Embed Size (px)
Citation preview

The changing shape of mitochondrialapoptosisMichał Wasilewski1 and Luca Scorrano1,2
1 Dulbecco-Telethon Institute, Venetian Institute of Molecular Medicine, Via Orus 2, 35129 Padova, Italy2 Department of Cell Physiology and Metabolism, University of Geneva Medical School, 1 Rue M. Servet, 1211 Geneve, Switzerland
Review
Mitochondria are key organelles in conversion of energy,regulation of cellular signaling and amplification of pro-grammed cell death. The anatomy of the organellematches this functional versatility in complexity and ismodulated by the concerted action of proteins thatimpinge on its fusion–fission equilibrium. A growingbody of evidence implicates changes in mitochondrialshape in the progression of apoptosis and, therefore,proteins governing such changes are likely candidatesfor involvement in pathogenetic mechanisms in neuro-degeneration and cancer. Here, we discuss the recentadvancements in our knowledge about the machinerythat regulates mitochondrial shape and on the roleof molecular mechanisms controlling mitochondrialmorphology during cell death.
Mitochondrial shapeMitochondria are essential organelles for the life and deathof a cell and participate in energy conversion, regulation ofsignaling cascades and apoptosis [1] (Box 1). Although theconventional image of these organelles is of a static nature,mitochondria can fuse and divide. This ability has beenknown for years but obscured by the fine yet static detailsprovided by electron microscopy. The advanced toolsof live-cell imaging and 3D reconstruction, as well aselectron tomography, have enabled us to fully appreciatetheir complexity and dynamic nature. The spectrum ofmitochondrial shapes ranges from spherical, grain-like,individual entities to long, branched filaments that, ulti-mately, might form one interconnected ‘megamitochon-drion’ per cell. Mitochondrial complexity begins at thelevel of their external appearance and continues to asimilarly complex and dynamic internal structure [2].
A major leap forward in understanding the mechanismsand consequences of mitochondrial fusion–fission camewith the discovery that the morphology of the organelle,like its ultrastructure, undergoes dynamic changes duringapoptosis. Interfering with these changes slows apoptoticprogression, indicating a functional role for mitochondrialshape in this process [3–5]. This finding instigated researchon the molecular details, mechanisms and pathophysiolo-gical consequences of mitochondrial shape changes (Box 2).Here, we review the current understanding of mitochon-drial fusion and fission in mammals and discuss evidencethat supports a role for morphological changes of mito-chondria in certain physiological and pathological pro-cesses, including apoptosis (Box 3).
Corresponding author: Scorrano, L. ([email protected]).
1043-2760/$ – see front matter � 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.tem.2009.
Mitochondrial fusion and fissionMitochondrial shape results from the balance of twoongoing antagonistic processes, fusion and fission. Wheneither process is blocked, the final morphology of themitochondria is the consequence of unopposed progressiontowards the other side of the equilibrium [6]. Members ofthe machinery regulating mitochondrial fusion have beenidentified by analyzing organelle morphology in Sacchar-omices cerevisiaemutants [7,8]. The shape of mitochondriadepends on several large ubiquitous GTPases with struc-tural homology to dynamins [9] that participate in fusion,fission and tubulation of biological membranes. Proteinsthat regulate mitochondrial shape share at least theGTPase domain and a C-terminal coiled-coil domain withthe prototypical dynamins, and these domains can functionas GTPase effectors or to mediate protein–protein inter-action [10]. The dynamin-related proteins that impinge onmitochondrial fusion are peculiar in that most of them areintegral membrane proteins. We next discuss the proper-ties of ‘mitochondria-shaping’ proteins of mammals inparticular. A scheme of the players known to participatein mitochondrial fission and fusion in mammalian cells ispresented in Figure 1 and 2, respectively.
FissionIn mammalian cells, mitochondrial fission depends ondynamin-related protein 1 (Drp1), a cytoplasmic largeGTPase similar to dynamin that mediates the fragmenta-tion of mitochondria and peroxisomes [11]. Drp1 translo-cates to mitochondria in response to cellular andmitochondrial cues. After mitochondrial dysfunction, cyto-plasmic Ca2+ rises, leading to activation of calcineurin anddephosphorylation of the conserved Ser637 of Drp1 [12],inducing translocation of Drp1 to mitochondria. Conflict-ing data exists surrounding the kinase responsible forphosphorylation of this residue. Protein kinase A wasreported to fulfill this function, linking mitochondrialmorphology to another crucial second messenger, cyclicAMP [13,14]. Alternatively, Ser637 could be phosphory-lated by calmodulin-dependent protein kinase Ia, althoughin this case, phosphorylation of Drp1 induced its mitochon-drial localization [15]. The phosphorylation status of thissite is dominant over that of Ser616 [12], which is con-trolled by cyclin-dependent kinase 1 to drive mitochondrialfission duringmitosis [16]. Mitochondrial Drp1 can then bestabilized on the surface of the organelle by SUMOylation[17,18], a process known to protect molecules from degra-dation by the ubiquitin-proteasome system.
03.007 Available online 31 July 2009 287

Box 1. Mitochondria
Mitochondria are double-membrane organelles, the origin of which
has been traced to an ancient prokaryotic endosymbiont. The outer
mitochondrial membrane (OMM) has a composition similar to that of
other eukaryotic membranes and originates from a vesicle engulfing
the endosymbiont. It is permeable to metabolites and small peptides
up to 3000 Da, thanks to the presence of voltage-dependent anion
channels. The inner mitochondrial membrane (IMM) has a clearer
prokaryotic origin and differs from the OMM in its physical properties
and composition. Its ion conductance and metabolite permeability are
tightly controlled. IMM comprises a larger fraction of proteins and
approximately 20% of cardiolipin, a lipid also found in bacterial
membranes, which is crucial for proper activity of several IMM
enzymes. IMM folds into a complicated network of tubules and
lamellae called cristae, reflecting its unique function. IMM is the site
of oxidative phosphorylation (OXPHOS), which provides most of the
ATP produced during aerobic metabolism. According to the chemios-
motic theory, the basic principle of OXPHOS is to pump protons
across the proton-impermeable IMM, generating an electrochemical
potential, which is then used to drive phosphorylation of ADP to ATP
by F1F0-ATPase synthase. Energy required for proton pumping is
provided by electrons from NADH and FADH produced during the
tricarboxylic acid cycle and other catabolic processes. The energy of
electrons, liberated in a number of redox reactions, is translated into
proton pumping by three large protein complexes in IMM (complex I,
III and IV), called together with the non-proton-pumping complex II,
the respiratory chain (RC) [1].
