Embed Size (px)
Citation preview

THE JOURNAL OF B~oraoorc~~ CHENIISTRY Vol. 238, No. 6, June 1963
Printed in U.S. A.
The Chemical Composition of Pneumococcal
C-Polysaccharide*
TEH-YUNG LIU AND EMIL C. GOTSCHLICH
From The Rockefeller Institute, New York 21, New York
(Received for publication, January 16, 1963)
The species-specific C-polysaccharide of the pneumococcus was first described by Tillett, Goebel, and Avery in 1930 (1). The chemical composition was examined by Heidelberger and Kendall (2) and by Goebel et al. (3) ; the latter study showed that the C-polysaccharide contained 4.47% phosphorus, 5.91% nitrogen, 50.6% reducing sugar, 21.8 % hexosamine, and 13.08% acetyl. Detailed information as to the chemical composition of the polysaccharide was not provided by these early studies, how- ever. It was felt that more thorough understanding of the chemistry of the C-polysaccharide might provide increased knowledge of the structure and metabolic activity of the pneumo- coccus, and might also shed light on the ability of this polysac- charide to combine with and precipitate the C-reactive protein which appears in human sera during infection.
The present report will describe investigations which have demonstrated the presence in the C-polysaccharide of amino sugars and amino acids. The chemical composition thus estab- lished indicates that the C-polysaccharide originates from the bacterial cell wall. One constituent of the polysaccharide, galactosamine 6-phosphate, has not been found before in bacterial cell walls.
EXPERIMENTAL PROCEDURE
Preparation of C-Polysaccharide-A method similar to the one described by Anderson and McCarty (4) was employed to pre- pare the C-polysaccharide from a strain of rough pneumococcus, designated R36A, which fails to produce type-specific capsular polysaccharide. The organisms were grown in 20-liter carboys containing 16 liters of filtered Todd-Hewitt medium. The broth was inoculated with 100 ml of a young culture and, after 6 hours of incubation at 37”, sterile 50% dextrose solution was added to a concentration of 2 $& dextrose to enhance growth. The culture was then maintained at pH 7.4 for 2 hours by three or four addi- tions of 2-g portions of autoclaved, solid sodium bicarbonate per liter of medium. The organisms were recovered by centrifuga- tion in an air-driven Sharples supercentrifuge. They were washed in the centrifuge with 0.9% NaCl solution. The bac- terial mass from each carboy was suspended in 250 ml of 0.9% NaCl solution and the organisms were lysed by adding 10% sodium deoxycholate solution to a concentration of 0.1%. The mixture was kept neutral to phenol red by the addition of 1 N
NaOH. After 15 minutes, 1 ml of 2 N acetic acid was added to each 100 ml of lysate, and the suspension was distributed into
* Supported in part by Grant RG-7256 from the United States Public Health Service and, from July 1,1962, by contract with the United States Army Medical Research and Development Com- mand, Office of the Surgeon General.
250-ml centrifuge bottles and placed in a 3-liter boiling water bath for 10 minutes. The heating caused the formation of a very voluminous precipitate which was separated from a clear yellowish supernatant by centrifugation. The precipitate, before being discarded, was washed with two 140-ml portions of 0.9% NaCl solution and the washings were added to the first supernatant. To the resulting supernatant was added 10% calcium chloride solution to a concentration of 1% CaC12, and absolute ethanol to a concentration of 20%. This procedure resulted in the precipitation of much of the remaining nucleic acids, which were removed by suction filtration through Celite 505. The concentration of alcohol in the filtrate was increased to 80% by the addition of absolute ethanol, and the precipitate that formed was recovered by centrifugation after refrigeration overnight. The precipitate was washed with 200-ml portions each of absolute ethanol, acetone, and ether and wss dried in a vacuum. This powder (approximately 2.5 g), which was only partially soluble, was suspended in water (200 ml) and was left standing overnight in the refrigerator to insure complete solution of the C-polysaccharide. The insoluble residue was discarded. After centrifugation, the supernatant was placed in a dialysis sac (Visking 24) and dialyzed for 48 hours against 4 liters of cold distilled water containing 20 ml of chloroform. The dialysis bag was then transferred to 2 liters of 0.01 M phosphate buffer, pH 7.4, containing 0.225% of NaCl and 0.1 To of MgCh.6HzO. After 6 hours at room temperature, 5 mg each of crystalline pancreatic ribonuclease and pancreatic deoxyribonuclease (Worthington Biochemical Corporation) were added to the contents of the dialysis bag and the dialysis was continued for 24 hours at 37”. A few milliliters of toluene and chloroform were added to prevent bacterial contamination. After 24 hours, 5 ml of thioglycolic acid neutralized to phenol red with N NaOH were added to the a-liter buffer bath, and a suspension containing 5 mg of crystalline streptococcal proteinase (5), kindly supplied by Dr. S. D. Elliott, was added to the contents of the dialysis sac. Incubation was continued for 24 hours at 37”. The ma- terial in the sac was treated by the method of Sevag (6) until no further protein was precipitated. Dialysis was then carried out against 4-liter volumes of cold distilled water changed twice a day for 3 to 4 days, and the material was lyophilized. About 500 mg of a white powder were obtained from 16 liters of original pneumococcal culture. Such preparations of C-polysaccharide contained less than 0.2% nucleic acid, as determined by measure- ment of ultraviolet absorption at 260 rnp, and reacted with antisera prepared against whole pneumococci, giving single lines of precipitation in double diffusion studies in agar. Before being subjected to the analytical procedures described below,
1928
by guest on April 1, 2019
http://ww
w.jbc.org/
Dow
nloaded from

June 1963 T.-Y. Liu and h’. C. Gotschlich 1929
the C-polysaccharide was desalted by gel filtration on a column, 4 X 50 cm, of Sephadex G-25 equilibrated with 50% acetic acid
(7). Analytical &‘ethocZs--Total nitrogen was determined by the
method of Kjeldahl. Total phosphorus in the C-polysaccharide was determined by the method of Fiske and SubbaRow (8) as modified by Umbreit, Burris, and Stauffer (9). The method of Chen, Toribara, and Warner (10) was used for the determina- tion of phosphorus in the samples isolated from the ion exchange column. The acetyl content was determined by the method of Elek and Harte (11).
