Embed Size (px)
Citation preview

ORIGINAL PAPER
The Down-Regulation of Neuroligin-2 and the Correlative ClinicalSignificance of Serum GABA Over-Expression in Hirschsprung’sDisease
Hongchao Yang • Jianyi Niu • Jian Wang •
Fan Zhang • Qiangye Zhang • Wentong Zhang •
Aiwu Li
Received: 23 January 2014 / Revised: 17 April 2014 / Accepted: 12 May 2014
� Springer Science+Business Media New York 2014
Abstract The goal of this study was to investigate the
expression level of neuroligin-2 in different colon tissue
segments of children with Hirschsprung’s disease (HSCR)
and the correlative clinical significance of serum Gamma-
Aminobutyric Acid (serum GABA) in HSCR. Neuroligin-2
was assessed by Immunohistochemistry staining method on
routine paraffin section from different colon tissue seg-
ments of HSCR (ganglionic colonic segment, transitional
colonic segment and aganglionic colonic segment). Wes-
tern-blot analysis and real-time fluorescence quantitative
PCR(qRT-PCR) were applied to compare and evaluate the
expression levels of neuroligin-2 from three segments of
HSCR, and we used Enzyme-linked Immunosorbent Assay
(ELISA) method to detect and compare the serum GABA
between HSCR and non-HSCR. Immunohistochemistry
staining demonstrated that intensive neuroligin-2 staining
was detected in the ganglion cells in the ganglionic colonic
and transitional colonic segments from the HSCR children;
however, neuroligin-2 staining was down-regulated
significantly in the aganglionic colonic segments. The
expression levels of neuroligin-2 mRNA and protein in the
aganglionic colonic segment were decreased compared to
the ganglionic colonic segment and transitional colonic
segment (P \ 0.05). And the level of serum GABA was
significantly higher in HSCR than that in non-HSCR. The
expression of neuroligin-2 varies from different segments
of HSCR. The down-regulation of neuroligin-2 in agan-
glionic colonic segments may be correlated with the
excessive intestine contraction and further result in HSCR.
The over-expression of serum GABA may be considered as
a new diagnostic method of HSCR.
Keywords Hirschsprung’s disease � Neuroligin-2 gene �Serum GABA
Introduction
Hirschsprung’s disease (HSCR), which is also called
aganglionosis, is a congenital disorder that affects 1 of
5,000 human births and is characterized by colonic stasis
due to the absence of enteric neurons in the distal gut [1],
leading to tonic contraction of the affected segment,
intestinal obstruction and massive distension of the proxi-
mal bowel. In 80 % of cases, the aganglionic tract involves
the short segment HSCR, while only 20 % of cases it
extends towards the long segment HSCR [2].
To date, the cause of HSCR still remains unclear, but
there is a common understanding that HSCR is a complex
disease influenced by multiple genetic factors and envi-
ronmental factors. Several genes such as ret proto-onco-
gene (RET), sex determining region Y-box 10(SOX10) and
endothelin receptor B (EDNRB), which play important
roles in the formation of ENS, have been identified
Hongchao Yang and Jianyi Niu have contributed equally to this work
and should be considered co-first authors.
H. Yang � J. Wang � Q. Zhang � W. Zhang � A. Li (&)
Department of Pediatric Surgery, Qilu Hospital, Shandong
University, 44#, Wenhua Xi Road, Jinan 250012, Shandong,
People’s Republic of China
e-mail: [email protected]
J. Niu
Qingzhou Clinical School, Weifang Medical University,
Qingzhou 262500, Shandong, People’s Republic of China
F. Zhang
Department of E.N.T, Qilu Hospital, Shandong University,
44#, Wenhua Xi Road, Jinan 250012, Shandong,
People’s Republic of China
123
Neurochem Res
DOI 10.1007/s11064-014-1334-y

involved in the pathogenesis of HSCR in human beings [3],
the alteration and abnormal expression of these genes,
however, only account for 30 % of the cases of HSCR [4].