Mitochondria contain an autonomous circular small genome that is
located in the internal compartment, the matrix. mtDNA encodes only
13 proteins, mitochondrial ribosomal RNA and transfer RNA, but
mutations in these genes result in severe diseases, mostly because of
impaired energy production.
Apart from OXPHOS, mitochondria participate in other important
metabolic pathways, such as fatty acid synthesis, gluconeogenesis,
steroidogenesis, synthesis of haeme and urea cycle [2]. Moreover,
mitochondria cooperate with the ER to control calcium homeostasis
and integrate information during apoptosis. The balance between
pro- and antiapoptotic proteins of the Bcl-2 family controls release of
cytochrome c, apoptosis-induction factor, Smac/DIABLO and en-
donuclease G from mitochondria. Sequestered in mitochondria,
these proteins carry out routine tasks (e.g. cytochrome c shuttles
electrons in RC). When released to the cytoplasm, they function as
proapoptotic factors, either by inducing apoptosome formation
(cytochrome c) or by a direct enzymatic activity (endonuclease G)
[74].
Review Trends in Endocrinology and Metabolism Vol.20 No.6
Fis1 is a membrane protein homogenously distributedin the outer mitochondrial membrane (OMM) via a trans-membrane domain located in the C-terminal region, andonly a small portion of the molecule protrudes into theintermembrane space (IMS) (Figure 1). The cytoplasmicregion contains six alpha helices, four of which (a2–a5)form two tetratricopeptide-repeat (TPR)-like domains, pre-dicted to allow protein–protein interactions [19]. Overex-pression of Fis1 induces mitochondrial fragmentation, butbecause it does not possess enzymatic activity, its roleis probably restricted to anchoring effector proteins tomitochondria. Accordingly, mitochondrial fragmentationcaused by Fis1 overexpression can be blocked by expres-sion of a dominant-negative mutant of Drp1 [20]. More-over, Drp1 and Fis1 seem to interact, as judged bycrosslinking and coimmunoprecipitation [21]. The Drp1–
Fis interaction is a transient event, and efficient fission
Box 2. Mitochondrial dynamics and bioenergetics
One of the most prominent functions of mitochondria is ATP
production by oxidative phosphorylation. Research on the relation-
ship between mitochondrial shape and function dates back to 1966,
when Hackenbrock [75] observed that mitochondria isolated from rat
livers shrunk and displayed a denser matrix during respiration. This
conformation was called ‘condensed’, as opposed to the ‘orthodox’
conformation of inactive mitochondria. The matrix conformation
transition was ascribed to osmotic changes caused by K+ movements
across the inner mitochondrial membrane and thought to be only an
artefact of isolated mitochondria. Indeed, mitochondria observed in
most mammalian tissues manifest orthodox conformation [76,77].
Nevertheless, mitochondria from muscles with high energy require-
ments seem to have condensed matrix [78]. Moreover, matrix
condensation is observed during apoptosis [79].
Overall, mitochondrial morphology depends on the energetic state
of the cell. In general, treatments that compromise mitochondrial
membrane potential induce mitochondrial fragmentation [12], an
observation that correlates well with data from other organisms.
Fusion of the outer membrane in yeast, for example, requires a
chemical component of electrochemical potential, and fusion of the
inner membrane depends on an electrical component [80]. Interest-
ingly, recovery of mitochondrial morphology after wash-out of
uncouplers requires de novo protein synthesis, suggesting a defect
288
requires dissociation of the complex. Accordingly, geneticmodifications of the N-terminal domain of Fis1, whichstabilize its binding to Drp1, abolish mitochondrial frag-mentation [22]. However, the absolute requirement of Fis1for binding Drp1 to mitochondria is somewhat weakenedby the observation that downregulation of Fis1 only par-tially diminishes Drp1 recruitment to mitochondria [6].Another question on themechanism of interaction betweenFis1 and Drp1 arises from the yeast model, in which thisinteraction requires the adaptor Mdv1 [23]. A mammalianorthologue for Mdv1 has not been yet identified, but con-sidering the conservation of fission mechanisms,additional elements could be involved.
Mechanism of fissionBy comparing Drp1 with the prototypical dynamins andconsidering data obtained in yeast, it was postulated that
in mitochondrial fusion [81] probably caused by Opa1 degradation
[42]. It is tempting to speculate that decreased potential blocks fusion
by Opa1 degradation and ultimately leads to mitochondrial fragmen-
tation by activating the fission machinery [12].
In turn, the bioenergetic state of a cell can be modified by changing
the levels of mitochondria-shaping proteins. Loss of fusion and
resulting mitochondrial fragmentation can cause mitochondrial
dysfunction, perhaps through the impaired exchange of matrix
material in fusion-incompetent mitochondria [82] and consequent
accumulation of stochastic functional errors. It is more difficult to
explain why failure to divide results in mitochondrial dysfunction [83].
Blockage of fission could impair elimination of defective elements of
the network by autophagy, leading to accumulation of dysfunctional
organelles and to dysfunction via secondary mechanisms, such as
those induced by reactive oxygen species [83,84].
Another emerging link between bioenergetics and mitochondrial
structure comes from studies of the F1F0-ATP synthase [85]. Dimer
formation of this enzyme does not influence its activity but does affect
cristae structure [85]. Accordingly, ribbons of enzyme dimers are
located in domains of high membrane curvature (e.g. cristae tubules
and on the sides of flat lamellar cristae) [86], and overexpression of
IF1, the natural inhibitor of F1F0-ATP synthase, stimulates both
enzyme dimerization and cristae biogenesis [87].

