Embed Size (px)
Citation preview

Journal of Thermal Biology 27 (2002) 17–27
The effect of reproductive condition on thermoregulation ina viviparous gecko from a cool climate
Jennifer Rocka,*, Alison Creea, Robin M. Andrewsb
aDepartment of Zoology, University of Otago, Box 56, Dunedin, New ZealandbDepartment of Biology, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061, USA
Received 19 August 2000; accepted 17 February 2001
Abstract
(1) Hoplodactylus maculatus, is a nocturnal, viviparous gecko that inhabits a cool-temperate region. (2) To determineif pregnant females expend more effort thermoregulating than non-pregnant females or males, we compared field bodytemperature ðTbÞ with microhabitat thermal maxima ðTtopÞ, thermal minima ðTbotÞ and selected temperature ðTselÞamong males, non-pregnant females and pregnant females. (3) An index of thermoregulatory effectiveness that
incorporated diurnal variation in Tsel was used to compare among reproductive conditions. (4) Relationships betweenTb, Ttop and Tbot varied between reproductive conditions and seasons, suggesting complex diurnal thermoregulatorystrategies in this nocturnal species. (5) Pregnant females achieved higher indices of thermoregulatory effort than males
in both seasons, with the highest value in late spring and a slightly lower index similar to that of non-pregnant femalesin summer. # 2002 Elsevier Science Ltd. All rights reserved.
Keywords: Body temperature; Thermoregulation; Nocturnality; Thermoregulatory index; Reproduction; Viviparity; Gekkonidae;
Hoplodactylus maculatus; Seasonal variation; Microhabitat selection
1. Introduction
Behavioural thermoregulation allows reptiles to mod-ify their body temperature ðTbÞ to accommodate theirreproductive status. For example, reproductive (gravid
or pregnant) females may have higher or lower bodytemperatures than non-reproductive females or males.Species that decrease Tb when reproductive tend to have
Tb’s > 308C when non-reproductive. In this situation,modifications of Tb with reproductive status enhanceembryonic survival (Beuchat, 1986; Mathies and An-
drews, 1997). On the other hand, species that increase Tbwhen reproductive tend to have Tb’s of 5308C whennon-reproductive. Under these circumstances, increasedTb accelerates the rate of embryonic development and
thus decreases the total length of gestation (Dorcas and
Peterson, 1998; Rosen, 1991; Slip and Shine, 1988; Tu
and Hutchison, 1994). Enhanced rates of embryonicdevelopment should be particularly beneficial forviviparous species that live in cold climates because thetime that females are burdened with young would be
reduced, and birth could occur at a time most conduciveto offspring survival (Andrews et al., 1997; Shine, 1983).Here, we examine the effects of reproductive condition
on thermoregulation in the common gecko Hoplodacty-lus maculatus. This species should be an ideal subject forsuch a study because of several unusual aspects of its
biology. First, H. maculatus is a member of an all-viviparous lineage of geckos restricted to New Zealandand New Caledonia (Kluge, 1967); geckos elsewhere inthe world are oviparous. The body temperatures of
females will thus affect embryonic development duringtheir entire period of development. Second, somepopulations of H. maculatus occupy very cool
environments (averaging 5188C even at the warmesttime of day in summer) where gestation lasts about 14
*Corresponding author. Fax: +64-3-479-7584.
E-mail address: [email protected]
(J. Rock).
0306-4565/02/$ - see front matter # 2002 Elsevier Science Ltd. All rights reserved.
PII: S 0 3 0 6 - 4 5 6 5 ( 0 1 ) 0 0 0 1 1 - 0

months (Cree, 1994; Cree and Guillette, 1995; Rock,1999). Third, H. maculatus is nocturnal, like most
geckos, and is active at Tb’s as low as 11.38C (Wernerand Whitaker, 1978). While active at night, commongeckos have little opportunity to attain Tb’s that differ
appreciably from ambient temperature. However, lim-ited research on nocturnal lizards has shown that theyare generally capable of some degree of thermoregula-tion during the day, and may achieve Tb’s similar to
some diurnal lizards (Dial, 1978; Kearney and Predavec,2000; Licht et al., 1966; Schlesinger and Shine, 1994).Common geckos are predominantly thigmothermic. In
southern New Zealand (Otago), they inhabit crevices onoutcrops of exfoliating schist rock (tors); they may beable to thermoregulate during the day through the
selection of retreat sites in favourable thermal micro-habitats. Common geckos thus live in habitats where theselection of high temperatures by reproductive females
would be particularly beneficial, but where nocturnalhabits and a paucity of daytime thermal opportunityshould make thermoregulation particularly challenging.Our recent study (Rock et al., 2000) on selected body
temperatures (Tsel; temperatures measured in thelaboratory on a thermal gradient) of common geckosin Otago showed that in summer, pregnant females
exhibit a mean Tsel as much as 88C higher than non-pregnant females and males, and as much as 108C higherthan the mean thermal maxima in gecko microhabitat.
The Tsel of both pregnant females and males is generallyhigher than microhabitat temperature, even during thewarmest times of day (Fig. 1A and B). Limited sampling
of field Tb (on two summer days) showed that commongeckos (particularly pregnant females) often have
difficulty achieving Tsel. Further, these analyses sug-gested that female Tb may be higher than male Tb, andthat both sexes are capable of thermoregulating
maximally to match or surpass some of the maximumtemperatures in their expected microhabitat (Rock et al.,2000).Our objective in the present study was to determine if
pregnant common geckos are able to thermoregulateeffectively in the field, and if this ability changes withstage of pregnancy and season. We addressed two
questions. The first is whether pregnant females havehigher Tb’s than males and non-reproductive femalesduring spring, summer and autumn. The second
question is how reproductive condition affects thermo-regulatory effectiveness. To address this latter question,we used a mathematical index of thermoregulation
based on the Tb a lizard is able to maintain relative toits Tsel, given the available environmental temperature(Hertz et al., 1993). Such thermoregulatory indices havepreviously been used to compare seasonal, geographic
and phylogenetic trends in reptile thermoregulation(Hertz et al., 1993; Christian and Weavers, 1996; D!ııaz,1997; Firth and Belan, 1998; L !oopez et al., 1998; Webb
and Shine, 1998; Kearney and Predavec, 2000). Theyhave not been used, however, to assess differences inthermoregulatory effort associated with reproduction.
We therefore asked, are pregnant geckos more effectiveat thermoregulation than non-pregnant females ormales?
