Embed Size (px)
Citation preview

Molecular & Biochemical Parasitology 120 (2002) 157–161
Short communication
The Cryptosporidium par�um ABC protein family�
Fernando Zapata a,1, Margaret E. Perkins b,2, Ynolde A. Riojas b,3, Teresa W. Wu b,4,Sylvie M. Le Blancq b,*
a Department of Pediatrics, Di�ision of Gastroenterology and Nutrition, Columbia Uni�ersity College of Physicians and Surgeons, New York, NY10032, USA
b Di�ision of En�ironmental Health Sciences, Mailman School of Public Health, Columbia Uni�ersity, New York, NY 10032, USA
Received 9 May 2001; accepted in revised form 7 December 2001
Keywords: ABC protein; Cryptosporidium par�um ; Drug resistance; Apicomplexa; MRP; MDR; Intron
www.parasitology-online.com.
Cryptosporidium par�um causes cryptosporidiosis, alife-threatening diarrheal disease in immunocompro-mized hosts, for which there is no consistently effectivetreatment [1]. Indeed, C. par�um exhibits an intrinsicresistance to standard drugs that are effective againstrelated organisms [2]. The basis of this phenotype is notknown, although, C. par�um ’s unique niche at thebrush border of infected epithelial cells may be a con-tributory factor. C. par�um infection starts by invasionof enterocytes by sporozoites, the motile extracellularstage released from infectious oocysts. The intracellularstages develop inside a vacuole at the apex of the hostcell. The main site of contact between the maturing
parasite and the host cell is an extensively folded mem-brane structure, called the feeder organelle.
ATP-binding cassette (ABC) proteins are key compo-nents of the cellular machinery for endobiotic andxenobiotic detoxification, and overexpression of certainABC proteins results in multiple drug resistance [3,4].Most ABC proteins are ATP-driven membrane trans-porters that translocate substrates to various intra- andextracellular compartments [3]. C. par�um ’s ABCproteins are likely to be involved in many essentialaspects of transport and may contribute to the intrinsicdrug resistance phenotype. We previously demonstratedthat SDZ-PSC833 and other cyclosporin analogs arepotent inhibitors of intracellular growth of C. par�umin vitro [5]. SDZ-PSC833 binds to the mammalian ABCprotein, MDR1 [6] and modulates its function as anefflux pump [7]. It is therefore possible that PSC833mediates its anticryptosporidial effects by modulatingthe activity of one or more of C. par�um ’s ABCproteins. Cryptosporidium ABC proteins are therefore‘drug targets of considerable interest’ [8]. We havepreviously characterized the first C. par�um ABCprotein, CpABC1, an �200 kDa protein found insporozoites and intracellular stages [9]. Here we presentan extension of the analysis of CpABC1, the character-ization of a new CpABC protein, CpABC2, and apreliminary assessment of the C. par�um ABC proteingene repertoire.
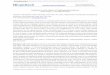
The localization of CpABC1 in sporozoites is shownin Fig. 1. CpABC1 is broadly distributed through thesporozoite with some concentration in the apical region
Abbre�iations: ABC, ATP-binding cassette; ALDP1,adrenoleukodystrophy protein; CFTR, cystic fibrosis transmembraneconductance regulator; IFA, immunofluorescence analysis; MDR,multidrug resistance protein; MRP, multidrug resistance-associatedprotein; ORF, open reading frame; PDR, pleiotropic drug resistanceprotein; RLI, RNAse L inhibitor; YEF3, yeast elongation factor 3.
� Note: Nucleotide sequence data reported in this paper are avail-able in the GenBank™ databases under the accession numbersU90628, AF315508, AF315509.
* Corresponding author. Present address: Doris Duke CharitableFoundation, 650 Fifth Avenue, 19th floor, New York, NY 10019,USA. Tel.: +1-212-974-7105; fax: +1-212-974-7950.
E-mail address: [email protected] (S.M. Le Blancq).1 F. Zapata, M.E. Perkins and Y.A. Riojas contributed equally to
the work reported in this manuscript.2 Present address: Harlem Hospital, New York, NY 10037, USA.3 Present address: New York City Department of Health, NY
10007, USA.4 Present address: Skirball Institute, New York University, NY
10016, USA.
