Embed Size (px)
Citation preview

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
Org. Divers. Evol. 6, Electr. Suppl. 8: 1 - 20 (2006)© Gesellschaft für Biologische SystematikURL: http://www.senckenberg.de/odes/06-08.htmURN: urn:nbn:de:0028-odes0608-9
The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in southern Luzon, PhilippinesSamuel W. James
Natural History Museum and Biodiversity Research Center, Dyche Hall, 1345 Jayhawk Drive, University of Kansas, Lawrence, KS 66045, USA
e-mail: [email protected]
Received 2 December 2004 • Accepted 11 August 2005
Abstract
An earthworm biodiversity survey of the Philippines has yielded 14 new species of the perichaetine megascolecid genus Pleionogaster, previously known from only a few species from scattered Philippine locations. Bicol, the southern peninsula of Luzon, has intact forests on several isolated volcanic peaks and other remote areas. Collections made in these forests yielded the following new species, here presented by type location: Mt. Malinao, Pleionogaster albayensis, P. bicolensis, P. castilloi, P. malinaoensis, P. tiwiensis; Mt. Isarog, P. ffitchae, P. isarogensis; Mt. Bulusan, P. bulusanensis, P. hongi, P. sorsogonensis; Catanduanes Island, P. nautsae, P. viracensis; Caramoan Peninsula, P. caramoanensis, P. nillosae. Most of the species were found only in the neighborhood of the type locality, but P. bicolensis occurs in two locations in northern Bicol. Intraspecific variation in P. castilloi was observed between northern and southern flanks of Mt. Malinao. The impor-tance of several previously overlooked Pleionogaster traits is demonstrated by their homogeneity within species reported here.
Keywords: Pleionogaster; Megascolecidae; Clitellata; Philippines; Luzon; Bicol
IntroductionThe perichaetine megascolecid genus Pleionogaster Michaelsen, 1892 has long been known from a few iso-lated collections made in the Philippines during the last two centuries (Easton 1979; James 2004), but its range and diversity remained unknown. In early 2001 the au-thor and a field team set out to conduct the first organi-zed collecting of earthworm material from the Philip-pines, and discovered many species of Pleionogaster in all parts of the island of Luzon, including Catandu-anes, an island united with Luzon at Pleistocene low sea levels (Heaney 1985; 1993). This paper is one of a series planned on new earthworm species discove-red by the Philippines Terrestrial Annelid and Gastro-pod Survey (PTAGS). Here I report the Pleionogaster species found in Bicol, a long southern peninsula of Luzon,plus Catanduanes Island, composed of lowland plains dotted with dormant and active volcanoes.
Pleionogaster is an interesting group of earthworms, from the standpoint of the systematics of the Phereti-ma complex (sensu Sims and Easton 1972) of the Me-gascolecidae. Several of its characteristics are unique among the genera of the complex, raising questions
about its placement in that group. Its members have a reduced esophageal gizzard in viii, all the anterior septa are present, and these are generally muscular an-terior of 9/10 or 10/11. There are paired meganephri-dia in the intestinal segments, but not elsewhere; these are connected to paired longitudinal ducts flanking the dorsal blood vessel. In addition there are regular ranks of micronephridia opening to corresponding rows of nephridiopores, again in the intestinal segments. Mi-cronephridia are present in the anterior segments but are placed differently. The intestine is supplied with several gizzards posterior to xxiv, followed by an aty-phlosolate section of intestine up to 25 segments long, at the end of which is a constriction and the typhlosolar origin, if a typhlosole is present. The supra-esophageal vessel extends posteriorly past the heart segments to xvii in most cases, and the vessels returning blood from the clitellar region (posterior latero-parietals) are from one to four pairs in segments xiv–xvii. In some cases the posteriormost pair of these vessels is connected to paired longitudinal vessels extending many segments back on the body wall. Most of these features have

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 2
been noted in older works on the genus, but some are reported here for the first time.
No other genera in the Pheretima complex share any of the unusual features of Pleionogaster, and neither do any other megascolecid genera found in the regi-on comprising the Philippine and Indonesian archi-pelagoes and mainland Southeast Asia west through Thailand. This poses a biogeographic and systematic puzzle, the solving of which will require additional ex-ploration. For now, the most likely location of related genera seems to be Australia, where Anisogaster Blake-more, 2000, Eastoniella Jamieson, 1977, Gastrodrilus Blakemore, 2000, Hickmaniella Jamieson, 1974, Pseu-docryptodrilus Jamieson, 1972, and Pseudonotoscolex Jamieson, 1971 share some characteristics with Pleio-nogaster (Jamieson 2000). The Indian genus Lampito Kinberg, 1866 has a similar arrangement of post-clitel-lar meganephridia and ducts (Gates 1972)
Ecologically, members of Pleionogaster are mostly endogeic, with generally long and slender, rarely pig-mented bodies, and always found in the soil, never in litter or arboreal habitats. Some of the larger species in which the anterior ends are darkly pigmented may be anecic.
Methods and materialOn the upper slopes of the Bicol volcanoes lie the re-maining primary montane and mossy forests of the area, and these were given priority in collecting. Collections were also carried out in two low-elevation sectors in some relatively undisturbed forest on Catanduanes Is-land and the nearby Caramoan Peninsula. Earthworms were collected by digging and handsorting soil, sear-ching organic soil and root mats from the tops of boul-ders, logs, roots and branches, dissecting epiphytes, primarily arboreal ferns, and searching the leaf axils of palms and Pandanaceae. All specimens examined for this paper were recovered from mineral soil, not from organic soil horizons, organic mats or other suspended soils, or plants.
Photographs were taken live of as many different species as could be distinguished in the field by in-spection with the unaided eye. Additional photomic-rographs were taken in the laboratory using a Nikon Coolpix 995 digital camera fitted to an adapter moun-ted on the phototube of a Leica M5 stereomicrosco-pe. Specimens were killed in 50% ethanol and fixed in 10% formaldehyde in the field. After at least 48 hours fixation they were rinsed in three changes of tap water with at least 12 hours between changes, and transferred to 80% ethanol. This removes residual formaldehyde, reduces health hazards to those working with the speci-mens, and may reduce DNA degradation. Duplicate sets of specimens were preserved in 95% ethanol for
molecular phylogenetic studies, and were archived at –20 ºC. All material was scored for external characters, drawings made by way of a drawing tube, and inter-nal anatomy studied by dorsal dissection. Setal counts were made over 1 mm each of a dorsal, lateral and ventral segmental equator arc, averaging these counts, and multiplying the average by the estimated circum-ference of the segment. This was necessary due to the extremely large numbers of setae (up to 300) crowded into the anterior setal rings.
Collecting site data include the acronym PTAGS (Philippine Terrestrial Annelid and Gastropod Survey) followed by a location number that uniquely identifies a location in the survey. Latitude and longitude are given in degrees and decimal minutes. Elevations were read from a GPS unit (Magellan Map410) if sufficient sa-tellites were detectable, or from an altimeter. The map datum used in the GPS readings was Luzon. Collectors listed are those working at the respective site.
Holotypes of the new species are deposited in the National Museum of the Philippines Annelid Collec-tion (NMA), P. Burgos St., Manila, Philippines. Ad-ditional depositories: KUNHM = Kansas University Natural History Museum, Lawrence, Kansas USA; UPLBMNH = University of the Philippines Los Baños Museum of Natural History, College, Laguna, Philip-pines.
Taxonomic sectionClitellata: Megascolecidae Rosa, 1891Genus Pleionogaster Michaelsen, 1892Perichaeta Beddard, 1886: 298 (in part).
Pleionogaster Michaelsen, 1892: 247; Beddard (1895: 433), Michaelsen (1896: 198), Easton (1979: 114), James (2004).
Plionogaster Michaelsen, 1900: 210; Stephenson (1930: 840), Stephenson (1933: 923), Gates (1943: 105), Jamieson (1971: 82).
Type species. Pleionogaster jagori Michaelsen, 1892 (Easton 1979).
Diagnosis. Perichaetine Megasolecidae with larger numbers of setae in the head segments than in post–cli-tellate segments, a reduced esophageal gizzard in viii, intestinal gizzards in the region xxiv–xxx, paired ente-roic stomate meganephridia and regular ranks of exoic micronephridia in post-clitellate segments, and a single pair of racemose prostates whose ducts are united with the vasa deferentia near the ental end of the duct. All known species have paired spermathecae in segments viii and ix.
Remarks. James (2004) discussed synonymy issues raised by Easton (1979) and made some changes to the genus definition in Easton (1979).

