Embed Size (px)
Citation preview

J Comp Physiol A (1996) 178:813-830 �9 Springer-Verlag 1996
M . S. Tu �9 M . H . D i c k i n s o n
The control of wing kinematics by two steering muscles of the blowfly (Calliphora vicina)
Accepted: 22 November 1995
Abstract We used a combination of high speed video and electrophysiological recordings to investigate the relationship between wing kinematics and the firing patterns of the first (bl) and second (b2) basalar muscles of tethered flying blowflies (Calliphora vicina). The bl typically fires once during every wing stroke near the time of the dorsal stroke reversal. The b2 fires either intermittently or in bursts that may be elicited by a visual turning stimulus. Sustained activation of the bl at rates near wing beat frequency appears necessary for the tonic maintenance of stroke amplitude. In addition, advances in the phase of b 1 activation were correlated with both increased wing protraction during the down- stroke and increased stroke amplitude. Similar kin- ematic alterations were correlated with b2 spikes, and consequently, both muscles may function in the control of turns toward the contralateral side. The effects of the two muscles were evident within a single stroke period and decayed quickly. Kinematic changes correlated with bl phase shifts were graded, suggesting a role in compensatory course stabilization. In contrast, b2 spikes were correlated with all-or-none changes in the wing stroke, a characteristic consistent with a role in mediating rapid turns towards or away from objects.
Key words K i n e m a t i c s - Insect flight �9 Motor control �9 Dip te ra . Muscle
Abbreviat ions bl first basalar muscle �9 b2 second basalar muscle �9 PWP pleural wing process - RS radial stop - S wing span - 13 angle between the
M. S. Tu 1 �9 M. H. Dickinson 2 (l:~) The Department of Anatomy and Organismal Biology, The University of Chicago, 1025 E 57th St, Chicago I1 60637, USA
Present addresses: Department of Zoology N J-15, University of Washington,
Seattle WA, 98195, USA 2 Department of Integrative Biology, University of California, Berkeley CA, 94720, USA
stroke plane and the longitudinal body axis - (I) stroke amplitude �9 dO stroke elevation - L wing length �9 tPbl phase of bl activation - udb2 phase of b2 activation �9 0 stroke deviation.
Introduction
Flies perform an extraordinary array of complex aerial maneuvers (Land and Collett 1974; Wagner 1986a, b, c), and understanding how flies accomplish these intricate behaviors is one important goal in the study of insect flight. The motor control of flight is a multi-step pro- cess. Complex interactions between the nervous system, muscles, and skeleton determine the motion of the wings during flight, and modulation of wing kinematics causes changes in aerodynamic forces that are essential for flight control. Recent studies have begun to clarify the relationship between specific kinematic parameters and aerodynamic force generation in flies (Zanker and G6tz 1990; Dickinson and G6tz 1993; Dickinson 1994; Dickinson and G6tz, unpublished). Relatively little is known, however, about how specific steering muscles affect the kinematics of the wing stroke.
Anatomical studies have identified an elaborate array of 17 pairs of synchronous muscles that are thought to function in flight by directly or indirectly altering the mechanics of the wing hinge (summarized in Wisser and Nachtigall 1984). These synchronous muscles are physiologically and anatomically distinct from the large, asynchronous muscles that generate the power for flight. Electrophysiological recordings dur- ing tethered flight in Drosophila (G6tz 1983; Heide 1983), Musca (Heide 1975; Egelhaaf 1989), and Calli- phora (Nachtigall and Wilson 1967; Heide 1971b, 1975, 1983) indicate that the activities of several identified steering muscles change during both spontaneous and visually-induced flight maneuvers. Mounting evidence suggests that the steering muscles control many

814 M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies
, m
Ba2Ba MP \
p S 3 ~
I mrn Fig. 1 The basalar muscles of Calliphora. The first (bl) and second (b2) basalar muscles have their origins on the walls of the thorax and insert on the inwardly projecting arm of the basalare (Ba). The triangular head region of the basalar is visible externally and articu- lates anteriorly with the mesopraescutum (MP) and the posterior margin of the dorsal episternum (dashed line). A tendon (not shown) passes from the basalar 2 (Ba2) to the radial stop (RS). Rostral and dorsal rotation of the basalare caused by contraction of the bl or the b2 pronates the basalar 2 and pulls the wing base forward. sp = spiracle, PWP = pleural wing process, PS = pleural suture, b3 = third basalar muscle. Modified from Tu and Dickinson (1994)
kinematic variables including wing beat frequency, stroke amplitude (Nachtigall and Wilson 1967; G6tz 1968; Heide 1983; Nachtigall and Roth 1983; Hengstenberg et al. 1986; G6tz 1987; Lehmann 1990) and the timing of wing rotation at ventral stroke reversal (Dickinson et al. 1993). However, with the exception of experiments by Lehmann (1990) on the second basalar muscle of Drosophila, neither recordings nor stimulation of the steering muscles during flight has been coupled with simultaneous measurements of wing kinematics.
In our attempt to understand how motor output from the nervous system controls flight maneuvers, we have focused on the first and second basalar muscles (bl and b2, nomenclature of Heide 1971a) in the blow fly Calliphora vicina. Both the bl and the b2 originate laterally and ventrally on the wails of the anterior thorax and insert on the inwardly projecting region of the nail-shaped basalare (Fig. 1). The triangular 'head' region of the basalare is visible externally and articu- lates with the thoracic cuticle immediately anterior to the wing hinge. During flight, the mechanical linkage of
the basalare to the wing hinge causes the basalare to pivot back and forth in phase with the up and down movements of the wings (Nalbach 1989). The anatom- ical arrangement of the bl and the b2 should enable both muscles to regulate either the mean basalare posi- tion and/or the dynamics of the basalare oscillations during flight. These changes in basalare motion could alter the kinematics of the wing stroke by altering the mechanical configuration of the wing hinge.
Although the bl and the b2 are anatomically situ- ated to produce similar mechanical effects on the wing hinge, their patterns of activity during flight differ con- siderably. The b2 is normally active only during turns towards the contralateral side and has been implicated in the control of wing stroke amplitude (Heide 1971b, 1975, 1983; G6tz 1983; Egelhaaf 1989; Lehmann 1990). The firing pattern of the bl is more complex and its function less well understood. Of the 6 direct steering muscles that have been recorded from during tethered flight, only the bl is continuously active. The other 5 muscles are selectively activated during turning ma- neuvers. During straight flight in both Calliphora and Drosophila, the bl fires a single action potential in nearly every wing beat cycle (Drosophila, Ewing 1979a, b; G6tz 1983; Calliphora, Heide 1971b, 1975, 1983; Kutsch and Hug 1981). Heide (1975) also reported bl activity near wing beat frequency in Musca, although Egelhaaf (1989) reported a much lower spike rate in the same species. In all species studied so far, the phase of bl activation shifts during visually-induced turns so that the bl on the outside of the turn fires in advance of the bl on the contralateral side (Heide 1983; G6tz 1983; Egelhaaf 1989). In addition, the spike frequency of the bl on the inside of a turn may decrease (Heide 1983; Egelhaaf 1989). In Musca, shifts in the frequency and phase of bl activation are correlated with fluctuations in yaw torque, suggesting that the bl can influence the production of aerodynamic forces (Egelhaaf 1989). During visually-induced turns in Drosophila, the phase relationships of the ventral stroke reversal change in a manner similar to the phase shifts in bl activation (Dickinson et al. 1993). This finding suggests that the bl might alter aerodynamic force production by regula- ting the timing of wing rotation.
When stimulated under isometric conditions at wing beat frequency, the bl of Calliphora reaches a nearly complete tetanus (Heide 1971b; Bergmann-Erb and Heide 1990; Tu and Dickinson 1994). From this result, it is not immediately obvious how changes in activation phase of the bl could cause changes in kinematics and force production. The isometric performance of any muscle, however, does not necessarily provide an accu- rate picture of its functional capabilities in the intact animal. In a previous study (Tu and Dickinson 1994), we found that the steady state work output of the bl from Calliphora during cyclic length oscillations de- pends strongly on the phase of activation. This result demonstrated that the bl has the capacity to regulate

M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies 815
wing kinematics even though it is normally active at frequencies that would produce tetanus under isomet- ric conditions. On the basis of these experiments, how- ever, we could not directly address the question of whether or not flies actually utilize phase-dependent changes in work output to control flight behavior. In the present study, we combined extracellular record- ings with high speed video in order to explicitly deter- mine the effects of bl and b2 firing phase on the wing kinematics of Calliphora vicina.
Materials and methods
Specimen preparation
Adult Calliphora vicina were taken from laboratory cultures 1 2 weeks after eclosion. Fly tethers were fashioned from # 0 insect pins. To form a stable area of attachment to the fly, we bent a hairpin loop in one end of the pin at a right angle to the pin shaft. Prior to tethering the fly, two pairs of insulated 28 gauge wires were secured to the tether above the attachment point of the fly with cyanoac- rylate adhesive. The ends of the wires attached to the tether were stripped to form terminals for the electrode wires. Short lengths (approximately 1 cm) of un-insulated 25.4 gm diameter platinum- iridium wire (A-M Systems) soldered to the terminals on the tether functioned as electrodes. The free ends of the electrode wires were sharpened by cutting the end of the wire at an acute angle with a razor blade on a brass block.
Flies were anesthetized by cooling them for 3~4 mins at - 4~ The tether was positioned over the dorsal midline of the mesopraes- cutum and secured to the fly with a small drop of a beeswax- collophonium mixture (beeswax: collophonium, 1 : 3) applied using an electrocautery tool (Model 100, Geiger-NY). To aid in visualizing the wing movements we applied a small spot of white water-based enamel paint to the wing tip immediately posterior to the j unction of vein R~+5 and the wing margin. Using cuticular landmarks to determine electrode placement, we implanted bipolar extracellular electrodes in the bl and the b2 on the right side of each fly. The pairs of electrodes were pushed through the cuticle of the fly and into the muscles, with the two wires aligned either perpendicular (bl) or parallel (b2) to the muscle fibers. Coagulation of haemolymph around the entrance site of the electrodes was sufficient to secure the electrodes in place. Following electrode implantation, we suspended each tethered fly in a rack above a 2 cm diameter polypropylene sphere floating in a small beaker of water. The flies were positioned at a height allowing them to walk on the sphere and to reach the wet surface of the sphere to drink. We fed the flies a concentrated sugar solution 1-2 times daily. In this manner, tethered flies with im- planted electrodes could be maintained for a week or longer. Fol- lowing electrode implantation, we allowed flies to recover for a min- imum of 8 h before experiments. Most of the data, however was collected from flies 2-9 days after electrode implantation. During this period we did not observe any obvious effects of the tethering procedure on the physical condition of the flies. In particular, where- as the wings of free-flying animals in our colony often become damaged after one to two weeks, the wings of the tethered flies remained in good condition over a similar period.
Wind and visual stimuli
Tethered Calliphora were more likely to sustain flight in an air current than they were in still air. Consequently, we positioned the flies facing a small open-throat wind tunnel fitted with an exit cone tapering to a diameter of 3.5 cm. The fan generated a nearly laminar
air current with a speed of approximately 2 ms- 1. We also provided a visual ground movement stimulus. A longitudinally stripped pat- tern cylinder mounted on a DC motor and positioned concentrically around a tubular, 60 W incandescent lamp projected a stripe pattern onto a horizontally placed sheet of translucent white plastic. Rota- tion of the cylinder produced front to back motion of the projected stripe pattern at an average speed of 0.3 ms 1.
Because of the high frame rate used to record video images, frame by frame analysis involved a practical compromise between the length and number of continuous flight sequences that we could analyze. Under these constraints, the frequency of spontaneous turning maneuvers during sustained tethered flight was generally too low to allow us to examine the full range of kinematic variation in the wing stroke. To increase the frequency of turning maneuvers, we provided the fly with a coarse visual turning stimulus by sus- pending a pendulum in front of the fly. The pendulum, constructed of a 7 • 0.8 cm cylindrical brass rod had an oscillation frequency of approximately 1.5 Hz. We positioned the pendulum in front and slightly above the animal so that its motion did not interefere with the wind stimulus. Despite the high light intensity required for high speed video imaging, the flies responded to the side-to-side motion of the pendulum with changes in wing stroke amplitude, head position and leg posture.