Hexosamine was determined by a modification of the Elson- Morgan reaction described by Rondle and Morgan (12). Re- ducing sugars were determined by the calorimetric modification of the Hagedorn-Jensen method (13) described by Rapport, Meyer, and Linker (14). Methyl pentose, hexose, and heptose were determined by the procedure of Dische and Shettles (15). Glucose was determined by the method which employs the Glucostat reagent (Worthington Biochemical Corporation). Glucuronic acid was determined by the Dische calorimetric carbazole method (16).
Paper Chromatography--For the detection of amino acids, 40 mg of C-polysaccharide were hydrolyzed in 6 N HCl for 18 hours at 105” and the hydrolysate was evaporated to dryness in a vacuum. The amino acids were separated by descending chro- matography on Whatman No. 1 paper for 70 hours in l-butanol- pyridine-water (6:4:3 by volume). Papers were sprayed with ninhydrin solution (Sigma Chemical Company). Glutamic acid and alanine isolated from the hydrolysate of C-polysac- charide by ion exchange chromatography (see section below) were also subjected to the same chromatographic procedure to confirm their identities.
Glucosamine and galactosamine were identified by the ninhy- drin oxidation procedure (17) carried out on a sample of C-poly- saccharides that had been hydrolyzed with 6 N HCl for 18 hours at 105”. The resulting pentoses were detected by ascending chromatography on Whatman No. 4 paper in I-butanol-acetic acid-water (4: 1: 1 by volume). The papers were sprayed with aniline hydrogen phthalate reagent (18) to detect reducing sugars. Glucosamine and galactosamine isolated from the C-polysaccharide by the ion exchange column (see section below) were also subjected to this identification procedure.
For the detection of glucose, a sample of C-polysaccharide was hydrolyzed with 1 N HCl for 2 hours at 100” and evaporated to dryness in a vacuum. Descending chromatograms on What- man No. 1 paper were run in I-butanol-acetic acid-water (450: 50 : 125 by volume) for 28 hours. The papers were sprayed with aniline hydrogen phthalate reagent (18).
Ion Exchange Chromatography-The amino acid and amino sugar content of acid hydrolysates were determined by ion exchange chromatography with the automatic recording equip- ment described by Spackman, Stein, and Moore (19). A sample of C-polysaccharide (about 10 mg) was hydrolyzed at 110” with 3 ml of 6 N HCl for 5, 12, and 22 hours in an evacuated, deaerated, sealed glass tube. The hydrolysate was evaporated to dryness in a rotary evaporator at 40” and the residue was dissolved in 5 ml of the pH 2.2 buffer used with the amino acid analyzer. The dark solution was filtered through sintered glass, and 2-ml portions were added to the 15- and 150-cm columns of the amino acid analyzer. For the isolation and identification of the various ninhydrin-positive substances, a sample of C-
polysaccharide (100 mg) was hydrolyzed with 6 N HCl (3 ml) for 2s hours at 110”. The hydrolysate was evaporated to dry- ness and the residue was transferred to a column of IR-120, 0.9 x 150 cm, with two l.O-ml portions of deionized water. The column was eluted with citrate buffer, pH 3.25, at 30” for 300 fractions of 1.0 ml and then at 50” with citrate buffer, pH 4.25, for another 300 fractions of 1.0 ml each. Peaks were 1 ocated by ninhydrin analysis of 100~~1 aliquots of each fraction. Individual components isolated in this manner were desalted by a procedure similar to the one of Dreze, Moore, and Bigwood (20) and were found to be free of other ninhydrin-positive ma- terial when rechromatographed. Galactosamine B-phosphate crystallized as the acetic acid was removed under vacuum. It was recrystallized from water (cf. (21)). The amino acid analyzer constant was 26.1 on an instrument for which the aspartic acid constant was 24.8. The glucosamine hydrochloride isolated exhibited [ar]:’ +73.7” (equilibrium value; c = 0.705 in water). Authentic n-glucosamine hydrochloride (Mann Research Laboratories, Inc.) exhibited [cu]:’ +71.2” (equilib- rium value; c = 1.069 in water). The isolated galactosamine hydrochloride and the standard sample (IMann Research Labora- tories, Inc.) exhibited [c$,’ +95.2” (equilibrium value; c = 0.352 in 0.1 N HCl) and [a]:’ Jr95.5” (equilibrium value; c = 0.450 in 0.1 N HCl), respectively.