Box 3. Diseases of mitochondria-shaping proteins
Charcot-Marie-Tooth
Charcot-Marie-Tooth (CMT) is an inherited peripheral neuropathy that
causes progressive deterioration of nerve conduction velocity in
peripheral neurons, clinically apparent as muscle atrophy and
sensory defects. CMT is a heterogeneous disease caused by
demyelination of nerves (CMTI, CMTIII and CMTIV) or axonal
degeneration without demyelination (CMTII). Mutations in Mfn2 are
associated with most cases of CMT type IIa (CMTIIa) [36]. Molecular
mechanisms underlying CMTIIa are not clear. Axonal degeneration
could be a consequence of insufficient energy supply caused by
improper mitochondria localization. In fact, Mfn2 dysfunction in
neurons leads to perinuclear aggregation of mitochondria and a
decrease in their axonal transport [82,88]. Alternatively, given the low
expression of Mfn1 in these cells, mutated Mfn2 might not be
functionally complemented by its Mfn1 homologue [82]. Accordingly,
aggregation of mitochondria is not observed in fibroblasts from
CMTIIa patients, which express higher levels of Mfn1 [89].
Mutations in GDAP1 are also associated with another type of CMT,
type IVa. GDAP1 is an integral protein of the outer mitochondrial
membrane expressed in neurons of the peripheral and central
nervous system. GDAP1 mutants have heterogeneous influence on
mitochondria morphology, ranging from fragmentation to perinuc-
lear aggregation [90]. The mechanism or mechanisms by which
mutations in GDAP1 cause CMT remain(s) elusive.
Autosomal dominant optic atrophy
Autosomal dominant optic atrophy (ADOA) is the most common
inherited optic neuropathy and is characterized by mild-to-moderate
progressive loss of visual acuity, often accompanied by impaired
color perception, central visual field defects and temporal optic disc
pallor. ADOA is caused by loss of retinal ganglion (RG) neurons and is
associated with Opa1 mutations [91,92]. ADOA is probably caused by
haploinsufficiency [93]; accordingly, decreased Opa1 levels are found
in samples collected from patients [94]. However, the relationship
between Opa1 levels and RG cell death remains unsolved. Some
evidence points to impaired oxidative phosphorylation as a possible
underlying mechanism, whereas other data suggest involvement of
mitochondrial dynamics and increased susceptibility to apoptosis [94].
Parkinson’s disease
Parkinson’s disease (PD) is a neurodegenerative disorder caused by
loss of dopaminergic neurons in the midbrain. The phosphatase and
tensin homolog-induced kinase 1 (Pink1) and Parkin, two genes
involved in the inherited form of PD, have been recently connected to
mitochondrial dynamics. Pink1 is a mitochondrial serine/threonine
kinase, and Parkin is a cytoplasmic E3 ubiquitin ligase. Null mutants
of Pink1 and Parkin in Drosophila melanogaster display mitochondrial
dysfunction and PD-like symptoms [95]. Of note, Parkin complements
loss of Pink1 [96]. Considerable interest was stirred by the finding that
induction of mitochondrial fission or attenuation of fusion amelio-
rates the inherited PD in these fly models [97], suggesting that Pink1
and Parkin might regulate mitochondrial shape. However, the main
question is whether the effects of Pink1 and Parkin on mitochondrial
morphology are direct or epiphenomena of a primary function; for
example, in the regulation of mitochondrial (dys)function and
degradation by autophagy. So far, the available evidence supports
the latter hypothesis. Loss of Pink1 is associated with defects in the
mitochondrial respiratory chain [98,99] and Parkin targets dysfunc-
tional mitochondria to autophagosomes, where they are degraded
[100]. Thus, fragmentation observed in Pink1-deficient cells might
well be a consequence of organelle dysfunction, and the finding that
induction of fusion worsens the phenotype could be a consequence of
impaired mitophagy that obviously requires organelles of discrete
size to be targeted to the autophagosomes.
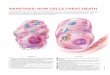
Figure 1. A cartoon depicting some of the players involved in fission of
mammalian mitochondria. The site of fission, represented by the ring around a
constricted mitochondrion, is boxed and enlarged. Drp1, dynamin-related protein
1; IMM, inner mitochondrial membrane; Mff, mitochondrial fission factor; Mtp18,
mitochondrial protein of 18 kDa; OMM, outer mitochondrial membrane.
Review Trends in Endocrinology and Metabolism Vol.20 No.6
Drp1 assembles in spirals around the fission sites, con-stricting them in a GTP-dependent manner [24]. However,the earlier model of action of prototypical dynamins hasbeen challenged recently by the discovery that fissiondepends on the concerted action of dynamin and of mech-anical forces acting on the membrane. This novel modelpostulates that fission is executed not only by the constrict-ing force generated by dynamin but also by lipid remodel-
ing and lateral tension, which in vivomight be provided bypulling the organelle along microtubules, its intracellulartracks [25,26]. It should be noted that Drp1 differs fromconventional dynamins because it does not possess a lipid-binding domain, perhaps explaining its requirement for amembrane receptor such as Fis1 (and perhaps others thatare yet unknown). Indeed, fission sites are often charac-terized by high Fis1 concentrations [27].
FusionFusion of the OMM is governed by two dynamin-relatedGTPases: mitofusin 1 (Mfn1) and mitofusin 2 (Mfn2). Theydisplay a high degree of homology and, accordingly, asimilar structure, composed of a terminal GTPase domain,two hydrophobic heptad repeats (HR) and two transmem-brane domains connected by a very short intramembranespace region (only five residues) [28]. Nevertheless, theyare not functionally equal. The GTPase activity of Mfn1 ismuch higher, although its affinity for GTP is lower, thanthat ofMfn2 [29]. Thus, they seem to have slightly differentroles in mitochondrial fusion. Mfn1 is responsible for mito-chondria tethering by antiparallel interaction of HR2 ofproteins from adjacent mitochondria [30]. The role of Mfn2is somewhat elusive, but this protein can be retrieved inhetero-oligomers with Mfn1 and perhaps participates inlater steps of mitochondrial fusion. Overexpression of aninactive mutant of Mfn2 leads to aggregation of mitochon-dria, as if their fusion was stalled at the step of initialtethering [31,32]. In addition, Mfn2 levels correlate withoxidative metabolism of skeletal muscle [33] and theproliferative ability of vascular smooth muscle cells by
289

Figure 2. A cartoon depicting some of the players involved in fusion of mammalian mitochondria. The inner (IMM) and outer (OMM) membranes of a mitochondrion are
depicted with the main players in mitochondrial fusion and regulation of the shape of cristae. The OMM of an adjacent fusing mitochondrion is also represented. Mfn,
mitofusin; Mib, mitofusins-binding protein; Opa1, optic atrophy 1.
