Embed Size (px)
Citation preview

ORIGINAL ARTICLE
The Cytomegalovirus Promoter-Driven Short Hairpin RNAConstructs Mediate Effective RNA Interference in ZebrafishIn Vivo
Jianguo Su & Zuoyan Zhu & Yaping Wang & Feng Xiong &
Jun Zou
Received: 30 July 2007 /Accepted: 28 September 2007 /Published online: 24 January 2008# Springer Science + Business Media, LLC 2007
Abstract The ability to utilize the RNA interference(RNAi) machinery for silencing target-gene expressionhas created a lot of excitement in the research community.In the present study, we used a cytomegalovirus (CMV)promoter-driven DNA template approach to induce shorthairpin RNA (shRNA) triggered RNAi to block exogenousEnhanced Green Fluorescent Protein (EGFP) and endoge-nous No Tail (NTL) gene expressions. We constructed threeplasmids, pCMV-EGFP-CMV-shGFP-SV40, pCMV-EGFP-CMV-shNTL-SV40, and pCMV-EGFP-CMV-shScrambled-SV40, each containing a CMV promoter driving an EGFPreporter cDNA and DNA coding for one shRNA under thecontrol of another CMV promoter. The three shRNA-generating plasmids and pCMV-EGFP control plasmidwere introduced into zebrafish embryos by microinjection.Samples were collected at 48 h after injection. Results wereevaluated by phenotype observation and real-time fluores-cent quantitative reverse-transcription polymerase chainreaction (Q-PCR). The shGFP-generating plasmid signifi-cantly inhibited the EGFP expression viewed under
fluorescent microscope and reduced by 70.05±1.26% ofexogenous EGFP gene mRNA levels compared withcontrols by Q-PCR. The shRNA targeting endogenousNTL gene resulted in obvious NTL phenotype of 30±4%and decreased the level of their corresponding mRNAs upto 54.52±2.05% compared with nontargeting controlshRNA. These data proved the feasibility of the CMVpromoter-driven shRNA expression technique to be used toinhibit exogenous and endogenous gene expressions inzebrafish in vivo.
Keywords CMV promoter . EGFP. No tail . RNAi .
shRNA . Zebrafish
Introduction
RNA interference (RNAi), a conserved antiviral immunity ofplants and animals (Ding et al. 2004), has been developed intoan effective method of sequence-specific gene knockdownfor analyzing gene functions in plants, invertebrates, andmammalian cells (Napoli et al. 1990; Fire et al. 1998;Elbashir et al. 2001). RNAi also holds great promise as apowerful therapeutic tool (Banan and Puri 2004; Kondragantiet al. 2006). The conserved RNAi pathway involves theprocessing of double-stranded RNA (dsRNA) duplexes into21-23 nucleotide (nt) molecules known as small interferingRNAs (siRNA) to initiate gene knockdown (Hannon 2002).The long dsRNA in lower eukaryotes, especially in themodel organism Caenorhabditis elegans, has been used todetermine gene functions (Ashrafi et al. 2003). However,long dsRNA in mammalian systems induces an antiviraldefence mechanism initiated by interferon (IFN), leading tononspecific translational shutdown and apoptosis (Gil andEsteban 2000).
Mar Biotechnol (2008) 10:262–269DOI 10.1007/s10126-007-9059-4
J. Su : Z. Zhu (*) :Y. Wang : F. XiongState Key Laboratory of Freshwater Ecology and Biotechnology,Institute of Hydrobiology, Chinese Academy of Sciences,Wuhan 430072, Chinae-mail: [email protected]
J. SuDepartment of Aquaculture,College of Animal Science and Technology,Northwest A&F University,Yangling 712100, China
J. ZouSchool of Biological Sciences, University of Aberdeen,Aberdeen AB24 2TZ, United Kingdom

Direct transfection of either chemically synthesized or invitro transcribed siRNAs of approximately 21 nt in lengthdoes not activate the IFN response but can induce reliableand efficient transient knockdown of target genes inmammalian cells (Tuschl 2002; Dykxhoorn et al. 2003).Compared with siRNA, the DNA-based vectors for expres-sion of short hairpin RNA (shRNA) offer additionaladvantages in silencing longevity, cost, and deliveryoptions (McIntyre and Fanning 2006). As a consequence,the development of shRNA molecules that are processedwithin the cell to produce active siRNA molecules hasprogressed rapidly (Brummelkamp et al. 2002). Such DNAexpression constructs have achieved highly gene knock-down efficiency without induction of the IFN response.
Expressed shRNA is transcribed in cells from a DNAtemplate as a single-stranded RNA molecule. Complemen-tary regions spaced by a small loop cause the transcript tofold back on itself, forming a short hairpin in a manneranalogous to natural micro-RNA. Recognizing and pro-cessing by the RNAi machinery convert the shRNA into thecorresponding siRNA. shRNA expression vectors havebeen engineered by using both viral and plasmid systems.These vectors often utilize promoters from a small class ofthe RNA polymerase III-type (pol. III) promoters(Schramm and Hernandez 2002) to drive the expressionof shRNA. Promoters of this type are preferred becausethey naturally direct the synthesis of small, highly abundantnoncoding RNA transcripts, with defined terminationsequences consisting of 4 to 5 thymidines (Ts) and haveno requirement for downstream termination elements(Geiduschek and Kassavetis 2001).
