Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Estuarine, Coastal and Shelf Science 115 (2012) 260e271
Contents lists available
Estuarine, Coastal and Shelf Science
journal homepage: www.elsevier .com/locate/ecss
The effect of salinity increase on the photosynthesis, growth and survival of theMediterranean seagrass Cymodocea nodosa
José M. Sandoval-Gil a,b, Lázaro Marín-Guirao a, Juan M. Ruiz a,*
a Seagrass Ecology Group, Instituto Español de Oceanografía, Centro Oceanográfico de Murcia, C/Varadero 1, 30740 San Pedro del Pinatar, Murcia, SpainbDepartamento de Ciencias del Mar y Biología Aplicada, Universidad de Alicante, Alicante, P.O. Box 99, 03080 Alicante, Spain
a r t i c l e i n f o
Article history:Received 20 April 2011Accepted 22 September 2012Available online 4 October 2012
Keywords:seagrassCymodocea nodosaphotosynthesishypersaline stressmesocosm
* Corresponding author.E-mail addresses: [email protected] (J.M
mu.ieo.es (J.M. Ruiz).
0272-7714/$ e see front matter � 2012 Elsevier Ltd.http://dx.doi.org/10.1016/j.ecss.2012.09.008
a b s t r a c t
There are major concerns in the Mediterranean Sea over the effects of hypersaline effluents fromseawater desalination plants on seagrass communities. However, knowledge concerning the specificphysiological capacities of seagrasses to tolerate or resist salinity increases is still limited. In this study,changes in the photosynthetic characteristics, pigment content, leaf light absorption, growth and survivalof the seagrass Cymodocea nodosa were examined across a range of simulated hypersaline conditions. Tothis end, large plant fragments were maintained under salinities of 37 (control ambient salinity), 39, 41and 43 (practical salinity scale) in a laboratory mesocosm system for 47 days. At the end of the exper-imental period, net photosynthesis exhibited a modest, but significant, decline (12e17%) in all testedhypersaline conditions (39e43). At intermediate salinity levels (39e41), the decline in photosyntheticrates was mainly accounted for by substantial increases in respiratory losses (approximately 98% of thecontrol), the negative effects of which on leaf carbon balance were offset by an improved capacity andefficiency of leaves to absorb light, mainly through changes in accessory pigments, but also in opticalproperties related to leaf anatomy. Conversely, inhibition of gross photosynthesis (by 19.6% compared tothe control mean) in the most severe hypersaline conditions (43) reduced net photosynthesis. In thistreatment, the respiration rate was limited in order to facilitate a positive carbon balance (similar to thatof the control plants) and shoot survival, although vitality would probably be reduced if such metabolicalterations persisted. These results are consistent with the ecology of Mediterranean C. nodosa pop-ulations, which are considered to have high morphological and physiological plasticity and a capacity togrow in a wide variety of coastal environments with varying salinity levels. The results from this studysupport the premise that C. nodosa has a higher tolerance to hypersaline conditions than the highlysensitive Posidonia oceanica, the other dominant Mediterranean seagrass, which is limited to marineenvironments with stable salinities.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Seagrass meadows represent the dominant habitat in marineinfralittoral environments of the Mediterranean Sea (Procacciniet al., 2003). The ecological and socio-economical relevance ofthese seagrass communities has been widely documented andrecognised worldwide, as has their high vulnerability to a variety ofanthropogenic disturbances concentrated in the coastal zone(Kenworthy et al., 2006; Short et al., 2007). In the MediterraneanSea, hypersaline plumes (i.e., brines) from seawater desalinationplants are recent human impacts that potentially threaten the
. Sandoval-Gil), juanm.ruiz@
All rights reserved.
conservation of seagrass habitats (Sánchez-Lizaso et al., 2008;Boudouresque et al., 2009; Ruiz et al., 2009a,b; Palomar andLosada, 2010). In effect, increases in seawater salinity have beenshown to alter seagrass communities in this coastal zone and others(Chesher, 1975; Tomasko et al., 2000; Fernández-Torquemada et al.,2005a; Gacia et al., 2007; Ruiz et al., 2009a,b). Consequently, anunderstanding of the mechanisms seagrass species employ totolerate salinity increases is crucial for forecasting and monitoringthe consequences of brine impacts on these vulnerable and valu-able ecosystems.
However, despite the fact that salinity is a major ecologicalfactor influencing seagrass distribution and structure worldwide(Chesher, 1975; Montague and Ley, 1993; Castriota et al., 2001;Einav et al., 2002; Fernández-Torquemada et al., 2005b; Gaciaet al., 2007), salinity tolerance ranges have only been established

J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271 261
for a few seagrass species. Furthermore, the structural and physi-ological mechanisms by which seagrasses cope with natural oranthropogenically induced changes in salinity regimes are stillpoorly understood (Drew, 1978a; Tyerman, 1989; for a review, see;Touchette, 2007).
The acquisition of particular adaptations in order to maintainosmotic equilibrium (e.g., osmoregulation) and key physiologicalfunctions (e.g. photosynthesis) is one of the basic propertiesenabling seagrasses to successfully evolve in marine environments(Arber, 1920; Kuo and Den Hartog, 2000). The physiologicalcapacity of seagrasses to tolerate increases in salinity is speciesspecific and closely related to the salinity characteristics of theenvironments in which they grow. Species limited to coastal envi-ronments with constant salinity levels are considered to be lesstolerant to changes in this parameter thanmore euryhaline species,which are able to grow in a broader range of coastal habitats withwidely different salinity levels (Tyerman, 1989; Kuo and DenHartog, 2000; Koch et al., 2007; Touchette, 2007). These twoextremes are represented by the distinct ecologies of the two mostcommon Mediterranean seagrasses, Posidonia oceanica (L.) Delileand Cymodocea nodosa (Ucria) Ascherson, and their differentialtolerances to hypersaline stress.
The distribution of the larger, slow-growing Posidonia oceanicais usually limited to open coastal waters with virtually constantsalinities (e.g., 36.5e38 on the southeastern coast of Spain: Ruizet al., 2009a,b; but see Meinesz et al., 2009; Tomasello et al.,2009). Conversely, the smaller, fast-growing Cymodocea nodosa iscommonly found in a wider range of marine environments withvarying salinity levels (den Hartog, 1970; Drew, 1978b; Procacciniet al., 2003; Boudouresque et al., 2009). Dense and highlyproductive C. nodosa meadows are common in open coastal waterswith constant salinities, estuaries with fluctuating salinities andtemperatures, and confined hypo- and hypersaline water bodiesalong the Mediterranean coast. Accordingly, C. nodosa has greatermorphological and functional plasticity than P. oceanica and hasbeen recognised as being a more eurybiontic species, betteradapted than P. oceanica to cope with changes in environmentalconditions (Drew, 1978b; Terrados and Ros, 1991; Pérez andRomero, 1994; Cebrián et al., 2000; Marbá and Duarte, 2001;Cancemi et al., 2002; Olesen et al., 2002; Agostini et al., 2003).
Thus, based on previous knowledge, it is expected that Cymo-docea nodosa meadows are more tolerant to natural or anthro-pogenically induced salinity increases than those of Posidoniaoceanica. Results obtained from recent studies are in agreementwith this general hypothesis. Plant growth, biomass andmortality inP. oceanica have been shown to be highly sensitive to very smallsalinity increases (39) above mean ambient values (Fernández-Torquemada and Sánchez-Lizaso, 2005; Gacia et al., 2007; Ruizet al., 2009a,b), while these same plant variables remained unaf-fected in C. nodosa at much higher salinity thresholds (greater than41) (Fernández-Torquemada and Sánchez-Lizaso, 2006; Pagès et al.,2010). This marked inter-specific difference in resisting hypersalinestress is likely to be determined by the physiological adaptivemechanisms involved in the capacity of each species to tolerate oravoidosmotic and ionic stress (e.g. Kramer andBoyer,1995;Verslueset al., 2006). The nature of such mechanisms remains unclear inseagrasses in general (Touchette, 2007) and has been virtuallyignored in the specific case of Mediterranean seagrasses. Amongthese mechanisms, photosynthesis is one of the most important,being generally affected by hypersaline conditions (Touchette,2007), although information on this topic is scarce and only avail-able for a few seagrass species. The limited evidence availableindicates that hypersalinity can reduce photosynthetic performancein seagrasses (and hence their growth and survival rates) by alteringthe structure of their photosynthetic apparatus (Iyer and Barnabas,
1993), its capacity to harvest light (McMillan and Moseley, 1967;Ralph, 1998) and its PSII photochemical efficiency (Ralph, 1998,1999; Koch et al., 2007), and/or by altering photosynthetic carbonmetabolism (e.g. Beer et al., 1980). Obtaining further informationregarding the effects of hypersalinity on the photosynthetic physi-ology of Mediterranean seagrass species is therefore crucial tounderstanding their specific tolerances to increased salinity, andalso to establishing scientific and technical criteria for preventingand controlling human impacts involving changes in seawatersalinity (e.g. desalination plants; Palomar and Losada, 2010).
In this study, we examined for the first time: (a) the behaviour ofthe photosynthetic physiology of Mediterranean Cymodocea nodosapopulations in response to sudden and chronic salinity increases,simulated in a laboratory mesocosm system, and (b) the relation-ship between leaf carbon balance and growth and shoot survival. Tothis end, pigment content (Chla, Chlb and total carotenoids), leafoptical properties, maximum photochemical efficiency (Fv/Fm),photosynthesiseirradiance curves (i.e. photosynthetic parameters),leaf growth and shoot features and survival were measured forC. nodosa plants under ambient salinity (i.e. 37) and compared withmeasurements performed on plants simultaneous exposed for 45days to various degrees of chronic salinity increase. This studyformed part of a broader experiment aimed at analysing thephysiological responses of Mediterranean seagrass species tosalinity increases, which simultaneously involved both Posidoniaoceanica and C. nodosa in order to provide reliable comparisonsbetween these species. Only data regarding C. nodosa photosyn-thesis (and respiration) are presented here, which are discussed inrelation to previously published data obtained during the sameexperiment for P. oceanica (Marín-Guirao et al., 2011).
