Embed Size (px)
Citation preview

462
Biochimica et Biophysics Actu, 620 (1980) 462-471 @ ElsevierlNorth-Holland Biomedical Press
BBA 57686
THE CONTRIBUTION OF SERUM PHOSPHATIDYLCHOLINE AND LYSOPHOSPHATIDYLCHOLINE TO LYMPH PHOSPHATIDYLCHOLINE
MARCIA STEERE and CHARLES M. MANSBACH, II
Veterans Administration Medical Center, Division of Gastroenterology, Department of Medicine, Duke University Medical Center, Durham, NC 27705 (U.S.A.)
(Received January 16th, 1980) (Revised manuscript received April lath, 1980)
Key words: PkosphutidyZckoIine; Lysophospkat~dyZcholine; Lipoprotein; Chylomicron; @at, Lympk~
Summary
1. The contribution of serum phosphatidylcholine and l-acyl lysophosphati- dylcholine to chylomicron and mesenteric lymph lipoproteins of heavier buoyant density was studied in rats with catheters placed in the jugular vein, duodenum, common bile duct and mesenteric lymph duct. The effect of includ- ing 10 mM phosphatidylcholine in the triolein emulsion infused into the duo- denum was also studied.
2. The intravenous infusion of phosphatidylcholine did not affect delivery of phosphatidylcholine into the lymph when phosphatidylcholine was included in the duodenal infusion. However, on intravenous lysophosphatidyl~holine infu- sion, phosphatidylcholine transport into the lymph was increased both in chylomicrons and the other lipoproteins found in the lymph when phosphati- dylcholine was included in the duodenal infusion.
3. The incorporation of serum phosphatidylcholine into chylomicron phos- phatidylcholine was minimal and decreased further by intraduoden~ phos- phatidylcholine infusion. Incorporation into the heavier lymph lipoproteins was <20X
4. The incorporation of serum lysophosphatidylcholine into chylomicrons was 17% at 4-6 h of infusion which was decreased by intraduoden~ phosphati- dylcholine as was the incorporation into the heavier lipoproteins of lymph.
5. It is concluded that serum phosphatidylcholine is a poor precursor of chylomicron phosphatidylcholine and that while lysophosphatidylcholine is a
Abbreviations: VLDL, very low density lipoprotein (d < 1.006); LDL, low density lipoprotein (1.006 < d <1.0-/O).

somewhat better precursor, its contribution to chylomicron phosphatidylcho- line is limited by its serum concentration.
Introduction
Interest in intestinal phosphatidylcholine metabolism stems from the im- portance of phosphatidylcholine not only as the major phospholipid of cell membranes 111 but also because of its importance as a constituent of lipopro- teins [ 21. For intestinal lipid transport, the major lipoprotein of interest is the chylomicron where phosphatidylcholine forms a minor part of the whole mole- cule but is a major component of its surface ‘membrane’ [3]. The origin of chylomicron phosphatidylcholine is important since its synthesis may limit tria- cylglycerol transport from the intestine under certain conditions [4,5].
In the bile duct cannulated rat, when excess phosphatidylcholine is provided in a triacylglycerol infusion, essentially all the chylomicron phosphatidylcho- line can be shown to be derived from absorbed phosphatidylcholine [6]. By contrast, when no phosphatidylcholine is included in the triacylglycerol infu- sion, chylomicron phosphatidylcholine is provided by de novo synthesis [6]. However, it was apparent from these studies [6] that the phosphatidylcholine associated with lipoproteins, whose buoyant densities were heavier than chylo- microns, was only in part derived from the sum of de novo synthesis and exo- genous phosphatidylcholine suggesting that considerable phosphatidylcholine was derived from other sources. An obvious possible extraintestinal reservoir for phosphatidylcholine is serum phosphatidylcholine and lysophosphatidyl- choline. This represents a large pool with an extraintestinal source for replenishment (liver) [ 71. Additionally, prior investigations have shown that both serum phosphatidylcholine and especially lysophosphatidylcholine can be delivered to the intestine [8]. Therefore, the present studies were undertaken to define the contribution of serum phosphatidylcholine and lysophosphatidyl- choline toward lymph phosphatidylcholine both in chylomicrons and the heavier lipoproteins. Additionally, the effect of including 10 mM phosphatidyl- choline in the triacylglycerol infusion on this contribution was investigated since this concentration of phosphatidylcholine in the duodenal infusion has been recently shown to profoundly affect de novo phosphatidylcholine syn- thesis and to alter intestinal microsomal composition and permeability [ 91.