Data acquisition
We recorded video images using a Kodak EktaPro Intensified Imager fitted with a Micro-Nikkor 200 mm lens and a 2 x telecon- vertor (Fig. 2a). The acquisition rate was 3000 frames s- 1 with an exposure time for each frame of 30 ~ts. We illuminated the wing stroke envelope using four fiber optic light sources. To avoid over- heating the fly, we fitted the lights with blue-green glass filters (Schott, BG-38). The fly was oriented with its right side towards the video camera and its sagittal plane perpendicular to the camera lens.
While video taping each flight sequence, we recorded simultan- eously from both the bl and b2. The timing of each muscle spike was recorded on the video tape as a flash in one of two LED's placed in the video field (Fig. 2). Signals from the bl and b2 electrodes were amplified (Model 1800, A-M Systems) and sent to one of two spike height window discriminators (Frederick Haer). The output from each window discriminator triggered one channel of a digital stimu- lator (Master-8, A.M.P.I.) which provided a 0.33 ms current pulse to the appropriate LED. We also recorded the extracellular potentials with a sampling rate of 8 kHz per channel using a 12-bit analog to digital converter (Digidatal200, Axon Instruments ) and data ac- quisition software (Axotape 2.0, Axon Instruments).
The digital memory of the EktaPro system limited the duration of continuous video recordings to 3.3 s. Each video sequence was transferred onto SVHS tape at 30 frames s- 1 for later analysis. After recording each series of flight sequences, we removed the fly and replaced it with a metal bar of known length to calibrate distance measurements. We sacrificed each fly by freezing and then measured the length (L) of the right wing from the base of the costal vein to the center of the white spot on the wing tip.
Data analysis
Images from sequences selected for detailed analysis were captured and stored on a computer (Power Macintosh 8100/80AV) and ana- lyzed using NIH Image software. The coordinate systems and the angles used to describe the wing kinematics are shown in Fig. 3. Using the coordinate system of the computer monitor, we digitized the horizontal (x) and vertical (z) coordinates of two points in each frame: the wing tip (identified by the white spot on the wing), and the wing base. Because the wing base was often difficult to identify in some frames, the wing base coordinates for the entire sequence were averaged and the mean wing base coordinate (Xo, Zo) was use in all
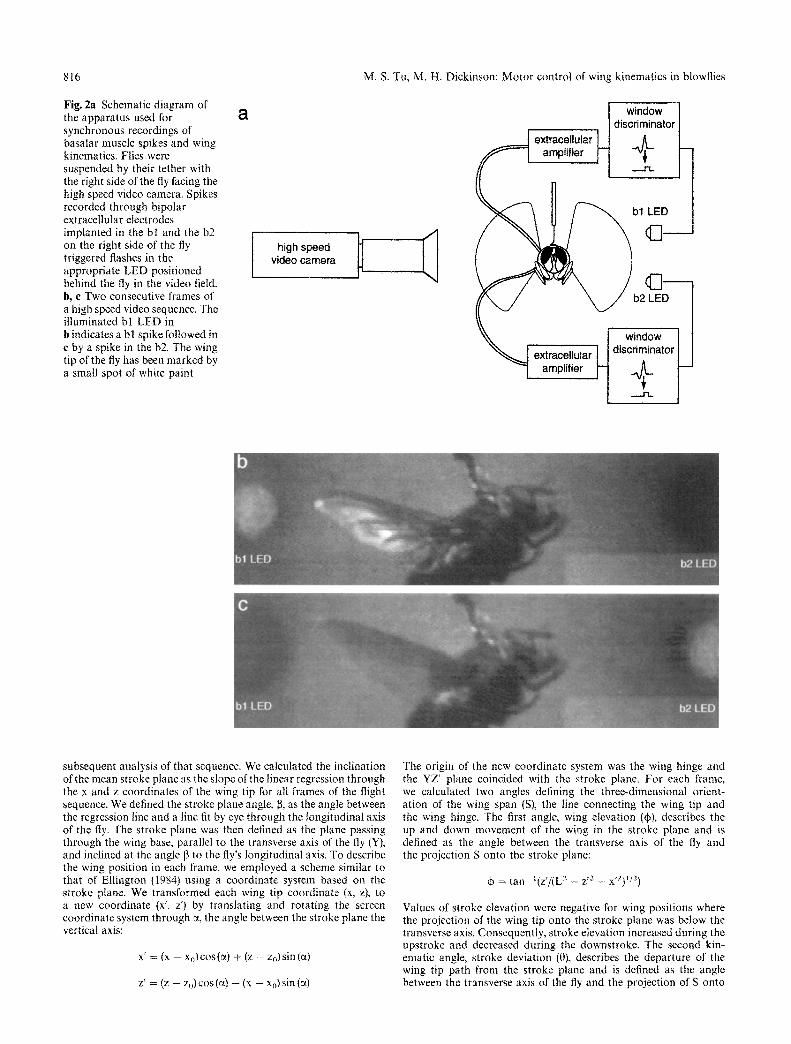
816
Fig. 2a Schematic diagram of the apparatus used for synchronous recordings of basalar muscle spikes and wing kinematics, Flies were suspended by their tether with the right side of the fly facing the high speed video camera. Spikes recorded through bipolar extracellular electrodes implanted in the bl and the b2 on the right side of the fly triggered flashes in the appropriate LED positioned behind the fly in the video field. b, e Two consecutive frames of a high speed video sequence. The illuminated bl LED in b indicates a bl spike followed in e by a spike in the b2. The wing tip of the fly has been marked by a small spot of white paint
a
high speed I video camera
M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies
window ~._~ extrace!lular I] disc'i~inat~ I
(/~--- ' amplifier q ~-- i~
\ t.. wi~176 I I ~'~.,,..] extracellular t I d~scr'm'na'~ I [ ~1 amplifier }'-1 .-~-'~L-~ ~'1
subsequent analysis of that sequence. We calculated the inclination of the mean stroke plane as the slope of the linear regression through the x and z coordinates of the wing tip for all frames of the flight sequence. We defined the stroke plane angle, 13, as the angle between the regression line and a line fit by eye through the longitudinal axis of the fly. The stroke plane was then defined as the plane passing through the wing base, parallel to the transverse axis of the fly (Y), and inclined at the angle 13 to the fly's longitudinal axis. To describe the wing position in each frame, we employed a scheme similar to that of Etlington (1984) using a coordinate system based on the stroke plane. We transformed each wing tip coordinate (x, z), to a new coordinate (x', z') by translating and rotating the screen coordinate system through 0~, the angle between the stroke plane the vertical axis:
x' = (x - xo)cos(~) + (z - Zo)sin (~)
z' = (z - Zo)COS(~X)- (x - xo) sin (a)
The origin of the new coordinate system was the wing hinge and the YZ' plane coincided with the stroke plane. For each frame, we calculated two angles defining the three-dimensional orient- ation of the wing span (S), the line connecting the wing tip and the wing hinge. The first angle, wing elevation (~), describes the up and down movement of the wing in the stroke plane and is defined as the angle between the transverse axis of the fly and the projection S onto the stroke plane:
~b = tan- l(z'/(LZ - z 'e - x'2) l/z)
Values of stroke elevation were negative for wing positions where the projection of the wing tip onto the stroke plane was below the transverse axis. Consequently, stroke elevation increased during the upstroke and decreased during the downstroke. The second kin- ematic angle, stroke deviation (0), describes the departure of the wing tip path from the stroke plane and is defined as the angle between the transverse axis of the fly and the projection of S onto

M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies 817
'N~ SP X' -- \ \ /
Fig. 3 Coordinate axes and angles used to describe wing kinematics. The angle [3 describes the inclination of the stroke plane (SP) to the longitudinal body axis of the fly (LA). The axis Z' lies in the stroke plane and is perpendicular to X'. Stroke elevation, qb, describes the angular position of the wing in the stroke plane relative to the transverse axis (Y) passing through the wing base. Values of stroke elevation were positive when the projection of the wing tip onto the stroke plane was above the transverse axis, and negative when the projection was below the transverse axis. Stroke deviation, 0, de- scribes the angular position of the wing above or below the stroke plane and was positive when the wing tip was above the stroke plane and negative when the wing tip was below the stroke plane
data set from 9 flies consisted of 143 flight sequences comprising 472 s of flight and 1.4 x 10 6 video frames. For detailed frame by frame analysis of muscle record- ing sequences, we selected one or two representative flight sequences of ls duration from each of 5 flies. In total, we digitized the wing tip position in 18,000 video frames from 6 flight sequences: 5 from sequences during which the bl was continuously active near wing beat frequency, and 1 additional sequence during which the bl was intermittently active. All results obtained from analysis of the selected sequences were qualitatively verified by examination of the remaining video records. In the 5 sequences with sustained bl activity, the aver- age wing beat frequency was 141 -t- 8 Hz (S.D., n = 707 cycles). The mean angle of the stroke plane, 13, for the same 5 sequences was 41.5~ 7 ~ At this wing beat frequency, the stroke cycle was sampled on average at a rate of 21 frames cycle-1, and measurements of the phase had an uncertainty of approximately + 9 ~ In the sequence during which the bl was intermittently active, the wing beat frequency was 119 Hz. The maximum variation in the calculated angles due to digitizing errors was 7 ~ for stroke deviation and 8 ~ for stroke elevation, determined by 5 repetitive analyses of the same video sequence. These errors were greatest near the dorsal and ventral stroke reversals.
Patterns of muscle activity
the X'Y plane:
0 = sin l(x'/(L2 -- z'2) 1/2)
Values of stroke deviation were positive when the wing tip was above the stroke plane and negative when the wing tip was below the stroke plane. Note that our nomenclature differs from that of Ellin- gton (1984) in that we use stroke elevation to describe the component of the wing motion in the stroke plane and stroke deviation to describe the component of wing motion perpendicular to the stroke plane. The bl and b2 firing times were determined by recording the frame number of each frame in which LED flashes were recorded. With the LED flash duration of 330 ps and the frame rate of 3000 frames s- ~, a single LED flash could be visible in at most two video frames. In the rare cases in which an LED was illuminated in two consecutive frames, the number of the first frame was taken as the time of muscle activation. For each sequence analyzed in detail, we used the digital recordings of the muscle potentials to verify the LED triggering. We defined stroke cycles by the local minimum values of stroke elevation. The phase of b l (Wbx) spikes and of b2 spikes (Wb2) were calculated from the time of the spike occurrence relative to the time of the preceding ventral stroke reversal, identified as a local minimum value of wing elevation. By our definition, a decrease in phase value corresponds to an advance in spike occurrence to a time earlier in the wing beat cycle.
Results
We recorded data only from flies that flew consistently with a tucked leg posture and a large wing stroke amplitude. These flies typically sustained flight until allowed to grasp a small piece of tissue paper. The total
In the absence of visual turning stimuli, or when the fly did not respond to visual stimulation, the bl fired a single spike during nearly every wing stroke, occa- sionally skipping 1 2 cycles (Figs. 4 and 9a, b). Under the same conditions, b2 activity occurred as single intermittent spikes. Continuous bl activity with occa- sional b2 spikes was typically correlated with large and bilaterally symmetrical wing stroke amplitudes with some jitter in the ventral extent of the wing stroke envelope.