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 3
Pleionogaster tiwiensis n. sp.(Fig. 1A, B)
Etymology. Named after Tiwi, the municipality clos-est to the type locality.
Type material. Holotype (NMA004134): adult, PTAGS 055, Albay Province, near Tiwi, montane for-est on north ridge of Mt. Malinao, 13° 25.98’ N, 123° 37.63’ E, 850m asl, 10 May 2001, leg. S.W. James & A. Castillo. Paratypes (KUNHM 002175): 4 adults, col-lecting data as for holotype.
Description. Unpigmented, body 94–103 mm x 3.1–3.5 (vii), 2.7–3.7 mm (xv), 3.3–3.9 mm (xxv); 215–285 segments, some specimens with regenerated segments; body cylindrical in cross-section; segments x–xiii bian-nulate, postclitellar segments faintly biannulate in ante-rior half of body. First dorsal pore 11/12 or 12/13; sper-mathecal pores paired in 7/8, 8/9, 0.08 circumference apart, female pore single in xiv, male pores crescentic paired in xviii on raised porophores, 0.12 circumfer-ence apart, 5–6 setae between male pores. Setae regu-larly distributed around segmental equators; estimated at 210 setae on vii, 114 setae on xxv; in vii no dorsal gap, in xx ZZ:YZ = 2–3, ventral gap AA:AB = 2. Cli-tellum annular 1/2xiii, xiv–1/2xvii; genital markings midventral broad pad xvii, paired presetal in line with male pores on xviii, narrow elongate midventral over equator, presetal portion of xx, xxi, midventral epider-mal thickenings of xxii–xxiv (Figure 1A). Body wall slightly depressed between male pores. Nephridiopores visible as gaps in setal rings of postclitellar segments, apparently 6–10 pores per segment but not always in regular rows.
Septa 5/6–8/9 thick, muscular, 9/10 thinner, remain-der membranous. Weak gizzard in viii with typical iri-descent outer wall but flaccid, esophagus vascularized, with low villous discontinuous vertical folds xiii–xvii, valvular xviii, intestinal origin xix, no caeca; thick in-testinal gizzards xxvii–xxx (4), xxv–xxviii (1); typhlo-sole xlvii–lxx, simple fold, 0.1–0.15 lumen diameter. Intestine heavily vascularized xx–xxvii, less vasculari-zed xxxi–xlv, xlvi; internal texture changes from rugo-se to smooth xlv, xlvi.
Hearts x–xiii esophageal, commissural vessels v–ix lateral. Supra-esophageal vessel x–xvii, efferent parie-to-esophageal vessels lacking, but vessels connecting extraesophageal vessel to body wall present in each of xiv–xvii, extra- esophageal vessels from vi to xiii, in xiii fuse to single midventral vessel on ventral esopha-geal wall, entering esophageal wall xviii.
Nephridia present as peptonephridia on anterior faces of 4/5, 5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall the-reafter; from xix 6–10 micronephridia per segment, 2 meganephridia per segment from xix posteriorly, the-
se attached to tubules adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermatheca with club-shaped ampulla, ampulla not sharply demar-ked from duct; simple club-shaped diverticulum joins duct near body wall (Figure 1B). Male sexual system holandric, testes and funnels enclosed in annular sacs in x, xi, these sacs encompass hearts, seminal vesicles; seminal vesicles xi, xii small, acinous; vasa deferen-tia very small, travel up prostatic ducts, adherent to surface thereof until entering duct at most half-way to male pore; each prostate racemose two- to four-lobed, occupying xviii; with muscular duct; copulatory bursae lacking.
Remarks. The efferent latero-parietal vessels of most Megascolecidae are formed from the coalescence of small vessels on the body wall of clitellar segments to a single pair of vessels that join the extraesophageal vessels or the esophageal wall, usually in the region of segments xiii–xvi. In P. tiwiensis this arrangement is not found, but instead there are four such paired con-nections to the extra-esophageals from the body wall of clitellar segments xiv–xvii.
Pleionogaster tiwiensis is one of many species with four intestinal gizzards in xxvii–xxx (Table 1). Among those it has similar numbers of micronephridia in the intestinal segments to P. viracensis and P. isarogensis. However, P. viracensis is pigmented and has a more posterior intestinal constriction, P. isarogensis lacks paired genital markings, has a more posterior intesti-nal origin, a more anterior intestinal constriction, and a single pair of latero-parietal vessels in xv rather than four pairs.
Pleionogaster albayensis n. sp.(Fig. 1 C, D)
Etymology. Named after the province of Albay in which the species was found.
Type material. Holotype (NMA 004135): adult, PTAGS 055, Albay Province, near Tiwi, montane for-est on north ridge of Mt. Malinao, 13° 25.98’ N, 123° 37.63’ E, 850m asl, 10 May 2001, leg. S.W. James & A. Castillo.
Description. Slight purplish-brown anterior-dorsal tint, dusky strip along mid-dorsal line, otherwise un-pigmented, body 94 mm x 2.7–3.2 mm (vii), 2.6–2.9 mm (xv), 3.0–3.7 mm (xxv); 159 + > 60 regenerated segments, regenerates very small and asetal, thus hard to count; body cylindrical in cross-section; segments viii, ix biannulate, x–xiii triannulate, anterior half of postclitellar segments triannulate. First dorsal pore 12/13; spermathecal pores paired in 7/8, 8/9, 0.2 cir-cumference apart; female pore single in xiv, male pore

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 4
openings crescentic, paired in xviii on porophores tilt-ed posteriorly, 0.18 circumference apart, 12 setae be-tween male pores. Setae regularly distributed around segmental equators; estimated at 148 setae on vii, 85 setae on xxv; no dorsal or ventral gaps. Clitellum an-nular xiv–xvii; genital markings paired presetal xvii, paired 17/18, paired 18/19 in depression posterior to male pores, paired 19/20, paired presetal xix, xx, un-paired midventral presetal xix (Figure 1C). Nephrid-iopores visible as gaps in setal rings and longitudinal musculature of postclitellar segments, 12 pores per segment in regular rows.
Septa 5/6–8/9 thick, muscular, 9/10 thinner, remain-der membranous. Weak gizzard in viii with typical iridescent outer wall but flaccid, esophagus valvular xix, intestinal origin xx, no caeca; thick intestinal giz-zards xxvii–xxx; typhlosole xlvii–lxxiii, simple low fold. Intestine internal texture changes from rugose to smooth xlv, coelomic fluid rich in oil within intestinal segments.
Hearts x–xiii esophageal, commissural vessels v–ix lateral. Supra-esophageal vessel xi–xvii, efferent pari-eto-esophageal vessels lacking, but vessels connecting extraesophageal vessel to body wall present in each of xiv–xvii, extra- esophageal vessels from vi to xiii, in xiii fuse to single midventral vessel on ventral esopha-geal wall, entering esophageal wall xviii.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall the-reafter; from xix 12 micronephridia per segment, 2 meganephridia per segment from xix posteriorly, the-se attached to paired tubules adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermatheca with elongate club-shaped ampulla; simple club-shaped diverticulum joins duct near body wall on medial side rather than usual lateral side, slightly shorter than am-pulla; no differentiation of duct from ampulla within coelom. (Figure 1D). Male sexual system holandric, testes and funnels enclosed in annular sacs in x, xi, the-se sacs encompass hearts, seminal vesicles; sacs for-med of membranes connecting septa 9/10 and 10/11, 10/11 and 11/12; seminal vesicles xi, xii large, dense, acinous; vasa deferentia small, adherent to ental half of prostatic duct, then fuse with duct; few micronephri-dia surrounding duct-body wall contact; each prostate racemose four-lobed, occupying xviii; with muscular duct; copulatory bursae lacking.
Remarks. Other species with four intestinal gizzards in xxvii–xxx and with dark pigmentation are P. bico-lensis, P. ffitchae and P. viracensis. All of these have more setae per segment, a more posterior intestinal
constriction, a more anterior intestinal origin, and the first two lack midventral genital markings (Table 1).
Pleionogaster bicolensis n. sp.(Fig. 1E, F)
Etymology. Named after the province of Bicol in southern Luzon, because the species was collected in several locations in this region.
Type material. Holotype (NMA 004136): adult, PTAGS 056, Albay Province, Barangay Jarod, upper montane forest on south ridge of Mt. Malinao, 13° 23.96’ N, 123° 37.16’ E, 1030m asl, 11 May 2001, leg. S.W. James & A. Castillo. Paratypes: one adult (KUN-HM 002176), one adult (UPLBMHN Z-NS-0083); col-lecting data as for holotype.
Other material. KUNHM 002177: 1 adult, PTAGS 055., Albay Province, near Tiwi, montane forest on north ridge of Mt. Malinao, 13° 25.98’ N, 123° 37.63’ E, 850m asl, 10 May 2001, leg. S.W. James & A. Cas-tillo. UPLBMNH Z-NS-0084: 1 adult, PTAGS 121, Camarines Norte Province, Caramoan Peninsula, low-elevation forest on karst, 13° 44.40’ N, 123° 54.38’ E, 245m asl, 21 May 2001, leg. Y. Hong, M. Levi, P. Nil-los. KUNHM 002178: 2 adults, PTAGS 122, Cama-rines Norte Province, Caramoan Peninsula, low-eleva-tion forest on karst, 13° 45.29’ N, 123° 53.81’ E, 346m asl, 22 May 2001, leg. Y. Hong, M. Levi, P. Nillos.
Description. Deep brown anterior dorsal pigmen-tation, body 155–293 mm x 5.5–6 mm (vii), 5.6–6.6 mm (xv), 5.6–6.8 mm (xxv); 340–370 segments, most specimens amputees; body cylindrical in cross-section; segment xi biannulate, triannulate xii–xxx exclusive of clitellum. First dorsal pore 12/13; spermathecal pores paired in 7/8, 8/9, 0.17 circumference apart, asetal sec-tions ventrally near spermathecal pores viii, ix; female pore single in xiv, male pore openings paired in xviii on porophores tilted posteriorly into sunken male field zone, 0.17 circumference apart, 8–10 setae between male pores. Setae regularly distributed around seg-mental equators; estimated at 130–220 setae on vii, 132–140 setae on xxv; no ventral gaps, dorsal gaps ZZ:YZ 2:1. Clitellum annular xiv–1/2xvii; genital mark-ings paired presetal xvii, xix; paired 17/18 (Figure 1E). Nephridiopores visible as gaps in setal rings and longi-tudinal musculature of postclitellar segments, 12 pores per segment in regular rows.
Septa 5/6–9/10 thick, muscular, others membranous. Weak gizzard in viii with typical iridescent outer wall but flaccid; esophagus valvular xviii, intestinal origin xix or xx, no caeca; thick intestinal gizzards xxvii–xxx; typhlosole lacking. Intestine internal texture changes from rugose to smooth at intestinal constriction lvii.
Hearts xi–xiii esophageal, x lateral, commissural vessels v–ix lateral. Supra-esophageal vessel xi–xvii,