Materials and Methods
Animal preparation. Male Sprague-Dawley rats, weighing 343 + 11 g were given 4 cannulae on the day prior to an experiment. PE 50 (Clay Adams Co., Parsippany, NJ) cannulae were placed in the right jugular vein, the mesenteric lymph duct [6] and the duodenum [6]. A PE 10 cannula was placed in the common bile duct proximal to the entry of the pancreatic duct. The rat was infused overnight with 0.15 M NaCl at 3.0 ml/h through the duodenal cannula and 0.2 ml/h through the jugular vein cannula using 2 constant infusion pumps (Harvard Apparatus Co., Millis, MA). Bile and lymph were allowed to drain freely. No food was offered. The next day, an infusion was begun through the

464
jugular vein cannula of [3ZP]phosphatidylcholine (1.6-3.2 - lo5 CPM, l-4 ~mol/ml) or lyso[32P]phosphatidylcholine (1.2-4.0 - 10’ CPM, l-4 pmol/ml) at 0.3 ml/h. One hour later, a sonified triolein emulsion infusion (triolein (from Sigma Chemical Co., St. Louis, MO), 72 mM; taurocholate (from Calbiochem, San Diego, CA), 20 mM; gum acacia 100 g/l; Tris-HCl (pH 7.0), 100 mM) was begun through the duodenal cannula at 4.5 ml/h. Where indicated 10 mM phos- phatidylcholine was included in the infusion. Lymph was collected on ice with- out heparin in 2-h increments for 6 h after the start of the triolein infusion. The lymph was defibrinated with a wooden applicator stick and centrifuged at 3 * lo6 g - min at 4°C (Beckman Instruments, Inc., Palo Alto, CA, model L2- 65B) using a SW50 rotor. The chylomicrons were removed using a tube slicing technique and the remainder of the lymph (the infranatant) collected sepa- rately. The experiment was terminated by an overdose of pentobarbital. When anesthesia was induced, heart blood was obtained.
In certain experiments, plasma and lymph were further fractionated into more specific lipoprotein components. To float the very low density lipopro- tein (VLDL) fraction, plasma or lymph was centrifuged for 2.3 - lo8 g * min at a density of 1.006. For low density lipoprotein (LDL), the samples were adjusted to a density of 1.070 with KBr and centrifuged at 2.3 - lo8 g - min. LDL, as was VLDL, was obtained by a tube slicing technique. The pellet from the d = 1.070 centrifugation was taken as ‘high density lipoprotein’ (HDL).
In other experiments, at the conclusion of lymph collection, the rat was sacrificed by an overdose of pentobarbital and the intestine removed to an iced glass plate. The mucosal surface was cleansed and the mucosa removed as pre- viously described [lo]. Its lipid content was extracted by the method of Folch et al. [ll].
Su bstru te preparations. [ 32P]Phosphatidylcholine was prepared by inserting common bile duct and duodenal cannulae into a rat. 5 mCi 32Pi was infused through the duodenal cannula in 1 h followed by an infusion of 1% tauro- cholate. Bile was collected for 24 h and extracted [ 111. The organic phase was collected and the phosphatidylcholine fraction isolated by silicic acid column chromatography [ 61. 1-Acyl lyso[ 32P]phosphatidylcholine was prepared from the 32P-labeled bile phosphatidylcholine by reaction with Nujc nuja venom (Sig- ma Chemical Co., St. Louis, MO) as previously described [lo]. The phosphati- dylcholine or lysophosphatidylcholine was prepared for intravenous infusion by sonification (Heat Systems-Ionics, Plain View, NY) for 1.5 min twice in 0.15 M NaCl in an ice bath. Samples of infusate were obtained in triplicate for determination of specific activity. Egg phosphatidylcholine was used for the intraduodenal perfusions. It was prepared as previously described [ 61. Triolein was washed repeatedly with ethanol to remove fatty acids. It was 96% pure.