Neuroligins are a family of postsynaptic transmembrane
proteins that bind to presynaptic neurexins [5], whereby
they form a trans-synaptic signal transduction complex and
mediate a bidirectional signaling between the presynaptic
axon and the postsynaptic target [6]. Presynaptic neurexins
and postsynaptic neuroligins likely work together with
other synaptic cell adhesion molecules of the cadherin and
ephrin families, and secreted proteins such as pentraxins, to
mediate central nervous system (CNS) synaptogenesis [7].
In the CNS, neurexins and neuroligins gained major
interest upon discovery that they induce formation and
function of synapses in recombinant non-neuronal cell
systems, and reports of preferential distribution of neurol-
igin-1 in glutamatergic synapses (excitatory synapses) and
neuroligin-2 in GABAergic synapses (inhibitory synapses)
[8–10] fuelled considerable efforts to elucidate their role in
synapse formation and specification. And it has been con-
firmed that [11] the expression level of glutamate and
GABA could reflect the expression level of neuroligin-1
and neuroligin-2, respectively.
Studies have demonstrated that there is bi-directional
communication between the enteric nervous system
(ENS) and the CNS [12]. And the brain is continuously
informed by afferent nerves detecting gut activity, nev-
ertheless it has been established that psychological state
has a great impact on gut function [13]. Whether neu-
roligins which expressed in CNS are expressed in ENS?
Our previous study had identified that [14, 15] neuroligins
were expressed on the ENS of human beings with HSCR
and the expression was down-regulated in aganglionic
colonic segments.
In the present study, we choose to further investigate the
expression of neuroligin-2 on ENS of HSCR and the serum
GABA related with neuroligin-2 and discuss whether the
pathogenesis of HSCR is involved with the abnormal
inhibitory synapses and whether the serum GABA could be
used as a new diagnostic method of HSCR.
Materials and Methods
Patients and Samples
Our study was approved by the ethics committee of Qilu
Hospital, Shandong University. The experimental samples
were collected from the surgical excision waste tissue and
preoperative routine blood collection. And no harm had
been done to patients at all, the special written consent was
not necessary according to the opinion of the ethics com-
mittee of Qilu Hospital, Shandong University.
Fifty two patients (2 months–5 years, 39 boys, and 13
girls) pathologically confirmed HSCR and 52 patients
(1–5 years, 48 boys, and 4 girls) with indirect inguinal
hernia (IIH) were regarded as control group were involved
in this study and they were all treated in Department of
Pediatric Surgery, Qilu Hospital, Shandong University
from September 2011 to June 2013. The cases of short-
segment type and long-segment type with HSCR were 42
and 10, respectively. All HSCR patients were treated with
Soave’s pull-through procedure. The children with HSCR
were not treated by any drugs before operative treatment.
Reagents
The detailed information of antibodies and primers was
listed in Tables 1 and 2, respectively and other reagents
were also available commercially: Protein extraction kit
(Beyotime, China), BCA Protein concentration determi-
nation kit (Beyotime, China), SDS-PAGE Gel Preparation
kit (Beyotime, China), Total RNA isolation kit (RNAiso
Plus, TaKaRa, Japan), Reverse transcription kit (Prime-
Script� RT reagent Kit with gDNA Eraser, TaKaRa,
Japan), SYBR� Premix Ex TaqTM Tli RNaseH Plus (Ta-
KaRa, Japan), Serum Gamma-Aminobutyric Acid ELISA
kit (E90900Ge, Uscn Life Science Inc., China).
Methods
Samples Preparation
Samples of aganglionic, transitional and ganglionic colon
segments were harvested from the surgical excision waste
colon of children patients with HSCR at the length of 3 cm
respectively. Specimens were divided into two parts. One
part was fixed in 10 % buffered formaldehyde for the
preparation of Immunohistochemistry. Immediately,
another part of fresh colon tissue was snap-frozen in liquid
nitrogen and stored at -80 �C in disinfected tubes until
used for the Quantitative real-time polymerase chain
reactions (qRT-PCR) assay and Western-blot analysis.