In response to the swinging pendulum, the b2 fired in bursts that were typically 10-30 cycles in duration with a spike frequency varying from 1 spike per cycle to 1 spike in 4 cycles (Fig. 9c). The b2 rarely (2 in 96 digitized cycles) fired more than once in the same cycle. Since a detailed analysis of the optomotor responses of the flies was not the immediate aim of the present study, we did not encode the motion of the pendulum on the video recordings for subsequent analysis. We did ob- serve, however, that bursts of spikes from the b2 occur- red roughly in phase with the pendulum swing. Bursts of activity in the right b2 were correlated with increased stroke amplitude on the right side. The strength of the response to the moving pendulum varied with time, among individuals, and with the amplitude of the pen- dulum swing. During b2 bursts, the bl continued to fire at or near wing beat frequency. Between bursts of b2 spikes, the activity pattern of the bl was qualitatively

818 M.S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies
90 r
0
l.xl.tl. M_x.u.l 1.1] LI l_t_ 1.1 ll__ J_t V - ' - r ' - V - ~ - ' ~ - r - r ' - ' - ~ - T - ~ - ~ - r ' ~ - y - ' ~ ] r - r - - ' - "~-~-V-~- ' - r~ "- ' -~-w ~ - ' - r ~ - " w--~-.-' r - r , - W - , * ~ ~ ] ,
50 ms
Fig. 4 Stroke elevation (~), stroke deviation (0), and extracellular recordings from the bl and the b2. The LED flashes reliably encoded the timing of muscle spikes. Spike times determined by LED flashes on the video record are indicated on the traces of stroke elevation for both the bl, (closed circles) and the b2, (open circles). In this sequence, the bl fired a single spike near the dorsal stroke reversal of nearly every wing stroke although there were occasional cycles in which the bl did not fire. The b2 was intermittently active. Note that spikes in the b2 were accompanied by a bl phase advance although the phase of bl spikes was occasionally advanced without an accom- panying b2 spike. The angle (ordinate) scale is the same for both stroke elevation and stroke deviation
indistinguishable from that observed during flight in the absence of the visual stimulus.
There were occasionally prolonged periods during which the bl was completely inactive or fired sporadi- cally regardless of the presence or absence of visual stimuli (Fig. 9d). This behavior often occurred towards the end of the 2-5 h required to record a set of flight sequences from one individual. This sharp drop in bl firing rate was always accompanied by a large decrease in wing stroke amplitude.
Both bl and the b2 spikes occurred most frequently near the transition from the upstroke to the down- stroke (Fig. 4). Activation of the b2 was nearly always accompanied by an advance in bl phase, although advances in bl phase also occurred independently orb2 activity. In the 5 digitized sequences in which the rate of bl activation was close to wing beat frequency, the mean phase values of bl spikes were 147 +_ 38 ~ (S.D. n = 555) for cycles in which only the bl fired, and 88 _+ 29 ~ (n = 96) for cycles in which both the bl and the b2 fired, measured with respect to ventral stroke reversal. The mean phase of b2 spikes was 122 _+ 22 ~ (n = 96). In the same 5 sequences, the mean phase of the maximum wing elevation (dorsal stroke reversal) was 160_ 7 ~ (n = 707), corresponding to a ratio of upstroke to downstroke duration of 0.8 : 1.
Wing tip trajectories
To examine graphically the changes in kinematics asso- ciated with variations in bl and b2 activity, we aver-
aged stroke cycles with similar muscle firing patterns (Fig. 5). We divided each flight sequence into individual stroke cycles and then sorted the cycles into three categories. The first category included cycles in which only the bl was active. A cycle was included in this group if it contained one and only one bl spike. We excluded cycles if they contained a b2 spike or if the b2 had fired in any of the preceding two cycles. Kinematic changes that were correlated with b2 spikes typically decayed within two cycles (see below). Consequently, we are confident that this criterion was sufficient to separate kinematic changes correlated with bl phase from the residual effects of previous b2 spikes. We further sub-divided the cycles containing only bl spikes into 5 bins according to the phase of bl activation. The second category included all cycles in which the b2 fired. In almost all cases, there was also a bl spike in the same cycle. Because of the relatively small number of b2 spikes in each sequence, we did not sort the b2 cycles into separate phase bins. The third category included all cycles in which neither the bl nor the b2 fired. We calculated cycle averages of the wing tip projection coordinates (x', z') for each of the 5 bl phase bins, the b2 cycles and the cycles without bl and b2 activity (Fig. 5, Table 1). For the cycle averaging, we obtained corresponding points in each cycle by evaluating cubic spline interpolations of the time series x'(t) and z'(t) at 24 equally spaced time values. In the data shown in Fig. 5, periods in which neither the bl nor the b2 fired did not exceed 2 stroke cycles.
Table 1 Mean bl phase (bias 1-5) and b2 phase (bin b2) values for the averaged wing stroke trajectories shown in Fig. 5
Mean phase _+ S.D.(n)
Bin a: C27.33 b: C28.13 c: C24.19 d: C24.21
0 (11) (16) (4) (13) 1 32_+8(28) 62_+11(6) 92+11(4) 76_+11(18) 2 151 • 5(27) 98 _+ 12(8) 130 _+ 7(3) 99 _+ 7(10) 3 170 _+ 7(20) 140 _+ 11(21) 162 _+ 8(19) 130 • 8(18) 4 189 -+ 0(3) 179 _+ 8(16) 191 _+ 10(14) 164 _+ 10(12) 5 211 _+ 8(3) 214 • 9(4) 219 _+ 10(14) 189 • 0(4)
b2 119 _+ 29(21) 124 _+ 36(21) 134 _+ 49(15) 89 + 32(34)

M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies 819
a b
, , x , ,
t~2
~'0
Fig. Sa~l B1 phase advance and activation of the b2 were both correlated with a rostral shift in the downstroke trajectory. The largest changes in the stroke kinematics tended to occur in cycles in which the b2 fired. Each of the 4 data sets shown is from a flight sequence in which the bl fired near wing beat frequency, and the duration of periods in which neither the bl nor the b2 fired did not exceed 2 cycles. In each case, we have plotted the coordinates of the wing tip position projected onto the plane of the video image. For reference, we have drawn silhouette of the fly behind each set of wing tip trajectories. The location of the wing base for each fly is indicated by a cross. Arrowheads indicate the direction of wing movement. Within each data set, each trace represents the average of all cycles in which neither the bl nor the b2 fired (0, dashed line), only the bl fired (1 5) or the b2 fired (b2). Each of the 5 traces for bl-only cycles represents the average of cycles falling into one of 5 phase bins. When neither the bl or the b2 fired, or when the bl fired near the dorsal stroke reversal (high phase values), the wing tip generally traced a figure-8. In some cases the rostral shift in the downstroke caused the figure-8 to disappear completely as either bl phase was advanced or the b2 fired. The mean phase values for each averaged stroke trajectory are given in Table 1
Changes in wing stroke kinematics were strongly correlated with variations in bl and b2 activity. When neither the bl nor the b2 fired, the wing tip nearly always traced a figure-8 pattern (Fig. 5a-d). Through- out the ventral loop of the figure-8, the upstroke path was anterior to the downstroke path. This basic stroke trajectory was modified when the bl was active. At the highest values of bl phase (longest delay relative to ventral stroke reversal), the wing tip trajectories were often similar to the figure-8 path seen in isolated cycles when neither the bl nor the b2 fired (Fig. 5a, b). In all flight sequences, however, the averaged downstroke trajectory was shifted rostrally for each incremental decrease (advance) in bl phase. A similar rostral shift in the downstroke occurred when the b2 fired. As in- dicated in Fig. 4, b2 spikes were often accompanied by a phase advance in the bl, and consequently we cannot ascribe the strong effect on wing beat trajectory to the b2 per se. In 4 of the 5 digitized sequences, the most extreme shift in the downstroke trajectory was asso- ciated with b2 activity, suggesting that the b2 may have
820 M. S, Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies
20
0
-20
20
0
-20
b l
40 "~' 4�9
2o
0 ~ 1 . �9
-20 , 0 90 180 270
gtbl (deg)
14"k~ 5 40 a \
o o
8
b2
4
. . . . .
_ . . "_ _ : . . _":-m-~.. �9149
-20 , , , 0 90 180 270
~b2 (deg)
3
n n
�9 �9 - _ 0 �9 . . . . . .
�9 o � 9
u ,,, i i
20
0
-20
= - . ~ ' " a ,
�9 5
20
0
-20
" " - i ' ~ , - ~
_ r - , - . - x __.e . . . . .
6
20
0
-20
0
1
r f j
90 180 270
7
�9 " : . t - k
i a i
0 90 180 270
2O
O,
-20'
8
. . . . . . 9 . . . . . .
O
90 180 270
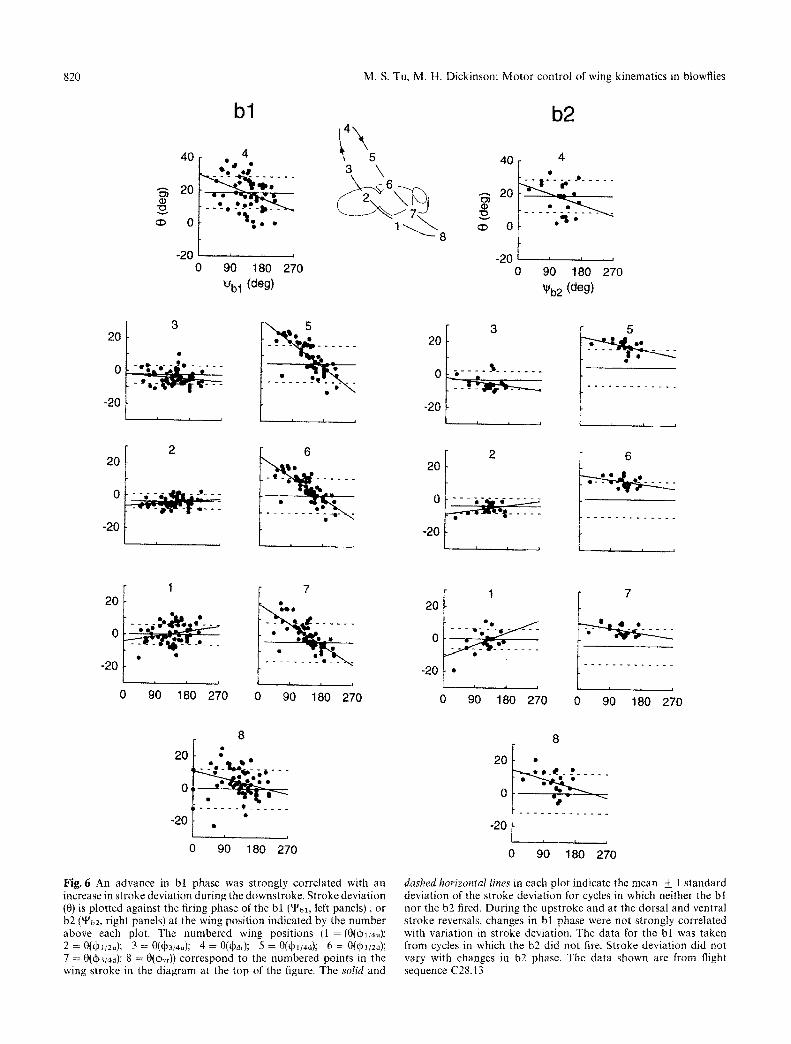
Fig. 6 An advance in bl phase was strongly correlated with an increase in stroke deviation during the downstroke. Stroke deviation (0) is plotted against the firing phase of the bl ('-l%x, left panels), or b2 (~b2, right panels) at the wing position indicated by the number above each plot. The numbered wing positions (1 = (0(qbl/4u); 2 = 0(~l/:u); 3 = 0(qb3/4u); 4 = 0(~r); 5 = 0(~w4a); 6 = 0(~1/2d); 7 = O(qb3/4a); 8 = O(d)vr)) correspond to the numbered points in the wing stroke in the diagram at the top of the figure�9 The solid and
20
0
-20 �9
0
1
i j
90 180 270
7
i I r
0 90 180 270
20
0
-20
0
8
" - ~ - -" . . . . .