For siRNA expression, the RNA polymerase II-type(pol. II) CMV promoter (human cytomegalovirus immedi-ate-early promoter) has several advantages over pol. IIIpromoters, such as U6 or H1. First, pol. II will toleratestrings of 4 or more Ts within the siRNA sequence, unlikepol. III, which will terminate transcription after incorpora-tion of a stretch of Ts. Second, the CMV promoter does notinterfere with other transcription events (such as expressionof the antibiotic resistance gene), making it easier toperform long-term gene silencing studies (Xia et al. 2002;Jonathan et al. 2007).
The zebrafish is a simple vertebrate that has manyattributes that make it ideal for the study of the immunesystem (Yoder et al. 2002). Long dsRNA causes nonspe-cific regression (Zhao et al. 2001), U6 promoter-drivenshRNA is not high efficient (Xie et al. 2005), and siRNAlast short time in vivo, which hinder the application ofRNAi technique in fish.
In the current study, we explored the CMV promoter todrive high-level expression of shRNA molecules in zebra-fish embryos. We constructed the plasmid vectors targetingthe exogenous expressing enhanced green fluorescent
protein (EGFP) gene and the endogenous no tail (NTL)gene. The inhibition assays used phenotype observation andreal-time fluorescent quantitative reverse-transcription po-lymerase chain reaction (Q-PCR).
Materials and Methods
Introducing CMV Promoter Sequence All the oligonucleo-tides in this study were synthesized on an appliedbiosystem model 380B automated DNA synthesizer byShanghai Sangon Biological Engineering Technology &Services Co., Ltd. (Table 1; Figure 1a–c). All the followinginserts, including the CMV promoter and SV40 transcrip-tional termination sequence produced by PCRs andshRNAs chemically synthesized, were sequenced on aModel 3730 DNA Sequence System (Shanghai InvitrogenCo., Ltd., China) to guarantee the correct sequences.
The CMV promoter was obtained and introduced restric-tion sites both upstream and downstream of the sequence byPCR, using the pCMV-EGFP plasmid (Clontech, USA) astemplate. A PCR sense primer was SCF93a with Xho I site,and an antisense primer was SCR94 with Hind III, Apa I,and Sal I sites (Table 1). The PCR product of 630 bp waspurified with gel extraction kit (Axygen), ligated intopMD18-T vector (TAKARA), transformed into TOP10competent cells. Three positive colonies were selected andsequenced for verification of the insert without mutation.The plasmid with correct insert was extracted and digestedwith Xho I and Hind III; meanwhile, the pCMV-EGFPplasmid was digested with the same enzymes. The targetfragments were purified, ligated with T4 ligase, and namedas pCMV-EGFP-CMV.
Inserting SV40 Transcriptional Termination Sequence ThePCR to get the SV40 transcriptional termination site was setup with pCMV-EGFP plasmid as template. The forward
Table 1 Oligonucleotides used in this study
Name Sequence
SCF93a CTCGAGTAGTTATTAATAGTAATCAATTACGSCR94a AAGCTTATGGGCCCGTCGACCGATCTGACGGTTC
ACTASVF81a GGGCCCAGCGGCCGCGACTCTAGATCATSVR95a AAGCTTGCAGTGAAAAAAATGCTTTATTTGTGSBAF86 GATGATGAAATTGCCGCACTGSBAR87 ACCAACCATGACACCCTGATGTSGF114 CAAGCAGAAGAACGGCATCASGR115 AGGTAGTGGTTGTCGGGCASNF122 CAGCACTGACAACCAGCAATCSNR123 GAACCCGAGGAGTGAACAGG
Mar Biotechnol (2008) 10:262–269 263263

primer was SVF81a with Apa I site, and the reverse primerwas SVR95a with Hind III site (Table 1). The amplicon wasperformed and confirmed as above. The plasmid wasdigested with Apa I and Hind III, and cloned into thepCMV-EGFP-CMV plasmid. The target fragments werepurified, ligated, and named as pCMV-EGFP-CMV-SV40.
