Embed Size (px)
Citation preview

�9 1991 by The Humana Press, Inc. All fights of any nature, whatsoever, reserved. 0163-4984/91/3103-0223 $02.20
The Effect of In Ovo Boron Supplementation on Bone Mineralization
of the Vitamin D-Deficient Chicken Embryo*
NANCY KING, 1 TED W. ODOM, .2 H. WAYNE SAMPSON, 3
AND ANDREW G. YERSIN 4
1United States Army Research Institute of Environmental/Vledicine; ~Department of Poultry Science, Texas Agricultural Experiment Station,
College Station, TX,. 3Department of Anatomy, College of Medicine, Texas A & /Vl University; and 4Dept. Poultry Science, Pennsylvania
State Univ., USDA-ARS, Food Animal Protection Research Laboratory
Received May 14, 1991; Accepted May 21, 1991
ABSTRACT
It has been hypo thes i zed that boron (B) is an essential e l ement for animals, but its action will vary greatly depend ing on the nutr i ture of the organism. One of the nutr ients implicated as having an interac- tion with boron is cholecalciferol (Vit D3). This s tudy was carried out to de te rmine if such an interact ion exists. The s tudy was conduc ted utilizing v i tamin D-deficient chicken embryos that were injected th rough the shell at 8 d of embryogenes i s with carrier (NaC1 and/or acetone), B (0.5 rag), B + Vit D3 (0.5 mg and 0.3 i~g, respectively), or Vit D 3 (0.3 or 1.5 p,g). The in oao concomitant adminis t ra t ion of boron and vi tamin D enhanced (p < 0.05) the hatchabili ty of the v i tamin D-deficient embryos . Fur thermore , boron and/or v i tamin D3 in- creased (p < 0.05) the percent of bone ash and decreased (p < 0.05) the exaggera ted height of the proliferative zone of the ep iphysea l
*The opinion or assertions contained herein are the private views of the authors and are not to be construed as official or as reflecting the views of the Department of the Army or the Department of Defense.
*Author to whom all correspondence and reprint requests should be addressed.
Biological Trace Element Research 223 VoL 31, 1991

224 King et aL
growth plate normally observed in vitamin D deficiency, suggesting a more rapid bone formation. The results provide further evidence supporting the hypothesis that boron plays a role in bone mineraliza- tion through an interaction with vitamin D.
Index Entries: Boron; chick embryo; vitamin D-deficient; bone mineralization; growth plate; in ovo.
INTRODUCTION
The essentiality of boron for higher plants was first demonstra ted by Agulhon in 1910 (1). However , it was not until 1923 that these findings were accepted as definite when Warington published similar results (2). Since then, scientists have tried to determine whe ther boron plays an essential role in animal nutrition as well (3-7).
Skinner and McHargue suggested the possibility of boron interac- tions with other nutrients when they reported an interaction between boron and potassium (6). More recently, results from Nielsen and Hunt ' s experiments have suggested that the presence or absence of other nu- trients, such as cholecalciferol, methionine, calcium, magnesium, man- ganese, a luminum, molybdenum, and perhaps others, may augment or mask the physiological effects exhibited by boron deficiency (8-13). Con- sequently, the physiological actions of boron are not consistently ob- served; therefore, boron has not been considered as an essential nutrient for animals (14-16).
Several factors favor the possibility that boron interacts with vitamin D. First, boron has a known affinity for organic compounds containing hydroxyl groups (17), second, there are a number of hydroxyl groups on the activated vitamin D molecule (18), and third, vitamin D has a lack of metabolic activity unless hydroxylated (18). Hunt ' s investigations have provided further evidence that an interaction exists (11-13,19). Specifi- cally, boron supplementat ion in the diet caused a decrease in elevated plasma glucose, [3-hydroxybutyrate, pyruvate, triglyceride, alkaline phosphatase, and T 3 normally associated with vitamin D 3 deficiency in chicks (19). Boron has also been found to enhance growth (8) and de- crease the growth plate height in the vitamin D-deficient chick (13). Calcium and phosphorus apparent absorption and balance were im- proved in vitamin D-deficient rats when boron was included in the diet (20). These observations led Hun t to hypothesize that boron may play a role in bone metabolism. Other studies have provided additional evi- dence support ing this hypothesis (21-28). The purpose of the present s tudy was to de termine whether an interaction between vitamin D and boron was evident from bone mineralization in vitamin D-deficient chick- en embryos.
