Embed Size (px)
Citation preview

The effect of additives on red cell 2,3 diphosphoglycerate levels in CPDA preservatives
s. VORA, c. WEST, AND E. BEUTLER
Forty-two chemical substances, chosen because they might influence red cell metab- olism, were screened for effect on red cell adenosine triphosphate and 2,3 diphos- phoglycerate (2,3 DPG) levels during storage in CPD or CPDA-1 at 4°C. Of these substances, six appeared on initial screening to increase 2,3 DPG levels during storage; on repeated examination, four compounds, i.e., oxalate, glyoxalate, ethyl oxaloacetate, and L-phenylalanyl-L-alanine, consistently increased 2,3 DPG levels during storage. It was shown that glyoxalate was converted rapidly to oxalate in blood, presumably through the lactate dehydrogenase reaction. Ethyl oxaloacetate is known to hydrolyze, giving rise to oxalate. Thus, the effect of both glyoxalate and ethyl oxaloacetate can be explained by the formation of oxalate, a compound already known to increase 2,3 DPG levels. The effect of L-phenylalanyl-L-alanine remains to be explained, but it may be hydrolyzed to L-alanine and L-phenylalanine, both of which are thought to have the capacity to in- crease red cell 2,3 DPG levels by inhibiting pyruvate kinase activity. TRANSFUSION 1989:29:226-229.
SINCE THE DISCOVERY that 2,3 diphosphoglycerate (2,3 DPG) modulates the oxygen dissociation curve of hemoglobin,',* many efforts have been made to design preservative solutions that prevent the rapid decline in the levels of this sugar phosphate in liquid-stored red cells (RBCs). Some of the modifications that have been introduced, such as the addition of inosine3 phosphoen~lpyruvate~ or modulation of the pH,S have been based upon well-established metabolic path- ways. Other additives have been discovered serendip- itously. Thus, dihydroxyacetone was added to RBCs6 before it was appreciated that these cells contained triokinase, the enzyme that phosphorylates this sugar to dihydroxyacetone phosphate,' a precursor of 2,3 DPG. Xanthone derivatives were known to have a direct effect on hemoglobin, and the fact that they improved 2,3 DPG preservation came as a surprise,8 which was explained later through the inhibitory ef- fect of these compounds on 2,3 DPG pho~phatase.~ The striking effect of oxalate on 2,3 DPG levels was appreciated only when it was found to be the active principle in ascorbate, in which it was present as a contaminant.10
With the hope of finding other substances that may be useful in maintaining 2,3 DPG levels during storage, we screened a large number of compounds
From the Department of Basic and Clinical Research, Research Institute of Scripps Clinic, La Jolla, California.
Supported in part by National Institute of Health grants RR00833 and HL25552 and by U.S. Army Medical Research Acquisition Activity Contract No. DAMD- 17-87-C-7074.
The views, opinions and/or findings contained in this report are those of the author(s) and should not be construed as an official Department of the Army position, policy, or decision unless so designated by other documentation.
Received for publication April 1, 1988; revision received July 28, 1988, and accepted August 18, 1988.
for their effect on RBC 2,3 DPG and adenosine triphosphate (ATP) levels of blood stored in CPDA- I solution. Some of the substances were chosen be- cause they might be inhibitors of pyruvate kinasel' and some because they might compete at the acti- vation site of diphosphoglycerate phosphatase with 2-phosphoglycolate, the most potent activator of the 2,3 DPG-degrading enzyme.
Materials and Methods In the initial screening studies, blood from normal volun-
teer donors was drawn into CPD or CPDA- 1 solution (Fen- wal Laboratories, Deerfield, IL). The blood was divided into four nearly equal portions and was distributed into plastic blood bags. The volume of each aliquot was measured by weighing the bag; minor differences in volume were cor- rected by the addition of 0.9 percent NaCl solution. The substances being studied were added to each of the aliquots in the saline. Samples without additions served as controls. The samples were mixed weekly. ATP and 2,3 DPG de- terminations were carried out weekly or semiweekly for at least 6 weeks by standard methods.12
In subsequent studies, 450 ml of blood was drawn into 6 3 ml of CPDA- 1, divided into 95- or 99-ml portions, and distributed into plastic blood bags. The substance under study was diluted in 1- or 5-ml 0.9 percent NaCl and added to each 95- or 99-ml aliquot of blood. Saline without preservative was added to the control.