0166-6851/02/$ - see front matter © 2002 Elsevier Science B.V. All rights reserved.PII: S 0 1 6 6 -6851 (01 )00445 -5

F. Zapata et al. / Molecular & Biochemical Parasitology 120 (2002) 157–161158
Fig. 1. Confocal images of immunofluorescence localization of CpABC1 and CpABC2 on C. par�um sporozoites. Sporozoites were excysted fromoocysts by incubation in PBS at 37 °C for 1 h. For IFA with Rab�GST-CpABC1 [9], smears of parasites were air-dried, fixed in 0.5%glutaraldehyde for 2 min and permeabilized in 0.4% Triton X-100 for 2 min followed by a wash in PBS. For IFA with Rab�GST-CpABC2,air-dried parasite smears were fixed in 0.5% glutaraldehyde for 1 min and permeabilized in 0.1% Triton-X for 2 min followed by a PBS wash.Smears were overlaid with Rab�GST-CpABC1 (diluted 1:75), Rab�GST-CpABC2 (diluted 1:75) or pre-immune rabbit serum (diluted 1:75) for1 h, washed in PBS, and incubated with FITC goat anti-rabbit antibody. Smears were overlaid with propidium iodide (2 �g ml−1) for 1 minfollowed by a wash in PBS. Preimmmune sera did not detect specific proteins in sporozoites (data not shown). IFA was carried out by confocalmicroscopy with a LSM 410 Zeiss Confocal Microscope. Confocal images are: panel A, Rab�CpABC2 (green) and nucleus (red); panel B,Rab�CpABC1 (green) and nuclei (red); panels C and D are the propidium iodide (red) and the FITC (green) signals shown in panel B displayedseparately. Green, FITC; red, propidium iodide.
and anterior to the nucleus (Fig. 1, panels B–D). Thepattern of fluorescence was punctate suggesting theprotein is compartmentalized.
Confocal immunofluorescence analysis (IFA) ofCpABC1 in intracellular stages of C. par�um is shownin Fig. 2. Studies on meronts released from their hostcells showed CpABC1 to be localized at the peripheryof the meront (Fig. 2B). Serial 1 �m sections throughthe sample are instructive (Fig. 2A). In the first sec-tions, at the base of the meront where no nuclei arevisible, CpABC1 is present and localized in a punctatepattern uniformly across the meront (Fig. 2, bottom ofpanel A). In the middle sections the punctate pattern ofCpABC1 is distributed around the periphery of themeront. At the top sections, CpABC1 signal is verylow. This is consistent with a hemispherical distributionof CpABC1 at the periphery of meronts. There was nopositive fluorescence when either infected Caco-2 cellswere stained with pre-immune serum or when IFA wasperformed on uninfected Caco-2 cells (data not shown).CpABC1 was previously shown to be localized at theboundary between host cells and the extracytoplasmicmeront [9]. The boundary localization of CpABC1 isclearly shown in the longitudinal section of the apicalregion of an infected Caco-2 cell (Fig. 2C). A diagramdepicting the localization of CpABC1 is shown in panelD.
The localization of CpABC1 to vesicle-like structuresin sporozoites (Fig. 1) suggests a pattern somewhatsimilar to that depicted for dense granules (described in[8]). Dense granule proteins are thought to be involvedin remodeling of the parasitophorous vacuole mem-brane [10]. Indeed, the distribution of CpABC1 in ahemispherical structure at the host–parasite boundaryof mature meronts suggests that it is a component of
one or more of the membranes at this interface, i.e. themeront plasma membrane, the parasitophorous vacuolemembrane, and the feeder organelle. CpABC1’s loca-tion suggests that it plays a role in the metabolicinteractions between the parasite and host cell duringparasite maturation.
CpABC2 is a single copy gene located on chromo-some 2 (data not shown). The complete CpABC2 se-quence was obtained from CpEST.453 [11], twooverlapping clones from a genomic library and by5’RACE, and is available under GenBank accessionnumber AF315508. CpABC2 has an uninterruptedopen reading frame (ORF) that encodes a putative1587-amino acid protein. A 5.3 kb CpABC2 transcriptwas identified in sporozoites by Northern analysis (datanot shown). A highly-specific rabbit antiserum, referredto as Rab�GST-CpABC2, was developed against theintracellular loop immediately preceding the C-terminalnucleotide binding domain in CpABC2 (amino acids1243–1323). Immunoblot analysis with Rab�GST-CpABC2 identified a �190 kDa protein in sporozoitesconsistent with the predicted molecular weight of 180.5kDa plus glycosylation (data not shown). IFA showedCpABC2 localized to the apical end of sporozoites (Fig.1, panel A). It was not possible to determine from theconfocal image if the protein was localized in the apicalorganelles or on the membranes surrounding these or-ganelles or the plasma membrane. The location ofCpABC2 in the apical region of sporozoites suggeststhat it is involved in invasion and/or the very earlystages of the establishment of the trophozoite. Thestorage of CpABC2 transcripts in the sporozoite wouldallow rapid de novo protein synthesis to supplement thestored proteins. CpABC2 was not detected either byimmunoblot or IFA in intracellular stages of C. par�um

F. Zapata et al. / Molecular & Biochemical Parasitology 120 (2002) 157–161 159
that were harvested from infected Caco-2 cells culturedfor either 24 or 42 h post-inoculation with oocysts (datanot shown).