Review Trends in Endocrinology and Metabolism Vol.20 No.6
sequestering the proto-oncogene Ras [34]. Mfn2 also con-trols the shape of the endoplasmic reticulum ER andtethers it to mitochondria [35]. Finally, Mfn2 mutationsare associated with Charcot-Marie-Tooth type IIa periph-eral neuropathy [36] (Box 3). Thus, it seems that Mfn2 hasa broader spectrum of functions than Mfn1. However, atleast in fibroblasts, Mfn1 (but not Mfn2) is required forfusion triggered by the inner membrane dynamin-relatedprotein Opa1 [37], further complicating the picture. Mfnscommunicate with Opa1, probably through the smallregion exposed to the IMS; a single mutation in this regioncompromises their pro-fusion ability [38].
Opa1 belongs to the family of dynamin-related proteins.It is anchored to the inner mitochondrial membrane (IMM)by a transmembrane domain located close to the N termi-nus, and most of the protein is exposed to the IMS. Inhumans, there are eight splice variants of Opa1, whereasin mice, there are only four [39]. All alternatively splicedexons are located between the GTPase domain and the Nterminus, close to the region inserted in the IMM. Alter-natively spliced Opa1 isoforms are subject to a complexpost-translational cleavage, represented experimentallyby five bands on a Western blot. Two higher molecularweight MW bands represent proteins integrated into theIMM, whereas three lower MW bands reflect forms thatcan be released into the IMS [40]. Controversies remain onhow the cleavage is exerted, but a unifying model has beenproposed in which the m-AAA protease paraplegin orAFG3L2 [40,41] or the i-AAA protease Yme1L [42,43]produce the lower MW forms of Opa1, on which the rhom-boid protease Parl acts to release soluble Opa1 to the IMSspace [44].
290
Two functions of Opa1 have been defined thus far. Opa1drives Mfn1-dependent fusion of mitochondria [37], per-haps contacting Mfn1 directly through its short IMSdomain [38]. Fusion requires both long and short formsof Opa1 [43], suggesting that different forms of Opa1govern mitochondrial fusion. Opa1 is also crucial forIMM structure. In fact, dynamics of IMM seem to dependexclusively on Opa1. Its downregulation causes theappearance of vacuolar cristae and widening of cristaejunctions [45,46], owing to disruption of a complex com-prising IMM and IMS forms of the protein [44,46](Figure 2).
Other proteins regulating mitochondrial dynamicsBesides the canonical mitochondria-shaping proteins,other factors have been suggested to regulate themorphology of the organelle.
Overexpression ofmitochondrial protein 18kDa (Mtp18)causes Drp1-dependent fragmentation, and its knockdownleads to highly interconnected mitochondria [47]. It hasbeen proposed, therefore, that Mtp18 acts downstream ofDrp1-Fis1, transmitting a division signal to the IMM[47,48]. Mtp18 has an antiapoptotic function; its silencingleads to cytochrome c release and cell death and sensitizescells to apoptotic stimuli. Knockdown of Mtp18 is a rareexample of increased susceptibility to apoptosis associatedwith highly interconnected mitochondria.
Endophilin B1 is a cytoplasmic protein involved inregulating membrane curvature that partially colocalizeswith mitochondria. Its downregulation causes dissociationof the OMM from the IMM, resulting in Drp1-dependentformation of OMM tubules and vesicles [49]. It was also

Review Trends in Endocrinology and Metabolism Vol.20 No.6
implicated in apoptotic signaling because it translocatesto mitochondria, where it mediates Bcl-2-associated Xprotein (Bax) activation, during cell death [50]. However,it is unclear whether membrane shaping by endophilin B1is necessary for Bax activation [51].
Mitofusin-binding protein (Mib) is a cytoplasmic proteinidentified through its interaction with Mfn1. It is postu-lated that Mib inhibits mitochondrial fusion by directlyinterfering with Mfn1 and possibly with Mfn2 [52].
Screens for mitochondrial morphology defects in Droso-phila melanogaster brought to light another candidatecomponent of the fission machinery. Mitochondrial fissionfactor, the human homolog of the identified protein, is anintegral protein of the OMM, and its knockdown inducesmitochondrial elongation and partly protects from frag-mentation caused by uncoupler or dominant-negativeMfn1, even though it does not interact with Drp1 or hFis1[53].
In addition to protein factors, the lipid milieu is alsoinvolved in the control of mitochondrial dynamics. Phos-phatidic acid, a fusogenic lipid produced in OMM fromcardiolipin by phospholipase D, is required for properfusion of mitochondria mediated by Mfns [54]. Moreover,recruitment of inositol 50-phosphatase synaptojanin 2A,another enzyme responsible for lipid remodeling, to mito-chondria causes mitochondria aggregation [55].
Apoptosis and changes in mitochondrial shapeAs we mentioned earlier, several reports indicate thatmitochondrial shape varies during apoptosis. However,whether these changes represent an epiphenomenon ofthe cell death cascade or play a key part in the amplifica-tion of this process remains the subject of much debate. Wenext review evidence implicating mitochondria-shapingproteins in cell death and the proposed mechanisms bywhich they potentially influence apoptosis.