L r k
90 180 270
dashed horizontal lines in each plot indicate the mean + 1 standard deviation of the stroke deviation for cycles in which neither the bl nor the b2 fired. During the upstroke and at the dorsal and ventral stroke reversals, changes in bl phase were not strongly correlated with variation in stroke deviation. The data for the bl was taken from cycles in which the b2 did not fire. Stroke deviation did not vary with changes in b2 phase. The data shown are from flight sequence C28.13
M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies 821
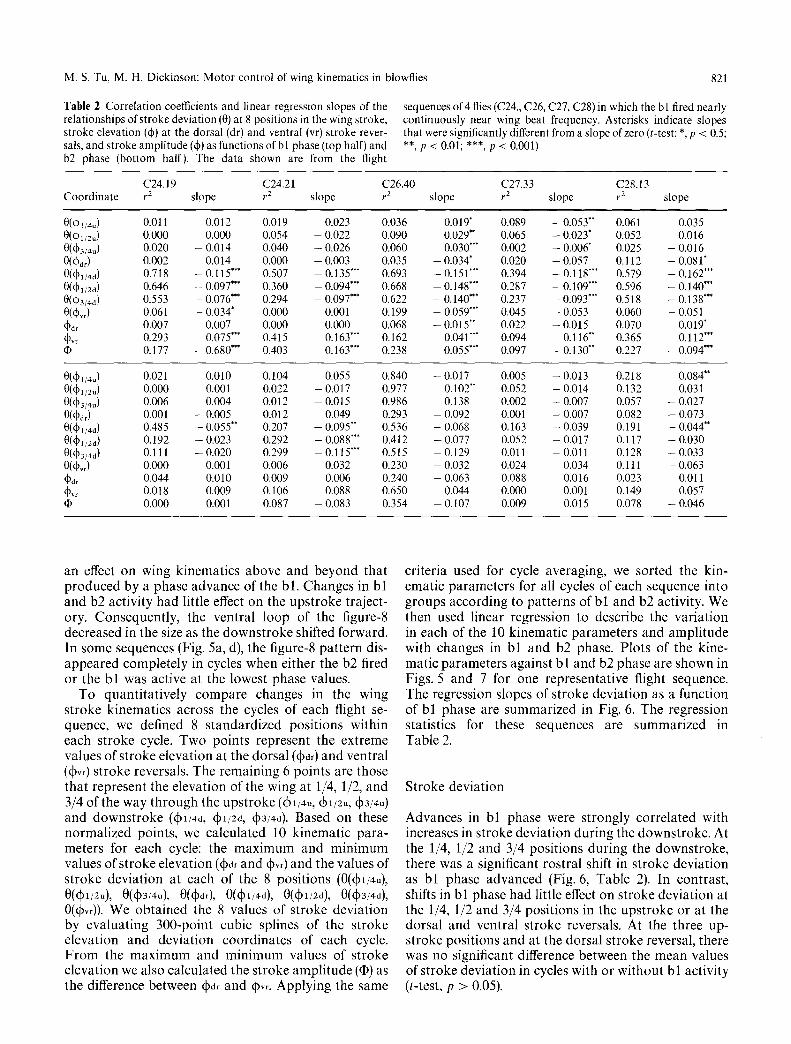
Table 2 Correlation coefficients and linear regression slopes of the relationships of stroke deviation (0) at 8 positions in the wing stroke, stroke elevation (qb) at the dorsal (dr) and ventral (vr) stroke rever- sals, and stroke amplitude (~) as functions of bl phase (top half) and b2 phase (bottom half). The data shown are from the flight
sequences of 4 flies (C24., C26, C27, C28) in which the bl fired nearly continuously near wing beat frequency. Asterisks indicate slopes that were significantly different from a slope of zero (t-test: *, p < 0.5; **, p < 0.01; ***, p < 0.001)
C24.19 C24.21 C26.40 C27.33 C28.13 Coordinate r 2 slope r z slope r 2 slope r 2 slope r 2 slope
0(c~1/4u ) 0.011 0.012 0.019 0.023 0.036 0.019" 0.089 - 0.053** 0.061 0.035 0(~1/2u) 0.000 0.000 0.054 - 0.022 0.090 0.029** 0.065 - 0.023* 0.052 0.016 0(4)314u) 0.020 - 0.014 0.040 - 0.026 0.060 0.030*** 0.002 - 0.006* 0.025 - 0.016 0(qbd~ ) 0.002 0.014 0.000 -- 0.003 0.035 -- 0.034* 0.020 -- 0.057 0.112 -- 0.081" 0(l~D1/4d ) 0.718 - 0.115"** 0.507 - 0.135"** 0.693 - 0.151'** 0.394 -- 0.118'** 0.579 -- 0.162"** 0(d01/2d) 0.646 -- 0.097*** 0.360 -- 0.094*** 0.668 -- 0.148"* 0.287 -- 0.109"** 0.596 -- 0.140"** 0((~)3,4d ) 0.553 -- 0.076*** 0.294 -- 0.097*** 0.622 - 0.140"** 0.237 - 0.093*** 0.518 - 0.138"** 0(~vr ) 0.061 - 0.034* 0.000 0.001 0.199 - 0.059*** 0.045 - 0.053 0.060 -- 0.051 ~dr 0.007 0.007 0.000 0.000 0.068 - 0.015 *+ 0.022 - 0.015 0.070 0.019" qbv~ 0.293 0.075*** 0.415 0.163"** 0.162 0.041'** 0.094 0.116"* 0.365 0.112"** q~ 0.177 - 0.680*** 0.403 - 0.163"** 0.238 - 0.055*** 0.097 - 0.130"* 0.227 - 0.094***
0((~li4u) 0.021 0.010 0.104 0.055 0.840 - 0.017 0.005 - 0.013 0.218 0.084** 0(qbli2u) 0.000 0.001 0.022 - 0.017 0.977 0.102"* 0.052 - 0.014 0.132 0.031 0((~3!4u ) 0.006 0.004 0.012 - 0.015 0.986 0.138 0.002 - 0.007 0.057 -- 0.027 0(Od,) 0.001 -- 0.005 0.012 0.049 0.293 -- 0.092 0.001 -- 0.007 0.082 -- 0.073 0(~l/4d) 0.485 -- 0.055** 0.207 -- 0.095** 0.536 -- 0.068 0.163 -- 0.039 0.191 -- 0.044** 0(l~)l/2d ) 0.192 - 0.023 0.292 - 0.088"** 0.412 -- 0.077 0.052 - 0.017 0.117 - 0.030 0(qb3i4d) 0.111 -- 0.020 0.299 -- 0.115"** 0.515 -- 0.129 0.011 -- 0.011 0.128 -- 0.033 0(~vr) 0.000 0.001 0.006 0.032 0.230 -- 0.032 0.024 0.034 0.111 -- 0.063 (~d, 0.044 0.010 0.009 0.006 0.240 -- 0.063 0.088 0.016 0.023 0.011 4)vr 0.018 0.009 0.106 0.088 0.650 0.044 0.000 0.001 0.149 0.057 @ 0.000 0.001 0.087 - 0.083 0.354 -- 0.107 0.009 0.015 0.078 -- 0.046
an effect on w ing k i n e m a t i c s a b o v e a n d b e y o n d t ha t p r o d u c e d b y a p h a s e a d v a n c e o f the b l . C h a n g e s in b l a n d b2 ac t iv i ty h a d l i t t le effect on the u p s t r o k e t ra jec t - ory. C o n s e q u e n t l y , the ven t r a l l o o p of the f igure-8 d e c r e a s e d in the size as the d o w n s t r o k e shif ted fo rward . In s o m e sequences (Fig. 5a, d), the f igure-8 p a t t e r n dis- a p p e a r e d c o m p l e t e l y in cycles when e i ther the b2 f ired o r the b l was ac t ive at the lowes t p h a s e values .
T o q u a n t i t a t i v e l y c o m p a r e changes in the wing s t r o k e k i n e m a t i c s ac ro s s the cycles of each fl ight se- quence , we def ined 8 s t a n d a r d i z e d p o s i t i o n s wi th in each s t r o k e cycle. T w o p o i n t s r ep re sen t the e x t r e m e va lues of s t r o k e e l e v a t i o n at the d o r s a l ((bur) a n d ven t ra l (qbvr) s t r o k e reversa ls . T h e r e m a i n i n g 6 p o i n t s a re those t ha t r e p r e s e n t the e l e v a t i o n of the wing at 1/4, 1/2, a n d 3/4 o f the w a y t h r o u g h the u p s t r o k e ((bl/4u, qbl/2u, d~3/4u) a n d d o w n s t r o k e (qbl/4a, qbl/2d, (b3/4d). Based on these n o r m a l i z e d po in t s , we c a l c u l a t e d i 0 k i n e m a t i c p a r a - me te r s for each cycle: the m a x i m u m a n d m i n i m u m va lues of s t r o k e e l e v a t i o n (qbar a n d ~w) a n d the va lues of s t r o k e d e v i a t i o n a t each o f the 8 p o s i t i o n s (0(qbl/4u), 0((~l/2u), 0((~)3/4u), 0((~dr), 0((~I/4-d), 0(+1/2d), 0((~)3/4-d), 0(40,r)). W e o b t a i n e d the 8 va lues o f s t r o k e d e v i a t i o n b y e v a l u a t i n g 300-po in t cub ic spl ines of the s t r o k e e l e v a t i o n a n d d e v i a t i o n c o o r d i n a t e s o f each cycle. F r o m the m a x i m u m a n d m i n i m u m va lues of s t r o k e e l e v a t i o n we a lso c a l c u l a t e d the s t r o k e a m p l i t u d e (O) as the d i f ference be tween ~/)dr a n d qbvr. A p p l y i n g the s a m e
c r i t e r ia used for cycle ave rag ing , we s o r t e d the k in - e m a t i c p a r a m e t e r s for all cycles of each sequence in to g r o u p s a c c o r d i n g to p a t t e r n s of b l a n d b2 ac t iv i ty . W e then used l inea r r eg re s s ion to desc r ibe the v a r i a t i o n in each o f the 10 k i n e m a t i c p a r a m e t e r s a n d a m p l i t u d e wi th c ha nge s in b l a n d b2 phase . P l o t s of the k ine- m a t i c p a r a m e t e r s a g a i n s t b l a n d b2 p h a s e a re s h o w n in Figs . 5 a n d 7 for one r e p r e s e n t a t i v e f l ight sequence . T h e r eg res s ion s lopes of s t r o k e d e v i a t i o n as a func t ion of b l p h a s e are s u m m a r i z e d in Fig . 6. The r eg res s ion s ta t i s t ics for these sequences a re s u m m a r i z e d in T a b l e 2.
S t r o k e d e v i a t i o n
A d v a n c e s in b l p h a s e were s t r o n g l y c o r r e l a t e d wi th inc reases in s t r o k e d e v i a t i o n d u r i n g the d o w n s t r o k e . At the 1/4, 1/2 a n d 3/4 p o s i t i o n s d u r i n g the d o w n s t r o k e , the re was a s ign i f ican t r o s t r a l shift in s t r o k e d e v i a t i o n as b l p h a s e a d v a n c e d (Fig. 6, T a b l e 2). In con t r a s t , shifts in b l p h a s e h a d l i t t le effect on s t r o k e d e v i a t i o n at the 1/4, 1/2 a n d 3/4 p o s i t i o n s in the u p s t r o k e o r a t the d o r s a l a n d ven t r a l s t r o k e reversals . A t the th ree up- s t r o k e p o s i t i o n s a n d at the d o r s a l s t r o k e reversa l , t he re was no s ign i f ican t d i f ference be tw e e n the m e a n va lues of s t r o k e d e v i a t i o n in cycles wi th o r w i t h o u t b l ac t iv i ty (t- test , p > 0.05).
822 M.S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies
0.05
0
-0.05
-0.1 o= -~ -0.15
-0.2
8
L I I I I I I I I I I , 1
1 2 3 4 5 6 7 8 1 2 3 4 wing positon
bl b2
@ dl.+. ~,~ 6 0
4 0
[ ,-.-, -20 + . . . . . . . . ; , . , _ _
t s . / " E l I . i F I I j r ~ ++++, -40 '~ �9 .:.~,,,,,~
6 o " '
I i i i
- - - 2 . - m _ . . . . . . .