shRNA Design and Expression Vectors Construct TheEGFP and NTL siRNA sequences used for constructionof shRNA vectors had previously been reported to beeffective in silencing gene expression (Tiscornia et al. 2003;Liu et al. 2005). shRNA oligonucleotides were designed asa synthetic duplex with overhanging ends identical to thosecreated by restriction enzyme digestion (upper oligo: Sal Iat the 5′ and Apa I at the 3′). The shRNA coding regioncontained a sense strand of 20 (EGFP), 19 (NTL), or 20(scrambled) nucleotide sequences followed by a short spacer(TTCAAGAGA), and the reverse complement sequence ofthe sense strand (Fig. 1a-c). Forward and reverse oligos forEGFP, Scrambled and NTL were SGFP97, SGFP98,SGFP97a, SGFP98a, SNTL120, and SNTL121, respectively(Fig. 1a-c). The nonspecific control was shuttling DNAsequence of siGFP and analyzed by BLASTN. Each oligowas suspended in water (50 mM) and 5 μl from each(SGFP97 and SGFP98 for EGFP) was put together, the mixwas heated to 95°C for 5 min, and then slowly equilibratedto room temperature. The duplex was inserted to the Sal Iand Apa I sites of pCMV-EGFP-CMV-SV40. Similarly, thescrambled and NTL shRNA vectors were produced,
respectively. The shRNA vectors for EGFP, scrambled, andNTL were confirmed by sequencing and called after pCMV-EGFP-CMV-shGFP-SV40 , pCMV-EGFP-CMV-shScrambled-SV40, and pCMV-EGFP-CMV-shNTL-SV40(Fig. 1d), respectively.
Microinjection of Zebrafish Embryos After artificial insem-ination, approximately 1 nl of shRNA expression vectorsolution (100 pg/nl) was microinjected into the embryo atthe 1- to 2-cell stage under a dissecting microscope using apulled microcapillary pipette as previously report (Wu et al.2006; Liu et al. 2006). We injected at least 450 embryosand cultured in three dishes for every group. Totally weused approximately 3,000 embryos. The injected anduninjected embryos were subsequently incubated in sterile0.3 × Danieau’s solution (19.3 nM NaCl, 0.23 mM KCl,0.13 mM MgSO4·7H2O, 0.2 mM Ca(NO3)2, 1.67 mMHEPES pH 7.2) at 28°C. We changed the solution andremoved dead embryos twice per day.
Phenotypic Analyses Phenotypes were evaluated at 48 hpf(hours post fertilization). Images of zebrafish embryos wererecorded by using an Olympus SZX12 fluorescent micro-scope and a digital camera (Liu et al. 2007). Featuresanalyzed for NTL were: disrupted notochord, abnormalsomites, and reduced tail (Halpern et al. 1993). EGFPexpression was monitored by using fluoresce microscopy.For phenotype count and fluorescent observation, we usedall the embryos.
Fig. 1 The constructs ofshRNA expression vectors. a.Illustration of shRNA expres-sion plasmid, containing CMVpromoter, SV40 transcriptiontermination sequence, shRNAtemplate, restriction sites, andthe skeleton component of orig-inal pCMV-EGFP plasmid. b–d.The transcription template se-quence of shGFP, shScrambled,shNTL unit, respectively
264 Mar Biotechnol (2008) 10:262–269

Q-PCR Assays Target mRNA levels in embryos werequantitatively analyzed by using Q-PCR. Zebrafish embryosinjected with pCMV-EGFP-CMV-shGFP-SV40, pCMV-EGFP-CMV-shNTL-SV40 , pCMV-EGFP-CMV-shScrambled-SV40, and pCMV-EGFP and uninjected con-trols were collected at 48 hpf, and total RNA was extractedfrom pools of 30 to 40 embryos per sample with triplicaterepeat using Trizol (Invitrogen). The RNA samples werefurther treated with RNase-free DNase I (Roche) to removecontaminated genomic DNA, followed by phenol/chloroformextraction and ethanol precipitation. A total of 1 μg RNAfrom each extraction was reverse transcribed with SuperscriptIII reverse transcriptase (Invitrogen) and random primers in a20-μl reaction volume according to the manufacturer’sinstructions. cDNA samples were diluted 1:5 before use inQ-PCR assays.
Q-PCRs were performed in an ABI Prism 7000Sequence Detection System (Applied Biosystems) usingthe following thermal cycling profile: 95°C 3 min, followedby 40 cycles of amplification (95°C 15 s, 58°C 15 s, 72°C45 s), followed by dissociation curve analysis to validatethe amplification of a single product. Each reactionconsisted of 4 μl of cDNA sample, 4 μl of nuclease-freewater, 10 μl of SYBR Green PCR master mix (Toyobo),and 1 μl of each primer set (2.5 μM). All reactions wereperformed in triplicate. All primers were designed toconform to a universal cycling program by the PrimerExpress 2.0 software (Applied Biosystems). The house-keeping gene β-actin was used as an internal standard tonormalize differences in template amounts. Primers for β-actin were upstream primer SBAF86 and downstreamprimer SBAR87 (Table 1). The PCR product size was135 bp. The forward primer sequence for EGFP wasSGF114 and the reverse primer was SGR115 (Table 1).The PCR product of the EGFP gene was 138 bp. NTLprimer sequences were forward SNF122 and reverseSNR123 (Table 1). The NTL amplicon was 126 bp.