The outer side of death: mitochondrial fragmentationThe original study associating mitochondrial fragmenta-tion with neuronal apoptosis [3] was further substantiatedby the discovery that when fragmentation is blocked by adominant-negative mutant of Drp1, progression towardscell death slows [4]. It has to be stressed that althoughmechanisms of apoptosis andmitochondrial fission overlapto some extent, they are not inevitably connected. Forexample, in Caenorhabditis elegans, mitochondria fissionis executed by full length DRP-1, whereas apoptosis isamplified by processing of DRP-1 by the caspase CED-3[56]. Drp1 forms foci in the mitochondrial membrane,ultimately colocalizing with scission sites. Interestingly,upon apoptotic stimulation, at least two other proteinscolocalize at these foci: Mfn2 and Bax [27]. Our under-standing of this remains elusive, but several lines of evi-dence suggest that foci serve as platforms for crosstalkbetween pro- and anti-apoptotic, as well as fusion andfission, molecules. For example, formation of fission sitescould be a requisite for Bax translocation to mitochondria[57]. In line with this, overexpression of Bcl-xL preventsapoptosis but not fragmentation [58]. Drp1 has beenreported to act downstream of Bax because its silencinghas no effect on Bax translocation [6,27]. This is corrobo-
rated by Drp1 ‘selectivity’ (i.e. Drp1 controls the release ofcytochrome c from mitochondria but does not control therelease of Smac/DIABLO) [59]. Under non-apoptotic con-ditions, the majority of Drp1 cycles between the cytoplasmandmitochondria, and the remaining 20% is stably bound tomitochondria; Bax translocation increases the percentage ofDrp1 that is stably bound to mitochondria. Interestingly,accumulation of Drp1 on the mitochondrial surface occursbetween completion of fission and initiation of cytochrome crelease [18]. This biphasic behavior suggests that althoughfission and cytochrome c release are dependent on the sameprotein and occur very close in time, they might be distinctevents. Dissociation between pro-fission and pro-apoptoticfunctions of Drp1 was also observed in experiments with anovel inhibitor of Drp1, which attenuated permeabilizationof the OMM in isolated mitochondria, even though fissiondid not take place in this system [60]. Several post-transla-tional modifications of Drp1 (including phosphorylation,ubiquitination and SUMOylation) influence its cellularlocalization. Phosphorylation of Ser637 by protein kinaseA prevents translocation of Drp1 tomitochondria andDrp1-dependent fission [14], ultimately slowing apoptosis [13]. Inresponse to a sustained increase in cytosolic Ca2+, Ser637 isdephosphorylated by calcineurin and Drp1 translocates tomitochondria [12]. SUMOylation of Drp1 controls Drp1levels and mitochondrial localization, as well as mitochon-drial fragmentation [17,18]. Drp1 can also be ubiquitinatedby membrane-associated RING-CH 5, which probably pro-motes translocation of Drp1 to mitochondria [61–63]. Thebalance between these post-translational modifications canultimately determine the morphology of the organelle.
Mfn2 is another proposed partner of Bax in the OMM.LikeDrp1, it colocalizes with Bax in foci [27]. Interestingly,point mutants that favor the GTP-bound form display ahomogenous distribution in the OMM [38], wheras inacti-vation of theGTPase domain blocksMfn2 from localizing tofoci [64]. In addition, Bax translocation is blocked whenMfn2 is restricted outside foci and, similarly, Mfn2 spreadshomogenously in the OMM in Bax/Bcl-2 homologousantagonist killer (Bak) deficient cells [38,64]. Overexpres-sion of wild-type Mfn2, localized in foci, triggers apoptosis[65]. In contrast, constitutively active mutants of Mfn2 donot differ from wild-type Mfn2 in their influence on mito-chondrial shape [66]. Thus, localization of Mfn2 in foci isimportant for Bax-mediated permeabilization of the OMM.An intriguing possibility is that Mfn2-enriched focirepresent Mfn2-mediated tethering between ER and mito-chondria [35], where lipids required for Bax-mediatedpermeabilization of the OMM [67] can be readily trans-ferred to mitochondria [68]. Mfn2 not only participates inBax-mediated permeabilization of the OMM but also inter-feres with complex survival signaling cascades, especiallyin cells of muscular lineage [34]. However, involvement ofMfn2 in ER–mitochondria tethering and in the regulationof survival signaling cascades seems to be independentfrom its mitochondrial localization and its effect ondynamics of the organelle [34,69], further stressing themultifaceted character of this protein.
One would expect, given the interaction between Fis1and Drp1 in mitochondrial fission, that this same inter-action occurs in apoptosis. However, it seems that cell
291

Review Trends in Endocrinology and Metabolism Vol.20 No.6
death evokes Fis1 overexpression independent of Drp1[20]. This concept is strengthened by the fact that apoptosisinduced by Fis1 does not involve Bax or Bak activation butis instead a Ca2+-dependent process involving events in theIMM. This is substantiated by the ability of scavengers ofreactive oxygen species and of mutations in the shortdomain of Fis1 in the IMS to block Fis1-induced apoptosis[70]. Accordingly, Fis1 promotes rearrangements in IMMstructure [21], suggesting crosstalk between pathways ofmitochondrial fission and changes in the ultrastructure ofmitochondria during cell death.
The inner side of death: cristae remodelingUnder basal conditions, the majority of cytochrome c islocated in the cristae where the complexes of the respir-atory chain reside. To enable its complete release duringapoptosis, cytochrome c is redistributed to the peripheralIMS in a process called cristae remodeling, characterizedby fusion of individual cristae and widening of the narrowcristae junctions [5].
Remodeling of the cristae occurs in response to a varietyof apoptotic stimuli, including members of the Bcl-2 familysuch as Bid [5], Bim [71] and Bik [72]. In the case of Bik,however, the effect seems indirect and mediated by Drp1-dependent mitochondrial fission, thereby linking the outerto the inner side of mitochondrial shape changes duringcell death. It has been questioned whether changes in theshape of the IMM are a late event following caspaseactivation or a required step for complete cytochrome crelease [73]. In accordance with its proposed function incristae biogenesis, Opa1 is the master regulator of cristaeshape and, therefore, of the rate and extent of cytochrome crelease during apoptosis [44,46,71]. This function dependson its GTPase activity and on the formation of an oligomerthat comprises both the membrane-bound and solubleforms of the protein [46]. The rhomboid protease Parlhas a crucial role in the proper assembly of the oligomer,which is destabilized early in the course of apoptosis,before the release of cytochrome c. In cells lacking Parl,oligomerization of Opa1 is impaired, resulting in a highersensitivity to apoptosis [44]. Thus, evidence supports thelatter hypothesis that cristae remodeling is associatedwithdisassembly of the Opa1-oligomer and required for cyto-chrome c release and the progression of the apoptoticcascade. However, a number of questions remain: how iscristae remodeling triggered? What are the functional con-sequences of this remodeling on mitochondrial function?Can the Opa1-dependent arm be exploited to devise noveltherapeutics to increase apoptosis in cancer cells?
Future perspectivesExtensive research on the role of mitochondrial dynamicsin cell death has led to the consensus that mitochondrialshape changes occur during apoptosis. However, given themultiple roles of mitochondria-shaping proteins in regulat-ing neuronal differentiation and dendrogenesis, lympho-cyte migration, and aging, genetic tools to conditionallyablate or overexpress mitochondrial fusion/fission playersremain an outstanding requirement to address the func-tional significance of these shaping proteins.
