Embed Size (px)
Citation preview

]. PhytOpalhology 123,259-272 (1988) © 1988 Paul Parey Scielllific Publishers, Berlin and Hamburg ISSN 0931-1785
Universitdt Konstanz, Fakultdt fur Biologie, Lehrstuhl fur Phytopathologie, D-7750 Konstanz
The Differentiation of Infection Structures as a Result of Recognition Events
between some Biotrophic Parasites and their Hosts
K. MENDGEN, A. SCHNEIDER, M. STERK and W. FINK
AUlhors' address: K. MENDGEN, A. SCHNEIDER, M. STERK, and W. FINK, Universilal Konslanz, Fakuhal fur Biologie, LehrSlUhl fur PhytOpalhologie, D-7750 KonSlanz (F.R.G.).
With 7 figures
Received March 24, 1988; accepted May 10, 1988
Abstract
MOSl uredospores of rUSl fungi develop infeclion SlrUClUres in a lypical pallern so lhal lhey can infecl lhe hosl plam. The funclion of lhese infeclion SlrUClures is divided intO lhe following three phases:
I. In lhe recognilion phase, lhe germ lube recognizes lhe cUlicle and lhe stOma. This process may occur independemly from lhe hOSl plant since copies of lhe cUlicle induce similar reaclions of lhe fungus. During fungal growlh on lhe epidermis, unspecific Slress responses of lhe planl are lriggered. .
2. In lhe signal phase, lhe fungal substOmalal vesicle and infeclion hypha(e) 'contacl lhe hOSl cells wilhin lhe leaf parenchyma. A signal from lhe hosl induces funher developmem of lhe fungus. HaustOrium mOlher cell differentialion is effecled and hauslorium formalion is inilialed. Al lhe same lime, lhe fungus suppresses lhe synlhesis of slfe;s melaboliles by lhe planl.
3. In lhe parasilic phase, lhe fungus penelrales lhe hOSl cell and complex imeraclions belween hOSl and parasile begin. A highly specialized interface around lhe haustOrium develops presumably in order to allow a more efficient nUlriem lransfer from hOSl IQ parasile. Eventual defence reaclions of lhe plant, generally on lhe race-cuilivar level, fail IQ be evoked or are suppressed in compalible combinalions.
Zusammenfassung
Die Ausbildung von Infektionsstrukturen als Folge von Erkennungsreaktionen zwischen einigen biotrophen Parasiten und ihren Wirtspflanzen
Del' Infeklionsverlauf del' dikaryolischen Uredospore del' ROslpilze wird in die folgenden drei Phasen umeneiil:
I. In der Erkennungsphase erkennl del' Keimschlauch die KUlikula und die Spailoffnung. Diesel' Yorgang kann auch unabhangig von del' Winspflanze ablaufen, da ein Abdruck der Pflanzen-
Presemed on lhe occasion of lhe 601h binhday of Prof. Dr. R. HEITEFUSS in GOllingen.
US. Copyrighl Clearance Center Code Stalemem 0931-1785/88/2303-0259$02.50/0

260 MENDGEN, SCHNEIDER, STERK and FINK
oberflache ahnliche Reize fur den Pilz abgibt. Wahrend des Wachstums des Pilzes auf der Epidermis wird bei der Wirtspflanze die Symhese von StreBmetaboliten ausgelost.
2. In der Signalphase nehmen das substomatare Vesikel und die Infektionshyphe des Pilzes Kontakt mit den Parenchymzellen im Blatt auf. Ein Signal der Wirtszellen induziert die weitere Entwicklung des Pilzes. Gleichzeitig unterdruckt der Pilz die weitere Synthese von aJlgemeinen StreBmetaboliten der Wirtspflanze.
3. In der parasitischen Phase dringt der Pilz in die Wirtszelle ein und komplizierte Wechselwirkungen zwischen Wirt und Parasit beginnen. Eine hochspezialisierte Imeraktionszone in der Umgebung des Haustoriums entwickelt sich, wahrscheinlich urn einen moglichst effektiven Transport von Metaboliten aus der Wirtszelle in das Pilzhaustorium zu ermoglichen. Weitere Abwehrreaktionen der Wirtszelle, hervorgerufen durch mehr oder weniger virulente Pilzrassen, werden vermieden oder unterdruckt.
Many plant pathogenic fungi differentiate specialized infection structures in order to penetrate into their host. In the monokaryotic phase, the basidiospore of the rust fungus Uromyces appendiculatus forms only a short germ tube before it differentiates an appressorium on the epidermis and perforates the cuticle. Within the epidermal cell, an intraepidermal vesicle develops from which hyphae start to colonize the host leaf (GOLD and MENDGEN 1984). The induction of these infection structures is nonspecific. The isolated cuticle of the host plant or even 5 % agar induce the formation of the intraepidermal vesicle (FREYTAG et al. 1988).
In the dikaryotic phase, most rust fungi develop more complex infection structures (Fig. 1 a): The germ tube crosses the array of wax crystals on the cuticle QOHNSON 1934) and grows towards a stoma. When the tip of the germ tube contacts the stomatal opening, appressorium formation begins. From the lower surface of the appressorium, an infection peg develops and penetrates into the substomatal cavity of the leaf where the substomatal vesicle with the infection hypha is formed. After close contact with the parenchymatous host cells, a haustorial mother cell develops which, in turn, differentiates a haustorium in the host cell. This sequence of specific infection structures seems to be the product of specific interactions between the host and the parasite. In this paper, three main phases, during which responses are triggered either by host and/or parasite, are discussed. These are the recognition, signal and parasitic phases (Fig. 1 b).