The blood samples were incubated at 37°C for 1 to 1.5 hours before storage. The internal pH of RBCs was mea- sured by freezing and thawing packed RBCs and measuring the pH of the lysate at 0°C with a pH meter (Corning model 150, Corning Glass Works, Scientific Instruments, Medford, MA). Oxalate determinations3 were carried out on plasma ultrafiltrates made from blood with added glyoxalate before storage and after I week.
Results Table 1 lists the 42 substances screened. In the prelim-
inary screening studies, oxalate, glyoxalate, ethyl oxaloac-
226
TRANSFUSION 1989-Vol. 29, No. 3
EFFECT OF ADDITIVES TO CPDA ON 2.3 DPG 221
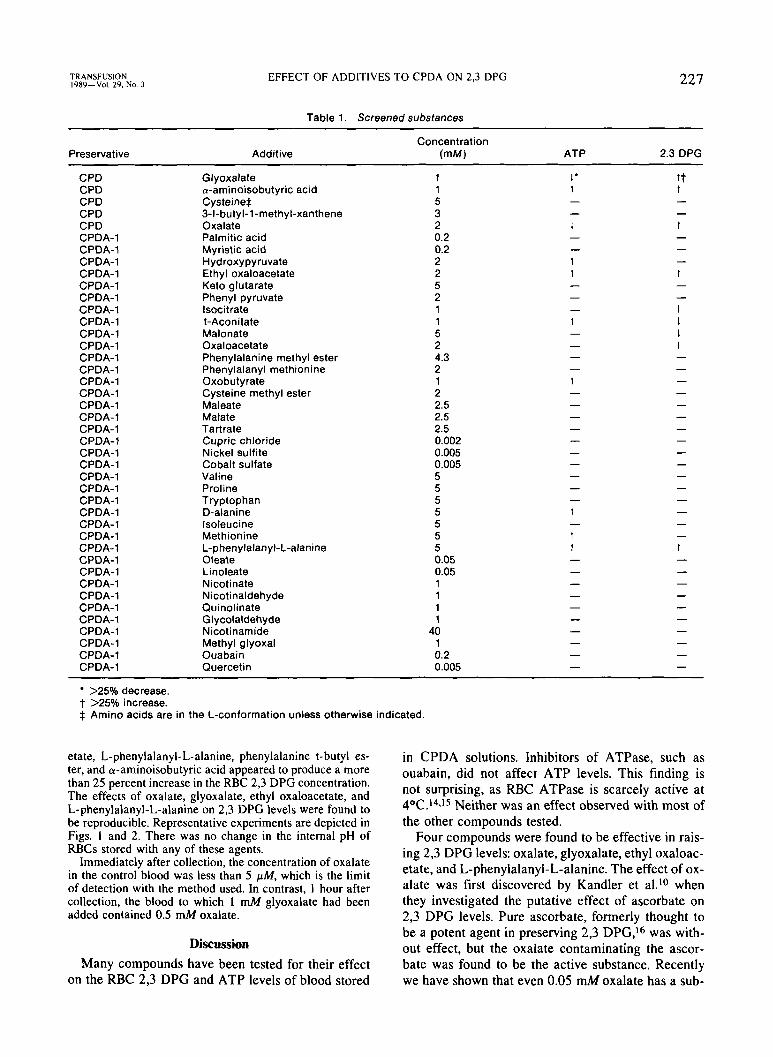
Table 1. Screened substances
Preservative Add it ive Concentration
(mM) ATP 2.3 DPG
CPD Glyoxalate 1 1' tt CPD a-aminoisobutyric acid 1 t t CPD CysteineS 5 CPD 3-I-butyl-1 -methyl-xanthene 3 CPD Oxalate 2 1 1 CPDA-1 Palmitic acid 0.2 CPDA-1 Myristic acid 0.2 CPDA-1 Hydroxypyruvate 2 1 CPDA-1 Ethyl oxaloacetate 2 t t CPDA-1 Keto glutarate 5 CPDA-1 Phenyl pyruvate 2
1 CPDA-1 lsocitrate 1 CPDA-1 t-Aconitate 1 t 1
1 CPDA-1 Malonate 5 1 CPDA-1 Oxaloacetate 2
CPDA-1 Phenylalanine methyl ester 4.3 CPDA-1 Phenylalanyl methionine 2 CPDA-1 Oxobutyrate 1 1 CPDA-1 Cysteine methyl ester 2
- - - -
- - - -
-
- - - - -
- - - - - -
- - - - - CPDA-1 Maleate 2.5