1.5 ml fresh blood sample was collected by preoperative
routine blood collection from all 104 patients (52 HSCR
and 52 IIH) and was allowed to clot for two hours at room
temperature before centrifugation (20 min, 1,0009g). Then
the serum samples were stored in aliquot at -80 �C and
prepared for ELISA.
Immunohistochemistry Staining
Paraffin-embedded blocks of different full-thickness colon
segments were sectioned with cross-sections at a thickness
of 4 lm, then heated to 65 �C, dewaxed in xylene (15 min,
3 times) and graded alcohols (100, 95, 80, 70 %; 5 min
Neurochem Res
123

each). Antigen retrieval was performed with 0.01 M citrate
buffer at pH 6.0 at 95 �C for 20 min. After incubation with
3 % H2O2 in methanol for 15 min at 37 �C to block
endogenous peroxidases, the sections were blocked with
6 % Bull Serum Albumin in PBS and incubated with pri-
mary antibody: anti-neuroligin-2 (1:50 dilution, goat
polyclonal, sc-14089, Santa Cruz Biotechnology, USA)
overnight at 4 �C. After washing with 0.1 M PBST (pH
7.4) (5 min, 3 times), antibodies were detected using the
Polink-2 plus� Polymer HRP Detection System for Goat
Primary Antibody (PV-9003, ZSGB-BIO, China) accord-
ing to the manufacturer’s instructions. The reaction was
visualized with the 3,30-diaminobenzidine tetrahydrochlo-
ride DAB Kit. Then sections were counterstained with
Harris’s haematoxylin, differentiated using hydrochloric
acid ethanol and blued using ammonia water. After dehy-
dration in graded alcohols and clearing with dimethylben-
zene, the slides were mounted with neutral gum and
examined using the Image-Pro� Plus analytical imaging
system (Media Cybernetics, Bethesda, MD, USA).
Western-Blot Analysis
20 mg frozen specimens from three different segments of
HSCR were homogenized. Then, 200 ll lysis buffer
(RIPA:PMSF 100:1, Cell Lysis Buffer, Beyotime, China)
was added to the homogenate. The supernatant containing
total protein from the tissue homogenate was separated by
centrifuging for 15 min at 12,000 r/min. Protein concentra-
tions were measured using a BCA protein kit to normalize the
amount of total protein. Samples containing equal amounts
of proteins (30 lg/sample) were separated on an 8 % SDS-
PAGE and then electrophoretically transferred to polyvi-
nylidene fluoride membranes (PVDF). After blocking with
6 % (w/v) nonfat milk and washing with Tris-buffered sal-
ine-Tween solution (TBST), membranes were incubated
with primary antibodies anti-neuroligin-2 (1:200, goat
polyclonal, sc-14089, Santa Cruz Biotechnology, USA) and
anti-b-actin (1:2,000, mouse polyclonal, ZSGB-BIO, China)
overnight at 4 �C. After washing, the blots were incubated
with HRP-conjugated secondary antibodies: rabbit anti-goat
IgG and goat anti-rat IgG (1:5,000, ZSGB-BIO, China) at
room temperature for 60 min, respectively. The membranes
were subsequently detected with the ECL system according
to the manufacturer’s instructions, and the expression levels
of neuroligin-2 were expressed by the relative gray values
(neuroligin-2 IOD/b-actin IOD), which was analyzed by the
Gel-Pro analyzer 4.0 Software.