2. Materials and methods
2.1. Field plant collection and mesocosm system
In October 2008, scuba divers collected large Cymodocea nodosafragments (shoots connected to basal rhizomes with intact rootsystems) from a dense, shallow meadow (at a depth of 5e6 m)colonising infralittoral sediments on the southeastern coast ofSpain (Isla Plana, Murcia Region; 37� 340 20.8600 N, 1� 120 28.1600 W).This locality was characterised by highly oligotrophic waters withconstant salinity (37e38 psu-Practical Salinity Scale) and seasonalfluctuations in downward irradiance (5.3e19.8 mol quanta day�1)and temperature (14.6e24.8 �C) (Ruiz et al., 2009a,b; unpublishedresults), which are typical conditions for shallow infralittoral habi-tats of the Spanish Mediterranean coast (Margalef, 1989; VargasYánez et al., 2010).
The plants were transported in large coolers to the laboratory,arrivingwithin 4 h of collection. Here theywere introduced into theaquaria of the mesocosm system for acclimatisation (see below).Five plant fragmentswereplaced in aplastic basket (22�40cmbaseand 10 cm in height) filled with coarse sediments, which had beenpreviously washed to remove animals and large organic particles.This represented the experimental unit (EU; Fig. 1). All plant frag-ments included the apical meristem and were carefully selected tohave comparable basal rhizomes, with similar lengths (35e45 cm)and numbers of shoots (5e10), in order to standardise the experi-ment. The overall number of shoots per EU was between 40 and 50,with a total of 24 EUs being prepared. Six EUs were randomlydispersed throughout the cross-section of each large aquarium ofthe mesocosm system (see below), together with EUs containingPosidonia oceanica fragments, which were simultaneously exam-ined in the same experiment (Marín-Guirao et al., 2011).
The mesocosm (Fig. 1) consisted of four large (1500 L), inde-pendent aquaria, each with an autonomous circuit of circulating

Fig. 1. General view of the mesocosms system (left) and C. nodosa in the experimental units (right).
J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271262
seawater and independent control of water flow, temperature,irradiance, pH and nutrients (for more details, see Marín-Guiraoet al., 2011). Illumination for each aquarium was provided bythree 400 W metal halide lamps (Aqualight-400). Temperatureswere controlled by a precise (�0.1 �C) automated system con-structed ad hoc for this and other mesocosm systems in our labo-ratory facilities. The seawater circulatory system permittedseparate control of water flow rates within each aquarium, whichwere adjusted to provide continuous seawater circulation andfiltration, and optimum seagrass photosynthesis (Enríquez andRodríguez-Román, 2006). Daily pH cycles were continuouslymonitored (Marín-Guirao et al., 2011). All aquaria exhibited regulardaily pH variations between 8.02 and 8.18, with maximum valuesoccurring during the light period and minimum values during thedark period, as a consequence of the photosynthetic activity of theseagrass. No significant differences were found in pH betweenaquaria throughout the experimental period (one-way ANOVA,p > 0.05), with the values being similar to those observed inCymodocea nodosa canopies at similar depths to the plant collectionsite (Invers et al., 1997).
The mesocosm circuits were filled with high-quality naturalseawater obtained from a nearby oligotrophic and unpollutedmarine protected area (Cabo de Palos-Hormigas Islands MarineReserve; 37� 370 57.800 N, 0� 410 21.400tiv W). Special care was takenwith seawater quality, which was controlled through continuousphysical and chemical filtration (Marín-Guirao et al., 2011). Thefiltration systems were cleaned daily and the seawater from themesocosm was partially replaced every week. Periodic analyses(approximately every 15 days) revealed that dissolved nutrient(nitrate and phosphate) concentrations were maintained withinlevels typically reported for oligotrophic coastal waters off thesoutheastern coast of Spain (unpublished results).
As explained here and in a previous related work (Marín-Guiraoet al., 2011), the precise control of environmental parameterswithin the mesocosm system was one of our major concerns. Theaimwas: (a) to maintain spatial homogeneity of such factors withinand between the aquaria; (b) to simulate an average environmentalbackground representative of that experienced by the plants innatural conditions, and (c) to avoid artificial conditions that couldcause additional stress for the plants. Previous and current exper-iments have demonstrated that this mesocosm system cansuccessfully maintain healthy plants of Cymodocea nodosa andPosidonia oceanica for periods of up to 4 months, with almost 100%survival and performances similar to those observed under naturalconditions. This is noteworthy, considering that performing suchexperiments under laboratory conditions is not feasible for mostseagrass species, especially the larger ones. Investigations involvingsuch species are usually undertaken in small aquaria on plantfragments of restricted size, which means the plants can only be
kept healthy for periods of about 2 or 3 weeks (e.g., Fernández-Torquemada and Sánchez-Lizaso, 2005; Cayabyab and Enríquez,2007; Pagès et al., 2010). This clearly limits the temporal scale ofexperimental manipulations and plant responses (Oksanen, 2001).In order to assess possible additional stress caused by the artificialconditions, measurements from the aquaria were compared withthose obtained simultaneously in the natural meadow (see Section2.7 and Table 6).
2.2. Experimental design
For plant acclimatisation, the EUs were maintained in theaquarium for 1 week prior to the start of the experimental treat-ments, under environmental conditions similar to those experi-enced by the natural population: a temperature of 21 � 0.1 �C,a salinity of 37 � 0.1 and a saturating irradiance of ca.300� 30 mmol quantam�2 s�1 (Marín-Guirao et al., 2011). A 12/12 hphotoperiod was selected, in order to provide a daily light exposureof 12.9 mol quanta m�2 day�1, which was close to that obtained byaveraging daily integrated light curves recorded in the field at thetop of the meadow leaf canopy (Marín-Guirao et al., 2011).
Following the acclimatisation period, salinity levels wereadjusted to obtain the experimental treatment selected for eachaquarium, while all other parameters (light, temperature, pH,nutrients and water flow) were kept constant. To this end, oneaquarium was maintained at ambient salinity (i.e., 37; the controltreatment), while salinity was increased in the other three aquar-iums to obtain hypersaline treatments of 39, 41 and 43. Salinitytreatments were randomly assigned to each large aquarium. Thesesalinity levels were achieved by adding high quality marine salt(Seachem Salt�) following the protocol described by Marín-Guiraoet al. (2011). These experimental salinity levels encompassed themeasured range experienced in the Mediterranean by Cymodoceanodosa under natural conditions (36.5e38; Sandoval-Gil et al.,2012) and when exposed to brine discharges (Fernández-Torquemada et al., 2005a; Fernández-Torquemada and Sánchez-Lizaso, 2006). The experimental conditions were maintained fora period of 47 days. We considered both the intensity and durationof the experiment to be sufficient to allow specific plant stressresponses, but not so great as to induce severe metabolic impair-ment and/or damage.
All plant response variables were measured at the end of theexperimental period. For each variable, except number of shoots(see below for the variable types), the average of two or threemeasurements performed on leaves from different shoots providedthe value used for each EU, which we considered the replicated unitin this experiment. Within EUs, each leaf came from a differentshoot, collected from a different plant fragment. Only healthyshoots (i.e. without wounds or herbivore damage) of a similar age

J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271 263
(3e5 years old) were selected. We avoided collection of the youn-gest (i.e. those closest to the apical shoot) and neighbouring shootswithin the same plant fragment, to circumvent any effects of spatialdependency due to internal resource gradients between sinks (i.e.apical, pioneer ramets) and source individuals (Marbá et al., 2002;Olivé et al., 2009). For each shoot, measurements and analyses werealways performed on mature photosynthetic tissues of leaf ranknumber 1 or 2.
In such an experimental design, differences in plant variablesbetween treatment levels may be subject to a risk of confounding,since all EUs from a particular treatment level were placed in thesame large aquarium (Quinn and Keough, 2002). In the presentstudy, this approach was unavoidable for several reasons: (a) theuse of smaller aquaria or tanks and plant fragments was notappropriate for the timescale and species examined; (b) replicationof such large aquaria has obvious economical and logisticalconstraints, and (c) other alternatives, such as field experiments,can be evenmore complex, since themanipulation of salinity in situis particularly complicated (e.g. Ruiz et al., 2009a,b). Given thesecircumstances, and based on our knowledge of the specific bio-logical and experimental systems, we followed the recommenda-tions of Oksanen (2001) and Quinn and Keough (2002), in order toidentify, control and reduce the probability that local factors otherthan the treatments would confound the effects of experimentaltreatments between the aquaria. In this sense, as explained above,the efficient control of environmental parameters within themesocosm system provided us confidence that factors other thansalinity did not differ between the aquaria. In addition, to avoidpossible influences of spatial environmental micro-gradients, therelative positions of the EUs within the aquaria were regularlychanged (approximately twice a week). Furthermore, some plantvariables (Fv/Fm, P vs. E curves, pigment content and leaf growth)measured during the acclimatisation period exhibited no signifi-cant differences between aquaria (ANOVA, p > 0.05). Thus, we areconfident that differences between treatments at the end of theexperimental period were due to divergence in time trajectoriesimposed by the treatments from the onset of the experiment andnot to pre-existing differences.
2.3. Chlorophyll fluorescence
Measurements of chlorophyll a fluorescence emissions wereperformed using a diving-PAM portable fluorometer (Walz,Germany) on dark-adapted leaves in order to calculate themaximum quantum yield of PSII (Fv/Fm), which representsa measure of the photochemical efficiency of the PSII with all reac-tion centres open (Schreiber, 2004; Larkum et al., 2006). Allmeasurements were performed on plants adapted to darknessovernight (i.e. before switching on the illumination system) toensure full oxidation of the reaction centres and primary electronacceptors. For the Fv/Fm measurements, minimum (F0) andmaximum (Fm) fluorescence were measured after exposing thephotosystems of dark-adapted leaves, pre-illuminated with a weakred modulated light, to a saturating pulse (0.8 s) of white light.During fluorescence determinations, each leaf was held in a DCL-8leaf-clip holder to maintain a constant distance between the leafand the fibre optic probe. Prior to the experiment, fluorescenceparameters measured along Cymodocea nodosa leaves confirmedthe typical variation pattern as a function of leaf age, as previouslydescribed for another seagrass species, Thalassia testudinum(Durako and Kunzelman, 2002; Enríquez et al., 2002). Since thissource of variation could mask possible treatment effects, fluores-cence measurements were performed at regular 1 cm intervals,from leaf base to apex, for all leaves present on a shoot (2e3 leavesper shoot). From this set of measurements, the maximum Fv/Fm
value was selected to represent the whole shoot. These measure-ments were performed on three randomly selected shoots fromeach EU on each sampling occasion (N¼ 6 replicates per treatment).Since the diving-PAM fluorometer allowed in vivo measurementswithout altering the state of the plants (Beer et al., 2001), thesemeasurements were performed once a week to assess variations inmean Fv/Fm values throughout the experimental period.