CPDA-1 Malate 2.5 CPDA-1 Tartrate 2.5 CPDA-1 Cupric chloride 0.002 CPDA-1 Nickel sulfite 0.005 CPDA-1 Cobalt sulfate 0.005 - - CPDA-1 Valine 5 CPDA-1 Proline 5 CPDA-1 Tryptophan 5 CPDA-1 D-alanine 5 t CPDA-1 lsoleucine 5 CPDA-1 Methionine 5 t CPDA-1 L-phenylalanyl-L-alanine 5 t t CPDA-1 Oleate 0.05 CPDA-1 Linoleate 0.05 CPDA-1 Nicotinate 1 CPDA-1 Nicotinaldehyde 1 CPDA-1 Quinolinate 1 CPDA-1 Glycolaldehyde 1 CPDA-1 Nicotinamide 40 CPDA-1 Methyl glyoxal 1 CPDA-1 Ouabain 0.2 CPDA-1 Quercetin 0.005
>25% decrease. t >25% increase. $ Amino acids are in the L-conformation unless otherwise indicated.
- - - - - - - - - - - - - -
- - -
-
- - - - - - - - - - - - - - - - - - - -
etate, L-phenylalanyl-L-alanine, phenylalanine t-butyl es- ter, and a-aminoisobutyric acid appeared to produce a more than 25 percent increase in the RBC 2,3 DPG concentration. The effects of oxalate, glyoxalate, ethyl oxaloacetate, and L-phenylalanyl-L-alanine on 2,3 DPG levels were found to be reproducible. Representative experiments are depicted in Figs. 1 and 2. There was no change in the internal pH of RBCs stored with any of these agents.
Immediately after collection, the concentration o f oxalate in the control blood was less than 5 pM, which i s the limit o f detection with the method used. In contrast, 1 hour after collection, the blood to which 1 mM glyoxalate had been added contained 0.5 mM oxalate.
Discussion Many compounds have been tested for their effect
on the RBC 2,3 DPG and ATP levels of blood stored
in CPDA solutions. Inhibitors of ATPase, such as ouabain, did not affect ATP levels. This finding is not surprising, as RBC ATPase is scarcely active at 40C.14J5 Neither was an effect observed with most of the other compounds tested.