RNA Isolation and qRT-PCR Assay
Total RNA was isolated from 25 mg specimens of three
different segments of HSCR by Total RNA Isolation Kit
(RNAiso Plus, TaKaRa, Japan) according to the manufac-
turer’s instructions. The quantity of RNA was assessed
spectrophotometrically. The OD260/280 of the RNA
Table 1 Detailed information
of antibodiesAntigen Primary antibody Dilution Applications Source
Neuroligin-2 Goat-anti-human polyclonal 1:50 Detect Nlgn-2 with
immunohistochemistry on
paraffin-embedded
sections
Santa Cruz,
USA
Neuroligin-2 Goat-anti-human polyclonal 1:200 Detect Nlgn-2 with
Western-blot
Santa Cruz,
USA
b-Actin Rat-anti-human Secondary
antibody
1:2,000 Western-blot internal
reference
ZSGB-BIO,
China
Anti-
neuroligin-2
Poly-HRP anti-goat IgG 1:200 Label Nlgn-2 on on
paraffin-embedded
sections
ZSGB-BIO,
China
Anti-
neuroligin-2
HRP-conjugated rabbit-anti-
goat IgG
1:5,000 Detect Nlgn-2 with
Western-blot
Santa Cruz,
USA
Anti-b-actin HRP-conjugated goat-anti-rat
IgG
1:5,000 Detect b-actin with
Western-blot
Santa Cruz,
USA
Table 2 Detailed information
of primers
F upstream primer, R
downstream primers
Primers Primer sequences (50 ? 30) Annealing temperature (�C) Product size (bp)
Neuroligin-2 F: CCAGTCTCCCGTCTACTTTTACA 59 316
R: CTGCTTCTCCTTGCTGTTGAAT
b-Actin F: AGCGAGCATCCCCCAAAGTT 60 285
R: GGGCACGAAGGCTCATCATT
Neurochem Res
123

samples ranged between 1.70 and 2.00. Then 1 lg of total
RNA was used in a 20 ll cDNA synthesis reaction at 37 �C
for 15 min, followed by 85 �C for 5 s, the SYBR� Premix Ex
Taq TM II(Perfect Real Time), which included 59 gDNA
Eraser Buffer 2 ll,gDNA Eraser 1 ll,Total RNA 1 lg, 59
PrimeScript� Buffer 2(for Real Time)4 ll,PrimeScript. RT
Enzyme Mix I 1 ll, RT Primer Mix 1 ll. The qRT-PCR
reactions were performed with reference to SYBR� Premix
Ex TaqTM (Tli RNaseH Plus) quantitative fluorescence kit in
Roche Applied Science LightCycler� 480 Real-time PCR
systems, and SYBR� Premix Ex TaqTM (Tli RNaseH Plus)
(29) 10 ll, PCR Forward and Reverse Primers (10 lM)
1 ll, cDNA 2 ll were added in the reverse transcription
system.
After the reaction, the Ct of neuroligin-2 mRNA from
each sample was measured and the 2-DDCt was calculated.
The results were expressed relative to the number of b-actin
transcripts used as an internal control. All experiments were
conducted in triplicate. The primers and annealing temper-
atures used for the amplification of human neuroligin-2 and
the endogenous control b-actin were shown in Table 2.
Enzyme-Linked Immunosorbent Assay
The 104 serum samples (52 HSCR and 52 IIH) stored in
aliquot at -80 �C were used for the detection of serum
GABA with reference to the Serum Gamma-Aminobutyric
Acid ELISA kit (E90900Ge, Uscn Life Science Inc.,
China) specification. Then after the reaction, the O.D.
value was measured at 450 nm and then the actual con-
centration of samples could be calculated according to the
standard concentration. Each sample was examined in
triplicates with the average value as the final result.
Statistical Analyses
GraphPad Prism� 5 for Windows software (La Jolla, CA,
USA) was used for the statistical procedures. The data were
expressed as mean ± SEM. Two group comparisons were
evaluated by unpaired t test and multiple comparisons were
analyzed by one-way ANOVA and the Tukey’s test. All
P values were two-sided and a value of P \ 0.05 was
considered statistically significant.