1. The recognition phase
After dissemination, the germination inhibitor in the rust uredospores diffuses away if there is sufficient moisture and the spore begins to germinate if light, temperature and water quality parameters allow this (see STAPLES and MAcKo 1984). The germ tube grows in close contact with the cuticle. In wheat, the wax crystal array of the host cuticle (LEwrs and DAY 1972), and the surface depressions located at anticlinal walls of epidermal cells, induce directed growth that increases the probability of the germ tube meeting a stoma. Gradients in pH at the leaf surface also influence the direction of germ tube growth of Uromyces viciae fabae (EDWARDS and BOWLING 1986). The tuning of germ tube tropism is very well adapted to the host plant surface since the different rust fungi find the
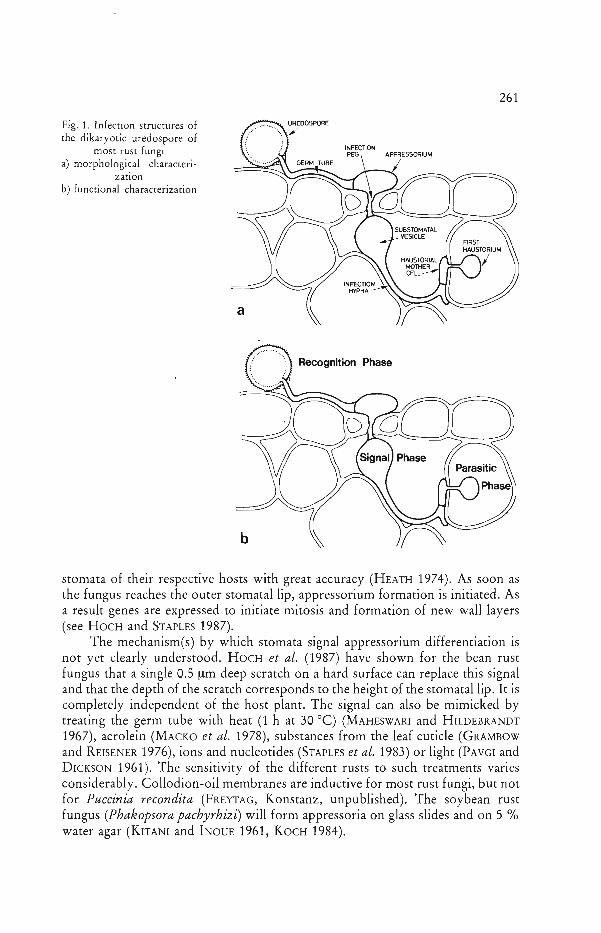
261 The Differentiation of Infection Structures
Fig. 1. Infection structures of the dikaryotic uredospore of
most rust fungi a) morphological characteri
zation b) functional characterization
Recognition Phase
stomata of their respective hosts with great accuracy (HEATH 1974). As soon as the fungus reaches the outer stomatal lip, appressorium formation is initiated. As a result genes are expressed to initiate mitosis and formation of new wall layers (see HOCH and STAPLES 1987).
The mechanism(s) by which stomata signal appressorium differentiation is not yet clearly understood. HoCH et al. (1987) have shown for the bean rust fungus that a single 0.5 f.lm deep scratch on a hard surface can replace this signal and that the depth of the scratch corresponds to the height of the stomatal lip. It is completely independent of the host plant. The signal can also be mimicked by treating the germ tube with heat (1 h at 30 QC) (MAHESWARI and HILDEBRANDT 1967), acrolein (MACKO et al. 1978), substances from the leaf cuticle (GRAMBow and REISENER 1976), ions and nucleotides (STAPLES et al. 1983) or light (PAVGI and DICKSON 1961). The sensitivity of the different rusts to such treatments varies considerably. Collodion-oil membranes are inductive for most rust fungi, but not for Puccinia recondita (FREYTAG, Konstanz, unpublished). The soybean rust fungus (Phakopsora pachyrhizi) will form appressoria on glass slides and on 5 % water agar (KITANI and INouE 1961, KOCH 1984).

262 MENDGEN, SCHNEIDER, STERK and FINK
STAPLES and HOCH (1982) first suggested that the sensing mechanism involves elements of the cytoskeleton of the hypha since as soon as appressorium formation is initiated, the microtubules and the F-actin cytoskeleton at the tip of the hypha are oriented parallel to the stomatal lips and a message starts (BOURETT et al. 1987). If there is such a signal chain between the hyphal tip and the nucleus to induce mitosis, the different modes of appressorium induction should either interfere with this signal chain or have the same effect on the hyphal tip. A second messenger system could be involved in this signal chain (STAPLES and HOCH 1987). We only know that the receptor at the hyphal tip is extremely sensitive, that the signal can be independent of the host (WYNN 1976), and is mediated by proteinase sensitive structures of the extracellular matrix of the germ tube (EpSTEIN et al. 1985).
During the germ tube growth on the leaf, the rust fungus elicits general defence reactions of the plant, such as the production of stress metabolites (YAMAMOTO et al. 1977) or the synthesis of ethylene (see PARADIES and ELSTNER 1980). Similarly, constituents of the germ tubes of rust fungi may induce the synthesis of phytoalexins in bean (Phaseolus vulgaris) leaves (HOPPE et al. 1980) or trigger the synthesis of lignin in wheat (Triticum aestivum) (MoERSCHBACHER et al. 1986). Chitin or chitosan also may elicit such defence reactions (YOUNG et al. 1982, DARVILL and ALBERSHEIM 1984). As with most other carbohydrate .. elicitors, the chitin detectable on the surface of the germ tubes of U. appendiculatus and U. vignae does not show race specific differences in its affinity to wheat germ agglutinin (Fig. 2). Also, the proteins with carbohydrate moieties in the germ tubes of P. graminis exhibit no race specificity (KIM and HOWES 1987) after two-dimensional gel electrophoresis.