Conjiguration of Amino Acids-Hog kidney n-amino acid oxidase, prepared by the method of IMassey, Palmer, and Vennett (22), was used to determine the quantity of n-amino acids present. The enzyme was diluted with crystalline ovalbu- min and lyophilized. The activity of the enzyme was about 3 units per mg. Salt-free preparations of alanine, glutamic acid, and serine (0.1 to 1.0 pmole), isolated from C-polysaccharide by the procedures described above, were treated with n-amino acid oxidase (1 mg in 0.5 ml of 0.05 M pyrophosphate buffer, pH 8.0) for 20 hours at room temperature, and the resulting mixture was placed on the 50-cm column of the amino acid analyzer (23) to determine by difference the quantity of each amino acid that had been decomposed by the enzyme. Control experiments with known quantities (1 pmole) of nn-glutamic acid, nn-serine, and nn-alanine were also carried out.
Periodate Oxidation cf Galactosamine 6-Phosphate-Oxidation of authentic and isolated samples (1 to 3 mg) of galactosamine 6-phosphate were carried out by the procedure of Distler, Merrick, and Roseman (21). Glycolaldehyde phosphate was identified by chromatography on a column, 0.9 X 10 cm, of Dowex l-X8, Cl- form (24). The effluent fractions were analyzed for glycolaldehyde (25) and for organic phosphate (10). Both samples yielded a single peak immediately after the change of eluent from 0.025 N HCl to 0.05 N HCl. The absorption spectrum of the green color developed with diphenylamine was determined between 480 rnp and 680 rnp on a Zeiss PlMQ II spectrophotometer, and found to be the same as that reported for glycolaldehyde (25).
RESULTS
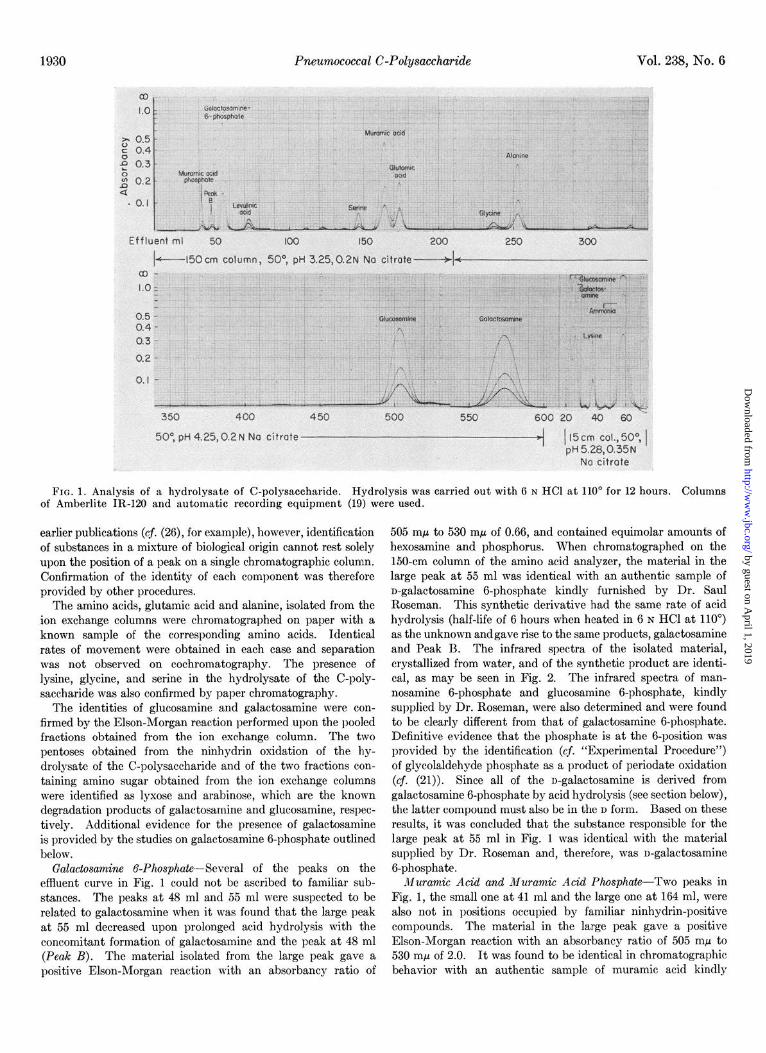
Ninhydrin-positive Substances of C-Polysaccharide-The ef- fluent curve obtained when a 12-hour acid hydrolysate of the C-polysaccharide was analyzed on the amino acid analyzer is shown in Fig. 1. Seven of the peaks correspond in position to those given by known substances; five to the amino acids, serine, glutamic acid, glycine, alanine, and lysine; and two to the amino sugars. glucosamine and galactosamine. As has been noted in
by guest on April 1, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Pneumococcal C-Polysaccharide Vol. 238, No. 6
FIG. 1. Analysis of a hydrolysate of C-polysaccharide. Hydrolysis was carried out with 6 N HCl at 110” for 12 hours. Columns of Amberlite IR-120 and automatic recording equipment (19) were used.
earlier publications (cj. (26), for example), however, identification of substances in a mixture of biological origin cannot rest solely upon the position of a peak on a single chromatographic column. Confirmation of the identity of each component was therefore provided by other procedures.
The amino acids, glutamic acid and alanine, isolated from the ion exchange columns were chromatographed on paper with a known sample of the corresponding amino acids. Identical rates of movement were obtained in each case and separation was not observed on cochromatography. The presence of lysine, glycine, and serine in the hydrolysate of the C-poly- saccharide was also confirmed by paper chromatography.