The repertoire of ABC protein genes in C. par�umwas estimated through the identification of putativeCpABC genes from 5020 C. par�um Genome SurveySequences (GSSs) [11]. Fifteen GSS clones, likely repre-
senting 7–9 independent ABC protein-encoding genes(see below), were identified in addition to CpABC1 andCpABC2. The 5020 GSSs total �3.4×106 bp of se-quence [11], i.e. �35% of the 10.4 Mb C. par�umgenome. Therefore, there are about 11 CpABC genes inapproximately one-third of the genome, and the totalcomplement of ABC proteins in C. par�um is likely tobe approximately 33. This is in accord with otherunicellular eukaryotes; for example, the yeast Saccha-romyces cere�isiae has 30 ABC protein-encoding genes[12]; and Plasmodium is predicted to have �30 [13].
We have constructed a complete ORF for one ofthese ABC protein genes, designated CpABC3.CpABC3 is located on chromosome 6 (data not shown).The sequence of CpABC3 (GenBank accession numberAF315509) was obtained from GSS clone CpG0689[11], two overlapping clones and a PCR product fromphage-based genomic libraries. The genomic sequenceof the CpABC3 gene was interrupted by a 56 nucleotidetype 0 intron that was spliced-out of the mRNA tran-script (data not shown). The ORF encoded a putative1394-amino acid protein with a predicted molecularweight of 156.7 kDa. Introns have been described inonly two of 25 (8%) C. par�um genes analyzed thus far,i.e. CpABC3 and �-tubulin [14]. Interestingly, this con-trasts to other apicomplexans, such as Plasmodiumfalciparum, in which approximately 50% of sequencedgenes have introns [15].
A CpABC3 transcript was not detected by Northernanalysis of RNA from sporozoites (data not shown).Expression of the CpABC3 gene was detected in intra-cellular stages of C. par�um through the use of RT-PCR of RNA from cultures that had been maintainedfor 48 h after inoculation with oocysts (data notshown). Monolayers of infected cells cultured for 48 hcontain developing sexual stages [16]. However, whiledetection of the transcript at this late time point couldbe due to stage-specific expression it is also possible
Fig. 2.
Fig. 2. Confocal images of immunofluorescence localization ofCpABC1 with Rab�CpABC1 and FITC goat anti-rabbit serum onmeronts from C. par�um-infected Caco-2 cells. Caco-2 cell cultureswere inoculated with oocysts and cultured for 42 h before harvesting[5,9]. Smears of cells and parasites were air-dried, fixed in EtOH for4 min and permeabilized in −20 °C acetone for 10 min. IFA wascarried out as described in Fig. 1. Smears were overlaid with propid-ium iodide (5 �g ml−1) for 1 min followed by a PBS wash. Panel A,1 �m serial sections through a released meront; panel B, sum ofsections of the meront sectioned in panel A; panel C, longitudinalsection at the apex of an infected Caco-2 cell; panel D, schematic ofthe images in panel C. The host cell is lightly shaded, the meront(dark shading) protrudes into the extracellular space (white). Thegreen band represents the CpABC1 FITC signal. The series of linesparallel to the host cell membrane indicate the relative position of thesections through the released meront shown in panel A. The sectionsand lines are from top to bottom and right to left, respectively.Green, FITC; red, propidium iodide; gray, phase-contrast image;purple, superimposed phase-contrast and FITC images.

F. Zapata et al. / Molecular & Biochemical Parasitology 120 (2002) 157–161160
that it reflects the increased overall number of parasites[5].