2. The signal phase
After the differentiation of the appressorium, a penetration peg develops from this appressorium, grows into the substomatal cavity of the leaf and further differentiates a substomatal vesicle. The cytoplasm accumulates in this vesicle which then elongates to form the infection hypha. A second round of nuclear division occurs. The fungus is in very close contact with the host parenchymatous cells and there seems to be an exchange of specific signals between host and parasite. Many arguments favor this assumption:
6-14 h after inoculation, when the rust fungus has differentiated the appressorium and proliferates in the substomatal chamber, the ethylene production induced by bean rust (u. appendiculatus) germ tubes is reduced (PARADIES et al. 1979, PARADIES and ELSTNER 1980). During the same period, in crown rust (Puccinia coronata) inoculated Avena leaves, the activity of phenylalanine ammonia-lyase (YAMAMOTO et al. 1977) and lipoxygenase (YAMAMOTO and TAN! 1986) decrease during development of the infection hypha. Wall deposits (ONOE et al. 1987) can be observed on those cells that surround the stomata (subsidiary cells). All these unspecific reactions of the host cells are suppressed during subsequent growth of the infection hypha and secondary hyphae in the leaves. At the same time that the non-specific host responses begin to be suppressed, the rust
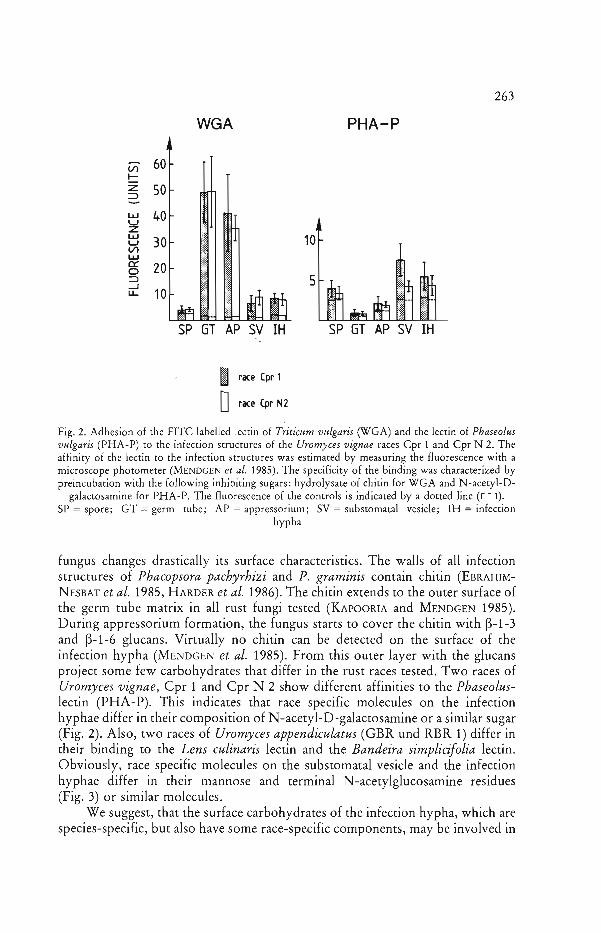
263 The Differentiation of Infection Structures
WGA PHA-P
V) 60 ~
z 50=> UJ U Z I.LJ U 10 V) I.LJ Cl: 0 :::> -l 5 I.L.
AP SV IH SP GT AP SV IH
I race (pr 1
orace (pr N2
Fig. 2. Adhesion of the FITC labelled lectin of Triticum vulgaris (WGA) and the lectin of Phaseolus vulgaris (PHA-P) to the infection structures of the Uromyces vignae races Cpr 1 and Cpr N 2. The affinity of the lectin to the infection structures was estimated by measuring the fluorescence with a microscope photometer (MENDGEN et al. 1985). The specificity of the binding was characterized by preincubation with the following inhibiting sugars: hydrolysate of chitin for WGA and N-acetyl-D
galactosamine for PHA-P. The fluorescence of the controls is indicated by a dotted line (r - 1). SP = spore; GT = germ tube; AP = appressorium; SV = subsromatal vesicle; IH = infection
hypha
fungus changes drastically its surface characteristics. The walls of all infection structures of Phacopsora pachyrhizi and P. graminis contain chitin (EBRAHIMN ESBAT et al. 1985, HARDER et al. 1986). The chitin extends to the outer surface of the germ tube matrix in all rust fungi tested (KAPOORIA and MENDGEN 1985). During appressorium formation, the fungus starts to cover the chitin with ~-1-3
and ~-1-6 glucans. Virtually no chitin can be detected on the surface of the infection hypha (MENDGEN et al. 1985). From this outer layer with the glucans project some few carbohydrates that differ in the rust races tested. Two races of Uromyces vignae, Cpr 1 and Cpr N 2 show different affinities to the Phaseoluslectin (PHA-P). This indicates that race specific molecules on the infection hyphae differ in their composition of N-acetyl-D-galactosamine or a similar sugar (Fig. 2). Also, two races of Uromyces appendiculatus (GBR und RBR 1) differ in their binding to the Lens culinaris lectin and the Bandeira simplicifolia lectin. Obviously, race specific molecules on the substomatal vesicle and the infection hyphae differ in their mannose and terminal N-acetylglucosamine residues (Fig. 3) or similar molecules.
We suggest, that the surface carbohydrates of the infection hypha, which are species-specific, but also have some race-specific components, may be involved in
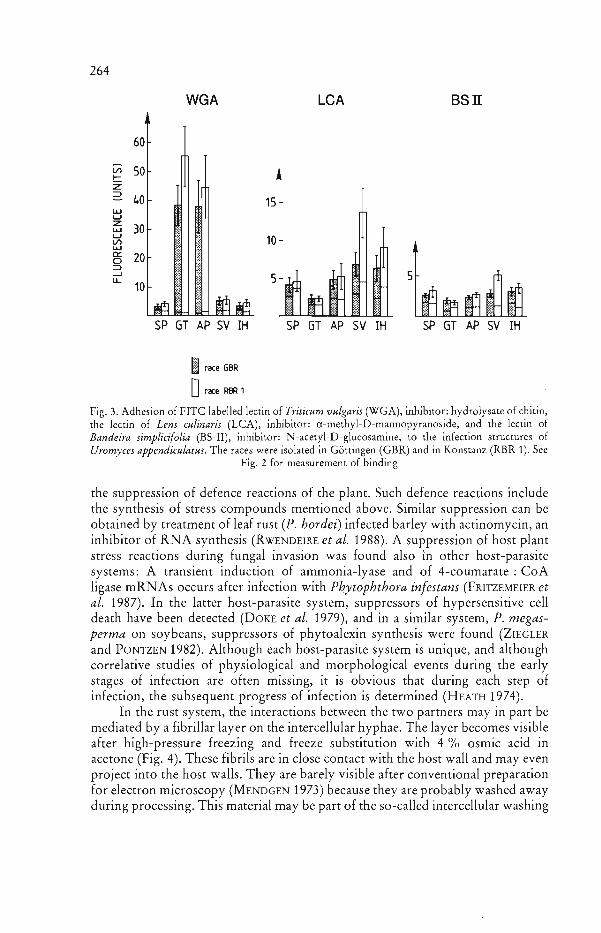
264 MENDGEN, SCHNEIDER, STERK and FINK
WGA LeA Bsn
60
l/l 50!::::: z ~ 40 15 I.LJ U
I.LJ z 30 U l/l 10 I.LJ t.. 0:: 0 20 $ ::::> --' f.u.. J 5
10
SP GT AP SV rH SP GT AP SV rH SP GT AP SV rH
I race GBR
orace RBR 1
Fig. 3. Adhesion of FITC labelled lectin of Triticum vulgaris (WGA), inhibitor: hydrolysate of chitin, the lectin of Lens culinaris (LCA), inhibitor: a-methyl-D-mannopyranoside, and the lectin of Bandeira simplicifolia (BS-II), inhibitor: N-acetyl-D-glucosamine, to the infection structures of Uromyces appendiculatus. The races were isolated in Gottingen (GBR) and in Konstanz (RBR 1). See
Fig. 2 for measurement of binding