The identities of glucosamine and galactosamine were con- firmed by the Elson-Morgan reaction performed upon the pooled fractions obtained from the ion exchange column. The two pentoses obtained from the ninhydrin oxidation of the hy- drolysate of the C-polysaccharide and of the two fractions con- taining amino sugar obtained from the ion exchange columns were identified as lyxose and arabinose, which are the known degradation products of galactosamine and glucosamine, respec- tively. Additional evidence for the presence of galactosamine is provided by the studies on galactosamine 6-phosphate outlined below.
Galactosamine 6-Phosphate--Several of the peaks on the effluent curve in Fig. 1 could not be ascribed to familiar sub- stances. The peaks at 48 ml and 55 ml were suspected to be related to galactosamine when it was found that the large peak at 55 ml decreased upon prolonged acid hydrolysis with the concomitant formation of galactosamine and the peak at 48 ml (Peak B). The material isolated from the large peak gave a positive Elson-Morgan reaction with an absorbancy ratio of
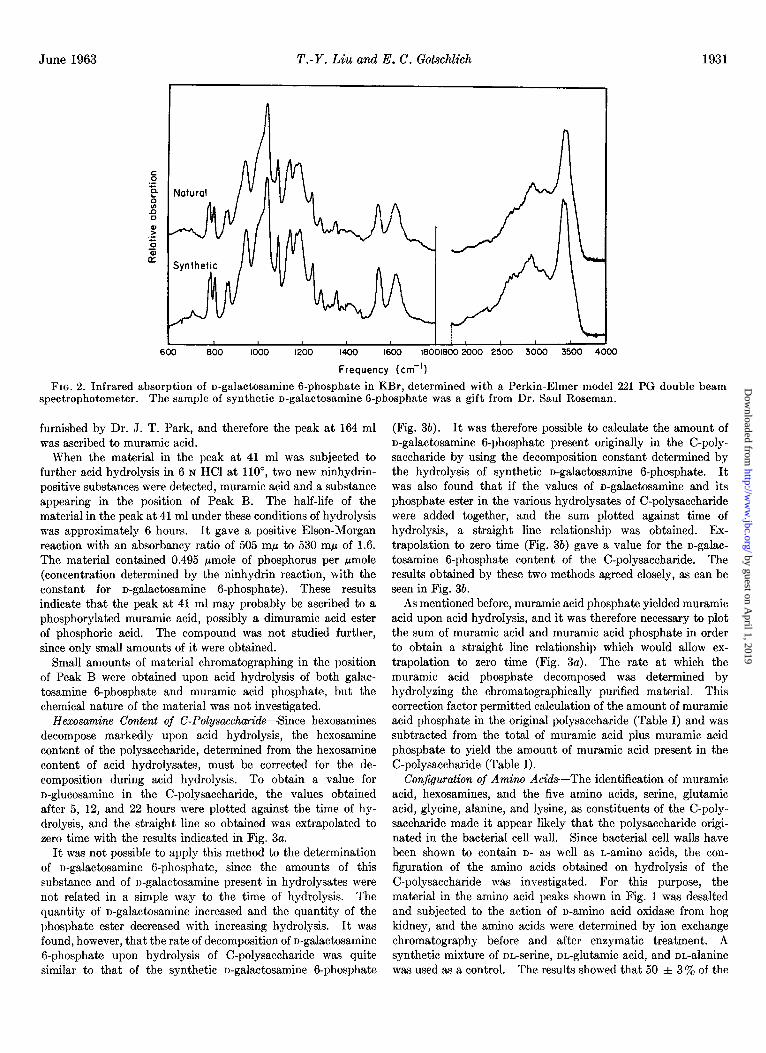
505 rnp to 530 rnp of 0.66, and contained equimolar amounts of hexosamine and phosphorus. When chromatographed on the 150-cm column of the amino acid analyzer, the material in the large peak at 55 ml was identical with an authentic sample of n-galactosamine 6-phosphate kindly furnished by Dr. Saul Roseman. This synthetic derivative had the same rate of acid hydrolysis (half-life of 6 hours when heated in 6 N HCl at 110’) as the unknown andgave rise to the same products, galactosamine and Peak B. The infrared spectra of the isolated material, crystallized from water, and of the synthetic product are identi- cal, as may be seen in Fig. 2. The infrared spectra of man- nosamine 6-phosphate and glucosamine 6-phosphate, kindly supplied by Dr. Roseman, were also determined and were found to be clearly different from that of galactosamine 6-phosphate. Definitive evidence that the phosphate is at the 6-position was provided by the identification (cf. “Experimental Procedure”) of glycolaldehyde phosphate as a product of periodate oxidation (cf. (21)). Since all of the n-galactosamine is derived from galactosamine 6-phosphate by acid hydrolysis (see section below), the latter compound must also be in the D form. Based on these results, it was concluded that the substance responsible for the large peak at 55 ml in Fig. 1 was identical with the material supplied by Dr. Roseman and, therefore, was n-galactosamine 6-phosphate.