Biological Trace Element Research Vol. 31, t991

In Ovo Boron Supplementation 225
MATERIALS AND METHODS
Production of a Vitamin D-Deficient Chicken Embryo
Vitamin D-deficient chicken embryos were produced by feeding laying hens a diet deficient in cholecalciferol, but supplemented with lo~, 25-dihydroxycholecalciferol (1 ~g/kg of diet) (Table 1). It has been dem- onstrated that this active form of vitamin D is not transferred in sufficient amounts from hen to egg (30-32). The hatching eggs in this s tudy were de termined to be vitamin D-deficient by the lower hatchability and bone ash when compared with hatching eggs from hens fed a s tandard laying hen diet (Table 2).
Experimental Protocol Two hundred vitamin D-deficient and 10 vitamin D-adequate hatch-
ing eggs were stored at 10~ for 4-5 d. The eggs were numbered, weighed, and placed in a forced-ventilation incubator (Petersime model 1, Petersime Incubator Co., Gettysburg, OH) at 37.6~ with relative humidi ty of 56%.
On day 8 of embryonic development , the eggs were candled. All infertile eggs and those exhibiting a dead embryo were discarded. The remaining vitamin D-deficient eggs were randomly placed in five treat- ment groups to receive carrier, boron, and/or one of two levels of cholecalciferol. Since cholecalciferol is hydrophobic while boron is hydro- phylic, each egg was injected in two sites using physiological saline (0.87% NaC1) and acetone as carriers. Ten vitamin D-adequate eggs were used as negative controls and, therefore, not assigned to any treatment group.
The injection sites were wiped with 80% alcohol, and each egg was injected onto the chorioallantoic membrane with an equal vol of fluid (100 ~L) as follows:
1. Acetone and NaC1 ( - D ) (vitamin D-deficient control) 2. 0.5 mg of B and acetone (B) 3. 0.5 mg of B and 0.3 ~g of cholecalciferol (B + D) 4. 0.3 ~g of cholecalciferol and NaC1 (0.3D) 5. 1.5 ~g of cholecalciferol and NaCI (1.5D)
The boron and physiological saline injections were prepared using borax and NaC1, respectively (Fisher Scientific, Fair Lawn, NJ). The vitamin D injections were prepared by dissolving cholecalciferol (vitamin D3, acti- vated 7-dehydrocholesterol, Sigma Chemical Company, St. Louis, MO) in acetone.
The injection sites were sealed with paraffin, and the eggs re turned to the incubator. On day 18 of embryonic development , the eggs were
Biological Trace Element Research Vol. 31, 1991

226 King e t aL
Table 1 Composition of Diets"
1,25-Vitamin Ingredients Laying hen, %" D 3, %'
Milo 9% - 67.379 Corn, yellow 65.555 - Soybean meal 48% 10.852 20.979 Oystershell 8.064 8.195 Monocalcium phosphate 1.776 1.737 Corn gluten meal 7.174 - Alfalfa meal 17% 5.683 - Sodium chloride 0.310 0.250 Fat, vegetable - 0.963 Fat, animal/vegetable blend 0.110 - MHA Met86 0.005 0.196 L Lysine 98% 0.172 - Vitamin premix 0.250 '''~' 0.250 r,~ Trace mineral mix #4'." 0.050 0.050
"Value given is percent of mix. "Standard laying hen diet adequate in all essential nutrients according to NRC (29). 'Standard laying hen diet with 1,25-Vitamin D3 (1 p~g/kg of diet) instead of vitamin D 3. '~Vitamin mix provides (per kilogram of diet): vitamin A 8800 IU, vitamin E 27.50 IU,
menadione 2.2 rag, menadione sodium bisulfite complex 6.6 mg, thiamine 1.1 mg, thiamine mononitrate 1.2 mg, riboflavin 6.6 rag, niacin 33 mg, choline 430 mg, choline chloride 495 rag, d-pantothenic acid 11 mg, Ca d-pantothenate 11.96 mg, pyridoxine 1.1 mg, pyridoxine HC1 1.34 rag, vitamin BK2 0.0165 mg, biotin 0.22 mg, folic acid 0.77 mg, ethoxyquin 55 mg, and vitamin D 3 2200 IU.