Fig. 1. Variation in microhabitat temperature and selected temperature ðTselÞ with time of day for H. maculatus at Macraes in threeseasons. Microhabitat temperatures are mean� central 50% of values recorded in three seasons (spring=September 1998 and
November 1997; summer=December–February 1997/1998; autumn=March–April 1998). Rock temperatures were measured at 2 h
intervals at two locations with Stowaway data loggers (Onset Computer Corporation, USA) fitted with external probes (for further
description see Rock et al., 2000). Ttoprock=the temperature of the underside of loose rock slab (60� 90� 8 cm l�w� d);
Tbottomrock=the temperature of the substrate rock beneath the top rock. Tsel is central 50% of values recorded at five times of day; data
are extracted from a separate study (Rock et al., 2000).
J. Rock et al. / Journal of Thermal Biology 27 (2002) 17–2718

2. Materials and methods
2.1. Study site and reproductive condition of geckos
The study site at Macraes Flat (hereafter Macraes) is
located in Eastern Otago, South Island, New Zealand at550–710m elevation in a grassland habitat. Commongeckos were captured by hand from crevices in tors.Males and females were differentiated visually (males
have a hemipenial sac), and female reproductivecondition was determined by gentle palpation of theabdominal region for follicles or ova/embryos (Cree and
Guillette, 1995). The reproductive cycle of Macraesgeckos has been described in detail elsewhere (Cree andGuillette, 1995; Rock et al., 2000). In brief, they
reproduce biennially according to the following schedule(Girling and Cree, 1995): In Year 1 of the pregnancycycle, females are in late vitellogenesis or early
pregnancy in early spring (September); they progressthrough pregnancy such that embryos reach about stage34 (Dufaure and Hubert, 1961) by mid-summer, and arefully developed (stage 40) by autumn (March). These
females remain in a late pregnant condition throughautumn and winter, giving birth in the following spring(September–November; Cree and Guillette, 1995; Rock,
1999). In Year 2 of the cycle, females are either in latepregnant or spent condition throughout spring, but byearly summer (December) they are generally all classified
as spent. As summer progresses, the proportion offemales in vitellogenesis increases, such that by autumnall non-pregnant females are vitellogenic. Ovulationoccurs the following spring. In this paper we focus
our analysis on Year 1 pregnant females, Year 2non-pregnant females and males.The reproductive cycle of male common geckos
at Macraes has not been described in detail, butprobably involves year-round spermatocytogenesisand spermiogenesis from summer to early autumn
(December–March; Rock et al., 2000; Cree,unpubl. obs.).
2.2. Measurement of field Tb, and gecko microhabitattemperatures
We measured the Tb of each gecko captured (usuallywithin 15 s, and always within545 s) by inserting a fine-gauge thermocouple thermometer (Cole-Palmer Digi-Sense1; calibrated to � 0.38C against an Ever Readyreference thermometer) approximately 3.5mm into thecloaca. Because of the small size of the geckos(68–85mm snout-to-vent-length (SVL) and 6–14 g)
extreme care was taken to minimise heat transfer. Weshielded the geckos from the wind and sun with a clothbag, and when Tb was being recorded, we held the
animals across the chest and hind limbs as far away fromthe cloaca/lower abdomen as possible. We recorded
most field Tb’s between 1020 and 1630 h, when geckoshave the greatest opportunity for thermoregulation; Tb’s
were obtained for 137 pregnant females, 115 non-pregnant females and 165 males. We collected daytimemeasurements in most months from November 1995
until April 1996, and in September and November of1996. This data we then subdivided by season (spring=September–November, summer=December–February,autumn=March–April). To estimate nocturnal activity
temperatures, we recorded Tb between 2230 and 0130 hin one summer month (December 1995; 7 pregnantfemales, 4 non-pregnant females and 11 males).
We also measured the immediate thermal opportunityfor each gecko caught. Common geckos were capturedfrom crevices under loose slabs of rock on the tors.
Within these crevices, which can be up to 30mm wide,geckos may vary their body posture to increase ordecrease contact with rock surfaces of varying tempera-
tures and so modify their Tb (Muth, 1977; Dial, 1978).Such thigmothermic basking has been observed incaptivity for H. maculatus, which adopts postures toflatten against the substrate, to elevate the body-trunk
off the substrate into the airspace of the crevice, toelevate the dorsum against the top rock, or to adhere tothe upper rock surface and to press the ventral side to
this surface (Rock, 1999). We therefore recorded thetemperature of the undersurface of the top rockimmediately above a gecko at its capture site (hereafter
indicated as Ttop), and the temperature of thebottom rock substrate (hereafter indicated as Tbot)immediately below the gecko to estimate the micro-habitat maximum and minimum for each gecko. Both
Ttop and Tbot were measured immediately after captureof the gecko, with the previously described thermo-couple thermometer.
2.3. Statistical analysis of field Tb and microhabitattemperature
We conducted all statistical analyses using DataDesk1 Version 4.1 (Data Descriptions, Inc., NY). Field
Tb’s were pooled across months within each season(spring, summer, autumn) for each reproductive condi-tion (pregnant female, non-pregnant female and male).
We used three separate two-way ANOVAs (withBonferroni post-hoc tests) to compare means of Tb,Ttop and Tbot among reproductive conditions andseasons. Significance was accepted at P40:05. Indivi-dual regression lines were fitted between Tb, Ttop andTbot at the capture site for each reproductive conditionin each season. To test for differences in slopes and
intercepts among reproductive conditions for eachseason, we performed ANCOVAs using Ttop andTbot as covariates; from these relationships we
inferred differences in thermoregulatory behaviour ofgeckos.
J. Rock et al. / Journal of Thermal Biology 27 (2002) 17–27 19

2.4. Calculation of an index of thermoregulation
For each reproductive condition, we calculated E, anindex representing the effectiveness of thermoregulation.Our calculations of E incorporate diel variation in Tsel,
as this parameter is known to cause significant variationin Tsel in this species (Rock et al., 2000) and in otherspecies of nocturnal geckos (Sievert and Hutchison,1988). Our calculations are thus modified slightly from
those of Hertz et al. (1993) and subsequent studies basedon the E index (e.g. Christian and Weavers, 1996; Webband Shine, 1998), which have assumed that this
parameter is constant.The thermoregulatory index is calculated as
E ¼ 1�dbde
such that E ¼ 0 when there is no thermoregulation andE approaches 1 when intensive thermoregulation isoccurring. The variables db and de are derived fromobservations of Tsel, Tb and Te (operative temperature,Hertz et al., 1993). Tsel was measured in the laboratory
at 1000, 1400 and 1700 h (Rock et al., 2000) and isexpressed here as the upper and lower bounds of thecentral 50% of values recorded at each time of day.