2.4. Respiration and photosynthetic rates
Photosynthesis-irradiance (P vs. E) curves for Cymodocea nodosaleaves were determined, following Walker (1987) and Cayabyaband Enríquez (2007), for two shoots from three of the six EUs,randomly selected from each aquarium (N ¼ 3 replicates pertreatment). Respiration and photosynthetic rates were measuredpolarographically, using a DW3 chamber with a Clark-type O2electrode system (Hansatech, UK) connected to a controlled-temperature circulating bath (P-Selecta, Spain). The incubationchamber was filled with filtered seawater and maintained at thesame temperature as the aquaria (21 �C). Leaf segments ofapproximately 0.6 cm2 from the middle portion of the first matureleaf were used for photosynthesis measurements on each shoot.This leaf portion represented the maximum Fv/Fm values, aspreviously measured on C. nodosa shoots.
Leaf segments were first incubated in darkness for 15 min, todetermine the initial dark respiration rate (initial-Rd), and thenexposed to increasing light intensities (E): 14, 20, 37, 66, 84, 143,250, 654, 1010, 1496 and 2271 mmol quanta m�2 s�1. After the finallight exposure, leaf segments were re-exposed to darkness, in orderto determine the final dark respiration rate (final-Rd). The leafsegments were illuminated using a tungsten-halogen light source(LS2; Hansatech, UK) and light intensities within the chamber werecalibrated using the cosine-corrected quantum sensor of thediving-PAM fluorometer. The seawater in the incubation chamberwas supplemented with 5 mM NaHCO3, to prevent carbon limita-tion, and N2 was bubbled through the chamber to maintain oxygenconcentrations within the 20e80% saturation range. Oxygen read-ings were continuously recorded on a PC using a transducer unit(Oxygraph; Hansatech, UK) connected to the electrode.
Oxygen variations were determined for each incubation intervalas mmol O2 cm�2 h�1 and plotted against their respective E values toconstruct a P vs. E curve, which exhibited the typical saturationkinetics of the photosynthetic response. The maximum rate of netphotosynthesis (net-Pmax) was determined by averaging themaximum values above the saturating irradiance (Ek). Grossphotosynthesis (gross-Pmax) was calculated as the sum of net-Pmaxand final-Rd. Photosynthetic efficiency (a, mmol O2 cm�2 h�1/mmol quanta m�2 s�1) was calculated as the slope of the regressionline fitted to the initial linear part of the P vs. E curve. Compensationirradiance (Ec) was obtained from the intercept of this regressionline with the X-axis. The Ek was calculated as the ratio Pmax/a. TheP:Rd ratio was used as a proxy for the daily leaf carbon balance, Pbeing calculated by multiplying gross-Pmax by the number of lighthours (i.e., 12), with Rd being the total respiration over a 24-hperiod, using final-Rd for the light period (12 h) and initial-Rd forthe dark period (12 h).
2.5. Pigment content and leaf optics
Pigment content measurements were performed on two shootsfrom each EU (N ¼ 6 replicates per treatment). The leaf portionsexamined were selected using the same criteria adopted for the Pvs. E curves (i.e. maximum Fv/Fm values). The pigments wereextracted from 0.8 cm2 leaf segments using 80% acetone, withMgCO3 solution added to prevent acidification of the extract
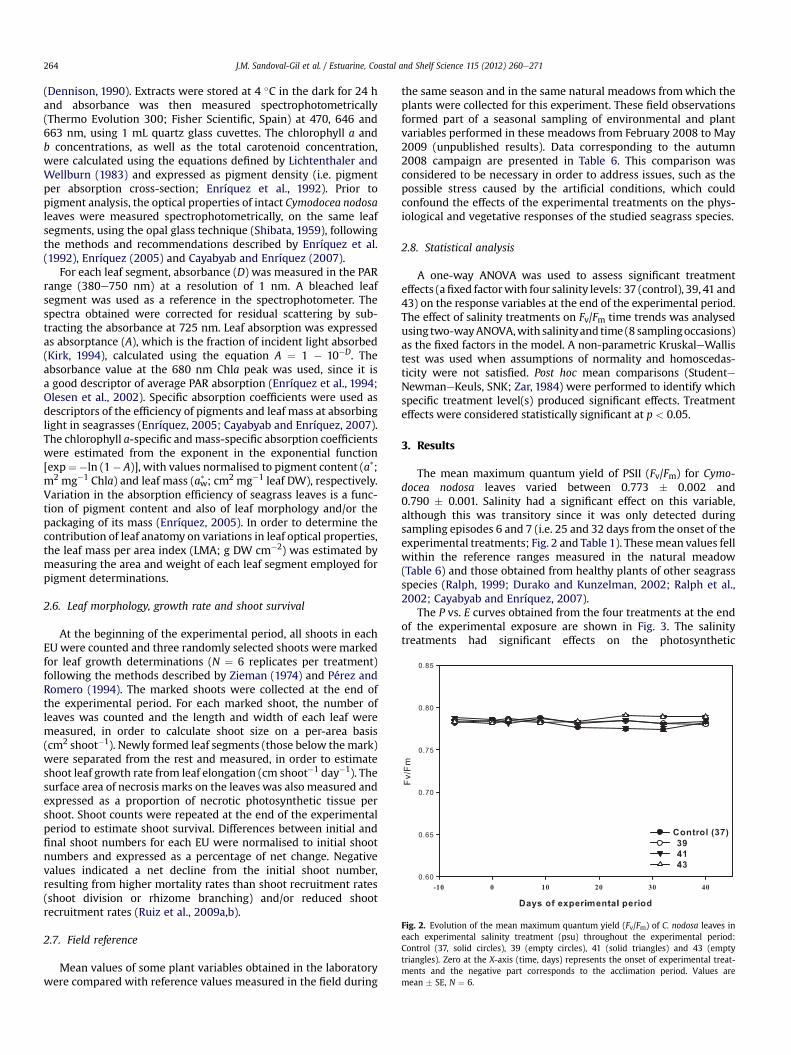
Fig. 2. Evolution of the mean maximum quantum yield (Fv/Fm) of C. nodosa leaves ineach experimental salinity treatment (psu) throughout the experimental period:Control (37, solid circles), 39 (empty circles), 41 (solid triangles) and 43 (emptytriangles). Zero at the X-axis (time, days) represents the onset of experimental treat-ments and the negative part corresponds to the acclimation period. Values aremean � SE, N ¼ 6.
J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271264
(Dennison, 1990). Extracts were stored at 4 �C in the dark for 24 hand absorbance was then measured spectrophotometrically(Thermo Evolution 300; Fisher Scientific, Spain) at 470, 646 and663 nm, using 1 mL quartz glass cuvettes. The chlorophyll a andb concentrations, as well as the total carotenoid concentration,were calculated using the equations defined by Lichtenthaler andWellburn (1983) and expressed as pigment density (i.e. pigmentper absorption cross-section; Enríquez et al., 1992). Prior topigment analysis, the optical properties of intact Cymodocea nodosaleaves were measured spectrophotometrically, on the same leafsegments, using the opal glass technique (Shibata, 1959), followingthe methods and recommendations described by Enríquez et al.(1992), Enríquez (2005) and Cayabyab and Enríquez (2007).
For each leaf segment, absorbance (D) was measured in the PARrange (380e750 nm) at a resolution of 1 nm. A bleached leafsegment was used as a reference in the spectrophotometer. Thespectra obtained were corrected for residual scattering by sub-tracting the absorbance at 725 nm. Leaf absorption was expressedas absorptance (A), which is the fraction of incident light absorbed(Kirk, 1994), calculated using the equation A ¼ 1 � 10�D. Theabsorbance value at the 680 nm Chla peak was used, since it isa good descriptor of average PAR absorption (Enríquez et al., 1994;Olesen et al., 2002). Specific absorption coefficients were used asdescriptors of the efficiency of pigments and leaf mass at absorbinglight in seagrasses (Enríquez, 2005; Cayabyab and Enríquez, 2007).The chlorophyll a-specific andmass-specific absorption coefficientswere estimated from the exponent in the exponential function[exp¼�ln (1� A)], with values normalised to pigment content (a*;m2 mg�1 Chla) and leaf mass (aw* ; cm2 mg�1 leaf DW), respectively.Variation in the absorption efficiency of seagrass leaves is a func-tion of pigment content and also of leaf morphology and/or thepackaging of its mass (Enríquez, 2005). In order to determine thecontribution of leaf anatomy on variations in leaf optical properties,the leaf mass per area index (LMA; g DW cm�2) was estimated bymeasuring the area and weight of each leaf segment employed forpigment determinations.
2.6. Leaf morphology, growth rate and shoot survival
At the beginning of the experimental period, all shoots in eachEU were counted and three randomly selected shoots were markedfor leaf growth determinations (N ¼ 6 replicates per treatment)following the methods described by Zieman (1974) and Pérez andRomero (1994). The marked shoots were collected at the end ofthe experimental period. For each marked shoot, the number ofleaves was counted and the length and width of each leaf weremeasured, in order to calculate shoot size on a per-area basis(cm2 shoot�1). Newly formed leaf segments (those below themark)were separated from the rest and measured, in order to estimateshoot leaf growth rate from leaf elongation (cm shoot�1 day�1). Thesurface area of necrosis marks on the leaves was also measured andexpressed as a proportion of necrotic photosynthetic tissue pershoot. Shoot counts were repeated at the end of the experimentalperiod to estimate shoot survival. Differences between initial andfinal shoot numbers for each EU were normalised to initial shootnumbers and expressed as a percentage of net change. Negativevalues indicated a net decline from the initial shoot number,resulting from higher mortality rates than shoot recruitment rates(shoot division or rhizome branching) and/or reduced shootrecruitment rates (Ruiz et al., 2009a,b).