Relative expression was calculated by using a modifiedcomparative cycle threshold (CT) method, in which CT wasdefined as the cycle numbers at which fluorescence reached aset threshold value. The differences in the CT value of thetarget genes from the corresponding internal control β-actingene, ΔCT (CTgene - CTactin), were calculated. The changesin ΔCT of the experiment group to the control group,ΔΔCT (ΔCTsample - ΔCTcontrol), were computed. Therelative expression level of the experiment group to thecontrol group was described using the equation 2-ΔΔCT, andthe value standed for a 1/n-fold difference relative to thecontrol. The expression of experiment group relative tothe control group was multiplied by 100% to simplify thepresentation of the data.
Statistical Analysis The data obtained from Q-PCR analysiswere subjected to an unpaired, two-tailed, Student’s t test.Error bars indicated standard errors. P<0.05 was consid-ered statistically significant, and P<0.01 was a verysignificant difference.
Results
Construction of shRNA Expression Vectors Using thepCMV-EGFP as template, PCR based cloning strategieswere performed to generate another set of CMV promoter,SV40 transcription termination sequence, and suitablerestriction sites. Insert the DNA template for expressingshRNA between CMV promoter and SV40 terminator toproduce shRNA expression vectors targeting EGFP andNTL. Similarly, the nonspecific control shRNA vector(pCMV-EGFP-CMV-shScrambled-SV40) also was generat-ed (Fig. 1d). All final shRNA expression constructsconsisted of a CMV promoter, shRNA sense sequence,loop sequence, shRNA antisense sequence, SV40 termina-tion sequence, and the pCMV-EGFP vector skeleton. Thetranscripted shRNA composed of two complementary 19-or 20-nucleotide sequence motifs in an inverted orientation,separated by a 9-bp spacer to form a hairpin dsRNA.
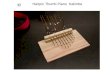
Activity of CMV Promoter Measured by Knockdown ofExogenous EGFP Expression To analyze the function ofCMV promoter, the level of EGFP expression in zebrafishembryos injected pCMV-EGFP-CMV-shGFP-SV40 wasdirectly compared with that injected pCMV-EGFP-CMV-shScrambled-SV40 and pCMV-GFP. Knockdown of EGFPin embryos was visualized by fluorescence microscopy(Fig. 2). Results showed that the fluorescence in theembryos injected pCMV-EGFP-CMV-shGFP-SV40exhibited reduction compared with that injected pCMV-EGFP-CMV-shScrambled-SV40 and pCMV-GFP.
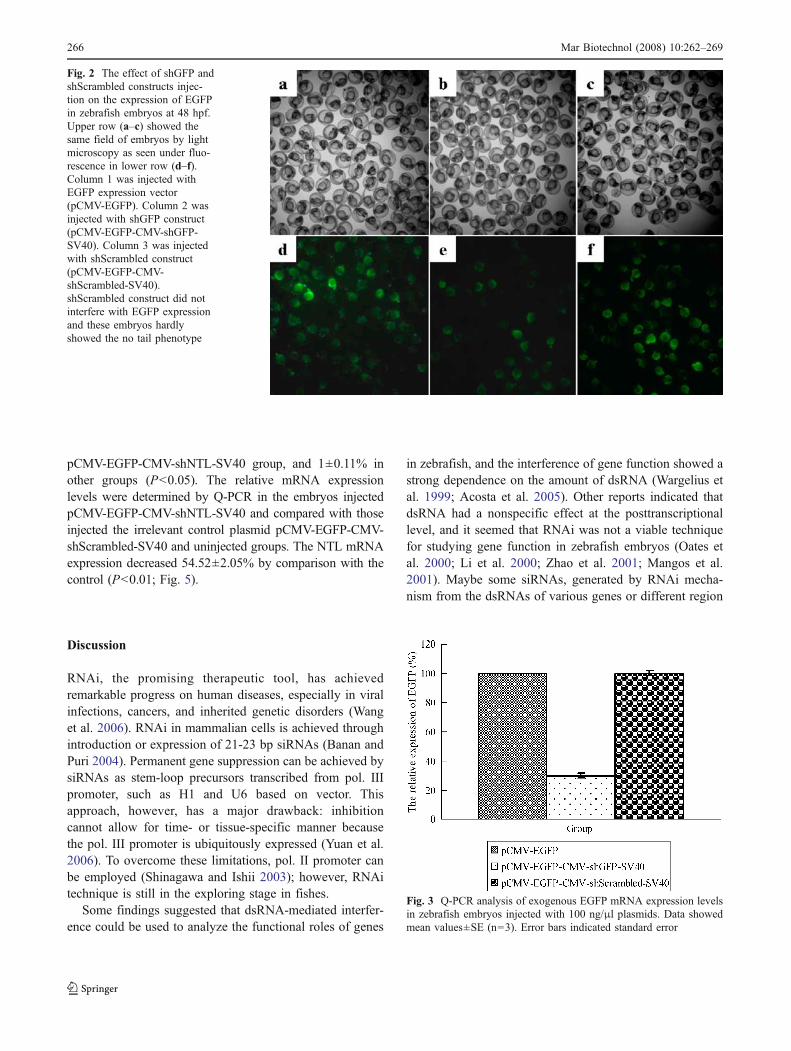
Q-PCR was used to quantify the EGFP mRNA transcript.The relative EGFP mRNA expression level reduced 70.05±1.76% (P<0.01) in the experiment group. The mRNAtranscripts were not significant differences between pCMV-EGFP-CMV-shScrambled-SV40 group and pCMV-GFPgroup (P>0.05; Fig. 3).