Four compounds were found to be effective in rais- ing 2,3 DPG levels: oxalate, glyoxalate, ethyl oxaloac- etate, and L-phenylalanyi-L-alanine. The effect of ox- alate was first discovered by Kandier et al.1° when they investigated the putative effect of ascorbate on 2,3 DPG levels. Pure ascorbate, formerly thought to be a potent agent in preserving 2,3 DPG,16 was with- out effect, but the oxalate contaminating the ascor- bate was found to be the active substance. Recently we have shown that even 0.05 mM oxalate has a sub-
228 VORA, WEST, AND BEUTLER TRANSFUSION Vol. 29. No. 3-1989
g15
0 1 0 2 0 30 40
t Control 0 2 mM Et-Oxalacetate + 2 . 5 mM Phenylalanyl-ala
$m k
3 0 < 1 V
0 1 0 20 30 40 50 STORAGE TIME (DAYS)
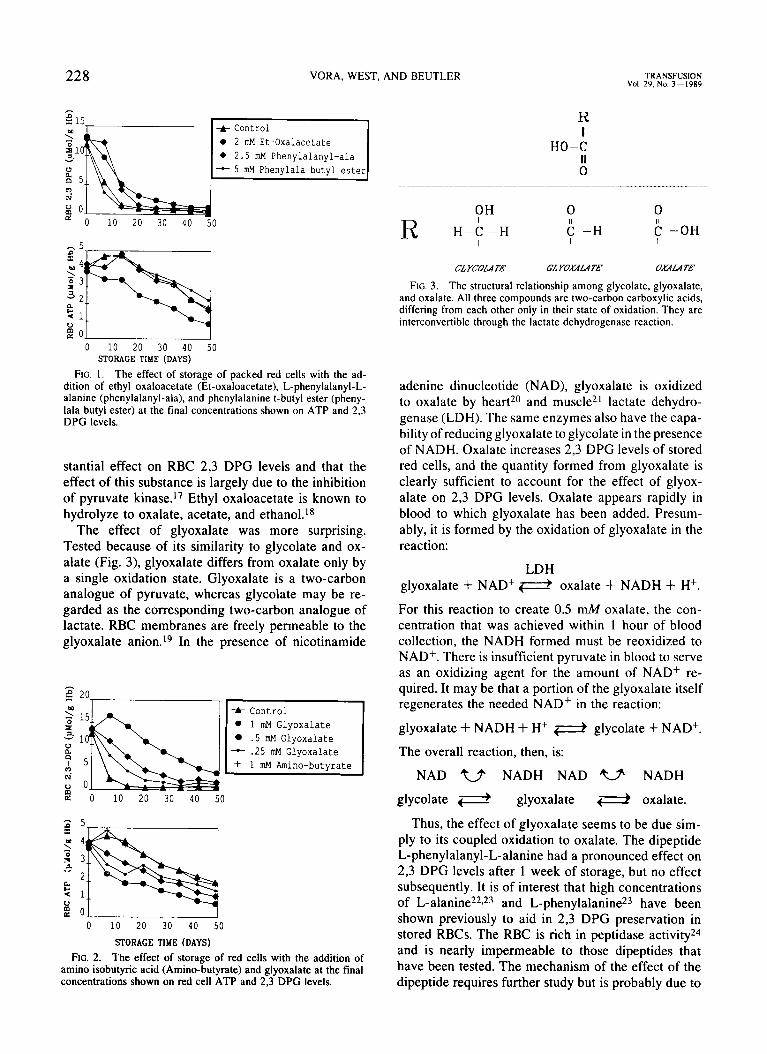
FIG. I . The effect of storage of packed red cells with the ad- dition of ethyl oxaloacetate (Et-oxaloacetate), L-phenylalanyl-L- alanine (phenylalanyl-ala), and phenylalanine t-butyl ester (pheny- lala butyl ester) at the final concentrations shown on ATP and 2,3 DPG levels.
stantial effect on RBC 2,3 DPG levels and that the effect of this substance is largely due to the inhibition of pyruvate kinase.” Ethyl oxaloacetate is known to hydrolyze to oxalate, acetate, and ethanol.I8
The effect of glyoxalate was more surprising. Tested because of its similarity to glycolate and ox- alate (Fig. 3), glyoxalate differs from oxalate only by a single oxidation state. Glyoxalate is a two-carbon analogue of pyruvate, whereas glycolate may be re- garded as the corresponding two-carbon analogue of lactate. RBC membranes are freely permeable to the glyoxalate anion.’g In the presence of nicotinamide
t Control 0 1 mM Glyoxalate
. 5 mM Glyoxalate - .25 mM Glyoxalate + 1 mM Amino-butyrate
0 1 0 20 30 40 50
0 10 20 30 40 50 STORAGE TIME (DAYS)
FIG. 2. The effect of storage of red cells with the addition of amino isobutyric acid (Amino-butyrate) and glyoxalate at the final concentrations shown on red cell ATP and 2,3 DPG levels.