2.7. Field reference
Mean values of some plant variables obtained in the laboratorywere compared with reference values measured in the field during
the same season and in the same natural meadows fromwhich theplants were collected for this experiment. These field observationsformed part of a seasonal sampling of environmental and plantvariables performed in these meadows from February 2008 to May2009 (unpublished results). Data corresponding to the autumn2008 campaign are presented in Table 6. This comparison wasconsidered to be necessary in order to address issues, such as thepossible stress caused by the artificial conditions, which couldconfound the effects of the experimental treatments on the phys-iological and vegetative responses of the studied seagrass species.
2.8. Statistical analysis
A one-way ANOVA was used to assess significant treatmenteffects (afixed factorwith four salinity levels: 37 (control), 39, 41 and43) on the response variables at the end of the experimental period.The effect of salinity treatments on Fv/Fm time trends was analysedusing two-wayANOVA,with salinityand time (8 samplingoccasions)as the fixed factors in the model. A non-parametric KruskaleWallistest was used when assumptions of normality and homoscedas-ticity were not satisfied. Post hoc mean comparisons (StudenteNewmaneKeuls, SNK; Zar, 1984) were performed to identify whichspecific treatment level(s) produced significant effects. Treatmenteffects were considered statistically significant at p < 0.05.
3. Results
The mean maximum quantum yield of PSII (Fv/Fm) for Cymo-docea nodosa leaves varied between 0.773 � 0.002 and0.790 � 0.001. Salinity had a significant effect on this variable,although this was transitory since it was only detected duringsampling episodes 6 and 7 (i.e. 25 and 32 days from the onset of theexperimental treatments; Fig. 2 and Table 1). Thesemean values fellwithin the reference ranges measured in the natural meadow(Table 6) and those obtained from healthy plants of other seagrassspecies (Ralph, 1999; Durako and Kunzelman, 2002; Ralph et al.,2002; Cayabyab and Enríquez, 2007).
The P vs. E curves obtained from the four treatments at the endof the experimental exposure are shown in Fig. 3. The salinitytreatments had significant effects on the photosynthetic

Table 1Summary of the ANOVA test performed to assess significant effects of salinitytreatments on the course of mean Fv/Fm values in C. nodosa leaves throughout theexperimental period. df ¼ degrees of freedom; MS ¼ mean squares; p ¼ probabilitylevel; ns ¼ not significant; *p < 0.01.
Effect df MS F p
Salinity (S) 3 0.000118 3.55 *Time (T) 7 0.000064 1.93 nsS � T 21 0.000069 2.08 *Residual 160 0.000033
J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271 265
parameters derived from the P vs. E curves (Fig. 4 and Table 2). Themean net-Pmax of plants from the control treatment was1.37� 0.002 mmol O2 cm�2 h�1, whereas this valuewas significantlydecreased, by 12e17%, in the hypersaline treatments. The meangross-Pmax demonstrated similar values between the 37 and 41salinity treatments, but was significantly decreased, by 19.6%, in themost severe salinity (43).
Dark respiration rates varied significantly between the treat-ments (Fig. 4 and Table 2). Mean initial-Rd was 16.9% and 41.5%higher in the 39 and 41 treatment conditions, respectively, than inplants from the control (�0.154 � 0.004 mmol O2 cm�2 h�1). Meanfinal-Rd exhibited a similar pattern of variation with salinity, butwith higher differences between the treatments: the mean valuesobtained from 39 to 41 were 75e85% higher than those measuredin the control (0.171 � 0.001 mmol O2 cm�2 h�1). Values of initial-and final-Rd were reduced by 11% and 17%, respectively, in plantsexposed to themost severe salinity treatment (43) in relation to thecontrol.
Relative to the mean control values, mean photosynthetic effi-ciency (a) was significantly increased in the 39 and 41 salinitytreatments (by 15e18.5%), while the saturation irradiance (Ek)decreased by approximately 28%. Compensation irradiances (Ec)were significantly different between all the treatments, althoughthe largest effect was observed in plants from the 43 salinitytreatment, in which this parameter was reduced by 23% relative tothe mean control value.
The P:Rd ratio indicated a highly positive daily carbon balance inthe control plants (4.008 � 0.09). This ratio was significantly lowerin the 39 and 41 salinities (31% and 33%, respectively), but similar to
Fig. 3. Oxygen evolution rates (mean � SE, N ¼ 3) as a function of irradiance obtainedfor C. nodosa leaves after 47 days of exposure to each experimental salinity treatment:Control (37, solid circles), 39 (empty circles), 41 (solid triangles) and 43 (emptytriangles). Values measured in the dark (0 mmol quanta m�2 s�1) correspond to initial-Rd values and those measured under illuminated conditions are net photosyntheticrates.
that detected in leaves from plants exposed to the most salinetreatment (43).
Pigment contents (Chla, Chlb and total carotenoids) in Cymo-docea nodosa leaves were also affected by the salinity treatments(Table 3). Mean Chla concentration decreased as salinity increased,especially in the most severe hypersaline treatment (43), wheremean values were significantly lower (18.2%) than the controlmean. The concentration of Chlb and the Chlb:a ratio bothincreased significantly in the 39 and 41 salinity treatments (by 6e9% and 5e7%, respectively), compared to the mean values of thecontrol and 43 treatments.
Total carotenoid concentrations were increased in the tissuesfrom all treatments, although the largest differences were in theleaves from the 39 and 41 treatments, which had mean valuessignificantly higher (by 10e17%) than the control. The car-otenoid:Chla ratio increased significantly (by 9e15%) in all hyper-saline treatments (Table 3).
The leaf optics results from Cymodocea nodosa leaves obtainedfrom each experimental treatment are shown in Fig. 5 and Table 4.The salinity treatments had significant effects on leaf absorptance(A), specific absorption coefficients (a* and aw* ) and the leaf massper area (LMA) index. A was 0.845 � 0.002 in plants from thecontrol treatment, which was slightly lower than that obtained bySilva and Santos (2003) from an Atlantic C. nodosameadow. SimilarA values were obtained from the most severe hypersaline condition(43), but Awas significantly higher (0.87e0.88) in plants exposed tointermediate salinity increases (i.e. 39 and 41). The mean value ofthe specific absorption coefficients showed a constant linearincrease between the 37 and 41 treatments (r ¼ 0.999, p < 0.05),but no further increase in this variable was observed in salinity 43when this coefficient was standardised to pigment content (a*) oreven lower values when it was relativised to leaf mass (aw* ). TheLMA of the leaf segments employed for pigment and absorptancemeasurements decreased significantly in the hypersaline treat-ments, indicating a lower degree of biomass packing in leavesunder hypersaline conditions.
Shoot size and leaf growth rates were not significantly affectedby the salinity treatments, although the lowest mean values wereobserved in shoots from the 41 and 43 salinity treatments (Table 5).The frequency distribution of the number of leaves per shoot ineach experimental treatment (Fig. 6) exhibited significant differ-ences between treatments (c2¼ 78.6, p< 0.0001), mainly due to anincrease in the percentage of shoots with only one leaf (the first andyoungest leaf) in the most severe hypersaline treatment (36% ofshoots in the 43 treatment compared with 10e23% in theremainder). Significant differences were also identified in the leafnecrotic area percentage, although there was no apparent rela-tionship with salinity. While somewhat higher mean values wereobserved in plants from themost severe salinity treatments (41 and43), leaf necrosis was always found in the apical parts of the oldestleaves, indicating leaf senescence. The net change in initial shootnumbers was negligible for all experimental treatments (Table 5).
Mean values of photosynthetic parameters (Pmax, a and Rd),pigment concentrations and leaf growth rates obtained from thecontrol treatment at the end of the experimental period were allwithin those ranges observed in the reference Cymodocea nodosameadow during the season in which the experiment was per-formed (Table 6).
4. Discussion
The physiological and vegetative behaviour of those plantsmaintained at ambient salinity (37; control treatment) in themesocosm system for 47 days was remarkably similar to thatdirectly observed in their natural meadow during the same season.

Fig. 4. Variations in photosynthetic parameters (mean � SE, N ¼ 3) of C. nodosa leaves derived from P vs. E curves with salinity treatments. Net-Pmax ¼ net maximum photosyntheticrate; gross-Pmax ¼ gross maximum photosynthetic rate; initial-Rd ¼ initial dark respiration rate; final-Rd ¼ final dark respiration rate; a ¼ photosynthetic efficiency; Ek ¼ saturationirradiance; Ec ¼ compensation irradiance.
J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271266
This indicated the reliability of the experimental system to repro-duce optimum environmental and physiological conditions forCymodocea nodosa in the laboratory. Shoot survival in this controltreatment was 100%, emphasising the success of the mesocosmsystem in maintaining plant vitality for experimental periods 3e5times longer than those employed in previous laboratory experi-ments involving other Mediterranean C. nodosa populations(Fernández-Torquemada and Sánchez-Lizaso, 2006; Pagès et al.,2010). More importantly, these results indicated that the artificialconditions elicited little or negligible stress on the observed plantbehaviour, which was, therefore, primarily a response to theexperimental treatments (salinity increases) and not to otherconfounding effects.
The photosynthetic physiology of the Cymodocea nodosa plantsused in this study was significantly altered following exposure tochronic salinity increases, above the average level at which they are
adapted to grow under natural conditions (i.e. infralittoral Medi-terranean habitats with a constant salinity of 37). Photosynthesisinhibition is a phenomenon generally observed in this (Drew,1978a), and other (Ogata and Matsui, 1964; Biebl and McRoy,1971; Kerr and Strother, 1985; Fernández-Torquemada et al.,2005b; Kahn and Durako, 2006; Koch et al., 2007), seagrassspecies when exposed to hypersaline stress. Accordingly, netphotosynthesis was partially and consistently inhibited in allhypersaline treatments in the present study, although the resultsobtained for other physiological variables suggested that the natureof the inhibition differed depending on the severity of the hyper-saline stress.