Muramic Acid and Muramic Acid Phosphate-Two peaks in Fig. 1, the small one at 41 ml and the large one at 164 ml, were also not in positions occupied by familiar ninhydrin-positive compounds. The material in the large peak gave a positive Elson-Morgan reaction with an absorbancy ratio of 505 rnp to 530 rnp of 2.0. It was found to be identical in chromatographic behavior with an authentic sample of muramic acid kindly
by guest on April 1, 2019
http://ww
w.jbc.org/
Dow
nloaded from
June 1963 T.-Y. Liu and E. C. Gokchlich 1931
600 800 1000 1200 1400 1600 160016002000 2500 3000 3500 4000
Frequency (cm-‘)
FIG. 2. Infrared absorption of u-galactosamine G-phosphate in KBr, determined with a Perkin-Elmer model 221 PG double beam spectrophotometer. The sample of synthetic n-galactosamine G-phosphate was a gift from Dr. Saul Roseman.
furnished by Dr. J. T. Park, and therefore the peak at 164 ml was ascribed to muramic acid.
When the material in the peak at 41 ml was subjected to further acid hydrolysis in 6 N HCl at 110”, two new ninhydrin- positive substances were detected, muramic acid and a substance appearing in the position of Peak B. The half-life of the material in the peak at 41 ml under these conditions of hydrolysis was approximately 6 hours. It gave a positive Elson-Morgan reaction with an absorbancy ratio of 505 rnp to 530 rnp of 1.6. The material contained 0.495 pmole of phosphorus per pmole (concentration determined by the ninhydrin reaction, with the constant for n-galactosamine g-phosphate). These results indicate that the peak at 41 ml may probably be ascribed to a phosphorylated muramic acid, possibly a dimuramic acid ester of phosphoric acid. The compound was not studied further, since only small amounts of it were obtained.
Small amounts of material chromatographing in the position of Peak B were obtained upon acid hydrolysis of both galac- tosamine 6-phosphate and muramic acid phosphate, but the chemical nature of the material was not investigated.
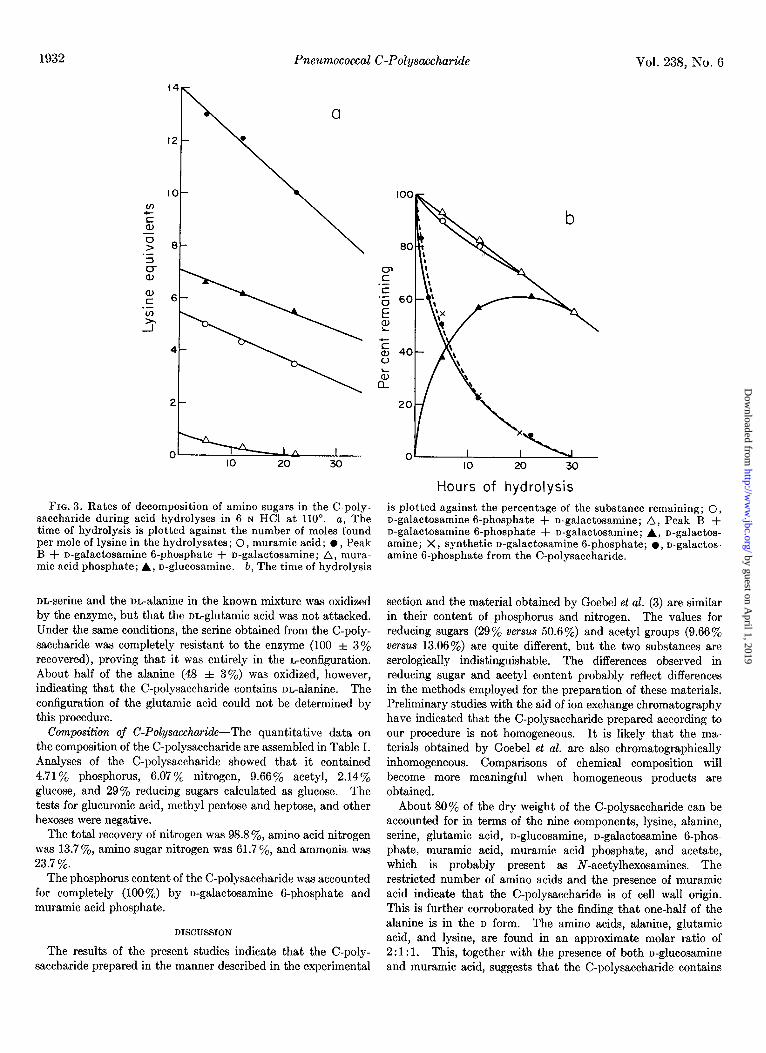
Hexosamine Content of C-Polysaccharide-Since hexosamines decompose markedly upon acid hydrolysis, the hexosamine content of the polysaccharide, determined from the hexosamine content of acid hydrolysates, must be corrected for the de- composition during acid hydrolysis. To obtain a value for n-glucosamine in the C-polysaccharide, the values obtained after 5, 12, and 22 hours were plotted against the time of hy- drolysis, and the straight line so obtained was extrapolated to zero time with the results indicated in Fig. 3a.