the suppression of defence reactions of the plant. Such defence reactions include the synthesis of stress compounds mentioned above. Similar suppression can be obtained by treatment of leaf rust (P. hordei) infected barley with actinomycin, an inhibitor of RNA synthesis (RWENDEIRE et al. 1988). A suppression of host plant stress reactions during fungal invasion was found also in other host-parasite systems: A transient induction of ammonia-lyase and of 4-coumarate : CoA ligase mRNAs occurs after infection with Phytophthora infestans (FRITZEMEIER et al. 1987). In the latter host-parasite system, suppressors of hypersensitive cell death have been detected (DoKE et al. 1979), and in a similar system, P. megasperma on soy beans, suppressors of phytoalexin synthesis were found (ZIEGLER and PONTZEN 1982). Although each host-parasite system is unique, and although correlative studies of physiological and morphological events during the early stages of infection are often missing, it is obvious that during each step of infection, the subsequent progress of infection is determined (HEATH 1974).
In the rust system, the interactions between the two partners may in part be mediated by a fibrillar layer on the intercellular hyphae. The layer becomes visible after high-pressure freezing and freeze substitution with 4 % osmic acid in acetone (Fig. 4). These fibrils are in close contact with the host wall and may even project into the host walls. They are barely visible after conventional preparation for electron microscopy (MENDGEN 1973) because they are probably washed away during processing. This material may be part of the so-called intercellular washing

265 The Differentiation of Infection Srructu res
Fig. 4. Electronmicrograph of an intercellular hypha of Puccinia coronata after high pressure freezing (see MOLLER and MOOR 1984) and freeze substitution in acetone for 24 h at -90°C, followed by a treatment with 4 % OsO, in acetone at -90°C, -60 cC, - 30°C and O°C during 24 h for each step. The wall of the fungus (w) is covered with a layer of fibrillar structures
(x 28,000)
fluid (IWF) which is obtained after infiltration of rust-infected wheat leaves with buffer and subsequent gentle centrifugation. ROHRINGER and MARTENS (1987) detected race specific proteins in this IWF. Some molecules bind to concavalin A which indicates that they also contain a-linked glucose or mannose and appear to originate from the fungus (HOLDEN and ROHRINGER 1985). Unfortunately, we have no proof that these differences are responsible for pathogenicity of the two races or whether they are part of gene specific interactions.
It is obvious from these studies that the contact of the rust hyphae with the host parenchymatous wall plays an important role in regulating the interaction between both partners. The molecules responsible for the interaction may be secreted by a special secretory system detected in infection hyphae and in all subsequen t stages of the fungus (KNAUF and MENDGEN 1988, WELTER et al. 1988). Since we have a species specific monoclonal antibody (Fig. 5) that recognizes molecules of only the substomatal vesicle and the infection hyphae, it seems possible to isolate and follow the synthesis and secretion of species-specific molecules. It is conceivable, that the species-specific molecules secreted at this stage of development suppress or elicit in the non-host plant the host defences. Such molecules may play a role in establishment of basic compatibility or elicitation of non-host resistance mechanisms (see HEATH 1987).
The plant also influences the development of the infection hypha. After the induction of the appressorium in vitro, some rust fungi (e.g. the soybean rust (Ph. pachyrhizi) and the coffee rust (Hemileia vastatrix) (S. FREYTAG, Konstanz,
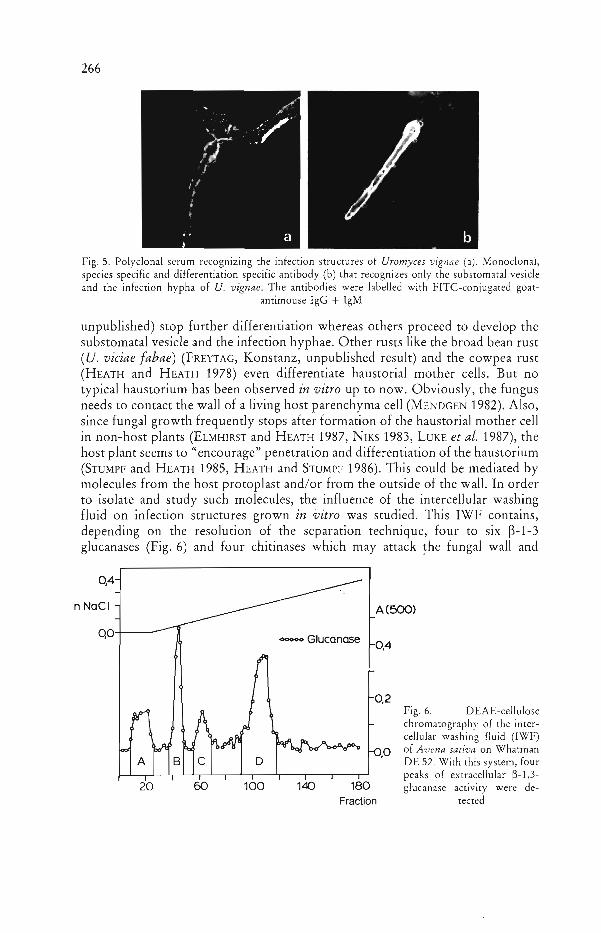
266 MENDGEN, SCHNEIDER, STERK and FINK
Fig. 5. Polyclonal serum recognizing the infection structures of Uromyces vignae (a). Monoclonal, species specific and differentiation specific antibody (b) that recognizes only the substomatal vesicle and the infection hypha of U. vignae. The antibodies were labelled with FITC-conjugated goat
antimouse IgG + IgM