Though we measured Tsel separately in both Januaryand February (mid- and late summer), there was littledifference in diurnal mean values between the two
months (Rock et al., 2000). We therefore pooled valuesfor January and February to calculate a central 50%range at each of three times of day. For Te (an
estimation of temperatures experienced by a non-thermoregulating gecko), we used a randomly selectedvalue from all Tbot’s recorded for each hour.Thus, the overall thermoregulatory index, E, is
composed of derived variables that reflect differentcomponents of thermoregulation. These include db (ameasure of the degree to which a gecko experiences Tboutside its Tsel range) and de (a measure of the thermalquality of the microhabitat). To calculate db for eachgecko, we calculated the absolute value of the deviation
of field Tb from the upper or the lower bound of the Tsel(central 50% range of Tsel measured at the time of daynearest the field Tb capture time). Specifically, we
compared field Tb’s obtained between 1000 and 1200 hto Tsel at 1000 h, Tb’s between 1201 and 1530 h to Tsel at1400 h and Tb’s between 1530 and 1700 h to Tsel at1700 h. Tb’s less than the lower bound of the central
50% of Tsel values were subtracted from the value of thelower bound, and Tb’s greater than the upper bound ofthe Tsel range (though rare) were subtracted from this
value. We assigned db ¼ 0 when Tb was within the upperand lower bounds of Tsel; thus, higher db’s representlarger differences between Tsel and Tb. Similarly, we
calculated values for de by determining absolute valuesfor the deviation of a randomly selected Tbot from the
upper, or more frequently lower, bound of Tsel at thenearest time of day. Consequently, high de values
indicate microhabitats of low thermal quality, i.e.requiring greater thermoregulatory effort for a geckoto achieve Tsel.
The values db, de and E each provide informationabout different aspects of thermoregulatory effective-ness. We therefore calculated their means for eachreproductive condition (pregnant females, non-pregnant
females and males) for late spring and summer. Withineach season, we used one-way ANOVAs (with Bonfer-roni post-hoc tests) to test the effect of reproductive
condition on db, de and E. Significance was accepted atP40:05.
3. Results
3.1. Observations of field Tb
Mean daytime Tb’s across the three seasons and
reproductive conditions ranged from 158C to 258C(Table 1); night-time Tb’s averaged 10.1� 0.38C insummer (December). Mean daytime Tb varied signifi-cantly with reproductive condition (F2; 416 ¼ 3:06,P40:05) and with season (F2; 416 ¼ 37:89, P40:0001).The interaction between reproductive condition andseason was also significant (F2; 416 ¼ 2:56, P40:05).Post-hoc tests revealed that pregnant females weresignificantly warmer than males in spring ðP40:01Þbut not in summer or autumn. There was no significant
difference in Tb between non-pregnant females andeither pregnant females or males in any season.
3.2. Observations of field Tb in relation to microhabitattemperature
To ascertain if there were differences in thermalmicrohabitat selection between reproductive conditions,
we compared seasonal means of thermal minima ðTbotÞand maxima ðTtopÞ recorded at gecko capture sites.Mean Tbot ranged from 138C to 178C, and was almostalways cooler than mean Ttop, which ranged from 158Cto 248C (Table 1). Tbot varied significantly betweenseasons (F2; 416 ¼ 31:84, P40:0001), with spring and
summer Tbot temperatures being significantly warmerthan autumn ðP40:0001Þ. There was no difference inTbot between reproductive conditions and the interactionbetween reproductive condition and season was also not
significant. Similarly, Ttop varied significantly betweenseasons (F2; 416 ¼ 25:12, P40:0001), with no significantdifference between reproductive conditions. However,
the interaction between reproductive condition andseason was significant (F2; 416 ¼ 2:94, P40:05). Inspring, pregnant females had marginally warmer mean
Ttop than males ðP40:05Þ; there was no differencebetween other reproductive conditions in other seasons.
J. Rock et al. / Journal of Thermal Biology 27 (2002) 17–2720
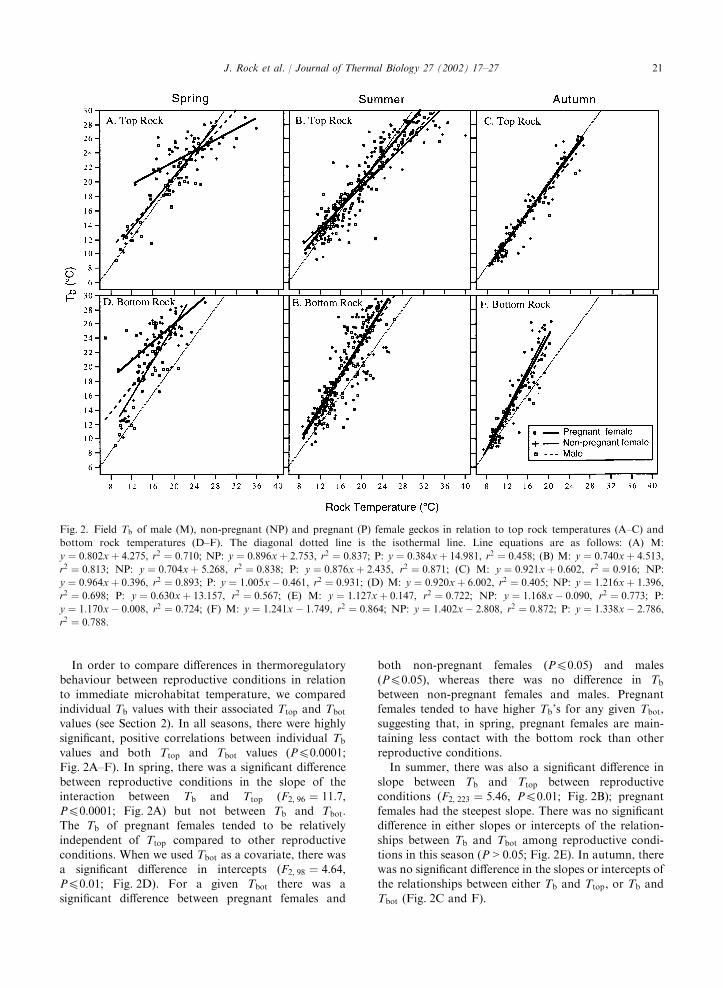
In order to compare differences in thermoregulatorybehaviour between reproductive conditions in relation
to immediate microhabitat temperature, we comparedindividual Tb values with their associated Ttop and Tbotvalues (see Section 2). In all seasons, there were highly
significant, positive correlations between individual Tbvalues and both Ttop and Tbot values (P40:0001;Fig. 2A–F). In spring, there was a significant difference
between reproductive conditions in the slope of theinteraction between Tb and Ttop (F2; 96 ¼ 11:7,P40:0001; Fig. 2A) but not between Tb and Tbot.The Tb of pregnant females tended to be relatively
independent of Ttop compared to other reproductiveconditions. When we used Tbot as a covariate, there wasa significant difference in intercepts (F2; 98 ¼ 4:64,P40:01; Fig. 2D). For a given Tbot there was asignificant difference between pregnant females and
both non-pregnant females ðP40:05Þ and malesðP40:05Þ, whereas there was no difference in Tbbetween non-pregnant females and males. Pregnantfemales tended to have higher Tb’s for any given Tbot,suggesting that, in spring, pregnant females are main-
taining less contact with the bottom rock than otherreproductive conditions.In summer, there was also a significant difference in
slope between Tb and Ttop between reproductiveconditions (F2; 223 ¼ 5:46, P40:01; Fig. 2B); pregnantfemales had the steepest slope. There was no significantdifference in either slopes or intercepts of the relation-
ships between Tb and Tbot among reproductive condi-tions in this season (P > 0:05; Fig. 2E). In autumn, therewas no significant difference in the slopes or intercepts of
the relationships between either Tb and Ttop, or Tb andTbot (Fig. 2C and F).