Results
Immunohistochemistry Staining
Immunohistochemistry were performed on paraffin-
embedded sections (from ganglionic, transitional and agan-
glionic segments), and light microscopy was used for
observation. The histological appearance and the expression
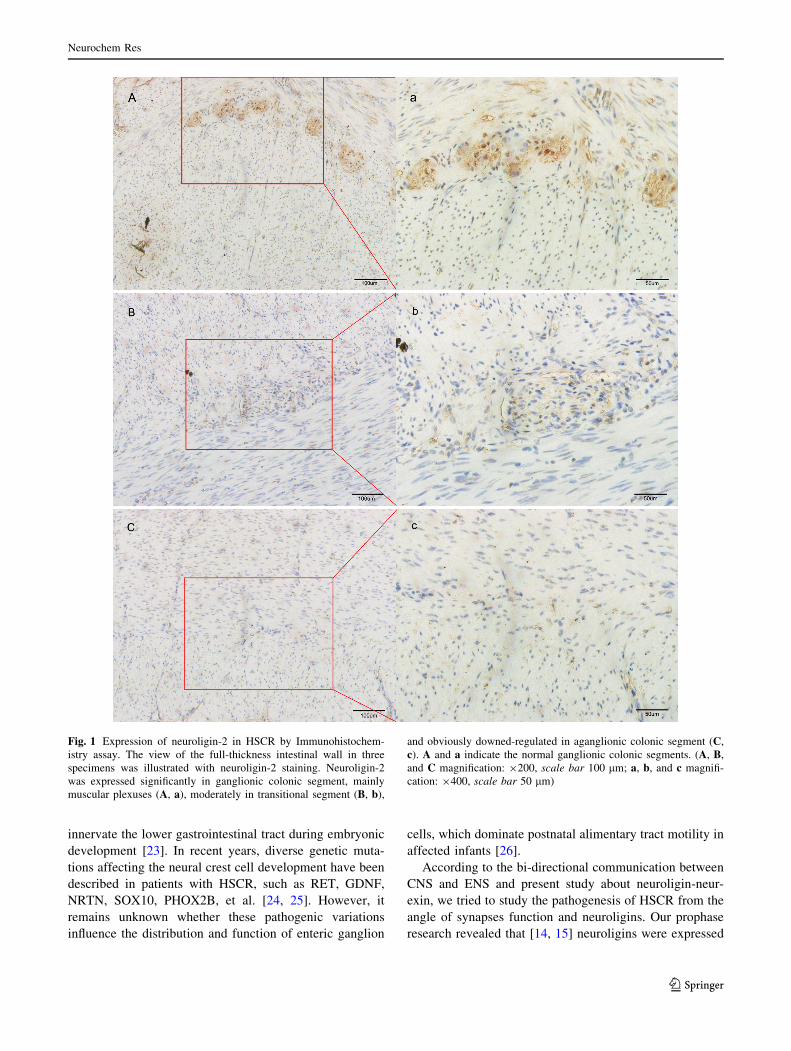
of neuroligin-2 were shown in Fig. 1. Intensive neuroligin-2
staining was detected in myenteric plexuses in the ganglionic
and transitional segments (Fig. 1A, a; B, b). The myenteric
plexuses were not formed in aganglionic segment, and
neuroligin-2 staining was down-regulated significantly in
these segments (Fig. 1C, c).
Western-Blot Analysis
Figure 2 represented that neuroligin-2 were expressed sig-
nificantly in ganglionic colonic segments (0.057 ± 0.0018,
n = 52), moderately in transitional colonic segments
(0.039 ± 0.0014, n = 52) and obviously downed-regulated
in aganglionic colonic segments (0.012 ± 0.0011, n = 52),
and the three difference of relative gray values had statistical
significance (P \ 0.05).
Real-Time Fluorescence Quantitative RT-PCR Assay
To obtain further insights into the expression of the target
genes, we performed qRT-PCR described above in the
method. As shown in Fig. 3, neuroligin-2 mRNA was
significantly expressed at obvious low levels in the agan-
glionic colonic segments [(1.20 ± 0.11) E-5, n = 52]
compared with those of normal ganglionic [(9.46 ± 0.21)
E-5, n = 52)] and transitional colon segments [(4.74 ±
0.23) E-5, n = 52; P \ 0.05].
Enzyme-Linked Immunosorbent Assay
As shown in Fig. 4, concentrations of serum GABA were
measured by ELISA. For serum GABA, there was a sig-
nificant difference in HSCR patients (23.69 ± 2.30) com-
pared to IIH patients (7.65 ± 1.16, P \ 0.05).