R I
HO-C II 0
OH 0 I1 0 II I
I C -OH I
R H-C-H 7 -H
CL YCOUTK GL YOxRL4TE OxRL4TK
FIG. 3. The structural relationship among glycolate, glyoxalate, and oxalate. All three compounds are two-carbon carboxylic acids, differing from each other only in their state of oxidation. They are interconvertible through the lactate dehydrogenase reaction.
adenine dinucleotide (NAD), glyoxalate is oxidized to oxalate by heart20 and muscle2’ lactate dehydro- genase (LDH). The same enzymes also have the capa- bility of reducing glyoxalate to glycolate in the presence of NADH. Oxalate increases 2,3 DPG levels of stored red cells, and the quantity formed from glyoxalate is clearly sufficient to account for the effect of glyox- alate on 2,3 DPG levels. Oxalate appears rapidly in blood to which glyoxalate has been added. Presum- ably, it is formed by the oxidation of glyoxalate in the reaction:
LDH glyoxalate + NAD+ oxalate -t NADH + Hf.
For this reaction to create 0.5 mM oxalate, the con- centration that was achieved within 1 hour of blood collection, the NADH formed must be reoxidized to NAD+. There is insufficient pyruvate in blood to serve as an oxidizing agent for the amount of NAD+ re- quired. It may be that a portion of the glyoxalate itself regenerates the needed NAD’ in the reaction:
glyoxalate + NADH + Hf glycolate + NADf.
The overall reaction, then, is:
NAD NADH NAD NADH
glycolate 7 glyoxalate oxalate.
Thus, the effect of glyoxalate seems to be due sim- ply to its coupled oxidation to oxalate. The dipeptide L-phenylalanyl-L-alanine had a pronounced effect on 2,3 DPG levels after 1 week of storage, but no effect subsequently. It is of interest that high concentrations of L - a l a n i r ~ e ~ ~ , * ~ and L-~henyla lan ine~~ have been shown previously to aid in 2,3 DPG preservation in stored RBCs. The RBC is rich in peptidase activity*4 and is nearly impermeable to those dipeptides that have been tested. The mechanism of the effect of the dipeptide requires further study but is probably due to

229 TRANSFUSION EFFECT OF ADDITIVES TO CPDA ON 2.3 DPG 1989-Vol. 29. No. 3
its hydrolysis to L-alanine and L-phenylalanine to which the RBC is permeable.
Acknowledgments The authors acknowledge the technical assistance of Mark Brlej.
References I . Benesch R, Benesch RE. The effect of organic phosphates
from the human erythrocyte on the allosteric properties of hemoglobin. Biochem Biophys Res Commun 1967;26: 162-7.
2. Chanutin A, Curnish RR. Effect of organic and inorganic phos- phates on the oxygen equilibrium of human erythrocytes. Arch Biochem Biophys 1967;121:96-102,
3. Chanutin A. Effect of storage of blood in ACD-adenine- inorganic phosphate plus nucleosides on metabolic intermedi- ates of human red cells. Transfusion 1967;7:409-19.
4. Hamasaki N, ldeguchi H, Ikehara Y. Regeneration of 2.3- bisphosphoglycerate and ATP in stored erythrocytes by phos- phoenolpyruvate: a new preservative for blood storage. Trans- fusion l981;2 1:39 1-6.
5. Beutler E, Meul A, Wood LA. Depletion and regeneration of 2,3-diphosphoglyceric acid in stored red blood cells. Transfu- sion 1969;9: 109- 14.
6. Brake JM, Deindoerfer FH. Preservation of red blood cell 2,3-diphosphoglycerate in stored blood containing dihydrox- yacetone. Transfusion 1973; I3:84-8.
7. Beutler E, Guinto E. Dihydroxyacetone metabolism by human erythrocytes: demonstration of triokinase activity and its char- acterization. Blood 1973;41:559-68.