"Nutrius, Inc., Brecksville, OH. ~Mount Air Vitamins, North Little Rock, AR. ."Vitamin mix provides same as stated above without vitamin D 3. J'Trace mineral mix provides the following in mg/kg of diet: manganese 68.35, iron
24.45, iodine 1.1, copper 4.4, zinc 55.05, and selenium 0.1.
candled again. On ly those eggs (n = 123) wi th a live embryo were t ransfer red to a ha tcher (Petersime model H-145) wi th a t empera tu re of 36.9~ and 69% relative humid i ty .
On the d a y of ha tch ing (21 d), morta l i ty rate was d e t e r m i n e d ([hatched eggs + t ransfer red eggs] x 100). All living chicks were we ighed , leg-banded, and t ransferred to the diagnost ic laboratory for t issue sampling.
Whole Blood and Plasma Anatysis
Whole blood (>500 I~L) was obta ined via cardiac punc tu r e in hep- a ranized syr inges . The blood was immedia t e ly ana lyzed for Ca 2 +, Na +, and K + (Chemly te Ion Selective Analyzer , Gilford Systems, Oberlin, OH). Remain ing blood samples were kept on ice and cent r i fuged (Model CR3000, Jouan Inc., Winchester , VA). Plasma was frozen for later analy- sis of Mg, total Ca, and Pi- Total p lasma Ca and Mg were de t e rmined by atomic absorp t ion spec t ropho tome t ry (Perkin-Etmer mode l 4000, Nor-
Biological Trace Element Research 'coL 31, ] 991

In Ovo Boron Supplementation
Table 2 Effect of 1,25-Vitamin D3 Supplementation
on Egg Hatchability and Bone Ash Percentage"
227
Diet Hatchability, %' Bone ash, %
Laying hen 93.33 _+ 3.76' 32.64 + 0.49 ' 1,25-Vitamin DJ' 62.64 + 2.93'- 30.40 + 0.292
"Data are mean +__ SEM. Means followed by different superscript numbers within a column differ significantly (p -< 0.05; Duncan's Multiple-Range Test).
~'Vitamin D3-deficient diet supplemented with 1,25-Vitamin D3 (1 ~g/kg of diet). 'Percent hatch from those eggs with a living embryo on day t8.
walk, CT) (33). The analysis of plasma Pi was performed using the Sigma Diagnostic procedure No. 360 UV (Sigma Diagnostics, St. Louis, MO) reading at 340 nm in a Beckman spectrophotometer model DU-6 (Beck- man Instruments, Inc., Irvine, CA).
Bone Analyses
Both legs were removed, and the muscle dissected from the bone and discarded. The left tibiae were measured for length and width; fat was extracted (Soxhlet Extraction Apparatus) with ether for 16 h and dried overnight in a drying oven at 102~ The dehydrated , defatted tibiae were weighed and ashed in a muffle furnace (Thermolyne Furnatol II, Thermolyne Corporation, Dubuque, IA) at 550~ for 18 h. The ash percentage of each bone was determined, and its mineral composit ion assessed by atomic absorption (Ca, Mg, Na, K) and calorimetrically (Pi) as described above.
Histology The right dissected tibiae were slit longitudinally and placed in
Carson's fixative. Six tibiae from each treatment group were chosen at random for histological and morphometr ic analysis. One-half of the proximal tibiae was dehydra ted with ethyl alcohol and embedded in paraffin. Two 5-~m sections were taken from each sample, and stained with hematoxylin and eosin. The growth-plate was traced indicating reserve, proliferative, and hyperthropic zone using a magnification of 14.75 x , and the height of each zone was measured at three intervals by an individual unfamiliar with the treatment distribution. The calculated average of the three measurements was used as the zone height.
Statistical Analyses
The data were statistically analyzed by the Statistical Analysis Sys- tem (34) utilizing the General Linear Model Procedure for an unbalanced analysis of variance (ANOVA) (35). A p value of <0.05 was considered statistically significant, and the means were parti t ioned by Duncan 's Multiple-Range Test (36).
Biological Trace Element Research Vol. 31, 1991

228 King et al.
RESULTS
Hatchability
Figure 1 shows that embryo hatchability was significantly higher (p < 0.05) for B + D than for 0.3D and B treatments. One hundred percent of the vitamin D-adequate embryos hatched.
Body Weight Neither the initial egg weights, body weights at hatching, nor the
calculated relative body weights ([body weight + initial egg weight] x 100) were significantly different among the treatments.