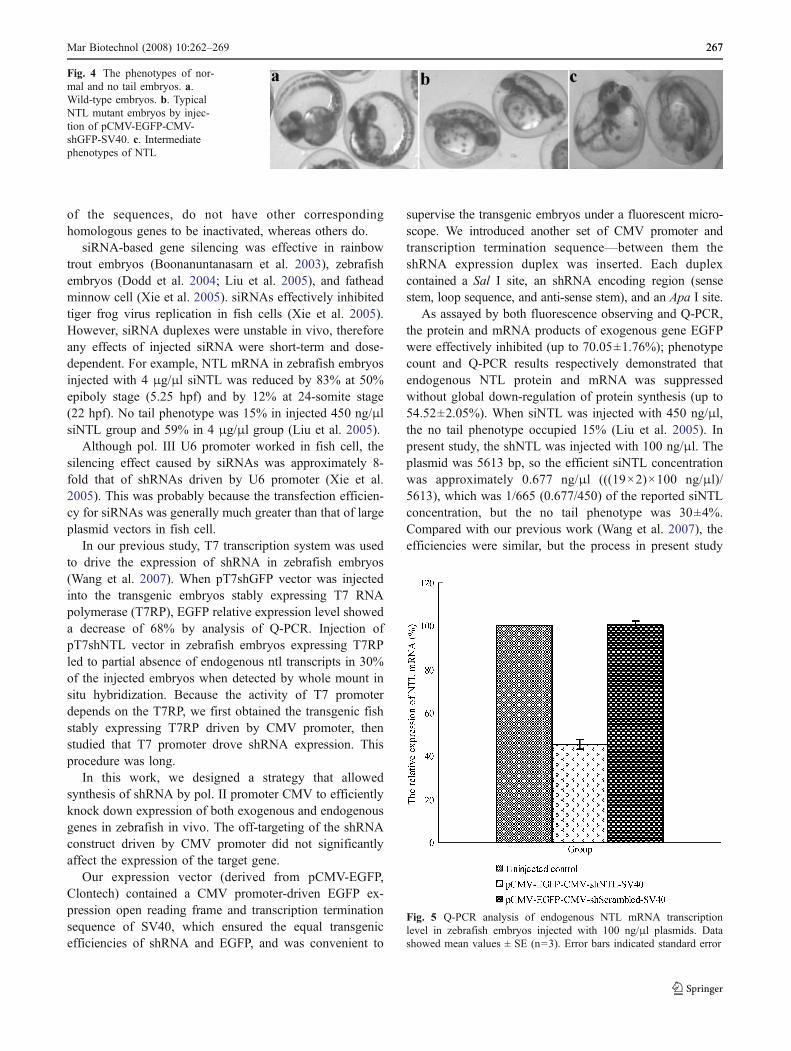
Function of CMV Promoter Measured by Knockdown ofEndogenous NTL Expression To further validate the functionof the CMV promoter for RNAi in fish, shRNA expressionvector targeting the endogenous NTL gene was produced. Theembryos of no tail phenotype (Fig. 4) were counted under adissection microscope, and occupied 30±4% at 48 hpf in
Mar Biotechnol (2008) 10:262–269 265265
pCMV-EGFP-CMV-shNTL-SV40 group, and 1±0.11% inother groups (P<0.05). The relative mRNA expressionlevels were determined by Q-PCR in the embryos injectedpCMV-EGFP-CMV-shNTL-SV40 and compared with thoseinjected the irrelevant control plasmid pCMV-EGFP-CMV-shScrambled-SV40 and uninjected groups. The NTL mRNAexpression decreased 54.52±2.05% by comparison with thecontrol (P<0.01; Fig. 5).
Discussion
RNAi, the promising therapeutic tool, has achievedremarkable progress on human diseases, especially in viralinfections, cancers, and inherited genetic disorders (Wanget al. 2006). RNAi in mammalian cells is achieved throughintroduction or expression of 21-23 bp siRNAs (Banan andPuri 2004). Permanent gene suppression can be achieved bysiRNAs as stem-loop precursors transcribed from pol. IIIpromoter, such as H1 and U6 based on vector. Thisapproach, however, has a major drawback: inhibitioncannot allow for time- or tissue-specific manner becausethe pol. III promoter is ubiquitously expressed (Yuan et al.2006). To overcome these limitations, pol. II promoter canbe employed (Shinagawa and Ishii 2003); however, RNAitechnique is still in the exploring stage in fishes.