Fig. 2. Field Tb of male (M), non-pregnant (NP) and pregnant (P) female geckos in relation to top rock temperatures (A–C) and
bottom rock temperatures (D–F). The diagonal dotted line is the isothermal line. Line equations are as follows: (A) M:
y ¼ 0:802xþ 4:275, r2 ¼ 0:710; NP: y ¼ 0:896xþ 2:753, r2 ¼ 0:837; P: y ¼ 0:384xþ 14:981, r2 ¼ 0:458; (B) M: y ¼ 0:740xþ 4:513,r2 ¼ 0:813; NP: y ¼ 0:704xþ 5:268, r2 ¼ 0:838; P: y ¼ 0:876xþ 2:435, r2 ¼ 0:871; (C) M: y ¼ 0:921xþ 0:602, r2 ¼ 0:916; NP:y ¼ 0:964xþ 0:396, r2 ¼ 0:893; P: y ¼ 1:005x� 0:461, r2 ¼ 0:931; (D) M: y ¼ 0:920xþ 6:002, r2 ¼ 0:405; NP: y ¼ 1:216xþ 1:396,r2 ¼ 0:698; P: y ¼ 0:630xþ 13:157, r2 ¼ 0:567; (E) M: y ¼ 1:127xþ 0:147, r2 ¼ 0:722; NP: y ¼ 1:168x� 0:090, r2 ¼ 0:773; P:y ¼ 1:170x� 0:008, r2 ¼ 0:724; (F) M: y ¼ 1:241x� 1:749, r2 ¼ 0:864; NP: y ¼ 1:402x� 2:808, r2 ¼ 0:872; P: y ¼ 1:338x� 2:786,r2 ¼ 0:788.
J. Rock et al. / Journal of Thermal Biology 27 (2002) 17–27 21

For all reproductive conditions in all seasons,coefficients of determination ðr2Þ were generally higherbetween Tb and Ttop than between Tb and Tbot. Tb’stypically fell close to the isothermal line for Ttop in allseasons (Fig. 2A–C). In contrast, almost all Tb values
(regardless of reproductive condition) lay above theisothermal line with Tbot in each season (Fig. 2D–F),suggesting that, by day, geckos (particularly pregnantfemales in spring and summer) engage in behaviours
that enhance contact with the top rock and minimisecontact with the bottom rock.
3.3. Index of thermoregulation
First we graphically compared values for Tb, Tbot and
Tsel in the seasons and months in which Tsel data wereavailable (Rock et al., 2000), i.e. late spring (November)and summer (January–February) (Fig. 3A and B; note
that values for Tb and Tbot are subsets of the datadepicted in Table 1). In late spring, both pregnantfemales and males had high Tsel bands (central
50%=24–298C and 26–288C respectively; Fig. 3A).Mean Tb for pregnant females (248C) was higher thanmean Tb for non-pregnant females (208C) and males(198C) and overlapped with Tsel, whereas mean Tb formales did not. Mean Tbot for pregnant females wasgenerally 28C warmer than for non-pregnant females ormales; mean Tbot values for pregnant females and males
were cooler than the lower limit of Tsel by 9–118C.In summer, Tsel for pregnant females (26–298C) was
consistently higher than that of both non-pregnant
females (21–268C) and males (20–268C) (Fig. 3B). MeanTb for pregnant females (218) was cooler than in latespring, and was 58C lower than the lowest bounds of
Tsel. In contrast, mean Tb for males and non-pregnant
Fig. 3. Values for Tb, Tsel and Tbot between geckos of different
reproductive conditions in months when Tsel was measured (late
spring=November, summer=January and February). Tbvalues are means�SE, Tbot values are means� SE; note that
these values are a subset of data from Table 1. P=pregnant,
NP=non-pregnant; late spring: P females n ¼ 29, NP femalesn ¼ 21, males n ¼ 45; summer: P females n ¼ 63, NP femalesn ¼ 46, males n ¼ 59. Tsel values are central 50%, calculatedfrom Rock et al. (2000). T
able1
MeansofdaytimefieldTbandmicrohabitattemperature(8C)ofH.maculatusfromMacraesa
Season[d]
Spring1995/96[4]
Summer1995/96[8]
Autumn1996[5]
Tb
Microhabitat
Tb
Microhabitat
Tb
Microhabitat
Tbot
Ttop
Tbot
Ttop
Tbot
Ttop
Males
22.0�0.8(50)
15.1�0.5
19.2�0.7
19.1�0.5(88)
16.8�0.4
19.8�0.7
14.5�0.9(27)
13.1�0.7
15.1�0.9
Females
Pregnant
24.1�0.5(29)
17.3�0.7
23.6�1.0
19.4�0.7(79)
16.6�0.5
19.4�0.7
14.8�1.0(29)
13.2�0.7
15.3�1.0
Non-pregnant
20.2�1.3(23)
15.4�0.9
19.2�1.2
20.1�0.7(62)
17.3�0.5
21.1�0.9
14.9�1.0(30)
12.6�0.7
15.1�1.0
aValuesaremeans�SE,withsamplesize(numberofgeckos)giveninparentheses.Withineachseasond=thenumberofsamplingdaysperseason;spring=September–
November,summer=December–February,autumn=March–April.
J. Rock et al. / Journal of Thermal Biology 27 (2002) 17–2722

females (both 218C) was slightly warmer than late springvalues, and converged with Tsel. Mean Tbot for pregnantfemales in summer (178C) remained similar in value tolate spring, whereas mean Tbot for non-pregnant females(178C) and males (188C) was several degrees warmer insummer than in late spring (though both values were still
2–48C cooler than the lower limit of Tsel).The indices of thermoregulation provided additional
insights into gecko behaviour. In spring, db and devalues were significantly lower in pregnant females thanin males (Table 2), confirming trends previouslydescribed in Fig. 3. A high level of thermoregulatory
effectiveness for pregnant females in spring is illustratedby the overall thermoregulatory index, E, which wassignificantly greater for pregnant females than for males(Table 2).