unpublished) stop further differentiation whereas others proceed to develop the substomatal vesicle and the infection hyphae. Other rusts like the broad bean rust (u. viciae fabae) (FREYTAG, Konstanz, unpublished result) and the cowpea rust (HEATH and HEATH 1978) even differentiate haustorial mother cells. But no typical haustorium has been observed in vitro up to now. Obviously, the fungus needs to contact the wall of a living host parenchyma cell (MENDGEN 1982). Also, since fungal growth frequently stops after formation of the haustorial mother cell in non-host plants (ELMHJRST and HEATH 1987, NIKS 1983, LUKE et al. 1987), the host plant seems to "encourage" penetration and differentiation of the haustorium (STUMPF and HEATH 1985, HEATH and STUMPF 1986). This could be mediated by molecules from the host protoplast and/or from the outside of the wall. In order to isolate and study such molecules, the influence of the intercellular washing fluid on infection structures grown in vitro was studied. This IWF contains, depending on the resolution of the separation technique, four to six ~-1-3
glucanases (Fig. 6) and four chitinases which may attack ~he fungal wall and
0,4
n NoCI A (500)
QO-t---~ <>000<> Glucanase 0.4
Fig. 6. DEAE-cellulose chromatograph:' of the Intercellular washing fluid (IWF) of Avena saliva on Whatman
A B C o DE 52. With this system, four peaks of extracellular ~-1,3
20 60 100 140 180 glucanase activity were de-Fraction tected
267 The Differentiation of Infection Structures
liberate "messenger" molecules (FINK et al. 1988). However, neither a deleterious effect on the infection hyphae nor the induction of haustoria has been observed (MENDGEN, unpublished result).
3. The parasitic phase
The parasitIc phase of the dikaryotic rust mycelium starts either with haustorial mother cell differentiation or with the penetration of the host cell wall. First, a haustorial neck with its neck ring forms within the host cell. This neck distally enlarges and the haustorial body differentiates (HARDER and CHONG 1984, HEATH and HEATH 1975, LITTLEFIELD and HEATH 1978). The host plant may accumulate very specific membrane structures around the haustorium. Complexes induced by P. graminis in wheat or oats are the same, but those induced by P. coronata in oats are very different. This means that fungus species-specific changes are induced in the host prQtoplast (CHONG and HARDER 1982, HARDER et al. 1978). In all rust fungi studied up to now, the haustorium itself is surrounded by the extrahaustorial membrane. Although this membrane is continuous with the host plasmalemma, it differs considerably from the normal plasmalemma. It has different affinities to some stains (see LITTLEFIELD and HEATH 1979, HARDER and CHONG 1984), a much lower sterol ~ontent (HARDER and MENDGEN 1982) and lacks intra membrane particles and ATPase activity (LITTLEFIELD and BRACKER 1972, SPENCER-PHILIPS and GAY 1981). The meaning of these qualities is not understood yet. Perhaps, the fungus induces a modification of the plasmalemma
10~ 1 .......� .,..!0 o~-W u -10 z w u If) -20 w c:: g -30
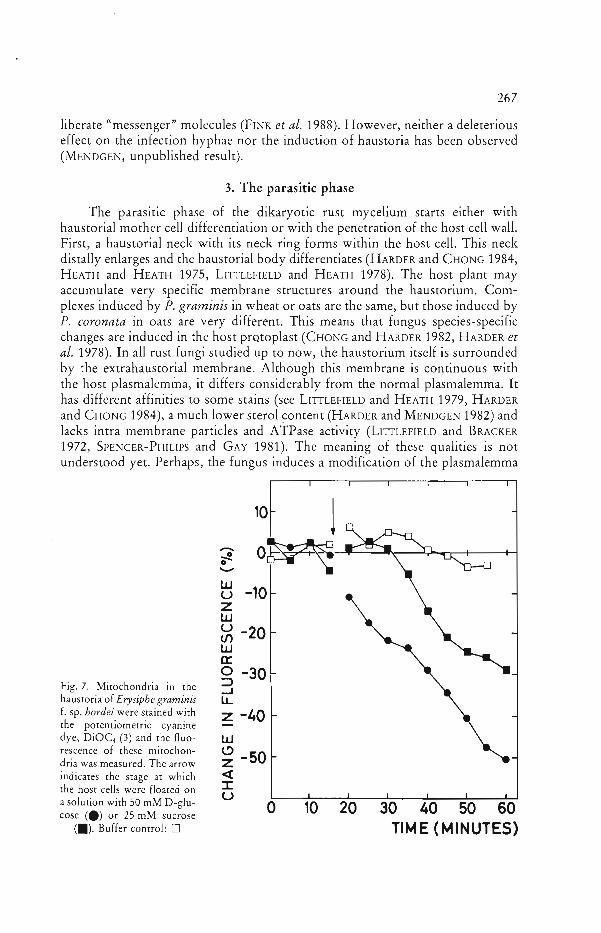
Fig. 7. Mitochondria in the ..J haustoria of Erysiphe graminis LL f. sp. hordei were stained with z -40 the potentiometric cyanine dye, DiOC, (3) and the fluo w rescence of these mitochon ~
dria was measured. The arrow z -50 indicates the stage at which <t
::r::the host cells were floated on Ua solution with 50 mM D-glu o 10 20 30 40 50 60 cose (e) or 25 mM sucrose
(.). Buffer control: 0 TIME (MINUTES)

268 MENDGEN, SCHNEIDER, STERK and FINK
around the haustorium that facilitates the flow of nutrients. In the rusts, we have no idea yet what nutrients are transferred from host to parasite (MENDGEN 1981).
In the case of the barley powdery mildew haustorium (Erysiphe graminis f. sp. hordei), we have evidence now that glucose is the sugar preferably transported from host to parasite. To show this, the membrane potential of haustorial mitochondria was studied with a potentiometric cyanine dye (BUSHNELL et al. 1987) which exhibits fluorescence in a polarized membrane. We assumed that the sugar which reaches the haustorium would first increase respiration there. As a consequence, the membrane potential of mitochondria increases. This increase of membrane potential was detected by measuring the quenching of fluorescence in mitochondria. An immediate effect was detected after feeding with glucose. The response was proportional to the glucose concentration added. Sucrose induced a reaction about 15-20 minutes later (Fig. 7). Galactose and L-glucose had no effect (MENDGEN and NASS 1988).