ABC proteins can be classified by sequence similarityinto many subfamilies. For example, S. cere�isiae ’s 30ABC proteins are grouped into six subfamilies: theMDR, PDR, MRP/CFTR, ALDP, YEF3 and RLIsubfamilies5 [12]. There are marked structure– functionrelationships within ABC protein subfamilies andacross phyla. For example, members of the MRP sub-family transport organic anions [17] and some arefunctional homologues, i.e. human MRP1 can comple-ment yeast MRP YCF1 [18]. The C. par�um ABCproteins were assigned to ABC protein subfamiliesthrough the use of BLAST searches of the S. cere�isiaedatabase for each CpABC gene and the putative GSSclones. CpABC1 and CpABC2 are members of theMRP subfamily and CpABC3 grouped with the MDRsubfamily (data not shown). Of the GSS clones ana-lyzed, 1 PDR, 1 ALDP, 1 MDR, and multiple MRPsequences were identified. The distribution of CpABCgenes among the subfamilies is intriguing: however, itshould be noted that the preponderance of MRP sub-family members in the one-third of the genome ana-lyzed thus far may not reflect the actual composition ofthe ABC protein family in C. par�um.
The functional equivalence of subfamily membersacross species and phyla suggests that, as MRP subfam-ily members, both CpABC1 and CpABC2 are organicanion transporters [19]. Substrates of four of the sixyeast MRPs have been identified. They includeoligomycin (YOR1) [20], glutathione S-conjugates(YCF1) [18,21], bile acids (YBT1) [22] and unconju-gated bilirubin (BPT1) [23]. YCF1 is a key player invacuolar detoxification of heavy metals [24], and YOR1mediates drug resistance through efflux across theplasma membrane [20]. Thus, CpABC1 and CpABC2may function as transporters of endogenous and xeno-biotic glutathione conjugates and it is reasonable toconjecture that they may contribute to C. par�um ’sintrinsic drug resistance phenotype. The localization ofCpABC1 to the primary exchange site between parasiteand host is consistent with such a role. The CpABC3protein is most similar to the S. cere�isiae MDR sub-family member, STE6. Other MDR subfamily mem-bers, such as the P. falciparum P-glycoproteinhomologue 1 (Pgh1), can functionally complement theSTE6 gene in S. cere�isiae [25]. Interestingly, mutationsin Pgh1 can confer resistance to the antimalarial drugs
mefloquine, quinine, and halofantrine [26], suggesting apossible role for CpABC3 in the management ofxenobiotics.
Our analysis suggests that C. par�um has approxi-mately 33 ABC protein genes. Studies to identify thesubstrates and transport activities of these proteins andtheir physiological roles will depend on heterologousexpression in tractable model systems. Such studies willbe crucial to understanding the contribution ofCpABCs to C. par�um ’s intrinsic drug resistance phe-notype and their potential as targets for rational drugdesign.
Acknowledgements
We thank the National Institutes of Health AIDSResearch and Reagents Program, Rick Nelson and BillStrong (C. par�um Genome Survey Sequence (GSS) andExpressed Sequence Tag (EST) projects) for reagents,and Richard Friedman (Comprehensive Cancer CenterComputer Facility) for sequence alignments. This workwas supported by National Institutes of Health GrantsAI 41351 and AI46287.
References
[1] Blagburn BL, Soave R. Prophylaxis and chemotherapy: humanand animal. In: Fayer R, editor. Cryptosporidium and cryp-tosporidiosis. Boca Raton, FL: CRC Press, 1997:111–28.
[2] Woods KM, Nesterenko MV, Upton SJ. Efficacy of 101 antimi-crobials and other agents on the development of Cryptosporidiumpar�um in vitro. Ann Trop Med Parasitol 1996;90:603–15.
[3] Bauer BE, Wolfger H, Kuchler K. Inventory and function ofyeast ABC proteins: about sex, stress, pleiotropic drug and heavymetal resistance. Biochim Biophys Acta 1999;1461:217–36.
[4] Borst P, Zelcer N, van Helvoort A. ABC transporters. BiochimBiophys Acta 2000;1486:128–44.
[5] Perkins ME, Wu TW, Le Blancq SM. Cyclosporin analogsinhibit in vitro growth of Cryptosporidium par�um. AntimicrobAgents Chemother 1998;42:843–938.
[6] Jette L, Murphy GF, Beliveau R. Drug binding to P-glyco-protein is inhibited in normal tissues following SDZ-PSC 833treatment. Int J Cancer 1998;76:729–37.
[7] Twentyman PR. Cyclosporins as drug resistance modifiers.Biochem Pharmacol 1992;43:109–17.
[8] Coombs GH. Biochemical peculiarities and drug targets in Cryp-tosporidium par�um : lessons from other coccidian parasites. Par-asitol Today 1999;15:333–8.