292
References1 Nicholls, D.G. and Ferguson, S.J. (2002) Bioenergetics. Academic
Press2 Dimmer, K.S. and Scorrano, L. (2006) (De)constructing mitochondria:
what for? Physiology 21, 233–2413 Martinou, I. et al. (1999) The release of cytochrome c from
mitochondria during apoptosis of NGF-deprived sympatheticneurons is a reversible event. J. Cell Biol. 144, 883–889
4 Frank, S. et al. (2001) The role of dynamin-related protein 1, amediator of mitochondrial fission, in apoptosis. Dev. Cell 1, 515–525
5 Scorrano, L. et al. (2002) A distinct pathway remodels mitochondrialcristae andmobilizes cytochrome c during apoptosis.Dev. Cell 2, 55–67
6 Lee, Y.J. et al. (2004) Roles of the mammalian mitochondrial fissionand fusion mediators Fis1, Drp1, and Opa1 in apoptosis. Mol. Biol.Cell 15, 5001–5011
7 Dimmer, K.S. et al. (2002) Genetic basis of mitochondrial function andmorphology in Saccharomyces cerevisiae. Mol. Biol. Cell 13, 847–853
8 McConnell, S.J. et al. (1990) Temperature-sensitive yeast mutantsdefective in mitochondrial inheritance. J. Cell Biol. 111, 967–976
9 Okamoto, K. and Shaw, J.M. (2005) Mitochondrial morphology anddynamics in yeast andmulticellular eukaryotes. Annu. Rev. Genet. 39,503–536
10 Praefcke, G.J. and McMahon, H.T. (2004) The dynamin superfamily:universal membrane tubulation and fission molecules?Nat. Rev. Mol.Cell Biol. 5, 133–147
11 Schrader, M. (2006) Shared components of mitochondrial andperoxisomal division. Biochim. Biophys. Acta 1763, 531–541
12 Cereghetti, G.M. et al. (2008) Dephosphorylation by calcineurinregulates translocation of Drp1 to mitochondria. Proc. Natl. Acad.Sci. U. S. A. 105, 15803–15808
13 Cribbs, J.T. and Strack, S. (2007) Reversible phosphorylation of Drp1by cyclic AMP-dependent protein kinase and calcineurin regulatesmitochondrial fission and cell death. EMBO Rep. 8, 939–944
14 Chang, C.R. and Blackstone, C. (2007) Cyclic AMP-dependent proteinkinase phosphorylation of Drp1 regulates its GTPase activity andmitochondrial morphology. J. Biol. Chem. 282, 21583–21587
15 Han, X.J. et al. (2008) CaM kinase I alpha-induced phosphorylation ofDrp1 regulates mitochondrial morphology. J. Cell Biol. 182, 573–585
16 Taguchi, N. et al. (2007) Mitotic phosphorylation of dynamin-relatedGTPase Drp1 participates in mitochondrial fission. J. Biol. Chem.282, 11521–11529
17 Harder, Z. et al. (2004) Sumo1 conjugates mitochondrial substratesand participates in mitochondrial fission. Curr. Biol. 14, 340–345
18 Wasiak, S. et al. (2007) Bax/Bak promote sumoylation of DRP1 and itsstable association with mitochondria during apoptotic cell death. J.Cell Biol. 177, 439–450
19 Suzuki, M. et al. (2003) The solution structure of humanmitochondriafission protein Fis1 reveals a novel TPR-like helix bundle. J.Mol. Biol.334, 445–458
20 James, D.I. et al. (2003) hFis1, a novel component of the mammalianmitochondrial fission machinery. J. Biol. Chem. 278, 36373–36379
21 Yoon, Y. et al. (2003) The mitochondrial protein hFis1 regulatesmitochondrial fission in mammalian cells through an interactionwith the dynamin-like protein DLP1. Mol. Cell. Biol. 23, 5409–5420
22 Yu, T. et al. (2005) Regulation of mitochondrial fission and apoptosisby the mitochondrial outer membrane protein hFis1. J. Cell Sci. 118,4141–4151
23 Hoppins, S. et al. (2007) The machines that divide and fusemitochondria. Annu. Rev. Biochem. 76, 751–780
24 Ingerman, E. et al. (2005) Dnm1 forms spirals that are structurallytailored to fit mitochondria. J. Cell Biol. 170, 1021–1027
25 Pucadyil, T.J. and Schmid, S.L. (2008) Real-time visualization ofdynamin-catalyzed membrane fission and vesicle release. Cell 135,1263–1275
26 Bashkirov, P.V. et al. (2008) GTPase cycle of dynamin is coupled tomembrane squeeze and release, leading to spontaneous fission. Cell135, 1276–1286
27 Karbowski, M. et al. (2002) Spatial and temporal association of Baxwith mitochondrial fission sites, Drp1, and Mfn2 during apoptosis. J.Cell Biol. 159, 931–938
28 Rojo, M. et al. (2002) Membrane topology and mitochondrial targetingof mitofusins, ubiquitous mammalian homologs of thetransmembrane GTPase Fzo. J. Cell Sci. 115, 1663–1674

Review Trends in Endocrinology and Metabolism Vol.20 No.6
29 Ishihara, N. et al. (2004) Mitofusin 1 and 2 play distinct roles inmitochondrial fusion reactions via GTPase activity. J. Cell Sci. 117,6535–6546
30 Koshiba, T. et al. (2004) Structural basis of mitochondrial tethering bymitofusin complexes. Science 305, 858–862
31 Chen, H. et al. (2003)MitofusinsMfn1 andMfn2 coordinately regulatemitochondrial fusion and are essential for embryonic development. J.Cell Biol. 160, 189–200
32 Eura, Y. et al. (2003) Twomitofusin proteins, mammalian homologuesof FZO, with distinct functions are both required for mitochondrialfusion. J. Biochem. 134, 333–344
33 Bach, D. et al. (2003) Mitofusin-2 determines mitochondrial networkarchitecture and mitochondrial metabolism. A novel regulatorymechanism altered in obesity. J. Biol. Chem. 278, 17190–17197
34 Chen, K.H. et al. (2004) Dysregulation of HSG triggers vascularproliferative disorders. Nat. Cell Biol. 6, 872–883
35 de Brito, O.M. and Scorrano, L. (2008) Mitofusin 2 tethersendoplasmic reticulum to mitochondria. Nature 456, 605–610