Some findings suggested that dsRNA-mediated interfer-ence could be used to analyze the functional roles of genes
in zebrafish, and the interference of gene function showed astrong dependence on the amount of dsRNA (Wargelius etal. 1999; Acosta et al. 2005). Other reports indicated thatdsRNA had a nonspecific effect at the posttranscriptionallevel, and it seemed that RNAi was not a viable techniquefor studying gene function in zebrafish embryos (Oates etal. 2000; Li et al. 2000; Zhao et al. 2001; Mangos et al.2001). Maybe some siRNAs, generated by RNAi mecha-nism from the dsRNAs of various genes or different region
Fig. 3 Q-PCR analysis of exogenous EGFP mRNA expression levelsin zebrafish embryos injected with 100 ng/μl plasmids. Data showedmean values±SE (n=3). Error bars indicated standard error
Fig. 2 The effect of shGFP andshScrambled constructs injec-tion on the expression of EGFPin zebrafish embryos at 48 hpf.Upper row (a–c) showed thesame field of embryos by lightmicroscopy as seen under fluo-rescence in lower row (d–f).Column 1 was injected withEGFP expression vector(pCMV-EGFP). Column 2 wasinjected with shGFP construct(pCMV-EGFP-CMV-shGFP-SV40). Column 3 was injectedwith shScrambled construct(pCMV-EGFP-CMV-shScrambled-SV40).shScrambled construct did notinterfere with EGFP expressionand these embryos hardlyshowed the no tail phenotype
266 Mar Biotechnol (2008) 10:262–269
of the sequences, do not have other correspondinghomologous genes to be inactivated, whereas others do.
siRNA-based gene silencing was effective in rainbowtrout embryos (Boonanuntanasarn et al. 2003), zebrafishembryos (Dodd et al. 2004; Liu et al. 2005), and fatheadminnow cell (Xie et al. 2005). siRNAs effectively inhibitedtiger frog virus replication in fish cells (Xie et al. 2005).However, siRNA duplexes were unstable in vivo, thereforeany effects of injected siRNA were short-term and dose-dependent. For example, NTL mRNA in zebrafish embryosinjected with 4 μg/μl siNTL was reduced by 83% at 50%epiboly stage (5.25 hpf) and by 12% at 24-somite stage(22 hpf). No tail phenotype was 15% in injected 450 ng/μlsiNTL group and 59% in 4 μg/μl group (Liu et al. 2005).
Although pol. III U6 promoter worked in fish cell, thesilencing effect caused by siRNAs was approximately 8-fold that of shRNAs driven by U6 promoter (Xie et al.2005). This was probably because the transfection efficien-cy for siRNAs was generally much greater than that of largeplasmid vectors in fish cell.
In our previous study, T7 transcription system was usedto drive the expression of shRNA in zebrafish embryos(Wang et al. 2007). When pT7shGFP vector was injectedinto the transgenic embryos stably expressing T7 RNApolymerase (T7RP), EGFP relative expression level showeda decrease of 68% by analysis of Q-PCR. Injection ofpT7shNTL vector in zebrafish embryos expressing T7RPled to partial absence of endogenous ntl transcripts in 30%of the injected embryos when detected by whole mount insitu hybridization. Because the activity of T7 promoterdepends on the T7RP, we first obtained the transgenic fishstably expressing T7RP driven by CMV promoter, thenstudied that T7 promoter drove shRNA expression. Thisprocedure was long.
In this work, we designed a strategy that allowedsynthesis of shRNA by pol. II promoter CMV to efficientlyknock down expression of both exogenous and endogenousgenes in zebrafish in vivo. The off-targeting of the shRNAconstruct driven by CMV promoter did not significantlyaffect the expression of the target gene.
Our expression vector (derived from pCMV-EGFP,Clontech) contained a CMV promoter-driven EGFP ex-pression open reading frame and transcription terminationsequence of SV40, which ensured the equal transgenicefficiencies of shRNA and EGFP, and was convenient to
supervise the transgenic embryos under a fluorescent micro-scope. We introduced another set of CMV promoter andtranscription termination sequence—between them theshRNA expression duplex was inserted. Each duplexcontained a Sal I site, an shRNA encoding region (sensestem, loop sequence, and anti-sense stem), and an Apa I site.
As assayed by both fluorescence observing and Q-PCR,the protein and mRNA products of exogenous gene EGFPwere effectively inhibited (up to 70.05±1.76%); phenotypecount and Q-PCR results respectively demonstrated thatendogenous NTL protein and mRNA was suppressedwithout global down-regulation of protein synthesis (up to54.52±2.05%). When siNTL was injected with 450 ng/μl,the no tail phenotype occupied 15% (Liu et al. 2005). Inpresent study, the shNTL was injected with 100 ng/μl. Theplasmid was 5613 bp, so the efficient siNTL concentrationwas approximately 0.677 ng/μl (((19×2)×100 ng/μl)/5613), which was 1/665 (0.677/450) of the reported siNTLconcentration, but the no tail phenotype was 30±4%.Compared with our previous work (Wang et al. 2007), theefficiencies were similar, but the process in present study
Fig. 5 Q-PCR analysis of endogenous NTL mRNA transcriptionlevel in zebrafish embryos injected with 100 ng/μl plasmids. Datashowed mean values ± SE (n=3). Error bars indicated standard error
Fig. 4 The phenotypes of nor-mal and no tail embryos. a.Wild-type embryos. b. TypicalNTL mutant embryos by injec-tion of pCMV-EGFP-CMV-shGFP-SV40. c. Intermediatephenotypes of NTL
Mar Biotechnol (2008) 10:262–269 267267

was simple. These results appeared that CMV-drivenshRNA vector was preferable to siRNA and T7 transcrip-tion system in inhibition assay in fish in vivo.