It was not possible to apply this method to the determination of n-galactosamine 6-phosphate, since the amounts of this substance and of n-galactosamine present in hydrolysates were not related in a simple way to the time of hydrolysis. The quantity of n-galactosamine increased and the quantity of the phosphate ester decreased with increasing hydrolysis. It was found, however, that the rate of decomposition of n-galactosamine 6phosphate upon hydrolysis of C-polysaccharide was quite similar to that of the synthetic n-galactosamine 6phosphate
(Fig. 3b). It was therefore possible to calculate the amount of n-galactosamine 6-phosphate present originally in the C-poly- saccharide by using the decomposition constant determined by the hydrolysis of synthetic n-galactosamine g-phosphate. It was also found that if the values of n-galactosamine and its phosphate ester in the various hydrolysates of C-polysaccharide were added together, and the sum plotted against time of hydrolysis, a straight line relationship was obtained. Ex- trapolation to zero time (Fig. 3b) gave a value for the n-galac- tosamine G-phosphate content of the C-polysaccharide. The results obtained by these two methods agreed closely, as can be seen in Fig. 3b.
As mentioned before, muramic acid phosphate yielded muramic acid upon acid hydrolysis, and it was therefore necessary to plot the sum of muramic acid and muramic acid phosphate in order to obtain a straight line relationship which would allow ex- trapolation to zero time (Fig. 3a). The rate at which the muramic acid phosphate decomposed was determined by hydrolyzing the chromatographically purified material. This correction factor permitted calculation of the amount of muramic acid phosphate in the original polysaccharide (Table I) and was subtracted from the total of muramic acid plus muramic acid phosphate to yield the amount of muramic acid present in the C-polysaccharide (Table I).
Conjiguraltin of Amino Acids--The identification of muramic acid, hexosamines, and the five amino acids, serine, glutamic acid, glycine, alanine, and lysine, as constituents of the C-poly- saccharide made it appear likely that the polysaccharide origi- nated in the bacterial cell wall. Since bacterial cell walls have been shown to contain D- as well as L-amino acids, the con- figuration of the amino acids obtained on hydrolysis of the C-polysaccharide was investigated. For this purpose, the material in the amino acid peaks shown in Fig. 1 was desalted and subjected to the action of n-amino acid oxidase from hog kidney, and the amino acids were determined by ion exchange chromatography before and after enzymatic treatment. A synthetic mixture of nn-serine, nn-glutamic acid, and nn-alanine was used as a control. The results showed that 50 f 3% of the
by guest on April 1, 2019
http://ww
w.jbc.org/
Dow
nloaded from
1932 Vol. 238, No. G
8-
6-
.
4-
2
0 1
Pneumococcal C-Polysaccharide
FIG. 3. Rates of decomposition of amino sugars in the C-poly- saccharide during acid hydrolyses in G N HCl at 110”. a, The time of hydrolysis is plotted against the number of moles found per mole of lysine in the hydrolysates ; 0, muramic acid; l , Peak B + n-galactosamine g-phosphate + n-galactosamine; A, mura- mic acid phosphate; A, n-glucosamine. b, The time of hydrolysis
nL-serine and the malanine in the known mixture was oxidized by the enzyme, but that the m-glutamic acid was not attacked. Under the same conditions, the serine obtained from the C-poly- saccharide was completely resistant to the enzyme (100 f 3% recovered), proving that it was entirely in the L-configuration. About half of the alanine (48 f 3%) was oxidized, however, indicating that the C-polysaccharide contains nL-alanine. The configuration of the glutamic acid could not be determined by this procedure.
Composition of C-Polysaccharide--The quantitative data on the composition of the C-polysaccharide are assembled in Table I. Analyses of the C-polysaccharide showed that it contained 4.71 y. phosphorus, 6.07% nitrogen, S.SSoil, acetyl, 2.14% glucose, and 29% reducing sugars calculated as glucose. The tests for glucuronic acid, methyl pentose and heptose, and other hexoses were negative.
The total recovery of nitrogen was 98.8 %, amino acid nitrogen was 13.7 ye, amino sugar nitrogen was 61.7 ye, and ammonia was 23.7 %.
The phosphorus content of the C-polysaccharide was accounted for completely (100%) by u-galactosamine 6phosphate and muramic acid phosphate.
DISCUSSION
The results of the present studies indicate that the C-poly- saccharide prepared in the manner described in the experimental
b
Hours of hydrolysis is plotted against the percentage of the substance remaining; 0, n-galactosamine 6-phosphate + n-galactosamine; A, Peak B + n-galactosamine B-phosphate + n-galactosamine; A, n-galactos- amine; X, synthetic n-galactosamine g-phosphate; l , n-galactos- amine B-phosphate from the C-polysaccharide.
section and the material obtained by Goebel et al. (3) are similar in their content of phosphorus and nitrogen. The values for reducing sugars (29% versus 50.6%) and acetyl groups (9.66% versus 13.06%) are quite different, but the two substances are serologically indistinguishable. The differences observed in reducing sugar and acetyl content probably reflect differences in the methods employed for the preparation of these materials. Preliminary studies with the aid of ion exchange chromatography have indicated that the C-polysaccharide prepared according to our procedure is not homogeneous. It is likely that the ma- terials obtained by Goebel et al. are also chromatographically inhomogeneous. Comparisons of chemical composition will become more meaningful when homogeneous products are obtained.