The events that induce this highly specialized interface between both partners and keep it in a balanced state, i.e. suppress host defence reactions and induce the flow of nutrients towards the parasite are completely unknown. In incompatible combinations, host defences generally begin at this stage of development. Protein synthesis again starts during differentiation of the rust haustorium and phenylalanine ammonia lyase, lipoxygenase and other enzyme activities increase (YAMAMOTO et al. 1977, YAMAMOTO and TAN! 1986). Also, phytoalexins are synthesized in oat tissue at this stage of the host-parasite interaction (MAYAM A 1982). Synthesis of phytoalexins, however, may not be correlated directly with resistance against bean rust infection (HOPPE et al. 1980). These reactions of the host seem to be regulated by the products of race specific genes (KEEN 1982). The effect can be abolished with antibiotics (TANI and YAMAMOTO 1978) and the plant becomes susceptible again. Similar events can be observed in barley powdery mildew (E. graminis f. sp. hordei) infected leaf tissue. Populations of mRNA increase during penetration of E. graminis and decrease after haustorium differentitation in a compatible combination (DAVIDSON et al. 1988). At the same stage of haustorial development, defence reactions against nonhost haustoria may be suppressed during the following one or two.days (KUNOH 1987). The regulation of such processes is still unknown. Perhaps, the extrahaustorial matrix that separates host and parasite contains the molecules that trigger or switch off the host responses. Unfortunately, these walls are not accessible to chemical analysis. Therefore, more studies are needed to prove that, during haustorium formation (i.e. the parasitic phase), a new type of interaction between host and parasite begins and is responsible for the defence reactions of the plant at the level of race specificity.
4. Conclusions
It is obvious that there are more or less specific interactions between host and parasite during the different stages of infection. Changes in the surface' structure of the cuticle may lead the fungus astray. During the initial contact in the substomatal cavity, signal molecules may stop or encourage the successful

269 The Differentiation of Infection Structures
colonization of the host. Even more delicate interactions occur during the parasitic phase when most race-specific interactions leading to compatibility or incompatibility are expressed. However, we have as yet no sound explanation for species- or race specific interactions. Also, we have no evidence for factors which lead to "horizontal" or "vertical" interactions (VANDERPLANK 1984). Perhaps a precise knowledge of the early interactions will improve our understanding of host and parasite and will explain the differences between species- and race specificity.
We want to thank the following colleagues and students. Dr. M. HEATH, TorontO, supplied the races of the cowpea rust and Dr. M. MULLER, ETH Zurich, allowed us to use the high pressure freezing instrument. Mrs. U. SCHWANZ produced the monoclonal antibodies and Drs. D. E. HARDER, F. H. J. RI]KENBERG, R. ROHRINGER and MARY ANN STUMPF helped with the manuscript. The Deutsche Forschungsgemeinschaft supported K. M. with grants.
Literature
BOURETT, T., H. C. HOCH, and R. C. STAPLES, 1987: Association of the microtubuli cytoskeletOn with the thigmotropic signal for appressorium formation in Uromyces. Mycologia 79, 540--549.
BUSHNELL, W. R., K. MENDGEN, and Z. Llu, 1987: Accumulation of potentiometric and other dyes in haustOria of Erysiphe graminis in living host cells. Physiol. Mol. Plant Pathol. 31, 237-250.
CHONG, J., and D. E. HARDER, 1982: Ultrastructure of haustOrium development in Puccinia coronata avenae: Some host responses. Phytopathology 72, 1527-1533.
DARVILL, A. G., and P. ALBERSHEIM, 1984: PhytOalexins and their e1icitors - a defence against microbial infection in plants. Annu. Rev. Plant Physiology 35, 243-275.
DAVIDSON, A. D., J. M. MANNERS, R. S. SIMPSON, and K. J. SCOTT, 1988: Altered host gene expression in near-isogenic barley conditioned by different genes for resistance during infection by Erysiphe graminis f. sp. hordei. Physiol. Mol. Plant Pathol. 32, 127-139.
DOKE, N., N. A. GARAS, and J. Kuc, 1979: Partial characterization and aspects of the mode of action of a hypersensitivity-inhibiting factor (HIF) isolated from Phytophthora infestans. Physiol. Plant Pathol. 15, 127-140.
EBRAHIM-NESBAT, F., H. H. HOPPE, and R. ROHRINGER, 1985: Lectin binding studies on the cell walls of soy bean rust (Phakopsora pachyrhizi Syd). Phytopath. Z. 114,97-107.
EDWARDS, M. c., and D. J. F. BOWLING, 1986: The growth of rust germ tubes towards stomata in relation to pH gradients. Physiol. Mol. Plant Pathol. 29, 185-196.
ELMHIRST, J. F., and M. C. HEATH, 1987: Interactions of the bean rust and cowpea rust fungi with species of the Phaseolus- Vigna plant complex. I. Fungal growth and development. Can. J. Bot. 65, 1096-1107.
EpSTEIN, L., L. LACETTI, R. C. STAPLES, H. C. HOCH, and W. A. HOOSE, 1985: Extracellular proteins associated with induction of differentiation in bean rust uredospore germlings. Phytopathology 75, 1073-1076.
FINK, W., M. LrEfLAND, and K. MENDGEN, 1988: Chitinases and ~-1,3-g1ucanases in the apoplastic compartment of oat leaves (Avena sativa L.). Plant Physiol. (in press).
FREYTAG, S., L. BRUSCAGLlONl, R. E. GOLD, and K. MENDGEN, 1988: Basidiospores of rust fungi (Uromyces species) differentiate infection structures in vitro. Exp. Mycology (in press).
FRITZEMEIER, K. H., C. CRETIN, E. KOMBRINK, F. ROHWER, J. TAYLOR, D. SCHEEL, and K. HAHLBROCK, J987: Transient induction of phenylalanine ammonia-lyase and 4-coumarate: CoA ligase mRNAs in potatO leaves infected with virulent or avirulent races of Phytophthora infestans. Plant Physiol. 85, 34-41.