Discussion
A number of studies have shown that the excitatory/
inhibitory synapse balance in the brain is believed to be a
key to neuronal plasticity mechanisms such as learning and
memory, and also a likely mediator of mental disorders
such as autism [16–20]. And a mismatch of neurexin and
neuroligin partners across synapses in the brain presumably
leads to loss of synaptic plasticity and/or erroneous wiring,
resulting in behavioral and cognitive deficiencies [21].
Normal gastrointestinal motility results from the coor-
dinated interplay of multiple cooperating mechanisms, both
intrinsic and extrinsic to the gastrointestinal tract [22].
HSCR is a developmental disorder characterized by the
absence of ganglion cells in the lower digestive tract.
Aganglionosis is attributed to a disorder of the enteric
nervous system (ENS) whereby ganglion cells fail to
Neurochem Res
123
innervate the lower gastrointestinal tract during embryonic
development [23]. In recent years, diverse genetic muta-
tions affecting the neural crest cell development have been
described in patients with HSCR, such as RET, GDNF,
NRTN, SOX10, PHOX2B, et al. [24, 25]. However, it
remains unknown whether these pathogenic variations
influence the distribution and function of enteric ganglion
cells, which dominate postnatal alimentary tract motility in
affected infants [26].
According to the bi-directional communication between
CNS and ENS and present study about neuroligin-neur-
exin, we tried to study the pathogenesis of HSCR from the
angle of synapses function and neuroligins. Our prophase
research revealed that [14, 15] neuroligins were expressed
Fig. 1 Expression of neuroligin-2 in HSCR by Immunohistochem-
istry assay. The view of the full-thickness intestinal wall in three
specimens was illustrated with neuroligin-2 staining. Neuroligin-2
was expressed significantly in ganglionic colonic segment, mainly
muscular plexuses (A, a), moderately in transitional segment (B, b),
and obviously downed-regulated in aganglionic colonic segment (C,
c). A and a indicate the normal ganglionic colonic segments. (A, B,
and C magnification: 9200, scale bar 100 lm; a, b, and c magnifi-
cation: 9400, scale bar 50 lm)
Neurochem Res
123

on the postsynaptic neurons and interstitial cells of Cajal in
mesenteric plexus of HSCR patients with a down-regulated
expression levels in the aganglionic colonic segment. On
the basis of our previous research, we went on our study
about the expression of neuroligin-2 and the serum GABA
related with neuroligin-2 and discussed whether the path-
ogenesis of HSCR was involved with the abnormal inhib-
itory synapses and whether the serum GABA could be used
as a new diagnostic method of HSCR.
Clinically, sometimes it is difficult for the diagnosis of
HSCR only based on the preoperative radiology, especially
for the newborn. So a new method to solve this problem must
be found. And, in this study, we tried to detect the serum
GABA which was based on the expression of neuroligin-2
and found its differential expression between HSCR and
non-HSCR which might be meaningful for the diagnosis of
HSCR. Our data showed that neuroligin-2 was expressed
significantly in ganglionic colonic segments, moderately
transitional colonic segments and down-regulated signifi-
cantly in aganglionic colonic segments. Moreover, the
expression level of serum GABA was obviously higher in
HSCR than that in non-HSCR. According to these, it can be
inferred that the abnormality of neuroligin-2 is closely
Fig. 2 Neuroligin-2 expression levels in children with HSCR by
Western blot. Protein extracts from normal ganglionic (1, 2),
transitional (3, 4), and aganglionic (5, 6) colon segments were probed
by Western blot analysis with neuroligin-2 antibodies in HSCR (a).