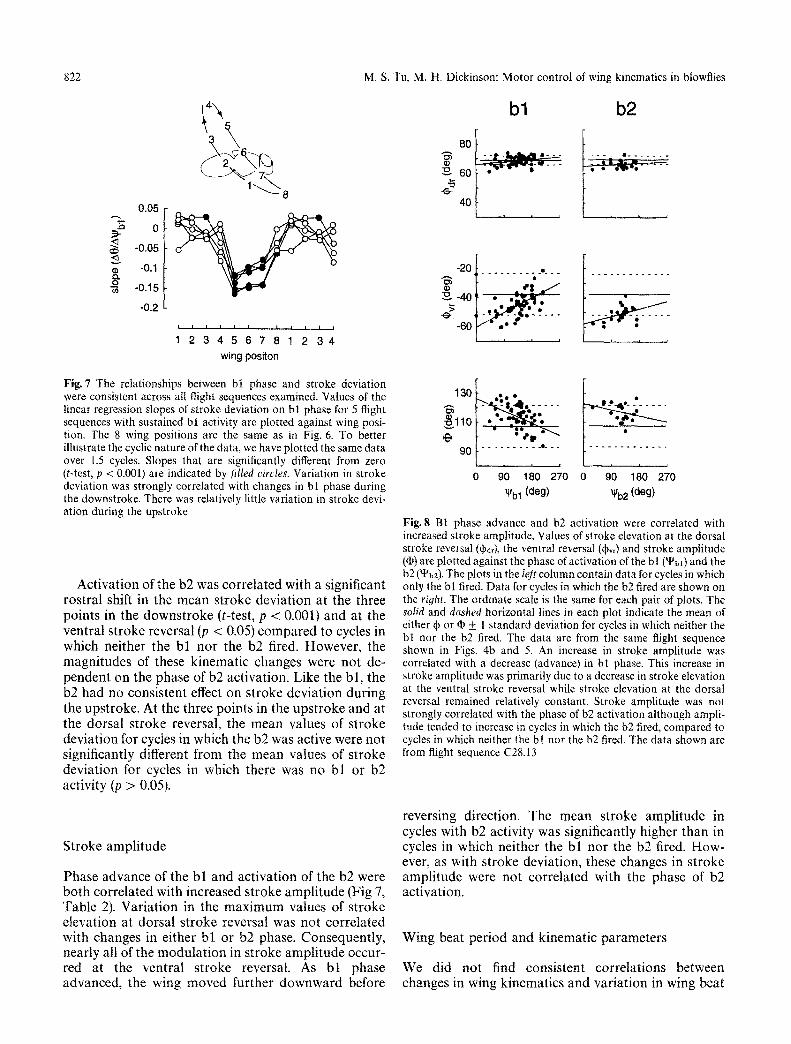
Fig.7 The relationships between bt phase and stroke deviation were consistent across all flight sequences examined. Values of the linear regression slopes of stroke deviation on bl phase for 5 flight sequences with sustained bl activity are plotted against wing posi- tion. The 8 wing positions are the same as in Fig. 6. To better illustrate the cyclic nature of the data, we have plotted the same data over 1.5 cycles. Slopes that are significantly different from zero (t-test, p < 0.001) are indicated by filled circles. Variation in stroke deviation was strongly correlated with changes in bl phase during the downstroke. There was relatively little variation in stroke devi- ation during the upstroke
Activat ion of the b2 was correlated with a significant rostrat shift in the mean s t roke deviat ion at the three points in the downs t roke (t-test, p < 0.001) and at the ventral s t roke reversal (p < 0.05) com pa red to cycles in which neither the b l nor the b2 fired. However , the magni tudes of these kinemat ic changes were not de- pendent on the phase of b2 activation. Like the b l , the b2 had no consistent effect on stroke deviat ion during the upstroke. At the three points in the ups t roke and at the dorsal s t roke reversal, the mean values of s t roke deviation for cycles in which the b2 was active were not significantly different f rom the mean values of s t roke deviat ion for cycles in which there was no b l or b2 activity (p > 0.05).
130
~110
90
�9 _ ' . .Ii 0 90 180 270 0 O0 180 270
~bl (deg) ~b2 (deg)
Fig. 8 B1 phase advance and b2 activation were correlated with increased stroke amplitude. Values of stroke elevation at the dorsal stroke reversal (qbdy, the ventral reversal (+v,) and stroke amplitude (q)) are plotted against the phase of activation of the b I (~bl) and the b2 (~b2). The plots in the left column contain data for cycles in which only the bl fired. Data for cycles in which the b2 fired are shown on the right. The ordinate scale is the same for each pair of plots. The solid and dashed horizontal lines in each plot indicate the mean of either ~ or q~ + 1 standard deviation for cycles in which neither the bl nor the b2 fired. The data are from the same flight sequence shown in Figs. 4b and 5. An increase in stroke amplitude was correlated with a decrease (advance) in bl phase. This increase in stroke amplitude was primarily due to a decrease in stroke elevation at the ventral stroke reversal while stroke elevation at the dorsal reversal remained relatively constant. Stroke amplitude was not strongly correlated with the phase of b2 activation although ampli- tude tended to increase in cycles in which the b2 fired, compared to cycles in which neither the bl nor the b2 fired. The data shown are from flight sequence C28.13
Stroke ampl i tude
Phase advance of the b l and act ivat ion of the b2 were bo th correlated with increased s t roke ampl i tude (Fig 7, Table 2). Variat ion in the m a x i m u m values of s troke elevation at dorsal s t roke reversal was not correlated with changes in either b l or b2 phase. Consequent ly, nearly all of the modu la t ion in s t roke ampl i tude occur- red at the ventral s t roke reversal. As b l phase advanced, the wing moved further downward before
reversing direction. The mean stroke ampl i tude in cycles with b2 activity was significantly higher than in cycles in which neither the b l nor the b2 fired. How- ever, as with stroke deviation, these changes in s t roke ampli tude were not correlated with the phase of b2 activation.
Wing beat per iod and kinemat ic pa ramete rs
We did not find consistent correlat ions between changes in wing kinematics and var ia t ion in wing beat
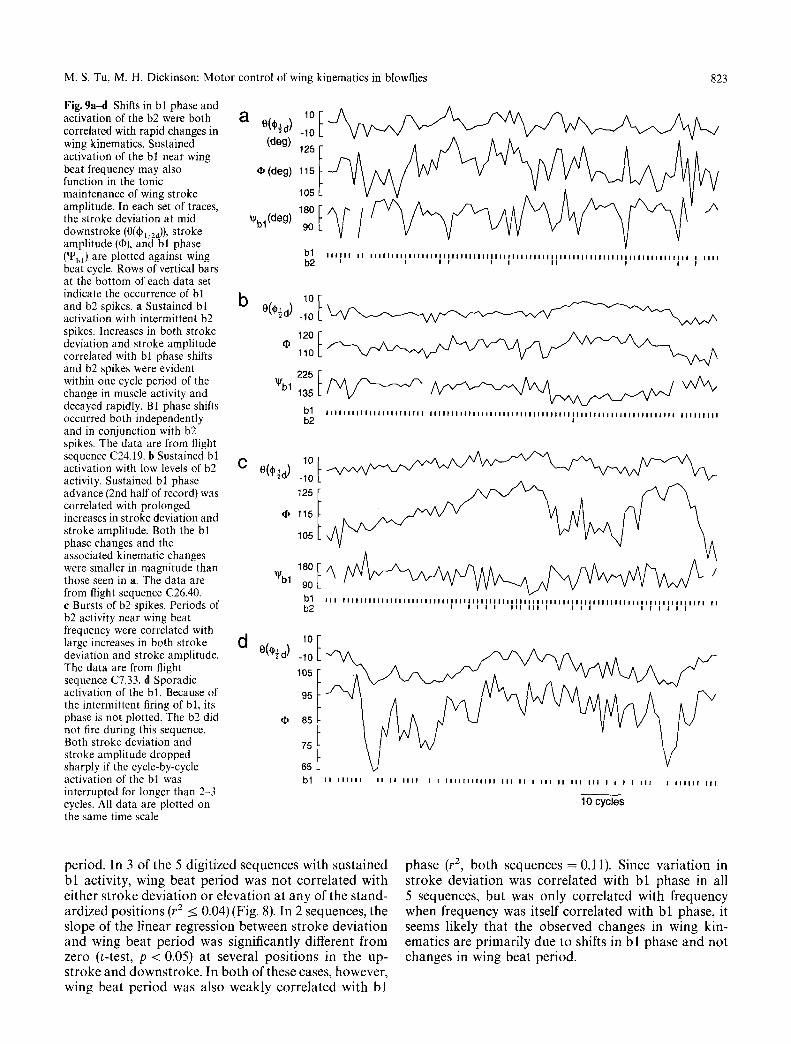
M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies 823
Fig. 9a~t Shifts in bl phase and activation of the b2 were both a 0t~'-dXW~l correlated with rapid changes in wing kinematics. Sustained (deg) activation of the b! near wing beat frequency may also �9 (deg) function in the tonic maintenance of wing stroke amplitude. In each set of traces, the stroke deviation at mid ~'bl(deg) downstroke (0(~1/2a)), stroke amplitude (q~), and bl phase (~bl) are plotted against wing beat cycle. Rows of vertical bars at the bottom of each data set indicate the occurrence of bl b and b2 spikes, a Sustained bl O(~d ) activation with intermittent b2 spikes. Increases in both stroke deviation and stroke amplitude correlated with bl phase shifts and b2 spikes were evident within one cycle period of the ~b l change in muscle activity and decayed rapidly. B1 phase shifts occurred both independently and in conjunction with b2 spikes. The data are from flight sequence C24.19. b Sustained bl activation with low levels of b2 C 0(O�89 activity. Sustained bl phase advance (2nd half of record) was correlated with prolonged increases in stroke deviation and stroke amplitude. Both the bl phase changes and the associated kinematic changes were smaller in magnitude than those seen in a. The data a r e "~I/bl from flight sequence C26,40. e Bursts of b2 spikes. Periods of b2 activity near wing beat frequency were correlated with large increases in both stroke d deviation and stroke amplitude. O,,v~d !/~._x The data are from flight sequence C7.33. d Sporadic activation of the bl. Because of the intermittent firing of bl, its phase is not plotted. The b2 did not fire during this sequence. Both stroke deviation and stroke amplitude dropped sharply if the cycle-by-cycle activation of the bl was interrupted for longer than 2-3 cycles. All data are plotted on the same time scale
125 f
115 I
105 L
180 f 90
bl , i J l l l I I l l l l l l l l l l l l l l l l l l l l l l l l l l i l J l l l ~ l l l l l l l l l l l l , l l , l l l r l t l l l l , , l l l j l , l r l l , l l , l l l j b2 I I ~ I I I I i i i
10
110
bl I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l I I I I I I I I I b2 i
lO
-lO 125
115
105
180
90 bl b2
10
-10
105
95
85
75
65 bl
, , , ' ' ' ' ' • ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' ' • ' • ' • ' • ' ' ' ' ' ' ' • ' • ' ' ' • ' • ' ' ' ' • ' ' • • • • • ' ' ' • ' ' ' ' • • ' • ' ' ' ' ' • ' ' ' ' • ' , , | I I I I I I I I I I I I
I I I I I I I I I I I I I I I I I I I I I I I I I l l l l l I I I I I I I I I I I I I I I I I I I I I I l l I I I I I l l | 1 1
10 cycles
period. In 3 of the 5 digitized sequences with sustained bl activity, wing beat period was not correlated with either stroke deviation or elevation at any of the stand- ardized positions (r 2 < 0.04) (Fig. 8). In 2 sequences, the slope of the linear regression between stroke deviation and wing beat period was significantly different from zero (t-test, p < 0.05) at several positions in the up- stroke and downstroke. In both of these cases, however, wing beat period was also weakly correlated with bl
phase (r 2, both sequences--0.11). Since variation in stroke deviation was correlated with bl phase in all 5 sequences, but was only correlated with frequency when frequency was itself correlated with bl phase, it seems likely that the observed changes in wing kin- ematics are primarily due to shifts in bl phase and not changes in wing beat period.