8. Hyde RM, Livingstone DJ, Patterson RA, Batchelor JF, King WR. Modification of the haemoglobin oxygen dissociation curve in whole blood by a compound with dual action. Lancet 1984;2:15-6.
9. Beutler E, Forman L, West C, Gelbart T. The mechanism of improved maintenance of 2,3-diphosphoglycerate in stored blood by the xanthone compound (2-(2-hydroxyethoxy)-6-( 1 - H-tetrazole-5-yl)xantheu-9-one (BW A440C). Biochem Phar- macol 1988;37:1057-60.
10. Kandler R, Grode G, Symbol R, Hickey G. Oxalate is the ac- tive component that produces increased 2,3-DPG in ascorbate stored red cells (abstract). Transfusion 1986;26:563.
1 I . Vora S. Red cell preservation: a novel proposal for the main- tenance of 2.3-DPC level. Blood 1986;68(Suppl 1):303a.
12. Beutler E. Red cell metabolism: a manual of biochemical meth- ods. 3rd ed. New York: Grune & Stratton, 1984.
13. Kohlbecker G, Butz M. Direct spectrophotometric determina-
tion of serum and urinary oxalate with oxalate oxidase. J Clin Chem Clin Biochem 1981;19:1103-6.
14. Wood L, Beutler E. Temperature dependence of sodium- potassium activated erythrocyte adenosine triphosphatase. J Lab Clin Med 1967;70287-94.
15. Beutler E, Kuhl W. Volume control of erythrocytes during storage: the role of mannitol. Transfusion 1988;28:353-7.
16. Wood LA, Beutler E. The effect of ascorbate on the mainte- nance of 2,3-diphosphoglycerate (2,3-DPC) in stored red cells. Br J Haematol 1973;25:611-8.
17. Beutler E, Forman L, West C. Effect of oxalate and malonate on red cell metabolism. Blood 1987;701389-93.
18. Windholz M, Budavari S, Blumetti RF, Otterbein ES, eds. The Merck index. Rahway, NJ: Merck & Co., 1983:554.
19. Hubbard AR, Sprandel U, Chalmers RA. Transport of oxalic acid, glycolic acid, glyoxylic acid and benzoic acid by resealed erythrocyte ‘ghosts’ prepared by a dialysis technique [proceed- ings]. Biochem SOC Trans 1979;7:958-60.
20. Warren WA. Catalysis of both oxidation and reduction of glyoxylate by pig heart lactate dehydrogenase isozyme I . J Biol Chem 1970245:1675-81,
21. Duncan RJS, Tipton KF. The oxidation and reduction of gly- oxylate by lactic dehydrogenase. Eur J Biochem 1969;l 1:58- 61.
22. Dawson RB, Ottinger WE, Chiu WM, Salib AA, Fagan DS, Keath JR. Control of red cell 2,3-DPG levels in vifro and a proposal for in vivo control in response to hypoxia and metabolic demand. In: Eaton JW, Kouzen DK, White JG, eds. Cellular and molecular aspects of aging: the red cell as a model. New York: Alan R. Liss, 1984:349-65.
23. Vora S. Metabolic manipulation of key glycolytic enzymes: a novel proposal for the maintenance of red cell 2.3- DPG and ATP levels during storage. Biomed Biochim Acta 1987;46:S285-9.
24. Pontremoli S, Sparatore B, Melloni E, et al. Differences and similarities among three acidic endopeptidases associated with human erythrocyte membranes. Molecular and functional stud- ies. Biochim Biophys Acta 1980;630313-22.
Shobhana Vora, MD, Assistant Member (deceased), Department of Basic and Clinical Research, Scripps Clinic and Research Foundation.
Carol West, Senior Research Assistant, Scripps Clinic and Research Foundation.
Ernest Beutler, MD, Member and Chairman, Department of Basic and Clinical Research (BCR-3). Scripps Clinic and Research Foundation, 10666 North Torrey Pines Road, La Jolla, CA 92037. [Reprint requests]





