About 80% of the dry weight of the C-polysaccharide can be accounted for in terms of the nine components, lysine, alanine, serine, glutamic acid, n-glucosamine, n-galactosamine 6-phos- phate, muramic acid, muramic acid phosphate, and acetate, which is probably present as N-acetylhexosamines. The restricted number of amino acids and the presence of muramic acid indicate that the C-polysaccharide is of cell wall origin. This is further corroborated by the finding that one-half of the alanine is in the D form. The amino acids, alanine, glutamic acid, and lysine, are found in an approximate molar ratio of 2: 1: 1. This, together with the presence of both n-glucosamine and muramic acid, suggests that the C-polysaccharide contains
by guest on April 1, 2019
http://ww
w.jbc.org/
Dow
nloaded from

*June 1963 T.-Y. Liu and I<. C. Gotschlich 1933
the classical cell wall mucopeptide consisting of repeating units of n-glucosamine and muramic acid with the amino acids in peptide linkage on the carboxyl group of the muramic acid. Furthermore, it has been shown that pneumococcal cell walls prepared by shaking with glass beads in a Mickle disintegrator contained the C-po1ysaccharide.l
ds was noted above, the phosphorus content (4.71% or 148 pmoles per 100 mg) can be completely accounted for as D-
galactosamine 6-phosphate (140 pmoles per 100 mg) and muramic acid phosphate (8 pmoles per 100 mg). From the data shown in Fig. 3b, it would appear that all of the n-galactosamine is phosphorylated in the original C-polysaccharide. The fact that no galactosamine was detectable by paper chromatography when the C-polysaccharide was hydrolyzed at 100” with 2 N HCl rather than 6 N HCl points to the same conclusion. At present, there is no information as to whether the phosphorus is in primary or secondary phosphate linkage. To the best of our knowledge this is the first report of the presence of n-galactosamine 6- phosphate as a component of a bacterial cell wa11.2 Other phosphorylated sugars or polyols, such as n-glucosamine 6- phosphate, ribitol phosphate, and glycerol phosphate, have assumed major importance in the chemistry of bacterial cell walls, especially since the discovery of teichoic acid (27, 28).
Approximately one-fifth of the muramic acid was found to be phosphorylated. Agren and de Verdier (29) found a substance which they tentatively identified as muramic acid g-phosphate in acid hydrolysates of Lactobacillus bifidus. The organisms had previously been extracted with organic solvents and hot trichloroacetic acid, and no information was given as to whether the muramic acid phosphate could have come from the cell walls. The identification of muramic acid phosphate in the hydrolysate of C-polysaccharide suggests that it may be a component of the cell wall of the pneumococcus.
From the data in Table I, it is clear that all of the components of the C-polysaccharide have not yet been identified. The products listed account for only about 80% of the weight of the material submitted to analysis. Moreover, the quantity of ammonia found in acid hydrolysates far exceeds that which could arise from amide nitrogen that might be present. It appears likely that acid hydrolysis has caused the decomposition of some as yet unidentified nitrogenous material with the formation of ammonia. Chromatographic fractionation of the C-polysaccharide, followed by detailed chemical analyses? is now in progress and may illuminate this question.
SUMMARY
;\ method for the preparation of C-polysaccharide from a strain of rough pneumococcus R36A has been described. Ion exchange chromatography on columns of Amberlite In-120 has proved to be effective for the isolation and characterization of ninhydrin-positive constituents of the C-polysaccharide. The results of the present studies indicate that the C-poly- saccharide contains four major amino acids and four amino sugars; these are lysine (1.29o/0), serine (0.27O/,), glutamic acid (1.60%), alanine (1.63 Yb), n-glucosamine (I 1.32%), n-galac- tosamine g-phosphate (34.82 CjL), muramic acid (11.28 %), and
1 Unpublished resu1t.s of J>r. M. McCarty. * It is, of course, conceivable that the phosphate could be at-
tached at some position other than C-6 in the C-polysaccharide, since galactosamine K-phosphate is isolated after vigorous acid hydrolysis and phosphate migration may have occurred.
TABLE I Chemical composition of C-polysaccharide
All values are given in terms of the anhydrous, ash-free C- polysaccharide. Amino acids and amino sugars were determined by ion exchange chromat,ography (19). The values for the stable amino acids, lysine, glutamic acid, glycine, and alanine, are the average of three determinations. The values for serine, n-galac- tosamine 6-phosphate, n-galactosamine, muramic acid phosphate, and muramic acid are extrapolated values (see the text and Pig. 3). The ammonia value was corrected for the amount of ammonia contributed by the decomposition of serine and the amino sugars, and is the average of three determinations. Acetyl, nitrogen, and phosphorus were determined as described in the text.
Composition
___- _____--
Lysine. Serine Glutamic acid. . Glycine. . . Alanine*............... Muramic acid phos-
phate................ Muramic acid.. n-Galactosamine 6-
phosphate. n-Glucosamine
NHa Glucose................ Acetyl.................
Total recovery.
Nitrogen.. . Phosphorus.
Weight per 100 mg o haunt per 100 sample mg of sample
mg /.lmoles
1.29 10.1 0.27 3.1 1.60 (ztO.05) 12.4 0.069 (*0.002 1.2 1.63 (ztO.05) 23.0
4.66 0.715 2.86
0.286 5.31
8.0 47.0
34.82 11.32
140.0 70.0
2.14
9.66
103.5 (A151 13.2
225
1.85
10.9
32.3 16.2
23.7
78.43 98.8
6.07 4.71
433
418
Per cent f total N
* Total alanine content. t Calculated as dimuramic acid ester of phosphate.
muramic acid phosphate (4.35y0). A small amount of glycine (0.069%) is also found in acid hydrolysates. The restricted number of amino acids and the presence of muramic acid, together with the finding that one-half of the alanine is in the D
form, indicate that the C-polysaccharide prepared by our pro- cedure is of cell wall origin. n-Galactosamine 6-phosphate, a major constituent of the C-polysaccharide, has been isolated and identified for the first time in bacterial cell walls. About 20% of the weight of the polysaccharide has not yet been accounted for. There is also more ammonia in acid hydrolysates than could arise from amide nitrogen. It therefore seems likely that some unidentified nitrogenous component is present.