36 Zuchner, S. et al. (2004) Mutations in the mitochondrial GTPasemitofusin 2 cause Charcot-Marie-Tooth neuropathy type 2A. Nat.Genet. 36, 449–451
37 Cipolat, S. et al. (2004) OPA1 requires mitofusin 1 to promotemitochondrial fusion. Proc. Natl. Acad. Sci. U. S. A. 101, 15927–15932
38 Neuspiel, M. et al. (2005) Activated Mfn2 signals mitochondrialfusion, interferes with Bax activation and reduces susceptibility toradical induced depolarization. J. Biol. Chem. 280, 25060–25070
39 Akepati, V.R. et al. (2008) Characterization of OPA1 isoforms isolatedfrom mouse tissues. J. Neurochem. 106, 372–383
40 Duvezin-Caubet, S. et al. (2007) OPA1 processing reconstituted inyeast depends on the subunit composition of the m-AAA protease inmitochondria. Mol. Biol. Cell 18, 3582–3590
41 Ishihara, N. et al. (2006) Regulation of mitochondrial morphologythrough proteolytic cleavage of OPA1. EMBO J. 25, 2966–2977
42 Griparic, L. et al. (2007) Regulation of themitochondrial dynamin-likeprotein Opa1 by proteolytic cleavage. J. Cell Biol. 178, 757–764
43 Song, Z. et al. (2007) OPA1 processing controls mitochondrial fusionand is regulated by mRNA splicing, membrane potential, and Yme1L.J. Cell Biol. 178, 749–755
44 Cipolat, S. et al. (2006) Mitochondrial rhomboid parl regulatescytochrome c release during apoptosis via Opa1-dependent cristaeremodeling. Cell 126, 163–175
45 Olichon, A. et al. (2003) Loss of OPA1 perturbates the mitochondrialinner membrane structure and integrity, leading to cytochrome crelease and apoptosis. J. Biol. Chem. 278, 7743–7746
46 Frezza, C. et al. (2006) OPA1 Controls Apoptotic Cristae RemodelingIndependently from Mitochondrial Fusion. Cell 126, 177–189
47 Tondera, D. et al. (2004) Knockdown of MTP18, a novelphosphatidylinositol 3-kinase-dependent protein, affectsmitochondrial morphology and induces apoptosis. J. Biol. Chem.279, 31544–31555
48 Tondera, D. et al. (2005) The mitochondrial protein MTP18contributes to mitochondrial fission in mammalian cells. J. CellSci. 118, 3049–3059
49 Karbowski, M. et al. (2004) Endophilin B1 is required for themaintenance of mitochondrial morphology. J. Cell Biol. 166, 1027–
103950 Takahashi, Y. et al. (2005) Loss of Bif-1 suppresses Bax/Bak
conformational change and mitochondrial apoptosis. Mol. Cell. Biol.25, 9369–9382
51 Etxebarria, A. et al. (2009) Endophilin B1/Bif-1 stimulates BAXactivation independently from its capacity to produce large scalemembrane morphological rearrangements. J. Biol. Chem. 284,4200–4212
52 Eura, Y. et al. (2006) Identification of a novel protein that regulatesmitochondrial fusion by modulating mitofusin (Mfn) protein function.J. Cell Sci. 119, 4913–4925
53 Gandre-Babbe, S. and van der Bliek, A.M. (2008) The novel tail-anchored membrane protein Mff controls mitochondrial andperoxisomal fission in mammalian cells. Mol. Biol. Cell 19, 2402–
241254 Choi, S.Y. et al. (2006) A common lipid links Mfn-mediated
mitochondrial fusion and SNARE-regulated exocytosis. Nat. CellBiol. 8, 1255–1262
55 Nemoto, Y. and De Camilli, P. (1999) Recruitment of an alternativelyspliced form of synaptojanin 2 to mitochondria by the interaction withthe PDZ domain of a mitocondrial outer membrane protein. EMBO J.18, 2991–3006
56 Breckenridge, D.G. et al. (2008)Caenorhabditis elegans drp-1 and fis-2regulate distinct cell-death execution pathways downstream of ced-3and independent of ced-9. Mol. Cell 31, 586–597
57 Yuan, H. et al. (2007) Mitochondrial fission is an upstream andrequired event for bax foci formation in response to nitric oxide incortical neurons. Cell Death Differ. 14, 462–471
58 Sheridan, C. et al. (2008) Bax- or Bak-induced mitochondrial fissioncan be uncoupled from cytochrome c release. Mol. Cell 31, 570–585
59 Estaquier, J. and Arnoult, D. (2007) Inhibiting Drp1-mediatedmitochondrial fission selectively prevents the release of cytochromec during apoptosis. Cell Death Differ. 14, 1086–1094
60 Cassidy-Stone, A. et al. (2008) Chemical inhibition of themitochondrial division dynamin reveals its role in Bax/Bak-dependent mitochondrial outer membrane permeabilization. Dev.Cell 14, 193–204
61 Nakamura, N. et al. (2006) MARCH-V is a novel mitofusin 2- andDrp1-binding protein able to change mitochondrial morphology.EMBO Rep. 7, 1019–1022
62 Yonashiro, R. et al. (2006) A novel mitochondrial ubiquitin ligaseplays a critical role in mitochondrial dynamics. EMBO J. 25, 3618–
362663 Karbowski, M. et al. (2007) The mitochondrial E3 ubiquitin ligase
MARCH5 is required for Drp1 dependent mitochondrial division. J.Cell Biol. 178, 71–84
64 Karbowski, M. et al. (2006) Role of Bax and Bak in mitochondrialmorphogenesis. Nature 443, 658–662
65 Huang, P. et al. (2007) Mitochondrial clustering induced byoverexpression of the mitochondrial fusion protein Mfn2 causesmitochondrial dysfunction and cell death. Eur. J. Cell Biol. 86,289–302
66 Jahani-Asl, A. et al. (2007) Mitofusin 2 protects cerebellar granuleneurons against injury induced cell death. J. Biol. Chem. 282, 23788–
2379867 Kuwana, T. et al. (2002) Bid, Bax, and lipids cooperate to form
supramolecular openings in the outer mitochondrial membrane.Cell 111, 331–342