824 M.S. Tu, M. H, Dickinson: Motor control of wing kinematics in blowflies
.-%
~D
v
10
0
-10
125
115
105
bl advance b2 spike
.s 120 f
30 I I I I I ! i I I I J
-5-4-3-2-1 0 1 2 3 4 5
cycle number
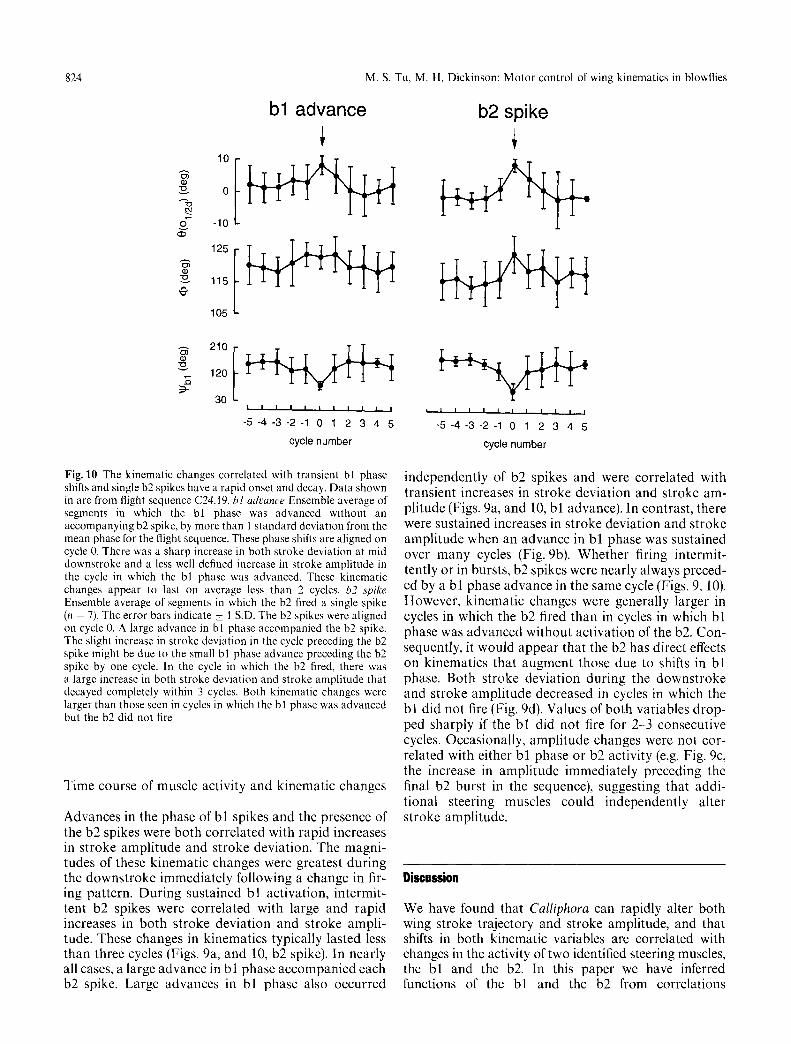
Fig. 10 The kinematic changes correlated with transient bl phase shifts and single b2 spikes have a rapid onset and decay. Data shown in are from flight sequence C24.19. bl advance Ensemble average of segments in which the bl phase was advanced without an accompanying b2 spike, by more than 1 standard deviation from the mean phase for the flight sequence. These phase shifts are aligned on cycle 0. There was a sharp increase in both stroke deviation at mid downstroke and a less well defined increase in stroke amplitude in the cycle in which the bl phase was advanced. These kinematic changes appear to last on average less than 2 cycles, b2 .spike Ensemble average of segments in which the b2 fired a single spike (n = 7). The error bars indicate + 1 S.D. The b2 spikes were aligned on cycle 0. A large advance in bl phase accompanied the b2 spike. The slight increase in stroke deviation in the cycle preceding the b2 spike might be due to the small bl phase advance preceding the b2 spike by one cycle. In the cycle in which the b2 fired, there was a large increase in both stroke deviation and stroke amplitude that decayed completely within 3 cycles. Both kinematic changes were larger than those seen in cycles in which the bl phase was advanced but the b2 did not fire
Time course of muscle activity and kinematic changes
Advances in the phase of bl spikes and the presence of the b2 spikes were both correlated with rapid increases in stroke amplitude and stroke deviation. The magni- tudes of these kinematic changes were greatest during the downstroke immediately following a change in fir- ing pattern. During sustained bl activation, intermit- tent b2 spikes were correlated with large and rapid increases in both stroke deviation and stroke ampli- tude. These changes in kinematics typically lasted less than three cycles (Figs. 9a, and 10, b2 spike). In nearly all cases, a large advance in bl phase accompanied each b2 spike. Large advances in bl phase also occurred
i I i i [ I I i i I I
-5-4-3-2-1 0 1 2 3 4 5
cycle number
independently of b2 spikes and were correlated with transient increases in stroke deviation and stroke am- plitude (Figs. 9a, and 10, bl advance). In contrast, there were sustained increases in stroke deviation and stroke amplitude when an advance in bl phase was sustained over many cycles (Fig. 9b). Whether firing intermit- tently or in bursts, b2 spikes were nearly always preced- ed by a bl phase advance in the same cycle (Figs. 9, I0). However, kinematic changes were generally larger in cycles in which the b2 fired than in cycles in which b l phase was advanced without activation of the b2. Con- sequently, it would appear that the b2 has direct effects on kinematics that augment those due to shifts in bl phase. Both stroke deviation during the downstroke and stroke amplitude decreased in cycles in which the bl did not fire (Fig. 9d). Values of both variables drop- ped sharply if the bl did not fire for 2-3 consecutive cycles. Occasionally, amplitude changes were not cor- related with either bl phase or b2 activity (e.g. Fig. 9c, the increase in amplitude immediately preceding the final b2 burst in the sequence), suggesting that addi- tional steering muscles could independently alter stroke amplitude.
Discussion
We have found that Calliphora can rapidly alter both wing stroke trajectory and stroke amplitude, and that shifts in both kinematic variables are correlated with changes in the activity of two identified steering muscles, the bl and the b2. In this paper we have inferred functions of the bl and the b2 from correlations

M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies 825
between patterns of endogenous muscle activity and changes in stroke kinematics. Consequently, our results strongly imply, but do not conclusively demonstrate, that changes in the patterns of bl and b2 activity cause changes in both stroke deviation and stroke amplitude. In addition, the observed variations in bl and b2 activity do not explain all of the observed variation in stroke kinematics. This is not surprising since the bl and the b2 are only two of approximately 14 pairs of synchronous steering muscles inserting directly on the sclerites of the wing hinge (Heide 1968, 1971a; Wisser and Nachtigall 1984). Besides the bl and the b2, the activity of only 4 of these muscles have been recorded during flight: the pterale muscles I1 and III1, and the muscles of the posterior notal wing process, hgl and hg3. Nachtigall and Wilson (1967) found that during spontaneous flight maneuvers by Muscina, Sarcophaga and Calliphora in which only one wing was beating, the b2 is active on the side of the beating wing while the hgl is active on the side of the non-beating wing. During visually-induced turning reactions by tethered Musca and Calliphora, the III1 and the b2 are active on the outside of the turn while the | 1 and the hg3 are active on the contralateral side (Heide 1971b, 1975; Egelhaaf 1989). Although in Drosophila, the III1 does not appear to function during visually-induced turning maneuvers, the b2 and the 11 appear to have the same functions as in Musca and Calliphora (Heide 1983; G6tz 1983). The results of these experiments suggest that stroke ampli- tude is increased by activation of the III1 and the b2, and decreased by activation of the 11, hg3 and hgl.
|n light of the known correlations between changes in stroke amplitude and activity by the | I l l , I1, hgl or hg3, it is unlikely that the amplitude changes observed in this study were caused exclusively by changes in bl and b2 activity (e.g. Fig 9c). Differences between the overall shapes of the wing tip trajectories among differ- ent flight sequences, even in data from the same fly (Fig. 5c, d), also suggest the influences of muscles be- sides the bl and the b2. Moreover, the effects of the III1, I1, hgl or hg3 on stroke deviation, and of the remaining steering muscles on both amplitude and stroke deviation, are currently unknown. The third basalar muscle (b3) is particularly intriguing in this respect since it is anatomically situated to act antagon- istically to both the bl and the b2. However, despite the potentially confounding effects of the other steering muscles, we were able to demonstrate consistent cor- relations between patterns of bl and b2 activity and changes in stroke kinematics. These results strongly imply that the bl and the b2, two of the largest steering muscles, play major roles in regulating both stroke amplitude and stroke deviation. In order to test the hypothesis that changes in the phase of the bl or b2 activation are sufficient to produce the changes in stroke deviation and amplitude reported here, we have performed experiments in which we stimulated either the bl or the b2 during flight. The results of these
stimulus experiments will be reported in a subsequent paper.
In the following sections, we will discuss the relation- ship between patterns of bl and b2 activity and flight behavior, and the mechanisms by which subtle changes in the firing phase of the bl could produce substantial changes in wing kinematics. We will also address the question of the extent to which the bl and the b2 might be functionally specialized to control different aspects of flight behavior. Finally, we will discuss the possible aerodynamic consequences of the kinematic alterations correlated with changes in b l and b2 activity.
B1 and b2 motor patterns and flight behavior
In this study, we did not attempt to correlate steering muscle activity with bilateral asymmetries in wing kin- ematics or with aerodynamic torque. Consequently we cannot directly link the observed patterns of muscle activity and wing kinematics to specific flight behav- iors. We can, however, make reasonable behavioral interpretations of our results. As the pendulum swung away from the flies, they increased stroke amplitude on the opposite side, consistent with the well characterized response to a small field visual stimulus (G6tz 1968, 1987; Nachtigall and Wilson 1967; Nachtigall and Roth 1983; Heide 1983; Hengstenberg et al. 1986). Changes in bl phase and b2 spikes are themselves indicative of turning reactions. During visually-induced turns in Calliphora and Musca, spikes in the bl on the outside of the turn are advanced in phase relative to spikes of the bl on the opposite side (Heide 1983; Egelhaaf 1989). In Musca, Calliphora and Drosophila, the b2 on each side of the fly is not normally active during straight flight but fires during turns to the contralateral side (G6tz 1983; Heide 1971b, 1975, 1983; Egelhaaf 1989). |n our data, bl activation late in each cycle combined with infrequent b2 spikes is likely to represent straight flight or turning reactions towards the ipsilateral side. In both Musca and Calliphora, delays in firing phase have been recorded from the bl on the outside of visually- induced turns (Heide 1983; Egelhaaf 1989). Low levels of activity are also expected from the b2 on the outside of a turn and during straight flight (Calliphora, Nach- tigall and Wilson 1967; Drosophila, Heide 1983). We found that the lowest values of stroke deviation and amplitude were correlated with the greatest delays in bl phase. Since we did not measure turning responses directly, we cannot determine whether these kinematics represent straight flight or turns towards the ipsilateral side.
Functions of the bl and b2
Our results suggest that both the bl and the b2 mediate rapid changes in stroke kinematics. We have also found

826 M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies
that maintenance of a large stroke amplitude may re- quire sustained activation of the bl. Under isometric conditions, the bl reaches a nearly complete tetanus when stimulated at wing beat frequency, suggesting that the bl functions primarily in the tonic control of wing kinematics (Heide 1971b; Bergmann-Erb and Heide 1990; Tu and Dickinson 1994). The bl might tonically regulate stroke amplitude by maintaining a rostral offset in the mean position of the basalare (Fig. 11). We have obtained preliminary evidence sup- porting this hypothesis by measuring the deflection of a laser beam reflected from a small mirror mounted on the external region of the basalare. These measure- ments have shown that during flight, the bl shortens tonically by 4% of its resting length with superimposed length oscillations of +_ 3% (Tu and Dickinson, unpub- lished).