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 5
efferent parieto-esophageal vessels from body wall of xv–xviii, intestinal segments to extraesophageal ves-sels in xv, segmental parietal vessel from body wall of xiv to extraesophageal vessels xiv; extra- esophageal vessels ventral-lateral to gut vi–xii, ventral to esopha-gus xiii–xviii, fuse to one midventral vessel xviii, en-ters esophageal wall at esophageal valve.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–xi, on body wall the-reafter; from xviii 12 micronephridia per segment, these connected to tubular sinus with connections to nephridiopores; 2 meganephridia per segment from xix posteriorly, these attached to paired tubules adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermathe-ca with horn-shaped ampulla thinner-walled than stout muscular duct; simple club-shaped diverticulum joins duct near body wall, slightly shorter than spermathecal duct (Figure 1F). Male sexual system holandric, tes-tes and funnels enclosed in horseshoe-shaped sacs in x, xi, these sacs encompass hearts, seminal vesicles; sacs formed of membranes connecting septa 9/10 and 10/11, 10/11 and 11/12; seminal vesicles xi, xii small, acinous; vasa deferentia slender, non-muscular; each prostate racemose rounded mass occupying xviii; with thick muscular duct; copulatory bursae lacking.
Remarks. Among Pleionogaster with four intestinal gizzards in xxvii–xxx and dark pigmentation, P. bico-lensis is most similar to P. ffitchae (Table 1). The lack of a typhlosole, the more posterior intestinal constric-tion, and the different array of latero-parietal vessels distinguish it from P. ffitchae. Distinction from P. al-bayensis is discussed under that species.
Pleionogaster bulusanensis n. sp.(Fig. 1G, H)
Etymology. Named after the mountain on which the species was discovered.
Type material. Holotype (NMA 004137): preclitel-late adult, PTAGS 051, Sorsogon Province, Bulusan National Park, dipterocarp forest at Bulusan Lake, 12° 45.32’ N, 124° 05.34’ E, 360m asl, 3 May 2001, leg. S.W. James, A. Castillo, K. James, P. James. Paratype (KUNHM 002179): 1 juvenile; collecting data as for holotype.
Description. Unpigmented, body 105 mm X 4.6 mm (vii), 4.6 mm (xxv); 220 segments (anterior plus posterior fragments of same worm), 317 segments in complete juvenile; body cylindrical. First dorsal pore 11/12; spermathecal pores paired in 7/8, 8/9, 0.15 cir-cumference apart; female pore single, male pore open-ings crescentic, paired in xviii on small porophores,
0.17 circumference apart, 20 setae between male pores; clitellum not developed. Setae regularly distributed around segmental equators; estimated 200 setae on vii, 144 setae on x, 120 setae on xxv; no dorsal, ventral gaps. Genital markings paired presetal xvii, postsetal xvii, paired presetal xix–xxi, all just medial to line of male pores (Figure 1G). Nephridiopores visible as gaps in setal rings, longitudinal musculature of postclitellar segments, 14 pores per segment in regular rows.
Septa 5/6–8/9 thick, muscular, 9/10 thinner, muscu-lar, remainder membranous. Weak gizzard in viii with typical iridescent outer wall but flaccid, esophagus valvular xix, intestinal origin xx, no caeca; thick intes-tinal gizzards xxvii–xxx; typhlosole lacking; intestine constricted lv, thereafter coated with brown chlorago-gen layer.
Hearts x–xiii lateral-esophageal with very small con-nections to dorsal vessel, commissural vessels vii–ix lateral; supra-esophageal vessel x–xvii, extra-esopha-geal vessels not traceable past viii.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall the-reafter; from xix 14 micronephridia per segment, two meganephridia per segment from xix posteriorly, these attached to paired ureters adherent to either side of dor-sal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermathe-ca blunt lanceolate ampulla, poorly differentiated duct; narrow digitate diverticulum half or less length of main spermathecal axis joins duct near body wall (Figure 1H). Male sexual system holandric, testes and funnels enclosed in annular sacs in x, xi, these sacs encompass hearts, dorsal vessel, seminal vesicles; sacs formed of membranes connecting septa 9/10 and 10/11, 10/11 and 11/12; seminal vesicles xi, xii slender arcs; each pros-tate racemose four- to five-lobed occupying xviii; duct muscular; vas deferens slender, non-muscular, joins ental end of prostatic duct but visible within duct wall over ental 2/3; copulatory bursae lacking.
Remarks. The arc-shaped seminal vesicles are un-usual. This species has a very large number of micro-nephridia per segment, relative to its body size. Other small-bodied species reported here tend to have low numbers of micronephridia (4–6 per segment), whereas large numbers (10–12) have been found in large-bod-ied species such as P. bicolensis. This indicates that the number of micronephridia may be size-related in most but not all species. The genital markings are more nu-merous and placed farther back in P. bulusanensis than in most other species encountered so far. Of those un-pigmented Pleionogaster with four intestinal gizzards in xxvii–xxx, it is the only species without a typhlosole

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 6
and has the posteriormost intestinal constriction (Table 1).Pleionogaster hongi n. sp.(Fig. 1I, J)
Etymology. Named after Dr. Hong Yong, a Korean earthworm specialist who is part of the PTAGS re-search team.
Type material. Holotype (NMA 004138): adult, PTAGS 051, Sorsogon Province, Bulusan National Park, dipterocarp forest at Bulusan Lake, 12° 45.32’ N, 124° 05.34’ E, 360m asl, 3 May 2001, leg. S.W. James, A. Castillo, K. James, P. James. Paratype (KUNHM 002180): adult; collecting data as for holotype.
Description. Faint pink anterior dorsal pigment, body 84, 87 mm x 3–3.5 mm (vii), 2.9–3.2 mm (xv), 3–3.4 mm (xxv); 190, 189 + 49 regenerated segments; body cylindrical. First dorsal pore 11/12; spermathecal pores paired in 7/8, 8/9, 0.18–0.19 circumference apart; female pore single, male pore openings crescentic, paired in xviii on small porophores facing posteriorly in sunken male field, 0.18–0.19 circumference apart, 10–12 setae between male pores; clitellum xiv–1/2xvii, annular. Setae regularly distributed around segmental equators; estimated 176 setae on vii, 104 setae on x, 80 setae on xxv; no dorsal, ventral gaps. Genital mark-ings paired presetal xvii, 17/18, xix–xxi (Figure 1I). Nephridiopores visible as gaps in setal rings, longitudi-nal musculature of postclitellar segments, 12 pores per segment in regular rows.
Septa 5/6–8/9 thick, muscular, 9/10, 10/11 thinner, muscular, remainder membranous. Weak gizzard in viii with typical iridescent outer wall but flaccid, eso-phagus valvular xix, intestinal origin xx, no caeca; in-testine vascularized xx–xxvi, thick intestinal gizzards xxvii–xxx; typhlosole simple fold ¼ lumen diameter xlvi–lxviii; intestine constricted 44/45, thereafter coa-ted with brown chloragogen layer.
Hearts x–xiii esophageal, commissural vessels vi–ix lateral; supra-esophageal vessel x–xvii, extra-esopha-geal vessels not traceable past viii, vessels from ventral esophagus xiv to body wall xv–xvii, paired longitudi-nal vessels on body wall xviii–xxiv.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall the-reafter; from xix 12 micronephridia per segment, two stomate meganephridia per segment from xix posteri-orly, these attached to paired ureters adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each sperma-theca cylindrical ampulla, poorly differentiated duct; club-shaped diverticulum half or less length of main spermathecal axis joins duct near body wall (Figure
1J). Male sexual system holandric, testes and funnels enclosed in annular sacs in x, xi, these sacs encompass hearts, dorsal vessel, seminal vesicles; sacs formed of membranes connecting septa 9/10 and 10/11, 10/11 and 11/12; seminal vesicles xi, xii slender arcs; each prosta-te racemose five-lobed occupying xviii; ectal 1/3 duct muscular; vas deferens slender, non-muscular, joins ental end of prostatic duct but visible within duct wall over ental 2/3; copulatory bursae lacking.
Remarks. The closest morphologically is P. bulu-sanensis, but P. hongi has pink pigmentation (versus brown in the other pigmented Bicol species), half as many setae between the male pores, an intestinal tran-sition anterior to that of P. bulusanensis, and a typhlo-sole (Table 1).
Pleionogaster ffitchae n. sp.(Fig. 2A, B)
Etymology. Named after Jana ffitch, who as an un-dergraduate student participated in the fieldwork and initial cataloguing of the 2001 expedition collections.
Type material. Holotype (NMA 004139): adult, PTAGS 060, Camarines Sur Province, montane forest on Mt. Isarog, 13° 39.90’ N, 123° 21.89’ E, 1100m asl, 14 May 2001, leg. Y. Hong, M. Levi, J. ffitch, P. Nillos, R. Abiada. Paratype (KUNHM 002181): adult; collec-ting data as for holotype.
Other material. UPLBMNH Z-NS-0085: adult, PTAGS 066, Camarines Sur Province, lower montane forest soils on Mt. Isarog, 13° 40.02’ N, 123° 21.16’ E, 920m asl, 17 May 2001, leg. S.W. James, Y. Hong, M. Levi, J. ffitch, P. Nillos, D. Franke, R. Abiada, R. Lazaro. KUNHM 002182: adult, PTAGS 069, Catan-duanes Province, low-elevation forest north of Baran-gay Summit, Buradan, 13° 46.0’ N, 124°16.0’ E, 275m asl, 22 May 2001, leg. S.W. James, P. James, J. James, K. James, J. ffitch, A. Castillo.
Description. Medium milky brown dorsal pigment, fading gradually towards tail, body 194, 170, 142 (am-putee) mm x 4.5–6 mm. (vii), 4.7–4.9 mm (xv), 4.6–5.8 mm (xxv); 285, 352, 210 (amputee) segments; body cylindrical. First dorsal pore 11/12, 12/13; spermathe-cal pores paired in 7/8, 8/9, 0.18–0.19 circumference apart bordered by thick lips on trailing edges of vii, viii, smaller lips on leading edges of viii, ix; female pore single, male pore openings crescentic, paired in xviii on small porophores facing posteriorly in sunken male field, 0.16–0.17 circumference apart, 8 setae between male pores; clitellum xiv–1/2xvii, annular. Setae regu-larly distributed around segmental equators; estimated 212–226 setae on vii, 150–170 setae on x, 100–110 se-tae on xxv; dorsal gaps variable ZZ = YZ, up to 2.5 YZ, ventral gaps AA:AB 1.5:1. Genital markings oval, paired, presetal xvii, rectangular paired 17/18, oblong,