Analytical procedures. Extracted lipids were dissolved in 0.5 ml chloroform/ methanol (2 : 1, v/v). Phosphatidylcholine and lysophosphatidylcholine were separated from other phospholipids by thin-layer chromatography (TLC) using the solvent system chloroform/methanol/NH,OH/H,O (150 : 75 : 9 : 3, v/v). The spots were identified by exposure to I2 vapor with the aid of authentic standards. Phosphatidylcholine was quantitated by a slight modification of the method of Marinetti [12] and radioactivity determined using the scintillators and solvent system previously described [6]. Blood volume was assumed to be 64 ml/kg [ 131.

Results
Intravenous phosphatidylcholine infusion The serum concentration of phosphatidylcholine was 0.51 f 0.16 mM and
had a specific activity of 11300 + 2254 dpm/pmol when [32P]phosphatidyl- choline was infused intravenously. On lipoprotein fractionation, 66% of the phosphatidylcholine radioactivity was found in the LDL fraction which also had the highest specific activity of all lipoprotein fractions (VLDL 9190 dpm/ pmol, LDL 15 806 dpm/l.tmol, HDL 5293 dpm/pmol, average of two determi- nations).
Lymph chylomicron phosphatidylcholine outputs for each 2-h period were similar to that previously reported [6] and were not materially affected by including 10 mM phosphatidylcholine in the triacylglycerol infusion (Table I). When the lymph infranatant was separated into different lipoprotein fractions, 65% of the radioactivity was in LDL, similar to what had been found in the plasma. However, the lymph differed considerably from the plasma in that no radioactivity was detected in the VLDL fraction. The specific activity for the LDL fraction in lymph was 1570 dpm/pmol and HDL 982 dpm/Ccmol.
The percentage of lymph phosphatidylcholine that was derived from serum phosphatidylcholine can be calculated by comparing the specific activity of serum phosphatidylcholine to that found in the lymph. As shown in Table II
TABLE I
PHOSPHATIDYLCHOLINE CONCENTRATION IN LYMPH AFTER THE INTRAVENOUS INFUSION
OF PHOSPHATIDYLCHOLINE (A) OR LYSOPHOSPHATIDYLCHOLINE (B)
Mesenteric lymph duct and duodenal fistulated rats were infused intravenously with phosphatidylcholine (A) or lysophosphatidylcholine (B) as indicated and intraduodenaIIy with a triolein emulsion with or without 10 mM phosphatidylcholine as shown. Lymph was collected and the chylomicron and lymph infranatant fraction isolated. The data are the means (2 1 S.E.) of four rats. + P < 0.03; * P < 0.02; 5 I’ < 0.01 (Student’s t-test for unpaired data comparing rats infused with or without 10 mM phosphatidylcho- line).