[9] Perkins ME, Riojas YA, Wu TW, Le Blancq SM. CpABC, aCryptosporidium par�um ATP-binding cassette protein at thehost–parasite boundary in intracellular stages. Proc Natl AcadSci USA 1999;96:5734–9.
[10] Dubremetz JF, Garcia-Reguet N, Conseil V, Fourmaux MN.Apical organelles and host-cell invasion by Apicomplexa. Int JParasitol 1998;28:1007–13.
[11] Strong WB, Nelson RG. Preliminary profile of the Cryptosporid-ium par�um genome: an expressed sequence tag and genomesurvey sequence analysis. Mol Biochem Parasitol 2000;107:1–32.
5 These designations reflect the most prominent mammalian(ALDP, MRP1, CFTR, MDR1, RLI) or yeast (PDR5, YEF1) mem-ber(s) of each subfamily. ALDP1, adrenoleukodystrophy protein;MRP1, multidrug resistance-associated protein 1; CFTR, cystic fibro-sis transmembrane conductance regulator, MDR1, multidrug resis-tance protein 1; RLI, RNAse L inhibitor; PDR5, pleiotropic drugresistance protein 5; YEF3, yeast elongation factor 3

F. Zapata et al. / Molecular & Biochemical Parasitology 120 (2002) 157–161 161
[12] Taglicht D, Michaelis S. Saccharomyces cere�isiae ABC proteinsand their relevance to human health and disease. MethodsEnzymol 1998;292:130–62.
[13] Gardner MJ, Tettelin H, Carucci DJ, et al. The malaria sequenc-ing project: complete sequence of Plasmodium falciparum chro-mosome 2. Parassitologia 1999;41:69–75.
[14] Widmer G, Tchack L, Chappell CL, Tzipori S. Sequence poly-morphism in the �-tubulin gene reveals heterogeneous and vari-able population structure in Cryptosporidium par�um. ApplEnviron Microbiol 1998;64:4477–81.
[15] Bowman S, Lawson D, Basham D, et al. The complete nucle-otide sequence of chromosome 3 of Plasmodium falciparum.Nature 1999;400:532–8.
[16] Bonnin A, Gut J, Dubremetz JF, Nelson RG, Camerlynck P.Monoclonal antibodies identify a subset of dense granules inCryptosporidium par�um zoites and gamonts. J Eukaryot Micro-biol 1995;42:395–401.
[17] Borst P, Evers R, Kool M, Wijnholds J. A family of drugtransporters: the multidrug resistance-associated proteins. J NatlCancer Inst 2000;92:1295–302.
[18] Tommasini R, Evers R, Vogt E, et al. The human multidrugresistance-associated protein functionally complements the yeastcadmium resistance factor 1. Proc Natl Acad Sci USA1996;93:6743–8.
[19] Borst P, Evers R, Kool M, Wijnholds J. The multidrug resis-tance protein family. Biochim Biophys Acta 1999;1461:347–57.
[20] Katzmann DJ, Hallstrom TC, Voet M, et al. Expression of anATP-binding cassette transporter-encoding gene (YOR1) is re-quired for oligomycin resistance in Saccharomyces cere�isiae.Mol Cell Biol 1995;15:6875–83.
[21] Li ZS, Szczypka M, Lu YP, Thiele DJ, Rea PA. The humanmultidrug resistance-associated protein functionally comple-ments the yeast cadmium resistance factor1. J Biol Chem1996;271:6509–17.
[22] Ortiz DF, St. Pierre MV, Abdulmessih A, Arias IM. A yeastATP-binding cassette-type protein mediating ATP-dependentbile acid transport. J Biol Chem 1997;272:15385–465.
[23] Petrovic S, Pascolo L, Gallo R, et al. The products of YCF1 andYLL015w (BPT1) cooperate for the ATP-dependent vacuolartransport of unconjugated bilirubin in Saccharomyces cere�isiae.Yeast 2000;16:561–71.
[24] Ghosh M, Shen J, Rosen BP. Pathways of As(III) detoxificationin Saccharomyces cere�isiae. Proc Natl Acad Sci USA1999;96:5001–6.
[25] Volkman SK, Cowman AF, Wirth DF. Functional complemen-tation of the ste6 gene of Saccharomyces cere�isiae with thepfmdr1 gene of Plasmodium falciparum. Proc Natl Acad Sci USA1995;92:8921–5.
[26] Reed MB, Saliba KJ, Caruana SR, Kirk K, Cowman AF. Pgh1modulates sensitivity and resistance to multiple antimalarials inPlasmodium falciparum. Nature 2000;403:906–9.