The relative gray values of neuroligin-2 expressed in Western-blot
was 0.057 ± 0.0018 in ganglionic colonic segments, 0.039 ± 0.0014
in transitional segments and 0.012 ± 0.0011 in aganglionic colonic
segments, and the difference of relative gray values had statistical
significance (0.057 ± 0.0018 vs. 0.039 ± 0.0014, * P \ 0.05;
0.057 ± 0.0018 vs. 0.012 ± 0.0011, * P \ 0.05; 0.039 ± 0.0014
vs. 0.012 ± 0.0011, * P \ 0.05) (b)
Fig. 3 Quantitative RT-PCR results of Neuroligin-2 expression in
different colonic segments. The Neuroligin-2 transcript was signif-
icantly decreased in aganglionic colonic segments compared with
other colonic segments (* P \ 0.05)
Fig. 4 Serum GABA in HSCR and IIH patients. We observed that
the level of serum GABA was significantly higher in HSCR
(23.69 ± 2.30) than that in IIH controls (7.65 ± 1.16, * P \ 0.05)
Neurochem Res
123

related to HSCR, and we conclude that the alterative and
abnormal expression of neuroligin-2 may play an important
role in the pathogenesis of HSCR through affecting the
inhibitory synaptic function. And the serum GABA may be
used as a new diagnostic method of HSCR.
Although our study does not provide direct evidence that
neuroligin-2 plays an important role in ENS development,
the neuroligin-2 expression levels in different segments can
be the basis for further investigations on the potential role
of neuroligin-2 and this study provides a new research
perspective for recognizing the molecular mechanism
underlying HSCR development. Of course, additional
studies are required to confirm the superior method for
diagnosing HSCR and we have confidence in the method of
the combination of research and clinic.
Conclusions
The results above show that neuroligin-2 is expressed in
human beings’ ENS, and for HSCR children, the expression
of neuroligin-2 from different colon segments may play an
important role in the pathogenesis of this disease: the obvi-
ously down-regulations of neuroligin-2 in aganglionic
colonic segment may be correlated with the excessive
intestine contraction and further result in HSCR. The
expression level of serum GABA coincided with neuroligin-
2 may be considered as a new diagnostic method of HSCR.
Acknowledgments We appreciate the financial support provided by
the National Natural Science Foundation of China (81270720) and
the Independent Innovation Foundation of Shandong University, II-
FSDU (2012ZD026). We thank all of the patients and their parents
involved in this study.
Conflict of interest The authors declare that there are no conflicts
of interest.
References
1. RobertsRR,Bornstein JC,BergnerAJ,YoungHM (2008)Disturbances
of colonic motility in mouse models of Hirschsprung’s disease. Am J
Physiol Gastrointest Liver Physiol 294(4):996–1008
2. Edery P, Pelet A, Mulligan LM, Abel L, Attie T et al (1994) Long
segment and short segment familial Hirschsprung’s disease: vari-
able clinical expression at the RET locus. J Med Genet
31(8):602–606
3. Heanue TA, Pachnis Vassilis (2007) Enteric nervous system
development and Hirschsprung’s disease: advances in genetic and
stem cell studies. Nat Rev Neurosci 8(6):466–479
4. Emison ES, McCallion AS, Kashuk CS (2005) A common sex-
dependent mutation in a RET enhancer underliea Hirschsprung
disease risk. Nature 434(7035):857–863
5. Sun M, Xing G, Yuan L, Gan G, Knight D et al (2011) Neuroligin
2 is required for synapse development and function at the dro-
sophila neuromuscular junction. J Neurosci 31(2):687–699
6. Ichtchenko K, Nguyen T, Sudhof TC (1996) Structures, alterna-
tive splicing, andneurexin binding of multiple neuroligins. J Biol
Chem 271:2676–2682
7. Siddiqui TJ, Craig AM (2011) Synaptic organizing complexes.
Curr Opin Neurobiol 21:132–143
8. Zhanyan Fu, Vicini Stefano (2009) Neuroligin-2 accelerates
GABAergic synapse maturation in cerebellar granule cells. Mol
Cell Neurosci 42(1):45–55
9. Varoqueaux F, Jamain S, Brose N (2004) Neuroligin 2 is
exclusively localized to inhibitory synapses. Eur J Cell Biol
83:449–456
10. Song JY, Ichtchenko K, Sudhof TC, Brose N (1999) Neuroligin 1
is a postsynaptic cell-adhesion molecule of excitatory synapses.