At intermediate salinity increases (39 and 41), gross photosyn-thesis remained close to control values and the reduction in netphotosynthesis was accounted for by the enhancement of darkrespiration rates observed only in these treatments. An increase in
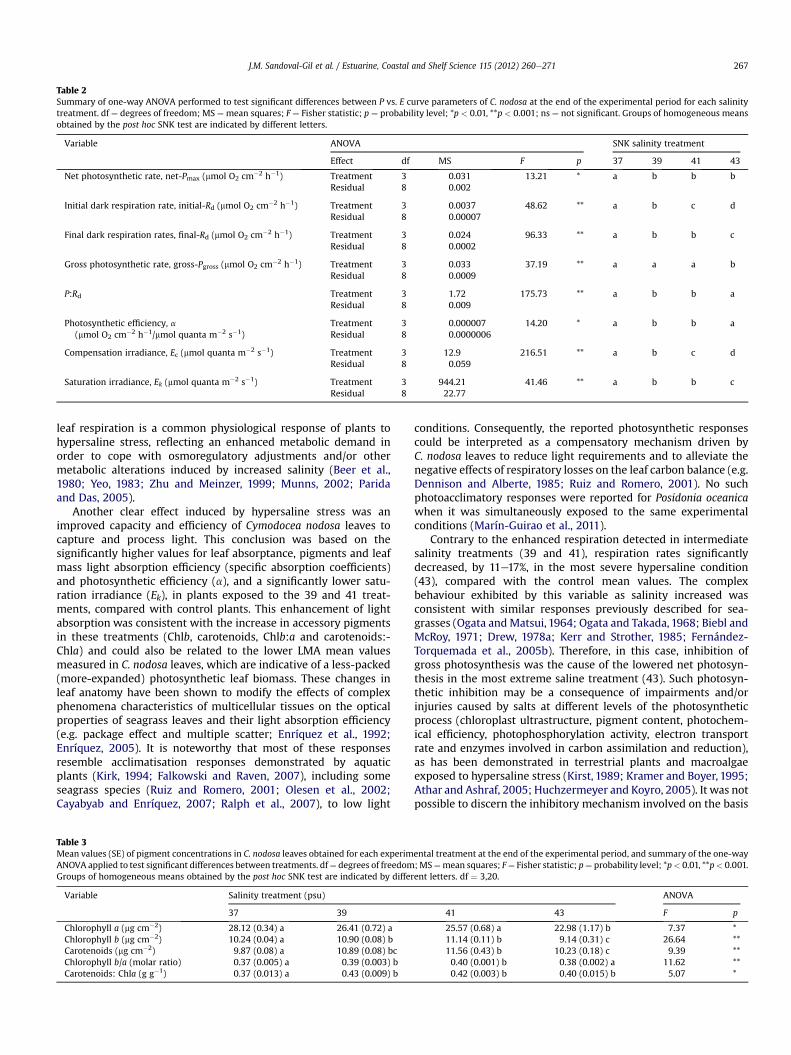
Table 2Summary of one-way ANOVA performed to test significant differences between P vs. E curve parameters of C. nodosa at the end of the experimental period for each salinitytreatment. df ¼ degrees of freedom; MS ¼mean squares; F ¼ Fisher statistic; p ¼ probability level; *p < 0.01, **p < 0.001; ns ¼ not significant. Groups of homogeneous meansobtained by the post hoc SNK test are indicated by different letters.
Variable ANOVA SNK salinity treatment
Effect df MS F p 37 39 41 43
Net photosynthetic rate, net-Pmax (mmol O2 cm�2 h�1) Treatment 3 0.031 13.21 * a b b bResidual 8 0.002
Initial dark respiration rate, initial-Rd (mmol O2 cm�2 h�1) Treatment 3 0.0037 48.62 ** a b c dResidual 8 0.00007
Final dark respiration rates, final-Rd (mmol O2 cm�2 h�1) Treatment 3 0.024 96.33 ** a b b cResidual 8 0.0002
Gross photosynthetic rate, gross-Pgross (mmol O2 cm�2 h�1) Treatment 3 0.033 37.19 ** a a a bResidual 8 0.0009
P:Rd Treatment 3 1.72 175.73 ** a b b aResidual 8 0.009
Photosynthetic efficiency, a(mmol O2 cm�2 h�1/mmol quanta m�2 s�1)
Treatment 3 0.000007 14.20 * a b b aResidual 8 0.0000006
Compensation irradiance, Ec (mmol quanta m�2 s�1) Treatment 3 12.9 216.51 ** a b c dResidual 8 0.059
Saturation irradiance, Ek (mmol quanta m�2 s�1) Treatment 3 944.21 41.46 ** a b b cResidual 8 22.77
J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271 267
leaf respiration is a common physiological response of plants tohypersaline stress, reflecting an enhanced metabolic demand inorder to cope with osmoregulatory adjustments and/or othermetabolic alterations induced by increased salinity (Beer et al.,1980; Yeo, 1983; Zhu and Meinzer, 1999; Munns, 2002; Paridaand Das, 2005).
Another clear effect induced by hypersaline stress was animproved capacity and efficiency of Cymodocea nodosa leaves tocapture and process light. This conclusion was based on thesignificantly higher values for leaf absorptance, pigments and leafmass light absorption efficiency (specific absorption coefficients)and photosynthetic efficiency (a), and a significantly lower satu-ration irradiance (Ek), in plants exposed to the 39 and 41 treat-ments, compared with control plants. This enhancement of lightabsorption was consistent with the increase in accessory pigmentsin these treatments (Chlb, carotenoids, Chlb:a and carotenoids:-Chla) and could also be related to the lower LMA mean valuesmeasured in C. nodosa leaves, which are indicative of a less-packed(more-expanded) photosynthetic leaf biomass. These changes inleaf anatomy have been shown to modify the effects of complexphenomena characteristics of multicellular tissues on the opticalproperties of seagrass leaves and their light absorption efficiency(e.g. package effect and multiple scatter; Enríquez et al., 1992;Enríquez, 2005). It is noteworthy that most of these responsesresemble acclimatisation responses demonstrated by aquaticplants (Kirk, 1994; Falkowski and Raven, 2007), including someseagrass species (Ruiz and Romero, 2001; Olesen et al., 2002;Cayabyab and Enríquez, 2007; Ralph et al., 2007), to low light
Table 3Mean values (SE) of pigment concentrations in C. nodosa leaves obtained for each experimANOVA applied to test significant differences between treatments. df¼ degrees of freedomGroups of homogeneous means obtained by the post hoc SNK test are indicated by differ
Variable Salinity treatment (psu)
37 39
Chlorophyll a (mg cm�2) 28.12 (0.34) a 26.41 (0.72) aChlorophyll b (mg cm�2) 10.24 (0.04) a 10.90 (0.08) bCarotenoids (mg cm�2) 9.87 (0.08) a 10.89 (0.08) bcChlorophyll b/a (molar ratio) 0.37 (0.005) a 0.39 (0.003) bCarotenoids: Chla (g g�1) 0.37 (0.013) a 0.43 (0.009) b
conditions. Consequently, the reported photosynthetic responsescould be interpreted as a compensatory mechanism driven byC. nodosa leaves to reduce light requirements and to alleviate thenegative effects of respiratory losses on the leaf carbon balance (e.g.Dennison and Alberte, 1985; Ruiz and Romero, 2001). No suchphotoacclimatory responses were reported for Posidonia oceanicawhen it was simultaneously exposed to the same experimentalconditions (Marín-Guirao et al., 2011).
Contrary to the enhanced respiration detected in intermediatesalinity treatments (39 and 41), respiration rates significantlydecreased, by 11e17%, in the most severe hypersaline condition(43), compared with the control mean values. The complexbehaviour exhibited by this variable as salinity increased wasconsistent with similar responses previously described for sea-grasses (Ogata andMatsui, 1964; Ogata and Takada, 1968; Biebl andMcRoy, 1971; Drew, 1978a; Kerr and Strother, 1985; Fernández-Torquemada et al., 2005b). Therefore, in this case, inhibition ofgross photosynthesis was the cause of the lowered net photosyn-thesis in the most extreme saline treatment (43). Such photosyn-thetic inhibition may be a consequence of impairments and/orinjuries caused by salts at different levels of the photosyntheticprocess (chloroplast ultrastructure, pigment content, photochem-ical efficiency, photophosphorylation activity, electron transportrate and enzymes involved in carbon assimilation and reduction),as has been demonstrated in terrestrial plants and macroalgaeexposed to hypersaline stress (Kirst, 1989; Kramer and Boyer, 1995;Athar and Ashraf, 2005; Huchzermeyer and Koyro, 2005). It was notpossible to discern the inhibitory mechanism involved on the basis
ental treatment at the end of the experimental period, and summary of the one-way;MS¼mean squares; F¼ Fisher statistic; p¼ probability level; *p< 0.01, **p< 0.001.ent letters. df ¼ 3,20.
ANOVA
41 43 F p
25.57 (0.68) a 22.98 (1.17) b 7.37 *11.14 (0.11) b 9.14 (0.31) c 26.64 **11.56 (0.43) b 10.23 (0.18) c 9.39 **0.40 (0.001) b 0.38 (0.002) a 11.62 **0.42 (0.003) b 0.40 (0.015) b 5.07 *

Fig. 5. Variations in optical properties (mean � SE, N ¼ 6) of C. nodosa leaves with salinity treatment: light absorption spectra and leaf absorptance (upper panels), and pigment-specific absorption coefficients (a*, aw* ) and leaf mass per area (LMA) or biomass packing (lower panels).
J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271268
of our results, and direct or indirect evidence is rather scarce forCymodocea nodosa (Drew, 1978a), as is the case for most seagrassspecies (for a review, see Touchette, 2007).
Beer et al. (1980) provided some experimental evidence that theactivity of the carbon-fixing enzyme RuBPcase present in theepidermis of Halodule uninervis was gradually inhibited by
Table 4Summary of the one-way ANOVA performed to test significant differences betweenleaf optical descriptors of C. nodosa at the end of the experimental period at eachsalinity treatment. df ¼ degrees of freedom; MS ¼ means squares; F ¼ Fisherstatistic; p¼ probability level; *p< 0.01, **p< 0.001. Groups of homogeneous meansobtained by the post hoc SNK test are indicated by different letters.