Bone There were also no significant differences for bone dry weight,
length, or width among treatments (Table 3). However, the ash percent- age was significantly higher (p < 0.05) for B + D compared with - D (vitamin D-deficient control) and B (Table 3). The ash percentage of the vitamin D-adequate chicks (32.77 _+ 0.51; 1l = 10) was similar to the B + D, 0.3D, and 1.5D treatments. The mineral composition of the bone ash was similar for all the treatments.
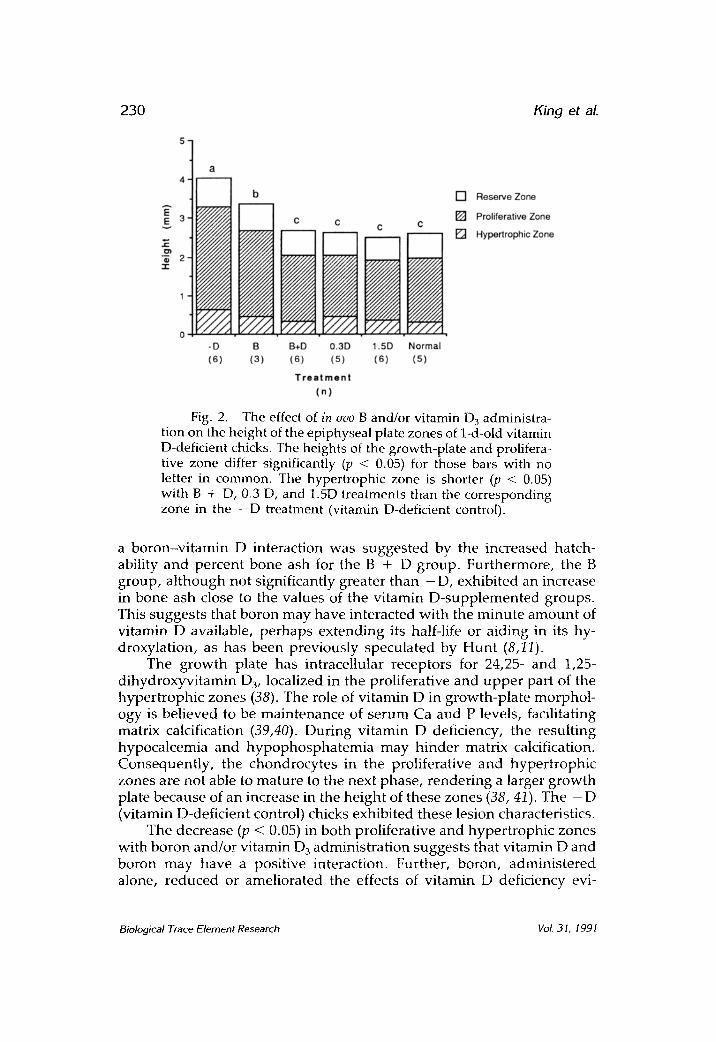
Histology Figure 2 depicts a reduced (p < 0.05) height of the epiphyseal
growth-plate caused by the boron and/or vitamin D injections. This decrease in growth plate height was similar to the growth plate of a normal (vitamin D-adequate) chick (Fig. 2). Analysis of the individual zones of the growth plate reveals that the difference in the overall plate height was accounted for by the effect of treatments in the proliferative and hypertrophic zones. The height of the proliferative zone exhibited the same pattern as the overall plate length with the treated groups having a shorter (p < 0.05) zone (Fig. 2). The changes in the hypertro- phic zone were less dramatic with groups containing vitamin D 3 hav- ing a smaller zone (p < 0.05) than the - D (vitamin D-deficient control), but the B-treated group was not significantly different from the other treatments.
Plasma
Ionized and bound Ca were similar among all of the treatments (Table 4). Total Ca was higher (p < 0.05) for B treatment than for the other treatments. Plasma Na § K +, Mg, and Pi w e r e not significantly different among treatments.
Biological Trace Element Research Vol. 31, 1991

In Ovo Boron Supplementat ion
90
70
-~ 50
3O
lO
a
L J J , !