Duodenal infusion Phosphatidylcholine (mM) after infusion for
A
O-2 h
Chylomicrons
2-4 h 4+?h
0 mM phosphatidylcholine 10 mM phosphatidylcholine
0.55 * 0.20 2.62 k 0.61 2.45 * 0.81 1.58 f 0.57 2.62 + 0.78 2.47 ?I 0.82
Lymph infranatant
0 mM phosphatidylcholine 0.14 f 0.06 0.15 f 0.06 0.15 C 0.06 10 mM phosphatidylcholine 0.16 f 0.02 0.17 f 0.03 0.16 ? 0.03
B Chylomicrons
0 mM phosphatidylcholine 10 mM phosphatidylcholine
0.96 k 0.45 1.92 f 0.55 1.92 f 0.55 3.10 + 0.40 + 6.57 f 0.76 s 6.88 + 0.36 0
Lymph infranatant
0 mM phosphatidylcholine 0.16 + 0.07 0.23 ? 0.08 0.18 f 0.07 10 mM phosphatidylcholine 0.46 f 0.05 * 0.55 + 0.06 + 0.55 k 0.08 *
466
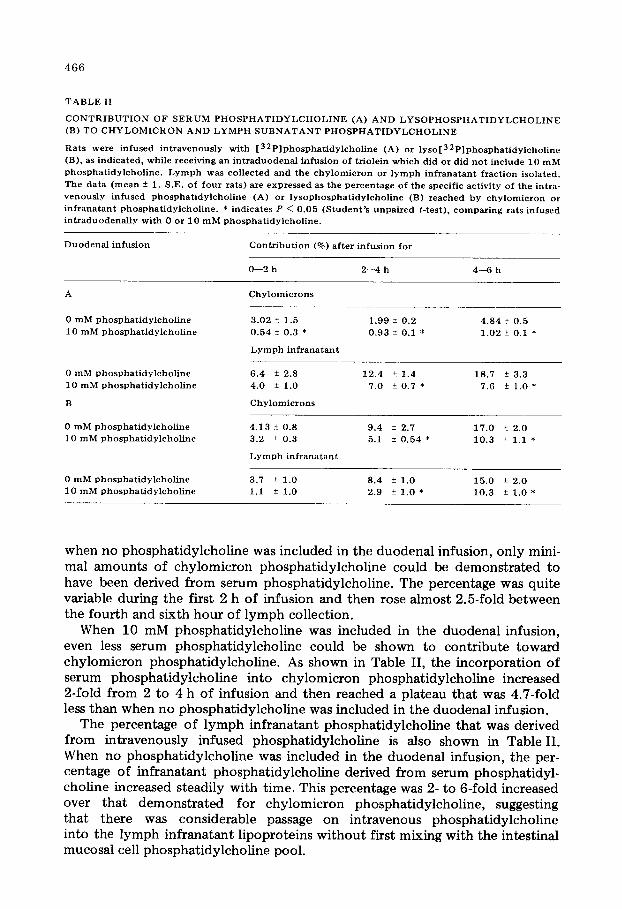
TABLE II
CONTRIBUTION OF SERUM PHOSPHATIDYLCHOLINE (A) AND LYSOPHOSPHATIDYLCHOLINE (B) TO CHYLOMICRON AND LYMPH SUBNATANT PHOSPHATIDYLCHOLINE
Rats were infused intravenously with [ 32Plphosphatidylcholine (A) or lyso[ 32Plphosphatidylcholine (B). as indicated, while receiving an intraduodenal infusion of triolein which did or did not include 10 mM phosphatidylcholine. Lymph was collected and the chylomicron or lymph infranatant fraction isolated. The data (mean k 1. S.E. of four rats) are expressed as the percentage of the specific activity of the intra- venously infused phosphatidylcholine (A) or lysophosphatidylcholine (B) reached by chylomicron or infranatant phosphatidylcholine. * indicates P < 0.05 (Student’s unpaired t-test), comparing rats infused intraduodenally with 0 or 10 mM phosphatidylcholine.
Duodenal infusion
A
0 mM phosphatidylcholine 10 mM phosphatidylcholine
O-2 h 2-4 h 4-6h
Chylomicrons - -.__.. _.