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 7
paired presetal xix–xxi (Figure 2A). Nephridiopores visible as gaps in setal rings, longitudinal musculature of postclitellar segments, 12 pores per segment in regu-lar rows.
Septa 5/6–9/10 thick, muscular, 10/11–16/17 tough but transparent, remainder membranous. Weak giz-zard in viii with typical iridescent outer wall but flac-cid, esophagus valvular xviii, intestinal origin xix, no caeca; intestine vascularized xix–xxvi, thick intestinal gizzards xxvii–xxx; typhlosole simple fold 1/6 lumen diameter 50/51 (1), liv (1)–lxxvii; intestine constricted xlix (1), 52/53 (1), thereafter coated with brown chlo-ragogen layer.
Hearts x–xiii esophageal, commissural vessels vi–ix lateral; supra-esophageal vessel x–xvii, extra-esopha-geal vessels traced from iii–ix, then on ventral eso-phageal surface x–xvi, segmental paired vessels from ventral esophagus xiii to body wall xiv–xvi, those of xvi connect to paired longitudinal vessels on body wall xviii–cv.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall the-reafter; from xix 12 micronephridia per segment, two stomate meganephridia per segment from xix posteri-orly, these attached to paired ureters adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermathe-ca long lanceolate ampulla, duct of similar structure; short basally-attached diverticulum (Figure 2B). Male sexual system holandric, testes and funnels enclosed in annular sacs in x, xi, these sacs encompass hearts, dorsal vessel, seminal vesicles; some individuals with U-shaped dorsally open sac in x; sacs formed of mem-branes connecting septa 9/10 and 10/11, 10/11 and 11/12; seminal vesicles xi, xii, thick ventral half, up-per portion slender arc; prostates racemose five-lobed occupying xviii; duct muscular; vas deferens slender, non-muscular; copulatory bursae lacking. Genital mar-king glands sessile, dense xvii–xxi corresponding in placement to external markings.
Remarks. Pleionogaster ffitchae is most similar to P. bicolensis; differences are discussed under the latter species.
Pleionogaster viracensis n. sp.(Fig. 2C, D)
Etymology. Named after Virac, the capitol city of Catanduanes Province and the large town nearest to the type locality.
Type material. Holotype (NMA 004140): adult, PTAGS 068, Catanduanes Province, low-elevation forest near Barangay Summit, Buradan, 13° 43.60’
N, 124° 17.14’ E, 210m asl, 21 May 2001, leg. S.W. James, P. James, J. James, K. James, J. ffitch, A. Castil-lo. Paratype (KUNHM 002183): adult; collecting data as for holotype.
Other material. UPLBMNH Z-NS-0086: adult, PTAGS 069, Catanduanes Province, low-elevation for-est north of Barangay Summit, Buradan, 13° 46’ N, 124°16’ E, 275m asl, 22 May 2001, leg. S.W. James, P. James, J. James, K. James, J. ffitch, A. Castillo. UPLBMNH Z-NS-0087: adult, PTAGS 071, Catan-duanes Province, lower riparian forest near Barangay San Miguel, Pangabinan, 13° 54.50 N, 124° 11 E, 212m asl, 23 May 2001, leg. S.W. James, J. ffitch, A. Castillo.
Description. Medium brown anterior dorsal pig-ment, diminishing to mid-dorsal stripe in post-clitellar segments, body 107–140 mm x 4–4.6 mm (vii), 3.1–3.8 mm (xv), 4.2–4.5 mm (xxv); 260–279 segments; body cylindrical. First dorsal pore 11/12; spermathecal pores paired in 7/8, 8/9, 0.13 circumference apart surrounded by thickened area about 2 mm wide from 1/2vii–1/2ix; female pore single, male pore openings crescentic, paired in xviii on small porophores facing posteriorly in sunken male field, 0.16–0.17 circumference apart, 10 setae between male pores; clitellum xiv–1/2xvii, annular. Setae regularly distributed around segmental equators; estimated 182–226 setae on vii, 150–160 se-tae on x, 138–146 setae on xxv; dorsal gaps variable ZZ = YZ, up to 1.5 YZ, no ventral gaps. Genital markings oval, paired, presetal xvii, rectangular paired 17/18, oblong, paired presetal xix–xxi, midventral at equator xviii (Figure 2C). Nephridiopores visible as gaps in se-tal rings, longitudinal musculature of postclitellar seg-ments, 10 pores per segment in regular rows.
Septa 5/6–7/8 thick, muscular, 8/9 thinly muscled, 9/10–16/17 membranous. Weak gizzard in viii with typical iridescent outer wall but flaccid, esophagus valvular xviii, intestinal origin xix, no caeca; intestine vascularized xix–xxvi, thick intestinal gizzards xxvii–xxx; typhlosole very low fold lii–lxcii, full size by lvi; intestine constricted lii (1), lvi (2), thereafter coated with brown chloragogen layer.
Hearts x–xiii esophageal, commissural vessels vi–ix lateral; supra-esophageal vessel x–xvii, extra-esopha-geal vessels traced from iii–ix, then on ventral eso-phageal surface x–xvii, segmental paired vessels from body wall to extra-esophageal vessels xiv; posterior la-tero-parietals to extra-esophageals xv, these connect to paired longitudinal vessels on body wall xviii–cv.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall the-reafter; from xix 10 micronephridia per segment, two stomate meganephridia per segment from xix posteri-

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 8
orly, these attached to paired ureters adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermathe-ca broad club-shaped ampulla with very short slightly differentiated muscular duct; short basally-attached diverticulum (Figure 2D). Male sexual system holan-dric, testes and funnels enclosed in annular sacs in x, xi, these sacs encompass hearts, seminal vesicles; slen-der arcuate seminal vesicles xi, xii; prostates racemose three- to four-lobed occupying xviii; duct muscular; vas deferens slender, non-muscular; copulatory bursae lacking.
Remarks. Among the brown-pigmented Pleionogas-ter with four intestinal gizzards in xxvii–xxx (Table 1), P. viracensis is most similar to P. albayensis and P. ffitchae. Pleionogaster albayensis has a lower number of setae and more anterior locations of the intestinal origin and constriction; P. ffitchae lacks mid-ventral genital markings, has more micronephridia per intes-tinal segment, and three pairs of latero-parietal vessels rather than two.
Pleionogaster isarogensis n. sp.(Fig. 2E, F)
Etymology. Named for Mt. Isarog, on which the spe-cies was found over a wide elevational range.
Type material. Holotype (NMA 004141): adult, PTAGS 065, Camarines Sur Province, upper montane forest soils on Mt. Isarog, 13° 39.75’ N, 123° 21.98’ E, 1500m asl, 16 May 2001, leg. S.W. James, Y. Hong, M. Levi, J. ffitch, P. Nillos, R. Abiada, R. Lazaro. Paratypes: 3 adults (KUNHM 002184), 4 adults (UPLBMNH Z-NS-0088); collecting data as for holotype.
Other material. UPLBMNH Z-NS-0089: 2 adults, PTAGS 059, Camarines Sur Province, soils of mon-tane forest on Mt. Isarog, 13° 39.79’ N, 123° 21.79’ E, 1330m asl, 13 May 2001, leg. S.W. James, Y. Hong, M. Levi, J. ffitch, D. Franke, P. Nillos, R. Abiada, R. Lazaro. KUNHM 002186: 2 adults, PTAGS 060, Ca-marines Sur Province, montane forest on Mt. Isarog, 13° 39.90’ N, 123° 21.89’ E, 1100m asl, 14 May 2001, leg. Y. Hong, M. Levi, J. ffitch, P. Nillos, R. Abiada. UPLBMNH Z-NS-0090: 1 adult, PTAGS 061, Cama-rines Sur Province, mossy forest soils at summit of Mt. Isarog, 13° 39.65’ N, 123° 22.28’ E, 1987m asl, 15 May 2001, leg. S.W. James, Y. Hong, M. Levi, J. ffitch, P. Nillos, R. Abiada, R. Lazaro. KUNHM 002187: 1 juvenile, PTAGS 062, Camarines Sur Province, in or-ganic mats in mossy forest at summit of Mt. Isarog, 13° 39.65’ N, 123° 22.28’ E, 1987m asl, 15 May 2001, leg. S.W. James, Y. Hong, M. Levi, J. ffitch, P. Nillos, R. Abiada, R. Lazaro. UPLBMNH Z-NS-0091: 2 adults, PTAGS 064, Camarines Sur Province, upper montane
forest soils on Mt. Isarog, 13° 39.70’ N, 123° 22.07’ E, 1745m asl, 16 May 2001, leg. S.W. James, Y. Hong, M. Levi, J. ffitch, D. Franke, P. Nillos, R. Abiada, R. Lazaro. KUNHM 002188: 2 adults, PTAGS 066, Ca-marines Sur Province, lower montane forest soils on Mt. Isarog, 13° 40.02’ N, 123° 21.16’ E, 920m asl, 17 May 2001, leg. S.W. James, Y. Hong, M. Levi, J. ffitch, P. Nillos, D. Franke, R. Abiada, R. Lazaro.
Description. Unpigmented, body 75–165 mm x 2.7–3.2 mm (vii), 2.2–3.0 mm (xv), 2.8–3.3 mm (xxv); 275–294 segments; body cylindrical. First dorsal pore 12/13; spermathecal pores paired in 7/8, 8/9, 0.09–0.11 circumference apart deep in furrows; female pore sin-gle, male pore openings crescentic, paired in xviii on small porophores, 0.11–0.12 circumference apart, 4–8 setae between male pores; clitellum xiv–1/2xvii, annu-lar. Setae regularly distributed around segmental equa-tors; estimated 170–186 setae on vii, 110–120 setae on x, 72–84 setae on xxv; asetal zones midventral in one or more of vii–ix. Genital markings oval, midventral, presetal xvii, xix, xx (all), some with additional mid-ventral GM xviii, epidermal thickenings ventral xxi, xxii; (Figure 2E). Nephridiopores visible as gaps in se-tal rings, longitudinal musculature of postclitellar seg-ments, 6 pores per segment in regular rows.
Septa 5/6–8/9 thick, muscular, 9/10 thinner, remain-der membranous. Weak gizzard in viii with typical iri-descent outer wall, esophagus valvular xviii, intestinal origin xx, no caeca; intestine vascularized xx–xxvi, thick intestinal gizzards xxvii–xxx with small vascu-lar network on anterior thin-walled half of gut in each giceriate segment; typhlosole simple fold 1/3 lumen diameter for first ten segments, then only 1/6 lumen diameter from xli, xliii–lxxviii; intestine constricted 40/41 or 41/42.
Hearts x–xiii esophageal, commissural vessels vii–ix lateral; supra-esophageal vessel x–xvii, extra-esophageal with branch from body wall of xiv, effe-rent posterior latero-parietal vessels travel up 15/16 to extra-esophageal vessel in xvi, these connected to paired longitudinal vessels on body wall xviii–lviii; not always visible past xxx.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall there-after; from xix 6–8 micronephridia per segment, two stomate meganephridia per segment from xix posteri-orly, these attached to paired ureters adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermathe-ca ovate ampulla; short basally-attached diverticulum (Figure 2F). Male sexual system holandric, testes and funnels enclosed in annular sac in x, U-shaped sac