Variable ANOVA SNK salinitytreatment
Effect df MS F p 37 39 41 43
Pigment specificabsorptioncoefficient (a*)
Treatment 3 0.000003 7.45 * a b b bResidual 20 0.00000036
Leaf mass-specificabsorptioncoefficient (a*w)
Treatment 3 0.002705 11.81 ** a b c bResidual 20 0.000229
Leaf mass perarea (LMA)
Treatment 3 0.000001 8.37 ** a b b bResidual 20 0.0000001
Absorptance (A) Treatment 3 0.001231 8.33 ** a b b aResidual 20 0.000148
increasing NaCl concentrations in in vitro assays. Inhibition ofphotosynthesis induced by hypersaline stress has also been asso-ciated with conformational changes of the photosynthetic appa-ratus (McMillan and Moseley, 1967; Jagels and Barnabas, 1989; Iyerand Barnabas, 1993; Ralph, 1998, 1999). Decreases in pigmentcontent and/or photochemical efficiency have been reported insome seagrass species in relation to hypersaline stress, asa symptom of damage to photosynthetic structures (e.g. Ralph,1998, 1999; Kamermans et al., 1999; Murphy et al., 2003; Kahnand Durako, 2006; Koch et al., 2007; Pagès et al., 2010), althoughsuch symptoms were only apparent under hypersaline conditionsfar more extreme than those studied here. Consistently, negativeeffects of hypersaline stress on the structure and function of thephotosynthetic apparatus of land plants usually occur in extremeconditions (Huchzermeyer and Koyro, 2005).
The decrease in chlorophyll density (Chla and Chlb) reported insalinity treatment 43 could also be interpreted in this context,although it could alternatively be a consequence of a metabolicdown-regulation of light harvesting, in order to adjust light captureto compensate for reduced carbon assimilation (Demmig-Adamsand Adams, 1996). In accordance with the latter possibility, Fv/Fmvalues provided no evidence that hypersaline treatments causedthe accumulated damage to the photosystems and antennaesystems, even in the most severe hypersaline treatment. Whateverthe mechanism involved, the partial inhibition of gross photosyn-thesis reported in the most severe hypersaline treatment should

Table 5Mean values (SE) of shoot variables of C. nodosa and summary of the one-way ANOVA and non-parametric KruskaleWallis H test performed to assess differences betweensalinity treatments measured at the end of the experimental period. ns ¼ not significant; df ¼ degrees of freedom; MS ¼ mean squares; F ¼ Fisher statistic; p ¼ probabilitylevel; p ¼ probability level; *p < 0.05; H ¼ KruskaleWallis statistic. Groups of homogeneous means obtained by the post hoc SNK test are indicated by different letters.
Variable Salinity treatment (psu) ANOVA KruskaleWallis test
37 39 41 43 F p H p
Shoot size (cm2 shoot�1) 3.13 (0.32) 3.28 (0.31) 3.02 (0.27) 2.66 (0.23) 1.08 nsLeaf growth rate (cm2 shoot�1 day�1) 0.064 (0.007) 0.068 (0.007) 0.053 (0.004) 0.061 (0.004) 0.74 ns% Leaf necrotic area 10.97 (3.5) ab 3.24 (1.23) a 20.07 (6.0) b 10.8 (4.1) ab 4.52 *% Shoot mortality 2.38 (1.06) 0 0 2.08 (1.93) 4.62 ns
J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271 269
contribute in the impairment of the leaf carbon balance. As clearlyindicated by the high leaf P:Rd ratio estimated from this treatment(similar to the control), this adverse situation was primarily coun-terbalanced by limiting the dark respiration rate to a level belowthat of the control mean. This particular respiration rate behaviourcould be interpreted as a disruption of cellular respiration or,alternatively, as a control mechanism to cope with the demands ofan altered metabolism, to enable survival under extremely stressfulconditions. A similar control of respiratory activity has been typi-cally described in terrestrial plants (Kramer and Boyer, 1995;Lambers et al., 2006) and marine macrophytes (Ogata andTakada, 1968; Kirst, 1989) following exposure to hypersalineconditions and has been associatedwith a general decline in carbonmetabolism and the starvation of respiratory substrates, or itsdiversion to osmoregulatory and osmoprotectant functions.
In addition to these possible acclimatisation mechanisms, ourresults suggest that Cymodocea nodosa may develop additionalmechanisms in order to help it cope with hypersaline stress. Forinstance, the increased total carotenoid content (and the car-otenoid:Chla ratio) observed in all the hypersaline treatmentscould be related to the activation of general protective mechanismsby the photosynthetic structures against photo-damage andoxidative stress induced by hypersaline stress (Demmig-Adams andAdams, 1996; Ralph et al., 2002; Athar and Ashraf, 2005; Parida andDas, 2005). Evidence of such mechanisms was found in the domi-nant Mediterranean seagrass Posidonia oceanica under the sameexperimental conditions (Marín-Guirao et al., 2011). In that sea-grass species, leaf light absorption, photosynthetic efficiency,photosynthetic rates (net and gross) and leaf carbon balance (withP:Rd 45e56% lower than control means) were all more negativelyaffected by the same hypersaline treatments than C. nodosa, even
Fig. 6. Percentage of C. nodosa shoots with 1, 2 or 3 leaves after 47 days of exposure toeach experimental treatment.
for the lowest salinity increase (39). These severe physiologicalalterations clearly interfered with the metabolic requirements forgrowth and survival in P. oceanica shoots. Comparatively, themetabolic carbon balance of C. nodosa leaves was much lessaffected within the same salinity range (31e33%, and only in theintermediate salinity treatments), with no apparent disruption ofleaf growth and shoot survival. Thus, under the same experimentalconditions, it is clear that P. oceanica is physiologically moresensitive to hypersaline stress than C. nodosa and that the plasticityof the photosynthetic physiology described in this study likelyrepresents an adaptation mechanism. Evidence of such differentialadaptation mechanisms was also found during the experimentdocumented here, relating to the role of other key physiologicalaspects in the responses of these seagrass species to hypersalinestress, such as the water relations (Sandoval-Gil et al., 2012).
Previous short-term laboratory experiments established thatCymodocea nodosa is able to survive and sustain growth in salinitiesof up to 41e44 (Fernández-Torquemada and Sánchez-Lizaso, 2006;Pagès et al., 2010). Our results are consistent with these earlierstudies, but also provide physiological evidence that these salinitylevels must be close to the plant’s physiological tolerance threshold.In effect, as explained before, the partial inhibition of grossphotosynthesis and the reduced respiratory activity reported from43 salinity treatment enables plants to survive in this severe stresscondition, but presumably with reduced vitality, since some of theinternal resources required for growth and biomass maintenancemust be reallocated to cope with stress metabolism (Lichtenthaler,1996). The leaf loss reported for shoots from this treatment isconsistent with such a situation, and others described for sea-grasses in relation to other stressors (e.g. light limitation; Ralphet al., 2007). Obviously, this tolerance threshold is only valid forC. nodosa populations growing in open coastal waters of theSpanishMediterranean Seawith a mean constant salinity of 37e38,and cannot be extrapolated to other habitats, regions or situationswith a different salinity regime. There are, for instance, highly
Table 6Reference values of some C. nodosa variables selected for this study measured in thefield. SE ¼ standard error.
Field reference values of C. nodosa
Maximum Minimum Mean (SE)
Net photosynthetic rate, net-Pmax
(mmol O2 cm�2 h�1)1.17 0.88 1.02 (0.06)
Photosynthetic efficiency, a(mmol O2 cm�2 h�1/mmol quantam�2 s�1)
0.015 0.014 0.014 (0.0004)
Initial dark respiration rate,initial-Rd (mmol O2 cm�2 h�1)
�0.15 �0.21 �0.18 (0.011)
Maximum quantum yield (Fv/Fm) 0.78 0.75 0.76 (0.002)Chlorophyll a (mg cm�2) 25.34 20.85 23.18 (0.7)Chlorophyll b (mg cm�2) 7.83 6.41 7.12 (0.23)Carotenoids (mg cm�2) 10.3 9.01 9.6 (0.22)Leaf growth rate
(cm2 shoot�1 day�1)0.064 0.02 0.041 (0.0057)

J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271270
productive C. nodosa populations in hypersaline lagoons in theMediterranean Sea (e.g. up to 47 in theMarMenor lagoon; Terradosand Ros, 1991) for which a higher tolerance threshold is expected(Fernández-Torquemada and Sánchez-Lizaso, 2011), as a result oflong-term genotypic selection (e.g. Tomasello et al., 2009) and thepossible existence of interactive effects between salinity and otherkey environmental factors that also vary between habitat types (e.g.Van Katwijk et al., 1999; Koch et al., 2007). However, this issuedeserves further research on C. nodosa, combining laboratoryexperiments and field work.
In conclusion, our study provides experimental evidence thatMediterranean populations of Cymodocea nodosa adapted tomarine infralittoral environments with stable salinity are physio-logically more tolerant than Posidonia oceanica to sudden andchronic increases in external salinity. This finding has severalgeneral ecological implications. Firstly, it is consistent with thedistinct biologies and ecologies exhibited by these two seagrassspecies, and thus explain their abundance and distributionthroughout the various marine habitats off the Mediterraneancoast. Secondly, it allows us to predict that natural populations ofC. nodosa adapted to Mediterranean open coastal environmentswill be more resistant to anthropogenically induced salinityincreases (e.g., brine discharges from desalination plants) thanneighbouring meadows of the more sensitive P. oceanica. None-theless, care must be taken when extrapolating conclusions fromcontrolled laboratory experiments performed on short to mediumtimescales to more complex environmental situations in the natureover extended periods.
Acknowledgements
This research was funded by two Spanish Government Minis-tries: the Ministerio de Medioambiente y Medio Rural y Marino(OSMOGRASS project no. 021/SGTB/2007/1.3) and theMinisterio deCiencia e Innovación (OSMOGRASS II project no. CTM2009-08413MAR), and a research grant was awarded to JM Sandoval bythe University of Alicante. We would like to thank Dr SusanaEnríquez (Universidad Nacional Autónoma de México) and MaríaJose Quiles (Universidad de Murcia, Spain) for their valuablesupport and advice regarding the use of respirometry and PAM-fluorometry techniques and interpretation of the photosyntheticresponses. Field and laboratory logistical support was provided byIEO technicians Rocío García Muñoz and Arantxa Ramos Segura andPhD student Jaime Bernardeau Esteller. Wewould also like to thankEmilio Cortés Melendreras (Marine Aquarium of the Universidad deMurcia) for providing technical support during the design anddeployment of the mesocosm system.
References
Agostini, S., Pergent, G., Marchand, B., 2003. Growth and primary production ofCymodocea nodosa in a coastal lagoon. Aquatic Botany 76, 185e193.
Arber, A., 1920. Water Plants. A Study of Aquatic Angiosperms. Cambridge Univer-sity Press, Cambridge.
