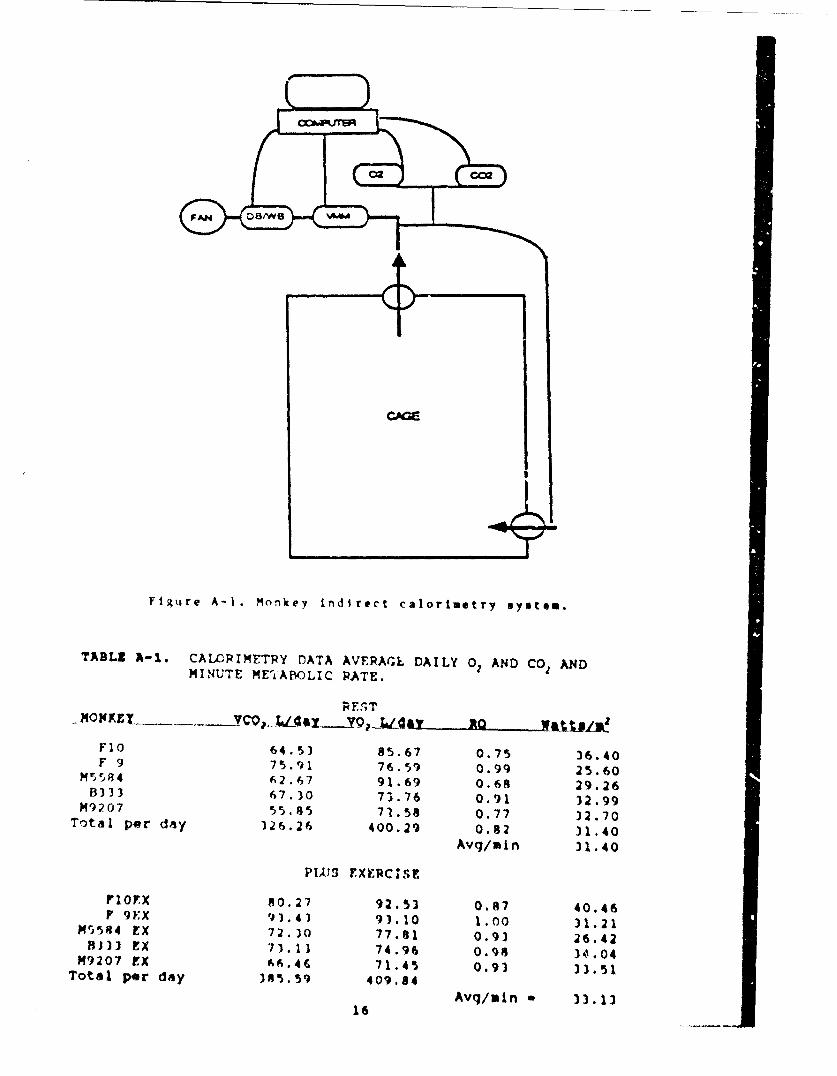
Figure A-I. Monkey indirect calorimetry system.
TABLZ A-i. CALCRIMET•Y DATA AVERAGL DAILY 0 AND CO, ANDMINUTE MEMABOLIC RATE.
FIO 64.53 85.67 0.75 36.40F 9 75.91 76.59 0.99 25.60f) 4 f2.67 91.69 0.68 29.26B333 67.30 73.76 0.91 32.99M9207 55.85 71.58 0.77 32.70Total per day 326.26 400.29 0.82 31.40
Avg/min 31.40
PUJ5 EXERCISE
riOX $0.27 92.53 0.87 40.467 9VX 91.43 93.10 1.00 31.21K5584 EX 72.30 77.81 0.93 26.42
RJJJ EX 73.11 74.96 0.98 34.04M9207 EX 6 .46 71.45 0.93 33.51Total pIr day 345.59 409.84
Avg/min * 33.1316

The results of the study using the newly designed continuousflow indirect calorimetry system for quantitation of energyexpenditure in unrestrained monkeys is significant in light of on-going studies of the effects of neuroactive agents on temperatureregulation, metabolism, and work tolerance.
This is the first study that reports resting metabolic ratedata in unrestziined patas monkeys. It is highly significant thatthe TQ and metabolic rate during daylight hours mea3ured with ournew calorimetry system are of the same order of magnitude as thosewhich we have previously reported (16, 18) in chair-restrained3nimals. It should also be noted that the RQ's recorded in theexercise experiment are those which one would predict in anexercising animal (Table 1). Furthermore, the metabolic ratequantitates for the first time that,with the exercise wheel, weare inducing metabolic rates that are approximately twice those ofthe resting level. The metabolic rates during the exerciue periodaere significantly higher than the daylight resting metabolicrates. Finally, it should be noted that in terms of 24 h energyexpenditure, 3 exercise periods of 3 h duration during a 24 hperiod did not result in a significant increase in the metabolicrate when averaged over a 24 h period (Table A-i).
It is clear that the technology that has been developed willbe very useful as we continue to examine the short- and long-termef.ects of neuroactive agents on thermoregulation, metabolism, andexercise tolerance in nonhuman primates.
17





