3.02 + 1.5 1.99 + 0.2 4.84 i 0.5
0.54 + 0.3 * 0.93 i- 0.1 * 1.02 f 0.1 *
Lymph infranatant
0 mM phosphatidylcholine 6.4 k 2.8 10 mM phosphatidylcholine 4.0 k 1.0
B Chylomicrons
0 mM phosphatidylcholine 10 mM phosphatidylcholine
4.13 i- 0.8 3.2 + 0.3
Lymph infranatant
0 mM phosphatidylcholine 10 mM phosphatidylcholine
3.7 + 1.0 8.4 f 1.0 15.0 * 2.0
1.1 + 1.0 2.9 t 1.0 * 10.3 + 1.0 *
Contribution (o/o) after infusion for
12.4 f 1.4 18.7 + 3.3 1.0 * 0.7 * 7.6 k 1.0 *
9.4 + 2.7 17.0 ? 2.0 5.1 i 0.54 * 10.3 f 1.1 *
when no phosphatidylcholine was included in the duodenal infusion, only mini- mal amounts of chylomicron phosphatidylcholine could be demonstrated to have been derived from serum phosphatidylcholine. The percentage was quite variable during the first 2 h of infusion and then rose almost 2.5-fold between the fourth and sixth hour of lymph collection.
When 10 mM phosphatidylcholine was included in the duodenal infusion, even less serum phosphatidylcholine could be shown to contribute toward chylomicron phosphatidylcholine. As shown in Table II, the incorporation of serum phosphatidylcholine into chylomicron phosphatidylcholine increased 2-fold from 2 to 4 h of infusion and then reached a plateau that was 4.7-fold less than when no phosphatidylcholine was included in the duodenal infusion.
The percentage of lymph infranatant phosphatidylcholine that was derived from intravenously infused phosphatidylcholine is also shown in Table II. When no phosphatidylcholine was included in the duodenal infusion, the per- centage of infranatant phosphatidylcholine derived from serum phosphatidyl- choline increased steadily with time. This percentage was 2- to 6-fold increased over that demonstrated for chylomicron phosphatidylcholine, suggesting that there was considerable passage on intravenous phosphatidylcholine into the lymph infranatant lipoproteins without first mixing with the intestinal mucosal cell phosphatidylcholine pool.

46’7
When 10 mM phosphatidylcholine was included in the duodenal infusate, serum phosphatidylcholine contributed less phosphatidylcholine to the lymph infranatant lipoproteins than when no phosphatidylcholine was included in the duodenal infusion. This reduction was significant at all time periods except for the O-2-h period in the lymph infranatant.
The quantity of phosphatidylcholine in the chylomicrons calculated to be derived from serum phosphatidylcholine at 4-6 h of infusion, represents only 0.9% of the total circulating phosphatidylcholine pool which is reduced to 0.3% when phosphatidylcholine was included in the duodenal perfusion. For the lymph infranatant this represents 0.3 and 0.1% of the circulating phospha- tidylcholine pool, respectively.
Mucosal specific activity was nearly the same as the averaged chylomicron specific activity (N = 3) 112 t 34 dpm/pmol vs. 103 f 29 dpm/pmol, respec- tively. This suggests that the phosphatidylcholine utilized for chylomicron phosphatidylcholine was well admixed with the total mucosal phosphatidylcho- line pool.
Intravenous lysophosphatidylcholine infusion When lys~[~~P]phosphatidylcholine was infused intravenously and no phos-
phatidylcholine infused intraduodenally, the serum phosphatidylcholine con- centration was 0.49 + 0.12 mM and the lysophosphatidylcholine concentra- tion was 0.13 + 0.05 mM. Serum lysophosphatidylcholine specific activity was 2966 * 503 dpm/pmol and phosphatidylcholine specific activity was 237 + 51 dpm/l-lmol. On including 10 mM phosphatidylcholine in the duodenal infusion, the serum phosphatidylcholine concentration increased to 0.93 f 0.17 mM but the lysophosphatidylcholine concentration remained nearly the same, 0.18 f 0.02 mM. The specific activity of serum lysophosphatidylcholine on including phosphatidylcholine in the intraduodenal infusion was 564 f 33 dpm/E.tmol and the serum phosphatidylcholine specific activity was 126 f 22 dpm/pmol. As shown in Table I, on inclusion of phosphatidylcholine in the duodenal infusion, the phosphatidylcholine transport in chylomicrons and heavier lipoproteins was significantly increased at each time period.