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 9
open dorsally in xi, these sacs encompass hearts, dor-sal vessel, seminal vesicles; sacs formed of membranes connecting septa 9/10,10/11; 10/11, 11/12; seminal vesicles xi, xii, loosely acinous; prostates racemose three- to four-lobed occupying xviii; ectal 1/3 duct muscular; vas deferens slender, non-muscular; copula-tory bursae lacking.
Remarks. Compared to other unpigmented Pleiono-gaster with four intestinal gizzards in xxvii–xxx, P. isarogensis has fewer setae per segment than P. bu-lusanensis and P. tiwiensis, more narrowly spaced male and spermathecal pores than P. bulusanensis, and consistently fewer micronephridia in the intesti-nal segments as well as a more anterior location of the intestinal constriction than both those species (Table 1). While P. tiwiensis has both midventral and paired genital markings, and P. bulusanensis has only paired markings, P. isarogensis has only midventral genital markings.
Pleionogaster malinaoensis n. sp.(Fig. 2G, H)
Etymology. Named after the mountain on which the material was collected.
Type material. Holotype (NMA 004142): adult, PTAGS 055, Albay Province, near Tiwi, montane for-est on north ridge of Mt. Malinao, 13° 25.98’ N, 123° 37.63’ E, 850m asl, 10 May 2001, leg. S.W. James & A. Castillo. Paratype (KUNHM 002189): adult; collecting data as for holotype.
Other material. KUNHM 002190: 2 adults, PTAGS 056, Albay Province, Barangay Jarod, upper mon-tane forest on south ridge of Mt. Malinao, 13° 23.96’ N, 123° 37.16’ E, 1030m asl, 11 May 2001, leg. S.W. James and A. Castillo.
Description. Unpigmented, body 97–110 mm x 2.8–3.2 mm (vii), 2.6–3.4 (xv), 3.5–4 mm (xxv); 227 segments, most specimens amputees; body cylindrical in cross-section; segments x–xiii triannulate, postclitel-lar segments faintly triannulate in anterior half. First dorsal pore 12/13; spermathecal pores paired in 7/8, 8/9, 0.10 circumference apart, female pores single in xiv, male pores crescentic paired in xviii on raised po-rophores, 0.09 circumference apart, 2–4 setae between male pores. Setae regularly distributed around segmen-tal equators; estimated at 150–160 setae on vii, setal density drops in ix, 100 setae on xxv; in vii no dorsal or ventral gaps, in xxv ZZ:YZ = 2, no ventral gap. Cli-tellum annular 1/2xiii, xiv–1/2xvii; genital markings broad midventral ix (1), x slightly wider than sperma-thecal pore spacing; midventral round white area xv (1) triangular pad xvi, paired presetal in line with male pores xix–xxii, or in less well developed specimens midventral epidermal thickenings on xxii–xxiii (Fig-
ure 2G). Nephridiopores visible as gaps in setal rings and longitudinal musculature of postclitellar segments, apparently 10–12 pores per segment but not always in regular rows.
Septa 5/6–8/9 thick, muscular, 9/10 thinner, remain-der membranous. Weak gizzard in viii with typical iridescent outer wall but flaccid, esophagus valvular xviii, intestinal origin xix or xx (1), no caeca; thick in-testinal gizzards xxv–xxix; typhlosole xlv, xlvii–lxiii, lxvii, lxxv simple fold, 0.1 lumen diameter. Intestine internal texture changes from rugose to smooth xliii, 43/44, xlvi.
Hearts x–xiii esophageal, commissural vessels v–ix lateral. Supra-esophageal vessel x–xvi, efferent parie-to-esophageal vessels lacking, but vessels connecting extraesophageal vessel to body wall present in each of xiv–xvii, extra- esophageal vessels from vi to xiii, in xiii fuse to single midventral vessel on ventral esopha-geal wall, entering esophageal wall xviii.
Nephridia present as peptonephridia on anterior face septum 5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall the-reafter; from xix 10–12 micronephridia per segment, 2 meganephridia per segment from xix posteriorly, the-se attached to tubules adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermatheca with ovate ampulla differentiated from narrower duct; simple club-shaped diverticulum joins duct near body wall, slightly longer than spermathecal duct (Figure 2H). Male sexual system holandric, testes and funnels enclosed in annular sacs in x, xi, these sacs encompass hearts, seminal vesicles; sacs formed of membranes connecting septa 9/10 and 10/11, 10/11 and 11/12; se-minal vesicles xi, xii small, acinous; vasa deferentia slender, non-muscular; each prostate racemose multi-lobed, occupying xviii; with slender muscular duct; copulatory bursae lacking.
Remarks. Other species with five gizzards in xxv–xxix are P. sorsogonensis and potentially P. castil-loi from the same mountain (Table 2). Pleionogaster malinaoensis has more intestinal micronephridia and more narrowly spaced male and spermathecal pores than those two species. Pleionogaster castilloi (see below) lacks paired genital markings; P. sorsogonen-sis has preclitellar genital markings. The former has a more posterior intestinal origin, the latter a more an-terior typhlosolar origin and spermathecal ampullae undifferentiated from the ducts. One individual from PTAGS055 had large numbers of small yellow nema-todes under the peritoneum on the inner body wall sur-face from xii and posteriorly.

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 10
Pleionogaster castilloi n. sp.(Fig. 2I, J)
Etymology. Named after Augusto Castillo, one of the PTAGS logistics coordinators, who discovered un-known reserves of strength climbing the crater wall of Mt. Malinao.
Type material. Holotype (NMA 004143): adult, PTAGS 055, Albay Province, near Tiwi, montane for-est on north ridge of Mt. Malinao, 13° 25.98’ N, 123° 37.63’ E, 850m asl, 10 May 2001, leg. S.W. James & A. Castillo. Paratype (KUNHM 002191): adult; collecting data as for holotype.
Other material. KUNHM 002192: 3 adults, PTAGS 056, Albay Province, Barangay Jarod, upper mon-tane forest on south ridge of Mt. Malinao, 13° 23.96’ N, 123° 37.16’ E, 1030m asl, 11 May 2001, leg. S.W. James and A. Castillo.
Description. Unpigmented, body 93–113 mm x 2.7–3.2 mm (vii), 2.6–2.9 mm (xv), 3.0–3.7 mm (xxv); 220–234 segments, most specimens amputees; body cylindrical in cross-section; segments x–xiii biannulate, anterior half of postclitellar segments triannulate. First dorsal pore 11/12, 12/13; spermathecal pores paired in 7/8, 8/9, 0.17 circumference apart, surrounded by thickened lips; female pores paired in xiv, male pore openings crescentic, paired in xviii on porophores tilt-ed posteriorly, slightly depressed, 0.17 circumference apart, 6–8 setae between male pores. Setae regularly distributed around segmental equators; estimated at 160–166 setae on vii, 70–80 setae on xxv; no dorsal or ventral gaps. Clitellum annular 1/3xiii–1/2xvii; genital markings broad midventral 17/18, 18/19, 19/20; long shallow indentations in raised areas, somewhat like the fusion of paired indented genital markings; mid-ventral epidermal thickenings on xxi–xxii (Figure 2I). Nephridiopores visible as gaps in setal rings and longi-tudinal musculature of postclitellar segments, 6 pores per segment in regular rows.
Septa 5/6–8/9 thick, muscular, 9/10 thinner, remain-der membranous. Weak gizzard in viii with typical iri-descent outer wall but flaccid, esophagus valvular xix, intestinal origin xx, no caeca; thick intestinal gizzards xxv–xxix, xxv–xxxi, xxvi–xxx, xxvi–xxx, xxvi–xxxi; typhlosole xlvii, xlviii–lxviii, lxx, lxvii, lxxv simple fold, 0.16 lumen diameter. Intestine internal texture changes from rugose to smooth xlvii, 45/46 (3), 47/48 (2); intestinal wall vascularized xxxii–xlvii.
Hearts x–xiii esophageal, commissural vessels v–ix lateral. Supra-esophageal vessel x–xviii, efferent pari-eto-esophageal vessels lacking, but vessels connecting extraesophageal vessel to body wall present in each of xiv–xvii, extra- esophageal vessels from vi to ix where they enter dorsal esophageal wall, in xiii fuse to single
midventral vessel on ventral esophageal wall, entering esophageal wall xviii.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall thereafter; from xix 6 micronephridia per segment, 2 meganephri-dia per segment from xix posteriorly, these attached to tubules adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermatheca with sagittate ampulla thinner-walled than duct; simp-le club-shaped diverticulum joins duct near body wall, slightly shorter than spermathecal duct (Figure 2J). Male sexual system holandric, testes and funnels en-closed in annular sacs in x, xi, these sacs encompass hearts, seminal vesicles; sacs formed of membranes connecting septa 9/10 and 10/11, 10/11 and 11/12; se-minal vesicles xi, xii small, acinous; vasa deferentia slender, non-muscular, adherent to ental 2/3 or more of prostatic duct, then fuse with duct; patch of microneph-ridia surrounding duct-body wall contact; each prostate racemose rounded mass, occupying xviii; with muscu-lar duct entering small muscular bulge at body wall; copulatory bursae lacking.
Remarks. This species is characterized by six micro-nephridia per segment, at the low end of the range, and relatively many intestinal gizzards, the number ranging from 5 to 7. The PTAGS 055 specimens from the north ridge of Mt. Malinao all had 5 gizzards and the intesti-nal transition at 45/46, rather than 6 or 7 gizzards and the intestinal transition at 47/48 as seen in the south ridge material (PTAGS 056). I have chosen to include these in one species based on the common number of micronephridia and the similarity in male field genital marking pattern. Distinction between P. castilloi and P. malinaoensis has been discussed under the latter spe-cies. The next most similar species is P. sorsogonensis, from which P. castilloi can be distinguished by the lack of preclitellar and paired postclitellar genital markings, the more posterior intestinal constriction, and a greater number of setae (Table 2).
Pleionogaster nillosae n. sp.(Fig. 3A, B)
Etymology. Named after Portia Nillos, one of the PTAGS logistics coordinators.
Type material. Holotype (NMA 004144): preclitel-late adult, PTAGS 122, Camarines Norte Province, Caramoan Peninsula, low-elevation forest on karst, 13° 45.29’ N, 123° 53.81’ E, 346m asl, 22 May 2001, leg. Y. Hong, M. Levi, P. Nillos. Paratype (KUNHM 002193): juvenile; collecting data as for holotype.
Description. Unpigmented, body 250 mm (extend-ed) x 5 mm (vii), 4.5 mm (xxv); 330 segments; body