GOLD,� R. E., and K. MENDGEN, 1984: Cytology of basidiospore germination, penetration and early colonization of Phaseolus vulgaris by Uromyces appendiculatus var. appendieulatus. Can. J. Bot. 62, 1989-2002.

270 MENDGEN, SCHNEIDER, STERK and FINK
GRAMIlOW, H. J., und H. J. REISENER, 1976: Differenzierung und Wachstum von Puccinia graminis f. sp. tritici in vitro. Ber. Dtsch. Bot. Ges. 89, 555-56l.
HARDER, D. E., R. ROHRINGER, D. J. SAMBORSKI, W. K. KIM, and J. CHONG, 1978: Electronmicroscopy of susceptible and resistant near isogenic (sr6/Sr6) lines of wheat infected by Puccinia graminis tritici. 1. The host-pathogen interface in the compatible (sr6/P6) interaction. Can. J. Bot. 56, 2955-2966.
- -, and K. MENDGEN, 1982: Filipin-sterol complexes in bean rust- and oat crown rust-fungal/plant interactions: Freeze-etch electron microscopy. Protoplasma 112,46--54. and J. CHONG, 1984: Structure and physiology of haustoria. In: BUSHNELL, W. R., and A. P. ROELFS (Eds), The Cereal Rusts, pp. 431-476. Academic Press, Orlando.
--, --, R. ROHRINGER, and W. KIM, 1986: Structure and cytochemistry of the walls ot urediospores, germ tubes, and appressoria of Puccinia graminis tritici. Can. J. Bot. 64, 476-485.
HEATH, M. c., 1974: Light and electron microscope studies of the interactions of host and non-host plants with cowpea rust - Uromyces phaseoli var. vignae. Physiol. Plant Pathol. 4, 403-414.
- -, and 1. B. HEATH, 1975: Ultrastructural changes associated with the haustorial mother cell during haustorium formation in Uromyces phaseoli var. vignae. Protoplasma 84, 297-314. and - -, 1978: Structural studies of the development of infection structures of cowpea rust, Uromyces phaseoli var. vignae. I. Nucleoli and nuclei. Can. J. Bot. 56, 648-66l. and M. A. STUMPF, 1986: Ultrastructural observations of penetration sites of the cowpea rust fungus in untreated and silicon-depleted French bean cells. Physiol. Mol. Plant Pathol. 29, 27-39. 1987: Host or nonhost resistance. In: ARNTZEN, CH. J., and C. RYAN (Eds), Molecular strategies for crop protection, pp. 25-34. Alan Liss, New York.
HOCH, H. c., and R. C. STAPLES, 1987: Structural and chemical changes among the rust fungi during appressorium development. Annu. Rev. Phytopathol. 25, 231-247.
- -, - -, B. WHITEHEAD, J. COMEAU, and E. D. WOLF, 1987: Signaling for growth orientation� and cell differentiation by surface topography in Uromyces. Science 235, 1659-1662.�
HOLDEN, D. W., and R. ROHRINGER, 1985: Proteins in intercellular washing fluid from noninocu�lated and rust-affected leaves of wheat and barley. Plant Physiol. 78, 715-723.
HOPPE, H. H., B. HDMME, and R. HEITEFUSS, 1980: Elictor induced accumulation of phytoalexins in healthy and rust infected leaves of Phaseolus vulgaris. Phytopath. Z. 97, 85-88.
JOHNSON, T., 1934: A tropic response in germ tubes of urediospores of Puccinia graminis tritici. Phytopathology 24, 8Q-82.
KAPOORIA, R. G., and K. MENDGEN, 1985: Infection structures and their surface changes during differentiation in Uromyces fabae. Phytopath. Z. 113, 317-323. ,
KEEN, N. T., 1982: Specific recognition in gene for gene host parasite system. Advances in Plant Pathology 1, 35-82.
KIM, W. K., and N. K. HOWES, 1987: Localization of gl¥.copeptides and race-variable polypeptides in urediosporelings and urediosporelings walls of Puccinia graminis cricici; affinity to concanavalin A, soybean agglutinin, and Lotus lectin. Can. J. Bot. 65, 1785-179l.
KITANI, K., and Y. INOUE, 1961: "Studies on the soybean rust and its control measure." Pan 1. Shekoku Agric. Exp. Stan., Zentsuji, Japan.
KNAUF, G., and K. MENDGEN, 1988: Secretion-systems and membrane-associated structures in rust fungi after high pressure freezing and freeze-fracturing. (submined.)
KOCH,� E., 1984: U ntersuchungen zur Biologie des Sojabohnenrostes (Phakopsora pachyrhizi Svd.) und zur Charakterisierung der Rostresistenz von Sojabohnenzuchdinien. Dissertation Uni"ersitat Gottingen.
KUNOH, H., 1987: Induced susceptibility and enhanced resistance at the cellular level in barley coleoptiles. In: NISHIMURA, 5., C. P. VANCE, and N. DOKE (Eds), Molecular determinants of plant diseases, pp. 59-73. Springer Verlag, Berlin.
LEWIS, B. c., and J. R. DAY, 1972: Behaviour of urediospore germ-tubes of Puccinia graminis tritici in relation to the fine structure of wheat leaf surfaces. Trans. Br. mycol. So';;. 58, 139-145.
LITTLEFIELD, 1.. J., and C. E. BRACKER, 1972: Ultrastructural specialization of the host-pathogen interface in rust infected flax. Protoplasm a 74, 271-305.
--, and M. C. HEATH, 1979: "Ultrastructure of Rust Fungi." Academic Press, New York.

271 The Differentiation of Infection Structures
LUKE, H. H., R. D. BARNETI, and P. L. PfAHLER, 1987: Xenoparasite - Nonhost Reactions in Puccinia-Gramineae Pathosystems. Phytopathology 77, 1488-1491.
MACKO, V., J. A. A. RENWICK, and J. F. RlSSLER, 1978: Acrolein induces differentiation of infection structures in the wheat stem rust fungus. Science 199,442---443.
MAHESHWARI, R., and A. C. HILDEllRANDT, 1967: Directional growth of the urediospore germ tubes and stomatal penetration. Nature (London) 214,1145-1156.