Our results demonstrate that in addition to tonically maintaining a large stroke amplitude, the bl could also produce more subtle changes in wing kinematics in response to changes in activation phase. Shifts in the phase of bl activation in response to visual turning stimuli are correlated with both changes in the timing of wing rotation at ventral stroke reversal and modulation of aerodynamic torque (Heide 1983; Egelhaaf 1989; Dickinson et. al. 1993). Together, these observations strongly implicate the bl in the control of turning maneuvers. However, the isometric characteristics of the bl, which are consistent with slow tonic regulation of the wing stroke, have been puzzling in light of this evidence suggesting that the bl mediates rapid changes in the stroke kinematics. Heide (1983) suggested that the small residual force oscillations seen under isometric conditions could function in the fine control of wing kinematics. However, the mechanism by which activation phase might affect these small force transients to produce changes in the wing stroke have remained unclear.
As a first step in understanding the role of the bl during flight, we previously measured the mechanical output of this muscle as it was subjected to phasic stimulation and cyclic length changes at wing beat frequency (Tu and Dickinson 1994). Under these simulated flight conditions, the bl maintained a con- tinuously high level of dynamic stiffness. Consequently, the work required to extend the bl in each cycle ex- ceeded the work performed by the muscle during shortening, and the net work output per cycle was negative. However, we also found that the dynamic stiffness, the peak force and the magnitude of mecha- nical energy dissipation all varied with changes in acti- vation phase, implying that shifts in bl phase could potentially produce changes in wing kinematics. However, because we measured the output of the bl under steady state conditions, we could not predict its dynamic response to transient shifts in activation phase. We were also unable to specify how changes in the output of the bl could be transmitted through the
complex mechanical linkages of the dipteran wing hinge to modify the motion of the wing.
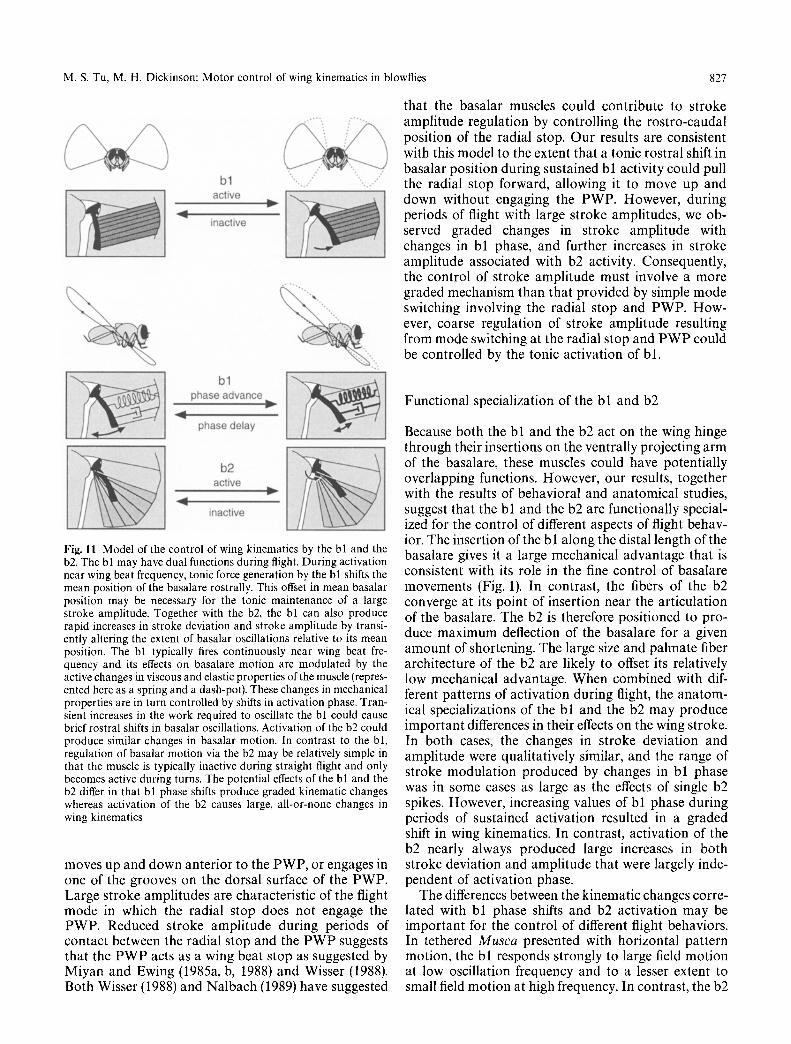
The results of the current study may help to clarify the functional importance of the phase-dependent changes in the mechanical properties of the bl. In this paper we have shown that advances in bl phase are indeed correlated with both increased stroke deviation and stroke amplitude. Moreover, these kinematic changes occur with a rapid onset and rapid decay, suggesting that the bl can alter wing kinematics on a cycle by cycle basis. Our measurements of negative work output by the bl imply that during flight, the musculo-skeletal system oscillating the bl must supply a net input of mechanical energy. Thus, the muscle may be approximated as a dissipative spring, with stiffness that can be modified on a cycle by cycle basis by changes in activation phase. During cyclical loading the work required to sustain bl length oscillations reaches a maximum when the muscle is activated in each cycle near its minimum length (Tu and Dickinson 1994). In the present study the maximal effects on stroke deviation occurred when the bl fired near the ventral stroke reversal. Due to the mechanical linkage of the basalar to the wing hinge, the bl should be maximally shortened at this wing position. Taken to- gether, these results imply that stroke amplitude and stroke deviation increase as the stiffness of the bl be- comes greater. In our muscle experiments, we held strain amplitude constant and measured the work re- quired to produce cyclic changes in bl length. During flight, if the magnitude of force oscillations acting on the basalare through the wing hinge remains relatively constant, then modulation bl stiffness should cause variations in the magnitude of oscillations of the ba- salare. In response to a phase advance, increased muscle stiffness should cause a decrease in the caudal extent of basalare oscillations. A rostral shift in the downstroke trajectory and increased stroke amplitude may therefore be caused by a forward shift in the mean basalare position (Fig. 11). Because of the anatomical position of the b2, activation of this muscle should cause a similar rostral shift in the mean position of the oscillating basalar. According to this model, advances in bl phase and activation of the b2 both produce similar alterations in wing kinematics because they cause similar changes in the motion of the basalare.
The head region of the basalare articulates at its posterior margin with the basalare 2 (Fig. 1). A tendon links the basalare 2 to the base of the radial vein so that rostral rotation of basalare exerts a forward pull on the wing base (Wisser and Nachtigall 1984; Nalbach 1989). The grooved tooth of the radial stop, a ventral projec- tion of the radial vein, suggests that it should articulate with one of the three peaks on the dorsal surface of the pleural wing process (PWP) (Miyan and Ewing, 1985a; Pfau 1987; Miyan and Ewing 1988; Wisser 1988; Nal- bach 1989). Nalbach (1989) found that flies can switch between flight modes in which the radial stop either
M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies 827
that the basalar muscles could contribute to stroke amplitude regulation by controlling the rostro-caudal position of the radial stop. Our results are consistent with this model to the extent that a tonic rostral shift in basalar position during sustained bl activity could pull the radial stop forward, allowing it to move up and down without engaging the PWP. However, during periods of flight with large stroke amplitudes, we ob- served graded changes in stroke amplitude with changes in bl phase, and further increases in stroke amplitude associated with b2 activity. Consequently, the control of stroke amplitude must involve a more graded mechanism than that provided by simple mode switching involving the radial stop and PWP. How- ever, coarse regulation of stroke amplitude resulting from mode switching at the radial stop and PWP could be controlled by the tonic activation of bl.
Fig. 11 Model of the control of wing kinematics by the bl and the b2. The bl may have dual functions during flight. During activation near wing beat frequency, tonic force generation by the bl shifts the mean position of the basalare rostrally. This offset in mean basalar position may be necessary for the tonic maintenance of a large stroke amplitude. Together with the b2, the bl can also produce rapid increases in stroke deviation and stroke amplitude by transi- ently altering the extent of basalar oscillations relative to its mean position. The bl typically fires continuously near wing beat fre- quency and its effects on basalare motion are modulated by the active changes in viscous and elastic properties of the muscle (repres- ented here as a spring and a dash-pot). These changes in mechanical properties are in turn controlled by shifts in activation phase. Tran- sient increases in the work required to oscillate the bl could cause brief rostral shifts in basalar oscillations. Activation of the b2 could produce similar changes in basalar motion. In contrast to the bl, regulation of basalar motion via the b2 may be relatively simple in that the muscle is typically inactive during straight flight and only becomes active during turns. The potential effects of the bl and the b2 differ in that bl phase shifts produce graded kinematic changes whereas activation of the b2 causes large, all-or-none changes in wing kinematics
moves up and down anterior to the PWP, or engages in one of the grooves on the dorsal surface of the PWP. Large stroke amplitudes are characteristic of the flight mode in which the radial stop does not engage the PWP. Reduced stroke amplitude during periods of contact between the radial stop and the PWP suggests that the PWP acts as a wing beat stop as suggested by Miyan and Ewing (1985a, b, 1988) and Wisser (1988). Both Wisser (1988) and Nalbach (1989) have suggested
Functional specialization of the bl and b2
Because both the bl and the b2 act on the wing hinge through their insertions on the ventrally projecting arm of the basalare, these muscles could have potentially overlapping functions. However, our results, together with the results of behavioral and anatomical studies, suggest that the bl and the b2 are functionally special- ized for the control of different aspects of flight behav- ior. The insertion of the bl along the distal length of the basalare gives it a large mechanical advantage that is consistent with its role in the fine control of basalare movements (Fig. 1). In contrast, the fibers of the b2 converge at its point of insertion near the articulation of the basalare. The b2 is therefore positioned to pro- duce maximum deflection of the basalare for a given amount of shortening. The large size and palmate fiber architecture of the b2 are likely to offset its relatively low mechanical advantage. When combined with dif- ferent patterns of activation during flight, the anatom- ical specializations of the bl and the b2 may produce important differences in their effects on the wing stroke. In both cases, the changes in stroke deviation and amplitude were qualitatively similar, and the range of stroke modulation produced by changes in bl phase was in some cases as large as the effects of single b2 spikes. However, increasing values of bl phase during periods of sustained activation resulted in a graded shift in wing kinematics. In contrast, activation of the b2 nearly always produced large increases in both stroke deviation and amplitude that were largely inde- pendent of activation phase.
The differences between the kinematic changes corre- lated with bl phase shifts and b2 activation may be important for the control of different flight behaviors. In tethered Musca presented with horizontal pattern motion, the bl responds strongly to large field motion at low oscillation frequency and to a lesser extent to small field motion at high frequency. In contrast, the b2

828 M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies
responds almost exclusively to small field motion at high frequency (Egelhaaf 1989). These results suggest that the bl functions primarily in compensatory course stabilization whereas the b2 functions in the control of turns towards objects. The large changes in wing kin- ematics correlated with b2 activation are consistent with a role in mediating voluntary turns during flight. In his studies of the free flight of Musca, Wagner (1986c) found that during pursuit behaviors, turns oc- cur as a series of rapid, large amplitude shifts in the angle of the body relative to the flight path. Further, activation of the b2 in Drosophila might be responsible for generating the rapid torque spikes that are observed in response to optomotor stimuli (Heisenberg and Wolf 1979, 1984). Our results suggest, however, that the bl and the b2 are not completely segregated in their function. In our experiments, b2 spikes were nearly always accompanied by an advance in bl phase, suggesting that the bl plays a role in augmenting or sustaining the effects of the b2. The bl receives input from wing beat synchronous afferents, most notably from the halteres (Heide 1983; Miyan and Ewing 1984; Fayyazuddin et al. 1994). Our observation that activation of the b2 nearly always coincides with a large phase advance of the bl implies that the descending visual input responsible for b2 activation might also delay or advance the activation of the bl by sensory afferents.