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 11
cylindrical in cross-section; postclitellar segments tri-annulate. First dorsal pore 12/13; spermathecal pores paired in 7/8, 8/9, 0.2 circumference apart; female pore not seen, male pore openings crescentic, paired in xviii on small flat porophores, 0.18 circumference apart, no setae between male pores. Setae regularly distributed around segmental equators; estimated at 314 setae on vii, 140 setae on xxv; no dorsal or ventral gaps. Genital markings faint, paired presetal xvii, xix; slight thicken-ing of epidermis over ventral surface of vii–ix (Figure 3A). Nephridiopores visible as gaps in setal rings of postclitellar segments, 10 pores per segment in regular rows.
Septa 5/6–9/10 thick, muscular, 10/11, 11/12 thinner, remainder membranous. Weak gizzard in viii with typi-cal iridescent outer wall but flaccid, esophagus valvular xix, intestinal origin xx, no caeca; six thick intestinal gizzards xxiv–xxix; typhlosole lacking. Intestinal tran-sition obscured by poor preservation, in l–lv region.
Hearts xi–xiii esophageal, commissural vessels v–x lateral. Supra-esophageal vessel xi–xvii, efferent parie-to-esophageal vessels not seen due to damage; segmen-tal parietal vessel from body wall of xiv to extraeso-phageal vessels xiv; extra-esophageal vessels not seen anteriorly, fused mid-ventral below esophagus xiii–xx.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall the-reafter; from xix 8–10 micronephridia per segment, 2 meganephridia per segment from xix posteriorly, these attached to paired ureters adherent to either side of dor-sal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermathe-ca with elongate horn-shaped ampulla, muscular duct; simple club-shaped diverticulum joins duct near body wall, much shorter than ampulla (Figure 3B). Male se-xual system holandric, testes and funnels enclosed in annular sacs in x, xi, these sacs encompass hearts, se-minal vesicles; sacs formed of membranes connecting septa 9/10 and 10/11, 10/11 and 11/12; seminal vesicles xi, xii slender arches; each prostate racemose hemis-pherical mass occupying xviii; with muscular S-curved duct; copulatory bursae lacking, slight glandular deve-lopment posterior to prostatic duct unions with body wall, suggestive of genital marking glands.
Remarks. Better characterization of this large, un-pigmented species was prevented by the poor state of preservation and by immaturity of the limited material obtained. For the time being it can be distinguished by its large size, comparable to P. bicolensis but lacking pigmentation, having fewer micronephridia per seg-ment, more setae per segment but none between the male pores, more gizzards (6) more anteriorly placed,
more anterior intestinal constriction, and different tes-tes sacs (Table 2). The faint genital markings may be due to immaturity.
Pleionogaster caramoanensis n. sp.(Fig. 3C, D)
Etymology. Named for the Caramoan Peninsula on the eastern side of Bicol.
Type material. Holotype (NMA 004145): adult, PTAGS 123, Camarines Norte Province, Caramoan Peninsula, low-elevation forest on karst, 13° 41.77’ N, 123° 54.02’ E, 350m asl, 23 May 2001, leg. Y. Hong, M. Levi, P. Nillos. Paratype (KUNHM 002194): adult; collecting data as for holotype.
Other material. UPLBMNH Z-NS-0092: 2 adults, PTAGS 121, Camarines Norte Province, Caramoan Peninsula, low-elevation forest on karst, 13° 44.40’ N, 123° 54.38’ E, 245m asl, 21 May 2001, leg. Y. Hong, M. Levi, P. Nillos. KUNHM 002195: 1 adult, PTAGS 128, Camarines Norte Province, Caramoan Peninsula, low-elevation forest on karst 13° 46.09’ N, 123° 55.08’ E, 31m asl, 24 May 2001, leg. Y. Hong, M. Levi, P. Nillos.
Description. Unpigmented, body 35–52 mm x 2–2.2 mm (vii), 2.1–2.4 mm (xv), 2.1–2.6 mm (xxv); 123–206 segments; body cylindrical to slightly oval in cross-section. First dorsal pore 12/13; spermathecal pores paired in 7/8, 8/9, 0.13 circumference apart; fe-male pore single, male pore openings crescentic, paired in xviii on small flat porophores, 0.15 circumference apart, 6 setae between male pores; clitellum annular xiv–xvii. Setae regularly distributed around segmental equators; estimated 170–250 setae on vii, 110–160 se-tae on x, 82–100 setae on xxv; no dorsal gaps, ventral gaps AA:AB = 2.5:1. Genital markings paired presetal, lateral to male pores xix, thickened white areas across ventral surface xix–xxi (Figure 3C). Nephridiopores visible as gaps in setal rings, longitudinal musculature of postclitellar segments, 4 pores per segment in regu-lar rows.
Septa 5/6–9/10 thick, muscular, 10/11 thinner, re-mainder membranous. Weak gizzard in viii with typi-cal iridescent outer wall but flaccid, esophagus valvular xviii (4), xix (1), intestinal origin 1/2xix (1), xix (4), no caeca; thick intestinal gizzards xxvii–xxix (1) or xxvi–xxviii (4); typhlosole low fold xxxix, xl–lviii. In-testine vascularized xix–xxvi, constricted xxxviii (1), xxxix (4).
Hearts x–xiii esophageal, commissural vessels ix lateral; vii, viii divide into several small vessels, appa-rently none reaching ventral vessel. Supra-esophageal vessel x–xvi, extra-esophageal vessels form from se-veral smaller vessel in iii, iv, branches dorsally to sup-ra-esophageal vessel in ix, other branch along ventral

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 12
esophageal wall to connect with efferent parieto-eso-phageal vessels from body wall of xvi, xvii.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall the-reafter; from xix 4–6 micronephridia per segment, 2 meganephridia per segment from xix posteriorly, these attached to paired ureters adherent to either side of dor-sal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermathe-ca with blunt cylindrical ampulla, clearly differentiated muscular duct; simple club-shaped diverticulum half or more length of main spermathecal axis joins duct near body wall via differentiated stalk, (Figure 3D). Male sexual system holandric, testes and funnels enclosed in annular sacs in x, xi, these sacs encompass hearts, dor-sal vessel, seminal vesicles; sacs formed of membranes connecting septa 9/10 and 10/11, 10/11 and 11/12; se-minal vesicles xi, xii loosely acinous; each prostate ra-cemose four-lobed occupying xviii; duct not muscular; vas deferens join prostatic duct one third distance from gland to body wall; copulatory bursae lacking.
Remarks. Pleionogaster caramoanensis is a small-bodied species with only three intestinal gizzards and only four micronephridia per intestinal segment. The other species with low numbers of nephridia are P. sor-sogonensis and P. isarogensis; both have many fewer setae per segment, and more gizzards (Tables 1, 2). The former has preclitellar genital markings, the latter only postclitellar midventral markings, whereas P. cara-moanensis has only postclitellar paired markings.
Pleionogaster sorsogonensis n. sp.(Fig. 3E, F)
Etymology. Named after the province that includes the type locality.
Type material. Holotype (NMA 004146): adult, PTAGS 051, Sorsogon Province, Bulusan National Park, dipterocarp forest at Bulusan Lake, 12° 45.32’ N, 124° 05.34’ E, 360m asl, 3 May 2001, leg. S.W. James, A. Castillo, K. James, P. James.
Description. Unpigmented, body 46 mm x 2.3 mm (vii), 2.6 mm (xxv); 117 segments (amputee); body cy-lindrical. First dorsal pore 13/14; spermathecal pores paired in 7/8, 8/9, 0.11 circumference apart; female pore not seen, male pore openings crescentic, paired in xviii on small porophores, 0.15 circumference apart, 2 setae between male pores; clitellum not developed. Setae regularly distributed around segmental equators; estimated 120 setae on vii, 114 setae on x, 72 setae on xxv; no dorsal, ventral gaps. Genital markings midven-tral 10/11, 11/12, xvi, xx–xxii; paired xvii, xix (Figure 3E). Nephridiopores visible as gaps in setal rings, lon-
gitudinal musculature of postclitellar segments, 4 pores per segment in regular rows.
Septa 5/6–9/10 thick, muscular, 10/11–12/13 thin-ner, remainder membranous. Weak gizzard in viii with typical iridescent outer wall but flaccid, esophagus valvular xviii, intestinal origin xix, no caeca; thick in-testinal gizzards xxv–xxix, gizzards occupy posterior half of each segment as usual but anterior half of gut wall xxv–xxix somewhat iridescent, also covered with dense segmental network of blood vessels connected to dorsal and ventral vessels; typhlosole low fold xl–xlvii, one fourth lumen diameter. Intestine constricted 38/39, thereafter coated with brown chloragogen layer.
Hearts x–xiii esophageal, commissural vessels vii–ix lateral; supra-esophageal vessel x–xiv, extra-esophage-al vessels not traceable; paired large longitudinal ves-sels mid-lateral on body wall from xviii posteriorly.
Nephridia present as peptonephridia on anterior face septa 5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall thereafter; from xix four micronephridia per segment, two clearly stomate meganephridia per segment from xix posteri-orly, these attached to paired ureters adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermatheca club-shaped ampulla, poorly differentiated duct; simple club-shaped diverticulum half or more length of main spermathecal axis joins duct near body wall (Figure 3F). Male sexual system holandric, testes and funnels enclosed in annular? or ventrally separated? sacs in x, xi, these sacs encompass hearts, dorsal vessel, seminal vesicles; sacs formed of membranes connecting sep-ta 9/10 and 10/11, 10/11 and 11/12; seminal vesicles xi, xii loosely acinous; each prostate racemose two to three-lobed occupying xviii; duct slightly muscular; vas deferens slender, non-muscular; copulatory bursae lacking.
Remarks. Pleionogaster sorsogonensis appears to be related to P. caramoanensis, with which it shares the same body size, pigmentation, and number of micro-nephridia (Table 2). Pleionogaster sorsogonensis has two more intestinal gizzards, fewer setae in the anterior segments, fewer setae between the male pores, differ-ent genital marking patterns on the male field and in the spermathecal segments, and spermathecae whose ducts and ampullae are not greatly differentiated.
Pleionogaster nautsae n. sp.(Fig. 3G, H)
Etymology. Named after Phyllis Nauts of Connecti-cut, USA, as a gift from her husband.
Type material. Holotype (NMA 004147): adult, PTAGS 071, Catanduanes Province, lower riparian for-