68 Vance, J.E. (1990) Phospholipid synthesis in a membrane fractionassociated with mitochondria. J. Biol. Chem. 265, 7248–7256
69 Guo, X. et al. (2007) Mitofusin 2 triggers vascular smooth muscle cellapoptosis viamitochondrial death pathway.Circ. Res. 101, 1113–1122
70 Alirol, E. et al. (2006) Themitochondrial fission protein hFis1 requiresthe endoplasmic reticulum gateway to induce apoptosis. Mol. Biol.Cell 17, 4593–4605
71 Yamaguchi, R. et al. (2008) Opa1-mediated cristae opening is Bax/Bakand BH3 dependent, required for apoptosis, and independent of Bakoligomerization. Mol. Cell 31, 557–569
72 Germain, M. et al. (2005) Endoplasmic reticulum BIK initiates DRP1-regulated remodelling of mitochondrial cristae during apoptosis.EMBO J. 24, 1546–1556
73 Sun, M.G. et al. (2007) Correlated three-dimensional light andelectron microscopy reveals transformation of mitochondria duringapoptosis. Nat. Cell Biol. 9, 1057–1065
74 Danial, N.N. and Korsmeyer, S.J. (2004) Cell death: critical controlpoints. Cell 116, 205–219
75 Hackenbrock, C.R. (1966) Ultrastructural bases for metabolicallylinked mechanical activity in mitochondria. I. Reversibleultrastructural changes with change in metabolic steady state inisolated liver mitochondria. J. Cell Biol. 30, 269–297
76 Perkins, G.A. et al. (2001) Membrane architecture of mitochondria inneurons of the central nervous system. J. Neurosci. Res. 66, 857–865
77 Perkins, G.A. et al. (2003) Three-dimensional analysis of mouse rodand cone mitochondrial cristae architecture: bioenergetic andfunctional implications. Mol. Vis. 9, 60–73
78 Schlame, M. et al. (1999) Microanalysis of cardiolipin in small biopsiesincluding skeletal muscle from patients with mitochondrial disease.J. Lipid Res. 40, 1585–1592
79 Gottlieb, E. et al. (2003) Mitochondrial membrane potential regulatesmatrix configuration and cytochrome c release during apoptosis. CellDeath Differ. 10, 709–717
293

Review Trends in Endocrinology and Metabolism Vol.20 No.6
80 Meeusen, S. et al. (2004) Mitochondrial fusion intermediates revealedin vitro. Science 305, 1747–1752
81 Ishihara, N. et al. (2003) Regulation of mitochondrial morphology bymembrane potential, and DRP1-dependent division and FZO1-dependent fusion reaction in mammalian cells. Biochem. Biophys.Res. Commun. 301, 891–898
82 Chen, H. et al. (2007) Mitochondrial fusion protects againstneurodegeneration in the cerebellum. Cell 130, 548–562
83 Parone, P.A. et al. (2008) Preventing mitochondrial fission impairsmitochondrial function and leads to loss of mitochondrial DNA. PLoSOne 3, e3257
84 Twig, G. et al. (2008) Fission and selective fusion governmitochondrial segregation and elimination by autophagy. EMBO J.27, 433–446
85 Paumard, P. et al. (2002) The ATP synthase is involved in generatingmitochondrial cristae morphology. EMBO J. 21, 221–230
86 Strauss, M. et al. (2008) Dimer ribbons of ATP synthase shape theinner mitochondrial membrane. EMBO J. 27, 1154–1160
87 Campanella, M. et al. (2008) Regulation of mitochondrial structureand function by the F1Fo-ATPase inhibitor protein, IF1.CellMetab. 8,13–25
88 Baloh, R.H. et al. (2007) Altered axonal mitochondrial transport in thepathogenesis of Charcot-Marie-Tooth disease from mitofusin 2mutations. J. Neurosci. 27, 422–430
89 Amiott, E.A. et al. (2008) Mitochondrial fusion and function inCharcot-Marie-Tooth type 2A patient fibroblasts with mitofusin 2mutations. Exp. Neurol. 211, 115–127
90 Niemann, A. et al. (2005) Ganglioside-induced differentiationassociated protein 1 is a regulator of the mitochondrial network: newimplications for Charcot-Marie-Tooth disease. J. Cell Biol. 170, 1067–
1078
294
91 Alexander, C. et al. (2000) OPA1, encoding a dynamin-relatedGTPase, is mutated in autosomal dominant optic atrophy linked tochromosome 3q28. Nat. Genet. 26, 211–215
92 Delettre, C. et al. (2000) Nuclear gene OPA1, encoding amitochondrial dynamin-related protein, is mutated in dominantoptic atrophy. Nat. Genet. 26, 207–210
93 Davies, V.J. et al. (2007) Opa1 deficiency in a mouse model ofautosomal dominant optic atrophy impairs mitochondrialmorphology, optic nerve structure and visual function. Hum. Mol.Genet. 16, 1307–1318
94 Spinazzi, M. et al. (2008) A novel deletion in the GTPase domain ofOPA1 causes defects in mitochondrial morphology and distribution,but not in function. Hum. Mol. Genet. 17, 3291–3302
95 Clark, I.E. et al. (2006) Drosophila pink1 is required for mitochondrialfunction and interacts genetically with parkin. Nature 441, 1162–
116696 Park, J. et al. (2006) Mitochondrial dysfunction in Drosophila
PINK1 mutants is complemented by parkin. Nature 441, 1157–
116197 Poole, A.C. et al. (2008) The PINK1/Parkin pathway regulates
mitochondrial morphology. Proc. Natl. Acad. Sci. U. S. A. 105,1638–1643
98 Gautier, C.A. et al. (2008) Loss of PINK1 causes mitochondrialfunctional defects and increased sensitivity to oxidative stress.Proc. Natl. Acad. Sci. U. S. A. 105, 11364–11369
99 Morais, V.A. et al. (2009) Parkinson’s disease mutations in PINK1result in decreased Complex I activity and deficient synaptic function.EMBO Mol Med 1, 1–14
100 Narendra, D. et al. (2008) Parkin is recruited selectively to impairedmitochondria and promotes their autophagy. J. Cell Biol. 183,795–803