Many of these trends were reversed in summer. Bothdb and de values varied significantly between reproduc-tive conditions (Table 2); however, post-hoc tests
showed that pregnant females had higher values of dband de than either males ðP40:0001Þ or non-pregnantfemales ðP40:0001Þ, with no significant difference
between males and non-pregnant females. These statis-tics also confirm trends seen in Fig. 3, with much of thedifference driven by the high narrow range of Tsel for
pregnant females during summer. Values for E alsovaried between reproductive conditions (Table 2),though not as markedly as in spring; pregnant femaleshad significantly higher values for E than males
ðP40:01Þ, with no significant difference between otherreproductive conditions.
4. Discussion
4.1. Observations of field Tb
Our field measurements confirm and extend observa-tions of low daytime Tb’s for common geckos of the H.
maculatus species complex at Macraes (Rock et al.,2000). Daytime field Tb’s varied seasonally as is oftenobserved in reptiles (Christian et al. (1983) and
Patterson (1992); reviewed in Christian and Bedford(1995) and Sch.aauble and Grigg (1998)). Nevertheless,even in spring and summer when thermoregulatory
opportunities were highest, daytime field Tb’s averagedonly 19–248C. These values are low compared todaytime Tb’s often >258C for many species of nocturnalgeckos (Licht et al., 1966; Valakos, 1989; Werner, 1990;Gil et al., 1994; Brown, 1996).The thermal limitations of the Macraes environment
are further emphasised by low night-time Tb’s. Thoughseveral attempts were made to capture emerged geckosat night, only two of 22 geckos were found emerged; theothers captured at night were beneath rock slabs.
Evidently emergence is irregular and perhaps dependenton warmer nights than we sampled. However, geckoscaptured under rocks at night were alert and mobile at
these low temperatures, despite having a mean Tb ofonly 108C (range 7–138C). Geckos captured in lateautumn (April/May) were also responsive and evasive at
daytime Tb’s as low as 4–78C, and laboratory observa-tions confirm that locomotion is possible at 58C (Rock,1999). Low diurnal field Tb’s, and the ability to be active
at cool night-time Tb’s, appear to be characteristicfeatures of the Gekkonidae (reviewed in Autumn, 1999),and the H. maculatus species complex appears to be aparticularly cold-tolerant group within this family
(Werner and Whitaker, 1978; Tocher, 1992).In our study, pregnant females had a mean Tb slightly
higher than males in spring, but not in summer or
autumn. Further, there was no significant differencebetween mean Tb for pregnant females and non-pregnant females in any season. This apparently limited
influence of reproductive condition on values of meanfield Tb for common geckos at Macraes contrasts withdata for many reptiles. Typically, Tb’s differ betweenmales and pregnant/gravid females and sometimes
Table 2
Means of db, de and E, with statistics for differences between reproductive conditions for two seasons: late spring (N)=November,
summer (J/F)=January and Februarya
Season Males Females Statistics
Pregnant Non-pregnant
Late spring (N) db 6.4� 0.7 0.7� 0.3 } F1, 73=40.692, P40:0001de 10.9� 0.5 6.7� 0.5 } F1, 73=33.567, P40:0001E 0.44� 0.05 0.91� 0.05 } F1, 73=39.7, P40:0001
Summer (J/F) db 2.4� 0.3a 5.6� 0.7b 2.4� 0.4a F2, 167=13.232, P40:0001de 2.6� 0.3a 9.4� 0.5b 3.9� 0.5a F2, 167=75.65, P40:0001E 0.08� 0.23a 0.52� 0.05b 0.37� 0.07a,b F2, 167=5.10, P40:01
aBecause Tsel was not available for non-pregnant females in late spring, db, de and E could not be calculated. Values are
means� SE; statistics are from one-way ANOVAs. Values with different letters differ significantly ðP40:05Þ.
J. Rock et al. / Journal of Thermal Biology 27 (2002) 17–27 23

between pregnant and non-pregnant females (Gravesand Duvall, 1993; Tu and Hutchison, 1994; Bl!aazquez,
1995; Dorcas and Peterson, 1998). Most of these species,however, are from warm temperate regions whereprecise thermoregulation is achievable. The small effect
of reproductive condition on mean daytime Tb’s atMacraes may reflect an environmental situation in whichthermoregulatory behaviour cannot typically compen-sate for low ambient temperature. This does not mean,
however, that thermoregulation is ineffective, and wewill consider the complex thermoregulatory strategiesexhibited by common geckos at Macraes in the
remainder of this discussion.
4.2. Observations of field Tb in relation to microhabitat
temperature
Our study revealed variation in the effect of micro-
habitat temperature on diurnal field Tb; it demonstratesthe importance of considering both factors whenevaluating thermoregulation in a nocturnal lizard. In
each season, mean values for daytime field Tb’s forcommon geckos were close to mean Ttop values andseveral degrees above mean Tbot values. These compar-isons suggest that daytime Tb’s of geckos are more
strongly influenced by the temperature of the rock abovethem than by the temperature of the rock below them.Furthermore, the difference in mean Tb between
pregnant females and males in spring is associated witha tendency for pregnant females to select retreat siteswith slightly warmer Ttop (perhaps thinner rocks).
Regression analysis confirmed that daytime Tb washighly correlated with both Ttop and Tbot, but in almostall cases the association was stronger with Ttop. DaytimeTb had a 1 : 1 relationship with Ttop, but typically
diverged from Tbot as conditions became warmer. Theseobservations imply that, as conditions warm up, geckosincrease their contact with the rock above them and
minimise contact with the substrate (e.g. by standing ontheir toes and pushing their dorsum against the rockabove). Similar thermoregulatory postures have been
observed for other nocturnal geckos. For example,Gehyra variegata basks indirectly under bark on treetrunks during the day and adopts bark or trunk
temperatures by changing its body posture (Bustard,1967). In both field and lab studies Coleonyx brevis hasbeen observed to thermoregulate by maximising contactwith warm rock surfaces directly overhead in a crevice
(Dial, 1978). On the occasion that Ttop becomesextremely warm at Macraes (32–408C), Tb’s fall belowTtop, suggesting that common geckos reduce contact
with the rock above as an upper level of thermalpreference is reached. The maximum values for daytimefield Tb’s of 28–308C are in accord with maximum Tselvalues of 288C recorded for this population (Rock et al.,2000).
This and a few other studies (Dial, 1978; Licht et al.,1966; Schlesinger and Shine, 1994; Kearney and
Predavec, 2000) demonstrate that, just because theyare not visible, researchers cannot assume that nocturnallizards do not engage in daytime thermoregulatory
behaviour. Further, the placement of copper models(Spencer and Grimmond, 1994) or data loggers (Vitt andSartorius, 1999) in the air spaces under rocks is unlikelyto provide reliable estimates of daytime body tempera-
ture because such models cannot replicate the variablepostures and movements used by nocturnal geckoswithin retreat sites.