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 13
est near Barangay San Miguel, Pangabinan, LMK039, 13° 54.5’ N, 124° 11.0’ E, 212m asl, 23 May 2001, leg. S.W. James, J. ffitch, A. Castillo. Paratypes (KUNHM 002196): 2 adults; collecting data as for holotype.
Other material. UPLBMNH Z-NS-0093: 2 adults, PTAGS 068, Catanduanes Province, low-elevation forest near Barangay Summit, Buradan, 13° 43.60’ N, 124° 17.14’ E, 210m asl, 21 May 2001, leg. S.W. James, P. James, J. James, K. James, J. ffitch, A. Cas-tillo. KUNHM 002197: 1 adult, PTAGS 070, Catan-duanes Province, lower montane forest near Barangay San Miguel, Pangabinan, no GPS data, 305m asl, 23 May 2001, leg. S.W. James, J. ffitch, A. Castillo.
Description. Unpigmented, body 158–295 mm x 3.3–4.6 mm (vii), 3.1–4.0 mm (xv), 3.5–4.3 mm (xxv); 286–343 segments; body cylindrical. First dorsal pore 12/13; spermathecal pores paired in 7/8, 8/9, 0.13 cir-cumference apart deep in furrows; female pore single, male pore openings crescentic, paired in xviii on small porophores, 0.12–0.15 circumference apart, 0–7 setae between male pores; pore orientation rotated so that crescent concavity directed antero-laterally rather than straight toward anterior; clitellum xiv–1/2xvii, annular. Setae regularly distributed around segmental equators; estimated 170–264 setae on vii, 150–196 setae on x, 92–106 setae on xxv; asetal zones midventral in one or more of vii–ix. Genital markings narrow transverse midventral extending to male pore lines presetal xvii, xix, latter with embedded paired marks, paired indented markings 17/18, broad epidermal thickenings ventral presetal, equatorial annuli of xx, xxi; ventral epidermal thickenings presetal ½ vii, viii to ½ ix, x (Figure 3G). Nephridiopores visible as gaps in setal rings, longitu-dinal musculature of postclitellar segments, 8 pores per segment in regular rows.
Septa 5/6–8/9 thick, muscular, 9/10/11 thinner, re-mainder membranous. Weak gizzard in viii with typi-cal iridescent outer wall, esophagus valvular xix or xx, intestinal origin xx or xxi, no caeca; intestine slightly vascularized xxi–xxiv, thick intestinal gizzards xxiv–xxix; typhlosole simple fold from liii, liv; intestine constricted 50/51 or 51/52.
Hearts x–xiii esophageal, commissural vessels vii–ix lateral; supra-esophageal vessel xi–xvii, extra-esophageal with branch from body wall of xiv, efferent posterior latero-parietal vessels travel up 15/16 to ex-tra-esophageal vessel in xv, these connected to paired longitudinal vessels on body wall xviii–xl; not always visible past xxx.
Nephridia present as peptonephridia on anterior faces septa 4/5/6, micronephridia preseptal clustered around commissural vessels vi–ix, on body wall there-after; from xix 8 micronephridia per segment, two sto-mate meganephridia per segment from xix posteriorly,
these attached to paired ureters adherent to either side of dorsal vessel.
Ovaries and funnels free in xiii, spermathecae paired in viii, ix without nephridia on ducts; each spermatheca club-shaped ampulla not differentiated from duct; short basally-attached diverticulum of similar shape (Figure 3H). Male sexual system holandric, testes, funnels en-closed in annular sacs in x, xi, these sacs encompass hearts, dorsal vessel, seminal vesicles; sacs formed of membranes connecting septa 9/10,10/11; 10/11, 11/12; seminal vesicles xi, xii, slender arcs with large dorsal block; prostates racemose occupying xviii; duct mus-cular; vas deferens slender,non-muscular, adheres to prostatic duct from duct-gland junction but visible on duct surface over ental one-third to one-half of duct; copulatory bursae lacking.
Remarks. Compared to P. nillosae, the other spe-cies with six intestinal gizzards in xxiv–xxix (see Table 2), P. nautsae has fewer setae and micronephridia per segment, more narrowly placed male and spermathe-cal pores, a typhlosole, and little or no differentiation of the spermathecal duct and ampulla. It is one of two species in Bicol with preclitellar midventral genital markings, the other being P. sorsogonensis which has half as many micronephridia, a more anterior intestinal constriction and paired post-clitellar genital markings.
DiscussionAmong those species for which many individuals were available for study, relative homogeneity of somatic and sexual characters is the rule. This is in contrast to the definition of P. horsti generated by synonymizing several species under this name (Easton 1979). Even so, the species described here would fall well outside that wide net.
In the course of this study, several characters not generally recorded in previous work proved useful in distinguishing species. In the post-giceriate intestine there is always a constriction, either side of which both internal and external intestinal surfaces differ. The in-ternal surface typically changes from rough and corru-gated anterior of the constriction to smooth posterior of it. The external intestinal surface is more densely covered with tan or brown chloragogen posterior of the constriction. In addition, the typhlosole (if present) al-ways begins after the constriction, whereas among the caecate genera of the Pheretima complex the typhlosole (if present) begins just after the caeca (range xxii–xx-vii) and thus many segments anterior to the typhloso-les of Pleionogaster species. No other known member of the Pheretima complex of genera (Sims and Easton 1972; Easton 1979) has these intestinal features, or the intestinal gizzards of Pleionogaster.

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 14
Along the body wall in the anterior intestinal seg-ments there is generally a longitudinal vessel on each side, extending from the region of segments xv–xviii back for 20–30 or even more segments. Sometimes this is not visible, probably due to lack of blood at fixati-on. Frequently this long vessel is clearly connected to a posterior latero-parietal vessel, which usually brings blood from the clitellar body wall to the extra-esopha-geal vessel (s) located on the ventral side of the eso-phagus. More rarely, this same vessel is seen to also connect to the prostate gland circulation. Among the Pheretima group of genera, I have seen this longitudi-nal vessel on the body wall of the intestinal segments in Pleionogaster only. Gates (1972) reports a similar vessel in Lampito, and I have seen a similarly placed vessel in a Honduran Ramiellona (James, unpublished data).
All Pleionogaster possess an arrangement of nephri-dia unique within the perichaetine Megascolecidae. Their micronephridia are arranged in regular rows with externally visible pores in the intestinal segments, and in clusters along the anterior septal faces and commis-sural vessels in the head segments. The nephropores in-terrupt the setal rings and longitudinal musculature, and so are easily seen. The number of micronephridia per segment appears to be relatively constant within a spe-cies, only P. tiwiensis varying by more than a pair, and varies among species (range 4–14). While it is general-ly true that larger species have more micronephridia per segment, there are exceptions, notably P. bulusanensis with 14 though not being particularly large-bodied. All species have a pair of stomate meganephridia in each intestinal segment, connected via tubules to ureters pa-rallel to either side of the dorsal vessel.
The testes sacs of Pleionogaster are often different from other perichaetine megascolecids in that some are composed of membranes joining the septa enclosing the testicular segments. In most other Pheretima group genera, and in the remaining Pleionogaster, the sacs do not enclose all the segmental contents, and usually only the testes and funnels. The Pleionogaster sacs are most often annular, but in some cases they are U-shaped, and open dorsally; in others the U is open ventrally, in which case I have described them as horseshoe-shaped. Annular sacs also occur in Philippine Pithemera and Polypheretima (James, unpublished data) but in these genera the sacs are never attached to both neighboring septa.
Upon consideration of these apparent synapomor-phies Pleionogaster has little in common with the ge-nera of the Pheretima complex, other than being peri-chaetine and having the same male terminalia. Even the latter show some variation from the common plan, as Pleionogaster have a vas deferens that remains separa-
te from and visible on the surface of the prostatic duct over the ental half or more of the duct, before it mer-ges and dives in to unite with the prostatic duct lumen. In the other genera, such as Pheretima and Amynthas, the vas deferens lumen joins the prostatic duct near the branching point of the prostatic duct as it receives the ductlets of the glandular lobes.
Pleionogaster is so far known only from the Philip-pines, and no closely similar genera have been disco-vered in other parts of SE Asia or the Australian regi-on, including New Guinea. How, and whence, it or its ancestor arrived on the Philippines and spread through-out the major islands without ending up anywhere else remains a mystery.
Even though the collecting teams made prodigious efforts at each site visited, only a few species were col-lected from more than one area. Pleionogaster bico-lensis seems to be widely distributed across the Bicol region, except for the extreme south where it was not found on Mt. Bulusan. Pleionogaster caramoanensis is abundant and widespread on the Caramoan Peninsula. This low-elevation limestone karst area was unsuitab-ly dry at the time of our visit, thus we have probab-ly not found all the species present at the Caramoan collection sites. Pleionogaster isarogensis was found at many collecting stations spanning a wide range of elevations within primary forest on Mt. Isarog, but not at any other location. Therefore, like most of the spe-cies described here, it is probably endemic to its own particular mountain. It is worth recalling here that even within the Mt. Malinao massif there is geographical variation of intestinal features in P. castilloi. The ex-tensive deforestation of the Philippines renders impos-sible any educated guesses about the original ranges of Bicol’s native earthworm species. It is possible that some survive in scattered spots of older secondary fo-rest or other areas with low disturbance frequency, but due to time and permit constraints we focused our ef-forts on easily defined locations where primary forest remains.
AcknowledgementsThis research was supported by the United States Nati-onal Science Foundation through grant DEB-0072764 to the author and Maharishi University of Manage-ment. John Stimson prepared the illustrations. Hendon Chubb made a contribution to the author’s research budget on behalf of his wife Phyllis Nauts. Thanks are due to the Philippine Department of Environment and Natural Resources, particularly to the Protected Areas and Wildlife Bureau staff who issued our permits, and numerous DENR provincial offices and their emplo-yees who worked with us in the field or simply pro-vided good information on access points and contacts