~ x x
-;-;-1 ,.-..] abe . ,
Z ";';'1 bo ",','~ - -
T r e a t m e n t
ab - - [ ] -D (n=31)
[ ] B (n=24)
[ ] B+D (n=21)
[ ] 0.3D (n=15)
f/,~/~,,~/~ [ ] 1.5D (n=22)
Fig. 1. The effect of in ovo B and/or vitamin D 3 ad- ministration on hatchability of vitamin D-deficient chicken embryos . Values are the mean + SEM. Bars with no letters in common differ significantly (p < 0.05) from each other.
Effect of In Ovo B on Tibial Characteristics
Table 3 and/or Vitamin D~ Administrat ion of 1-D-Old Vitamin D-Deficient Chicks"
229
Width, Trea tment n Weight, mg ~' Length, mm mm Ash, %
- D 12 59.74 _+ 2.65' 30.6 + 0.3' 1.5 + 0.0' 28.69 _+ 0.91 ~ B 5 59.15 _+ 2.16' 30.5 + 0.1' 1.4 + 0.0' 29.56 _+ 0.87 -~,~ B + D 15 62.03 + 1.56' 31.0 + 0.3 ~ 1.4 + 0.0' 33.00 +_ 0.6@ 0.3D 5 60.80 _+ 3.15' 30.6 +_ 0.4' 1.5 _+ 0.0' 32.02 _+ .042 ~.z 1.5D 12 61.36 + 2.52' 30.2 + 0.4' 1.5 _+ 0.0' 31.58 _+ 0.61 ~.~-
"Data are mean _+ SEM. Means fo l lowed by different superscr ipt n u m b e r s wi thin a co lumn differ s ignif icantly (p < 0.05; D u n c a n ' s Mul t ip le-Range Test).
~'Dried, defat ted left tibia.
DISCUSSION
O t h e r e x p e r i m e n t s c o n d u c t e d in this l a b o r a t o r y h a v e s u g g e s t e d a b o r o n - d o s e r e s p o n s e , w i t h 0.5 m g o f b o r o n p r o v i d i n g o p t i m a l r e s p o n s e (28). T h e r e f o r e , 0.5 m g of b o r o n was c h o s e n as the b o r o n d o s e to de t e r - m i n e w h e t h e r a v i t a m i n D - b o r o n i n t e r a c t i o n exists . Based o n the a m o u n t o f v i t a m i n D in a c h i c k e n egg , 1.5 Dg of v i t a m i n D w a s c o n s i d e r e d " a d e q u a t e , " w h e r e a s 0.3 p.g w a s a " l o w " d o s e of v i t a m i n D.
A 25-27% b o n e a s h v a l u e is c o n s i d e r e d rachi t ic b y the A s s o c i a t i o n of Official A g r i c u l t u r e C h e m i s t ( A O A C ) ch ick a s say (37). T w e n t y - e i g h t pe r - c en t b o n e a sh for the - D ( v i t a m i n D-de f i c i en t con t ro l ) s u p p o r t e d a v i t a m i n D d e f i c i e n c y in the e m b r y o s u s e d in this s t u d y . T h e poss ib i l i ty of
Biological Trace Element Research VoL 31, 1991
230 King et aL
:ta I I E~, 3 I~ 1
. . , r
|
-D (6)
c c
i
NA
</,,'A B B+D 0.3D
(3) (6) (s)
T r e a t m e n t (n)
c c
I;-,x///)'.,~
z.///, r / / / , ; 1.5D Normal (6) (s)
[ ] Reserve Zone
[ ] Proliferative Zone
[ ] Hypertrophic Zone
Fig. 2. The effect of in ovo B and/or vitamin D3 administra- tion on the height of the epiphyseal plate zones of 1-d-old vitamin D-deficient chicks. The heights of the growth-plate and prolifera- tive zone differ significantly (p < 0.05) for those bars with no letter in common. The hypertrophic zone is shorter (p < 0.05) with B + D, 0.3 D, and 1.5D treatments than the corresponding zone in the - D treatment (vitamin D-deficient control).
a bo ron-v i t amin D interaction was sugges ted by the increased hatch- ability and percent bone ash for the B + D group. Fur thermore , the B group, a l though not significantly greater than - D, exhibited an increase in bone ash close to the values of the vi tamin D-supp lemen ted groups. This sugges ts that boron may have interacted with the m i n u t e a m o u n t of vi tamin D available, pe rhaps ex tend ing its half-life or aiding in its hy- droxylation, as has been previously specula ted by Hun t (8,11).