Proc Natl Acad Sci USA 96:1100–1105
11. Hoon M, Bauer G, Fritschy JM, Moser T, Falkenburger BH,
Varoqueaux F (2009) Neuroligin 2 controls the maturation of
GABAergic synapses and information processing in the retina.
J Neurosci 29:8039–8050
12. Graf ER, Kang Y, Hauner AM, Craig AM (2006) Structure
function and splice site analysis of the synaptogenic activity of
the neurexin-1 b LNS domain. J Neurosci 26(16):4256–4265
13. Wouters MM, Boeckxstaens GE (2011) Neuroimmune mecha-
nisms in functional bowel disorders. Neth J Med 69(2):55–61
14. Wang J, Mou Y, Zhang Q, Zhang F, Yang H et al (2013)
Expression and significance of neuroligins in myenteric cells of
cajal in Hirschsprung’s disease. PLoS One 8(6):e67205
15. Zhang Q, Wang J, Li A, Liu H, Zhang W et al (2013) Expression
of neurexin and neuroligin in the enteric nervous system and their
down-regulated expression levels in Hirschsprung disease. Mol
Biol Rep 40(4):2969–2975
16. Chih B, Engelman H, Scheiffele P (2005) Control of excitatory
and inhibitory synapse formation by neuroligins. Science
307(5713):1324–1328
17. Graf ER, Zhang X, Jin SX, Linhoff MW, Craig AM (2004)
Neurexins induce differentiation of GABA and glutamate post-
synaptic specializations via neuroligins. Cell 119(7):1013–1026
18. Levinson JN, El-Husseini A (2005) Building excitatory and
inhibitory synapses: balancing neuroligin partnerships. Neuron
48(2):171–174
19. Prange O, Wong TP, Gerrow K, Wang YT, El-Husseini A (2004)
A balance between excitatory and inhibitory synapses is con-
trolled by PSD-95 and neuroligin. Proc Natl Acad Sci USA
101(38):13915–13920
20. Sudhof TC (2008) Neuroligins and neurexins link synaptic
function to cognitive disease. Nature 455(7215):903–911
21. Biswas S, Reinhard J, Oakeshott J, Russell R, Srinivasan MV
et al (2010) Sensory regulation of neuroligins and neurexin I in
the honeybee brain. PLoS One 5(2):e9133
22. Du P, O’Grady G, Davidson JB, Cheng LK, Pullan AJ (2010)
Multiscale modeling of gastrointestinal electrophysiology and
experimental validation. Crit Rev Biomed Eng 38(3):225–254
23. Tam PK, Garcia-Barcelo M (2009) Genetic basis of Hirsch-
sprung’s disease. Pediatr Surg Int 25(7):543–558
24. Amiel J, Sproat-Emison E, Garcia-Barcelo M, Lantieri F, Bur-
zynski G et al (2008) Hirschsprung disease, associated syndromes
and genetics: a review. J Med Genet 45(1):1–14
25. Brooks AS, Bertoli-Avella AM, Burzynski GM, Breedveld GJ,
Osinga J et al (2005) Homozygous nonsense mutations in
KIAA1279 are associated with malformations of the central and
enteric nervous systems. Am J Hum Genet 77(1):120–126
26. Shimotake T, Tomiyama H, Aoi S, Iwai N (2003) Discrepancy
between macroscopic and microscopic transitional zones in Hir-
schsprung’s disease with reference to the type of RET/GDNF/
SOX10 gene mutation. J Pediatr Surg 38(5):698–701
Neurochem Res
123











![Intestinal pseudo-obstruction: adult Hirschsprung’s ...radiologyupdate.org/f/2018/06/Intestinal pseudo...ious causes [4]. One of them is Hirschsprung’s disease (HD), which is considered](https://img.pdfslide.us/doc/110x75/5e4e08035522ee140639de6b/intestinal-pseudo-obstruction-adult-hirschsprungas-pseudo-ious-causes.jpg)
