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 15
for the collecting sites. The National Power Corpora-tion (NAPOCOR) provided access to the Mt. Malinao area within their geothermal development zone. Hong Yong, Augusto Castillo, Portia Nillos, Matthew Levi, Jana ffitch, Dorothy Franke, Joy James, Pearl James,
Katharine James and David James provided assistance in the field. Finally, we thank the people of Barangay Summit, Catanduanes, for opening their homes to us during our stay there.
ReferencesBeddard, F.E., 1895. A Monograph of the Order of Oligo-
chaeta. Clarendon Press, Oxford, UK.Easton, E.G., 1979. A revision of the ‘acaecate’ earthworms
of the Pheretima group (Megascolecidae: Oligochaeta): Archipheretima, Metapheretima, Planapheretima, Pleionogaster and Polypheretima. Bull. Brit. Mus. Nat. Hist. (Zool.) 35, 1–128.
Gates, G.E., 1943. On some American and oriental earthworms part II. Family Megascolecidae. Ohio J. Sci. 43, 99–116.
Gates, G.E., 1972. Burmese earthworms. An introduction to the systematics and biology of megadrile oligochaetes with special reference to Southeast Asia. Trans. Am. Phil. Soc. 62, 1–326.
Heaney, L.R., 1985. Zoogeographic evidence for midd-le and late Pleistocene land bridges to the Philippine islands. Mod. Quat. Res. SE Asia 9, 127–143.
Heaney, L.R., 1993. Biodiversity patterns and the conser-vation of mammals in the Philippines. Asia Life Sci. 2, 261–274.
James, S.W., 2004. New species of Amynthas, Pheretima and Pleionogaster (Clitellata: Megascolecidae) of the Mt. Kitanglad Range, Bukidnon, Mindanao Island, Phil-ippines. Raffles Bull. Zool. 52, 289–313.
Jamieson, B.G.M., 1971. A review of the megascoleco-id earthworm genera (Oligochaeta) of Australia. Part I—Reclassification and checklist of the megasolecoid genera of the world. Proc. R. Soc. Qld. 82, 75–86.
Jamieson, B.G.M., 2000. Native Earthworms of Australia. PDF document on CD-ROM. Science Publishers, Inc., Enfield, New Hampshire, USA.
Michaelsen, W., 1892. Terricolen der Berliner zoologischen Sammlung, II. Arch. Naturg. 59, 209–261.
Michaelsen, W., 1896. Oligochäten Kükenthal—Ergebnisse einer zoologischen Forschungsreise in den Molukken und in Borneo. Abh. Senckenb. Ges. 23, 192–243.
Sims, R.W., Easton, E.G., 1972. A numerical revision of the earthworm genus Pheretima auct. (Megascolecidae: Oligochaeta) with the recognition of new genera and an appendix on the earthworms collected by the Royal Society North Borneo Expedition. Biol. J. Linn. Soc. 4, 169–268.
Stephenson, J., 1930. The Oligochaeta. Clarendon Press, Oxford, UK.
Stephenson, J., 1933. Oligochaeta from Australia, North Carolina, and other parts of the world. Proc. Zool. Soc. Lond. 1932, 899–941.

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 16
tiwiensis albayensis bicolensis bulusan-ensis
hongi ffitchae viracensis isarogensis
Pigmentation none brown brown none pink brown brown none
Setae vii, xxv 210, 114 148, 85 130–220, 140
200, 120 176, 104 220, 160 200, 140 180, 76
Spermathecal pore spacing
0.08 0.20 0.17 0.15 0.18 0.18 0.13 0.10
Male pore spacing 0.12 0.18 0.17 0.17 0.18 0.16 0.16 0.11
Setae betw. male pores
5, 6 12 8–10 20 10–12 8 10 4–8
Midventral genital marks
yes yes none none none none yes yes
Paired genital marks
yes yes yes yes yes yes yes none
Nephridia/seg. 6–10 12 12 14 12 12 10 6
Intestinal orgin xix xx xix or xx xx xx xix xix xx
Typhlosolar origin xlvii xlvii none none xlvi l lii xli
Testes sacs annular annular open ventral annular annular annular annular annular x, U-shaped xi
Spermath. duct differentiated
no no yes no no no yes yes
Latero-parietal vessels
xiv–xvii xiv–xvii xiv, xv xiv–xvi xiv–xvii xiv–xvi xiv, xv xv
Table 1: Pleionogaster species with four intestinal gizzards in xxvii–xxx.

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 17Table 2. Pleionogaster species with three, five, or six intestinal gizzards.
malinaoensis castilloi nillosae caramoanensis sorsogonensis nautsae
Pigmentation none none none none none none
Setae vii, xxv 160, 100 160, 80 314, 140 170–250, 110–160
120, 72 170–260, 100
Spermathecal pore spacing
0.10 0.17 0.20 0.13 0.11 0.13
Male pore spacing 0.09 0.17 0.18 0.15 0.15 0.13
Setae betw. male pores
2–4 6–8 0 6 2 0–7
Midventral genital marks
yes yes ? none yes yes
Paired genital marks yes none yes yes yes none
Nephridia/seg. 10–12 6 10 4 4 8
Intestinal orgin xix xx xx xix xix xx, xi
Typhlosolar origin xlv xlv or xlviii
none xl xl liii
Gizzards 5 5–7 6 3 5 6
Testes sacs annular annular annular annular annular annular
Spermath. duct differentiated
yes yes yes yes no no
Latero-parietal vessels
xiv–xvii xiv–xvii xiv, ? xvi, xvii ? xv
Table 2: Pleionogaster species with three, five, or six intestinal gizzards.

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 18
5m
m
5m
m
5m
m
5m
m
10m
m
1m
m
1 mm
1m
m
2m
msp
sp
sp
sp
sp3
mm
xviii
xviii
xviii
xviii
xviii
A
B
C
D
E
F
G
H
I
J
Fig. 1: A, B. Pleionogaster tiwiensis sp.n.; (A) ventral view, (B) spermatheca. C, D. Pleionogaster albayensis sp.n.; (C) ventral view, (D) spermatheca. E, F. Pleionogaster bicolensis sp.n.; (E) ventral view, (F) spermatheca. G, H. Pleiono-gaster bulusanensis sp.n.; (G) ventral view, (H) spermatheca. I, J. Pleionogaster hongi sp.n.; (I) ventral view, (J) sper-matheca. Abbreviation: sp=spermathecal pores.
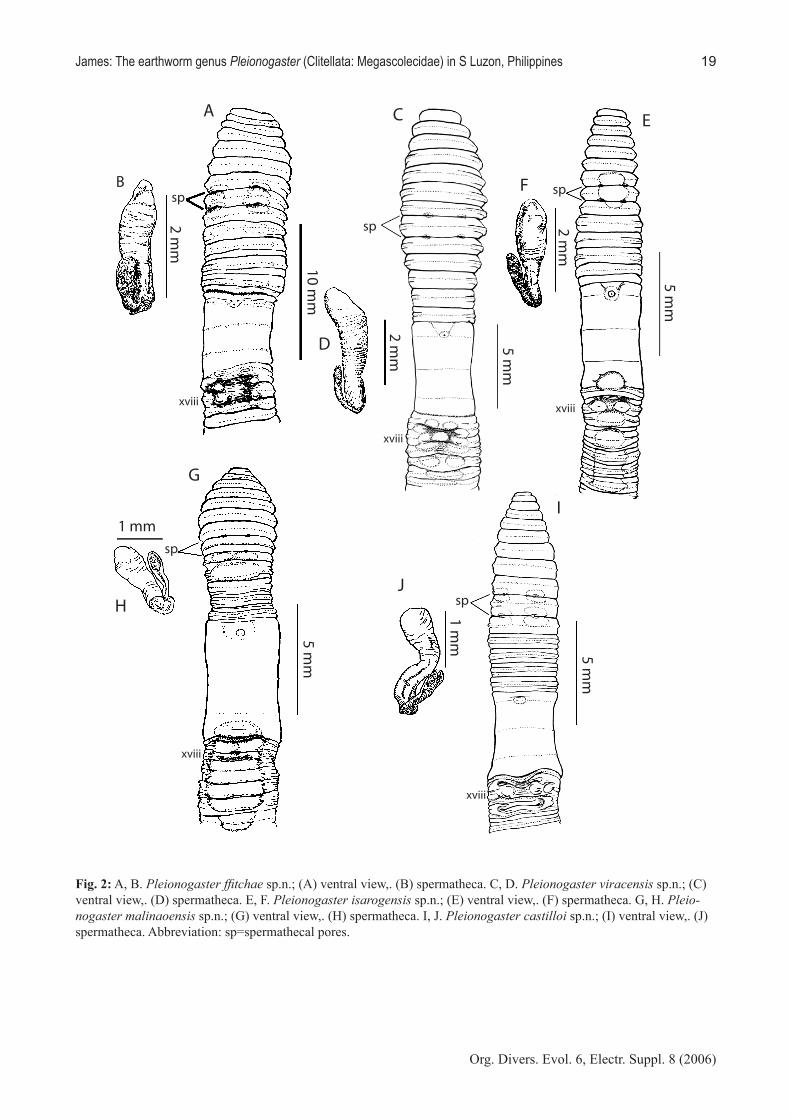
Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 19
B
A C E
G
H
I
J
sp
2m
m
F
2m
m
10m
m
D
2m
m
1 mm
1m
m5m
m
5m
m
5m
m
5m
m
sp
sp
sp
sp
xviii
xviii
xviii
xviii
xviii
Fig. 2: A, B. Pleionogaster ffitchae sp.n.; (A) ventral view,. (B) spermatheca. C, D. Pleionogaster viracensis sp.n.; (C) ventral view,. (D) spermatheca. E, F. Pleionogaster isarogensis sp.n.; (E) ventral view,. (F) spermatheca. G, H. Pleio-nogaster malinaoensis sp.n.; (G) ventral view,. (H) spermatheca. I, J. Pleionogaster castilloi sp.n.; (I) ventral view,. (J) spermatheca. Abbreviation: sp=spermathecal pores.

Org. Divers. Evol. 6, Electr. Suppl. 8 (2006)
James: The earthworm genus Pleionogaster (Clitellata: Megascolecidae) in S Luzon, Philippines 20
B
AC
E
G
H
sp
1m
m
F
3m
m
10m
m
D
3m
m
1m
m
5m
m
5m
m
5m
m
sp
sp
sp
xviii
xviii
xviii
xviii
Fig. 3: A, B. Pleionogaster nillosae sp.n.; (A) ventral view,. (B) spermatheca. C, D. Pleionogaster caramoanensis sp.n.; (C) ventral view,. (D) spermatheca. E, F. Pleionogaster sorsogonensis sp.n.; (E) ventral view,. (F) spermatheca. G, H. Pleionogaster nautsae sp.n.; (G) ventral view,. (H) spermatheca. Abbreviation: sp=spermathecal pores.





