Shown in Table II is the percentage of chylomicron and lymph infranatant phosphatidylcholine derived from serum lysophosphatidylcholine. This per- centage increased at each time period. In contrast to intravenous phosphatidyl- choline perfusion, the percentage of chylomicron phosphatidylcholine derived from intravenously infused lysophosphatidylcholine was considerably greater, 4-lo-fold at 4-6 h of perfusion. Also unlike the results obtained with intra- venous phosphatidylcholine, the percentage of chylomicron phosphatidylcho- line and lymph infranatant phosphatidylcholine derived from serum lysophos- phatidylcholine were nearly the same. The inclusion of 10 mM phosphatidyl- choline in the duodenal infusate substantially reduced the incorporation of radiolabel into both chylomicron phosphatidylcholine and lymph infranatant phosphatidylcholine at the later 2 h periods.
The percentage of the total circulating lysophosphatidylcholine pool calcu- lated to have been distributed to chylomicron phosphatidylcholine at 4-6 h of infusion was 13% when no phosphatidylcholine was included in the duo- denal perfusate and 6% when it was. In the lymph infranatant, 0.8 and 1.3%

468
respectively, of the lysophosphatidylcholine circulating pool was thus distri- buted.
Discussion
Potential sources for chylomicron phosphatidylcholine include de novo syn- thesis, utilization of absorbed dietary or biliary phosphatidylcholine, cellular membrane phosphatidylcholine and phosphatidylcholine from serum. It is clear from previous work that de novo synthesis and intestinal luminal phosphatidyl- choline provide the majority of chylomicron phosphatidylcholine [ 61. Never- theless, serum’ is potentially a considerable source of phosphatidylcholine for chylomicron phosphatidylcholine and therefore its contribution is worth deter- mining. Lysophosphatidylcholine, whose concentration in serum is much smaller than phosphatidylcholine, has previously been demonstrated to be an intestinal phosphatidylcholine precursor [S] _
The potential contribution of serum phosphatidylcholine and lysophosphati- dylcholine to chylomicron phosphatidylcholine, is limited by the fact that of the amount delivered to the intestine least would be distributed to the mature mucosaI epithelial cells, the site of chylomicron formation [lo]. For example, serum fatty acids were chiefly found to be taken up by the cryptal cells of the intestine rather than the cells at the villus tip [ 141. One cause for this distribu- tion is the counter current nature of blood flow in the intestine [ 151. Another, but unexplored cause, could be a difference in penetrability of the cellular membrane of cryptal cells vs. mature enterocytes by ampipathic lipids as sug- gested by alterations in the enzymic constituents of plasma membrane vesicles from cryptal and villus cells [ 161.
Stein and Stein [ 81, on injecting [ 32P]phosphatidylcholine intravenously, found only a small (-4%) fraction to be taken up by the small intestine in 150 min with even less phosphatidyl~hol~e localized to the intestine at earlier time periods. This percentage of total injected phosphatidylcholine delivered to the intestine is very close to the percentage contribution of serum phosphatidyl- choline to chylomicron phosphatidylcholine as shown in the present study even though a constant infusion of .phosphatidyl~hol~e was given rather than a bolus injection.