MAYAMA, S., 1983: The role of avenalumin in the resistance of oats to crown rust. Memoirs of Faculty of Agriculture, Kagawa University 42, 1--64.
MENDGEN, K., 1973: Feinbau der [nfektionsstrukturen von Ummyces phaseoli. Phytopath. Z. 78, 109-120. 1981: Nutrient uptake in rust fungi. Phytopathology 71, 983-989. 1982: Differential recognition of the outer and inner walls of epidermal cells by a rust fungus. Naturwissenschaften 69, 502-503. M. LANGE, and K. BRETscHNElDER, 1985: Quantitative estimation of the surface carbohy�drates on the infection structures of ruSt fungi with enzymes and lectins. Arch. Microbiol. 140,� 307-311.� and P. NASS, 1988: The activity of powdery mildew haustoria after feeding the host cells with� different sugars, as measured witli"a potentiometric cyanine dye. Planta 174, 283-288.�
MOERSCHllACHER, B., K. H. KOGEL, U. NOLL, and H. J. REISENER, 1986: An elicitor of the hypersensitive lignification response in wheat leaves isolated from the rust fungus Puccinia graminis f. sp.· tritici. l. Partial purification and characterisation. Z. Naturforsch. 41 c, 830-838.
MULLER, M., and H. MOOR, 1984: Cryofixation of thick specimens by high pressure freezing. In: Science of Biological Specimen Preparation, SEM [nc., AMF O'Hare (Chicago), U.5.A., 131-138.
NIKS, R. E., 1983: Haustorium formation by Puccinia hordei in leaves of hypersensitive, partially resistant, and nonhost plant genotypes. Phytopathology 73, 64-66.
ONOE,� T., T. TANl, S. MlNAGAWA, and H. SAGAWA, 1987: Ultrastructural changes of stomata in relation to specificity of rust fungi. In: NISHIMURA, S., C. P. VANCE, and N. DOKE (Eds), Molecular determinants of plant diseases, pp. 29-45. Springer Verlag Berlin.
PARADIES, J., B. HUMME, H. H. HOPPE, R. HEITEFUSS, and E. F. ELSTNER, 1979: Induction of ethylene formation in bean (Phaseolus vulgaris) hypocotyl segments by preparations isolated from germ tube cell walls of Uromyces phaseoli. Planta 146, 193-197. unci E. F. ELSTNER, 1980: Wirt-Parasit Bezichungen: Untersuchungen zur [nduktion der Athylenbildung in Hoheren Pflanzen und zur Rolle des Athylens bei der Auspriigung von Krankheitssymptomen und der EinJeitung von Abwehrreaktionen. Ber. Dtsch. Bot. Ges. 93, 635-657.
PAVGI,� M. 5., and J. G. DlcKsON, 1961: [nfluence of environmental factors on development of infection structures of Puccinia sorghi. Phytopathology 51,224-226.
ROHRJNGER, R., and J. W. MARTENS, 1987: [nfection-related proteins in intercellular washing fluids from stem rust-affected leaves: race-associated protein differences revealed by PAGE and Con A blotting. Physiol. Mol. Plant Pathol. 31,375-386.
RWENDEIRE, ABEL j., and M. J. BARNEs, 1988: The initial response of barley to challenge by leaf rust (Puccinia hordel Otth): some factors in RNA metabolism related to resistance. Physiol. Mol. Plant Pathol. 32, 115-126.
SPENCER-PH III PS, P. T. N., and J. L. GAY, 1981: Domains of ATPase in plasma membranes and transport through infected plant cells. New Phytologist 89, 393-400.
STAPLES, R. c., and H. C. HOCH, 1982: A possible role for microtubules in the induction of nuclear division in bean rust uredospore germlings. Exp. Mycology 6, 293-302. H. j. GRAMllOW, and H. C. HOCH, 1983: Potassium ion induces rust fungi to develop infection structures. Exp. Mycology 7, 40---46. and V. MACKO, 1984: Germination of urediospores and differentiation of infection structures. In: BUSHNELL, W. R., and A. P. ROELFS (Eds), The Cereal Rusts, pp. 255-289. Academic Press, Orlando. and H. C. HOCH, 1987: Infection structures - form and function. Exp. Mycology 11, 163-169.

272 MENDGEN, SCHNEIDER, STERK and FINK, The Differentiation of Infection Structures
STUMPF, M. A., and M. C. HEATH, 1985: Cytological studies of the interactions between the cowpea rust fungus and silicon-depleted French bean plants. Physiol. Plant Pathol. 27, 369-385.
TANI, T., and H. YAMAMOTO, 1978: Nuclei acid and protein synthesis in association with the resistance of oat leaves to crown rust. Physiol. Plant Pathol. 12, 113-121.
VANDERPLANK, J. E., 1984: Disease Resistance in Plants. Academic Press, Orlando. WELTER, K., M. MULLER, and K. MENDGEN, 1988: The hyphae of Uromyces appendiculatus within
the leaf tissue after high pressure freezing and freeze substitution. Protoplasma (in press). WYNN, W. K., 1976: Appressorium formation over stomata by the bean rust fungus: Response to a
surface contact stimulus. Phytopathology 66, 136-146. YAMAMOTO, H., H. HOKIN, T. TANI, and G. KADOTA, 1977: Phenylalanine ammonia-lyase in
relation to the crown rust resistance of oat leaves. Phytopath. Z. 90, 203-211. - -, and T. TANI, 1986: Possible involvement of lipoxygenase in the mechanism of resistance of
oats to Puccinia coronata avenae. J. Phytopathology 116, 329-337. YOUNG, D. H., H. KOHLE, and H. KRAUSS, 1982: Effect of chitosan on membrane permeability
of suspension-cultured Glycine max and Phaseolus vulgaris cells. Plant Physiology 70, 1449-1454.
ZIEGLER, E., and R. PONTZEN, 1982: Specific inhibition of glucan-elicited glyceollin accumulation in soy beans by an extracellular mannan-glycoprotein of Phytophthora megasperma f. sp. glycinea. Physiol. Plant Pathol. 20, 321-331.
..�





