Athar, H., Ashraf, M., 2005. Photosynthesis under drought stress. In: Pessarakli, M.(Ed.), Hand Book of Photosynthesis, second ed. C.R.C. Press, New York, USA,pp. 795e810.
Beer, S., Eshel, A., Waisel, Y., 1980. Carbon metabolism in seagrasses III. Activities ofcarbon-fixing enzymes in relation to internal salt concentrations. Journal ofExperimental Botany 31, 1027e1033.
Beer, S., Björk, M., Gademann, R., Ralph, P., 2001. Measurements of photosyntheticrates in seagrasses. In: Short, F.T., Coles, R.G. (Eds.), Global Seagrass ResearchMethods. Elsevier, Amsterdam, pp. 183e198.
Biebl, R., McRoy, C.P., 1971. Plasmatic resistance and rate of respiration andphotosynthesis of Zostera marina at different salinities and temperatures.Marine Biology 8, 48e56.
Boudouresque, C.F., Bernard, G., Pergent, G., Shili, A., Verlaque, M., 2009. Regressionof Mediterranean seagrasses caused by natural processes and anthropogenicdisturbances and stress: a critical review. Botanica Marina 52, 395e418.
Cancemi, G., Buia, M.C., Mazzella, L., 2002. Structure and growth dynamics ofCymodocea nodosa meadows. Scientia Marina 66 (4), 365e373.
Castriota, L., Beltrano, A.M., Giambalvo, O., Vivona, P., Sunseri, G., 2001. A One-yearStudy of the Effects of a Hyperhaline Discharge from a Desalination Plant on theZoobenthic Communities in the Ustica Island Marine Reserve (Southern Tyr-rhenian Sea), vol. 36. CIESM, Monaco.
Cayabyab, N.M., Enríquez, S., 2007. Leaf photoacclimatory responses of the tropicalseagrass Thalassia testudinum under mesocosm conditions: a mechanisticscaling-up study. New Phytologist 176, 108e123.
Cebrián, J., Pedersen, M.F., Kroeger, K.D., Valiela, I., 2000. Fate of production of theseagrass Cymodocea nodosa in different stages of meadow formation. MarineEcology Progress Series 204, 119e130.
Chesher, R.H., 1975. Biological impact of a large-scale desalination plant at KeyWest, Florida. In: Ferguson, E.J., Johanes, R.E. (Eds.), Tropical Marine Pollution.Elsevier Scientific Publishing Company, Amsterdam, pp. 99e181.
Demmig-Adams, B., Adams, W.W., 1996. The role of xanthophyll cycle carotenoidsin the protection of photosynthesis. Trends in Plant Science 1, 21e26.
den Hartog, C., 1970. The Sea-grasses of the World. North-Holland PublicationsCompany, Amsterdam.
Dennison, W.C., 1990. Chlorophyll content. In: Phillips, R.C., McRoy, C.P. (Eds.),Seagrass Research Methods. UNESCO, Paris, France, pp. 83e86.
Dennison, W.C., Alberte, R.S., 1985. Role of daily light period in the depth distri-bution of Zostera marina, eelgrass. Marine Ecology Progress Series 25, 5le62.
Drew, E.A., 1978a. Carbohydrate and inositol metabolism in seagrass Cymodoceanodosa. New Phytologist 81, 249e264.
Drew, E.A., 1978b. Factors affecting photosynthesis and its seasonal variation in theseagrass Cymodocea nodosa (Ucria) Aschers, and Posidonia oceanica (L.) Delile inthe Mediterranean. Journal of Experimental Marine Biology and Ecology 31,173e194.
Durako, M.J., Kunzelman, J.I., 2002. Photosynthetic characteristics of Thalassia tes-tudinum measured in situ by pulse-amplitude modulated (PAM) fluorometry:methodological and scale-based considerations. Aquatic Botany 73, 173e185.
Einav, R., Harussi, K., Perry, D., 2002. The footprint of desalination processes on theenvironment. Desalination 152, 141e154.
Enríquez, S., 2005. Light absorption efficiency and the package effect in the leaves ofthe seagrass Thalassia testudinum. Marine Ecology Progress Series 289, 141e150.
Enríquez, S., Rodríguez-Román, A., 2006. Effect of water flow on the photosynthesisof three marine macrophytes from a fringing-reef lagoon. Marine EcologyProgress Series 323, 119e132.
Enríquez, S., Agustí, S., Duarte, C.M., 1992. Light absorption by seagrass Posidoniaoceanica leaves. Marine Ecology Progress Series 86, 201e204.
Enríquez, S., Agustí, S., Duarte, C.M., 1994. Light absorption by marine macrophytes.Oecologia 98, 121e129.
Enríquez, S., Merino, M., Iglesias-Prieto, R., 2002. Variations in the photosyntheticperformance along the leaves of the tropical seagrass Thalassia testudinum.Marine Biology 140, 891e900.
Falkowski, P., Raven, J., 2007. Aquatic Photosynthesis, second ed. PrincetonUniversity Press, Princeton, 484 pp.
Fernández-Torquemada, Y., Sánchez-Lizaso, J.L., 2005. Effects of salinity on leafgrowth and survival of the Mediterranean seagrass Posidonia oceanica (L.)Delile. Journal of Experimental Marine Biology and Ecology 320, 57e63.
Fernández-Torquemada, Y.F., Sánchez-Lizaso, J.L., 2006. Effects of salinity on growthand survival of Cymodocea nodosa (Ucria) Ascherson and Zostera noltii Horne-man. Biologia Marina Mediterranea 13, 46e47.
Fernández-Torquemada, Y.F., Sánchez-Lizaso, J.L., 2011. Responses of two Medi-terranean seagrasses to experimental changes in salinity. Hydrobiologia 669,21e33.
Fernández-Torquemada, Y.F., Sánchez-Lizaso, J.L., González-Correa, J.M.G., 2005a.Preliminary results of the monitoring of the brine discharge produced by theSWRO desalination plant of Alicante (SE Spain). Desalination 182, 579e590.
Fernández-Torquemada, Y.F., Durako, M.J., Sánchez-Lizaso, J.L., 2005b. Effects ofsalinity and possible interactions with temperature and pH on growth andphotosynthesis of Halophila johnsonii Eiseman. Marine Biology 148, 251e260.
Gacia, E., Invers, O., Manzanera, M., Ballesteros, E., Romero, J., 2007. Impact of thebrine from a desalination plant on a shallow seagrass (Posidonia oceanica)meadow. Estuarine, Coastal and Shelf Science 72, 579e590.
Huchzermeyer, B., Koyro, H.W., 2005. Salt and drought stress effects on photosyn-thesis. In: Pessarakli, M. (Ed.), Handbook of Photosynthesis, second ed. CRCPress, Taylor & Francis Publishing Company, Florida, pp. 1065e1092.
Invers, O., Romero, J., Pérez, M., 1997. Effects of pH on seagrass photosynthesis:a laboratory and field assessment. Aquatic Botany 59, 185e194.
Iyer, V., Barnabas, A.D., 1993. Effects of varying salinity on leaves of Zostera capensisSetchell. I. Ultrastructural changes. Aquatic Botany 46, 141e153.
Jagels, R., Barnabas, A., 1989. Variation in leaf ultrastructure of Ruppia maritima L.along a salinity gradient. Aquatic Botany 33, 207e221.
Kahn, A.E., Durako, M.J., 2006. Thalassia testudinum seedling responses to changesin salinity and nitrogen levels. Journal of Experimental Marine Biology andEcology 335, 1e12.
Kamermans, P., Hemminga, M.A., De Jong, D.J., 1999. Significance of salinity andsilicon levels for growth of a formerly estuarine eelgrass (Zostera marina)population (Lake Grevelin, The Netherlands). Marine Biology 133, 527e539.
Kenworthy, W.J., Wyllie-Echeverria, S., Coles, R.G., Pergent, G., Pergent-Martini, C.,2006. Seagrass conservation biology: an interdisciplinary science for protectionof the seagrass biome. In: Larkum, A.W.D., Orth, R.J., Duarte, C.M. (Eds.), Sea-grasses: Biology, Ecology and Conservation. Springer, Dordrecht, pp. 595e623.

J.M. Sandoval-Gil et al. / Estuarine, Coastal and Shelf Science 115 (2012) 260e271 271
Kerr, E.A., Strother, S., 1985. Effects of irradiance, temperature and salinity onphotosynthesis of Zostera muelleri. Aquatic Botany 23, 177e183.
Kirk, J.T.O., 1994. Light and Photosynthesis in Aquatic Ecosystems, second ed.University of Cambridge Press, Cambridge, UK, England, 509 pp.
Kirst, G.O., 1989. Salinity tolerance of eukaryotics marine algae. Annual Review ofPlant Physiology and Plant Molecular Biology 40, 21e53.
Koch, M.S., Schopmeyer, S.A., Kyhn-Hansen, C., Madden, C.J., Peters, J.S., 2007. Tropicalseagrass species tolerance to hypersalinity stress. Aquatic Botany 86, 14e24.
Kramer, P.J., Boyer, J.S., 1995. Water Relations of Plants and Soils. Academic Press,San Diego.
Kuo, J., Den Hartog, C., 2000. Seagrasses: a profile of an ecological group. BioliaMarina Mediterranea 7, 3e17.
Lambers, H., Chapin, F.S., Pons, T.L., 2006. Plant Physiological Ecology. Springer-Verlag, New York.
Larkum, A.W.D., Drew, E.A., Ralph, P.J., 2006. Photosynthesis and metabolisms inseagrasses at the cellular level. In: Larkum, A.W.D., Orth, R.J., Duarte, C.M. (Eds.),Seagrasses: Biology, Ecology and Conservation, pp. 323e345. The Netherlands.
Lichtenthaler, H.K., Wellburn, A.R., 1983. Determination of total carotenoids andchlorophyll a and b of leaf extracts in different solvents. Biochemical SocietyTransactions 603, 591e592.
Lichtenthaler, H.K., 1996. Vegetation stress: an introduction to the stress concept inplants. Plant Physiology 148, 4e14.
Marbá, N., Duarte, C.M., 2001. Growth and sediment space occupation by seagrassCymodocea nodosa roots. Marine Ecology Progress Series 224, 291e298.