The g rowth plate has intracellular receptors for 24,25- and 1,25- dihydroxyvitamin D3, localized in the proliferative and u p p e r part of the hyper t rophic zones (38). The role of vi tamin D in growth-pla te morphol - ogy is bel ieved to be main tenance of se rum Ca and P levels, facilitating matrix calcification (39,40). During vi tamin D deficiency, the result ing hypocalcemia and h y p o p h o s p h a t e m i a may h inder matrix calcification. Consequent ly , the chondrocytes in the proliferative and hyper t roph ic zones are not able to mature to the next phase, render ing a larger g rowth plate because of an increase in the he ight of these zones (38, 41). The - D (vitamin D-deficient control) chicks exhibited these lesion characteristics.
The decrease (p < 0.05) in both proliferative and hyper t roph ic zones with boron and/or vi tamin D 3 adminis t ra t ion suggests that vi tamin D and boron may have a positive interaction. Further, boron, adminis te red alone, r educed or ameliorated the effects of vi tamin D deficiency evi-
Biological Trace Element Research Vol. 31, 1991

in Ovo Boron Supplementation
Table 4 Effect of In Ovo B and/or Vitamin D3 Administrat ion
on Plasma Calcium of 1-D-Old Vitamin D-Deficient Chicks"
231
Treatment Ionized Ca, mmol/L Bound Ca, mmol/L" Total Ca, mmol/L
- D 1.29 _+ 0.03 ' 0 .40 -+ 0.20 ' 1 .42 + 0.132 n = t 3 n = 5 n = 12
B 1.49 • 0 .12 ' - 2 .33 • 0 .40 ' n = 2 n = 2
B + D 1.34 + 0.04 ' 0 .37 • 0 .12 ' 1 .52 + 0.162 n = 8 n = 2 n = 1 3
0 . 3 D 1.39 _+ 0.05' - 0 .86 -+ 0.99 ~ n = 3 n = 4
1 .5D 1.36 + 0 .04 ' 0 .37 + 0 .16 ' 1 .43 • 0 .15 -~ n = 9 n = 3 n = 7
"Data are mean _+ SEM. M e a n s fo l lowed by a d i f fe ren t supe r sc r ip t n u m b e r wi th in a co lumn differ s ignif icant ly (p < 0.05; D u n c a n ' s Mul t ip le -Range Test).
~'Bound Ca = total Ca - ion ized Ca.
denced by a smaller (p < 0.05) proliferative zone, and a qualitative decrease in the hypertrophic zone height.
The results of this s tudy concur with those previously reported by Hunt (13), providing further evidence that boron and vitamin D may interact. Boron's involvement in bone metabolism is suggested by the results; however , the character of its role is not as yet well defined. Whether vitamin D deficiency augments the need for boron (8), or boron insufficiency aggravates vitamin D deficiency remains to be elucidated. It is possible that the interaction may be expressed through another metab- olite, such as Ca, P, Mg, or others. Furthermore, the results obtained in studies utilizing vitamin D-adequate chicks (13,28) have suggested that a fine line exists be tween the positive and negative effects of boron supple- mentat ion dependen t on vitamin D nutriture. These results indicate that, al though boron supplementat ion could be of some benefit for certain parts of the world population, it may not be for others.
ACKNOWLEDGMENTS
The authors are grateful to Ruth Albright and Deborah K. McGuinn for the technical assistance provided, and to Lori White for typing this manuscript.
REFERENCES
1. H. Agulhon, Ann. Inst. Pasteur 34, 321 (1910). 2. K. Warington, Ann. Bot. 37, 629 (1923). 3. E. Hove, C. A. Elvehjem, and E. B. Hart, Am. J. Physiol. 127, 689 (1939).
Biological Trace Element Research VoL 31, 1991

232 King et aL
4. E. Orent-Keiles, Proc. Soc. Exp. Biol. Med. 44, 199 (1940). 5. J. D. Teresi, E. Hove, C. A. Elvehjem, and E. B. Hart, Am. ]. Physiol. 140, 513
(1944). 6. J. T. Skinner and J. S. McHargue, Am. J. Physiol. 143, 385 (1945). 7. R. H. Follis, Am. J. Physiol. 150, 520 (1947). 8. C. D. Hunt and F. H. Nielsen, Trace Element Metabolism in Man and Animals,
J. McC. Howell, J. M. Gawthorne, and C. L. White, eds., Springer-Verlag, Berlin, Heidelberg, New York, pp. 597-600 (1982).