Our study is one of the first to demonstrate thatreproductive condition affects the diurnal thermoregula-tion of nocturnal geckos. In spring, the correlation
between daytime Tb and retreat site temperature incommon geckos varied significantly among reproductiveconditions. Pregnant females had a shallower slope in
the relationship between Tb and Ttop than did non-pregnant females or males, although mean Ttop selectedby pregnant females was significantly warmer than Ttopselected by other reproductive conditions. In other
words, daytime Tb was more independent of Ttop forpregnant females than for males and non-pregnantfemales. Pregnant females also had a higher intercept in
the relationship between Tb and Tbot than did males andnon-pregnant females. These observations imply thatpregnant females in spring are selecting warmer sites and
achieving higher Tb’s than males and non-pregnantfemales, but doing it by other means than by simplymatching Tb to Ttop. They may move within or betweenretreat sites more frequently, maintain less contact with
the substrate and/or engage more readily in discreetsolar basking at the edge of their crevices in order tomaintain Tb’s within a narrower range. Discreet basking
has occasionally been observed in the wild for commongeckos (Werner and Whitaker, 1978; J. Rock, pers. obs.)and for other species of nocturnal geckos (Gil et al.,
1994; Brown, 1996), but whether this behaviour varieswith season or reproductive condition has not yet beentested. In both summer and autumn, the temperature of
microhabitats selected did not differ between reproduc-tive conditions. However, in summer pregnant femaleshad Tb’s significantly closer to Ttop than did non-pregnant females or males. This suggests that they were
thermoregulating maximally within their crevice relativeto males and non-pregnant females. These seasonaldifferences observed for pregnant females but not non-
pregnant females or males suggest that some aspect ofthe progression of embryogenesis affects thermoregula-tion in H. maculatus.
4.3. Index of thermoregulation
Our study is the first to use the E index to evaluate theeffect of reproductive condition on the effectiveness of
J. Rock et al. / Journal of Thermal Biology 27 (2002) 17–2724

thermoregulation. Components of the index of thermo-regulation provide a good synopsis of thermoregulatory
effectiveness between pregnant females, non-pregnantfemales and male common geckos. In particular, theyconfirm that field Tb’s often fall substantially below Tsel(central 50%) as predicted for this species (Rock et al.,2000) and observed in other species of nocturnal geckos(Licht et al., 1966; Gil et al., 1994; Brown, 1996;Angilletta and Werner, 1998; Christian et al., 1998).
Mean Tb for H. maculatus overlapped with Tsel in only afew instances, and the overlap was limited to the bottomend of the Tsel band. Values for db and de (Tsel � Tb and
Tsel � Tbot; respectively) indicate that in late spring, bothTb and microhabitat temperatures for pregnant femaleswere generally closer to Tsel than were those of males.
The E index, which combines db and de, illustrates thateffectiveness of thermoregulation is dramatically higherfor pregnant females than males in late spring. Because
Tsel was not available for non-pregnant females inspring, a full comparison between reproductive condi-tions cannot be made. However, the similarity of Tb andTbot values between non-pregnant females and males,
relative to values for pregnant females, suggests that anE index for non-pregnant females would not be as highfor pregnant females in this season.
By summer, db and de values indicate that males andnon-pregnant females achieved Tb’s significantly closerto Tsel, with greater similarity between Tsel and Tbot;than did pregnant females. Although values of E forboth pregnant females and males are much lower insummer than in late spring, pregnant females achievedhigher indices of thermoregulatory effectiveness than
males. Though there was no significant differencebetween pregnant females and non-pregnant females insummer, non-pregnant females did not have E values
significantly different from those of males. Thus in bothseasons, pregnant females expended greater thermore-gulatory effort to maintain high Tb’s relative to their
desired Tsel under the available microhabitat tempera-tures.An unexpected finding of this study was that in
summer, when Tsel for pregnant females is at its highestand least variable, mean Tb for pregnant females wascooler than in late spring; this contributed to a lowervalue for E. Seasonal variation in thermoregulatory
effectiveness (measured with a similar index of thermo-regulation) also occurs in the nocturnal gecko, Christi-nus marmoratus (Kearney and Predavec, 2000).
However, in this species the highest E index (for malesand females combined) occurs in summer. The Tb valuefor H. maculatus may be strongly influenced by the
vagaries of weather at Macraes, and thus temperaturesat the retreat site, at the times and days of sampling.Though continuous data logging of rock temperatures in
a retreat site confirms that daytime temperatures aregenerally warmer in summer than in spring (Fig. 1),
there is considerable variation among days and thermalconditions can change dramatically even within a day
(Rock et al., 2000). Clearly, the most informative studyof thermoregulation in common geckos would begenerated by continuous monitoring both of microha-
bitat with numerous data loggers monitoring many rocksurfaces and air spaces within the tor complexes, and ofTb with radio telemetry implants. Unfortunately, theextremely heterogeneous and stochastic thermal envir-
onment, and small body size and crepuscular habits ofcommon geckos make such a study extremely difficult.We can, however, make some predictions about
thermoregulatory strategy for common geckos in coldclimates such as Macraes.
4.4. Thermoregulatory strategies of H. maculatus
Common geckos at Macraes have developed complex
thermoregulatory strategies to deal with the overall lowtemperature. They are not simply substrate thermo-conformers, as they have been described in the past
(Spencer and Grimmond, 1994). Our data suggest thatcommon geckos are diurnal thigmotherms, employing avariety of thermoregulatory behaviours including pos-tural or positional adjustments within their retreat sites,
movements between retreat sites and opportunisticmarginal basking. The effectiveness of such thermo-regulation varies with reproductive condition in com-
mon geckos. Pregnant females, rather than beingencumbered (as has been suggested for other viviparouslizards, Andrews et al., 1997) are generally more effective
at thermoregulation than other reproductive conditions.The value of maternal thermoregulation in viviparousspecies is well known for diurnal lizards (e.g. Shine andHarlow, 1993). Our data suggest that in a thermally
challenging habitat such as Macraes, thermoregulationis particularly critical for pregnant geckos to maintain asuitable Tb for embryogenesis. This premise is supported
by a laboratory study in which cool temperatures(daytime maximum 208C) resulted in lower success ofpregnancies of Macraes geckos (i.e. higher rates of
resorptions, abortions or stillbirths) than higher tem-peratures (Rock, 1999).Despite the value of thermoregulation for geckos,
particularly pregnant females, at cool sites such asMacraes, it is unclear what thermoregulatory strategywould be most effective to achieve maximal thermo-regulation. The thermal microhabitat within tor com-
plexes is extremely heterogeneous, both within andbetween days, in the maximum and minimum valuesreached and the duration of these values (Rock et al.,
2000). For a gecko, this means that position in the torcomplex at any particular time is critical to theeffectiveness of thermoregulation, both in amplitude
and duration of Tb values attained. One thermoregula-tory strategy might be to gamble on finding high
J. Rock et al. / Journal of Thermal Biology 27 (2002) 17–27 25

temperatures during the day under thin rocks, whichwould heat relatively rapidly under sunny conditions.