Marbá, N., Hemminga, M.A., Mateo, M.A., Duarte, C.M., Mass, Y., Terrados, J.,Gacia, E., 2002. Carbon and nitrogen translocation between seagrass ramets.Marine Ecology Progress Series 226, 287e300.
Margalef, R., 1989. Western Mediterranean. Omega, Barcelona.Marín-Guirao, L., Sandoval-Gil, J.M., Ruiz, J.M., Sánchez-Lizaso, J.L., 2011. Photo-
synthesis, growth and survival of the Mediterranean seagrass Posidonia oce-anica in response to simulated salinity increases in a laboratory mesocosmsystem. Estuarine, Coastal and Shelf Science. http://dx.doi.org/10.1016/j.ecss.2011.01.003.
McMillan, C., Moseley, F.N., 1967. Salinity tolerances of five marine spermatophytesof Redfish Bay, Texas. Ecology 48, 503e506.
Meinesz, A., Cirik, S., Akcali, B., Javel, F., Migliaccio, M., Thibaut, T., Yüksek, A.,Procaccini, G., 2009. Posidonia oceanica in the Marmara Sea. Aquatic Botany 90,18e22.
Montague, C.L., Ley, J.A., 1993. A possible effect of salinity fluctuation on abundanceof benthic vegetation and associated fauna in north-eastern Florida Bay. Estu-aries 16, 703e717.
Munns, R., 2002. Comparative physiology of salt and water stress. Plant, Cell andEnvironment 25, 239e250.
Murphy, L.R., Kinsey, S.T., Durako, M.J., 2003. Physiological effects of short-termsalinity changes on Ruppia maritima. Aquatic Botany 75, 293e309.
Ogata, E., Matsui, T., 1964. Photosynthesis in several marine plants of Japan asaffected by salinity, drying and pH, with attention to their growth habitat.Botanica Marina 8, 199e217.
Ogata, E., Takada, H., 1968. Studies on the relationship between the respiration andthe changes in salinity in some marine plants in Japan. Journal of ShimonsekiUniversity of Fisheries 16, 67e88.
Oksanen, L., 2001. Logic of experiments in ecology: is pseudoreplication a pseu-doissue? Oikos 94, 27e38.
Olesen, B., Enríquez, S., Duarte, C.M., Sand-Jensen, K., 2002. Depth-acclimation ofphotosynthesis, morphology and demography of Posidonia oceanica and Cym-odocea nodosa in the Spanish Mediterranean Sea. Marine Ecology ProgressSeries 236, 89e97.
Olivé, I., García-Sánchez, M.P., Brun, F.G., Vergara, J.J., Pérez-Llorens, J.L., 2009.Interactions of light and organic matter under contrasting resource simulatedenvironments: the importance of clonal traits in the seagrass Zostera noltii.Hydrobiologia 629, 199e208.
Pagès, J.F., Pérez, M., Romero, J., 2010. Sensitivity of the seagrass Cymodocea nodosato hypersaline conditions: a microcosm approach. Journal of ExperimentalMarine Biology and Ecology 386, 34e38.
Palomar, P., Losada, I.J., 2010. Desalination in Spain: recent developments andrecommendations. Desalination 255, 97e106.
Parida, A.K., Das, A.B., 2005. Salt tolerant and salinity effects on plants. A review.Ecotoxicology and Environmental Safety 60, 324e349.
Pérez, M., Romero, J., 1994. Growth dynamics, production and nutrient status of theseagrass Cymodocea nodosa in a Mediterranean semi-estuarine environment.PSZNI Marine Ecology 15, 51e64.
Procaccini, G., Buia, M.C., Gambi, M.C., Perez, M., Pergent, G., Pergent-Martini, C.,Romero, J., 2003. Seagrasses of the western mediterranean. In: Green, E.P.,Short, F.T. (Eds.), World Atlas of Seagrasses. University of California Press, LosAngeles, California, pp. 48e52.
Quinn, G.P., Keough, M.J., 2002. Experimental Design and Data Analysis for Biolo-gists. Cambridge University Press, Cambridge.
Ralph, P.J.,1998. Photosynthetic responses ofHalophila ovalis (R. Br.)Hook. f. to osmoticstress. Journal of Experimental Marine Biology and Ecology 227, 203e220.
Ralph, P.J., 1999. Photosynthetic response of Halophila ovalis (R. Br.) Hook. f. tocombined environmental stress. Aquatic Botany 65, 83e96.
Ralph, P.J., Polk, S.M., Moore, K.A., Orth, R.J., Smith Jr., W.O., 2002. Operation of thexanthophyll cycle in the seagrass Zostera marina in response to variable irra-diance. Journal of Experimental Marine Biology and Ecology 271, 189e207.
Ralph, P.J., Durako, M.J., Enríquez, S., Collier, C.J., Doblin, M.A., 2007. Impact of lightlimitation on seagrasses. Journal of Experimental Marine Biology and Ecology350, 176e193.
Ruiz, J.M., Romero, J., 2001. Effects of in situ experimental shading on the Medi-terranean seagrass Posidonia oceanica. Marine Ecology Progress Series 215,107e120.
Ruiz, J.M., Boudouresque, C.F., Enríquez, S., 2009a. Mediterranean seagrasses.Botanica Marina 52, 369e381.
Ruiz, J.M., Marín-Guirao, L., Sandoval-Gil, J.M., 2009b. Responses of the Mediter-ranean seagrass Posidonia oceanica to in situ simulated salinity increase.Botanica Marina 52, 459e470.
Sánchez-Lizaso, J.L., Romero, J., Ruíz, J.M., Gacia, E., Buceta, J.L., Invers, O.,Torquemada, Y.F., Más, J., Ruíz-Mateo, A., Manzanera, M., 2008. Salinity toler-ance of the Mediterranean seagrass Posidonia oceanica: recommendations tominimise the impact of brine discharges from desalination plants. Desalination221, 602e607.
Sandoval-Gil, J.M., Marín-Guirao, L., Ruiz, J.M., 2012. Tolerance of Mediterraneanseagrasses (Posidonia oceanica and Cymodocea nodosa) to hypersaline stress:water relations and osmolyte concentrations. Marine Biology 159, 1129e1141.
Schreiber, U., 2004. Pulse-amplitude-modulation (PAM) fluorometry and saturationpulse method: an overview. In: Papageorgiou, G.C., Govindjee (Eds.), Chloro-phyll a Fluorescence: a Signature of Photosynthesis. Springer, Dordrecht,pp. 279e319.
Shibata, K., 1959. Spectrophotometry of translucence biological materials opal glasstransmission method. Methods of Biochemical Analysis 7, 77e109.
Short, F., Carruthers, T., Dennison, W., Waycott, M., 2007. Global seagrass distribu-tion and diversity: a bioregional model. Journal of Experimental Biology andEcology 350, 3e20.
Silva, J., Santos, R., 2003. Daily variation patterns in seagrass photosynthesis alonga vertical gradient. Marine Ecology Progress Series 257, 37e44.
Terrados, J., Ros, J., 1991. Production dynamics in a macrophyte-dominatedecosystem: the Mar Menor coastal lagoon (SE Spain). Oecologia Aquatica 10,255e270.
Tomasello, A., Di Maida, G., Calvo, S., Pirrotta, M., Borra, M., Procaccini, G., 2009.Seagrass meadows at the extreme of environmental tolerance: the caseof Posidonia oceanica in a semi-enclosed coastal lagoon. Marine Ecology 30,288e300.
Tomasko, D.A., Blake, N.J., Dye, C.W., Hammond, M.A., 2000. Effects of the disposalos reverse osmosis seawater desalination discharges on a seagrass meadow(Thalassia testudinum) offshore of Antigua, West Indies. In: Bortone, S.A. (Ed.),Seagrasses: Monitoring, Ecology, Physiology and Management. CRC Press, BocaRaton, FL, pp. 99e112.
Touchette, B.W., 2007. Seagrassesalinity interactions: physiological mechanismsused by submersed marine angiosperms for a life at sea. Journal of Experi-mental Marine Biology and Ecology 350, 194e215.
Tyerman, S.D., 1989. Solute and water relations of seagrasses. In: Larkum, A.W.D.,Mc Comb, A.J., Sheperd, S.A. (Eds.), Biology of Seagrasses: a Treatise on theBiology of Seagrasses with Special Reference to the Australian Region. Elsevier,Amsterdam, pp. 723e759.
Van Katwijk, M.M., Schmitz, G.H.W., Gasseling, A.P., van Avesaath, P.H., 1999. Effectsof salinity and nutrient load and their interaction on Zostera marina. MarineEcology Progress Series 190, 155e165.
Vargas Yánez, M., García Martínez, M.C., Moya Ruiz, F., Tel, E., Parrilla, G., FraileNuez, E., Lavín, A., García, M.J., 2010. Cambio climático en el Mediterráneoespañol. Colección Temas de Oceanografía, second ed. Instituto Español deOceanografía, Ministerio de Ciencia e Innovación, Madrid, 176 pp.
Verslues, P.E., Agarwal, M., Katiyar-Agarwal, S., Zhu, J., Zhu, J.K., 2006. Methods andconcepts in quantifying resistance to drought, salt and freezing, abiotic stressesthat affect plant water status. Plant Journal 45, 523e539.
Walker, D.A., 1987. The Use of the Oxygen Electrode and Fluorescence Probes inSimple Measurements of Photosynthesis. Oxygraphics, Sheffield, UK.
Yeo, A.R., 1983. Salinity resistance: physiologies and prices. Physiologia Plantarum58, 214e222.
Zar, J.H., 1984. Statistical significance of mutation frequencies, and the power ofstatistical testing using the Poisson-distribution. Biometrical Journal 26, 83e88.
Zhu, J., Meinzer, F.C., 1999. Efficiency of C-4 photosynthesis in Atriplex lentiformisunder salinity stress. Australian Journal of Plant Physiology 26, 79e86.
Zieman, J.C., 1974. Methods for the study of the growth and production of turtlegrass Thalassia testudinum König. Aquaculture 4, 139e143.
Glossary
P vs. E: photosynthesis versus irradiance curvesPAM: pulse amplitude modulated fluorometerPAR: photosynthetic active radiation (l ¼ 400e700 nm)Chla: chlorophyll aChlb: chlorophyll b





