9. F. H. Nielsen, T. R. Shuler, T. J. Zimmerman, and E. O. Uthus, Biol. Trace Element Res. 17, 91 (1988).
10. F. H. Nielsen, T. R. Shuler, T. J. Zimmerman, and E. O. Uthus, May, nesium 7, 133 (1988).
11. C. D. Hunt and F. H. Nielsen, Trace Elements in Man and Animals, vol. 6, L. S. Hurley, C. L. Keen, R. B. L6nnerdal, and R. B. Rucker, eds., Plenum, New York, pp. 275-276 (1988).
12. C. D. Hunt, Biol. Trace Min. Res. 22, 201 (1989). 13. C. D. Hunt, FASEB J. 4, A645 (1990). 14. F. H. Nielsen, Annu. Rev. Nutr. 4, 21 (1984). 15. F. H. Nielsen, Nutr. Rev. 46, 337 (1988). 16. F. H. Nielsen, Nutr. Today Jan/Feb, 4 (1988). 17. C. A. Zittle, Adv. Enzymol. 12, 493 (1951). 18. M. Audran, Mayo Clin. Proc. 60, 851 (1985). 19. C. D. Hunt, FASEB J. 3, A760 (1989). 20. M. Hegsted, M. J. Keenan, F. Slyer, and P. Wozniak, Biol. Trace Min. Res.
28, 243 (1991). 21. F. H. Nielsen, C. D. Hunt, L. M. Mullen, and J. R. Hunt, FASEB J. 1, 394
(1987). 22. M. A. Bock, M. Powey, and M. Ortiz, FASEB ]. 4, A520 (1990). 23. R. E. Newnham, Trace Element Metabolism in Man and Animals, J. McC.
Howell, J. M. Gawthorne, and C. L. White, eds., Springer-Verlag, Berlin Heidelberg, New York, pp. 400-402 (1982).
24. M. A. Elliot, and H. M. Edwards, Proceedings of the Southern Poultry Science Society, Eleventh Meeting, Atlanta, GA, pp. 14 (1990).
25. T. F. Brown, M. E. McCormick, D. R. Morris, and L. K. Zeringue, Nutr. Res. 9, (1989).
26. S. L. Pardue, T. W. Odom, N. King, and H. W. Sampson, FASEB J. 3, A1072 (1989).
27. S. L. Pardue, S. H. Frappier, C. Garza, N. King, and T. W. Odom, Poult. Sci. 68 {Suppl. 1), 196 (1989).
28. N. King, T. W. Odom, H. W. Sampson, and S. L. Pardue, Biol. Trace Element Res. (in press).
29. Subcommittee on Poultry Nutrition, Committee on Animal Nutrition, Board on Agriculture, National Research Council, Nutrient Requirement of Poultry 8th revised ed. National Academy Press, Washington DC, (1984).
30. L. E. Hart, H. K. Schnoes, and H. F. DeLuca, Arch. Biochem. Biophys. 250, 426 (1986).
31. J. H. Soares, M. R. Swerdel, and M. A. Ottinger, Poult. Sci. 58, 1004 (1979). 32. M. L. Sunde, C. M. Turk, and H. F. DeLuca, Science 200, 1067 (1978). 33. Analytical Methods for Atomic Absorption Spectrophotometry, 0303-0152. Perkin-
Elmer, Norwalk, CT (1982). 34. SAS Proprietary Software Release 6.03, SAS Institute, Cary, NC (1987).
Biological Trace Element Research VoL 31, 1991

in Ovo Boron Supplementation 233
35. R. G. D. Steel and J. H. Torrie, Principles and Procedures of Statistics, McGraw- Hill, New York, NY (1960).
36. D. B. Duncan, Biometrics 11, 1 (1955). 37. Official Methods of Analysis, S. Williams, ed., Association of Official Analyti-
cal Chemists, VA (1984). 38. J. P. Iannotti, Orthop. Clin. North Am. 21, 1 (1990). 39. S. S. Jande and I. R. Dickson, Acta Anat. 108, 463 (1980). 40. P. H. Long, S. R. Lee, G. N. Rowland, and W. M. Britton, Avian Dis. 28, 933
(1984). 41. R. M. Leach and C. V. Gay, J. Nutr. 117, 784 (I987).
Biological Trace Element Research VoL 31, 1991





