However, geckos in this microhabitat would also riskeither overheating during the hottest part of some days,or risk predation when shuttling to cooler retreats.
Further, if they remained associated with thin rocksovernight, they would experience colder temperaturesthan those typically available in deep rock creviceswhich typically experience a lag in cooling after dark
(Werner, 1990). An alternative strategy would be forgeckos to hedge their bets and remain in more uniformlywarm microhabitats (under thicker rocks, perhaps
moving upwards within the tor complex if several hoursof warm temperatures occurred). Such an opportunisticstrategy has been observed in gravid Thamnophis elegans
(Huey et al., 1989), although this species inhabits a morethermally hospitable habitat in which accurate thermo-regulation during pregnancy may be less critical than it
is for H. maculatus. Retreat site selection by lizards canbe influenced by many factors (Huey et al., 1989). Inother nocturnal geckos, factors such as microclimaticand social quality (Schlesinger and Shine, 1994), as well
as predation risk (Downes and Shine, 1998) are knownto influence retreat selection. For H. maculatus, selectionof retreat sites for optimal thermal qualities may also be
compromised by the site’s proximity to food sources,e.g. fruiting shrubs are most accessible to geckos that arenot sheltered deep within the tor complex (J. Rock, pers.
obs.). Further, the likelihood of disturbance also mayaffect selection of retreat site; much of the tor habitat ofcommon geckos is grazed by stock that, at highdensities, disturb loose rocks on and around tors (J.
Rock, pers. obs.).In conclusion, though heat flux in a rocky micro-
habitat is complex, nocturnal ectotherms in retreats
within this habitat can thermoregulate effectively.Accordingly, the animal’s position within the rockhabitat should be directly associated with its thermo-
regulatory need (Huey et al., 1989). This need may varywith seasonal ecological constraints, such as thosediscussed above, or be driven by endogenous physiolo-
gical cycles, that can vary seasonally and daily (Firthand Belan, 1998). In nocturnal geckos, both metabolismand optimal growth rates have been shown to bedependent on diurnal thermoregulation involving thig-
mothermic basking (Bustard, 1967; Autumn and DeNardo, 1995). These processes, in turn, may haveextensive effects on geographic range and life history
characteristics, such that nocturnal lizards in coolclimates are predicted to be slow growing and long-lived (Autumn and De Nardo, 1995). H. maculatus
fulfils this prediction as it has slow growth rates andgreat longevity, as well as one of the slowest reproduc-tive cycles known amongst reptiles (Cree, 1994; Cree and
Guillette, 1995; Rock, 1999). Our study demonstratesthat reproductive condition places significant demands
on the efficiency of thermoregulation in nocturnalreptiles.
Acknowledgements
Research was supported by the Division of Sciences,University of Otago to AC and JR, and the NationalScience Foundation grant BR-9022425 to RMA. We
thank K. and M. Philip for access to field sites, theDepartment of Conservation for permits, and thevolunteers who helped us capture geckos. Funding for
HOBO and Stowaway data loggers was provided by theMiss E.L. Hellaby Indigenous Grasslands ResearchTrust. We especially thank R. Shine and M. Kearney
for comments on the content of this manuscript, andtwo anonymous reviewers for their valuable suggestions.
References
Andrews, R.M., Mendez de la Cruz, F.R., Santa Cruz, M.V.,
1997. Body temperatures of female Sceloporus grammicus:
thermal stress or impaired mobility? Copeia 1, 108–115.
Angilletta Jr., M.J., Werner, Y.L., 1998. Australian geckos do
not display diel variation in thermoregulatory behavior.
Copeia 3, 736–742.
Autumn, K., 1999. Secondarily diurnal geckos return to cost of
locomotion typical of diurnal lizards. Physiol. Biochem.
Zool. 72, 339–351.
Autumn, K., De Nardo, D.F., 1995. Behavioral thermoregula-
tion increases growth rate in a nocturnal lizard. J. Herpetol.
29, 157–162.
Beuchat, C., 1986. Reproductive influences on the thermo-
regulatory behaviour of a live-bearing lizard. Copeia 4,
971–979.
Bl!aazquez, M.C., 1995. Body temperature, activity patterns and
movements by gravid and non-gravid females of Malpolon
monspessulanus. J. Herpetol. 29, 264–266.
Brown, R.P., 1996. Thermal biology of the gecko Tarentola
boettgori: comparisons among populations from different
elevations within Gran Canaria. Herpetologica 52, 396–405.
Bustard, H.R., 1967. Activity cycle and thermoregulation in the
Australian gecko Gehyra variegata. Copeia 4, 753–758.
Christian, K.A., Bedford, G., 1995. Seasonal changes in
thermoregulation by the frillneck lizard, Chlamydosaurus
kingii, in tropical Australia. Ecology 76, 124–132.
Christian, K.A., Bedford, G., Green, B., Schultz, T., Newgrain,
K., 1998. Energetics and waterflux of the marbled velvet
gecko (Oedura marmorata) in tropical and temperate
habitats. Oecologia 116, 336–342.
Christian, K.A., Tracy, C.R., Porter, W.P., 1983. Seasonal
shifts in body temperature and use of microhabitats by
Galapagos land iguanas (Conolophus pallidus). Ecology 64,
463–468.
Christian, K.A., Weavers, B.W., 1996. Thermoregulation of
monitor lizards in Australia: an evaluation of methods in
thermal biology. Ecol. Monogr. 66, 139–157.
J. Rock et al. / Journal of Thermal Biology 27 (2002) 17–2726

Cree, A., 1994. Low annual reproductive output in female
reptiles from New Zealand. N. Z. J. Zool. 21, 351–372.
Cree, A., Guillette Jr., L.J., 1995. Biennial reproduction with a
fourteen-month pregnancy in the gecko Hoplodactylus
maculatus from southern New Zealand. J. Herpetol. 29,
163–173.
Dial, B.E., 1978. The thermal ecology of two sympatric,
nocturnal Coleonyx (Lacertilia: Gekkonidae). Herpetologica
34, 194–201.
D!ııaz, J.A., 1997. Ecological correlates of the thermal quality of
an ectotherm’s habitat: a comparison between two temperate
lizard populations. Functional Ecol. 11, 79–89.
Dorcas, M.E., Peterson, C.R., 1998. Daily body temperature
variation in free-ranging rubber boas. Herpetologica 54,
88–103.