Shape of the wing tip path and flight forces
Reports of conflicting wing tip paths of flying Diptera under different experimental conditions have generated some confusion in the field of insect flight. The reported wing tip paths vary in their basic shape (figure-8 vs. open ellipsoid) and in the relative positions of the upstroke and downstroke. They have been reported for different species, from tethered flight in still and in moving air, from free forward flight, and from free hovering flight (free flying Calliphora, Drosophila, Simulium, and Tipula, Ennos 1989; tethered Muscina and Musca, Hollick 1940; tethered Phormia, Nachtigall 1966; tethered Calliphora, Wood 1970; tethered Drosophila, Zanker 1990a, b; Zanker and G6tz 1990; see discussion by Ellington 1984). Much of the debate over the different wing tip paths has focused on differ- ences between free and tethered flight, tethering pro- cedure (e.g. abdominal vs. thoracic) and the effects of still or moving air on tethered flight. As important as these differences in flight conditions may be, many of the apparent conflicts may represent different points along the continuum of kinematics under the active control by the fly. Flies rarely fly in straight lines and must therefore constantly modulate the kinematics of the wing stroke to generate aerodynamic torques. In addition, extensive studies of optomotor responses in flies have shown that flies actively modulate the kin-
ematics of their wing stroke during spontaneous ma- neuvers and in response to visual stimuli (Heisenberg and Wolf 1984). Much of the apparently conflicting data for wing tip paths may therefore arise from at- tempts to describe a typical or steady-state stroke path rather than the range of possible modulation under controlled variation of stimulus conditions. We have shown that Calliphora can actively control stroke devi- ation in addition to stroke amplitude. The fly can shift its wing tip trajectory from a figure-8 to an open loop by modulating the phase of bl activation or by activat- ing the b2. Moreover, these kinematic changes are not due to random variation but appear to function in the control of specific behaviors.
The existing confusion over wing tip paths may ob- scure fundamentally important size and species specific differences in the motor control of flight. Under care- fully controlled stimulus conditions, Zanker (1990b) described the kinematic modulation of the wing stroke of Drosophila in response to different wind and visual stimuli. Under all conditions, the wing path of Drosophila differs from that reported here for Calli- phora in that the upstroke path lies anterior to the path of the downstroke. This basic difference in the wing stroke could represent fundamental differences in the kinematics required for controlled generation of aero- dynamic forces at different body sizes. We cannot yet determine the aerodynamic consequences of the kin- ematic differences seen in Drosophila and Calliphora, or of the variation in wing kinematics observed within each species. However, a recent modeling study has shown that subtle changes in the upstroke and down- stroke trajectories can have important consequences for aerodynamic force production (Dickinson 1994). Precise control of the stroke trajectory may therefore be equally as important for flight steering as control of wing stroke amplitude.
It is interesting to note that the alterations in wing kinematics correlated with modulation of bl and b2 activity were primarily restricted to changes in the downstroke. In comparison, the upstroke was much less variable and also shorter in duration. Although we did not quantify the more subtle changes in kinematics such as angle of attack, our results suggesting that the bl and the b2 only affect the downstroke trajectory corroborate the findings of recent aerodynamic ana- lyses of insect flight. Recent flow visualization studies indicate that like most birds, flies produce a single vortex loop during each stroke from vorticity generated solely by the downstroke and the ventral stroke rever- sal (Musca, Grodinsky and Morozov 1993; Drosophila, Dickinson and G6tz, unpublished). Thus, in contrast to previous theories of dipteran flight (Nachtigall 1966, 1981), the upstroke may not contribute significantly to the generation of flight forces. Under these aero- dynamic constraints, it seems reasonable that during flight maneuvers, flies would modulate the kinematics of the downstroke but not the upstroke.

M. S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies 829
The flight control system of flies involves complex interactions between motor output from the nervous system, the physiological properties of specific steering muscles, the anatomical arrangement of these muscles, and the mechanics of the wing hinge. We have shown that tonic force generation in response to sustained activation of the bl may indeed function in the tonic maintenance of wing stroke amplitude. At the same time, the phase sensitivity of the mechanical properties of the b l during cyclic length oscillations appears to give the bl the additional capacity to produce phasic and graded changes in wing kinematics. The b2 differs markedly from the bl in that it is activated only during specific behaviors, and kinematic changes correlated with b2 spikes tend to be all or none rather than graded. Although both the bl and the b2 act on the wing hinge by regulating the oscillations of the basalare, the functional specializations of these two muscles appear to provide the fly with a flexible control system capable of mediating both com- pensatory course stabilization and voluntary turns towards objects.
Acknowledgements We thank Fritz-Olaf Lehmann for assistance in automating the video analysis and Wai Pang Chan for general technical assistance. This work was supported by NIH predoctoral training grant T32-GM-07839 to M.S.T., NSF grant IBN-9208765 to M.H.D., and a David and Lucille Packard Foundation Fellow- ship to M.H.D.
References
Bergmann-Erb D, Heide G (1990) Kontraktionsmodus direkter Flugsteuermuskeln yon Calliphora. Thieme, Stuttgart
Dickinson MH (1994) The effects of wing rotation on unsteady aerodynamic performance at low Reynolds numbers. J Exp Biol 192:179 206
Dickinson MH, G6tz KG (1993) Unsteady aerodynamic perfor- mance of model wings at low Reynolds numbers. J Exp Biol 174: 45 64
Dickinson MH, Lehmann F-O, G6tz KG (1993) The active control of wing rotation by Drosophila. J Exp Biol 182:173 189
Egelhaaf M (1989) Dynamic properties of two control systems un- derlying visually guided turning in house-flies. J Comp Physiol A 161:777 783
Ellington CP (1984) The aerodynamics of hovering insect flight III. Kinematics. Phil Trans R Soc Lond B 305:41 78
Ennos AR (1989) The kinematics and aerodynamics of the free flight of some Diptera. J Exp Biol 142:49 85
Ewing AW (1979a) The neuromuscular basis of courtship song in Drosophila: the role of direct and axillary wing muscles. J Comp Physiol 130:87-93
Ewing AW (1979b) The role of feedback during singing and flight in Drosophila melano.qaster. Physiol Entomol 4:329-337
Fayyazuddin A, Chan WP, Jordan CE, Dickinson MH (1994) The role of haltere afferents in the activity of a steering muscle in the blowfly, Calliphora vicina. Soc Neurosci Abstr 20: 1595
G6tz KG (1968) Flight control in Drosophila by visual perception of motion. Kybernetik 4:199 208
G6tz KG (1983) Bewegungssehen und Flugsteuerung bei der Fliege Drosophila. In: Nachtigall W (ed) BIONA- report 2. Fischer, Stuttgart, pp 21 34
G6tz KG (1987) Course-control, metabolism and wing interference during ultralong tethered flight in Drosophila melanogaster. J Exp Biol 128:35-46
Grodinsky DL, Morozov PP (1993) Vortex formation during tethered flight of functionally and morphologically two-winged insects, including evolutionary considerations on insect flight. J Exp Biol 182:11-40
Heide G (1968) Flugsteuerung durch nicht-fibrill~re Flugmuskeln bei der Schmeil3fliege Calliphora. Z Vergl Physiol 59:456460
Heide G (1971a) Die Funktion der nicht-fibrillS_ren Flugmuskeln bei der Schmeil3fliege Calliphora. Teil I: Lage, Insertionsstellen und Innervierungsmuster der Muskeln. Zool Jb Physiol 76:87 98
Heide G (1971b) Die Funktion der nicht-fibrill~iren Flugmuskeln bei der Schmeif3fliege Calliphora. Teil II: Muskul~ire Mechanismen der Flugsteuerung und ihre nerv6se Kontrolle. Zool Jb Physiol 76:99 137
Heide G (1975) Properties of a motor output system involved in the optomotor response in flies. Biol Cybern 20:99 112
Heide G (1983) Neural mechanisms of flight contol in Diptera. In: Nachtigall W (ed) B1ONA-report 2. Fischer, Stuttgart, pp 35-52
Heisenberg M, Wolf R (1979) On the fine structure of yaw torque in visual flight orientation of Drosophila melanogaster. J Comp Physiol 130:113 130
Heisenberg M, Wolf R (1984) Vision in Drosophila. Springer, Berlin Hengstenberg R, Sandeman DC, Hengstenberg B (1986) Compensa-
tory head roll in the blowfly Calliphora during flight. Proc R Soc Lond B 227:455-482
Hollick FSJ (1940) The flight of the dipterous fly Muscina sabulans Fall6n. Phil Trans R Soc Lond B 230:357-390
Kutsch W, Hug W (1981) Dipteran flight motor pattern: Invariabili- ties and changes during postlarval development. J Neurobio112: 1 14
Land MF, Collett TS (1974) Chasing behaviour of houseflies (Fannia canicularis), J Comp Physiol 89:331-357
Lehmann F-O (1990) Flugsteuerung bei der Fliege Drosophila: Elek- trische Erregung des Sternobasalarmuskels in Abh~ngigkeit von der Flfigelschlagphase. Diplomarbeit Universit~it Tiibingen
Miyan JA, Ewing AW (1984) A wing synchronous receptor for the dipteran flight motor. J Insect Physiol 30:567 574
Miyan JA, Ewing AW (1985a) How Diptera move their wings: a re-examination of the wing base articulation and muscle systems concerned with flight. Phil Trans R Soc Lond B 311: 271-302
Miyan JA, Ewing AW (1985b) Is the 'click' mechanism of dipteran flight an artifact of CCh anaesthesia? J Exp Biol 116:313-322
Miyan JA, Ewing AW (1988) Further observations on dipteran flight: details of the mechanism. J Exp Biol 136:229-241
Nachtigall W (1966) Die Kinematik der Schlagfliigelbewegungen von Dipteren. Methodische und analytische Grundlagen zur Biophysik des Insektenflugs. Z Vergl Physiol 52:155 211
Nachtigall W (1981) Insect flight aerodynamics. In: Herreid CF II, Fourtner CR (eds) Locomotion and energetics in arthropods. Plenum, New York, pp 127 162
Nachtigall W, Roth W (1983) Correlations between stationary measurable parameters of wing movement and aerodynamic force production in the blowfly (Calliphora vicina R.-D.). J Comp Physiol 150:251 260
Nachtigall W, Wilson DM (1967) Neuro-muscular control of dip- teran flight. J Exp Biol 47:77 97
Nalbach G (1989) The gear change mechanism of the blowfly (Calli- phora erythrocephala) in tethered flight. J Comp Physiol A 165: 321-331
Pfau HK (1987) Critical comments on a 'novel' mechanical model of dipteran flight' (Miyan & Ewing, 1958). J Exp Biol 128:463468
Tu MS, Dickinson MH (1994) Modulation of negative work output from a steering muscle of the blowfly Calliphora vicina. J Exp Biol 192:207-224
Wagner H (1986a) Flight performance and visual control of flight of the free-flying housefly (Musca domestica L.) I. Organization of the flight motor. Phil Trans R Soc Lond B 312:527 551

830 M.S. Tu, M. H. Dickinson: Motor control of wing kinematics in blowflies
Wagner H (1986b) Flight performance and visual control of flight of the free-flying housefly (Musca domestica L.) II. Pursuit of targerts. Phil Trans R Soc Lond B 312:553-579
Wagner H (1986c) Flight performance and visual control of flight of the free-flying housefly (Musca domestica L.) III. Interactions between angular movement induced by wide- and smallfield stimuli. Phil Trans R Soc Lond B 312:581 595
Wisser A (1988) Wing beat of Calliphora erythrocephala: turning axis and gearbox of the wing base (Insecta, Diptera). Zoomorpho- logy 107:359-369
Wisser A, Nachtigall W (1984) Functional-morphological investiga- tion on the flight muscles and their insertion points in the
blowfly Calliphora erythrocephala (Insecta, Diptera). Zoomor- phology 104:188-195
Wood J (1970) A study of the instantaneous air velocities in a plane behind the wings of certain Diptera flying in a wind tunnel. J Exp Biol 52:17 25
Zanker JM (1990a) The wing beat of Drosophila melanogaster I. Kinematics. Phil Trans R Soc Lond B 327:1 18
Zanker JM (1990b) The wing beat of Drosophila melanogaster III. Control. Phil Trans R Soc Lond B 327:45-64
Zanker JM, G6tz KG (1990) The wing beat of Drosophila melano- gaster II. Dynamics. Phil Trans R Soc Lond B 327:1944





