It is possible that the physicochemical form of the infused phosphatidylcho- line, which circulated with a buoyant density similar to LDL and presumably contained the lipid composition found in lipoprotein X [17], was an inappro- priate vehicle for the presen~tion of phosphatidylcholine to the intestine for chylomicron synthesis. This is unlikely because of three factors. Firstly, this method of phosphatidylcholine infusion has been shown to be effective in delivering phosphatidylcholine at least to the liver [ 181 where it is taken up in enough quantity to exert significant physiologic effects. Secondly, the injected radiolabeled phosphatidylcholine liposomes obviously exchanged phosphatidyl- choline with all lipoprotein classes in the serum. This phosphatidylcholine exchange in the vascular space has been suggested to be facilitated by a phos- pholipid exchange protein [ 191. Thus, in the present case, although LDL was most heavily labeled, all lipoprotein classes were, in fact, labeled and thus if lipoproteins other than LDL were large contributors to chylomicron phosphati-

469
dylcholine, more heavy labeling of chylomicron phosphatidylcholine would have been expected. That circulating lipoprotein lipids can contribute to lymph chylomicron lipid is demonstrated by cholesterol since nearly all chylomicron cholesterol has been shown to be derived from lipoprotein cholesterol in man and 30% in the rat [20]. Lastly, it is obvious that much larger quantities of labeled phosphatidylcholine exuded from the vascular space and were trans- ported in the lymph infranatant predominantly again in LDL than appeared in chylomicron phosphatidylcholine. This indicates that the intestine was exposed to considerably higher specific activity phosphatidylcholine than was in fact taken up and utilized for chylomicron formation.
The suitability of biliary phosphatidylcholine as a chylomicron phosphati- dylcholine precursor has been previously demonstrated [6]. In fact biliary phosphatidylcholine was found to be the preferred chylomicron phosphatidyl- choline precursor when compared to potato phosphatidylcholine whose acyl group composition more closely resembles chylomicron phosphatidylcholine [61.
When 10 mM phosphatidylcholine was included in the duodenal infusion, the contribution of serum phosphatidylcholine toward both chylomicron phos- phatidylcholine and the phosphatidylcholine of lipoproteins of heavier buoyant density was depressed. The most likely cause of this effect is the 2-fold increase in microsomal phosphatidylcholine content when 10 mM phosphatidylcholine is infused into the duodenum [9]. Any phosphatidylcholine entering the mucosal cell would have to traverse the microsomal membrane in order to be incorporated into chylomicrons. In doing so, it would be exposed to and pre- sumably would exchange with a phosphatidylcholine pool twice as large as in microsomes from rats with no phosphatidylcholine in their intraduodenal infu- sion. This would suggest an apparent 2-fold suppression in incorporation. Since a 4.7-fold suppression was found in the 4-6 h collection period, an increase in microsomal phosphatidylcholine pool size may not be the whole explanation. Another possibility is that intraduodenal phosphatidylcholine perfusion inhibits the intestinal cell permeability to phosphatidylcholine liposomes or that the phosphatidylcholine content of cellular membranes other than micro- somes is also increased. This latter possibility is not likely since the phosphati- dylcholine content of the whole mucosal cells is unaffected by intraduodenal phosphatidylcholine perfusion at the concentration employed [ 91.
Intravenous lyso [ 32P]phosphatidylcholine perfusion resulted in the radio- labeling of phosphatidylcholine within the vascular compartment as well. This could be due to the conversion of lysophosphatidylcholine to phosphatidylcho- line directly by a poorly characterized enzyme present in serum [21] or due to the entry of the lysophosphatidylcholine into liver with subsequent synthesis of phosphatidylcholine and its transport as lipoprotein phosphatidylcholine. By whatever method phosphatidylcholine was labeled, it is not likely to have significantly hindered the interpretation of the results which was based on the assumption that lysophosphatidylcholine is the predominant species labeled. This thesis is supported by the comparative specific activities in which it is evi- dent that lysophosphatidylcholine is 5-lo-fold more heavily labeled that phos- phatidylcholine in the serum and by the results on lysophosphatidylcholine perfusion which are significantly different than when phosphatidylcholine was infused.

470
Of particular interest is the finding that intravenous lysophosphatidylcholine caused significant changes in phosphatidylcholine metabolism. Phosphatidylcho- line concentrations in the serum, chylomicrons and lymph infranatant were all significantly elevated when 10 mM phosphatidylcholine was included in the duodenal perfusion as compared to when no phosphatidylcholine was present. This cannot be accounted for by the quantity of lysophosphatidylcholine infused and therefore must indicate either a facilitation of the passage into the lymph of phosphatidylcholine synthesized from absorbed lysophosphatidylcho- line or an enhanced conversion of absorbed lysophosphatidylcholine into phos- phatidylcholine and its subsequent transport into the lymph. The latter possi- bility is the more likely since phosphatidylcholine transport in the lymph is un- affected by the inclusion of phosphatidylcholine in the duodenal infusion in the absence of intravenously infused lysophosphatidylcholine.