Downes, S., Shine, R., 1998. Heat, safety or solitude? Using
habitat selection experiments to identify a lizard’s priorities.
Anim. Behav. 55, 1387–1396.
Dufaure, J.P., Hubert, J., 1961. Table de d!eeveloppement du
l!eezard vivipare: Lacerta (Zootoca) vivipara Jacquin. Arch.
Anat. Microscop. Morphol. Exp. 50, 309–328.
Firth, B.T., Belan, I., 1998. Daily and seasonal rhythms in
selected body temperatures in the Australian lizard Tiliqua
rugosa (Scincidae): field and laboratory observations.
Physiol. Zool. 71, 303–311.
Gil, M.J., Guerrero, F., P!eerez-Mellado, V., 1994. Diel variation
in preferred body temperatures of the Moorish gecko
Tarentola mauritanica during summer. Herpetol. J. 4, 56–59.
Girling, J.E., Cree, A., 1995. Plasma corticosterone levels are
not significantly related to reproductive stage in female
common geckos (Hoplodactylus maculatus). Gen. Comp.
Endocrinol. 100, 273–281.
Graves, B.M., Duvall, D., 1993. Reproduction, rookery use,
and thermoregulation in free-ranging, pregnant Crotalus
v.viridis. J. Herpetol. 27, 33–41.
Hertz, P.E., Huey, R.B., Stevenson, R.D., 1993. Evaluating
temperature regulation by field-active ectotherms: the fallacy
of the inappropriate question. Am. Nat. 142, 796–818.
Huey, R.B., Peterson, C.R., Arnold, S.J., Porter, W.P., 1989.
Hot rocks and not-so-hot rocks: retreat-site selection by
garter snakes and its thermal consequences. Ecology 70,
931–944.
Kearney, M., Predavec, M., 2000. Do nocturnal ectotherms
thermoregulate? A study of the temperate gecko Christinus
marmoratus. Ecology 81, 2984–2996.
Kluge, A.G., 1967. Higher taxonomic categories of Gekkonid
lizards and their evolution. Bull. Am. Mus. Nat. Hist. 135,
1–59.
Licht, P., Dawson, W.R., Shoemaker, H., Main, A.R., 1966.
Observations on the thermal relations of Western Australian
lizards. Copeia 1, 97–110.
L !oopez, P., Salvador, A., Mart!ıın, J., 1998. Soil temperature, rock
selection, and the thermal ecology of the amphisbaenian
reptile Blanus cinereus. Can. J. Zool. 76, 673–679.
Mathies, T., Andrews, R.M., 1997. Influence of pregnancy on
the thermal biology of the lizard, Sceloporus jarrovi: why do
pregnant females exhibit low body temperatures? Functional
Ecol. 11, 498–507.
Muth, A., 1977. Body temperatures and associated postures of
the zebra-tailed lizard, Callisaurus draconoides. Copeia 1,
122–125.
Patterson, J.W., 1992. Seasonal variation in field body
temperatures of the lizard Mabuya striata punctatissima.
Amphibia–Reptilia 13, 243–250.
Rock, J., 1999. The relationship between thermal biology and
female reproduction in the viviparous gecko, Hoplodactylus
maculatus. Unpublished Ph.D. Thesis, University of Otago,
Dunedin, New Zealand.
Rock, J., Andrews, R.M., Cree, A., 2000. Effects of reproduc-
tive condition, season and site on selected temperatures of a
viviparous gecko. Physiol. Biochem. Zool. 73, 344–355.
Rosen, P.C., 1991. Comparative field study of thermal
preferenda in garter snakes (Thamnophis). J. Herpetol. 25,
301–312.
Sch.aauble, C.S., Grigg, G.C., 1998. Thermal ecology of the
Australian agamid Pogona barbata. Oecologia 114,
461–470.
Schlesinger, C.A., Shine, R., 1994. Selection of diurnal retreat
sites by the nocturnal gekkonid lizard Oedura lesueurii.
Herpetologica 50, 156–163.
Shine, R., 1983. Reptilian reproductive modes: the oviparity–
viviparity continuum. Herpetologica 39, 1–8.
Shine, R., Harlow, P., 1993. Maternal thermoregulation
influences offspring viability in a viviparous lizard. Oecologia
96, 122–127.
Sievert, L.M., Hutchison, V.H., 1988. Light versus heat:
thermoregulatory behaviour in a mocturnal lizard (Gekko
gecko). Herpetologica 44, 266–273.
Slip, D., Shine, R., 1988. Reptilian endothermy: a field study of
thermoregulation by brooding diamond pythons. J. Zool.
216, 367–378.
Spencer, N.J., Grimmond, N.M., 1994. Influence of elevation
on the thermoregulation of two sympatric lizards. N. Z. J.
Zool. 21, 379–385.
Tocher, M.D., 1992. Paradoxical preferred body temperatures
of two allopatric Hoplodactylus maculatus (Reptilia: Gekko-
nidae) populations from New Zealand. N. Z. Nat. Resour.
19, 53–60.
Tu, M., Hutchison, V.H., 1994. Influence of pregnancy on
thermoregulation of water snakes (Nerodia rhumbifera). J.
Therm. Biol. 19, 255–259.
Valakos, E., 1989. Thermal ecology of Crytodactylus kotschyi
(Steindachner, 1870) (Sauria-Gekkonidae) in the insular
ecosystems of the Aegean. Herpetol. J. 1, 396–399.
Vitt, L.J., Sartorius, S.S., 1999. HOBOs, Tidbits and lizard
models: the utility of electronic devices in field studies of
ectotherm thermoregulation. Functional Ecol. 13, 670–674.
Webb, J.K., Shine, R., 1998. Thermoregulation by a nocturnal
elapid snake (Hoplocephalus bungaroides) in southeastern
Australia. Physiol. Zool. 71, 680–692.
Werner, Y.L., 1990. Habitat dependent thermal regimes of two
Hawaiian geckos (Reptilia: Gekkonidae). J. Therm. Biol. 15,
281–290.
Werner, Y.L., Whitaker, A.H., 1978. Observations and com-
ments on the body temperatures of some New Zealand
reptiles. N. Z. J. Zool. 5, 375–393.
J. Rock et al. / Journal of Thermal Biology 27 (2002) 17–27 27





![Neonatal Thermoregulation - University of · PDF fileNeonatal Thermoregulation Julia Petty. ... A care study. Journal of Neonatal Nursing. ... 5 Thermoregulation [Compatibility Mode]](https://img.pdfslide.us/doc/110x75/5aafe83f7f8b9a6b308de3c0/neonatal-thermoregulation-university-of-thermoregulation-julia-petty-a-care.jpg)