Acknowledgments-The authors wish to acknowledge with thanks the many suggestions and considerable assistance given them by Dr. Stanford Xoore, Dr. William H. Stein, and Dr. Maclyn McCarty. We greatly appreciate the cooperation of Dr. Saul Roseman in providing us with samples of synthetic galactosamine, glucosamine, and mannosamine 6-phosphates, and of Dr. J. T. Park in sending us a sample of synthetic muramic acid. We also thank Miss Barbara Bowman for her skillful assistance with the experiments and Mr. S. Theodore Bella for his expert performance of the microanalyses. We are indebted
by guest on April 1, 2019
http://ww
w.jbc.org/
Dow
nloaded from

1934 Pneumococcal C-Polysaccharide Vol. 238, No. 6
to Dr. Herbert Jaffe for the infrared spectra of n-galactosamine 12. RONDLE, C. J. M., AND MORGAN, W. T. J., Biochem. J., 61,
G-phosphate. The n-amino acid oxidase was a gift from Dr. 2. 586 (1955).
Bohak, and the streptococcal proteinase was supplied by Dr. S. 13. HAGEDORN, H. C., AND JENSEN, B. N., Biochem. Z., 135, 46
(1923). D. Elliott, to whom we also express our gratitude. 14. RAPPORT, M. M., MEYER, K., AND LINKER, A., J. Biol. Chem.,
1.
2.
3.
4.
5. 6. 7.
8.
9.
10.
11.
REFERENCES
TILLETT, W. S., GOEBEL, W. F., AND AVERY, 0. T., J. Exptl. Med., 62, 895 (1930).
HEIDELBERGER, M., AND KENDALL, F. E., J. Exptl. Med., 63, 625 (1931).
GOEBEL, W. F., SHEDLOVSKY, T., LAVIN, G. I., AND ADAMS, M. H., J. Biol. Chem., 148, 1 (1943).
ANDERSON, H. C., AND MCCARTY, M., J. Exptl. Med., 93, 25 (1951).
ELLIOTT, S. D., J. Exptl. Med., 92,201 (1950). SEVAG, M. G., Biochem. Z., 273, 419 (1934). CRESTFIELD, A. M., MOORE, S., AND STEIN, W. H., J. Biol.
Chem., 238, 622 (1963). FISKE, C. H., ANI) SUBBARO~, Y., J. Biol. Chem., 66, 375
(1925). UMBREIT, W. W., BURRIS, R. H., AND STAUFFER, J. L., Muno-
metric techniques and related methods for the study of tissue metabolism, Burgess Publishing Company, Minneapolis, 1945.
CHEN, P. S., JR., ToRIB.~R.~, T. Y., AND WARNER, H., Anal. Chem., 28, 1756 (1956).
ELEK, A., AND HARTE, R. A., Ind. Eng. Chem., Anal. Ed., 8, 267 (1936).
186, 615 (1950). 15. DISCHE, Z., AND SHETTLES, L. B., J. Biol. Chem., 176, 595
(1948). 16. DISCHE, Z., J. Biol. Chem., 167, 189 (1947). 17. STOFFYN, P. J., AND JEANLOZ, R. W., Arch. Biochem. Biophys.,
62, 373 (1954). 18. PARTRIDGE, S. M., Nature, 164, 443 (1949). 19. SPACKMAN. D. H., STEIN, W. H., AND MOORE, S., Anal. Chem.,
30, 1190 ‘(1958): 20. DR&ZE. A.. MOORE. S.. AND BIGWOOD. E. J.. Anal. Chim. Acta.
11,554 (1954). ’ ’ I ,
21. DISTLER, J. J., MERRICK, J. M., AND ROSEMAN, S., J. Biol. Chem., 230, 497 (1958).
22. MASSEY, V., PALMER, G., AND VENNETT, R., Biochim. et Bio- phys. Acta, 48, 1 (1961).
23. STARK, G. R., AND SMYTH, D. G., J. Biol. Chenz., 238, 214 (1963).
24. BYRNE, W. L., AND LARUY, H. A., Biochim. et Biophys. Acta, 14, 495 (1954).
25. DISCHE, Z., AND BORENFREUND, E., J. Biol. Chem., 180, 1297 (1949).
26. STEIN, W. H., AND MOORE, S., J. Biol. Chem., 176,337 (1948). 27’. RODE. E.. AND GOEBEL. W. F.. J. Exvtl. Med.. 116, 73 (1962). 28. BAD&LEG, J., Federation Proc:, 21, lb84 (1962). 29. ~GREN, G., AND DE VERDIER, C. H., Acta Chem. &and., 12,
1927 (1958).
by guest on April 1, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Teh-Yung Liu and Emil C. GotschlichThe Chemical Composition of Pneumococcal C-Polysaccharide
1963, 238:1928-1934.J. Biol. Chem.
http://www.jbc.org/content/238/6/1928.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/238/6/1928.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 1, 2019
http://ww
w.jbc.org/
Dow
nloaded from





