Normally the CMV promoter is long lasting, because theconstruction can integrate the genome, continuously tran-script RNA, and inherit next generation. Cecropin B, aninsect antimicrobial peptide, was expressed in medaka,driven by a CMV promoter. The F2 transgenic fish hadacquired elevated resistance to bacterial infection (Sarmasiket al. 2002). In present study, we combined the activity ofCMV promoter with shRNA sequence. Actually, we alsotested the expressions at day 5 after injection. The inhibitionefficiency reduced a little. To compare with other resultsmentioned, we just showed the results at 48 h after injection.
Considered together, the CMV promoter, the pol. IIpromoter, could be used in the construction of plasmidbased shRNA expression vector. The CMV promoter-driven shRNA constructs could lead to effectively andspecifically inhibition of exogenous and endogenous geneexpressions in zebrafish in vivo. These vectors efficientlyinduced RNAi in zebrafish through production of shRNAmolecules targeted at the exogenous expressing reportergene EGFP, and the endogenous zebrafish NTL gene. Thatthe use of this promoter sequence and the shRNA vectorcloning strategy described will be advantageous in RNAifunctional genomic experiments and an important step inthe development of novel RNAi technology for immunetherapeutics in the transgenic delivery of shRNA moleculesin fishes. This method may provide a novel perspective forthe application of RNAi technology in suppressing geneexpression in fish.
Acknowledgements The authors thank Miss Yuejiao Lu for criti-cally reading the manuscript. The technical assistance provided byMing Li, Na Wang, Jun Dai, Shangping Chen, and other laboratorymembers was greatly appreciated. This work was supported by(30428024 and 30540084) from National Natural Science Foundationof China, (KSCX2-YW-N-021) from Chinese Academy of Sciences,(08080262, 08080245, and 01140508) from Northwest A & FUniversity in China, (20070410298) from China Postdoctoral ScienceFoundation and (2007FB09) from Institute of Hydrobiology, CAS.
References
Acosta J, Carpio Y, Borroto I, Gonzalez O, Estrada MP (2005)Myostatin gene silenced by RNAi show a zebrafish giantphenotype. J Biotechnol 119:324-331
Ashrafi K, Chang FY, Watts JL, Fraser AG, Kamath RS, Ahringer J,Ruvkun G (2003) Genome-wide RNAi analysis of Caenorhabdi-tis elegans fat regulatory genes. Nature 421:268-272
Banan M, Puri N (2004) The ins and outs of RNAi in mammaliancells. Curr Pharm Biotechnol 5:441-450
Boonanuntanasarn S, Yoshizaki G, Takeuchi T (2003) Specific genesilencing using small interfering RNAs in fish embryos. BiochemBiophys Res Commun 310:1089-1095
Brummelkamp TR, Bernards R, Agami R (2002) A system for stableexpression of short interfering RNAs in mammalian cells.Science 296:550-553
Ding S, Li H, Lu R, Li F, Li W (2004) RNA silencing: a conservedantiviral immunity of plants and animals. Virus Res 102:109-115
Dodd A, Chambers SP, Love DR (2004) Short interfering RNA-mediated gene targeting in the zebrafish. FEBS Lett 561:89-93
Dykxhoorn DM, Novina CD, Sharp PA (2003) Killing the messenger:short RNAs that silence gene expression. Nat Rev Mol Cell Biol4:457-467
Elbashir SM, Harborth J, Lendeckel W, Yalcin A, Weber K, Tuschl T(2001) Duplexes of 21-nucleotide RNAs mediate RNA interfer-ence in cultured mammalian cells. Nature 411:494-498
Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC(1998) Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391:806-811
Geiduschek EP, Kassavetis GA (2001) The RNA polymerase IIITranscription Apparatus. J Mol Biol 310:1-26
Gil J, Esteban M (2000) Induction of apoptosis by the dsRNA-dependent protein kinase (PKR): mechanism of action. Apoptosis5:107-114
Halpern ME, Ho RK, Walker C, Kimmel CB (1993) Induction ofmuscle pioneers and floor plate is distinguished by the zebrafishno tail mutation. Cell 75:99-111