The increase seen in serum phosphatidylcholine when phosphatidylcholine was included in the duodenal infusion is not readily understood. It is possible that not all lymph was drained from the intestine by the fistula and therefore that the greatly increased lymph phosphatidylcholine entered the vascular space.
It can be seen from these studies that both circulating phosphatidylcholine and lysophosphatidylcholine can be precursors of lymph infranatant phosphati- dylcholine. The actual contribution is inversely correlated with the presence of phosphatidylcholine in the intestinal lumen. In the absence of luminal phos- phatidylcholine, when intestinal microsomes are not loaded with phosphatidyl- choline [9], the sum of the contributions of circulating phosphatidylcholine and lysophosphatidylcholine to lymph infranatant phosphatidylcholine is 34% of the phosphatidylcholine present (4-6 h of perfusion). This is close to the 50% of lymph infranatant phosphatidylcholine which cannot be shown to be syn- thesized by the intestinal mucosa [ 61.
Acknowledgements
This work was supported in part by the Medical Research Service, Veterans Administration, and by the National Institutes of Health Research Grant AM 17526,
References
1 Nilsson, O.S. and DaIIner, G. (1977) Biochim. Biophys. Acta 464.453-458
2 Scmu, A.M. and Wisdom, C. (1972) Annu. Rev. Biochem. 41, 703-730
3 Zilversmit, D.B. (1965) J. CIin. Invest. 44.1610-1622
4 Sabesin. S.M., Holt. P.R. and Clark, S.B. (1975) Lipids 12. 840-846
5 Tso, P.. Lam, J. and Simmonds, W.J. (1978) Biochim. Biophys. Acta 528, 364-372
6 Mansbach. C.M., II (1977) J. CIin. Invest. 60.411420
7 Schumaker, V.N. and Adams, G.H. (1969) Annu. Rev. Biochem. 38,113-136
8 Stein. Y. and Stein, 0. (1966) Biochim. Biophys. Acta 116.95-107
9 Mansbach, C.M., II and Parthasarathy, S. (1979) J. Biol. Chem. 254,9688-9694
10 Mansbach, C.M.. II (1973) Biochim. Biophys. Acta 296. 386400
11 Folch, J., Less, M. and SloaneStanley, G.H. (1957) J. Biol. Chem. 226, 497-509
12 Marinetti, G.V. (1962) J. Lipid Res. 3, l-20
13 Altman. P. and Dittmer. D. (1974) Biology Data Book. 2nd edn.. Vol. 3, p. 1847. The Federation of
American Societies for Experimental Biology, Bethesda, MD

471
14 GangI. A. and Ockner, R.K. (19’75) J. CIin. Invest. 55.803-813
15 Lanciault, G. and Jacobson, E.D. (1976) Gastroenterology 71,851-873 16 Gratecos, G., Knibiehler, M., Benoit, V. and Semeriva. M. (1978) Biochim. Biophys. Acta 512, 508-
524
17 Manzato. E.. FelIini. R.. Baggio. G., Walch, S., Neubeck. W. and Seidel., D. (1976) J. Clin. Invest. 57, 1248-1260
18 Jakoi, L. and Quarfordt, S.H. (1974) J. Biol. Chem. 249. 5840-5844 19 Eisenberg, S. (1978) J. Lipid Res. 19, 229-236 20 Quintao, E.C.R.. Drewiacki, A., Stechhahn, K.. de Farina, E.C. and Sipohi, A.M. (1979) J. Lipid Res.
20.941--951 21 Subbaiah, P.V. and Bagdade, J.D. (1978) Life Sci. 22.1971-1978