Hannon GJ (2002) RNA interference. Nature 418:244-251Jonathan EP, Foster JS, Kestler D, Alan S, Wall JS (2007) Inhibition
of Bence-Jones protein synthesis by RNA interference. JImmunol 178:S88-S89
Kondraganti S, Gondi CS, McCutcheon I, Dinh DH, Gujrati M,Rao JS, Olivero WC (2006) RNAi-mediated downregulationof urokinase plasminogen activator and its receptor in humanmeningioma cells inhibits tumor invasion and growth. Int JOncol 28:1353-1360
Liu J, Sun Y, Wang N, Wang Y, Zhu Z (2006) Upstream regulatoryregion of zebrafish lunatic fringe: isolation and promoteranalysis. Mar Biotechnol 8:357-365
Li YX, Farrell MJ, Liu R, Mohanty N, Kirby ML (2000) Double-stranded RNA injection produces null phenotypes in zebrafish.Dev Biol 217:394-405
Liu WY, Wang Y, Qin Y, Wang YP, Zhu ZY (2007) Site-directed geneintegration in transgenic zebrafish mediated by cre recombinaseusing a combination of mutant lox sites. Mar Biotechnol 9:418-420
Liu WY, Wang Y, Sun YH, Wang Y, Wang YP, Chen SP, Zhu ZY(2005) Efficient RNA interference in zebrafish embryos usingsiRNA synthesized with SP6 RNA polymerase. Dev GrowthDiffer 47:323-331
Mangos S, Vanderbeld B, Krawetz R, Sudol K, Kelly GM (2001) Ranbinding protein RanBP1 in zebrafish embryonic development.Mol Reprod Dev 59:235-248
McIntyre GJ, Fanning GC (2006) Design and cloning strategies forconstructing shRNA expression vectors. BMC Biotechnol 6:1-8
Napoli C, Lemieux C, Jorgensen R (1990) Introduction of a ChimericChalcone Synthase Gene into Petunia Results in Reversible Co-Suppression of Homologous Genes in trans. Plant Cell 2:279-289
Oates AC, Bruce AE, Ho RK (2000) Too much interference: injectionof double-stranded RNA has nonspecific effects in the zebrafishembryo. Dev Biol 224:20-28
Sarmasik A, Warr G, Chen T (2002) Production of transgenic medakawith increased resistance to bacterial pathogens. Mar Biotechnol4:310-322
Schramm L, Hernandez N (2002) Recruitment of RNA polymerase IIIto its target promoters. Genes Dev 16:2593-2620
Shinagawa T, Ishii S (2003) Generation of Ski-knockdown mice byexpressing a long double-strand RNA from an RNA polymeraseII promoter. Genes Dev 17:1340-1345
268 Mar Biotechnol (2008) 10:262–269

Tiscornia G, Singer O, Ikawa M, Verma IM (2003) A general methodfor gene knockdown in mice by using lentiviral vectorsexpressing small interfering RNA. Proc Natl Acad Sci USA100:1844-1848
Tuschl T (2002) Expanding small RNA interference. Nat Biotechnol20:446-448
Wang N, Sun Y, Liu J, Wu G, Su J, Wang Y, Zhu Z (2007) Knockdown of gfp and no tail expression in zebrafish embryo by invivo-transcribed short hairpin RNA with T7 plasmid system. JBiomed Sci 14:767-776
Wang L, Wu G, Yu L, Yuan J, Fang F, Zhai Z, Wang F, Wang H(2006) Inhibition of CD147 expression reduces tumor cellinvasion in human prostate cancer cell line via RNA interference.Cancer Biol Ther 5:608-614
Wargelius A, Ellingsen S, Fjose A (1999) Double-stranded RNAinduces specific developmental defects in zebrafish embryos.Biochem Biophys Res Commun 263:156-161
Wu Y, Zhang G, Xiong Q, Luo F, Cui C, Hu W, Yu Y, Su J, Xu A,Zhu Z (2006) Integration of double-fluorescence expressionvectors into zebrafish genome for the selection of site-directedknockout/knockin. Mar Biotechnol 8:304-311
Xia H, Mao Q, Paulson H, Davidson B (2002) siRNA-mediated genesilencing in vitro and in vivo. Nat Biotechnol 20:1006-1010
Xie J, Lu L, Deng M, Weng S, Zhu J, Wu Y, Gan L, Chan SM, He J(2005) Inhibition of reporter gene and Iridovirus-tiger frog virusin fish cell by RNA interference. Virology 338:43-52
Yoder JA, Nielsen ME, Amemiya CT, Litman GW (2002) Zebrafish asan immunological model system. Microbes Infect 4:1469-1478
Yuan J, Wang X, Zhang Y, Hu X, Deng X, Fei J, Li N (2006) shRNAtranscribed by RNA Pol II promoter induce RNA interference inmammalian cell. Mol Biol Rep 33:43-49
Zhao Z, Cao Y, Li M, Meng A (2001) Double-stranded RNAinjection produces nonspecific defects in zebrafish. Dev Biol229:215-223
Mar Biotechnol (2008) 10:262–269 269269